Back to Journals » Drug Design, Development and Therapy » Volume 12
Antioxidative stress effects of vitamins C, E, and B12, and their combination can protect the liver against acetaminophen-induced hepatotoxicity in rats
Authors Abdulkhaleq FM, Alhussainy TM, Badr MM, Abu Khalil AA, Gammoh O, Ghanim BY ![]() , Qinna NA
, Qinna NA ![]()
Received 28 April 2018
Accepted for publication 27 July 2018
Published 18 October 2018 Volume 2018:12 Pages 3525—3533
DOI https://doi.org/10.2147/DDDT.S172487
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Professor Manfred Ogris
Farah M Abdulkhaleq,1 Tawfiq M Alhussainy,1 Mujtaba M Badr,2 Asad A Abu Khalil,2 Omar Gammoh,3 Bayan Y Ghanim,2 Nidal A Qinna1,2
1Department of Pharmacology and Biomedical Sciences, Faculty of Pharmacy and Medical Sciences, University of Petra, Amman, Jordan; 2University of Petra Pharmaceutical Center (UPPC), University of Petra, Amman, Jordan; 3Department of Pharmacy, Faculty of Health Sciences, American University of Madaba, Madaba, Jordan
Background: Several vitamins, including C, E, and B12, have been recognized as antioxidants and have shown hepatoprotective effects against the hepatotoxicity caused by acetaminophen (APAP) overdose. The current investigation aims to study the effect of these vitamins and their combination in protecting the liver from APAP hepatotoxicity in rats.
Materials and methods: An in vitro model of freshly isolated rat hepatocytes was utilized for assessing hepatocyte mitochondrial activity conducted by cell proliferation assay (MTT). The isolated hepatocytes were treated with vitamin C, vitamin E, vitamin B12 and their combination, with and without further addition of toxic concentrations of APAP. In addition, an in vivo experiment was carried out on Sprague Dawley rats treated intraperitoneally for 8 days with emulsions of the vitamins or their combination prior to injecting them with APAP.
Results: In vitro results showed that vitamins C and B and the combination preparation significantly increased the percentage of hepatocyte mitochondrial activity, both with and without the addition of APAP (P<0.01). The mitochondrial activity in the isolated cultured hepatocytes was further enhanced with APAP addition. In vivo, the vitamins and their combination effectively reduced APAP-induced serum liver enzymes levels, namely ALT, AST, and ALP, and also attenuated oxidative stress and lipids peroxidation confirmed by the results of glutathione, superoxide dismutase, and maloondialdehyde.
Conclusion: Pretreatment with vitamins C, E, B12, or their combination was found to be beneficial in preventing in vivo hepatic oxidative stress induced by APAP overdose. Vitamin C on its own showed superior protection against APAP-induced liver injury in rats compared to the other vitamins. The proliferation of APAP-intoxicated liver cells in vitro was highest when protected with the vitamins’ combination.
Keywords: glutathione, hepatocytes, proliferation, hepatoprotective, superoxide dismutase, primary cell culture
Introduction
Drug-induced hepatotoxicity is encountered in clinical practice and rises within easily accessible compounds like over-the-counter drugs. Acetaminophen, N-acetyl-para-aminophenol (APAP) or paracetamol, was originally introduced to the market as an analgesic drug by Von Mering in 1893,1 mostly used as an analgesic and antipyretic for relieving mild and moderate pain and fever.2,3 APAP-induced liver injury is translated to acute liver failure, with admission of more than 30,000 patients every year.4 The spectrum of injury caused by APAP triggers the liver all the way down to its hepatocytes and nuclear molecules.5 Interestingly, infants and young children are less susceptible to acute APAP toxicity due to age-associated differences in drug metabolism.6
At therapeutic levels, most of the administered dose is normally metabolized by glucuronidation and sulfonation (Phase II) to produce inactive nontoxic metabolites that are easily excreted by kidney.7 On the other hand, a small portion is metabolized by oxidation (Phase I) through CYP2E1, a member of the CYP family,8 to a highly toxic metabolite, namely, N-acetyl-p-benzoquinoneimine (NAPQI), a reactive metabolite that depletes glutathione (GSH) and covalently binds to mitochondrial proteins. NAPQI is efficiently detoxified by conjugation with GSH. However, at toxic doses, GSH is depleted by the conjugation reaction, and NAPQI covalently binds to proteins to produce reactive oxygen species (ROS). ROS induce oxidative stress leading to lipid peroxidation, mitochondrial dysfunction, disruption of calcium, and nitric oxide homeostasis, and finally, cell death by apoptosis and necrosis.9
Antioxidants may exert their effects on biological systems by several mechanisms including electron donation, chelation of metal ions, regulation of gene expression, or by acting as co-antioxidants.10,11 Among biologically important antioxidants, vitamins C, E, and B12 function mainly by two principle mechanisms of action; initially by donating electrons to free radicals: the chain-breaking mechanism. The second mechanism involves removal of initiators of reactive oxygen or nitrogen species, acting as secondary antioxidants by quenching a chain-initiating catalyst.12–14
Vitamin C (VC), also known as ascorbic acid, is a potent naturally occurring water-soluble antioxidant.15 Vitamin E (VE), a potent lipid-soluble antioxidant,16 is found in the plasma and tissues as an isomer, namely, α-tocopherol, the most important antioxidant in cell membranes.17 It also regulates cell proliferation, platelet aggregation, and NADPH-oxidase activation, as well as the expression of genes responsible for its own metabolism, cell adhesion, inflammation, and fibrosis.18 Vitamin B12 (VB12) (cyano cobalamin) is a water-soluble vitamin essential for maintaining the normal functions of the peripheral nervous system19 and the brain, as it plays a considerably critical role in the myelination of the white matter and nerves.20 The potential antioxidative properties and the ability of cobalamins in regulating inflammatory cytokines and have been previously reported.13 Furthermore, the synthetic cyanocobalamin is one of cobalamin forms that is available commercially as a dietary supplement.21
Since APAP-induced liver injury represents the most frequent cause of drug-induced liver failure,10 and such injury is mainly attributed to acute induction of oxidative stress in hepatocytes, the aim of the present study was to evaluate the hepatoprotective effects of VC, VE, and VB12 on APAP-induced hepatotoxicity. Moreover, we also wanted to investigate whether a preparation comprising a combination of all three vitamins could augment the expected hepatic protection.
Materials and methods
Materials and chemicals
All the materials used for the experiment were of analytical grade. VC, VE, and VB12, EDTA, MgCl2, FBS, streptomycin, penicillin, RPMI-1640 medium, olive oil, and GSH ELISA assay kit were all purchased from Sigma Aldrich (St Louis, MO, USA). Hank’s balanced salt solution (HBSS) was obtained from Invitrogen (Carlsbad, CA, USA). Collagenase II and L-glutamine were purchased from Gibco BRL (Gaithersburg, MD, USA). Tween 80 and DMSO were obtained from Scharlan Chemie S.A. (Barcelona, Spain), whereas Tris base and TACS® MTT kit were purchased from Promega (Madison, WI, USA) and Trevigen (Gaithersburg, MD, USA), respectively. Acetaminophen and isoflorane were kind gifts from the Jordanian Pharmaceutical Manufacturing Company JPM (Amman, Jordan) and Hikma Pharmaceuticals (Amman, Jordan), respectively. ELISA kits for superoxide dismutase (SOD) and malondialdehyde (MDA) were purchased from MyBioSource, (San Diego, CA, USA).
Experimental animals
Adult male Sprague Dawley rats with average weights of 240±20 g were housed at the Animal House Unit of the University of Petra (Amman, Jordan). Rats were kept in air-conditioned environment under controlled temperatures, humidity, and photoperiod cycles and acclimatized for 10 days before experimenting. Rats were fasted overnight (for 18–22 hours) with free access to water, unless otherwise stated. All experiments were approved by the Research Council at the Faculty of Pharmacy and Medical Sciences (approval number: 15/2/2016) and conducted in accordance with the University of Petra Institutional Guidelines on Animal Use, which adopts the guidelines of the Federation of European Laboratory Animal Science Association.
Preparation of buffers
Liver perfusion buffers were freshly prepared under sterile techniques (Al Shaker et al25). Perfusion buffer I was prepared by adding 0.9 mM MgCl2 (95.211 g/mol), 0.5 mM EDTA (292.24 g/mol), and 0.5 mM Tris base (121.14 g/mol) to 0.5 L HBSS (without Ca2+ and Mg2+). Perfusion buffer II was prepared by adding 0.5 mM Tris base to 0.5 L HBSS (with Ca2+ and Mg2+). Perfusion buffer II with collagenase II was prepared by dissolving appropriate amount of collagenase II equivalent to 1,000 IU in 300 mL of perfusion buffer II. RPMI medium was prepared by the addition of 2 mM of L-glutamine, 5% FBS, 100 IU/mL penicillin, and 100 mg/mL of streptomycin to RPMI-1640 medium.
Preparation of emulsions and suspensions
Owing to the nature and solubility of the tested vitamins, an oil-in-water (o/w) emulsion was found to be a suitable carrier for the vitamins and their combination. All vitamins were prepared as o/w emulsions, using 10% olive oil as the oily phase and 2.5% Tween 80 as the emulsifying agent. Primary emulsions were prepared using dry gum method under the same conditions. VC and VB12, being water-soluble vitamins, were dissolved in distilled water (the aqueous phase). On the other hand, VE, being a lipid-soluble vitamin, was dissolved in olive oil followed by the addition of Tween 80. The resultant oily phase was then mixed with the aqueous phase using a vortex mixer. The final volume of each prepared emulsion was completed with sufficient quantity of distilled water. All preparations were freshly made before use.
The in vivo dose selections of 500 mg/kg VC, 200 mg/kg VE, 10 mg/kg VB12, and their combination were based on previous studies.19,22,23
An oral dose of 2,800 mg/kg/d of APAP was used to induce hepatotoxicity in rats.9 APAP stock was freshly prepared before use by weighing appropriate amount of the drug and suspending it in distilled water for in vivo testing or in RPMI media for in vitro intoxications of isolated hepatocytes with a concentration of 10 mM APAP.24
Surgical and liver perfusion protocols
In order to perfuse the liver with collagenase enzyme, a small animal general anesthesia system (SomnoSuite, Kent Scientific Corporation, Torrington, CT, USA) linked to an oxygen concentrator (Dual Flow Oxygen Concentrator, Longfian Scitech Co., Ltd. Hebei, People’s Republic of China), a water bath (Elmasonic S, Elma, Singen, Germany), a peristaltic perfusion pump (Langer Instruments Corporation, Boonton, NJ, USA), and a portable phlegm suction unit (Ningbo Sofine Electric Co., Ltd., Zhejiang, People’s Republic of China) were used.
The adult rat was placed on a surgical board (Plas Labs, Lansing, MI, USA), above a heating pad maintained at 37°C, and anesthetized by isoflurane (5% for induction and 2.5% for maintenance).25 A midline longitudinal incision was made to expose the rat’s viscera. Hepatic portal vein was sighted and cannulated with an 18-gauge angiocath (Becton Dickinson, Mountain View, CA, USA) and perfused with a prewarmed (37°C) perfusion buffer I at a flow rate of 10 mL/min. The perfused buffer was allowed to efflux from the body by cutting the inferior vena cava from the right kidney side. Thereafter, perfusion was switched to perfusion buffer II plus collagenase II, and the flow rate was increased to 25 mL/min and proceeded until the liver became mushy and pale in color. The liver was then dissected and placed into a sterile stoppered container with buffer II plus collagenase II, to be transferred to a sterile cell culture hood.
Hepatocyte isolation
Liver cells were dispersed gently under laminar flow, using sterile cell scrapers (Fisher Scientific Ltd., Loughborough, UK) in a sterile petri dish containing buffer II plus collagenase II. Connective tissues and undigested tissue fragments were removed from the cell suspension and centrifuged at 100 RCF for 3 minutes at 4°C. Cells were then resuspended in RPMI medium, and centrifuged at 200 RCF for 10 minutes at 4°C. After centrifugation, the supernatant was aspirated again, and cells were resuspended in 10 mL warm RPMI medium and centrifuged at 200 RCF for 10 minutes at 15°C. Finally, the resultant cell suspension was counted under light microscope using a hemocytometer (Marienfeld-Superior Co., Lauda-Königshofen, Germany).
Hepatocyte culture
Hepatocytes were diluted with appropriate amount of warm RPMI medium to the desired concentration in order to reach 70%–80% confluence. Hepatocytes were seeded into sterile 96 well plates and allowed to settle for 30 minutes before incubation in a humidified CO2 incubator overnight at 37°C to allow cells to recover and grow in monolayer prior to testing.
In vitro testing protocol and cell proliferation assay
After 24 hours of incubation, the seeded plates were treated with the freshly prepared emulsions, at concentrations containing 500 μM VC,26 180 μM VE,27 100 μM VB12,19 or their combination, with or without further addition of APAP, followed by 3 hours incubation. Later, RPMI media with and without 10 mM APAP24 was added to its corresponding hepatocyte-containing wells and further incubated for 48 hours. For assessing the mitochondrial activity for cultured hepatocytes, which also resembles cellular proliferation, TACS MTT cell proliferation assay was performed according to the manufacturer’s instructions (Trevigen TACS MTT assay and Promega Glomax® Microplate Reader).
In vivo protocols of hepatoprotection
An in vivo study was performed on 56 adult male Sprague Dawley rats divided randomly into seven groups (n=8) and treated for 8 days. Control groups were injected intraperitoneally with placebo o/w emulsion (Groups I and II), whereas the other groups were treated with 500,28 20023, and 1029 mg/kg/d of VC, VE, and VB12, respectively (Groups III, IV, and V). Groups VI and VII were treated with the combination of the three vitamins either intraperitoneally or orally, respectively, by suspending equivalent amount of vitamins to obtain the same doses per vitamin as mentioned for the preceding treatment groups. On the other hand, systemic liver toxicity was induced by administering APAP (2,800 mg/kg) twice,9 at days 7 and 8, for all groups except Group I (negative control). At day 9, all rats were sacrificed and blood samples were withdrawn by cardiac puncture and left at room temperature for 15 minutes to clot completely. Blood samples were centrifuged at 6,000 rpm for 10 minutes at room temperature to obtain serum and stored at −20°C until requested for analysis. In addition, serum samples were referred for analysis of liver function biomarkers (ALT, AST, ALP, albumin, and cholesterol) at Al-Mahabbah Hospital, Madaba, Jordan. Stress biomarkers (SOD, MDA, and total GSH) were assessed in-house using ELISA kits, according to the manufacturers’ procedures and instructions.
Data analysis
Each data point represents the mean±the standard error of the mean (SEM). Statistical comparisons were made using SPSS statistical software (version 20, IBM Company, Armonk, NY, USA), using one-way ANOVA followed by Tukey’s post Hoc Multiple Comparison test to compare between groups. P-values <0.05 were considered statistically significant (P<0.05).
Results
Cotreatment of hepatocytes with VC, VE, and VB12 enhances mitochondrial activity
The selected concentration of 10 mM of APAP significantly decreased the cellular proliferation of the isolated rat hepatocytes approximately by half (Figure 1). Figure 1 illustrates the mitochondrial activity of isolated rat hepatocytes treated with the vitamins when examined on nonintoxicated isolated hepatocytes compared to APAP-intoxicated cells. The tested vitamins did not show any notable proliferation changes in healthy isolated hepatocytes except for VE that significantly reduced hepatocyte mitochondrial activity (P<0.01). On the other hand, when APAP-intoxicated hepatocytes were pretreated with the vitamins or their combination, the mitochondrial activity was significantly increased (P<0.01), and highest proliferation activity was observed in the cells treated with the combination.
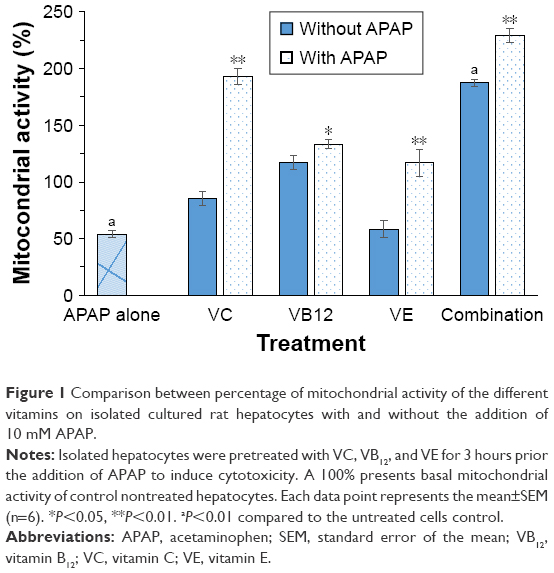 | Figure 1 Comparison between percentage of mitochondrial activity of the different vitamins on isolated cultured rat hepatocytes with and without the addition of 10 mM APAP. |
VC, VE, and VB12 ameliorate APAP-induced hepatotoxicity
As illustrated in Table 1, treating rats with a high hepatotoxic dose of APAP caused significant increase in serum levels of ALT, AST, and ALP (P<0.01), although it did not cause any pronounced change in serum cholesterol and albumin levels. The effect of VC on protecting the liver from APAP toxicity was quite noticeable, where it significantly reduced serum levels of all tested liver enzymes (P<0.01). On the other hand, VE and VB12 caused a statistically significant decrease (P<0.01) in the levels of ALT and ALP. A decrease in AST levels was also noticed, although not statistically significant. Orally administered combination prior APAP intoxication in rats decreased the overproduction of ALT, AST, and ALP in their serum while intraperitoneal administered combination reduced only ALT and AST levels (Table 1).
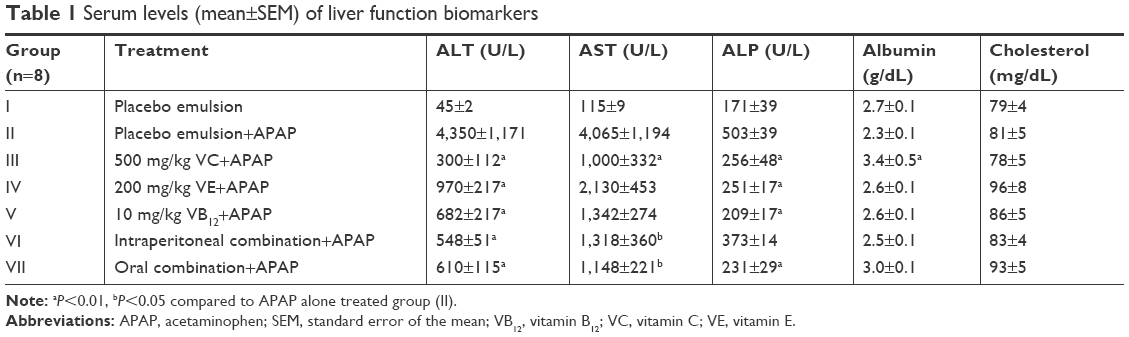 | Table 1 Serum levels (mean±SEM) of liver function biomarkers |
Vitamins C, E, and B12 ameliorate APAP-induced oxidative stress
Serum levels of MDA, SOD, and total GSH are illustrated in Figures 2–4, respectively. When rats were intoxicated with an overdose of APAP, a marked increase in serum MDA, SOD, and total GSH levels was observed (P<0.01). Pretreatment of rats with VC, VE, VB12, or their combinations prior to APAP intoxication has significantly prevented such increase (P<0.01). Although the decrease in APAP-induced lipid peroxidation, observed by MDA formation, is lowered by pretreatment with mentioned vitamins, they did not show complete reversal down to control basal levels. The significant reversal of APAP-induced increase in GSH to norm was observed in both VC and VB12. However, all vitamin reversed SOD production in APAP-injected rats.
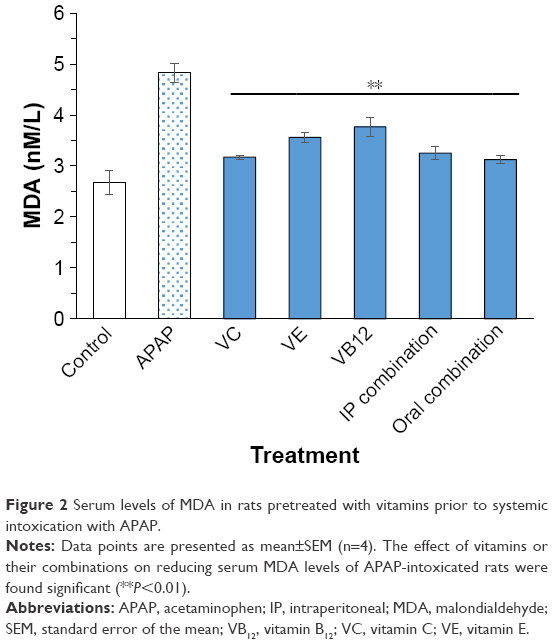 | Figure 2 Serum levels of MDA in rats pretreated with vitamins prior to systemic intoxication with APAP. |
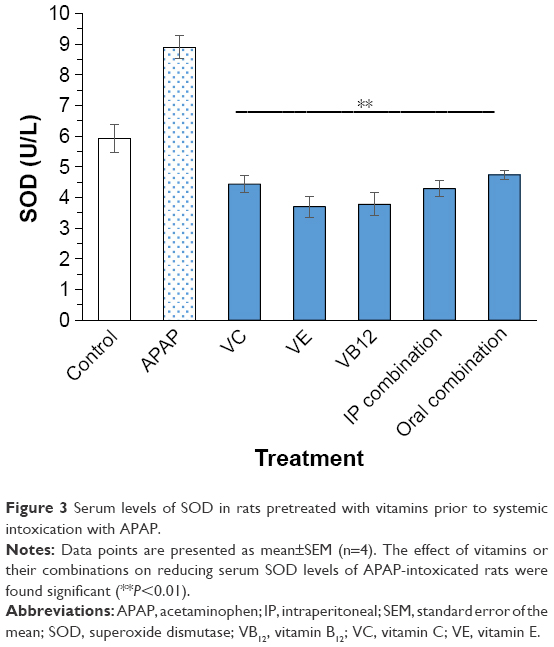 | Figure 3 Serum levels of SOD in rats pretreated with vitamins prior to systemic intoxication with APAP. |
Discussion
Vitamins have been reported to have pharmacologically protective effects owing to their antioxidant and various cell regulatory roles. APAP, in animal models, can induce hepatic injury, usually pointed out as hepatocyte apoptosis and necrosis. High doses of APAP have also been shown to decrease hepatocyte viability and survival.30 Having known that mice are more susceptible to APAP-induced injury and rats show more resistance unless high doses are employed,31 the current study investigates the potential hepatoprotective effects of specific vitamins in inhibiting such injury. It was demonstrated that the tested vitamins, namely, VC, VE, and VB12, significantly reduced APAP-induced liver injury in Sprague Dawley rats. Their combination also significantly enhanced cell viability and mitochondrial activity in vitro for both APAP-treated and untreated hepatocytes. All vitamins, alone or in combination, exerted statistically significant increase in cellular activity and proliferation in APAP-treated cell sections of ~2-fold in comparison to untreated control cells (P<0.01). Nevertheless, the results indicated that VC and VB12 had a moderate influence on mitochondrial activity when tested on normal hepatocytes (Figure 1). Such an enhancement might be sufficient to protect the cells from oxidative stress and excess free radical and ROS formation, as reported in previous studies.18,32
Liver transaminases, namely, ALT and AST, remain the gold standards for the assessment of liver injury and have been used for a long time as the biomarkers of choice.33 These enzymes are released into blood after hepatic cell injury, and their elevated serum levels are indicative of cellular leakage and loss of functional integrity of hepatocyte cell membranes.2 The increase in ALT levels is more specific for liver damage than AST, since it is mainly found in the liver, whereas AST is also found in other tissues such as heart, skeletal muscles, kidney, red blood cells, and brain.34 Intoxication of rats with APAP highly increased the levels of serum ALT and AST, indicating liver injury. However, pretreatment of rats with 500 mg/kg VC, 200 mg/kg VE, 10 mg/kg VB12, or their combination, daily for 8 days, significantly inhibited the increase of ALT serum levels; however, VC and the combination group significantly decreased AST serum levels unlike VE and VB12, which showed insignificant effect. These results are in line with many previously reported studies32,35,36 where such reduction in liver enzymes indicates the possession of membrane-stabilizing activity in the tested vitamins and might indicate their ability to induce regeneration of liver cells, therefore reducing the leakage of the liver enzymes into the blood.
In contrast to ALT and AST, serum level of ALP is related to the function of hepatic cell rather than its integrity.2,33 In this study, ALP levels were significantly elevated in APAP group (Group II), while pretreatment with the vitamins or their combinations inhibited such increase, although not statistically significant in the intraperitoneal combination group (Group VI).
Usually, injury of the liver inhibits synthesis of major proteins including albumin. A decrease in albumin levels was noted in rats treated with high doses of APAP alone. This can be endorsed to APAP-induced disruption in hepatocyte integrity, which either in turn leads to decreased hepatic capacity for protein synthesis, or increased proteolysis and degradation activities. VC showed the highest protection against APAP-induced albumin deprivation unlike the other groups, which exhibited an insignificant effect. This might be linked to its considerable ability to protect hepatocytes from injury, thereby reserving its role in albumin levels modulation.
Besides albumin, cholesterol is one of many molecules synthesized by the liver and thereafter released to the bloodstream.33 To date, there is no clear and constant published report confirming the effect of APAP overdose on modulating in vivo cholesterol levels. Some previous studies reported an increase in cholesterol serum levels after APAP overdose,32 while others claimed decreased levels,37 whereas few other reports claimed no remarkable change.38 This variability in results might be related to the dose and frequency of APAP administration which affects the extent of liver damage on many enzymes including cholesterol 7-α-hydroxylase (CYP7A1), the rate-limiting enzyme in bile acid biosynthesis that converts cholesterol to bile acid.38 However, in this study, no noticeable change in cholesterol levels was observed after administration of the selected toxic dose of APAP.
It is well documented that liver tissue contains relatively high content of polyunsaturated fatty acids, which are very sensitive to peroxidative damage.39 Peroxidation of polyunsaturated fatty acids results in MDA formation, a biochemical indicator of necrosis.40,41 MDA is a biomarker for oxidative stress that indicates a state of damage when detected in serum. The presented results show that treatment of rat hepatocytes with APAP markedly increased MDA levels by nearly 2-fold, whereas pretreatment with vitamins C, E, and B12 or their combinations (intraperitoneal and oral) significantly inhibited such an increase. The observed protective effect of these vitamins against lipid peroxidation could be related to their antioxidant effects, which assist in the preservation of membrane integrity through radical scavenging.18
The exact mechanism of APAP-induced oxidative stress is yet not fully elucidated. However, upon an APAP overdose, the reactive metabolite of APAP, namely NAPQI, induces oxidative stress and increases ROS production in the cell leading to its destruction and death. It is highly recognized that SOD and GSH are antioxidants that protect cells against the increased ROS and oxidative insult following APAP administration.
SOD catalyzes the dismutation of superoxide radical to hydrogen peroxide.42 In the present work, oral administration of a toxic dose of APAP to Sprague Dawley rats for 2 consecutive days caused significant elevation of serum SOD levels. This elevation may postulate an over release or expression of SOD in the intoxicated rats as a defense mechanism to overcome increased ROS formation and compensate the oxidative stress evolved. On the other hand, daily pretreatment with VC, VE, and VB12 or their combination over 8 consecutive days significantly reversed APAP-induced increase in SOD levels, thus indicating their potential radical scavenging effects that compensate the role of SOD in defending APAP-induced oxidative stress.
Concerning the involvement of GSH in APAP hepatotoxicity, results showed an increase in serum GSH levels in rats after APAP administration (Figure 4), while pretreatment with the three vitamins solely or their combination effectively reversed such increase down to the control levels, except for VE that significantly inhibited the injury but did not fully protect the liver from the insult. The abnormal increase in GSH content after a high dose of APAP may be due to another indirect resistance mechanism achieved by rats to overcome the toxicity and increased formation of ROS. The decrease in serum total GSH levels with the vitamins and their combinations may be due to their ability to neutralize the ROS and, as a result, preventing them from depleting GSH stores. It has been previously reported that high doses of VC are capable of normalizing GSH levels in rats injected with toxic doses of APAP,43,44 and that is in line with our results. As discussed in the case of SOD earlier, the observed elevation of GSH might also be attributed to a compensatory defense mechanism against stress induced by APAP overdose. Such results are in line with the previous work of Aycan et al,45 on Wistar rats where serum ALT, AST, GSSG, and SOD activity, as well as the serum and tissue MDA levels were found to be higher in the APAP group (500 mg/kg) than in the control group.
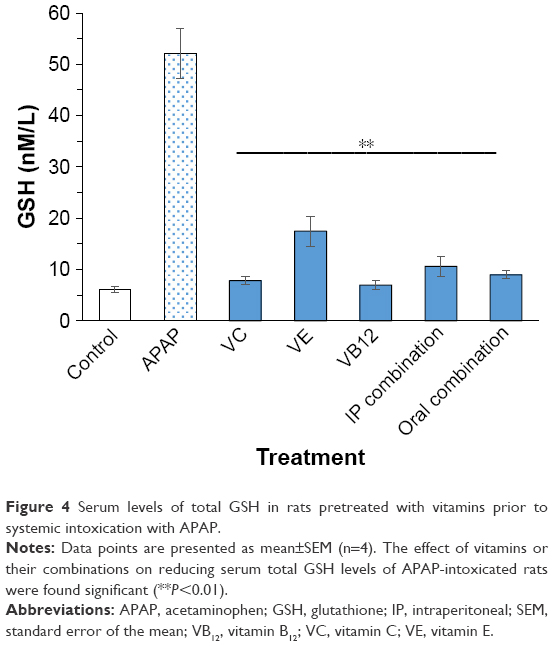 | Figure 4 Serum levels of total GSH in rats pretreated with vitamins prior to systemic intoxication with APAP. |
Conclusion
Pretreatment with VC, VE, and VB12 or their combination was shown to protect the livers of rats, both in vitro and in vivo, and prevented APAP-induced liver damage. The survival of APAP-intoxicated liver cells in vitro was highest when protected with the vitamin combination followed by VC, VB12, and VE, respectively. Yet, it seems that VC on its own may have a superior protection against APAP-induced liver injury compared with the other vitamins. VC has also shown significant protection actions on the isolated liver cells. The outcome of this research suggests that the use of one or more of the investigated vitamins can protect from hepatotoxicity or liver-induced injury which may occur from APAP overdose; however, such claims must be tested clinically. Therefore, future investigations are needed to approve the potential protective effects of the tested vitamins on APAP-induced injury clinically in humans compared to other available APAP antidotes and further elucidate the molecular mechanisms behind such capability.
Acknowledgments
The authors would like to acknowledge Deanship of Scientific Research at the University of Petra, Amman, Jordan, for their support and funding for this project (grant number 3/4/2016 and 9/4/2017). Mujtaba M Badr, is currently affiliated with the College of Pharmacy and Nutrition, University of Saskatchewan, Saskatoon, SK, Canada.
Disclosure
The authors report no conflicts of interest in this work.
References
Prescott LF. Paracetamol: past, present, and future. Am J Ther. 2000;7(2):143–147. | ||
Eesha BR, Mohanbabu Amberkar V, Meena Kumari K, et al. Hepatoprotective activity of Terminalia paniculata against paracetamol induced hepatocellular damage in Wistar albino rats. Asian Pac J Trop Med. 2011;4(6):466–469. | ||
Athersuch TJ, Antoine DJ, Boobis AR, et al. Paracetamol metabolism, hepatotoxicity, biomarkers and therapeutic interventions: a perspective. Toxicol Res. 2018;7(3):347–357. | ||
Blieden M, Paramore LC, Shah D, Ben-Joseph R. A perspective on the epidemiology of acetaminophen exposure and toxicity in the United States. Expert Rev Clin Pharmacol. 2014;7(3):341–348. | ||
Ruepp SU, Tonge RP, Shaw J, Wallis N, Pognan F. Genomics and proteomics analysis of acetaminophen toxicity in mouse liver. Toxicol Sci. 2002;65(1):135–150. | ||
Heubi JE, Barbacci MB, Zimmerman HJ. Therapeutic misadventures with acetaminophen: hepatoxicity after multiple doses in children. J Pediatr. 1998;132(1):22–27. | ||
McGill MR, Sharpe MR, Williams CD, Taha M, Curry SC, Jaeschke H. The mechanism underlying acetaminophen-induced hepatotoxicity in humans and mice involves mitochondrial damage and nuclear DNA fragmentation. J Clin Invest. 2012;122(4):1574–1583. | ||
Ben-Shachar R, Chen Y, Luo S, Hartman C, Reed M, Nijhout HF. The biochemistry of acetaminophen hepatotoxicity and rescue: a mathematical model. Theor Biol Med Model. 2012;9(9):55. | ||
Qinna NA, Shubbar MH, Matalka KZ, Al-Jbour N, Ghattas MA, Badwan AA. Glucosamine enhances paracetamol bioavailability by reducing its metabolism. J Pharm Sci. 2015;104(1):257–265. | ||
Yin H, Cheng L, Holt M, Hail N, Maclaren R, Ju C. Lactoferrin protects against acetaminophen-induced liver injury in mice. Hepatology. 2010;51(3):1007–1016. | ||
Lü J-M, Lin PH, Yao Q, Chen C. Chemical and molecular mechanisms of antioxidants: experimental approaches and model systems. J Cell Mol Med. 2010;14(4):840–860. | ||
Chen ZJ. Ubiquitin signalling in the NF-kappaB pathway. Nat Cell Biol. 2005;7(8):758–765. | ||
Birch CS, Brasch NE, Mccaddon A, Williams JH. A novel role for vitamin B(12): Cobalamins are intracellular antioxidants in vitro. Free Radic Biol Med. 2009;47(2):184–188. | ||
Saito Y, Shichiri M, Hamajima T, et al. Enhancement of lipid peroxidation and its amelioration by vitamin E in a subject with mutations in the SBP2 gene. J Lipid Res. 2015;56(11):2172–2182. | ||
Adikwu E, Deo O. Effect of vitamin C (ascorbic acid). Pharmacol Pharm. 2013;4(1):84–92. | ||
Traber MG, Stevens JF, Vitamins C. Vitamins C and E: beneficial effects from a mechanistic perspective. Free Radic Biol Med. 2011;51(5):1000–1013. | ||
Dufour JF, Clavien PA, Graf R, Trautwein C, editors. Signaling Pathways in Liver Diseases. New York: Springer; 2005. | ||
González R, Cruz A, Ferrín G, et al. Nitric oxide mimics transcriptional and post-translational regulation during α-tocopherol cytoprotection against glycochenodeoxycholate-induced cell death in hepatocytes. J Hepatol. 2011;55(1):133–144. | ||
Nishimoto S, Tanaka H, Okamoto M, Okada K, Murase T, Yoshikawa H. Methylcobalamin promotes the differentiation of Schwann cells and remyelination in lysophosphatidylcholine-induced demyelination of the rat sciatic nerve. Front Cell Neurosci. 2015;9:298. | ||
Gupta L, Gupta RK, Gupta PK, Malhotra HS, Saha I, Garg RK. Assessment of brain cognitive functions in patients with vitamin B12 deficiency using resting state functional MRI: A longitudinal study. Magn Reson Imaging. 2016;34(2):191–196. | ||
Stewart JS, Roberts PD, Hoffbrand AV. Response of dietary vitamin-b12 deficiency to physiological oral doses of cyanocobalamin. Lancet. 1970;296(7672):542–545. | ||
Mardones L, Zúñiga FA, Villagrán M, et al. Essential role of intracellular glutathione in controlling ascorbic acid transporter expression and function in rat hepatocytes and hepatoma cells. Free Radic Biol Med. 2012;52(9):1874–1887. | ||
Ming Z, Fan YJ, Yang X, Lautt WW, Wayne Lautt W. Synergistic protection by S-adenosylmethionine with vitamins C and E on liver injury induced by thioacetamide in rats. Free Radic Biol Med. 2006;40(4):617–624. | ||
Lewerenz V, Hanelt S, Nastevska C, El-Bahay C, Röhrdanz E, Kahl R. Antioxidants protect primary rat hepatocyte cultures against acetaminophen-induced DNA strand breaks but not against acetaminophen-induced cytotoxicity. Toxicology. 2003;191(2–3):179–187. | ||
Al Shaker HA, Qinna NA, Badr M, et al. Glucosamine modulates propranolol pharmacokinetics via intestinal permeability in rats. Eur J Pharm Sci. 2017;105(May):137–143. | ||
Montecinos V, Guzmán P, Barra V, et al. Vitamin C is an essential antioxidant that enhances survival of oxidatively stressed human vascular endothelial cells in the presence of a vast molar excess of glutathione. J Biol Chem. 2007;282(21):15506–15515. | ||
Nizam N, Discioglu F, Saygun I, et al. The effect of α-tocopherol and selenium on human gingival fibroblasts and periodontal ligament fibroblasts in vitro. J Periodontol. 2014;85(4):636–644. | ||
Li R, Shen L, Yu X, Ma C, Huang Y. Vitamin C enhances the analgesic effect of gabapentin on rats with neuropathic pain. Life Sci. 2016;157:25–31. | ||
Ahmad A, Afroz N, Gupta UD, Ahmad R. Vitamin B 12 supplement alleviates N′-nitrosodimethylamine-induced hepatic fibrosis in rats. Pharm Biol. 2014;52(4):516–523. | ||
Hinson JA, Roberts DW, James LP. Mechanisms of acetaminophen-induced liver necrosis. Handb Exp Pharmacol. 2010;196:369–405. | ||
Mcgill MR, Williams CD, Xie Y, Ramachandran A, Jaeschke H. Acetaminophen-induced liver injury in rats and mice: comparison of protein adducts, mitochondrial dysfunction, and oxidative stress in the mechanism of toxicity. Toxicol Appl Pharmacol. 2012;264(3):387–394. | ||
Otuechere CA, Abarikwu SO, Rufai MA, Ohiozoje AE, Martins E, Farombi EO. Protective effects of vitamin C against propanil-induced hepatotoxicity in wistar rats. Asian Pac J Trop Dis. 2012;2(Suppl 1):S212–S217. | ||
Washington IM, Van Hoosier G. Clinical biochemistry and hematology. In: Suckow MA, Stevens KA, Wilson RP, editors. The Laboratory Rabbit, Guinea Pig, Hamster, and Other Rodents. New York: Elsevier; 2012:57–116. | ||
Amacher DE. A toxicologist’s guide to the preclinical assessment of hepatic microsomal enzyme induction. Toxicol Mech Methods. 2006;16(7):385–394. | ||
Cuce G, Çetinkaya S, Koc T, et al. Chemoprotective effect of vitamin E in cyclophosphamide-induced hepatotoxicity in rats. Chem Biol Interact. 2015;232:7–11. | ||
Abdel-Azeem AS, Hegazy AM, Ibrahim KS, Farrag AR, El-Sayed EM. Hepatoprotective, antioxidant, and ameliorative effects of ginger (Zingiber officinale Roscoe) and vitamin E in acetaminophen treated rats. J Diet Suppl. 2013;10(3):195–209. | ||
Anantha KC, Siva RC, Manohar RA, Reddy C, Am R, Dak C. Hepatoprotective effect of biherbal ethanolic extract against paracetamol-induced hepatic damage in albino rats. J Ayurveda Integr Med. 2012;3(4):198. | ||
Ramirez MI, Karaoglu D, Haro D, Barillas C, Bashirzadeh R, Gil G. Cholesterol and bile acids regulate cholesterol 7 alpha-hydroxylase expression at the transcriptional level in culture and in transgenic mice. Mol Cell Biol. 1994;14(4):2809–2821. | ||
Catalá A. Five decades with polyunsaturated Fatty acids: chemical synthesis, enzymatic formation, lipid peroxidation and its biological effects. J Lipids. 2013;2013:710290. | ||
Jaeschke H, Gores GJ, Cederbaum AI, Hinson JA, Pessayre D, Lemasters JJ. Mechanisms of hepatotoxicity. Toxicol Sci. 2002;65(2):166–176. | ||
Ho E, Karimi Galougahi K, Liu CC, Bhindi R, Figtree GA. Biological markers of oxidative stress: Applications to cardiovascular research and practice. Redox Biol. 2013;1(1):483–491. | ||
Mirochnitchenko O, Weisbrot-Lefkowitz M, Reuhl K, Chen L, Yang C, Inouye M. Acetaminophen toxicity. Opposite effects of two forms of glutathione peroxidase. J Biol Chem. 1999;274(15):10349–10355. | ||
El-Ridi MR, Rahmy TR. Action of vitamin C against acetaminophen-induced hepatorenal toxicity in rats. J Toxicol. 2000;19(3–4):275–304. | ||
Mottot G, Margetts G. Effect of vitamin c on acute paracetamol poisoning (Paracetamol overdose; antidotes; vitamin C) Paracetamol is a safe antipyretic-analgesic drug when used at recommended dose levels. However, in cases of overdosage it can induce severe and sometimes. Toxicol Lett. 1983;18(1–2):153–156. | ||
Aycan IÖ, Tüfek A, Tokgöz O, et al. Thymoquinone treatment against acetaminophen-induced hepatotoxicity in rats. Int J Surg. 2014;12(3):213–218. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.