Back to Journals » Neuropsychiatric Disease and Treatment » Volume 21
Antidepressant Effect of Short-Chain Fatty Acids on Chronic Unpredictable Mild Stress in Model Rats And Their Influence on the Oral-Gut Microbiota
Authors Li Y, Cao J, Yang Z, Chen H
Received 2 September 2025
Accepted for publication 9 December 2025
Published 17 December 2025 Volume 2025:21 Pages 2855—2868
DOI https://doi.org/10.2147/NDT.S560434
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Taro Kishi
Yan Li,1,* Jiajun Cao,2,* Zhiyue Yang,2 Hongli Chen1
1Department of Stomatology, Fenyang College of Shanxi Medical University, Luliang, Shanxi, People’s Republic of China; 2Department of Clinical Medicine, Fenyang College of Shanxi Medical University, Luliang, Shanxi, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yan Li, Department of Stomatology, Fenyang College of Shanxi Medical University, Luliang, Shanxi, People’s Republic of China, Tel +86 182 3411 9809, Email [email protected]
Purpose: To investigate the effects of short-chain fatty acids (SCFAs) on depression-like behaviors and the oral gut microbiota in rats subjected to chronic unpredictable mild stress (CUMS) using behavioral tests and high-throughput sequencing of 16SrRNA.
Methods: Sprague-Dawley rats were randomly divided into Control, CUMS, and SCFAs groups. Except for the Control group, all rats underwent CUMS modeling and drug administration for 28 days. Behavioral tests assessed depression-like behaviors, while 16SrRNA sequencing analyzed oral and gut microbiota changes. Histopathological examination of the colon tissues and immunohistochemical analysis of the tight junction protein ZO-1 were performed.
Results: Interventions using SCFAs can alleviate symptoms in rats with CUMS-induced depression. Analysis of microbial diversity revealed significant differences in both Alpha and Beta diversities of the gut microbiota among all three groups. The composition of the microbiota showed that, at the phylum level, in the intestinal microbiota, the Firmicutes/Bacteroidetes (F/B) ratio in the SCFAs group was significantly lower than that in the CUMS group. At the genus level, Lactobacillus was the dominant bacterium in the gut. Rothia was predominant among the oral bacteria. Linear discriminant effect size (LEfSe) analysis revealed that, in the gut, the relative abundances of Bifidobacterium, Collinsella, and Sphingobium increased in the SCFAs group. In the oral cavity, the relative abundance of Prevotella increased in the CUMS group but decreased in the SCFAs group. The histopathological examination of the colon revealed that, compared with the CUMS group, in the SCFAs group, the inflammatory cells in the lamina propria decreased, and the tight junction protein ZO-1 in colonic epithelium cells increased.
Conclusion: SCFAs intervention ameliorated depression-like behaviors, modulated gut microbiota composition, enhanced ZO-1 expression, reduced gut inflammation, and promoted immune function, thereby restoring oral gut microbiota homeostasis. This study elucidated SCFAs’ therapeutic potential of SCFAs in depression and provided a theoretical foundation for their clinical application in antidepressant treatments.
Keywords: 16SrRNA sequencing, depression-like behaviors, gut inflammation, antidepressants
Introduction
Major depressive disorder (MDD), a complex mental disorder characterized by persistent depressed mood, diminished interest, and anhedonia, affected an estimated 279 million individuals globally in 2019.1 It’s global prevalence has continued to exhibit a significant upward trend in recent years.2 The pathogenesis of depression is multifactorial.3 Among them, dysbiosis of the gut microbiota can cause the occurrence of MDD through the gut-brain axis.4,5 Furthermore, an increasing amount of evidence have shown that patients with MDD often suffer from oral symptoms such as periodontal disease and tooth decay.6–8 The oral microbiota can alter the composition of the gut microbiota through the gut-brain axis. It can be seen that the imbalance of oral-gut microbiota is closely related to the occurrence and development of MDD. The microbial-oral-gut-brain axis has gradually become the focus of studying the pathogenesis and potential treatment pathways of depression.
Under physiological conditions, the microbiota and the host maintain a dynamic symbiotic equilibrium. Disruption of microbial homeostasis may compromise systemic stability, thereby predisposing the organism to localized or systemic pathologies.9,10 Supplementation of the organism with metabolites helps to restore homeostasis of the host microbiota. Short-chain fatty acids (SCFAs), including acetate, propionate, and butyrate, are the final metabolites produced by the gut microbiota through the fermentation of dietary fibers. They are the energy sources for cells. Beyond their bioenergetic functions, SCFAs exert pleiotropic regulatory effects on critical physiological and pathophysiological processes, including host energy metabolism, endocrine secretion dynamics, and immunoinflammatory responses.11,12 Emerging evidence indicates that SCFAs are pivotal signaling mediators in the pathophysiology and progression of multiple neurodegenerative disorders, including MDD.13,14
In our study, we constructed a CUMS depressed rat model, observed its antidepressant effect by gavage of SCFAs, observed its regulation of oral-gut microbiota, and explored the potential effects of SCFAs in the treatment of depression, providing a better theoretical basis for the choice of SCFAs in antidepressant treatment.
Materials and Methods
Experimental Reagents, Animal Grouping and Drug Treatment
Sodium acetate 50 mg/(kg·d), sodium propionate 100 mg/(kg·d) and sodium butyrate 50 mg/(kg·d) were purchased from Sigma Company (USA). A 1:1:1 mixed solution of SCFAs was prepared based on rat weight 1 mL/100 g. Total 25 male adult SD rats with weight 180±20 g were purchased from Weitonglihua Experimental Animal Technology Co., Ltd. (Beijing, China). Rats were raised in SPF-grade animal rooms, and 25 SD rats were divided into three groups: Control group (n=8): no treatment was performed; CUMS group (n=10): before daily stress, normal saline was administered by gavage with normal saline; SCFAs group (n=7): before daily stress, the mixed solution of SCFAs was administered by gavage at 1 mL/100 g, and continuous administration was administered for 28 days.
CUMS Model Construction
In the model preparation combined with single-cage solitary breeding, rats experienced chronic and unpredictable mild stimulation, and simulated patients with depressive disorders became ill under different stress stimuli. Stress methods included hot and cold stimulation, light and dark stimulation, bound stimulation, noise stimulation, tail stimulation, foreign body stimulation, humid lining, water, and fasting; one to two stress methods were randomly selected every day, and the stress methods were not repeated for 3 consecutive days, with a modeling duration of 28 days.15
Body Weight Measurement
The body weights of all rats were recorded daily to monitor longitudinal changes throughout the experimental period.
Behavioral Test
Behavioral tests were performed after the modeling and drug administration was completed. Before the test, all rats were allowed to adapt to the behavioral laboratory for 2 hours and returned to their original mouse cages after the experiment was completed. The weight and behavioral tests of the rats were conducted in a double-blind manner: The experimenters assigned the rats random numbers, and the behavioral scores were evaluated by researchers who were unaware of the groups.
Sucrose Preference Test (SPT)
The entire experiment lasted three days: on the first day, rats were raised in a single cage, and two bottles of equal amounts of 1% sucrose water were placed in each cage; on the second day, two bottles of equal amounts of drinking water were replaced for adaptive training; on the third day, one bottle of drinking water and one bottle of 1% sucrose water were placed in each cage, and the water bottle position was exchanged for 12 h to avoid position preference. After 24 h, sugar water and drinking water consumption were recorded, and the sugar water preference coefficient was calculated (sweet water consumption/total liquid consumption × 100%).
Open Field Test (OFT)
The activity site was an open spontaneously movable box (100×100×40 cm), and the rats were placed in a box. The OFT-100 rat opening activity experimental system was purchased from Taimeng Software Co., Ltd. (Chengdu, China), infrared cameras were used to record the number of rearings, total distance, and distance in the outer area of the rats in the box within 5 min.16
Collection of Oral Microbiota and Fecal Specimens From Rats
Using the cotton swab method, the head of the swab was placed in full contact with the buccal mucosa, tongue, and floor of the mouth of the rat for 30s. The swab was cut off and the swab head was placed in a sterilized Eppendorf tube. After the behavioral tests, the rats were placed in clean cages lined with sterilized disinfectant filter paper. Fecal samples were collected immediately after defecation. All specimens were rapidly cryopreserved at −80°C.
Oral and Gut Microbiota 16SrRNA Sequencing and Bioinformatics Bata Analysis
16S rRNA belongs to second-generation sequencing and can conduct sequencing analysis on the V3-V4 two variable regions. It can only be used for species identification analysis at the genus level. The DNA was extracted using a genomic DNA extraction kit. To ascertain the molecular size, the isolated DNA was subjected to 0.8% agarose gel electrophoresis and a Nanodrop was used to quantify the DNA. The V3-V4 variable region sequences of the 16S rRNA gene of bacteria were targeted, and the primers 338F (5′-ACTCCTACGG GAGGCAGCA-3′) and 806R (5′-GGACTACHVGG GTWTCTAAT-3′) were used to prepare the PCR amplification and sequencing library using the Illumina NovaSeq PE250. The samples were retested and supplemented after the original high-throughput sequencing data were filtered based on sequence quality.
Analyses were performed using the QIIME2 software, including the removal of repeat sequences and clustering. Representative sequences were annotated based on the Greengenes database and the taxonomic information of all ASVs in each specimen was recorded. Based on the distribution of ASV in different samples, the alpha diversity level of each sample was evaluated, and a beta diversity analysis was performed using the principal coordinate analysis (PCoA) method based on the Bray-Curtis distance matrix. The LEfSe analysis method was used to identify different bacterial genera.
HE Staining of Colon Tissue
Pentobarbital sodium was purchased from Ekear Biotechnology Co., Ltd. (Shanghai, China). The rats were euthanized by intraperitoneal injection of 6% pentobarbital sodium. Carefully observe the rat for 60 seconds to confirm that its breathing and heartbeat have stopped and there was no reflex, then quickly performed the dissection. The rat colon tissues were separated, and a portion of the tissues was fixed with 4% paraformaldehyde and subsequently embedded in paraffin to make sections (5 μm). Colonic tissues were dissected from rat specimens and fixed in 4% paraformaldehyde. After dewaxing and dehydration, hematoxylin and eosin (H&E) staining was performed, and pathological changes in the colon tissue were observed under an optical microscope.
Immunohistochemical Staining and Analysis of Tight Junction Proteins in Colon Tissue
A ZO-1 immunohistochemistry kit was purchased from Bioss Biotechnology Co., Ltd. (Beijing, China). The tissue was repaired with an antigen repair solution, cooled, and a drop of goat serum was added for mounting. Subsequently, ZO-1 (diluted at a ratio of 1:1000) was added and incubated overnight at 4°C. The next day, the corresponding secondary antibody (dilution ratio of 1:5000) was added. The samples were then incubated in the dark for 2 h. DAB staining was performed, followed by hematoxylin staining, gradient ethanol dehydration, xylene clearing, and neutral resin mounting. The positive staining area of ZO-1 was observed under a microscope, and the percentage of the positive staining area was calculated using ImageJ software.
Statistical Analysis
The IBM SPSS22.0 software, version 27.0 (NY, USA) was used for the statistical analysis. When the weight and behavioral test results of the three groups of rats met the normal distribution and homogeneity of variance, a one-way analysis of variance was used. Differences were considered statistically significant at P < 0.05.
Results
Body Weight Changes
Prior to model establishment, no statistically significant differences in body weights were observed among the three groups (Figure 1A). After the administration was completed, there were significant differences in body weight between the Control and CUMS groups (P < 0.0001), between the Control and SCFAs groups (P < 0.0001), and between the CUMS and SCFAs groups (P = 0.0057). Specifically, the body weight of the CUMS group was significantly lower than that of the Control group, whereas the body weight of the rats increased after SCFA administration (Figure 1B).
 |
Figure 1 The results of body weight and behavioral tests in three groups of rats. (A) Baseline weight of the three groups of rats; (B) Body weight of the three groups of rats after intervention; (C) Sucrose preference of the three groups of rats; (D) Number of rearing times in the three groups of rats; (E) Total distance of the three groups of rats; (F) Distance in outer area of the three groups of rats. (*P<0.05,** P<0.01, ***P<0.001,****P<0.0001). |
Behavioral Test Results
SPT
Compared to the Control group, the sugar water preference coefficient of the CUMS group decreased significantly (P = 0.00017). Compared to the CUMS group, the coefficient of the SCFAs group increased significantly (P = 0.00807) (Figure 1C). The results indicated that the CUMS model rats exhibited a lack of pleasure, while intervention with SCFAs alleviated this symptom.
OFT
The number of rears in the CUMS group was significantly lower than that in the Control group (P=0.0002). Compared with the Control group, the SCFAs group showed differences (P = 0.00479).
Compared with the CUMS group, the SCFAs group had an increasing trend, however, there was no statistical significance (Figure 1D); In terms of total distance, there were significant differences between the CUMS group (P < 0.0001) and the SCFAs group (P = 0.00286) compared to the Control group. Moreover, there was a significant difference between the CUMS and SCFAs groups (P = 0.00018) (Figure 1E). In terms of distance in the outer area, the CUMS group (P<0.0001) and SCFAs (P=0.00259) was significantly different from the Control group, and there was also a significant difference between the CUMS and SCFAs groups (P=0.01079) (Figure 1F). The results of the OFT experiment showed that CUMS-depressed rats had obvious depression-like behavior, mainly manifested in the reduction in the number of rearings, total distance, and distance in the outer area, while SCFAs intervention improved this symptom.
The above data can be found in Table 1.
 |
Table 1 The Results of Body Weight and Behavioral Tests in Three Groups of Rats |
Gut Microbiota Analysis
Gut Microbiota Alpha and Beta Diversity Analysis
Alpha diversity analysis revealed statistically significant differences (Chao1 index, P = 0.00025; observed species index, P = 0.00035) in the gut microbiota of the three groups of rats. The values of both indices were significantly lower than those of the Control group. However, in the SCFAs group, both the index values increased (Figure 2A). Beta diversity analysis revealed a significant separation trend among the samples from the three groups, indicating that there were significant differences in the gut microbiota among the Control, CUMS, and SCFAs groups (Figure 2B).
 |
Figure 2 Alpha and Beta diversity analysis of the gut microbiota. (A) Alpha diversity analysis of gut microbiota across the three experimental rat groups; (B) Beta diversity analysis of gut microbiota across the three experimental rat groups. (G stands for gut microbiota). (*P<0.05,***P<0.001). |
Analysis of Oral and Gut Microbiota Composition
There were differences in the composition of the oral and gut microbiota among the three groups of rats. At the phylum level, within the gut microbiota, the CUMS group exhibited relative abundances of 88.31% for Firmicutes and 8.11% for Bacteroidetes, whereas the SCFAs group demonstrated proportions of 85.13% and 11.61%, respectively. Compared to the CUMS group, the SCFAs group exhibited a reduction in the Firmicutes/Bacteroidetes (F/B) ratio (Figure 3A). At the genus level, Lactobacillus constituted the predominant bacterial genus in the gut microbiota. Its relative abundance was significantly higher in the SCFAs group (34.50%) than that in the CUMS group (23.64%) (Figure 3B). Within the oral microbiota, Rothia emerged as the dominant genus with a significantly higher relative abundance in the SCFAs group (29.07%) than in the CUMS group (19.26%) (Figure 3C).
 |
Figure 3 Gut and oral microbiota composition analysis. (A) Composition and relative abundance of gut microbiota at the phylum level across the three rat groups; (B) Composition and relative abundance of gut microbiota at the genus level across the three rat groups; (C) Composition and relative abundance of oral microbiota at the genus level across the three rat groups; (G stands for gut microbiota and O stands for oral microbiota). |
Differential Species Analysis
LEfSe analysis of the gut microbiota identified 20 characteristic species with significant differences (LDA > 1, Figure 4A). Among these, six species were enriched in the SCFAs group: Streptococcus, Collinsella, Marvinbryantia, Sphingobium, Bifidobacterium, Marivita; nine species were enriched in the CUMS group: Turicibacter, Enterococcus, Bacteroides, Helicobacter, Paenibacillus, Butyricimonas, Massilia, Phenylobacterium, Anaeroplasma; and five species were enriched in the Control group: rc4_4, Lactococcus, Sutterella, Aerococcus, Psychrobacter.
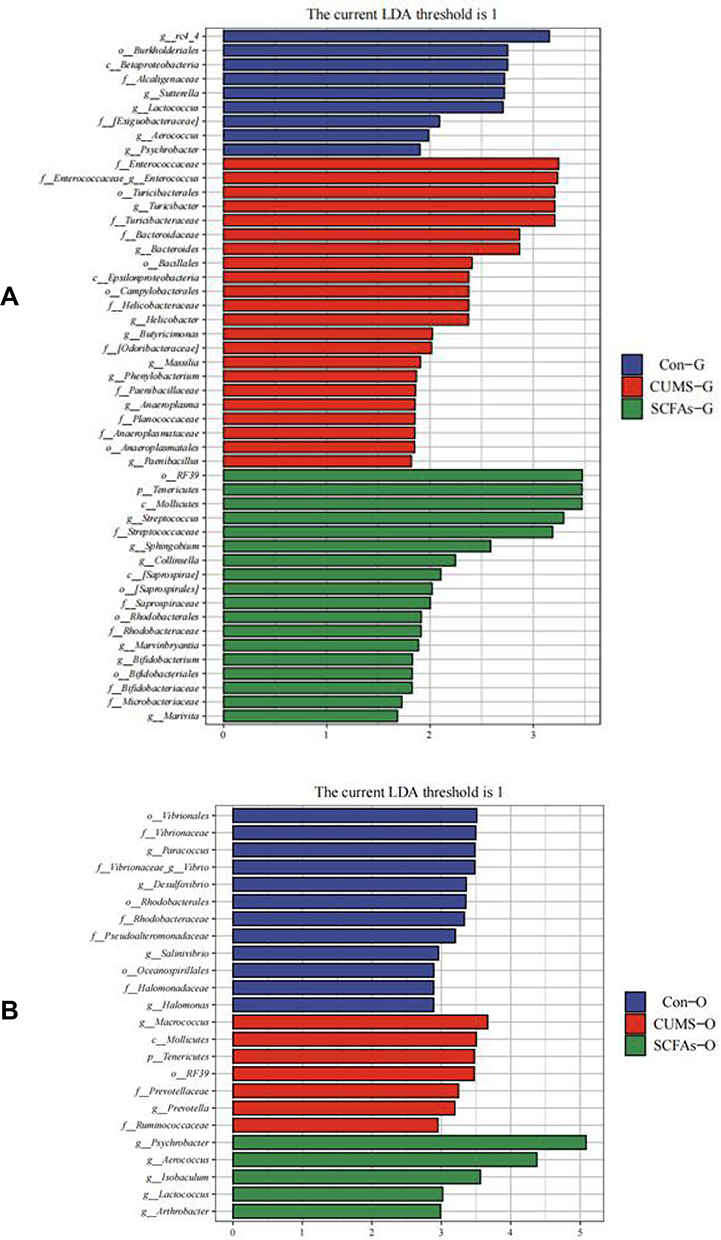 |
Figure 4 Taxonomic biomarkers found by LEfSe in three groups. (A)Taxonomic biomarkers found by LEfSe in gut microbiota; (B) Taxonomic biomarkers found by LEfSe in oral microbiota. (G stands for gut microbiota and O stands for oral microbiota). |
LEfSe analysis of the oral microbiota identified 12 characteristic species with significant differences (LDA > 1; Figure 4B). Among these, five species were enriched in the SCFAs group: Psychrobacter, Aerococcus, Isobaculum, Arthrobacter, Lactococcus; two species were enriched in the CUMS group: Macrococcus, Prevotella; and five species were enriched in the Control group: Vibrio, Paracoccus, Desulfovibrio, Halomonas, Salinivibrio.
Pathological Changes and the Expression of the Tight Junction Protein ZO-1 in Colon Tissue
In the Control Group, the colonic specimens demonstrated structural integrity with well-demarcated stratification of the mucosa, submucosa, muscularis propria, and serosa. Colonic mucosal epithelial cells were closely arranged, the number of goblet cells was normal, and no obvious infiltration of inflammatory cells was observed (Figure 5A). In the CUMS Group, the number of goblet cells in the colonic mucosa decreased and a large number of inflammatory cells were observed in the lamina propria (Figure 5B). In the SCFAs group, the number of goblet cells and mucus secretion increased, and inflammatory cell infiltration decreased (Figure 5C).
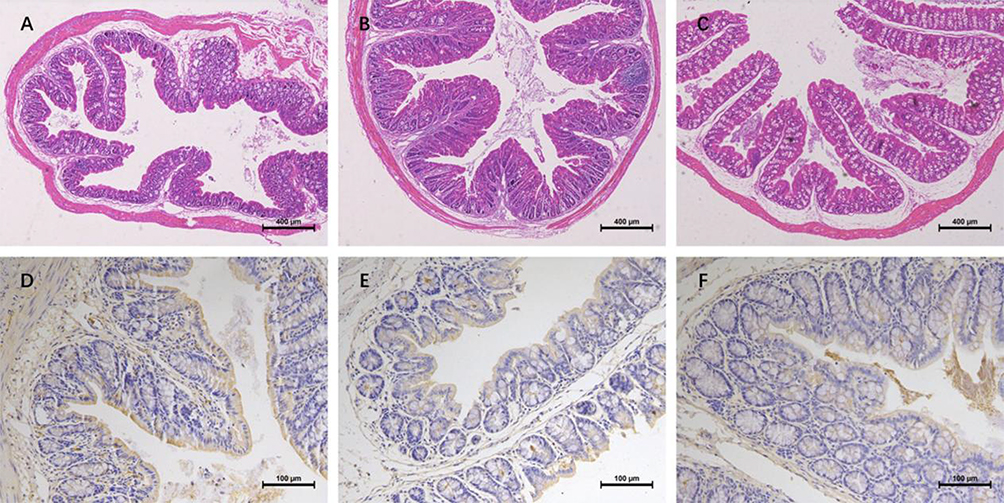 |
Figure 5 HE staining in colon tissue and Immunohistochemical staining of tight junction proteins. (A) HE staining of colon tissues in rats from Control group; (B) HE staining of colon tissues in rats from CUMS group; (C) HE staining of colon tissues in rats from SCFAs group (×5 magnification); (D) Detection of positive expression of ZO-1 in colon tissues of rats from Control group; (E) Detection of positive expression of ZO-1 in colon tissues of rats from CUMS group; (F) Detection of positive expression of ZO-1 in colon tissues of rats from SCFAs group (×20 magnification). |
The immunohistochemical results showed that, in the Control group, continuous positive signals of the tight junction protein ZO-1 were evenly distributed between the colonic epithelial cells. The percentage of positively stained areas was 31.32% (Figure 5D). In the CUMS group, the positive signal was weakened and the tight junction structure was loose. The percentage of positively stained areas was 24.27% (Figure 5E). The positive signal continuity in the SCFAs group improved continuously, the tight junction structure became more compact, and the percentage of positively stained area increased to 28.60% (Figure 5F).
Discussion
The oral cavity is the pivotal organ for mastication, digestion, and food deglutition. Oral saliva harbors a substantial microbial load, which is subsequently translocated to the gastrointestinal tract via swallowing, thereby establishing extensive crosstalk and interconnection between the oral and gut microbiota. The oral cavity and the intestinal tract are the main habitats of the human microbiome. The oral gut microbiota can influence diseases through various pathways such as immunity, metabolism, and endocrinology. Consequently, dysbiosis within either the oral or gut microbiota significantly influences the disease initiation, progression, and clinical outcomes.17 The central nervous system can establish a bidirectional regulatory effect between the brain and the oral-gut microbiota through various pathways such as nerves, immunity, and endocrinology.18 MDD is a chronic mental disorder that seriously threatens human health. It imposes a heavy burden on patients themselves, their families, and society as a whole, and has become a widespread public health issue of concern in today’s society. Dysbiosis of the oral gut microbiota is closely associated with the occurrence and development of MDD.6,8
The behavioral results of this study suggest that SCFAs can effectively alleviate depression-like behaviors in CUMS-induced depressed rats, suggesting that SCFAs may represent a potential option for treating depression.
The Influence of SCFAs on the Oral-Gut Microbiota
16SrRNA sequencing revealed differences in the Alpha and Beta diversity of the gut microbiota among the three rat groups. In the alpha diversity analysis, compared to the Control group, the number of species in the gut microbiota of rats in the CUMS group decreased, whereas the number of species increased in the SCFAs group. The greater the diversity of microbiota, the more beneficial it is to the health of the host. In our study, the diversity of the gut microbiota in the SCFAs group was greater than that in the CUMS group; this result was consistent with that of Mann’s study.19
Differences were recorded in the composition of the oral and gut microbiota among the three groups of rats. At the phylum level within the gut microbiota, SCFA intervention significantly reduced the relative abundance of Firmicutes while elevating Bacteroidetes representation, consequently diminishing the F/B ratio relative to the CUMS group. At the genus level, Lactobacillus constituted 23.34% of the gut microbiota in the CUMS group, which increased to 34.50% in the SCFAs group. The research team from the University of Virginia discovered that chronic stress stimulation would lead to a significant reduction in Lactobacillus in the gut microbiota of mice, and a significant decrease in the level of cytokine IFN-γ in the mouse serum.20 Lactobacillus can alleviate depression caused by chronic stress by maintaining the level of IFN-γ. Maintaining Lactobacillus colonization and a stable level of IFN-γ is necessary to prevent and treat MDD. In our study, after intervention with SCFAs, the proportion of Lactobacillus in the gut microbiota increased, which was beneficial for alleviating depression. Concurrently, in the oral microbiota, at the genus level, compared with the CUMS group, the abundance of Rothia increased in the SCFAs group. Rothia can maintain the balance of oral microbiota. They occupy specific sites and prevent harmful exogenous bacteria from colonizing the oral mucosa and surface of the teeth, making it difficult for harmful bacteria to attach. Rothia’s metabolites can regulate pH of the oral environment. The alkaline substances produced can neutralize acidic substances, inhibit the growth of acidophilic bacteria, prevent the oral environment from becoming overly acidic, and reduce the risk of tooth decay. Furthermore, Rothia, the resident bacteria in the oral cavity, can stimulate the oral immune system, keeping it in an alert state, enhancing the local immune defense of the oral cavity, improving the body’s ability to recognize and eliminate pathogens, maintaining oral immune homeostasis, and preventing the occurrence of inflammatory diseases.21
LEfSe analysis of the gut microbiota revealed that, at the genus level, in the SCFAs group, the relative abundances of Bifidobacterium, Collinsella, and Sphingobium increased. As an important probiotic for the human body, Bifidobacterium can produce SCFAs, inhibit intestinal immune inflammatory responses, restore intestinal barrier function, and regulate systemic immunity.22,23 The probiotics that produce SCFAs primarily include Lactobacillus, Bifidobacterium and Butyricum. Previous studies have shown that bifidobacteria can produce the neurotransmitter homovanillic acid, which inhibits autophagic cell death and protects hippocampal neurons, thereby alleviating MDD.24 The analysis of LEfSe in the oral microbiota revealed that the relative abundance of Prevotella increased in the CUMS group and decreased in the SCFAs group. Prevotella infections are associated with periodontitis and periapical periodontitis. They can enter the bloodstream by releasing pro-inflammatory factors IL-6 and TNF-α, thereby exacerbating the systemic inflammatory response. Könönen et al discovered that the Prevotella genus in the oral cavity may increase the risk of depression by reducing the conversion of nitrate to nitric oxide, leading to decreased neural plasticity and exacerbated neural inflammation.25 This finding is consistent with the results of our study.
The Effects of SCFAs on Intestinal Tissues
The intestinal barrier is an important physiological defense mechanism that regulates and maintains homeostasis of the internal environment of the body. It mainly functions through mechanical barriers composed of tight junction proteins and intestinal epithelial cells, immune barriers consisting of immune cells and cytokines, microbial barriers based on the gut microbiota, and chemical barriers, including mucins and antimicrobial peptides.26 Maintaining the integrity of the intestinal barrier is vital for body health. Tang et al discovered that the gut microbiota of depressed mice was imbalanced, and the tight junction proteins of the intestinal barrier were disrupted, resulting in complete damage to the intestinal barrier and systemic inflammatory responses.27 Our study found that in the CUMS group, the ZO-1 tight junction protein of the colon tissue was loose, resulting in impaired intestinal barrier function. Harmful microorganisms, toxins, and metabolites enter systemic circulation, overactivate immune cells, and induce intestinal immune imbalance. Inflammatory cells infiltrating the lamina propria are also observed. In the SCFAs group, the intestinal tight junction structure became more compact, the intestinal barrier was restored, and the infiltration of inflammatory cells decreased. Th17/Treg cells play an important role in ameliorating intestinal inflammation.
Our study revealed that SCFA intervention induced compositional and diverse alterations in the oral gut microbiota of CUMS depressive rats. Two possible reasons for this scenario are as follows: First, the gut houses 70% of the host’s immune cells and is the largest immune organ. After SCFAs intervention, the abundance of beneficial bacteria in the gut increased, whereas that of harmful bacteria decreased. The regulation of T cell differentiation by the gut microbiota leads to a reduction in the production of Th17 cells and an increase in the production of Treg cells. The secretion of anti-inflammatory cytokines such as IL-10 is increased. Immune cells and factors diffuse through the circulatory system into the oral microenvironment, enhancing local immunity in the mouth. The oral microbiota gradually tended towards a stable state. Second, SCFAs can be absorbed into the bloodstream, pass through the blood-brain barrier, activate receptors in the brain, affect brain function, and thereby alleviate depression-like behaviors. They can also reach the mouth through the circulatory system, regulate the expression of antibacterial peptides in gingival junctional epithelial cells, reduce oral inflammation, and gradually balance the oral microbiota.
The oral gut microbiota can affect MDD through various pathways, such as immunity, metabolism, and endocrinology. Studies have shown that by supplementing with probiotics, it is possible to regulate the stability of the microbiome and promote a healthy metabolism, thereby improving MDD.20,28,29 However, since most of the live probiotics need to be stored at a low temperature of 2–8°C, they will lose their activity if the temperature exceeds 40°C, and also be eroded by various digestive fluids such as gastric acid and bile, resulting in a significant reduction in the number of some active probiotics. These probiotics cannot attach to or colonize the intestinal tract to perform their functions. Therefore, directly supplementing their metabolites, such as SCFAs, may be a better approach.
Limitation
The sample size of this study was small, and the dynamic effects of SCFAs on oral and gut microbiota were not observed. Furthermore, the lack of direct detection of inflammatory cytokines in serum/cerebrospinal fluid (such as IL-6, TNF-α, IFN-γ) or SCFAs levels is a deficiency.
Conclusion
Our study demonstrated that SCFAs can alleviate the depressive-like behaviors of CUMS rats, increase the number of beneficial bacteria in the gut, reduce gut inflammation, increase the expression of the intestinal tight junction protein ZO-1, enhance immunity, and gradually stabilize the oral and gut microbiota of CUMS-depressed rats. Given the potential antidepressant effects of SCFAs, its supplementation may be an effective strategy for treating depression.
Ethical Statement
This study complied with the Chinese Guidelines for Ethical Review of Laboratory Animals (GB/T 35892-2018) and was approved by the Medical Ethics Committee of Fenyang College, Shanxi Medical University (Approval No. 2024016).
Acknowledgments
We sincerely thank all the participants of this research for their significant contributions to the project.
Funding
This study was financially supported by the Science and Technology Innovation Project of Higher Education Institutions in Shanxi Province (Grant No. 2023L477) and the Research Project of Fenyang College of Shanxi Medical University (Grant No. 2025A08).
Disclosure
All authors hereby declare that they have no conflicts of interest regarding this study.
References
1. GBD 2019 Mental Disorders Collaborators. Global, regional, and national burden of 12 mental disorders in 204 countries and territories, 1990-2019: a systematic analysis for the global burden of disease study 2019. Lancet Psychiatry. 2022;9(2):137–150. doi:10.1016/S2215-0366(21)00395-3
2. Wang L, Zhou L, Zhu Y, et al. Trends in prescription therapy for adolescents with depression in nine major areas of China during 2017-2021. Front Psychiatry. 2023;14:1175002. doi:10.3389/fpsyt.2023.1175002
3. Cui L, Li S, Wang S, et al. Major depressive disorder: hypothesis, mechanism, prevention and treatment. Signal Transduct Target Ther. 2024;9(1):30. doi:10.1038/s41392-024-01738-y
4. Teng T, Huang F, Xu M, et al. Microbiota alterations leading to amino acid deficiency contribute to depression in children and adolescents. Microbiome. 2025;13(1):128. doi:10.1186/s40168-025-02122-w
5. Zhu S, Li X, Yu Y, et al. EZH2-mediated H3K27me3 links microbial inosine loss to depression: a gut-brain epigenetic switch. Theranostics. 2025;15(18):9969–9986. doi:10.7150/thno.120824
6. Zeng Y, Jia X, Li H, et al. Oral microbiota among treatment-naïve adolescents with depression: a case-control study. J Affect Disord. 2025;375:93–102. doi:10.1016/j.jad.2025.01.089
7. O’Neil A, Berk M, Venugopal K, Kim SW, Williams LJ, Jacka FN. The association between poor dental health and depression: findings from a large-scale, population-based study (the NHANES study). Gen Hosp Psychiatry. 2014;36(3):266–270. doi:10.1016/j.genhosppsych.2014.01.009
8. Wang G, Cao L, Li S, et al. Gut microbiota dysbiosis-mediated ceramides elevation contributes to corticosterone-induced depression by impairing mitochondrial function. NPJ Biofilms Microbiomes. 2024;10(1):111. doi:10.1038/s41522-024-00582-w
9. de Vos WM, Tilg H, Van Hul M, Cani PD. Gut microbiome and health: mechanistic insights. Gut. 2022;71(5):1020–1032. doi:10.1136/gutjnl-2021-326789
10. Barlow JT, Leite G, Romano AE, et al. Quantitative sequencing clarifies the role of disruptor taxa, oral microbiota, and strict anaerobes in the human small-intestine microbiome. Microbiome. 2021;9(1):214. doi:10.1186/s40168-021-01162-2
11. Blaak EE, Canfora EE, Theis S, et al. Short chain fatty acids in human gut and metabolic health. Benef Microbes. 2020;11(5):411–455. doi:10.3920/BM2020.0057
12. Li L, Ma L, Fu P. Gut microbiota-derived short-chain fatty acids and kidney diseases. Drug Des Devel Ther. 2017;11:3531–3542. doi:10.2147/DDDT.S150825
13. Chang L, Wei Y, Hashimoto K. Brain-gut-microbiota axis in depression: a historical overview and future directions. Brain Res Bull. 2022;182:44–56. doi:10.1016/j.brainresbull.2022.02.004
14. Dalile B, Van Oudenhove L, Vervliet B, Verbeke K. The role of short-chain fatty acids in microbiota-gut-brain communication. Nat Rev Gastroenterol Hepatol. 2019;16(8):461–478. doi:10.1038/s41575-019-0157-3
15. Song N, Liu Z, Gao Y, Lu S, Yang S, Yuan C. NAc-DBS corrects depression-like behaviors in CUMS mouse model via disinhibition of DA neurons in the VTA. Mol Psychiatry. 2024;29(5):1550–1566. doi:10.1038/s41380-024-02476-x
16. Tang M, Jiang P, Li H, et al. Antidepressant-like effect of n-3 PUFAs in CUMS rats: role of tPA/PAI-1 system. Physiol Behav. 2015;139:210–215. doi:10.1016/j.physbeh.2014.11.054
17. Cheung MK, Tong S, Wong M, et al. Extent of oral-gut transmission of bacterial and fungal microbiota in healthy chinese adults. Microbiol Spectr. 2023;11(1):e0281422. doi:10.1128/spectrum.02814-22
18. Ochoa-Repáraz J, Kasper LH. The microbiome and neurologic disease: past and future of a 2-way interaction. Neurotherapeutics. 2018;15(1):1–4. doi:10.1007/s13311-018-0604-9
19. Mann ER, Lam YK, Uhlig HH. Short-chain fatty acids: linking diet, the microbiome and immunity. Nat Rev Immunol. 2024;24(8):577–595. doi:10.1038/s41577-024-01014-8
20. Merchak AR, Wachamo S, Brown LC, et al. Lactobacillus from the altered schaedler flora maintain IFNγ homeostasis to promote behavioral stress resilience. Brain Behav Immun. 2024;115:458–469. doi:10.1016/j.bbi.2023.11.001
21. Tsuzukibashi O, Uchibori S, Kobayashi T, et al. Isolation and identification methods of Rothia species in oral cavities. J Microbiol Methods. 2017;134:21–26. doi:10.1016/j.mimet.2017.01.005
22. Young SL, Simon MA, Baird MA, et al. Bifidobacterial species differentially affect expression of cell surface markers and cytokines of dendritic cells harvested from cord blood. Clin Diagn Lab Immunol. 2004;11(4):686–690. doi:10.1128/CDLI.11.4.686-690.2004
23. Chen J, Chen X, Ho CL. Recent development of probiotic bifidobacteria for treating human diseases. Front Bioeng Biotechnol. 2021;9:770248. doi:10.3389/fbioe.2021.770248
24. Zhao M, Ren Z, Zhao A, et al. Gut bacteria-driven homovanillic acid alleviates depression by modulating synaptic integrity. Cell Metab. 2024;36(5):1000–1012.e6. doi:10.1016/j.cmet.2024.03.010
25. Könönen E, Fteita D, Gursoy UK, Gursoy M. Prevotella species as oral residents and infectious agents with potential impact on systemic conditions. J Oral Microbiol. 2022;14(1):2079814. doi:10.1080/20002297.2022.2079814
26. Suzuki T. Regulation of the intestinal barrier by nutrients: the role of tight junctions. Anim Sci J. 2020;91(1):e13357. doi:10.1111/asj.13357
27. Tang E, Hu T, Jiang Z, et al. Isoquercitrin alleviates lipopolysaccharide-induced intestinal mucosal barrier damage in mice by regulating TLR4/MyD88/NF-κB signaling pathway and intestinal flora. Food Funct. 2024;15(1):295–309. doi:10.1039/d3fo03319h
28. Liu P, Liu Z, Wang J, et al. Immunoregulatory role of the gut microbiota in inflammatory depression. Nat Commun. 2024;15(1):3003. doi:10.1038/s41467-024-47273-w
29. Nikolova VL, Smith M, Hall LJ, Cleare AJ, Stone JM, Young AH. Perturbations in gut microbiota composition in psychiatric disorders: a review and meta-analysis. JAMA Psychiatry. 2021;78(12):1343–1354. doi:10.1001/jamapsychiatry.2021.2573
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.