Back to Journals » Neuropsychiatric Disease and Treatment » Volume 21
Anxiolytic Effects of Natural Products: Pathological Mechanisms, Molecular Targets, Therapeutic Prospects
Authors Yu X, Li G, Liu J, Chen Y, Wang F
Received 1 October 2025
Accepted for publication 19 November 2025
Published 29 November 2025 Volume 2025:21 Pages 2623—2653
DOI https://doi.org/10.2147/NDT.S571692
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Roger Pinder
Xiaowen Yu,1 Guoqiang Li,2 Juan Liu,3 Yinling Chen,4 Feng Wang4
1Division 4 of Neurology Department, Affiliated Hospital of Shandong University of Traditional Chinese Medicine, Jinan, People’s Republic of China; 2Department of Pain Medicine, The First Affiliated Hospital of Shandong First Medical University and Shandong Provincial Qianfoshan Hospital, Jinan, People’s Republic of China; 3Department of Traditional Chinese Medicine, Shandong Rehabilitation Hospital, Jinan, People’s Republic of China; 4Department of Neurology, Shandong Second Provincial General Hospital, Jinan, People’s Republic of China
Correspondence: Feng Wang, Email [email protected]
Abstract: Anxiety disorders are complex neuropsychiatric conditions with incompletely understood mechanisms, which hinders the development of targeted therapies. Natural products have attracted attention as promising anxiolytic candidates due to their multi-component nature, synergistic effects, and ability to modulate multiple molecular targets and signaling pathways. However, a systematic integration of anxiety pathology with the mechanisms of natural products is still lacking. This review summarizes recent advances in the molecular and cellular basis of anxiety and explores how natural products (such as flavonoids, terpenoids, and alkaloids) interact with key genes, receptors, and intracellular signaling pathways. It also addresses current research limitations, including insufficient depth of mechanistic studies, lack of standardized models, and challenges in clinical translation. By synthesizing mechanistic and therapeutic evidence, this work aims to support the development of effective, evidence-based natural anxiolytics with improved clinical applicability.
Keywords: anxiety disorder, pathological mechanism, natural product, anxiolytic, clinical translation
Introduction
Anxiety disorders, including panic disorder (with or without agoraphobia), generalized anxiety disorder, social anxiety disorder, specific phobias, and separation anxiety disorder, are the most prevalent mental health conditions globally, contributing to substantial healthcare costs and a significant disease burden. Large-scale population studies suggest that up to 33.7% of individuals will experience anxiety-related symptoms at some point in their lives, with global prevalence estimates ranging widely from 3.8% to 25%. Furthermore, chronic comorbid conditions affect up to 70% of those with anxiety disorders, exacerbating the impact on overall health. These disorders can severely impair daily functioning and quality of life. In clinical practice, first-line treatments typically include selective serotonin reuptake inhibitors, serotonin-norepinephrine reuptake inhibitors, and benzodiazepines. While these conventional treatments can effectively relieve symptoms, provide symptomatic relief, approximately one-third of patients do not respond adequately, highlighting the limitations in current treatment options and the need for more effective therapies.1–4
More importantly, as a complex mental health condition, the molecular and neurobiological mechanisms underlying anxiety disorders remain poorly understood, posing significant challenges to the development of targeted therapeutic strategies. Consequently, a systematic analysis of the mechanisms involved in the occurrence and progression of anxiety disorders, the development of new anti-anxiety drugs with enhanced safety and efficacy, and the identification of their molecular targets and signaling pathways have become key scientific challenges in the field of mental health research, which urgently require innovative breakthroughs.
Natural products are metabolites derived from a wide range of sources, including animals, plants, insects, microorganisms, marine organisms, and even human and animal bodies. Their unique and multifaceted mechanisms of action enable them to overcome the limitations of conventional Western medicines, which often target single molecular pathways. This makes natural products a promising avenue for developing treatments for complex diseases. International research shows that the majority of new chemical entity drugs are either directly derived from natural products or are structural derivatives, underscoring their critical role in modern drug discovery. As a result, natural products not only serve as a valuable resource for disease treatment but also drive innovation in drug development, continually unlocking significant market potential and industrial value in the healthcare sector.5,6
Recent advancements in molecular biology technologies have greatly enhanced our understanding of the pathological mechanisms underlying anxiety disorders. Meanwhile, natural products have demonstrated unique advantages in anti-anxiety treatment due to their ability to regulate multiple molecular targets. However, several key challenges persist in current research, including fragmented insights into the underlying mechanisms, difficulty in identifying active pharmacological metabolites, and poorly defined target networks. These limitations have significantly hindered the development of natural product-based anti-anxiety therapies.
This review systematically explored the cascade of mechanisms underlying anxiety disorders through a comprehensive analysis of relevant literature and examined the intervention properties of natural products from multiple perspectives. Additionally, it provided an in-depth evaluation of the limitations in current research, laying the groundwork for the development and clinical translation of precision anti-anxiety therapies based on natural products.
Review Method
To investigate the pathological mechanisms of anxiety disorders and the potential role of natural products in their treatment, we conducted a comprehensive literature search across PubMed, Web of Science, and ScienceDirect databases. The search terms included: “natural product,” “anxiety,” “anxiolytic,” “anxiety disorder,” “neuroinflammation,” “neuroplasticity,” “synaptic plasticity,” “microbial-gut-brain axis,” “glial cell,” “astrocyte,” “mitochondrion,” “neurogenesis,” “autophagy,” “epigenetics,” “glutamate,” “hypothalamic-pituitary-adrenal axis,” and “microglia.” Articles were independently reviewed by two researchers based on titles, abstracts, and full texts, in accordance with predefined inclusion and exclusion criteria. The inclusion criteria were: (1) original research articles published in English, and (2) studies investigating the mechanisms of natural products in the treatment of anxiety disorders. The exclusion criteria were: (1) articles published in languages other than English; (2) gray literature; (3) editorials; and (4) duplicate publications.
Pathological Mechanism of Anxiety Disorder
Neuroinflammation
Neuroinflammation is primarily characterized by the activation of innate immune cells in the CNS, leading to the release of pro-inflammatory cytokines and other bioactive substances. Neuroinflammation plays an important role in the pathogenesis of anxiety disorders, with microglia (resident immune cells of CNS) being especially involved. For example, in Rev-erbα knockout mice, increased levels of pro-inflammatory cytokines in the hippocampus, along with dysfunctional microglial activity, have been linked to anxiety-like behaviors.7,8 Additionally, a study demonstrated that a deficiency in the ten-eleven translocation 2 (TET2) protein promotes anxiety-like behaviors by activating the NLR family pyrin domain containing 3 (NLRP3)/interleukin-1β (IL-1β) inflammatory pathway in microglia.9 In addition, dysfunction of the P2X7 receptor (P2X7R), a key regulator of neuroinflammation, has been associated with emotional disturbances, such as anxiety, through psychological stress-induced demyelination in the amygdala. This effect is mediated via the activation of the P2X7R/NLRP3 inflammasome cascade.10,11 Moreover, ATP-induced activation of P2X7Rs in the hippocampus stimulates microglia, contributing to pain-related anxiety and depression-like behaviors through IL-1β signaling.12
Moreover, microglia also mediate anxiety induced by environmental factors. For instance, exposure to polystyrene nanoplastics during early adulthood activates both microglia and astrocytes in mice, resulting in persistent neuroinflammation and anxiety-like behavior that lasts up to ten months post-exposure.13 In endocrine-related conditions, hyperglycemia upregulates the expression of the neuronal transcription factor tonicity-responsive enhancer binding protein, which in turn promotes increased expression of C-C motif chemokine ligand 2 (CCL2) in the medial prefrontal cortex (PFC) and ventral hippocampus. CCL2 activates microglia through its receptor CCR2, triggering the release of pro-inflammatory cytokines and inducing anxiety-like behavior.14 Treatment with extracellular vesicles derived from BV2 cells significantly reduced pro-inflammatory gene expression in microglia and alleviated anxiety-like behavior.15 Additionally, acid sphingomyelinase-regulated anxiety-like behavior has been linked to microglial activation via the Toll-like receptor (TLR) signaling pathway.16 Adipose triglyceride lipase (ATGL)-mediated lipolysis of triglycerides leads to the production of inflammatory lipid mediators, which stimulate microglial inflammatory responses. Inhibiting ATGL reduces neuroinflammation and improves behavioral abnormalities, including anxiety.17 Furthermore, the pro-inflammatory cytokines interleukin-17A (IL-17A) and interleukin-17C (IL-17C) induce anxiety-like behavior by increasing the firing frequency of IL-17RA-positive neurons in the basolateral amygdala (BLA), while the anti-inflammatory cytokine interleukin-10 (IL-10) exerts anxiolytic effects by reducing the firing frequency of these neurons via IL-10RA signaling18 (Figure 1).
 |
Figure 1 Neuroinflammation in anxiety disorders. Abbreviations: CNS, central nervous system; NLRP3, NLR Family Pyrin Domain Containing 3; IL-1β, interleukin-1β; BDNF, brain-derived neurotrophic factor; ROS, reactive oxygen species; ATP, adenosine triphosphate. |
Overall, neuroinflammatory responses play an important role in the pathological development of anxiety disorders, particularly through inflammatory signaling mechanisms mediated by microglia. Among these mechanisms, the P2X7 signaling pathway is a key regulator of microglial activation, and its dysregulated activation has been implicated in the onset of anxiety-like behaviors by promoting the release of pro-inflammatory cytokines. However, the specific regulatory mechanisms of other neuroinflammation-related signaling pathways in anxiety disorders, beyond the P2X7 pathway, are not yet fully understood. Further systematic investigations are required to elucidate the spatiotemporal roles of these pathways in the occurrence and development of anxiety disorders.
Neuroplasticity
Synaptic Plasticity
Neuroplasticity, a fundamental concept in neuroscience, refers to the brain’s ability to change and adapt through the continuous formation of new neural connections. A critical aspect of neuroplasticity is synaptic plasticity, which involves dynamic modifications at the synapse level and underpins learning and memory processes.19 Disruption of synaptic plasticity plays a significant role in the pathological development of anxiety disorders. Anxiety-like behavior is often associated with the loss of synaptic plasticity and the downregulation of synaptic plasticity regulatory factors.20,21 Abnormal expression of genes related to synaptic plasticity, inflammation, and oxidative stress has been observed in anxiety-like behaviors induced by ovarian hormone withdrawal in mice.22 In addition, in a lipopolysaccharide (LPS)-induced model of depression and anxiety, synaptic loss in the hippocampal dentate gyrus (DG) and impaired microglial phagocytic function were observed in mice.23 Mechanistic studies have shown that, in anxiety-like behaviors, the deficiency of hippocampal Tachykinin receptor 3 (TACR3) disrupts long-term potentiation in the DG.24 The complex formed by tropomyosin receptor kinase C (TrkC) and presynaptic protein tyrosine phosphatase sigma (PTPσ) plays a critical role in excitatory synaptic maturation by regulating synaptic protein phosphorylation. Dysfunction of this complex can lead to synaptic developmental defects and anxiety-related behaviors.25 Moreover, indoleamine 2,3-dioxygenase 1 (IDO1) has been implicated in chronic migraine pain sensitivity and comorbid anxiety, as it regulates microglial activation and synaptic pruning in the anterior cingulate cortex (ACC).26
Gamma-aminobutyric acid (GABA) is a non-protein amino acid and a major neurotransmitter that plays a crucial role in promoting relaxation, alleviating depression and insomnia, and regulating synaptic transmission and neuronal development.27 GABA is closely related to synaptic plasticity and anxiety. A reduction in the number of GABAergic interneurons can impair cortical inhibitory neurotransmission, potentially leading to an imbalance between excitation and inhibition (E/I) in the PFC.28 Moreover, maternal immune activation (MIA) disrupts the GABAergic inhibitory balance in hippocampal CA1 pyramidal neurons of offspring, resulting in neurodevelopmental abnormalities and anxiety-like behavior.29 Further research has demonstrated that G protein-coupled GABAB receptors in dopamine neurons of the ventral tegmental area (VTA) interact with hyperpolarization-activated cyclic nucleotide-gated (HCN) channels at GABAergic synapses. This interaction serves as a negative feedback mechanism that limits the duration of inhibitory postsynaptic potentials (IPSPs), thereby helping to mitigate the development of anxiety.30 Chronic stress, on the other hand, increases Src kinase activity and tyrosine phosphorylation of calmodulin, which disrupts the interaction between MyosinVa and Neuroligin2, leading to reduced GABA transmission and heightened anxiety-like behavior.31
Neurogenesis
Neurogenesis refers to the generation of functional neurons from neural progenitor cells in the hippocampal DG, contributing to the repair and maintenance of neuronal circuits.32 Disruptions in neurogenesis can play a significant role in the development of anxiety-like behaviors. For example, the absence of serine/threonine protein kinase 24 (STK24), a protein involved in cell apoptosis, spinal cord synapse formation, and neuronal migration, disrupts hippocampal development, reduces adult hippocampal neurogenesis, and induces anxiety-like behavior.33 In antibiotic-treated mice exhibiting anxiety-like behavior, inhibition of the glycogen synthase kinase-3β (GSK-3β)/cyclic adenosine monophosphate (cAMP) response element binding protein (CREB) pathway, along with reduced brain-derived neurotrophic factor (BDNF) signaling, results in decreased neurogenesis in the ventral hippocampal DG.34 Additionally, mechanistic studies have shown that premature reduction of lamin B1 in adult neural stem/progenitor cells induces age-related anxiety-like behavior. Although the absence of lamin B1 initially promotes neurogenesis, it ultimately depletes the stem cell pool and reduces the survival of newly formed neurons, leading to a significant decline in neurogenic capacity.35 Interestingly, neurogenesis regulation has demonstrated potential anti-anxiety effects, such as the use of near-infrared light (940 nm/40 Hz) to enhance neurogenesis in the hippocampus.36
Glutamate System
Glutamate is the primary neurotransmitter in the vertebrate brain, with approximately one glutamatergic synapse per cubic millimeter of neural fibers. Upon presynaptic release, glutamate rapidly diffuses across the synaptic cleft, with only a small fraction binding to receptors at the postsynaptic site, thereby initiating neural effects.37 However, excessive glutamate input can lead to excitotoxicity, resulting in neuronal cell death following acute brain injury.38 Maternal-infant separation has been shown to alter the expression of metabotropic glutamate receptor 5 (mGluR5) and influence anxiety-related behaviors in adult rats.39 The hippocampus plays an important role in anxiety induced by peripheral inflammation; for example, activation of metabotropic glutamate receptor 1 in colitis mice triggers anxiety-like behaviors through overactivation of hippocampal glutamate synapses.40 In a chronic restraint stress anxiety model, enhanced presynaptic glutamate release from unidirectional projection neurons in the dorsomedial PFC (dmPFC) to the BLA results in an excitation/inhibition (E/I) imbalance favoring excitation.41 Long-term consumption of low-dose aspartame has also been shown to induce anxiety-like behaviors in both male and female mice by upregulating glutamate signaling-related genes in the amygdala while downregulating GABA-related genes, leading to increased excitability in the amygdala.42 Additionally, reducing glutamate levels, such as through pyruvate-mediated clearance of blood glutamate, has been shown to alleviate anxiety symptoms following traumatic brain injury.43
In summary, neuroplasticity plays a pivotal role in the pathophysiology and treatment of anxiety disorders, particularly through mechanisms involving synaptic plasticity, hippocampal neurogenesis, and the imbalance between the GABAergic inhibitory system and the glutamatergic excitatory system. Intervention strategies targeting these neuroplasticity mechanisms hold significant promise as emerging approaches for anti-anxiety therapies. Future research should focus on exploring the interactions between different neuroplasticity processes and developing a precise intervention framework based on multimodal regulation of plasticity. These advancements will be crucial not only for deepening our understanding of the underlying mechanisms but also for translating these insights into effective clinical applications for anxiety disorders.
Hypothalamic-Pituitary-Adrenal (HPA) Axis
The HPA axis is a complex network of positive and negative feedback mechanisms involving the pituitary gland, adrenal glands, and hypothalamus. In response to external stressors, the paraventricular nucleus of the hypothalamus releases corticotropin-releasing hormone (CRH), which stimulates the pituitary to secrete adrenocorticotropic hormone (ACTH). ACTH, in turn, prompts the adrenal cortex to produce glucocorticoids, including cortisol. Cortisol exerts its effects through a negative feedback mechanism that prevents excessive activation of the HPA axis, helping to maintain homeostasis. Both positive and negative feedback processes regulate essential physiological functions, such as immunity, fertility, and the body’s stress response. Disruptions in these processes are implicated in the pathophysiology of anxiety disorders,44,45 with HPA axis dysregulation commonly observed in animal models. Notably, elevated levels of CRH and CRHR1 proteins have been found in the amygdala, contributing to the progression of anxiety-like behaviors.46,47
Mechanistic studies have demonstrated that transgenic mice overexpressing myristoylated alanine-rich C-kinase substrate-like 1 (MARCKSL1) exhibit significant anxiety-like behaviors. These behaviors are characterized by increased activity of CRH neurons in the paraventricular nucleus (PVN) of the hypothalamus, elevated CRH mRNA expression, and higher plasma cortisol levels, suggesting hyperactivity of the HPA axis.48 In gender-specific studies, female mice exhibited more pronounced anxiety-like behaviors immediately after stress exposure. Furthermore, the oval nucleus of the bed nucleus of the stria terminalis in female mice showed both higher levels and prolonged release of corticotropin-releasing factor (CRF) compared to males. CRF neurons in females also exhibited significantly higher expression levels of CRF receptor 1 (CRFR1) than those in males.49 In addition, regulation of the HPA axis, such as through the use of dexmedetomidine, has been shown to alleviate acute stress-induced anxiety-like behaviors in mice by inhibiting the neuronal activity of CRH in the PVN50 (Figure 2).
 |
Figure 2 HPA axis in anxiety disorders. Abbreviations: CRH, corticotropin-releasing hormone; ACTH, adrenocorticotropic hormone. |
In summary, the occurrence and development of anxiety disorders are closely associated with dysfunction of the HPA axis, particularly with overactivity of CRH and the heightened expression of CRFR1 in females compared to males. This gender-specific difference may provide a neurobiological basis for the observed gender disparities in the onset of anxiety disorders. In addition, intervention strategies targeting CRH regulation have shown promising potential for the treatment of anxiety. However, the molecular mechanisms underlying HPA axis dysfunction in anxiety disorders remain insufficiently understood. Further research is needed, especially to elucidate the causal relationship between HPA axis dysregulation and the pathological processes of anxiety. These studies will provide the theoretical foundation necessary for the development of precise diagnostic and therapeutic strategies.
Microbial-Gut-Brain Axis
The microbial population in the human gastrointestinal system is diverse, and its extensive genetic and metabolic capacity plays a crucial role in various aspects of human biology, including development, aging, disease, and health maintenance. One of the most significant interactions is the “microbiota-gut-brain” axis, which facilitates bidirectional communication between the gut microbiota and the CNS.51 Growing evidence links gut microbiota composition with anxiety and depression disorders. For example, patients with anxiety disorders often exhibit a higher relative abundance of pro-inflammatory bacteria and a lower abundance of short-chain fatty acid (SCFA)-producing bacteria.52 In addition, dysbiosis of the gut microbiota in individuals with ulcerative colitis (UC), such as the presence of Enterococcus mundtii, has been linked to anxiety and depression-like behaviors. This is thought to occur via the downregulation of colon neuropeptide Y (NPY) expression, leading to reduced peripheral and central NPY levels. Research shows that the reduction of NPY is a driving factor for anxiety like behavior.53 Further investigations into UC-related anxiety have revealed that gut microbiota dysbiosis, metabolic disturbances, and a decrease in immune proteins contribute to systemic inflammation, which affects CNS function through the microbiota-gut-brain axis.54 In preclinical studies, maternal-infant separation has been shown to disrupt gut microbiota composition in mice, significantly reducing the abundance of Lactobacillus reuteri and impairing the expression of intestinal amino acid transporters. This reduction in amino acids, particularly glutamine, limits the synthesis of glutamate (Glu) and GABA in the medial PFC (mPFC), resulting in synaptic dysfunction, social deficits, and anxious behavior. It is also a driving factor for anxiety like behavior.55 Moreover, exposure to 3.5 GHz radiofrequency radiation has been shown to induce metabolic disorders by causing gut microbiota dysbiosis, activating NLRP3-dependent pyroptosis in hippocampal CA3 neurons, and ultimately contributing to anxiety-like behaviors in mice.56
On the other hand, emerging evidence suggests that the “microbiota-gut-brain” axis also plays a key role in mediating anti-anxiety effects. For instance, short-term psychological stress can induce intestinal permeability, exposing microbial products that activate intestinal immune responses. This activation promotes the differentiation of T helper 17 cells, which rely on IL-1β signaling to produce IL-22. Elevated IL-22 then crosses the damaged blood-brain barrier (BBB) and enters the brain septum, where it directly interacts with neurons expressing IL-22 receptors, inhibiting their activation and alleviating anxiety-like behavior.57 Additionally, intestinal-derived lactate has been shown to shift tryptophan metabolism from the neurotoxic kynurenine pathway toward the synthesis of 5-hydroxytryptamine (5-HT)), by upregulating tryptophan hydroxylase (TPH1) and inhibiting IDO1. Meanwhile, Akkermansia muciniphila enhances propionic acid production in a lactate-enriched environment, collectively alleviating anxiety behavior in mice while repairing intestinal barrier function.58 Furthermore, fluoxetine, a commonly used anti-anxiety and anti-depressant drug, has been shown to increase the abundance of Lactobacillus species in the gut, which in turn regulates the expression of serotonin receptors (5-HT1A, 5-HT2C) and melanocortin 4 receptors via the enteric nervous system.59 In conclusion, accumulating research highlights the critical role of the “microbiota-gut-brain” axis in the onset and progression of anxiety disorders. Targeted modulation of the gut microbiota and its interaction with the CNS may offer promising therapeutic strategies for the development of novel anti-anxiety treatments, grounded in a deeper understanding of its pathophysiological mechanisms (Figure 3).
 |
Figure 3 Microbial-gut-brain axis in anxiety disorders. |
Mitochondrial Quality Control
In eukaryotic cells, mitochondria are the primary organelles responsible for energy production, playing a crucial role in regulating cellular metabolism. Beyond energy production, mitochondria are essential for various biological functions, including redox homeostasis, immune responses, differentiation, and cell proliferation. The mitochondrial quality control (MQC) system, which includes processes such as mitochondrial biogenesis, dynamics, and autophagy, is continuously modulated by a variety of physiological signals and external stimuli.60 Mitochondrial dysfunction is frequently observed in individuals with high anxiety traits or anxiety disorders, manifesting as abnormal energy metabolism, increased oxidative stress, dysregulated neurosteroid synthesis, and alterations in mitochondrial DNA copy number.61 Large-scale data analysis, such as that using the UK Biobank, suggests that mitochondrial function may mediate the effects of behavioral factors (such as alcohol consumption and smoking) on mental health through mechanisms like oxidative stress and impaired energy metabolism. For example, alcohol consumption has been shown to increase mitochondrial heterogeneity, leading to mitochondrial dysfunction and exacerbating anxiety and depression.62 Moreover, chronic ethanol exposure induces mitochondrial damage and the release of mitochondrial DNA, activating the cyclic GMP-AMP synthase (cGAS)/stimulator of interferon genes (STING) pathway. This activation triggers microglial activation and neuroinflammation in the PFC, contributing to anxiety-like behaviors.63
Further molecular biology studies involving neurons have highlighted the critical role of Mitofusin-2 (Mfn2) in the nucleus accumbens in regulating anxiety and depression-like behaviors. Mfn2 influences mitochondrial structure and the function of medium spiny neurons (MSNs), which are key to these behaviors.64 In addition, the expression of Mfn2 in D2 MSNs of the nucleus accumbens is a key determinant of social advantage, as it helps maintain mitochondrial function and neuronal excitability.65 Recent studies have also shown that polystyrene nanoparticles, with varying surface modifications, can accumulate in the mouse brain following oral exposure. This accumulation leads to abnormal mitochondrial morphology, decreased membrane potential, and reduced ATP production. The imbalance causes excessive mitochondrial fission and neuronal apoptosis, a process mediated by Dynamin-related protein 1 (Drp1), ultimately triggering anxiety-like behaviors.66
In terms of treatment, Early Handling has been shown to exert anti-anxiety effects in high-anxiety mice by regulating mitochondrial dynamics, specifically the balance between mitochondrial fission and fusion in the hypothalamus, as well as modulating energy metabolism pathways.67 Additionally, running track exercise enhances the Sirtuin 1 (SIRT1)-dependent proliferator-activated receptor gamma coactivator-1 alpha (PGC1α)/nuclear respiratory factor 1 (NRF1)/mitochondrial transcription factor A (TFAM) axis, which promotes hippocampal mitochondrial biogenesis and alleviates anxiety and depression-like behaviors in APP/PS1 mice.68 These findings suggest that targeting mitochondrial function regulation, such as modulating the balance of mitochondrial dynamics, could offer a promising strategy for developing anti-anxiety treatments.
Glial Cells
Glial cells are found in all organisms with a CNS. As the brain becomes more complex, the number, diversity, and function of glial cells have increased significantly.69 They are primarily categorized into astrocytes, microglia, oligodendrocytes, and oligodendrocyte precursor cells, collectively accounting for approximately half of the total cells in the mammalian CNS. Glial cells provide essential nutritional and metabolic support for neurons.70 Abnormal glial function plays a crucial role in the pathological development of anxiety. For instance, obesity can lead to the aging of astrocytes and microglia surrounding the lateral ventricles in mice, causing these aged cells to secrete pro-inflammatory cytokines that inhibit neural stem cell activity, reducing neurogenesis and contributing to anxiety-like behaviors.71 Additionally, dysregulated circadian rhythms can impair oligodendrocyte myelin formation via overexpression of brain and muscle ARNT-like protein-1 (Bmal1), which is linked to anxiety and depression-like behaviors.72 Mice subjected to unpredictable chronic mild stress and prolonged social isolation exhibit anxiety and depression-like behaviors, along with abnormal activation of microglia and astrocytes in the hippocampal DG region, which in turn impairs hippocampal neurogenesis.73 Furthermore, astrocyte-specific knockout of the melatonin receptor 2 (Mtnr1b) disrupts the excitation/inhibition (E/I) balance by downregulating the GABAergic signaling pathway, inducing anxiety-like behaviors.74 The absence of nucleotide transporters in astrocytes also leads to increased anxiety and depression-like behaviors and reduced reward motivation.75 Furthermore, disruptions in the mechanistic target of rapamycin (mTOR) complexes 1 and 2 (mTORC1 and mTORC2) in astrocytes of the VTA are associated with anxiety and depression, with mTORC2 dysfunction specifically contributing to anxiety-like behavior in mice.76
These findings highlight the pathological role of abnormal glial cell function in anxiety-like behaviors, suggesting that targeted regulation of glial cells may offer promising therapeutic benefits. For example, ATP released by hippocampal astrocytes has been shown to enhance excitatory synaptic transmission in granule cells of the dentate gyrus (DG), contributing to anxiety-like effects.77 In addition, activation of the Gi-coupled G protein-coupled receptor (Gi-GPCR) signaling pathway in astrocytes alleviates anxiety phenotypes in Sapap3 gene knockout mice.78 Additionally, activating cannabinoid receptor 1 in astrocytes has been demonstrated to reduce anxiety and depression-like behaviors by regulating BBB function.79 Microglia, through CX3C chemokine ligand 1 (CX3CL1)-C-X3-C motif chemokine receptor 1 (CX3CR1)-dependent phagocytosis, inhibit GABACeA neuronal activity, promoting the resolution of anxiety after acute stress.80 Therefore, glial cells, especially astrocytes, play an essential regulatory role in the pathophysiology of anxiety disorders. Targeting specific molecular pathways within these cells holds significant potential for developing effective therapeutic strategies. While preclinical animal models have shown promising results, the translational application of these therapeutic targets requires further systematic validation, as there is considerable species heterogeneity in glial cell function.
Other Types
Autophagy is the primary intracellular degradation mechanism through which cytoplasmic materials are transported to lysosomes for breakdown. Beyond merely eliminating cellular debris, autophagy acts as a dynamic system, recycling components to produce new building blocks and energy essential for cellular regeneration and homeostasis.81 For instance, when combined with a high-calorie self-service diet, alpha-ketoglutarate exacerbates anxiety-like behaviors and upregulates autophagy-related gene expression.82 Additionally, nicotine activates the phosphatidylinositol-3-kinase (PI3K)/protein kinase B (Akt)/mechanistic target of rapamycin (mTOR) pathway, enhancing autophagy and alleviating hippocampal neuron damage and anxiety-like behaviors induced by chronic stress.83 Furthermore, the NLRP3 inflammasome-specific inhibitor MCC950 has been shown to improve cognitive function and reduce anxiety-like behaviors in Alzheimer’s disease model rats by inhibiting NLRP3 inflammasome-mediated neuroinflammation and excessive autophagy.84
DNA methylation regulates a wide range of biological activities, including embryonic development, transcription, chromatin structure, genomic imprinting, and chromosomal integrity.85 However, studies have shown that paclitaxel inhibits the expression of DNA methyltransferase 3a (DNMT3a), leading to DNA hypomethylation. This alteration results in the upregulation of GABA_A receptor β1 subunits and other related components in the mPFC, enhancing inhibitory synaptic transmission and mediating chemotherapy-related neuropathic pain and anxiety-like behaviors.86 In addition, mice exposed to social stress during adolescence exhibit increased anxiety-like behaviors in adulthood, a significant reduction in the low-frequency amplitude of mPFC activity, and a marked increase in histone H3K9 methylation levels in this region.87
Overall, the pathological mechanisms underlying anxiety disorders may be associated with abnormal autophagy regulation and the dysregulation of epigenetic modifications, including DNA methylation and histone methylation. However, current research has not yet fully elucidated the precise mechanisms through which these biological processes contribute to anxiety disorders. Further investigation into the molecular regulatory networks involved, as well as potential intervention strategies targeting these pathways, holds significant promise. In-depth studies could pave the way for the development of novel therapeutic targets for anxiety disorders in the future (Figure 4).
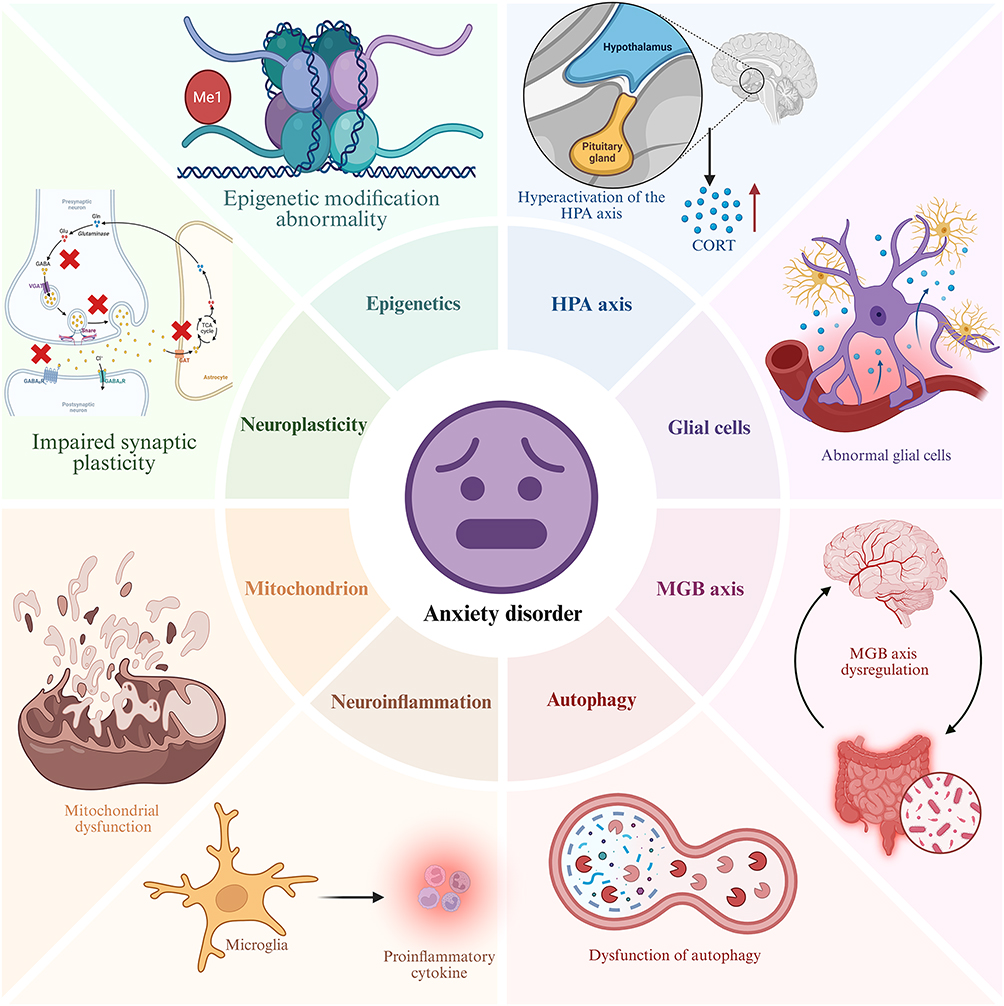 |
Figure 4 Pathological mechanism of anxiety disorder. Abbreviations: HPA, hypothalamic-pituitary-adrenal; MGB, Microbial-gut-brain. |
Molecular Mechanism of Natural Products
Flavonoids
Flavonoids, a diverse group of plant-derived secondary metabolites, are known for their broad pharmacological benefits, including anti-cancer, antioxidant, anti-inflammatory properties, and their ability to reduce vascular fragility.88 These compounds have also shown potential in the treatment of anxiety disorders. For instance, 7,8-dihydroxyflavone, a flavonoid, has been found to regulate glutamate transmission via the tropomyosin-related kinase B (TrkB) receptor in the BLA, helping alleviate alcohol-induced anxiety-like behavior caused by chronic alcohol exposure.89 Another flavonoid, cynaroside, derived from natural plants, has been shown to effectively improve anxiety-like behaviors in mice subjected to chronic unpredictable mild stress (CUMS). Its mechanism involves inhibiting the polarization of microglia toward the M1 pro-inflammatory phenotype, reducing neuroinflammation, and mitigating ferroptosis, which contributes to its anxiolytic effects.90 Additionally, Ziziphus jujuba Mill., a traditional Chinese medicine (TCM) known for its sedative properties, contains the flavonoid spinosin, which regulates the extracellular signal-regulated kinase 1/2 (ERK1/2)-CREB-BDNF signaling pathway, helping to alleviate anxiety-like behaviors.91
Astragalin, a natural flavonoid found in various edible plants such as green tea, mulberries, and dodder,92 has been shown to exert its anti-anxiety effects by modulating the ACC and lateral hypothalamus through the C-X-C chemokine receptor type 4 (CXCR4)-Beclin1/VPS34 signaling pathway. This modulation inhibits neuronal excitability and enhances autophagy, ultimately reducing pain sensitivity and anxiety-like behavior in mice.93 Another flavonoid, daidzin, derived from Glycine max (L). Merr., exhibits anti-anxiety effects through its antioxidant and anti-inflammatory properties, along with its ability to regulate the cholinergic system.94 Icariin, extracted from epimedium sagittatum (Siebold & Zucc). Maxim., alleviates anxiety-like behavior induced by hematopoietic shock in mice by inhibiting astrocyte activation, likely through NFκB-induced epigenetic reprogramming.95 In addition, cyanidin has been found to inhibit the IL-17A signaling pathway, reduce downstream inflammatory responses, and regulate glutamate transporter function in the amygdala, providing potential therapeutic benefits for anxiety caused by methylenedioxypyrovalerone (MDPV) withdrawal.96
Luteolin, a flavonoid found in various fruits, vegetables, and botanical drugs (such as Lonicera japonica Thunb), has demonstrated significant neuroprotective effects in multiple contexts, including inflammation, oxidation, and cancer.97 In animal models of latent depression, luteolin has been shown to regulate glycerophospholipid metabolism in both the hippocampus and PFC, contributing to the alleviation of anxiety and depression-like behaviors.98 Additionally, luteolin modulates the NFκB/NLRP3 inflammasome axis, reducing anxiety-like behaviors in rats subjected to sleep deprivation.99 It also exhibits therapeutic benefits in managing chronic pain-induced anxiety and depression-like symptoms, likely through its antioxidant, anti-inflammatory, and neuroprotective properties in both the hippocampus and PFC.100 In Parkinson’s disease models, luteolin has shown to have both anti-inflammatory and antioxidant effects, improving anxiety and depression-like behaviors in Parkinson’s rats.101
Quercetin, a widely recognized flavonoid, has shown promise in reducing anxiety through multiple mechanisms. It exerts its effects primarily by inhibiting neuroinflammation and microglial activation, preventing microglia from transforming into the M1 phenotype—a state associated with inflammation. This modulation occurs through the regulation of the TLR4 pathway, which is involved in immune responses and neuroinflammation. Additionally, quercetin has been found to reshape gut microbiota homeostasis, further contributing to the regulation of brain metabolism, which may help alleviate anxiety symptoms.102,103 Although quercetin is considered safe by the Food and Drug Administration for use under normal conditions (since 2010), it is still recommended to consult a doctor before taking high doses of quercetin, especially if combined with other medications, to avoid any potential interactions or side effects.104
Phenolic
Polyphenols, which are bioactive metabolites found in plants, offer a wide range of health benefits through various mechanisms, including cardiovascular protection, modulation of the gut microbiota, anti-inflammatory, antioxidant, and anti-obesity effects. TCM, an integral part of global healthcare, serves as an important source of natural medicinal resources that can inspire the development of modern pharmacological therapies. Among TCM materials, pony bark stands out as a representative medicinal plant, with its active metabolite, Paeonol, demonstrating significant effects on the CNS according to modern pharmacological studies.105 In the context of anxiety, Paeonol has been shown to effectively reverse anxiety-like behaviors in ovariectomized mice. This effect is attributed to its ability to restore the expression of the G protein-coupled receptor 30 (GPR30) in the PFC and hippocampus, and to enhance the activity of BDNF in these regions. Additionally, Paeonol activates the PI3K/Akt/mTOR signaling pathway, which plays a crucial role in regulating neuronal survival and synaptic plasticity.106 In addition, Paeonol inhibits the activation of the metabotropic glutamate receptor 5 (mGluR5) and gamma-aminobutyric acid type B receptor subunit 2 (GABBR2), and modulates β-arrestin2, thereby influencing the cAMP-protein kinase A (PKA) signaling cascade. This modulation helps alleviate anxiety symptoms, especially in conditions such as premenstrual dysphoric disorder and psychiatric symptoms associated with periodic hormonal changes. Paeonol’s action on these pathways also plays a role in preventing hippocampal injury.107
Preclinical studies indicate that Salidroside, a promising metabolite derived from Rhodiola rosea L., exerts a variety of pharmacological effects, including anti-inflammatory, antioxidant, and neuroregenerative properties through multiple signaling pathways.108 In a post-traumatic stress disorder (PTSD) model, Salidroside has been shown to reduce neuroinflammation, enhance hippocampal synaptic plasticity, and prevent hippocampal neuron apoptosis, thus helping to treat PTSD and alleviate anxiety-like behaviors.109 Curcumin, a natural polyphenolic metabolite extracted from turmeric, is a potent antioxidant and anti-inflammatory agent.110 However, its clinical application is limited by poor water solubility, instability, and low bioavailability. To address these challenges, there has been growing interest in synthesizing novel curcumin analogues to enhance its efficacy. For example, the curcumin analogue NL04 has been found to alleviate central sensitization in bone cancer pain by inhibiting the activation of the NLRP3 inflammasome, reducing IL-1β release, and significantly improving anxiety-like behaviors.111 Additionally, the polyphenol punicalin, derived from Punica granatum L., has demonstrated the ability to suppress anxiety induced by LPS through the TLR4/NFκB signaling pathway.112
Resveratrol, a well-known natural phenolic metabolite found in various foods (for example, grapes, etc),113 has been shown to exert anti-anxiety effects in the social isolation stress model by regulating the NFκB/NLRP3 signaling pathway.114 In addition, resveratrol enhances brain antioxidant levels, restores monoamine balance, and mitigates HPA axis dysfunction, providing notable neuroprotective effects.115 In adolescent models of social isolation, resveratrol alleviates anxiety-like behavior by modulating spinal plasticity and improving mitochondrial function in the nucleus accumbens of female mice.116 Furthermore, resveratrol has been shown to reduce neuroinflammation and microglial activation in the hippocampus in experimental cerebral palsy, thereby protecting against memory impairments and anxiety-like behaviors.117 These studies collectively highlight resveratrol’s significant anti-anxiety effects through a multi-target regulatory mechanism across different animal models. However, the bioavailability of resveratrol following oral administration remains low (<5%), and high doses may lead to nephrotoxicity, liver damage, and other potential side effects.118 Future research should focus on optimizing its pharmacokinetics, investigating its toxicity mechanisms, and developing novel delivery systems to enhance its therapeutic potential.
Terpenoids
Terpenes, a family of naturally occurring organic chemicals, are found in plants, insects, and microbes. β-Sitosterol, the most abundant plant sterol, is present in nearly all plant-based foods and some traditional Chinese botanical drugs. Due to its broad spectrum of biological functions, β-sitosterol has been used to treat various diseases.119 Recent studies suggest that β-sitosterol can alleviate anxiety symptoms associated with migraines, with its mechanism involving the improvement of oxidative/nitrosative stress and the enhancement of mitochondrial function.120 Geniposide, the primary bioactive metabolite of Gardenia jasminoides J.Ellis., is a naturally occurring iridoid glycoside with a variety of pharmacological properties, including anti-inflammatory, antioxidant, neuroprotective, and anti-diabetic effects.121 In a diabetes animal model, Bo et al demonstrated that Geniposide regulates CREB-mediated adult neurogenesis, which helps treat diabetes-related depression and anxiety.122
Glycyrrhizin, a triterpenoid glycoside found in the roots of licorice plants, is the primary active metabolite of Glycyrrhiza glabra L and exhibits a range of pharmacological activities.123 Glycyrrhizin has been shown to alleviate chronic hypoxia-induced white matter injury and oxygen-glucose deprivation-induced oligodendrocyte damage while improving anxiety-like behavior through the inhibition of the HMGB1/TLR4 pathway, exerting anti-inflammatory effects.124 In addition, glycyrrhizin enhances anxiety-like behavior by modulating Glutamate transporter 1 and Period (Per) 1/2-dependent pathways.125 Saffron, originally cultivated in East and Middle Eastern regions and later in Mediterranean countries,126 contains carotenoids that alleviate depression and anxiety in UCMS-induced rats. The mechanism underlying these effects involves the regulation of the BDNF-ERK-CREB signaling pathway.127 Moreover, quinoa saponin may serve as a natural dietary supplement for treating and preventing anxiety by regulating the gut-brain axis and inhibiting activation of the TLR4/MyD88/NFκB pathway.128
Artemisinin, derived from Artemisia annua L., is well-known for its potent anti-malaria properties. Recent studies have shown that artemisinin administration can alleviate PTSD-like symptoms in single prolonged stress rats, including anxiety, cognitive impairments, reduced socialization, and improvements in synaptic plasticity, while also reducing neuronal apoptosis.129 In PTSD animal models exhibiting anxiety, sodium aescinate has been shown to improve pathological behaviors by inhibiting NLRP3 inflammasome activation in the Dorsal Raphe Nuclei (DRN), reducing DRN apoptosis and alleviating mitochondrial damage.130 Limonene, a monoterpene found in citrus fruit peel oils,131 alleviates anxiety-related behaviors by modulating dopaminergic and GABAergic neuron activity via the A2A receptor.132 Similarly, geraniol, another natural monoterpene, has demonstrated anti-inflammatory, antioxidant, neuroprotective, and anti-cancer properties.133 It alleviates anxiety-like behavior in rats by reducing oxidative stress, repairing hippocampal neurotransmission, and normalizing cortical electroencephalography wave patterns following a single electric shock.134 In addition, the anti-anxiety effects of geraniol may be linked to its gastric healing properties, promoting gastric mucosa repair and reducing the recurrence of injury.135
Paeoniflorin, the main active metabolite of Paeonia lactiflora Pall.,136 alleviates visceral hypersensitivity and anxiety in irritable bowel syndrome (IBS)-like rats by regulating HPA axis activity and the BDNF/TrkB/phosphorylase Cγ1 (PLCγ1) signaling pathway.137 Furthermore, paeoniflorin demonstrates favorable toxicological characteristics, exhibiting low toxicity in acute toxicity tests without signs of genetic toxicity or mutagenicity.138 However, to ensure its clinical safety, continuous monitoring is essential, alongside further exploration of its mechanisms, particularly regarding pharmacokinetic interactions and adaptive responses during long-term use. These investigations would provide valuable scientific insights, facilitating a more comprehensive risk-benefit assessment for clinical applications.
Polysaccharide
Polysaccharides, along with proteins and nucleic acids, are essential biomacromolecules that play crucial roles in the growth and development of organisms. They are important metabolites of cell membranes and microbial cell walls in both higher plants and animals.139 Fucoidan, a polymer composed of l-fucose and l-fucosyl-4-sulfate, is naturally found in marine resources. Fucoidan has been shown to inhibit P-selectin, thereby preventing neutrophil aggregation at injury sites.140 Research by Li et al demonstrated that fucoidan downregulates the NLRP3 inflammasome pathway, increases mRNA levels of BDNF and PSD95 in the cerebral cortex and hippocampus, and regulates the intestinal microbiota. Additionally, fucoidan enhances intestinal barrier and BBB permeability, restores the abnormal structure of the intestinal microbiota, and alleviates UC and associated anxiety-like behaviors.141 Cichorium intybus L. oligosaccharides (CIOs), derived from chicory, are a mixture of oligosaccharides and polysaccharides. Treatment with CIOs increases 5-HT levels, reduces damage to hippocampal neurons, and upregulates the BDNF/ERK and PI3K/AKT/mTOR signaling pathways. These effects collectively contribute to the anti-anxiety and anti-depressive properties of CIOs in CUMS mice.142
Inulin, a soluble dietary fiber found in various plants as a reserve polysaccharide,143 has been shown to alleviate anxiety-like behaviors induced by constipation. This effect is mediated through several mechanisms, including the inhibition of neuroinflammation, prevention of synaptic ultrastructure damage, upregulation of tight junction protein expression, regulation of gut microbiota, and an increase in the concentration of SCFAs in fecal matter.143,144 Therefore, natural polysaccharides like fucoidan and inulin exert anti-anxiety effects via multi-target, multi-pathway regulatory mechanisms. These bioactive polysaccharides hold promise as new candidates for anti-anxiety therapies, potentially expanding the medicinal value of natural products. Future research should focus on exploring the structure-activity relationships, dose-response mechanisms, and synergistic effects of these polysaccharides, providing a solid theoretical foundation for the development of novel anti-anxiety treatment strategies based on polysaccharide molecules.
Alkaloid
Alkaloids, a class of nitrogen-containing organic metabolites found in plants, fungi, and some animals, are known for their alkaline characteristics and significant biological activities. Coptisine, a benzyltetrahydroisoquinoline alkaloid, is a key metabolite of Coptis chinensis Franch.145 In the LPS-induced anxiety model, administration of 60 mg/kg of coptisine significantly alleviates anxiety-like behavior. This effect is attributed to coptisine’s regulation of the Warburg effect in microglia via pyruvate kinase isoform M2.146 Berberine, a bioactive metabolite found in various Chinese botanical drugs, has been traditionally used for treating conditions like diarrhea and dysentery. In recent years, it has gained attention for its broad pharmacological effects and health benefits.147 In the context of anti-anxiety research, berberine has been shown to significantly alleviate anxiety-like behavior in mice with chronic pain. This effect is mediated by the inhibition of neurons projecting from the ventral lateral thalamus to the cingulate anterior cortex (Cg2).148
Moreover, berberine has low toxicity at conventional doses, offering clinical benefits without significant side effects, although mild gastrointestinal reactions have been reported in some patients.149 However, further research is needed to investigate the effects of other alkaloid-based natural products on normal physiological functions and tissue structures when exerting anti-anxiety pharmacological effects. A particularly important area for exploration is the systematic evaluation of their safe therapeutic window for selective action on the CNS.
Botanical Drugs Extracts
Botanical drugs extracts, which are concentrated active metabolites isolated from various plant parts (such as roots, stems, leaves, flowers, and fruits) through physical or chemical methods, are rich in bioactive compounds like alkaloids, flavonoids, and polysaccharides. These extracts are widely used in medicine, health products, cosmetics, and food. Hibiscus syriacus Linnaeus flower (HSF), also known as Mugunghwa, is an ornamental shrub and the national flower of South Korea. The extract of HSF and its saponins have been shown to alleviate sleep disorders and reduce anxiety-like behaviors. This effect is believed to be mediated by the regulation of GABA receptors, ventrolateral preoptic nucleus (VLPO) neuron activity, and neuropeptide expression.150 Passiflora incarnata L., a perennial plant commonly used in TCM, has long been recognized for its ability to alleviate stress. Recent studies have confirmed its effectiveness in treating insomnia, anxiety, and depression.151 Specifically, the hydroalcoholic extract of Passiflora incarnata L has been shown to improve Valproic acid-induced autism and anxiety-like behaviors through its antioxidant and neuroprotective properties.152 Echinacea purpurea (L). Moench., a member of the Asteraceae family, is widely known for its ability to regulate inflammatory conditions, such as skin inflammation, sore throats, and gum swelling. Recent studies suggest that its extract can help alleviate brain damage induced by Bifenthrin, reduce oxidative stress and improve behavioral disorders, including anxiety.153 In clinical trials, it has been found that kava (Piper methylsticum G. Forst) extract has a certain therapeutic effect on acute situational anxiety and is relatively safe.154 In addition, hops (Humulus lupulus L.) dry extract can alleviate patients’ anxiety.155
Essential Oils
Aromatherapy is a holistic treatment practice that uses the aromatic essences of plant essential oils, which have gained significant attention for their therapeutic potential in promoting overall health. Essential oils derived from plant metabolites have played an important role in aromatherapy as a complementary medical treatment approach.156 Bergamot essential oil (BEO), an extract from the bergamot fruit, has demonstrated significant neuroprotective effects.157 In a study by Zhu et al, inhalation of 1.0% BEO was found to significantly reduce anxiety-like behaviors. This effect was primarily mediated through the activation of anterior olfactory nucleus (AON) Glu neurons, which project to ACCGABA neurons, thereby indirectly inhibiting the activity of ACCGlu neurons, ultimately alleviating anxiety-like behavior.158 Patchouli essential oil has been shown to improve stress-induced anxiety and depression-like behaviors by regulating the gut microbiota, SCFAs production, and regulating neurotransmitter levels.159 Perilla frutescens (L). Britton essential oil has also been reported to alleviate behavioral disorders, including anxiety-like behaviors, in social stress mice. Its mechanism of action is thought to involve the regulation of cortisol levels, modulation of hippocampal neurotransmitters, and activation of the ERK signaling pathway.160 Citrus reticulata Blanco pericarpium viride is widely used in TCM, often in combination with other herbal metabolites to treat anxiety and depression. The essential oil derived from Citrus reticulata Blanco pericarpium viride has been shown to regulate olfactory glutamate levels, inhibit neurotoxicity caused by excessive activation of N-methyl-D-aspartate receptors (NMDAR), promote neurogenesis, and alleviate anxiety.161
Other Types
Ruscogenin, a major bioactive steroidal saponin derived from Ophiopogon japonicus (Thunb). Ker Gawl., has been shown to exert anti-inflammatory and anti-anxiety effects.162 In models of chronic pain-induced anxiety disorders, ruscogenin regulates the TLR4-mediated NFκB/mitogen-activated protein kinase (MAPK)/NLRP3 signaling pathway, effectively reducing inflammation and anxiety-like behaviors.163 Betaine, also known as trimethylglycine, is widely distributed across animals, plants, and microorganisms. It plays a crucial role as an osmoprotectant and methyl donor in various physiological processes.164 Betaine’s anti-anxiety effects are attributed to its ability to inhibit DNA damage and mitochondrial dysfunction, which blocks the cGAS/STING pathway, thereby promoting hippocampal neurogenesis and alleviating anxiety symptoms.165 Isochlorogenic acid A, a phenolic acid found in various medicinal plants, is known for its protective effects on organs such as the lungs, liver, and intestines. In the context of anxiety, isochlorogenic acid A exerts its effects through the BDNF/Nuclear factor erythroid-related factor 2 (Nrf2)/Glutathione peroxidase (GPx4) signaling pathways, mitigating lead-induced neuroinflammation, ferroptosis, oxidative stress, and anxiety-like behavior.166 Shikonin, a naphthoquinone metabolite isolated from the Lithospermum erythrorhizon Siebold & Zucc root, is primarily recognized for its potent anti-tumor activity. In anxiety disorders, shikonin reduces the expression of pro-inflammatory cytokines (IL-1β, IL-6, and TNF-α) in the hippocampus, contributing to its anti-anxiety effects167,168 (Figure 5 and Table 1).
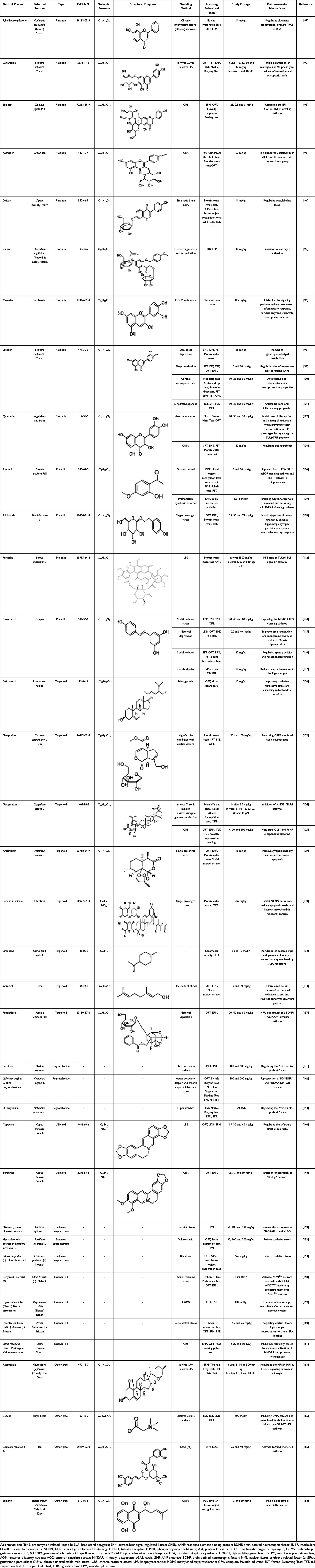 |
Table 1 Natural Product Research Information |
 |
Figure 5 Anxiolytic molecular mechanisms of natural products. Abbreviations: ERK, extracellular signal regulated kinase; CREB, cAMP response element binding protein; BDNF, brain-derived neurotrophic factor; NLRP3, NLR Family Pyrin Domain Containing 3; TLR4, toll-like receptor 4; PI3K, phosphatidylinositol-3-kinase; Akt, protein kinase B; mTOR, mechanistic target of rapamycin; NF-κB, nuclear factor-kappa B; Nrf2, nuclear factor erythroid-related factor 2; GPx4, glutathione peroxidase. |
Limitations and Challenges
Anxiety disorders, being complex mental health conditions, remain incompletely understood in terms of their pathophysiological mechanisms, which presents significant challenges to conventional single-target drug therapies. These challenges often include limited efficacy and undesirable side effects. In this context, natural products offer distinct advantages for anti-anxiety therapy, as they facilitate broader regulation and personalized adaptation, consistent with the principles of precision medicine advocated by contemporary healthcare practices. By constructing a multi-target dynamic regulatory network, a more biologically adaptive treatment system can be developed. Integrating traditional medical wisdom with modern technological advancements provides a promising model for overcoming the deficiencies of synthetic, single-target drugs. This novel research framework holds significant potential for the development of new treatment strategies that optimize efficacy, stability, and individual variability, ultimately paving the way for breakthroughs in the treatment of mental illnesses.
However, current research on the anti-anxiety effects of natural products faces a significant gap in clinical translation. Despite the substantial body of preclinical evidence demonstrating the anti-anxiety potential of these natural products, most findings remain confined to experimental models. This limitation can be attributed to several factors, including heterogeneity in research methodologies, individual differences in clinical samples, and the absence of standardized quality control systems for natural products. More importantly, there is a critical shortage of high-quality, evidence-based studies, particularly those with rigorous randomized controlled trials (RCTs). This gap has led to a lack of sufficient data regarding the efficacy, stability, dose-response relationships, and safety profiles of natural products in clinical settings.
Nevertheless, no pharmacologically active natural compounds are entirely free from non-specific off-target effects on normal tissues.169 In the process of alleviating anxiety, natural products may carry the risk of toxicity or adverse effects. However, existing research still lacks a systematic evaluation of potential adverse reactions and long-term risks associated with natural products, particularly when multiple metabolites are used in combination. This absence of a comprehensive safety evaluation system could significantly limit the clinical translation of natural products and hinder their broader therapeutic application.
In addition, some natural products have inherent physicochemical limitations, including poor chemical stability, low bioavailability, and limited ability to cross the BBB. These factors significantly hinder the targeted delivery of active metabolites to the CNS. Furthermore, natural resources such as Chinese herbal medicines, which are derived from traditional medicinal systems, often face challenges like heterogeneous cultivation conditions, non-standardized processing methods, and batch-to-batch variability in active metabolites. The lack of well-defined quality control standards limits their clinical applicability and complicates the investigation of their pharmacological mechanisms.
Importantly, while classic behavioral paradigms using rodents, such as open-field experiments and elevated plus-maze tests, can quantify anxiety-like behaviors, they are insufficient in capturing the complex psychological dimensions of human-specific anxiety disorders. In addition, species differences between rodents and humans, in terms of neurobiology and the pathological evolution of anxiety, prevent these models from fully replicating the dynamic development of complex psychiatric symptoms. Many experimental models focus on short-term anxiety induced by acute stress but fail to systematically simulate chronic anxiety or comorbid depression. Consequently, the biological validity and clinical relevance of these models remain limited, posing challenges for effectively translating animal-based research into clinical applications.
Conclusion and Future Directions
Future research should prioritize the development of a clinical translation research framework for natural products. This involves systematically advancing multi-center randomized controlled trials and objectively assessing the clinical efficacy of natural products in treating anxiety, utilizing a standardized evaluation system (including anxiety-related cognitive dimension scales and biomarker panels). Additionally, it is crucial to gradually improve a collaborative model that combines natural product therapies with traditional therapies. Adaptive clinical trial designs should be employed to validate the clinical benefits of combination therapies (eg, combined administration of natural products with demonstrated preclinical efficacy and classic anti-anxiety agents such as benzodiazepines). Simultaneously, it is essential to closely monitor both the metabolic status and safety profiles of these treatments in patients, ensuring their long-term efficacy and patient safety.
In addition, further research is needed to explore advanced targeted delivery systems, such as the development of intelligent delivery platforms using nanocarriers and BBB penetration technologies. These systems can enhance the concentration and duration of action of active metabolites in the CNS, ultimately improving therapeutic efficacy. Meanwhile, it is crucial to strengthen the quality control evaluation system for TCM, establishing a standardized quality control framework across the entire TCM production and application chain. This will help enhance the precision and reliability of natural product-based therapies, providing a solid scientific foundation for their clinical translation.
Importantly, in the future, integrating cutting-edge technologies such as single-cell transcriptome sequencing and spatial multi-omics is crucial for systematically elucidating the dynamic molecular mechanisms underlying the pathological progression of anxiety disorders. This approach will facilitate the development of a phased dynamic model of chronic anxiety progression and identify critical time windows for potential optimal interventions using natural products. Additionally, combining human-derived organoid models with targeted gene-editing technologies will allow for the exploration of cross-species mechanisms and the optimization of treatment strategies.
In summary, there is a persistent and urgent need for systematic investigations into the pathological mechanisms of anxiety disorders and the development of targeted interventions in both preclinical and clinical contexts. Such research will not only establish a theoretical basis for novel drug discovery but also facilitate the clinical translation of natural products with anxiolytic potential, thereby shortening the journey from laboratory findings to practical clinical applications. Although natural products have demonstrated considerable promise in treating anxiety disorders, their efficacy and safety require further validation through well-designed human clinical trials.
Data Sharing Statement
No data was used for the research described in the article.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the Qilu Biancang TCM Talent Cultivation Project (Lu Wei Han [2024] No. 78).
Disclosure
All authors declare that there are no conflicts of interest. Figure 1 (https://BioRender.com/3fixn01), Figure 2 (https://BioRender.com/gpdzi3z), Figure 3 (https://BioRender.com/o3rh5z1), Figure 4 (https://BioRender.com/bg6zgux) and Figure 5 (https://BioRender.com/ya4zvk8) were created by BioRender (https://www.biorender.com/).
References
1. Bandelow B, Michaelis S. Epidemiology of anxiety disorders in the 21st century. Dialogues Clin Neurosci. 2015;17(3):327–335. doi:10.31887/DCNS.2015.17.3/bbandelow
2. Kandola A, Vancampfort D, Herring M, et al. Moving to beat anxiety: epidemiology and therapeutic issues with physical activity for anxiety. Curr Psychiatry Rep. 2018;20(8):63. doi:10.1007/s11920-018-0923-x
3. Tyrer P, Baldwin D. Generalised anxiety disorder. Lancet. 2006;368(9553):2156–2166. doi:10.1016/S0140-6736(06)69865-6
4. O’Leary KB, Khan JS. Pharmacotherapy for anxiety disorders. Psychiatr Clin North Am. 2024;47(4):689–709. doi:10.1016/j.psc.2024.04.012
5. Xing P, Zhong Y, Cui X, Liu Z, Wu X. Natural products in digestive tract tumors metabolism: functional and application prospects. Pharmacol Res. 2023;191:106766. doi:10.1016/j.phrs.2023.106766
6. Yuan S, Ma T, Zhang YN, Wang N, Baloch Z, Ma K. Novel drug delivery strategies for antidepressant active ingredients from natural medicinal plants: the state of the art. J Nanobiotechnology. 2023;21(1):391. doi:10.1186/s12951-023-02159-9
7. Guo B, Zhang M, Hao W, Wang Y, Zhang T, Liu C. Neuroinflammation mechanisms of neuromodulation therapies for anxiety and depression. Transl Psychiatry. 2023;13(1):5. doi:10.1038/s41398-022-02297-y
8. Chen R, Routh BN, Straetker JE, et al. Microglia depletion ameliorates neuroinflammation, anxiety-like behavior, and cognitive deficits in a sex-specific manner in Rev-erbα knockout mice. Brain Behav Immun. 2023;114:287–298. doi:10.1016/j.bbi.2023.08.029
9. Gao Z, Lv H, Wang Y, Xie Y, Guan M, Xu Y. TET2 deficiency promotes anxiety and depression-like behaviors by activating NLRP3/IL-1β pathway in microglia of allergic rhinitis mice. Mol Med. 2023;29(1):160. doi:10.1186/s10020-023-00757-9
10. Sidoryk-Węgrzynowicz M, Strużyńska L. Astroglial and microglial purinergic P2X7 receptor as a major contributor to neuroinflammation during the course of multiple sclerosis. Int J Mol Sci. 2021;22(16):8404. doi:10.3390/ijms22168404
11. Li Y, Zhang Y, Lin D, Fu X, Jing C. Demyelination of the amygdala mediates psychological stress-induced emotional disorders partially contributed by activation of P2X7R/NLRP3 cascade. Brain Behav Immun. 2025;124:365–375. doi:10.1016/j.bbi.2024.12.023
12. Chen LQ, Lv XJ, Guo QH, et al. Asymmetric activation of microglia in the hippocampus drives anxiodepressive consequences of trigeminal neuralgia in rodents. Br J Pharmacol. 2023;180(8):1090–1113. doi:10.1111/bph.15994
13. Shan S, Cheng D, Li H, et al. Short-term PS-NP exposure in early adulthood induces neuronal damage in middle-aged mice via microglia-mediated neuroinflammation. J Hazard Mater. 2025;489:137615. doi:10.1016/j.jhazmat.2025.137615
14. Pan K, Gao Y, Zong H, et al. Neuronal CCL2 responds to hyperglycaemia and contributes to anxiety disorders in the context of diabetes. Nat Metab. 2025. doi:10.1038/s42255-025-01281-2
15. Rinaldi A, Balietti M, Principi E, et al. BV2-derived extracellular vesicles modulate microglia inflammatory profile, neuronal plasticity, and behavioural performances in late adult mice. Brain Behav Immun. 2024;122:58–74. doi:10.1016/j.bbi.2024.08.012
16. Yuan H, Xu Y, Jiang H, et al. Acid sphingomyelinase modulates anxiety-like behavior likely through toll-like receptor signaling pathway. Mol Brain. 2025;18(1):8. doi:10.1186/s13041-025-01178-x
17. Robb JL, Boisjoly F, Machuca-Parra AI, et al. Blockage of ATGL-mediated breakdown of lipid droplets in microglia alleviates neuroinflammatory and behavioural responses to lipopolysaccharides. Brain Behav Immun. 2025;123:315–333. doi:10.1016/j.bbi.2024.09.027
18. Lee B, Kwon JT, Jeong Y, et al. Inflammatory and anti-inflammatory cytokines bidirectionally modulate amygdala circuits regulating anxiety. Cell. 2025;188(8):2190–2202.e15. doi:10.1016/j.cell.2025.03.005
19. Magee JC, Grienberger C. Synaptic plasticity forms and functions. Annu Rev Neurosci. 2020;43:95–117. doi:10.1146/annurev-neuro-090919-022842
20. Sasibhushana RB, Shankaranarayana Rao BS, Srikumar BN. Anxiety-, and depression-like behavior following short-term finasteride administration is associated with impaired synaptic plasticity and cognitive behavior in male rats. J Psychiatr Res. 2024;174:304–318. doi:10.1016/j.jpsychires.2024.04.029
21. Li S, Xu W, Gong L, et al. Subchronic nonylphenol exposure induced anxiety-like behavior and decreased expressions of regulators of synaptic plasticity in rats. Chemosphere. 2021;282:130994. doi:10.1016/j.chemosphere.2021.130994
22. Fu Q, Qiu R, Yao T, et al. Music therapy as a preventive intervention for postpartum depression: modulation of synaptic plasticity, oxidative stress, and inflammation in a mouse model. Transl Psychiatry. 2025;15(1):143. doi:10.1038/s41398-025-03370-y
23. Han QQ, Shen SY, Liang LF, Chen XR, Yu J. Complement C1q/C3-CR3 signaling pathway mediates abnormal microglial phagocytosis of synapses in a mouse model of depression. Brain Behav Immun. 2024;119:454–464. doi:10.1016/j.bbi.2024.04.018
24. Wojtas MN, Diaz-González M, Stavtseva N, et al. Interplay between hippocampal TACR3 and systemic testosterone in regulating anxiety-associated synaptic plasticity. Mol Psychiatry. 2024;29(3):686–703. doi:10.1038/s41380-023-02361-z
25. Khaled H, Ghasemi Z, Inagaki M, et al. The TrkC-PTPσ complex governs synapse maturation and anxiogenic avoidance via synaptic protein phosphorylation. EMBO J. 2024;43(22):5690–5717. doi:10.1038/s44318-024-00252-9
26. Hu J, Ji WJ, Liu GY, et al. IDO1 modulates pain sensitivity and comorbid anxiety in chronic migraine through microglial activation and synaptic pruning. J Neuroinflammation. 2025;22(1):42. doi:10.1186/s12974-025-03367-w
27. Tufail T, Ain HBU, Virk MS, et al. GABA (γ-aminobutyric acid) enrichment and detection methods in cereals: Unlocking sustainable health benefits. Food Chem. 2025;464(Pt 3):141750. doi:10.1016/j.foodchem.2024.141750
28. Hu L, Qiu MJ, Fan WJ, et al. Characterization of GABAergic marker expression in prefrontal cortex in dexamethasone induced depression/anxiety model. Front Endocrinol. 2024;15:1433026. doi:10.3389/fendo.2024.1433026
29. Griego E, Cerna C, Sollozo-DuPont I, Fuenzalida M, Galván EJ. Maternal immune activation alters temporal Precision of spike generation of CA1 pyramidal neurons by unbalancing GABAergic inhibition in the Offspring. Brain Behav Immun. 2025;123:211–228. doi:10.1016/j.bbi.2024.09.012
30. Pérez-Garci E, Pysanenko K, Rizzi G, et al. Binding of HCN channels to GABAB receptors in dopamine neurons of the VTA limits synaptic inhibition and prevents the development of anxiety. Neurobiol Dis. 2025;206:106831. doi:10.1016/j.nbd.2025.106831
31. Pandey S, Han W, Li J, et al. Reversing anxiety by targeting a stress-responsive signaling pathway. Proc Natl Acad Sci U S A. 2024;121(31):e2400078121. doi:10.1073/pnas.2400078121
32. Fang S, Wu Z, Guo Y, et al. Roles of microglia in adult hippocampal neurogenesis in depression and their therapeutics. Front Immunol. 2023;14:1193053. doi:10.3389/fimmu.2023.1193053
33. Wu KY, Tsao CH, Su NC, Deng SM, Huang GJ. Stk24 deficiency causes disrupted hippocampal neurogenesis and anxiety-like behavior in mice. Commun Biol. 2025;8(1):663. doi:10.1038/s42003-025-08035-6
34. Takahashi K, Kurokawa K, Miyagawa K, Mochida-Saito A, Takeda H, Tsuji M. Repeated exposure to antibiotics exhibits anxiety-like behaviors with a reduction in neurogenesis in the ventral hippocampus of dentate gyrus. Neurosci Lett. 2025;849:138131. doi:10.1016/j.neulet.2025.138131
35. Bedrosian TA, Houtman J, Eguiguren JS, et al. Lamin B1 decline underlies age-related loss of adult hippocampal neurogenesis. EMBO J. 2021;40(3):e105819. doi:10.15252/embj.2020105819
36. Qi X, Xu Z, Liu X, et al. Near-infrared light induces neurogenesis and modulates anxiety-like behavior. Stem Cell Res Ther. 2024;15(1):494. doi:10.1186/s13287-024-04114-3
37. Aggarwal A, Liu R, Chen Y, et al. Glutamate indicators with improved activation kinetics and localization for imaging synaptic transmission. Nat Methods. 2023;20(6):925–934. doi:10.1038/s41592-023-01863-6
38. Verma M, Lizama BN, Chu CT. Excitotoxicity, calcium and mitochondria: a triad in synaptic neurodegeneration. Transl Neurodegener. 2022;11(1):3. doi:10.1186/s40035-021-00278-7
39. Tsotsokou G, Nikolakopoulou M, Kouvelas ED, Mitsacos A. Neonatal maternal separation affects metabotropic glutamate receptor 5 expression and anxiety-related behavior of adult rats. Eur J Neurosci. 2021;54(2):4550–4564. doi:10.1111/ejn.15358
40. Wang JM, Wang YM, Zhu YB, et al. Peripheral inflammation triggering central anxiety through the hippocampal glutamate metabolized receptor 1. CNS Neurosci Ther. 2024;30(4):e14723. doi:10.1111/cns.14723
41. Liu WZ, Zhang WH, Zheng ZH, et al. Identification of a prefrontal cortex-to-amygdala pathway for chronic stress-induced anxiety. Nat Commun. 2020;11(1):2221. doi:10.1038/s41467-020-15920-7
42. Jones SK, McCarthy DM, Vied C, Stanwood GD, Schatschneider C, Bhide PG. Transgenerational transmission of aspartame-induced anxiety and changes in glutamate-GABA signaling and gene expression in the amygdala. Proc Natl Acad Sci U S A. 2022;119(49):e2213120119. doi:10.1073/pnas.2213120119
43. Frank D, Gruenbaum BF, Shelef I, et al. Blood glutamate scavenging as a novel glutamate-based therapeutic approach for post-traumatic brain injury anxiety and social impairment. Transl Psychiatry. 2023;13(1):41. doi:10.1038/s41398-023-02329-1
44. DeMorrow S. Role of the hypothalamic-pituitary-adrenal axis in health and disease. Int J Mol Sci. 2018;19(4):986. doi:10.3390/ijms19040986
45. Balthazar L, Lages YVM, Romano VC, Landeira-Fernandez J, Krahe TE. The association between the renin-angiotensin system and the hypothalamic-pituitary-adrenal axis in anxiety disorders: a systematic review of animal studies. Psychoneuroendocrinology. 2021;132:105354. doi:10.1016/j.psyneuen.2021.105354
46. Belo-Silva AE, de Gusmão Taveiros Silva NK, Marianno P, et al. Effects of the combination of chronic unpredictable stress and environmental enrichment on anxiety-like behavior assessed using the elevated plus maze in Swiss male mice: Hypothalamus-Pituitary-Adrenal Axis-mediated mechanisms. Horm Behav. 2024;162:105538. doi:10.1016/j.yhbeh.2024.105538
47. Zhu J, Wang C, Wang Y, et al. Electroacupuncture alleviates anxiety and modulates amygdala CRH/CRHR1 signaling in single prolonged stress mice. Acupunct Med. 2022;40(4):369–378. doi:10.1177/09645284211056352
48. Tanaka T, Shimizu S, Ueno M, Fujihara Y, Ikawa M, Miyata S. MARCKSL1 regulates spine formation in the amygdala and controls the hypothalamic-pituitary-adrenal axis and anxiety-like behaviors. EBioMedicine. 2018;30:62–73. doi:10.1016/j.ebiom.2018.03.018
49. Zhang N, Zhao S, Ma Y, et al. Hyperexcitation of ovBNST CRF neurons during stress contributes to female-biased expression of anxiety-like avoidance behaviors. Sci Adv. 2024;10(19):eadk7636. doi:10.1126/sciadv.adk7636
50. Qiu G, Wang P, Rao J, et al. Dexmedetomidine inhibits paraventricular corticotropin-releasing hormone neurons that attenuate acute stress-induced anxiety-like behavior in mice. Anesthesiology. 2024;140(6):1134–1152. doi:10.1097/ALN.0000000000004982
51. Loh JS, Mak WQ, Tan LKS, et al. Microbiota-gut-brain axis and its therapeutic applications in neurodegenerative diseases. Signal Transduct Target Ther. 2024;9(1):37. doi:10.1038/s41392-024-01743-1
52. Cao Y, Cheng Y, Pan W, Diao J, Sun L, Meng M. Gut microbiota variations in depression and anxiety: a systematic review. BMC Psychiatry. 2025;25(1):443. doi:10.1186/s12888-025-06871-8
53. Joo MK, Lee JW, Woo JH, Kim HJ, Kim DH, Choi JH. Regulation of colonic neuropeptide Y expression by the gut microbiome in patients with ulcerative colitis and its association with anxiety- and depression-like behavior in mice. Gut Microbes. 2024;16(1):2319844. doi:10.1080/19490976.2024.2319844
54. Yuan X, Chen B, Duan Z, et al. Depression and anxiety in patients with active ulcerative colitis: crosstalk of gut microbiota, metabolomics and proteomics. Gut Microbes. 2021;13(1):1987779. doi:10.1080/19490976.2021.1987779
55. Zhu J, Zhong Z, Shi L, et al. Gut microbiota mediate early life stress-induced social dysfunction and anxiety-like behaviors by impairing amino acid transport at the gut. Gut Microbes. 2024;16(1):2401939. doi:10.1080/19490976.2024.2401939
56. Zhou GQ, Wang X, Gao P, et al. Intestinal microbiota via NLRP3 inflammasome dependent neuronal pyroptosis mediates anxiety-like behaviour in mice exposed to 3.5 GHz radiofrequency radiation. Sci Total Environ. 2024;927:172391. doi:10.1016/j.scitotenv.2024.172391
57. Xia M, Lu J, Lan J, et al. Elevated IL-22 as a result of stress-induced gut leakage suppresses septal neuron activation to ameliorate anxiety-like behavior. Immunity. 2025;58(1):218–231.e12. doi:10.1016/j.immuni.2024.11.008
58. Pan M, Qian C, Huo S, et al. Gut-derived lactic acid enhances tryptophan to 5-hydroxytryptamine in regulation of anxiety via Akkermansia muciniphila. Gut Microbes. 2025;17(1):2447834. doi:10.1080/19490976.2024.2447834
59. Lee YB, Cho YJ, Kim JK. The unique role of fluoxetine in alleviating depression and anxiety by regulating gut microbiota and the expression of vagus nerve-mediated serotonin and melanocortin-4 receptors. Biomed Pharmacother. 2025;182:117748. doi:10.1016/j.biopha.2024.117748
60. Liu BH, Xu CZ, Liu Y, et al. Mitochondrial quality control in human health and disease. Mil Med Res. 2024;11(1):32. doi:10.1186/s40779-024-00536-5
61. Filiou MD, Sandi C. Anxiety and brain mitochondria: a bidirectional crosstalk. Trends Neurosci. 2019;42(9):573–588. doi:10.1016/j.tins.2019.07.002
62. Zhang H, Chen Y, Zhang J, et al. Assessing the joint effects of mitochondrial function and human behavior on the risks of anxiety and depression. J Affect Disord. 2023;320:561–567. doi:10.1016/j.jad.2022.09.157
63. Zhao W, Zhao S, Wei R, et al. cGAS/STING signaling pathway-mediated microglial activation in the PFC underlies chronic ethanol exposure-induced anxiety-like behaviors in mice. Int Immunopharmacol. 2024;134:112185. doi:10.1016/j.intimp.2024.112185
64. Gebara E, Zanoletti O, Ghosal S, et al. Mitofusin-2 in the nucleus accumbens regulates anxiety and depression-like behaviors through mitochondrial and neuronal actions. Biol Psychiatry. 2021;89(11):1033–1044. doi:10.1016/j.biopsych.2020.12.003
65. Ghosal S, Gebara E, Ramos-Fernández E, et al. Mitofusin-2 in nucleus accumbens D2-MSNs regulates social dominance and neuronal function. Cell Rep. 2023;42(7):112776. doi:10.1016/j.celrep.2023.112776
66. Ma Y, Xu D, Wan Z, et al. Exposure to different surface-modified polystyrene nanoparticles caused anxiety, depression, and social deficit in mice via damaging mitochondria in neurons. Sci Total Environ. 2024;919:170739. doi:10.1016/j.scitotenv.2024.170739
67. Thomou C, Nussbaumer M, Grammenou E, et al. Early handling exerts anxiolytic effects and alters brain mitochondrial dynamics in adult high anxiety mice. Mol Neurobiol. 2024;61(12):10593–10612. doi:10.1007/s12035-024-04116-5
68. Wang Y, Zhang R, Jiang Y, Liao J, Mu L, Hu M. Hippocampal SIRT1 signaling mediates the ameliorative effect of treadmill exercise on anxiety- and depression-like behavior in APP/PS1 mice. Front Aging Neurosci. 2024;16:1489214. doi:10.3389/fnagi.2024.1489214
69. Freeman MR, Rowitch DH. Evolving concepts of gliogenesis: a look way back and ahead to the next 25 years. Neuron. 2013;80(3):613–623. doi:10.1016/j.neuron.2013.10.034
70. Liu Y, Shen X, Zhang Y, et al. Interactions of glial cells with neuronal synapses, from astrocytes to microglia and oligodendrocyte lineage cells. Glia. 2023;71(6):1383–1401. doi:10.1002/glia.24343
71. Ogrodnik M, Zhu Y, Langhi LGP, et al. Obesity-induced cellular senescence drives anxiety and impairs neurogenesis [published correction appears in Cell Metab. 2019;29(5):1233. doi: 10.1016/j.cmet.2019.01.013]. Cell Metab. 2019;29(5):1061–1077.e8. doi:10.1016/j.cmet.2018.12.008
72. Zuo Y, Hou Y, Wang Y, Yuan L, Cheng L, Zhang T. Circadian misalignment impairs oligodendrocyte myelination via Bmal1 overexpression leading to anxiety and depression-like behaviors. J Pineal Res. 2024;76(1):e12935. doi:10.1111/jpi.12935
73. Du Preez A, Onorato D, Eiben I, et al. Chronic stress followed by social isolation promotes depressive-like behaviour, alters microglial and astrocyte biology and reduces hippocampal neurogenesis in male mice. Brain Behav Immun. 2021;91:24–47. doi:10.1016/j.bbi.2020.07.015
74. Meng Z, Guo S, Dong X, et al. Astrocyte-Ablation of Mtnr1b increases anxiety-like behavior in adult male mice. J Integr Neurosci. 2023;22(6):154. doi:10.31083/j.jin2206154
75. Huang Q, Lee HH, Volpe B, et al. Deletion of murine astrocytic vesicular nucleotide transporter increases anxiety and depressive-like behavior and attenuates motivation for reward. Mol Psychiatry. 2025;30(2):506–520. doi:10.1038/s41380-024-02692-5
76. Zheng Z, Zhou H, Yang L, Zhang L, Guo M. Selective disruption of mTORC1 and mTORC2 in VTA astrocytes induces depression and anxiety-like behaviors in mice. Behav Brain Res. 2024;463:114888. doi:10.1016/j.bbr.2024.114888
77. Cho WH, Noh K, Lee BH, et al. Hippocampal astrocytes modulate anxiety-like behavior. Nat Commun. 2022;13(1):6536. doi:10.1038/s41467-022-34201-z
78. Soto JS, Neupane C, Kaur M, Pandey V, Wohlschlegel JA, Khakh BS. Astrocyte Gi-GPCR signaling corrects compulsive-like grooming and anxiety-related behaviors in Sapap3 knockout mice. Neuron. 2024;112(20):3412–3423.e6. doi:10.1016/j.neuron.2024.07.019
79. Dudek KA, Paton SEJ, Binder LB, et al. Astrocytic cannabinoid receptor 1 promotes resilience by dampening stress-induced blood-brain barrier alterations. Nat Neurosci. 2025;28(4):766–782. doi:10.1038/s41593-025-01891-9
80. Chen D, Lou Q, Song XJ, et al. Microglia govern the extinction of acute stress-induced anxiety-like behaviors in male mice. Nat Commun. 2024;15(1):449. doi:10.1038/s41467-024-44704-6
81. Mizushima N, Komatsu M. Autophagy: renovation of cells and tissues. Cell. 2011;147(4):728–741. doi:10.1016/j.cell.2011.10.026
82. Demianchuk O, Bayliak M, Vatashchuk M, et al. Alpha-ketoglutarate promotes anxiety, activates autophagy, and suppresses antioxidant enzymes in the cerebral cortex of female mice on cafeteria diet. Brain Res Bull. 2025;222:111255. doi:10.1016/j.brainresbull.2025.111255
83. Xiao X, Shang X, Zhai B, Zhang H, Zhang T. Nicotine alleviates chronic stress-induced anxiety and depressive-like behavior and hippocampal neuropathology via regulating autophagy signaling. Neurochem Int. 2018;114:58–70. doi:10.1016/j.neuint.2018.01.004
84. Naeem A, Prakash R, Kumari N, et al. MCC950 reduces autophagy and improves cognitive function by inhibiting NLRP3-dependent neuroinflammation in a rat model of Alzheimer’s disease. Brain Behav Immun. 2024;116:70–84. doi:10.1016/j.bbi.2023.11.031
85. Meng H, Cao Y, Qin J, et al. DNA methylation, its mediators and genome integrity. Int J Biol Sci. 2015;11(5):604–617. doi:10.7150/ijbs.11218
86. Tian L, Li XH, Zhao YL, et al. DNMT3a Downregulation Ttriggered Upregulation of GABAA receptor in the mPFC promotes paclitaxel-induced pain and anxiety in male mice. Adv Sci. 2025;12(5):e2407387. doi:10.1002/advs.202407387
87. Wang J, Zhang W, Xu H, et al. The changes of histone methylation induced by adolescent social stress regulate the resting-state activity in mPFC. Research. 2023;6:0264. doi:10.34133/research.0264
88. Sun Q, Liu Q, Zhou X, et al. Flavonoids regulate tumor-associated macrophages - from structure-activity relationship to clinical potential (Review). Pharmacol Res. 2022;184:106419. doi:10.1016/j.phrs.2022.106419
89. Wang N, Liu X, Li XT, et al. 7,8-dihydroxyflavone alleviates anxiety-like behavior induced by chronic alcohol exposure in mice involving tropomyosin-related kinase B in the Amygdala. Mol Neurobiol. 2021;58(1):92–105. doi:10.1007/s12035-020-02111-0
90. Zhou Y, Huang Y, Ye W, Chen Z, Yuan Z. Cynaroside improved depressive-like behavior in CUMS mice by suppressing microglial inflammation and ferroptosis. Biomed Pharmacother. 2024;173:116425. doi:10.1016/j.biopha.2024.116425
91. Guo Y, Wei F, Lv Y, et al. Spinosin improves anxiety disorders in mice with chronic restraint stress via the ERK1/2-CREB-BDNF pathway. Int Immunopharmacol. 2024;143(Pt 2):113517. doi:10.1016/j.intimp.2024.113517
92. Chen J, Zhong K, Qin S, et al. Astragalin: a food-origin flavonoid with therapeutic effect for multiple diseases. Front Pharmacol. 2023;14:1265960. doi:10.3389/fphar.2023.1265960
93. Lin J, Zhang W, Wang S, et al. Astragalin inhibits neuronal excitability and activates neuronal autophagy in the ACC and LH of CFA mice to alleviate inflammatory pain and pain-related emotions. Int Immunopharmacol. 2025;148:114115. doi:10.1016/j.intimp.2025.114115
94. Zahoor M, Farhat SM, Khan S, Ahmed T. Daidzin improves neurobehavioral outcome in rat model of traumatic brain injury. Behav Brain Res. 2024;472:115158. doi:10.1016/j.bbr.2024.115158
95. Zhang DX, Jia SY, Xiao K, et al. Icariin mitigates anxiety-like behaviors induced by hemorrhagic shock and resuscitation via inhibiting of astrocytic activation. Phytomedicine. 2024;128:155507. doi:10.1016/j.phymed.2024.155507
96. Inan S, Meissler JJ, Shekarabi A, et al. Cyanidin prevents MDPV withdrawal-induced anxiety-like effects and dysregulation of cytokine systems in rats. Brain Res. 2023;1806:148310. doi:10.1016/j.brainres.2023.148310
97. Kou JJ, Shi JZ, He YY, et al. Luteolin alleviates cognitive impairment in Alzheimer’s disease mouse model via inhibiting endoplasmic reticulum stress-dependent neuroinflammation. Acta Pharmacol Sin. 2022;43(4):840–849. doi:10.1038/s41401-021-00702-8
98. Wu X, Xu H, Zeng N, et al. Luteolin alleviates depression-like behavior by modulating glycerophospholipid metabolism in the hippocampus and prefrontal cortex of LOD rats. CNS Neurosci Ther. 2024;30(3):e14455. doi:10.1111/cns.14455
99. Xiong F, Lv X. Luteolin reversed anxiety and depressive-like behavior via modulation of the NF-κB/NLRP3 inflammasome axis in the hippocampus of rats subjected to sleep deprivation. Iran J Basic Med Sci. 2024;27(8):1050–1058. doi:10.22038/IJBMS.2024.75068.16277
100. Mokhtari T, Lu M, El-Kenawy AE. Potential anxiolytic and antidepressant-like effects of luteolin in a chronic constriction injury rat model of neuropathic pain: role of oxidative stress, neurotrophins, and inflammatory factors. Int Immunopharmacol. 2023;122:110520. doi:10.1016/j.intimp.2023.110520
101. She R, Zhang Z, Han M, et al. Luteolin exhibits anxiolytic and antidepressant potential in Parkinson’s disease rat: antioxidant and anti-inflammatory effects. Rejuvenation Res. 2025;28(2):67–82. doi:10.1089/rej.2024.0045
102. Wang N, Li F, Du J, et al. Quercetin protects against global cerebral ischemia‒reperfusion injury by inhibiting microglial activation and polarization. J Inflamm Res. 2024;17:1281–1293. doi:10.2147/JIR.S448620
103. Li B, Yan Y, Zhang T, et al. Quercetin reshapes gut microbiota homeostasis and modulates brain metabolic profile to regulate depression-like behaviors induced by CUMS in rats. Front Pharmacol. 2024;15:1362464. doi:10.3389/fphar.2024.1362464
104. Zou H, Ye H, Kamaraj R, Zhang T, Zhang J, Pavek P. A review on pharmacological activities and synergistic effect of quercetin with small molecule agents. Phytomedicine. 2021;92:153736. doi:10.1016/j.phymed.2021.153736
105. Wu R, Liu Y, Zhang F, et al. Protective mechanism of Paeonol on central nervous system. Phytother Res. 2024;38(2):470–488. doi:10.1002/ptr.8049
106. Kang WC, Lee YS, Park K, et al. Paeonol alleviates postmenopause-induced neuropsychiatric symptoms through the modulation of GPR30 in ovariectomized mice. J Ethnopharmacol. 2024;327:118063. doi:10.1016/j.jep.2024.118063
107. Wei E, Gao A, Mu X, et al. Paeonol ameliorates hippocampal neuronal damage by inhibiting GRM5/GABBR2/β-arrestin2 and activating the cAMP-PKA signaling pathway in premenstrual irritability rats. Brain Res Bull. 2023;205:110830. doi:10.1016/j.brainresbull.2023.110830
108. Yang S, Wang L, Zeng Y, et al. Salidroside alleviates cognitive impairment by inhibiting ferroptosis via activation of the Nrf2/GPX4 axis in SAMP8 mice. Phytomedicine. 2023;114:154762. doi:10.1016/j.phymed.2023.154762
109. Chen XD, Wei JX, Wang HY, et al. Effects and mechanisms of salidroside on the behavior of SPS-induced PTSD rats. Neuropharmacology. 2023;240:109728. doi:10.1016/j.neuropharm.2023.109728
110. Nanavati K, Rutherfurd-Markwick K, Lee SJ, Bishop NC, Ali A. Effect of curcumin supplementation on exercise-induced muscle damage: a narrative review. Eur J Nutr. 2022;61(8):3835–3855. doi:10.1007/s00394-022-02943-7
111. Chen L, Hua B, He Q, et al. Curcumin analogue NL04 inhibits spinal cord central sensitization in rats with bone cancer pain by inhibiting NLRP3 inflammasome activation and reducing IL-1β production. Eur J Pharmacol. 2024;970:176480. doi:10.1016/j.ejphar.2024.176480
112. Chen P, Guo Z, Lei J, Wang Y. Pomegranate polyphenol punicalin ameliorates lipopolysaccharide-induced memory impairment, behavioral disorders, oxidative stress, and neuroinflammation via inhibition of TLR4-NF-кB pathway. Phytother Res. 2024;38(7):3489–3508. doi:10.1002/ptr.8219
113. Wu SX, Xiong RG, Huang SY, et al. Effects and mechanisms of resveratrol for prevention and management of cancers: An updated review. Crit Rev Food Sci Nutr. 2023;63(33):12422–12440. doi:10.1080/10408398.2022.2101428
114. Zarebavani M, Baghaei Naeini F, Farahvash A, Moradi F, Dashti N. Resveratrol attenuates chronic social isolation stress-induced affective disorders: involvement of NF-κB/NLRP3 axis. J Biochem Mol Toxicol. 2023;37(5):e23311. doi:10.1002/jbt.23311
115. Shukla P, Akotkar L, Aswar U. Resveratrol attenuates early life stress induced depression in rats: behavioural and neurochemical evidence. Neurosci Lett. 2024;820:137606. doi:10.1016/j.neulet.2023.137606
116. Zhao J, Ye L, Liu Z, et al. Protective effects of resveratrol on adolescent social isolation-induced anxiety-like behaviors via modulating Nucleus accumbens spine plasticity and mitochondrial function in female rats. Nutrients. 2022;14(21):4542. doi:10.3390/nu14214542
117. Calado CMSDS, Manhães-de-Castro R, da Conceição Pereira S, et al. Resveratrol reduces neuroinflammation and hippocampal microglia activation and protects against impairment of memory and anxiety-like behavior in experimental cerebral palsy. Mol Neurobiol. 2024;61(6):3619–3640. doi:10.1007/s12035-023-03772-3
118. Shaito A, Posadino AM, Younes N, et al. Potential adverse effects of resveratrol: a literature review. Int J Mol Sci. 2020;21(6):2084. doi:10.3390/ijms21062084
119. Wang H, Wang Z, Zhang Z, Liu J, Hong L. β-Sitosterol as a promising anticancer agent for chemoprevention and chemotherapy: mechanisms of action and future prospects. Adv Nutr. 2023;14(5):1085–1110. doi:10.1016/j.advnut.2023.05.013
120. Vafaei A, Vafaeian A, Iranmehr A, Nassireslami E, Hasannezhad B, Hosseini Y. Effects of β-sitosterol on anxiety in migraine-induced rats: the role of oxidative/nitrosative stress and mitochondrial function. CNS Neurosci Ther. 2024;30(9):e14892. doi:10.1111/cns.14892
121. Zhu D, Ni Y, Chen C, Dong Z, Wang L, Zhang W. Geniposide ameliorates diabetic nephropathy in type 2 diabetic mice by targeting AGEs-RAGE-dependent inflammatory pathway. Phytomedicine. 2024;135:156046. doi:10.1016/j.phymed.2024.156046
122. Sun B, Jia X, Yang F, Ren G, Wu X. CREB-mediated generation and neuronal growth regulates the behavioral improvement of geniposide in diabetes-associated depression mouse model. Neurosci Res. 2021;165:38–44. doi:10.1016/j.neures.2020.05.003
123. Mou Y, Liao W, Li Y, et al. Glycyrrhizin and the related preparations: an inspiring resource for the treatment of liver diseases. Am J Chin Med. 2024;52(2):315–354. doi:10.1142/S0192415X24500149
124. Lou J, Ding B, Fang M, et al. Glycyrrhizin attenuates white matter injury by inhibiting neuroinflammation through the HMGB1/TLR4 pathway. Mol Neurobiol. 2025;62(5):6070–6087. doi:10.1007/s12035-024-04657-9
125. Ma S, Chong Y, Zhang R, et al. Glycyrrhizic acid treatment ameliorates anxiety-like behaviour via GLT1 and Per1/2-dependent pathways. J Ethnopharmacol. 2024;328:118013. doi:10.1016/j.jep.2024.118013
126. El Midaoui A, Ghzaiel I, Vervandier-Fasseur D, et al. Saffron (Crocus sativus L.): a source of nutrients for health and for the treatment of neuropsychiatric and age-related diseases. Nutrients. 2022;14(3):597. doi:10.3390/nu14030597
127. Mohammadi S, Naseri M, Faridi N, et al. Saffron carotenoids reversed the UCMS-induced depression and anxiety in rats: behavioral and biochemical parameters, and hippocampal BDNF/ERK/CREB and NR2B signaling markers. Phytomedicine. 2023;119:154989. doi:10.1016/j.phymed.2023.154989
128. Lan Y, Wang X, Yan F, et al. Quinoa saponin ameliorates lipopolysaccharide-induced behavioral disorders in mice by inhibiting neuroinflammation, modulating gut microbiota, and counterbalancing intestinal inflammation. J Agric Food Chem. 2025;73(8):4700–4715. doi:10.1021/acs.jafc.5c00296
129. Liu Q, Ding X, Wang Y, et al. Artemisinin reduces PTSD-like symptoms, improves synaptic plasticity, and inhibits apoptosis in rats subjected to single prolonged stress. Front Pharmacol. 2024;15:1303123. doi:10.3389/fphar.2024.1303123
130. Mei T, Ma L, Kong F. Sodium aescinate improve behavioral performance by inhibiting dorsal raphe nucleus NLRP3 inflammasome in Post-traumatic stress disorder Rat Model. Biochem Biophys Res Commun. 2023;671:166–172. doi:10.1016/j.bbrc.2023.06.004
131. Araújo-Filho HG, Dos Santos JF, Carvalho MTB, et al. Anticancer activity of limonene: a systematic review of target signaling pathways. Phytother Res. 2021;35(9):4957–4970. doi:10.1002/ptr.7125
132. Song Y, Seo S, Lamichhane S, et al. Limonene has anti-anxiety activity via adenosine A2A receptor-mediated regulation of dopaminergic and GABAergic neuronal function in the striatum. Phytomedicine. 2021;83:153474. doi:10.1016/j.phymed.2021.153474
133. Yang YY, Du LX, Zhu JY, et al. Antipruritic effects of geraniol on acute and chronic itch via modulating spinal GABA/GRPR signaling. Phytomedicine. 2023;119:154969. doi:10.1016/j.phymed.2023.154969
134. Nisar R, Inamullah A, Ghalib AUF, et al. Geraniol mitigates anxiety-like behaviors in rats by reducing oxidative stress, repairing impaired hippocampal neurotransmission, and normalizing brain cortical-EEG wave patterns after a single electric foot-shock exposure. Biomed Pharmacother. 2024;176:116771. doi:10.1016/j.biopha.2024.116771
135. Venzon L, Meurer MC, Dos Santos França TC, et al. Geraniol accelerates the gastric healing, minimizes ulcers recurrence, and reduces anxiolytic-like behavior in ulcerated rodents by oral or inhaled route. Inflammopharmacology. 2022;30(6):2331–2344. doi:10.1007/s10787-022-01068-x
136. Wang XL, Wang YT, Guo ZY, et al. Efficacy of paeoniflorin on models of depression: a systematic review and meta-analysis of rodent studies. J Ethnopharmacol. 2022;290:115067. doi:10.1016/j.jep.2022.115067
137. Liang R, Ge W, Song X, et al. Paeoniflorin alleviates anxiety and visceral hypersensitivity via HPA axis and BDNF/TrkB/PLCγ1 pathway in maternal separation-induced IBS-like rats. Curr Mol Pharmacol. 2024. doi:10.2174/0118761429280572240311060851
138. Ou X, Yu Z, Pan C, et al. Paeoniflorin: a review of its pharmacology, pharmacokinetics and toxicity in diabetes. Front Pharmacol. 2025;16:1551368. doi:10.3389/fphar.2025.1551368
139. Yu Y, Shen M, Song Q, Xie J. Biological activities and pharmaceutical applications of polysaccharide from natural resources: a review. Carbohydr Polym. 2018;183:91–101. doi:10.1016/j.carbpol.2017.12.009
140. Huerta MÁ, Tejada MÁ, Nieto FR. Fucoidan as a promising drug for pain treatment: systematic review and meta-analysis. Mar Drugs. 2024;22(7):290. doi:10.3390/md22070290
141. Li S, Qian Q, Xie Y, et al. Ameliorated effects of Fucoidan on dextran sulfate sodium-induced ulcerative colitis and accompanying anxiety and depressive behaviors in aged C57BL/6 mice. J Agric Food Chem. 2024;72(25):14199–14215. doi:10.1021/acs.jafc.4c03039
142. Luo Y, Bei X, Zhang Y, et al. Cichorium intybus L. Oligo-Polysaccharides (CIO) exerts antianxiety and antidepressant effects on mice experiencing behavioral despair and chronic unpredicted mild stress. Foods. 2025;14(1):135. doi:10.3390/foods14010135
143. Qin YQ, Wang LY, Yang XY, et al. Inulin: properties and health benefits. Food Funct. 2023;14(7):2948–2968. doi:10.1039/d2fo01096h
144. Zou H, Gao H, Liu Y, et al. Dietary inulin alleviated constipation induced depression and anxiety-like behaviors: involvement of gut microbiota and microbial metabolite short-chain fatty acid. Int J Biol Macromol. 2024;259(Pt 2):129420. doi:10.1016/j.ijbiomac.2024.129420
145. Lu Q, Tang Y, Luo S, Gong Q, Li C. Coptisine, the characteristic Constituent from Coptis chinensis, exhibits significant therapeutic potential in treating cancers, metabolic and inflammatory diseases. Am J Chin Med. 2023;51(8):2121–2156. doi:10.1142/S0192415X2350091X
146. Qi Y, Zhao X, Wu W, et al. Coptisine improves LPS-induced anxiety-like behaviors by regulating the Warburg effect in microglia via PKM2. Biomed Pharmacother. 2025;183:117837. doi:10.1016/j.biopha.2025.117837
147. Wang K, Yin J, Chen J, Ma J, Si H, Xia D. Inhibition of inflammation by berberine: molecular mechanism and network pharmacology analysis. Phytomedicine. 2024;128:155258. doi:10.1016/j.phymed.2023.155258
148. He M, Chen YX, Feng PP, et al. Berberine alleviates chronic pain-induced anxiety-like behaviors by inhibiting the activation of VLT-projecting cACC (Cg2) neurons. Commun Biol. 2024;7(1):1651. doi:10.1038/s42003-024-07372-2
149. Imenshahidi M, Hosseinzadeh H. Berberine and barberry (Berberis vulgaris): a clinical review. Phytother Res. 2019;33(3):504–523. doi:10.1002/ptr.6252
150. Kim YR, Lee SY, Lee SM, Shim I, Lee MY. Effect of Hibiscus syriacus Linnaeus extract and its active constituent, saponarin, in animal models of stress-induced sleep disturbances and pentobarbital-induced sleep. Biomed Pharmacother. 2022;146:112301. doi:10.1016/j.biopha.2021.112301
151. Janda K, Wojtkowska K, Jakubczyk K, Antoniewicz J, Skonieczna-żydecka K. Passiflora incarnata in neuropsychiatric disorders–a systematic review. Nutrients. 2020;12(12):3894. doi:10.3390/nu12123894
152. Amini F, Amini-Khoei H, Haratizadeh S, et al. Hydroalcoholic extract of Passiflora incarnata improves the autistic-like behavior and neuronal damage in a valproic acid-induced rat model of autism. J Tradit Complement Med. 2023;13(4):315–324. doi:10.1016/j.jtcme.2023.02.005
153. Abdel-Wahhab KG, Sayed RS, El-Sahra DG, Hassan LK, Elqattan GM, Mannaa FA. Echinacea purpurea extract intervention for counteracting neurochemical and behavioral changes induced by bifenthrin. Metab Brain Dis. 2024;39(1):101–113. doi:10.1007/s11011-023-01303-6
154. Kuchta K, Hladikova M, Thomsen M, Nahrstedt A, Schmidt M. Kava (Piper methysticum) extract for the treatment of nervous anxiety, tension and restlessness. Drug Res. 2021;71(2):83–93. doi:10.1055/a-1268-7135
155. Kyrou I, Christou A, Panagiotakos D, et al. Effects of a hops (Humulus lupulus L.) dry extract supplement on self-reported depression, anxiety and stress levels in apparently healthy young adults: a randomized, placebo-controlled, double-blind, crossover pilot study. Hormones. 2017;16(2):171–180. doi:10.14310/horm.2002.1738
156. Vora LK, Gholap AD, Hatvate NT, et al. Essential oils for clinical aromatherapy: a comprehensive review. J Ethnopharmacol. 2024;330:118180. doi:10.1016/j.jep.2024.118180
157. Chang J, Yang H, Shan X, et al. Bergamot essential oil improves CUMS-induced depression-like behaviour in rats by protecting the plasticity of hippocampal neurons. J Cell Mol Med. 2024;28(8):e18178. doi:10.1111/jcmm.18178
158. Zhu MY, Dong WY, Guo JR, et al. A neural circuit for bergamot essential oil-induced anxiolytic effects. Adv Sci. 2025;12(1):e2406766. doi:10.1002/advs.202406766
159. Ouyang P, Kang D, You W, Shen X, Mo X, Liu Y. Pogostemon cablin essential oil affects anxiety- and depressive-like behaviors and the gut microbiota in chronic unpredictable mild stress model rats. Front Nutr. 2024;11:1303002. doi:10.3389/fnut.2024.1303002
160. Nguyen LTH, Nguyen NPK, Tran KN, Shin HM, Yang IJ. Intranasal administration of the essential oil from Perillae Folium ameliorates social defeat stress-induced behavioral impairments in mice. J Ethnopharmacol. 2024;324:117775. doi:10.1016/j.jep.2024.117775
161. Chen J, Liu Y, Song Y, et al. Neuro-stimulating effect of Citri Reticulata Pericarpium Viride essential oil through regulating Glu/NMDAR on olfactory bulb to improve anxiety-like behavior. J Ethnopharmacol. 2024;331:118332. doi:10.1016/j.jep.2024.118332
162. Wu Y, Yu X, Wang Y, et al. Ruscogenin alleviates LPS-triggered pulmonary endothelial barrier dysfunction through targeting NMMHC IIA to modulate TLR4 signaling [published correction appears in Acta Pharm Sin B. 202212(7):3198-3199. doi: 10.1016/j.apsb.2022.04.004]. Acta Pharm Sin B. 2022;12(3):1198–1212. doi:10.1016/j.apsb.2021.09.017
163. Qi JY, Jin YC, Wang XS, et al. Ruscogenin exerts anxiolytic-like effect via microglial NF-κB/MAPKs/NLRP3 signaling pathways in mouse model of chronic inflammatory pain. Phytother Res. 2024;38(11):5417–5440. doi:10.1002/ptr.8325
164. Zhao G, He F, Wu C, et al. Betaine in inflammation: mechanistic aspects and applications. Front Immunol. 2018;9:1070. doi:10.3389/fimmu.2018.01070
165. Liu W, Zhong X, Yi Y, et al. Prophylactic effects of betaine on depression and anxiety behaviors in mice with dextran sulfate sodium-induced colitis. J Agric Food Chem. 2024;72(38):21041–21051. doi:10.1021/acs.jafc.4c05547
166. Guo JT, Li HY, Cheng C, et al. Isochlorogenic acid A ameliorated lead-induced anxiety-like behaviors in mice by inhibiting ferroptosis-mediated neuroinflammation via the BDNF/Nrf2/GPX4 pathways. Food Chem Toxicol. 2024;190:114814. doi:10.1016/j.fct.2024.114814
167. Ni M, Zhou J, Zhu Z, et al. Shikonin and cisplatin synergistically overcome cisplatin resistance of ovarian cancer by inducing ferroptosis via upregulation of HMOX1 to promote Fe2+ accumulation. Phytomedicine. 2023;112:154701. doi:10.1016/j.phymed.2023.154701
168. Shi C, Qi Z, Yang C, Luo S, Huang S, Luo Y. Shikonin ameliorates depressive- and anxiogenic-like behaviors in rats via the suppression of inflammation in the hippocampus. Neurosci Lett. 2024;837:137893. doi:10.1016/j.neulet.2024.137893
169. Guo C, Huang Q, Wang Y, et al. Therapeutic application of natural products: NAD+ metabolism as potential target. Phytomedicine. 2023;114:154768. doi:10.1016/j.phymed.2023.154768
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.