Back to Journals » Journal of Inflammation Research » Volume 18
Anti-PS IgG Immune Complexes Impair Macrophage Phagocytosis in SLE via LOX-Dependent Oxidative Stress
Authors Guan H, Huang L, Liu Y, Zhu E, Chen L, Li W, Wu H, Zhang X, Qin R, Zheng J ![]() , Mo Y, Zhong M, Xu B, Dai X, Wei Q, Chen Y, Wang Q, Zheng Z, Ma K, Tang C
, Mo Y, Zhong M, Xu B, Dai X, Wei Q, Chen Y, Wang Q, Zheng Z, Ma K, Tang C ![]()
Received 12 March 2025
Accepted for publication 9 August 2025
Published 22 August 2025 Volume 2025:18 Pages 11521—11538
DOI https://doi.org/10.2147/JIR.S527859
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 5
Editor who approved publication: Dr Tara Strutt
Hui Guan,1– 3,* Liqi Huang,1,* Yu Liu,1,* Enyi Zhu,4 Lefeng Chen,5 Weijie Li,2 Haiqi Wu,2 Xiaoying Zhang,6 Rencai Qin,2 Jingpeng Zheng,2 Yingqian Mo,5 Ming Zhong,1 Bihua Xu,7 Xiaoyan Dai,8 Qi Wei,2 Yunwei Chen,2 Qingwen Wang,7 Zhihua Zheng,1 Kongyang Ma,2 Chun Tang1
1Department of Nephrology, Center of Kidney and Urology, the Seventh Affiliated Hospital, Sun Yat-Sen University, Shenzhen, 518107, People’s Republic of China; 2Centre for Infection and Immunity Studies, School of Medicine, The Sun Yat-sen University, Shenzhen, 518107, People’s Republic of China; 3Department of Radiation Oncology, The First Affiliated Hospital of Zhengzhou University, Zhengzhou, 450052, People’s Republic of China; 4The Division of Nephrology, The Sixth Affiliated Hospital, Sun Yat-Sen University, Guangdong Provincial Key Laboratory of Colorectal and Pelvic Floor Diseases, Guangzhou, 510000, People’s Republic of China; 5Department of Rheumatology, Sun Yat-sen Memorial Hospital of Sun Yat-Sen University, Guangzhou, 510120, People’s Republic of China; 6Health Management Center, the Seventh Affiliated Hospital, Sun Yat-Sen University, Shenzhen, 518107, People’s Republic of China; 7Department of Rheumatology and Immunology, Peking University Shenzhen Hospital, Shenzhen, 518036, People’s Republic of China; 8Clinical Research Institute, Hengyang Medical School, the second Affiliated Hospital, University of South China, Hengyang, 421002, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Chun Tang, Email [email protected] Kongyang Ma, Email [email protected]
Purpose: Systemic lupus erythematosus (SLE) is a severe autoimmune disease with systemic complications mediated by immune-complex formation. The elevated level of anti-phosphatidylserine (PS) IgG has been implicated in SLE pathogenesis. In this study, we aimed to explore the effector mechanisms of PS immune-complex during lupus development.
Patients and Methods: Serological profiles of immune-complexes in SLE patients were analyzed. Immunofluorescence staining showed PS-IgG immune-complex deposition in kidney biopsies of lupus nephritis patients. C57BL/6J mice were immunized with PS for immune-complex and renal function assessment. The roles of PS-IgG immune-complex and lysyl oxidase (LOX) were validated from SLE PBMCs, THP-1 cell line and PS-immunized lupus mice. The intracellular reactive oxygen species (ROS) levels, and phagocytosis function were examined by flow cytometry in SLE PBMCs, THP-1 cell line and PS-immunized lupus mice. For in vitro treatment, the effects of antioxidant N-acetylcysteine (NAC) and LOX inhibitor β-Aminopropionitrile (BAPN) were verified in THP-1 cell line and cells from PS-immunized lupus mice.
Results: SLE and lupus nephritis (LN) patients showed significant elevated circulating and glomerular PS-IgG immune-complex levels. ROC analysis indicated PS-IgG immune-complex as a strong biomarker in SLE and LN. Mechanistically, induced macrophages from SLE patients treated with PS-IgG immune-complex significantly increased cytoplasmic ROS levels, elevated LOX expression and exhibited dampened phagocytotic function. In mice, PS immunization triggered PS-IgG immune complex formation, increased LOX expression, immune-complex deposited glomerular nephritis, and impaired phagocytotic function of macrophages. NAC and BAPN treatment restored the phagocytotic function of human and murine macrophages.
Conclusion: Our results indicate that PS-IgG immune-complex can directly impair macrophage phagocytotic functions via LOX mediated-oxidative stress and may serve as a novel biomarker for SLE.
Keywords: systemic lupus erythematosus, anti-phospholipid antibodies, immune complex, macrophages
Introduction
Systemic lupus erythematosus (SLE) is a chronic systemic, autoimmune, inflammatory connective tissue disease with multiple organ damages that predominantly affects women of childbearing age.1 Renal damage is one key clinical feature in patients with SLE.2 Up to 40% of SLE patients progress to lupus nephritis (LN) with immune-complex deposited glomerular dysfunction, and 10–20% eventually progress to end-stage renal disease.3 However, the underlying mechanisms of immune-complex formation and deposition in LN patients remains partially understood.
Disruption of immune tolerance and activation of autoimmunity plays a key role in the pathogenesis of SLE.4 During SLE pathogenesis, autoantigens released by dead cells or neutrophil extracellular traps stimulate autoreactive B cell expansion and autoantibodies production.5,6 The formation and pathogenic effects of immune complex (IC) has been indicated in the pathogenesis of SLE.7 Recently, we reported the pathogenic role of B-1 cell-derived anti-phosphatidylserine (PS) IgG in SLE.6 However, the effector functions of PS-IgG IC during lupus development remains largely unclear. Macrophages in SLE exhibit a deficient phagocytic capacity, struggling to effectively clear apoptotic cells and immune complexes, which may lead to the accumulation of cellular debris and the initiation of autoimmune responses. They also oversecrete various pro-inflammatory cytokines like IL-1β, TNF-α, and IL-6, promoting inflammation and disease progression.8,9 M Herrmann et al reported the diminished phagocytotic function of macrophages in patients with SLE.10,11 Previous studies also indicated the clearance of anti-dsDNA IC in macrophages via FcgRI signaling pathway.12 However, the underlying mechanisms remain largely unclear. Herein, we aim to evaluate the role of ant-PS antibodies and PS-IgG IC plays in the macrophage phagocytosis dysfunction during lupus pathogenesis.
Reactive oxygen species (ROS) play a critical role in physiological signaling by acting as second messengers in processes such as vascular tone regulation, immune defense, and redox homeostasis, often generated by enzymes like NADPH oxidase and NO synthase.13 At low concentrations, ROS regulate cell proliferation, differentiation, and survival by modulating redox-sensitive pathways, including MAPK and NF-κB activation, while excessive ROS can induce oxidative stress and pathological conditions.14 Also, ROS play a crucial role in modulating the phagocytic function of macrophages. As key components of the innate immune system, macrophages utilize ROS to enhance their ability to engulf and destroy pathogens. Recent studies have highlighted the dual role of ROS in both promoting phagocytosis and maintaining the balance between antimicrobial activity and self-damage.15 Lysyl oxidase (LOX) is a copper-dependent enzyme that catalyzes the oxidative cross-linking of collagen and elastin in the extracellular matrix (ECM), essential for tissue integrity and elasticity.16,17 LOX is involved in the cross-linking of extracellular matrix proteins, and its activity can modulate the oxidative environment within cells. In autoimmune diseases (such as rheumatoid arthritis and systemic sclerosis), chronic inflammation leads to excessive cross-linking and fibrosis of the ECM, and the abnormal expression of LOX is the core driving factor of this process. LOX is regulated by TGF-β and feedback-enhances its signal, promoting fibroblast activation and Th17 cell differentiation, which is the key pathological mechanism of rheumatoid arthritis.17 These studies suggest that LOX may be involved in the pathogenesis of autoimmune diseases. Studies have indicated that LOX inhibitor BAPN can inhibit ROS production and leptin-induced profibrotic mediators.18 LOX was also reported to promote GBM progression by driving macrophage infiltration.19 Understanding the interplay between LOX-mediated ROS and macrophage dysfunction is crucial for developing targeted therapeutic strategies in inflammatory diseases, like SLE, which needs to be further explored.
Our study firstly identified the diagnostic values and underlying mechanism of the PS-IgG IC in SLE. Previous study highlighted the pathogenic roles of anti-dsDNA IgG in lupus nephritis, here, we proved the direct deposition of PS-IgG IC in renal glomeruli of SLE patients and animal models. The PS-IgG IC has been also indicated as a strong biomarker in SLE and LN by ROC analysis. Mechanically, PS-IgG IC in SLE induced-macrophages significantly increases the cytoplasmic ROS levels and LOX expression, and dampens phagocytotic function. Moreover, antioxidant N-acetylcysteine (NAC) and LOX inhibitor β-Aminopropionitrile (BAPN) dramatically restore the ROS levels and phagocytotic function of human and murine macrophages. Thus, PS-IgG immune-complex impaired macrophage phagocytotic functions via LOX medicated oxidative stress. Our study might provide a novel biomarker and guide the diagnosis and clinical treatment of SLE.
Materials and Methods
Patients
Patients meeting the 2019 European League Against Rheumatism/American College of Rheumatology criteria for SLE were recruited.20 Following patients were excluded from the study: (1) drug-induced SLE; (2) pregnant or lactating patients; (3) receiving high-dose glucocorticoid or plasma exchange therapy within six months; (4) receiving anti-B cell therapy (including Rituximab, Belimumab, and Telitacicept) within six months; (5) history of malignancy within five years (except cured basal cell carcinoma of the skin or carcinoma in situ of the cervix); (6) having the active infectious disease. Healthy volunteers were recruited as healthy controls (HC). Samples and information from SLE patients and HC were collected from the Seventh Hospital of Sun Yat-sen University, the Sun Yat-sen Memorial Hospital of Sun Yat-sen University, and the Shenzhen Hospital of Peking University. Peripheral blood mononuclear cells (PBMCs) and serum from SLE patients and HC were collected by density gradient centrifugation. Baseline data were collected from SLE patients, including age, gender, coagulation function, renal function, autoantibody levels, and complement levels. Disease activity was assessed according to the systemic lupus erythematosus disease activity index 2000 (SLEDAI-2000).21 A SLEDAI score of 0–4 indicated a stable disease, and a score greater than 4 indicated the active SLE.22 Briefly, forty-eight peripheral blood samples were collected from SLE patients. Of the patients, 43 (89.6%) were female and 5 (10.4%) were male with a mean age of 36 years. Among them, 29 (60.4%) were LN patients, 19 (39.6%) were patients with non-lupus nephritis. Additionally, 22 (45.8%) were in the active stage. Hematological analysis, including serum albumin, and hemoglobin levels. A total of 26 blood specimens were collected from healthy controls (HC), with a mean age of 40 years. Among them, 19 (73.1%) were female, and 7 (26.9%) were male. The characteristics of SLE patients and HC has been shown in Table S1. All experiments involving human samples were approved by the Ethics Committee of the Seventh Hospital of Sun Yat-sen University (KY-2021-097-01), the Ethics Committee of Sun Yat-sen Memorial Hospital of Sun Yat-sen University (SYSEC-KY-KS-2021-339) and the Ethics Committee of Shenzhen Hospital of Peking University (2021–043) and conducted in accordance with the Declaration of Helsinki. All patients and healthy controls have signed the informed consent form.
Animal Model
SLE predominantly affects women, with a ratio of about 9 women to 1 man.23 Therefore, we chose female mice as our animal model. Female C57BL/6J mice were housed in an SPF-grade environment at the Experimental Animal Center of Sun Yat-sen University. The animal experiments were conducted in compliance with ethical standards as approved by the Animal Experimental Ethics Committee of Sun Yat-sen University, under approval number SYSU-IACUC-2021-000882. Ethical guidelines were strictly followed during the entire experimental process, which included Laboratory animal—Guideline for ethical review of animal welfare (GB T 35892–2018), Laboratory animal—Guidelines for euthanasia (GB∕T 39760–2021), and Guidelines for the Ethical Termination Evaluation of Animal Experiments (RBT 173–2018). For mouse model generation, PS immunized mice and lupus mice were generated as previously.6,24 At week 16, macrophage aggregation to the peritoneal cavity in PS-immunized mice by intraperitoneal injection of 4% sterile thioglycolic acid broth (BD Biosciences, 211716, USA). Mice were euthanized and blood, kidney, bone marrow, and peritoneal cells were harvested after three days. Serum creatinine levels were measured according to the manufacturer’s instructions (Nanjing Jiancheng, C011-2-1, China). Five to nine mice were used in each group, and mice were randomly assigned to experimental groups using a simple random sampling method, which was blind to the experimenters.
Serum Cytokines Assay
According to the manufacturer’s instructions, a human inflammation panel (BioLegend, 740809, USA) was used to detect serum cytokines in SLE patients, including monocyte chemoattractant protein-1 (MCP-1), interleukin-1β (IL-1β), interferon-α2 (IFN-α2), IFN-γ, tumor necrosis factor-α (TNF-α), IL-6, IL-8, IL-10, IL-12p70, IL-17A, IL-18, IL-23, and IL-33. All samples were examined using a BD LSR Fortessa flow cytometer (BD Biosciences, USA), and data were analyzed using LEGENDplex software (BioLegend, USA).
Enzyme-Linked Immunosorbent Assay (ELISA)
Anti-phosphatidylserine/prothrombin (PS/PT) antibody levels (J&L Biological, JL14171, JL48056, JL14166, China), anti-prothrombin (PT) antibody levels (J&L Biological, JL14185, JL14191, JL14197, China), and anti-dsDNA IgG levels (J&L Biological, JL13373-96T, China) in serum of SLE patients and HC (1:10 dilution) were detected using ELISA kits according to the manufacturer’s instructions. ELISA plates were coated with the antigen overnight at 4°C. Plates were incubated with phosphate buffer saline (PBS) containing 0.5% bovine serum albumin (BSA), 0.5% gelatin, and 0.5% Tween-20 for one hour to block the non-specific binding sites. PBS-diluted samples (1:10 dilution for humans, and 1:200 dilution for mice) were added and incubated for two hours. Then, horseradish peroxidase (HRP) conjugated antibodies were added and incubated for two hours. Next, tetramethylbenzidine (TMB) reagent was added and incubated for 4–30 min at 37°C. Finally, the chromogenic reaction was terminated with a stop solution. The optical density (OD) of each well at 450 nm was immediately recorded by Synergy H1 microplate reader (BioTek, Vermont, USA). Human anti-PS IgA/IgG/IgM antibodies were detected using BSA-PS antigen (Cloud-Clone, CPB881Ge11, China) with the addition of HRP goat anti-human IgA antibody/biotin anti-human IgG and HRP-streptavidin/biotin anti-human IgM and HRP-streptavidin. Similarly, mouse anti-PS IgG antibodies were detected using BSA-PS antigen with the addition of biotin anti-mouse IgG and HRP-streptavidin. Mouse anti-dsDNA IgG antibodies were detected by DNA antigen (Solarbio, D8020, China) combined with biotin anti-mouse IgG and HRP-streptavidin. For validating the serum levels of anti-PS IgG immune complexes, we used rabbit anti-PS coating antibody (Cloud-Clone Corp., Wuhan, China) for the binding of PS epitopes in serum PS-IgG IC, then we used the detecting antibodies of anti-human/mouse IgG to binding the IgG epitopes in serum PS-IgG IC by ELISA assay and confocal microscopy. The detailed flowchart for the validation of PS-IgG IC were shown in Figure S1.
Immunofluorescence Staining
Kidney biopsy specimens from LN patients and controls were collected for immunofluorescence staining was performed. For immune fluorescent staining, paraffin sections were stained by goat anti-human IgG primary antibody, followed by HRP rabbit anti-goat secondary antibody combined with CY3-TSA (Servicebio, G1223, China). PS staining was detected by rabbit anti-PS primary antibody, followed by HRP goat anti-rabbit secondary antibody combined with 488-TSA (Servicebio, G1231, China). Cell nuclei were stained with DAPI (Servicebio, G1012, China). For mouse kidney sections, rabbit anti-PS antibody combined with CY3-goat anti-rabbit labeled PS, Alexa Fluor 488 goat anti-mouse IgG labeled IgG were applied. Besides, primary antibody included F4/80 (MedChemexpress, HY-P80665, USA), LOX (Proteintech, 17958-1-AP, China), and secondary antibodies included Goat anti-Rat IgG (H+L) Cross-Adsorbed Secondary Antibody, Alexa Fluor™ 594 (Thermo Fisher, A11007, USA) and CoraLite488-conjugated Goat Anti-Rabbit IgG (H+L) antibody (Proteintech, SA00013-2, China). All images were captured by Pannoranmic microscopy (3Dhistech, Hungary) and analyzed using the ImageJ software (version 1.53k, USA). The PS and IgG co-localization/IgG positive area ratio represented the contribution of PS-IgG IC deposition.
Renal Pathological Staining
Paraffin sections of mouse kidneys were used for hematoxylin-eosin (HE) staining, and captured with a DM6B microscope (Leica, Germany). At least ten visible fields were randomly selected from each section to assess glomerular activity scores as reported previously.25
Cell Culture
PBMCs from SLE patients and mouse peritoneal macrophages were cultured in RPMI 1640 medium containing 10% fetal bovine serum and 1% penicillin/streptomycin antibodies. Monocytes differentiation from human peripheral blood to macrophages was induced using 25 ng/mL of macrophage-stimulating factor (M-CSF; Novoprotein, C417, China). THP-1 cell line was purchased from HyCyte Company (TCH-C361, Jiangsu, China) and cultured in a medium containing 10% fetal bovine serum, 1% penicillin/streptomycin, and 50 μm β-mercaptoethanol in RPMI 1640 medium. Phorbol 12-myristate 13-acetate (PMA; GLPBIO, GN10444, USA) was used for THP-1 differentiation. For PS-IgG IC treatment, plates were coated with sterile PS (50 μg/mL) overnight at 4°C, followed by washing with PBS. Plates were then coated with purified IgG from SLE patients (100 μg/mL) and incubated at 37°C for 2 h and then washed with PBS for three times to exclude the non-specific binding. N-acetylcysteine (NAC; Beyotime, ST1546, China) was used to scavenge intracellular ROS; β-Aminopropionitrile (BAPN; MedChemExpress, HY-Y1750, USA) was used to inhibit LOX.
Flow Cytometry
Single cell suspensions of PBMCs, THP-1 cells, kidney, peritoneal cavity, and bone marrow of mice were prepared. Dead cells were excluded using Zombie Aqua fixable viability kit (BioLegend, 423102, USA). TruStain FcX was added to block the non-specific binding. Cells were then labeled with fluorescence-conjugated antibodies as indicated (Table S2). DCFDA/H2DCFDA-cellular ROS assay kit (Abcam, ab113851, UK) was used to label intracellular ROS. MitoSpy (BioLegend, 424805, USA) and MitoSOX (ThermoFisher Scientific, M36008, USA) were used for labeling mitochondria and mitochondrial superoxide, respectively. PHrodo Red Zymosan Bioparticles (ThermoFisher Scientific, P35364, USA) were incubated with macrophages to assess the phagocytotic function. Data were collected using a CytoFLEX flow cytometer (Beckman Coulter, USA) and analyzed by FlowJo software (version 10.8.1, Tree Star, USA).
Serum IgG Purification
Serum IgG protein from SLE patients was purified by affinity chromatography as previously reported (Figure S2).26 Briefly, serum was loaded onto a Protein G agarose purification column (Beyotime, P2009, China). IgG protein was eluted with Glycine eluent (50 mmol/l, pH 2.7) and neutralized immediately with the neutral buffer and then dialyzed with PBS. Toxin Eraser endotoxin removal kit (Genscript, L00338, China) was used to remove the endotoxin from purified SLE IgG, and the protein was concentrated by ultrafiltration. Protein concentration was determined using a BCA kit (CWBIO, CW0014S, China). Then, the purified proteins were used for ELISA assay and cell culture.
RNA Sequencing
RNA preparation, library construction and Illumina PE150 sequencing were performed by IGE Biotechnology Institute (Guangzhou, China). The sequencing error rate of each base was obtained by converting the sequencing Phred value (Phred score, Qphred) through the formula Qphred = −10log10(e). The Phred value was calculated during the base identification process based on a model that predicts the probability of base discrimination errors. Under normal circumstances, the sequencing error rate at each base position should be lower than 0.5%. The low-quality sequences were filtered out by using tools such as FastQC and Trimmomatic, and the results were verified to meet the standards through re-examination. Statistical analysis was conducted on the original data and the high-quality data (clean reads) obtained through filtering, resulting in the basic information of the data. Clean reads data were aligned to the reference genome using the Hisat2 software, with the mapped rate greater than 96%. Differential gene expression was quantified by edgeR, and P < 0.05 and |log2FoldChange| >1 was used as the criteria for significance of differences. Distribution of differential genes was demonstrated by volcano plot. Enrichment analysis of differential genes was performed based on the Kyoto Encyclopedia of Genes and Genomes (KEGG, http://www.genome.jp/) database.
Western Blotting
Cell lysates were prepared by lysing cells in RIPA buffer containing protease and phosphatase inhibitors to prevent protein degradation. Protein concentrations were determined using the BCA Protein Assay Kit (NCM Biotech, WB6501, China). Samples were mixed with loading buffer, heated at 95°C for 5 minutes, and subjected to SDS-PAGE on a 10% acrylamide gel. Proteins were transferred to nitrocellulose membranes, which were blocked with 5% non-fat milk in TBST for 1 hour at room temperature to prevent nonspecific binding. Then incubate with primary antibodies for anti-LOX (Proteintech, 17958-1-AP, China) or anti-Vinculin (Proteintech, 66305-1-Ig, China) overnight at 4°C, followed by three washes with TBST. Secondary antibodies, including anti-rabbit IgG-HRP antibody (1:2000, ABclonal, AS014) or anti-mouse IgG-HRP antibody (1:2000, ABclonal, AS003) were applied for 1 hour at room temperature. After additional washes, bands were detected using a chemiluminescent substrate and imaged with a Bio-Rad ChemiDoc XRS+ System.
Quantitative Real-Time PCR
Total RNA was extracted from cells using the NcmSpin RNA Kit (NCM Biotech, M5105, China) according to the manufacturer’s protocol. The quality and concentration of the RNA were assessed using a NanoDrop spectrophotometer. Reverse transcription was performed using the Evo M-MLV RT Mix Kit with gDNA Clean for qPCR Ver.2 (Accurate Biology, AG11728, China) with oligo(dT) primers to synthesize cDNA from 1 µg of total RNA. Quantitative real-time PCR (RT-qPCR) was conducted using the SYBR Green Premix Pro Taq HS qPCR Kit (Accurate Biology, AG11701, China) on a QuantStudio 3 Real-Time PCR System. Primer sequences were designed as follows: LOX: forward 5’-TTCCTGCGCTCAGTAACCT-3’ reverse 5’-CAGGTCTGGGAGACCGTACT-3’; GAPDH: forward 5’-AGAAGGCTGG GGCTCATTTG-3’ reverse 5’-AGGGGCCATCCACAGTCTTC-3’. Relative gene expression was calculated using the 2^(-ΔΔCt) method with GAPDH as the reference gene.
Statistical Analysis
Mechanism diagram was created by Figdraw (Agreement number: OOUYI68a0d). All data in this study were shown as mean ± standard deviation (SD). The Mann–Whitney U-test was used to compare two groups of continuous variables with non-normal distribution in humans, and the Student’s t-test was used to compare two groups of continuous variables with normal distribution in mice. A one-way ANOVA test was used for comparing multiple groups. The Pearson correlation coefficient test was used to analyze the correlation of two continuous variables. Statistical results were plotted by GraphPad Prism software (version 9.0, GraphPad Software Inc., CA, USA). P > 0.05, no significance (ns); P < 0.05 was defined as a statistically significant difference. *P < 0.05; **P < 0.01; and ***P < 0.001.
Results
Elevated Circulating PS-IgG Immune Complex Levels in SLE Patients
The pathogenic roles of anti-phosphatidylserine IgG in human and murine lupus has been reported in our previous study.6 We then detected the levels of anti-PS IgG and PS-IgG IC in SLE patients. The ELISA results demonstrated elevated levels of anti-PS IgG and PS-IgG IC in SLE patients (Figure 1A and B), indicating the existence of circulating anti-PS IgG and PS-IgG IC formation in patients with SLE. Moreover, PS-IgG IC levels in SLE patients were positively correlated with anti-PS IgG levels and anti-dsDNA IgG levels, but negatively correlated with C4 levels (Figure 1C and E). Then, SLE patients were categorized into high and low-level anti-PS IgG subgroup based on the median serum anti-PS IgG levels. Further analysis on the comparison of clinical and laboratory indices between the two groups showed that the high-level anti-PS IgG subgroup had higher levels of total IgG, anti-PS IgA, anti-dsDNA IgG, SLEDAI, pyuria, PS-IgG IC, and lower levels of C3 and C4 than those in low-level anti-PS IgG subgroup (Table S3).
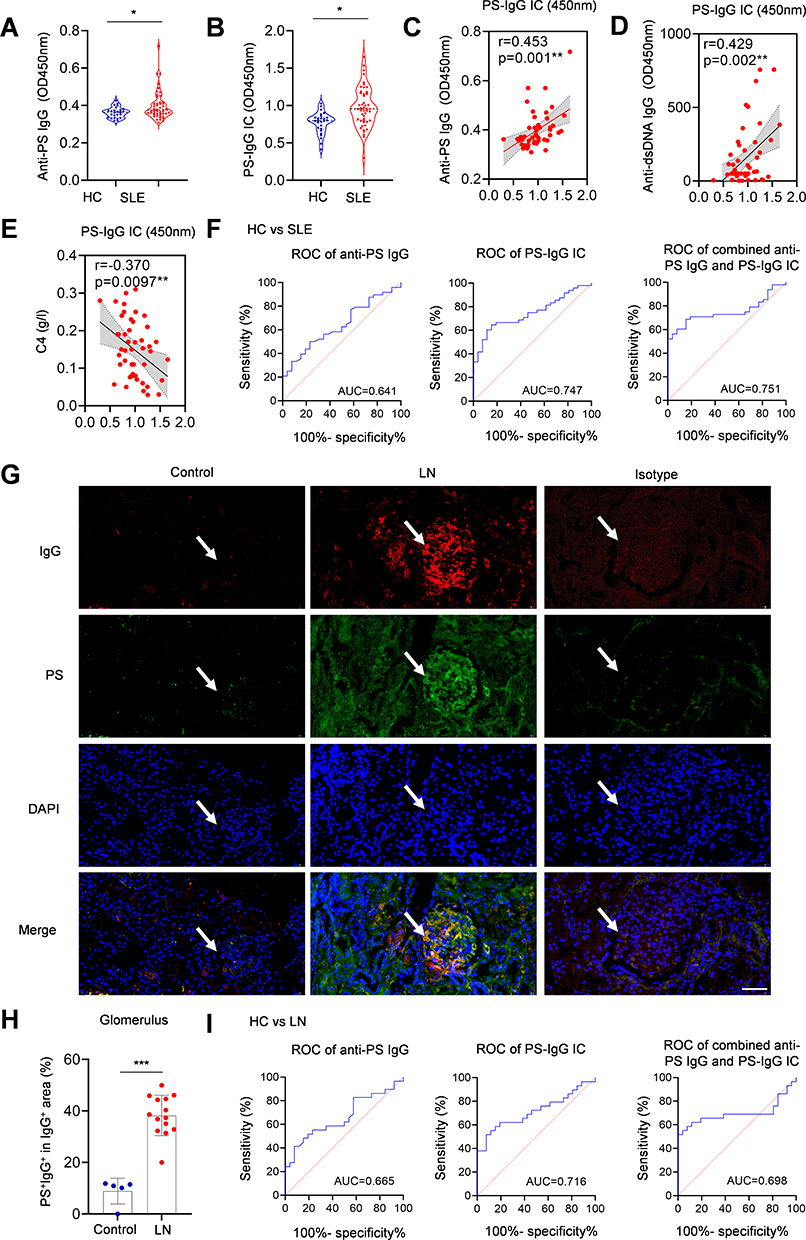 |
Figure 1 PS-IgG IC level was elevated in SLE and LN patients. (A) Anti-PS IgG and (B) PS-IgG IC levels in SLE patients and controls. Serum levels of PS-IgG IC and the correlation between serum PS-IgG IC level and (C) anti-PS IgG, (D) anti-dsDNA, and (E) C4 in SLE patients (n = 48). (F) ROC analysis of serum anti-PS IgG and PS-IgG IC for the diagnosis of SLE. (G) Deposition of PS-IgG IC in glomeruli of LN patients and controls (Scale bar, 20 mm); The white arrows indicate the glomerular area. (H) Comparison of glomerular deposition of PS and IgG colocalized region in LN patients (n = 14) and controls (n = 5). (I) ROC analysis of serum anti-PS IgG and PS-IgG IC for diagnosing of LN. Data were expressed as mean ± SD. *, P<0.05; ***, P <0.001. |
To evaluate anti-PS IgG and PS-IgG IC formation during lupus development, the anti-PS IgG and PS-IgG IC diagnostic effectiveness in SLE were analyzed using the receiver operating characteristic (ROC) curve. The area under the curve (AUC) for anti-PS IgG, PS-IgG IC, and their combination in diagnosing SLE was 0.641, 0.747, and 0.751, respectively (Figure 1F). PS-IgG IC showed higher AUC values than anti-PS IgG in the diagnosis of SLE.
Increased Glomerular Deposition of PS-IgG IC in LN Patients
To determine the role of PS-IgG IC in LN development, kidney biopsies from LN patients and controls were examined for PS-IgG IC deposition by assessing PS and IgG co-localization. Examination of immunofluorescent images showed the colocalization of PS and IgG staining, indicating the PS-IgG IC deposition in the glomeruli in LN patients (Figure 1G). Statistical analysis showed that PS-IgG IC deposition in the glomeruli was significantly more severe in LN patients compared with controls (Figure 1H). To evaluate the contributions of PS-IgG IC in LN development, the ROC method was used to analyze the efficacy of anti-PS IgG and PS-IgG IC for the diagnosis of LN. The AUC of anti-PS IgG, PS-IgG IC, and combination of both for the diagnosis of LN was 0.665, 0.716, and 0.698, respectively (Figure 1I). Interestingly, PS-IgG IC also showed higher AUC values than anti-PS IgG in the diagnosis of LN.
PS-IgG IC Impairs Macrophages Functions in SLE Patients
To determine the potential cells affected by PS-IgG IC in SLE, we first analyzed the relationship between PS-IgG IC levels and peripheral blood counts in SLE patients, which suggested that PS-IgG IC was negatively correlated with circulating monocyte counts (Figure 2A). No significant correlations were observed between circulating PS-IgG IC levels and leukocyte, lymphocyte, neutrophil, or platelet counts (Figure 2A). The SLE IgG were purified as indicated and PS-IgG IC were generated. Freshly prepared SLE PBMCs were then stimulated with vehicle control, PS antigen, IgG antibody, and PS-IgG IC, respectively. The numbers of CD14+ monocytes, CD4+ T cells, and CD19+ B cells were detected by flow cytometry. Results showed that PS-IgG IC stimulation could selectively decrease the frequencies of CD14+ monocytes in SLE PBMCs (Figure 2B and C), while the frequencies of CD4+ T cells and CD19+ B cells showed no significant changes (Figure S3A). Moreover, the correlation analysis between PS-IgG IC levels and peripheral cytokine levels in SLE patients suggested that PS-IgG IC was not significantly correlate with inflammatory cytokines such as MCP-1, IL-6, IL-8, and IL-10, suggesting that PS-IgG IC may affect monocyte function through a non-inflammatory pathway (Figure S4).
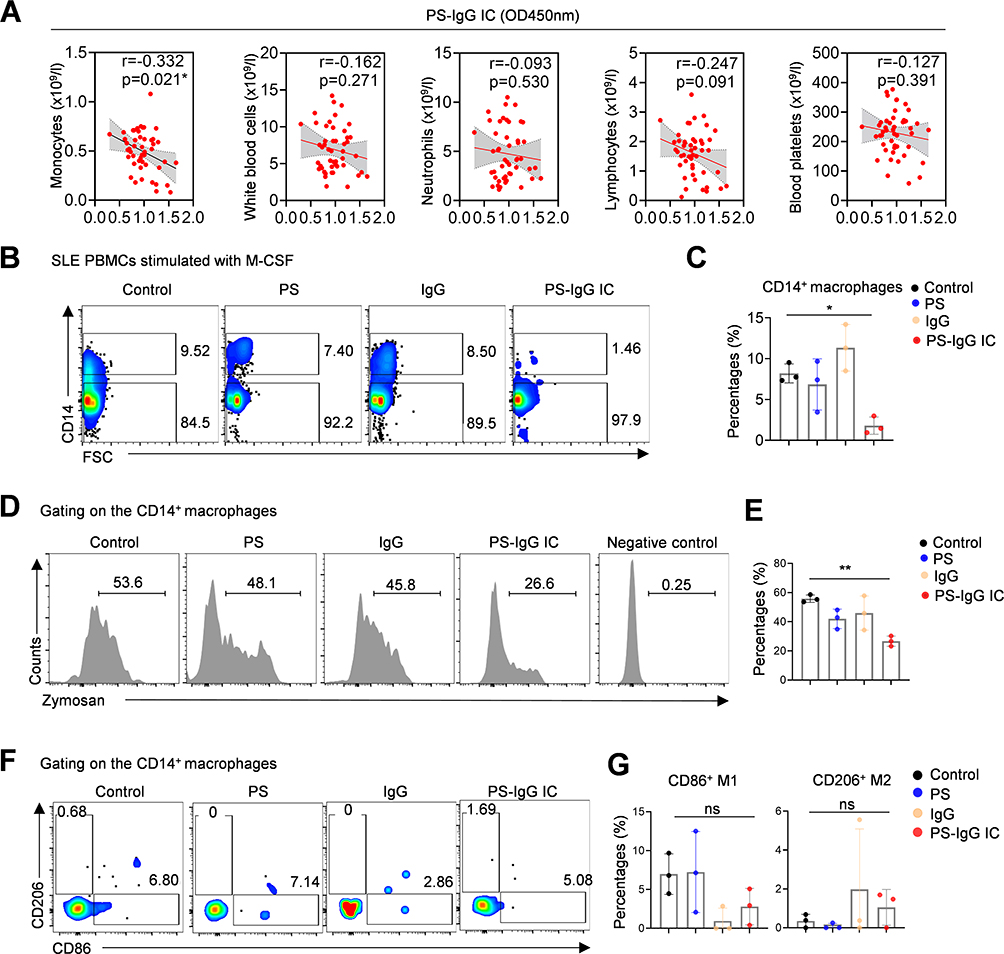 |
Figure 2 The PS-IgG IC treatment impairs phagocytotic function of SLE macrophages. (A) Correlation between serum PS-IgG IC and blood cell counts in SLE patients. Flow cytometric images and data plots show (B and C) the proportion of CD14+ macrophages, (D and E) phagocytosis of Zymosan and (F and G) CD206-, CD86-positive subsets. PBMCs from SLE patients were polarized by M-CSF for 5 days, then treated with PS, purified IgG or PS-IgG IC for 12 hours. Data were expressed as mean ± SD; ns, P > 0.05; *, P <0.05; **, P <0.01. |
 |
Figure 3 PS-IgG IC triggers oxidative stress in SLE macrophage. PBMCs from SLE patients were polarized by M-CSF for 5 days, then treated with PS, purified IgG or PS-IgG IC for 12 hours. Flow cytometric images and data plots show intracellular ROS (A and B), mitochondrial superoxide, and frequencies of mitochondria (C–F) in macrophages derived from SLE PBMCs. PBMCs from SLE patients were polarized by M-CSF for 5 days, then treated with PS-IgG IC or NAC combined PS-IgG IC for 12 hours. Flow cytometric images and data plot show intracellular ROS (G) and phagocytosis of Zymosan (H) in macrophages derived from SLE PBMCs. Data were expressed as mean ± SD; ns, P > 0.05; *, P <0.05; **, P <0.01. |
Then, monocytes in SLE PBMCs were induced to differentiate into macrophages with M-CSF. The induced SLE macrophages were treated with PS antigen, IgG protein, and PS-IgG IC, respectively. The effect of PS-IgG IC on macrophage function, including M1/M2 polarization and phagocytosis were then detected by flow cytometry. PS-IgG IC treatment significantly impaired the phagocytotic function of macrophages which detected with PHrodo Red Zymosan Bioparticles (Figure 2D and E). However, The PS-IgG IC treatment showed non-significant effect on the polarization of macrophages toward CD86+ M1 and CD206+ M2 subsets (Figure 2F and G).
PS-IgG IC Impairs Macrophages Functions in SLE Patients via Oxidative Stress
The ROS levels were then detected in SLE macrophages. Approximated 2-fold increase of intracellular ROS was detected by DCF staining in SLE macrophages with PS-IgG IC treatment (Figure 3A and B). Interestingly, no significant changes in mitochondrial superoxide and mitochondrial by MitoSOX and mitochondrial numbers by MitoSpy were detected by flow cytometry (Figure 3C and F), indicating that increase intracellular ROS may not be generated by mitochondria. While NAC, a ROS inhibitor reducing the thickness of the mucus in cytoplasm, could effectively reduce the ROS levels in PS-IgG IC treated SLE macrophages (Figure 3G). In addition, NAC treatment could restore the dampened phagocytotic functions of SLE macrophages (Figure 3H). Moreover, we detected the effects of PS-IgG IC in a monocyte cell line as well. THP-1 was stimulated with PMA to induce the differentiation toward macrophages, and the function of PS-IgG IC on macrophages was detected by flow cytometry. Insistently, the results showed that PS-IgG IC stimulation of THP-1 did not significantly affect the polarization toward M1/M2 (Figure S3B). However, it caused an increase in the intracellular total ROS level and impaired phagocytic function of THP-1 cells (Figure S3C and D).
PS-IgG IC Inhibits the Phagocytotic Function of Macrophages via LOX
To determine the potential cells affected by PS-IgG IC in SLE, SLE PBMCs were stimulated with PS-IgG IC treatment. Then, control and PS-IgG IC-treated SLE PBMCs were then subjected to RNA extraction and bulk RNA-sequencing analysis. The volcano plot showed the upregulation of LOX, CXCL2, IL35G, HBA1, CSF3, and downregulation of FcγR1A, LILPB5, CNTF, S100A12 in SLE PBMCs with PS-IgG IC stimulation (Figure 4A). The KEGG enrichment analysis showed that differentially expressed genes were enriched in the FcγR-mediated phagocytosis signaling pathway, IL-17 signaling pathways, and JAK-STAT signaling pathways in SLE PBMCs upon PS-IgG IC treatment (Figure 4B). This was consistent with the above findings that the phagocytosis function of macrophages decreased with PS-IgG IC stimulation. LOX has also been proved to be a chemical inducer of macrophages. Western blotting results validated the elevated LOX expression in PS-IgG IC treated SLE PBMCs (Figure 4C and D).
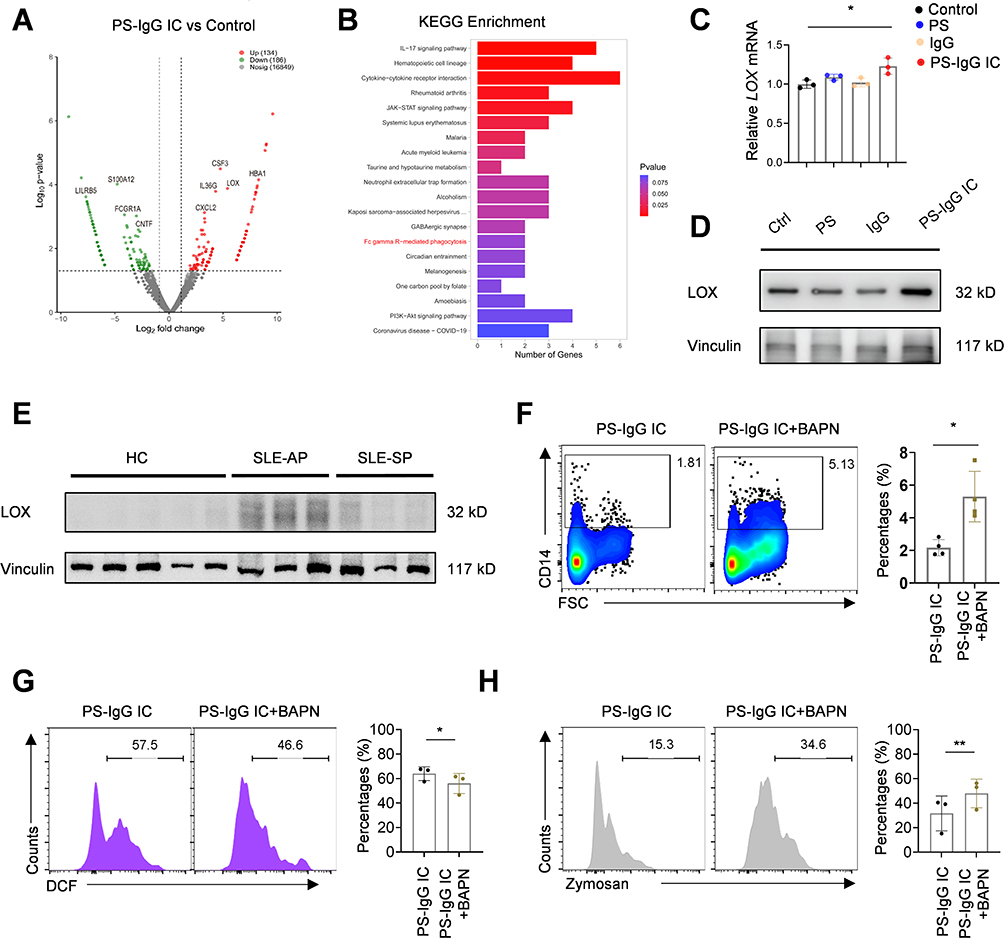 |
Figure 4 LOX can restore the phagocytosis function of macrophages. The volcano map (A), and KEGG enrichment analysis (B) of differentially expressed genes stimulated by PS-IgG IC in PBMCs compared to the control group. (C) RT-qPCR and (D) Western blot for LOX expression in PBS, PS, IgG and PS-IgG IC treated THP-1 cells. (E) Western blot for LOX expression in PBMCs of healthy controls, SLE patients with active period, and SLE patients with stable period. Flow cytometric images and data plot show CD14+ monocytes (F), intracellular ROS (G) and phagocytosis of Zymosan (H) in THP-1 cells. Data were expressed as mean ± SD; ns, P > 0.05; *, P <0.05; **, P < 0.01. |
We also found that LOX protein expression was significantly upregulated in SLE patients with active period compared with healthy controls (Figure 4E). To explore whether LOX participated ROS-mediated the impaired phagocytotic function of macrophages, we used LOX inhibitor BAPN to treat the PS-IgG IC stimulated THP-1 cells. Flow cytometer analysis suggested that BAPN treatment could increase the percentage of CD14+ monocytes in PS-IgG stimulated THP-1 cells (Figure 4F). We also observed that BAPN restored the ROS levels and phagocytotic function of THP-1 cells (Figure 4G and H). These data suggests that LOX mediates impaired phagocytic function of macrophages induced by PS-IgG IC.
PS Immunization Triggers Glomerular Immune Complex-Deposited Nephritis in Mice
The elevated serum levels of PS-IgG IC were detected in mice with chromatin-induced lupus (Figure 5A). To reveal the importance of PS-specific autoimmune response and PS-IgG IC formation during lupus development, we immunized mice with PS antigen (Figure 5B). PS mice showed significantly higher serum anti-PS IgG, PS-IgG IC, anti-dsDNA IgG, and serum creatinine levels compared to controls (Figure 5C–F). For renal assessment, the PS mice had more severe renal inflammatory cell infiltration (Figure 5G). Immunofluorescence results indicated that PS-IgG deposition in the kidneys was more severe in PS mice than in controls (Figure 5G and H). Besides, we also found that LOX was highly expressed in kidney macrophages of PS mice (Figure 5I). Our findings highly mimic the symptoms of SLE patients with renal damages, indicating the key roles of PS-specific autoimmune response and PS-IgG IC formation during LN development.
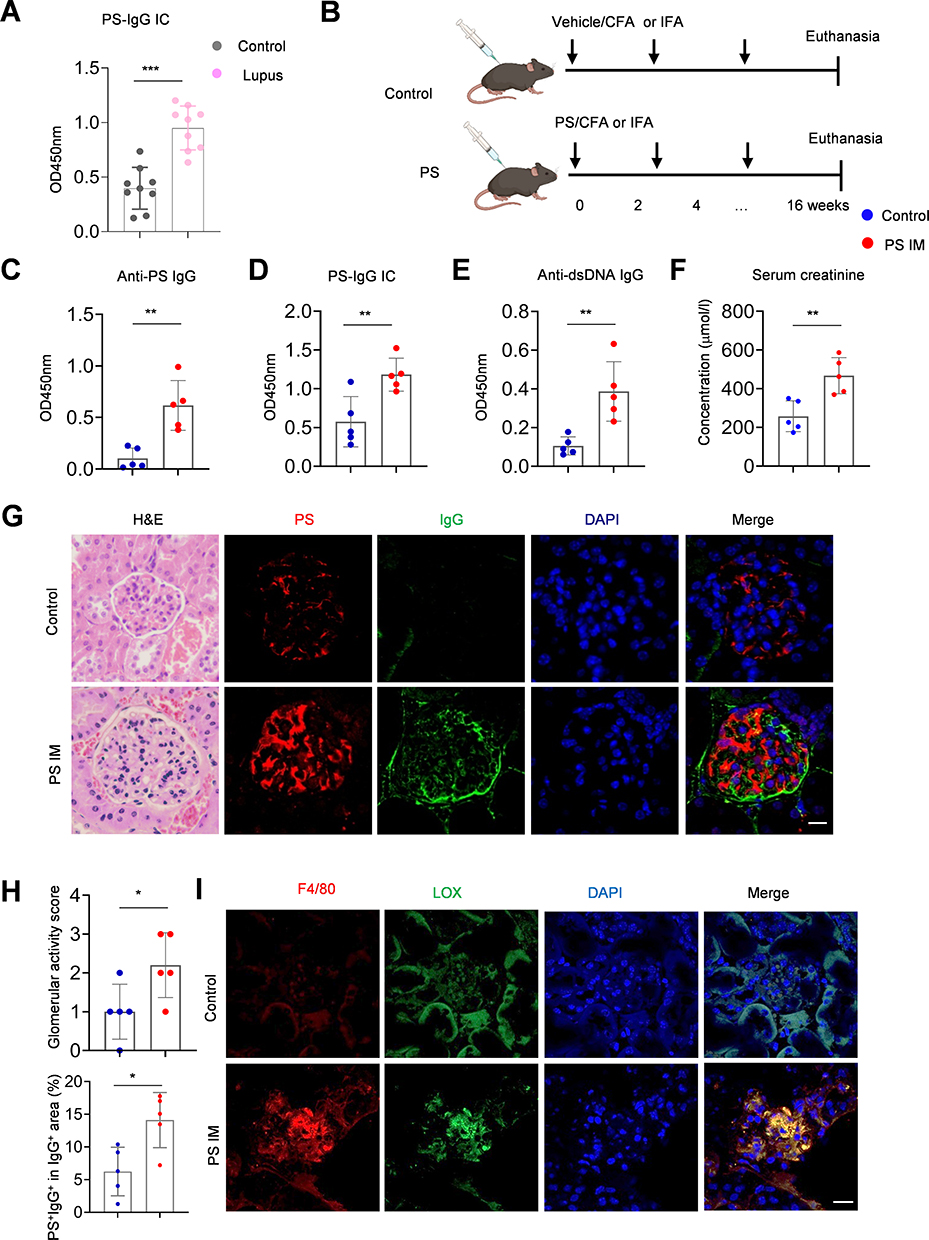 |
Figure 5 PS immunization triggers PS-IgG IC formation and renal injury in mice. (A) Serum PS-IgG immune complex levels in control and lupus mice (n = 9). (B) Flowchart shows the research strategy in generating PS-immunized mice. Data plots show the (C) serum anti-PS IgG, (D) PS-IgG IC, (E) anti-dsDNA IgG levels, and (F) creatinine levels in PS-immunized and control mice (n = 5 per group). (G) Representative images show H&E and confocal images of PS-IgG IC deposition in glomeruli of PS-immunized and control mice (n = 5 per group, Scale bar, 20μm). (H) Data plot shows the statistical analysis of PS-IgG IC deposition in glomeruli of PS-immunized and control mice. (I) Immunofluorescence for LOX expression and its co-localization with kidney macrophages in PS-immunized and control mice. Data were expressed as mean ± SD. *, P <0.05; **, P < 0.01, ***, P <0.001. |
Inhibition of ROS Restores Macrophage Function in PS-Immunized Mice
Then we analyzed the proportions, intracellular ROS, and phagocytosis of mononuclear macrophages in the bone marrow, kidney, and peritoneal cavity of PS and control mice by flow cytometry. Leukocytes were labeled with CD45, neutrophils were excluded with Ly6G, and macrophages were labeled with CD11b and F4/80 (Figure 6A). The results showed that there was reduction in the percentage of macrophages in peritoneal cavity and elevation in kidneys between the PS mice and control groups (Figure 6B). While, the total intracellular ROS levels of macrophages were significantly higher in the peritoneal cavity and kidney of mice with the PS immunization group (Figure 6C). Phagocytotic functions of macrophages in the peritoneal cavity were significantly reduced in the PS-immunized mice as well group (Figure 6D). The ex vivo addition of NAC in purified peritoneal macrophages could significantly inhibit intracellular ROS and restore the phagocytotic function of macrophages from PS-immunized mice (Figure 6E). Consistently, BAPN treatment can also restore the ROS level and phagocytotic function of macrophages from PS-immunized mice (Figure 6F and G).
 |
Figure 6 Impaired macrophage function in PS-immunized mice via oxidative stress. (A) Flow cytometric images show the CD45+CD11b+Ly6G−F4/80− monocyte and CD45+CD11b+Ly6G−F4/80+ macrophages, the intracellular ROS levels of macrophages in the kidney, peritoneal cavity, and bone marrow of mice. (B) Data plots show the statistics of macrophages in PS-immunized and control mice (n = 5 per group). (C) Data plots show the statistics of intracellular ROS levels in macrophages in PS-immunized and control mice (n = 5 per group). (D) Flow cytometric images and data plot show the phagocytosis of Zymosan in peritoneal macrophages of PS-immunized and control mice (n = 5 per group). (E) Flow cytometric images and data plot show the phagocytosis of Zymosan in PS-immunized peritoneal macrophages with or without NAC treatment (n = 5 per group). (F) Flow cytometric images and data plot show the intracellular ROS levels in PS-immunized peritoneal macrophages with or without BAPN treatment (n = 4 per group). (G) Flow cytometric images and data plot show the phagocytosis of Zymosan in PS-immunized peritoneal macrophages with or without BAPN treatment (n = 4 per group). Data were expressed as mean ± SD. Student’s t-test was used for comparison between two groups. ns, P > 0.05; *, P <0.05; **, P <0.01. |
In conclusion, our findings in SLE patients and PS-immunized mouse model both highlight the key pathogenic roles of PS-specific autoimmune response and PS-IgG IC during lupus nephritis development. Importantly, anti-oxidative drugs and LOX inhibitors may effectively rescue the immune-complex mediated macrophage dysfunction for the treatment of SLE (Figure 7).
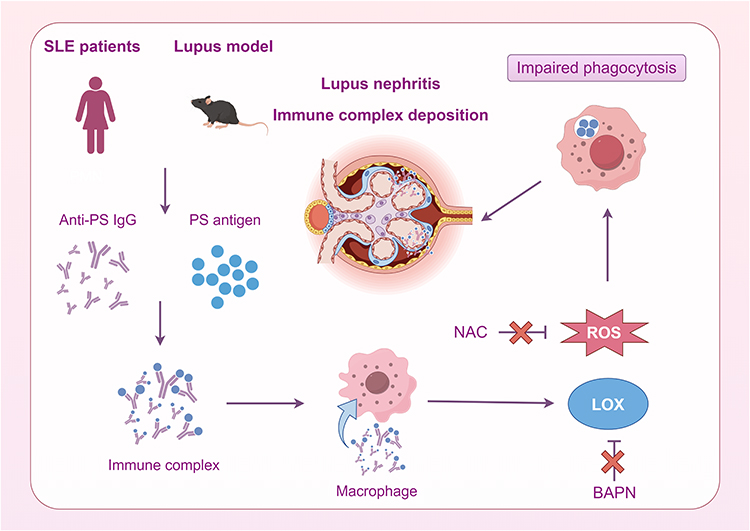 |
Figure 7 The mechanism diagram shows that in SLE patients and lupus model, anti-PS IgG can form PS-IgG IC with PS antigen, which induces oxidative stress and LOX of macrophages, decreases phagocytosis, increases circulating immune complexes, and finally mediates the occurrence of immune complex deposition glomerulonephritis. NAC and BAPN can reduce the ROS of macrophages and restore their phagocytic function. |
Discussion
SLE is an autoimmune disease with multiple organ damages, including the skin, nerves, gastroenterology, and kidney.27 The traditional treatment regimen for LN is mainly glucocorticoid combined with immunosuppressive therapy. However, due to the complex pathological mechanism of LN and individual differences in patients, only 30–50% of patients achieve remission after standard treatment. Therefore, it is necessary to develop new diagnostic biomarkers and explore new therapeutic targets to improve the prognosis of LN patients. Anti-phospholipid antibodies are closely associated with thrombocytopenia, hemolytic anemia, and epilepsy, and their role and mechanism in LN remain unclear. In the study, we explored the role of a novel anti-phospholipid antibody, anti-PS IgG, in the pathogenesis of SLE. Our results showed indicated the elevated serum anti-PS IgG antibodies in SLE patients, as previously reported.28,29 Besides, we reported that the PS-IgG IC with PS antigens caused LOX mediated oxidative stress and reduced phagocytosis in macrophages, decreased clearance of autoantigens, and ultimately lead to the immune complex deposition glomerulonephritis. Antioxidant NAC and LOX inhibitor BAPN can inhibit the intracellular ROS levels and restore the phagocytosis of macrophages.
Anti-phospholipid antibodies (aPLs) are a group of autoantibodies against various negatively charged phospholipids, phospholipid-binding proteins, and phospholipid-protein complexes, which are closely associated with thrombocytopenia, hemolytic anemia, neuropsychiatric lupus, arteriovenous thrombosis, pulmonary hypertension, and recurrent miscarriage in SLE patients.30 Vinicius Domingues et reported that the risk of acute and chronic microvascular renal lesions in aPL-positive patients was 3–5 times that of aPL-negative patients.31 Here, we showed that anti-PS IgG antibodies and PS-IgG IC were significantly higher in SLE compared with controls. Moreover, PS-IgG IC was found to correlate well with anti-dsDNA IgG and C4. Anti-PS IgG levels are markedly elevated in SLE patients compared to healthy controls and show a positive correlation with SLEDAI scores. Suggesting the anti-PS IgG contributed multiple organ damage in SLE, such as kidney, skin, joint, central nerves system and so on. Our previous study indicated the especially elevated anti-PS IgG levels in patients with LN.6 The underlying mechanisms may include immune complex formation, inflammatory cytokine secretion, neutrophil extracellular trap (NET) release and endothelial injury.32 Then, we further divided the SLE patients into high anti-PS IgG and low anti-PS IgG subgroups according to the median anti-PS IgG level and found that the high-level anti-PS IgG subgroup had higher disease activities and poorer renal function. Our ROC analysis demonstrated that PS-IgG IC alone achieved an AUC of 0.747 for SLE diagnosis and 0.716 for LN diagnosis, outperforming anti-PS IgG (AUC 0.641 for SLE; 0.665 for LN) and showing medium efficacy. Critically, value of PS-IgG IC lies in its unique mechanistic link to macrophage dysfunction, whereas anti-dsDNA IgG and complement (C3, C4) primarily reflect general immune activation. Collectively, these results suggest that elevated levels of anti-PS IgG in SLE patients are closely related to disease activity and renal injury and deserve further study.
To further clarify how PS-IgG IC is involved in and regulates SLE activity and renal injury, we investigated from the perspective of increased immune complex formation and impaired clearance. Previous studies have reported that elevated circulating immune complexes in SLE patients can lead to impaired phagocytosis of macrophages. The pathogenic effects of immune complex deposition in human lupus nephritis recruited monocytes and induced inflammatory responses in glomeruli.33 In our study, PS-IgG IC was found to be negatively correlated with counts of peripheral blood monocytes and not significantly correlated with other immune cells. Then SLE PBMCs were stimulated with PS-IgG IC, and flow cytometry revealed a significant decrease in the number of monocytes, while the number of T cells and B cells did not change significantly. The above results suggested that PS-IgG IC mainly acted by acting on monocytes. The abnormity of monocytes and macrophages has been indicated in human SLE.34
Previous studies have reported impaired macrophage phagocytosis of autoantigens and apoptotic cells in SLE patients.10,35,36 Nathan Richoz et al found that macrophages in the kidney are of different origins, one derived from monocyte-derived macrophages and one from tissue-resident macrophages. The former expresses FcγR, which internalizes circulating IC and presents IC-associated antigens. The latter has a limited ability to take up IC, express monocyte chemokines, and regulate monocyte recruitment to the kidney.37 Therefore, we extracted peripheral blood monocytes, induced differentiation into macrophages, and performed in vitro experiments to clarify the effect of PS-IgG IC on monocyte-derived macrophages. In this study, PS-IgG IC stimulated macrophages stimulated by PS-IgG IC produced elevated ROS level, accompanied by diminished phagocytosis. We observed the similar phenomenon in the THP-1 cell line. The above results suggest that PS-IgG IC could mediate impaired phagocytosis by elevating ROS in macrophages. Jing et al reported that IgG immune complexes could induce metabolic reprogramming of glycolysis in macrophages, secrete inflammatory cytokines such as IL-1β, IL-6, and TNF-α, and induce ROS production in a process that depends on hypoxia-inducible factor-1α.38 Several studies have suggested that ROS is elevated in macrophages phagocytosing IgG-containing immune complexes, which seems to be contrary to our findings.39,40 However, this distinction aligns with the dual nature of ROS: while physiological ROS bursts are essential for efficient phagocytosis, sustained ROS elevation disrupts redox balance and impairs macrophage function (causing cause oxidative stress and induce DNA damage and apoptosis)41—consistent with our rescue experiments using ROS scavengers (NAC). The therapeutic effects of NAC have been reported in patients with lupus nephritis and lupus mouse models.42,43 Here, we observed that phagocytosis and intracellular ROS levels of macrophage were restored after NAC treatment.
LOX plays an important role in the assembly of extracellular matrix related to catalytic activity, which is crucial for cell fate, differentiation and communication in the process of development, tissue maintenance and repair. LOX has also been shown to be a chemical inducer of macrophages. Chen et al found that LOX promoted macrophage infiltration through the β1 integrin-PYK2 pathway, recruiting macrophages to glioblastoma.19 In addition, it has been found that the methylation level of LOX promoter region is decreased and the expression is increased in rats exposed to dexamethasone during pregnancy, resulting in increased ROS production.44 Our transcriptome sequencing found that PS-IgG IC stimulation can increase the LOX levels in PBMCs of SLE patients, and Western blot analysis confirmed that PS-IgG IC can also increase the LOX protein expression, which was elevated in PBMCs of SLE patients with active period. In addition, we found that the LOX inhibitor BAPN can restore PS-IgG IC-mediated impairment of macrophage phagocytosis function and elevated ROS levels.
There are several lupus mouse models. For instance, MRL/lpr mice can be used to study genetic factors that lead to spontaneous lupus in mice.45 A chemical inducer, pristane, can also be used as an environmental factor to break immune tolerance for lupus development.46 In addition, the induced lupus mouse models can be established by subcutaneous injection of nuclear protein of apoptotic cells into C57BL/6J mice as we reported previously.25 In this study, we induced C57BL/6J mice to produce anti-PS IgG and PS-IgG IC by subcutaneous immunization of PS antigen to further explore the pathogenic mechanism of PS-IgG IC-mediated renal injury in mice. However, it is unclear whether the degree of macrophage infiltration in the kidney is affected by PS-IgG IC. Besides, it is not known whether PS-IgG IC affects the secretion of cytokines by macrophages in addition to the phagocytosis function.
Our study firstly identified the diagnostic values and underlying mechanism of the PS-IgG IC in SLE. Previous study highlighted the pathogenic roles of anti-dsDNA IgG in lupus nephritis, here, we proved the direct deposition of PS-IgG IC in renal glomeruli of SLE patients and animal models. Currently, the anti-Human Phosphatidylserine Recombinant Antibody (Bavituximab) have been primarily investigated for the treatment of advanced cancers, such as Unresectable Hepatocellular Carcinoma (HCC), Advanced Non-Small-Cell Lung Cancer (NSCLC) with ongoing trials. Our study may indicate the potential side effects of PS-targeted therapy in further. Our study provides PS-IgG IC as a novel biomarker for the diagnosis of SLE. Moreover, the antioxidant NAC and LOX inhibitor BAPN dramatically restore the PS-IgG IC impaired phagocytotic function of human and murine macrophages, indicating novel therapeutic targets of SLE by restoring the macrophage phagocytotic function. NAC and BAPN—though mechanistically effective—their clinical translation requires cautious consideration. The efficacy and safety profiles of novel therapeutic agents must be rigorously evaluated in human contexts.
This study still has some limitations. In further, we still need to assess the contribution of anti-PS IgG and PS-IgG IC in other organ-specific or systemic autoimmune diseases, such as type 1 diabetes, rheumatoid arthritis, multiple sclerosis and so on. Moreover, the deposition of PS-IgG IC in other organs, including skin, joint and central nerves system should also be detected in SLE patients and animal models. Proposed studies should also integrate the PS-IgG IC monitoring with single-cell RNA sequencing to investigating the potential targeted cells and mechanisms in SLE. The sample size of human and animal subjects recruited in this study was limited. In future, we still need to recruit newly diagnosed or relapsing SLE patients with PS-IgG IC targeted therapies.
Conclusion
Our study firstly identified the diagnostic values and underlying mechanism of the PS-IgG IC in SLE. In summary, our results suggest that PS-IgG IC can cause oxidative stress and impair their phagocytosis via elevated LOX. NAC and BAPN treatment dramatically restore the PS-IgG IC impaired phagocytotic function of human and murine macrophages, indicating novel therapeutic targets of SLE by restoring the macrophage phagocytotic function. Future validation in the underlying mechanisms of PS-IgG IC may provide novel targets for the treatment of SLE.
Abbreviations
SLE, Systemic lupus erythematosus; LN, lupus nephritis; IC, immune complex; PS, 5- phosphatidylserine; ROS, Reactive oxygen species; LOX, Lysyl oxidase; BAPN, β-Aminopropionitrile; NAC, N-acetylcysteine; HC, healthy controls; PBMCs, Peripheral blood mononuclear cells; MCP-1, chemoattractant protein-1; IL-1β, interleukin-1β; PBS, phosphate buffer saline; BSA, bovine serum albumin; HRP, horseradish peroxidase; TMB, tetramethylbenzidine; OD, optical density; HE, hematoxylin-eosin; PAS, periodic acid-schiff; SD, standard deviation; aPLs, Anti-phospholipid antibodies; OR, odds ratio.
Data Sharing Statement
The raw data involved in the study can be obtained by contacting the corresponding authors. All original data for RNA-sequencing was uploaded at Sequence Read Archive (SRA) database (PRJNA951058). The raw data of flow cytometry and ELISA were supplemented in Table S4. The other raw data involved in this article will be shared upon reasonable request to the corresponding author.
Study Approval
Samples and information from SLE patients and HC were collected from the Seventh Hospital of Sun Yat-sen University, the Sun Yat-sen Memorial Hospital of Sun Yat-sen University, and the Shenzhen Hospital of Peking University. The study has received ethical approval from the Sun Yat-sen Memorial Hospital (SYSEC-KY-KS-2021-339), the Seventh Affiliated Hospital of Sun Yat-sen University (KY-2021-097-01) and the Shenzhen Hospital of Peking University (2021-043). The patients provided their written informed consent to participate in this study. Animal experiments were approved by the Institutional Animal Care and Use Committee of Sun Yat-sen University (SYSU-IACUC-2021-000882).
Acknowledgments
We thank the SLE patients and healthy volunteers for their support of the study.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the Shenzhen Medical Research Fund (A2402020), China Postdoctoral Science Foundation (No. GZC20232407), National Nature Science Foundation of China (81901635, 82171782, 81974046, 82170690, 81974253), Shenzhen Science Foundation for Distinguished Young Scholars (RCJC20210706092009004), Guangdong Pearl River Talent Program (2021QN02Y108), Shenzhen Key Laboratory for Systems Medicine in Inflammatory Diseases (ZDSYS20220606100803007), the Natural Science Foundation of Guangdong (2019A1515011112), the Key Project of Basic Research of Shenzhen Science and Technology Innovation Commission (JCYJ20200109140203849, JCYJ20230807110715030), Shenzhen Key Laboratory of Immunity and Inflammatory Diseases (ZDSYS20200811143756018), Medical Scientific Research Foundation of Guangdong Province (A2021234), Research Foundation of Peking University Shenzhen Hospital (JCYJ2020008).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Kiriakidou M, Ching CL. Systemic lupus erythematosus. Ann Intern Med. 2020;172(11):ITC81–ITC96. doi:10.7326/AITC202006020
2. Durcan L, O’Dwyer T, Petri M. Management strategies and future directions for systemic lupus erythematosus in adults. Lancet. 2019;393(10188):2332–2343. doi:10.1016/S0140-6736(19)30237-5
3. Almaani S, Meara A, Rovin BH. Update on Lupus Nephritis. Clin J Am Soc Nephrol. 2017;12(5):825–835. doi:10.2215/CJN.05780616
4. Pan L, Lu MP, Wang JH, et al. Immunological pathogenesis and treatment of systemic lupus erythematosus. World J Pediatr. 2020;16(1):19–30. doi:10.1007/s12519-019-00229-3
5. Tsokos GC, Lo MS, Reis PC, Sullivan KE. New insights into the immunopathogenesis of systemic lupus erythematosus. Nat Rev Rheumatol. 2016;12(12):716–730. doi:10.1038/nrrheum.2016.186
6. Ma K, Du W, Wang S, et al. B1-cell-produced anti-phosphatidylserine antibodies contribute to lupus nephritis development via TLR-mediated Syk activation. Cell Mol Immunol. 2023;20(8):881–894. doi:10.1038/s41423-023-01049-2
7. Thanadetsuntorn C, Ngamjanyaporn P, Setthaudom C, et al. The model of circulating immune complexes and interleukin-6 improves the prediction of disease activity in systemic lupus erythematosus. Sci Rep. 2018;8(1):2620. doi:10.1038/s41598-018-20947-4
8. Di Benedetto P, Ruscitti P, Vadasz Z, et al. Macrophages with regulatory functions, a possible new therapeutic perspective in autoimmune diseases. Autoimmun Rev. 2019;18(10):102369. doi:10.1016/j.autrev.2019.102369
9. Moulton VR, Suarez-Fueyo A, Meidan E, et al. Pathogenesis of human systemic lupus erythematosus: a cellular perspective. Trends Mol Med. 2017;23(7):615–635. doi:10.1016/j.molmed.2017.05.006
10. Herrmann M, Voll RE, Zoller OM, et al. Impaired phagocytosis of apoptotic cell material by monocyte-derived macrophages from patients with systemic lupus erythematosus. Arthritis Rheum. 1998;41(7):1241–1250. doi:10.1002/1529-0131(199807)41:7<1241::AID-ART15>3.0.CO;2-H
11. Liang CL, Jiang H, Feng W, et al. Total glucosides of paeony ameliorate pristane-induced lupus nephritis by inducing PD-1 ligands(+) macrophages via activating IL-4/STAT6/PD-L2 signaling. Front Immunol. 2021;12:683249. doi:10.3389/fimmu.2021.683249
12. Craig ML, Bankovich AJ, McElhenny JL, Taylor RP. Clearance of anti-double-stranded DNA antibodies: the natural immune complex clearance mechanism. Arthritis Rheum. 2000;43(10):2265–2275. doi:10.1002/1529-0131(200010)43:10<2265::AID-ANR14>3.0.CO;2-J
13. Checa J, Aran JM. Reactive oxygen species: drivers of physiological and pathological processes. J Inflamm Res. 2020;13:1057–1073. doi:10.2147/JIR.S275595
14. Harris IS, DeNicola GM. The complex interplay between antioxidants and ROS in cancer. Trends Cell Biol. 2020;30(6):440–451. doi:10.1016/j.tcb.2020.03.002
15. Wang P, Geng J, Gao J, et al. Macrophage achieves self-protection against oxidative stress-induced ageing through the Mst-Nrf2 axis. Nat Commun. 2019;10(1):755. doi:10.1038/s41467-019-08680-6
16. Lucero HA, Kagan HM. Lysyl oxidase: an oxidative enzyme and effector of cell function. Cell Mol Life Sci. 2006;63(19–20):2304–2316. doi:10.1007/s00018-006-6149-9
17. Laczko R, Csiszar K. Lysyl Oxidase (LOX): functional contributions to signaling pathways. Biomolecules. 2020;10(8):1093. doi:10.3390/biom10081093
18. Martinez-Martinez E, Rodriguez C, Galan M, et al. The lysyl oxidase inhibitor (beta-aminopropionitrile) reduces leptin profibrotic effects and ameliorates cardiovascular remodeling in diet-induced obesity in rats. J Mol Cell Cardiol. 2016;92:96–104. doi:10.1016/j.yjmcc.2016.01.012
19. Chen P, Zhao D, Li J, et al. Symbiotic macrophage-glioma cell interactions reveal synthetic lethality in PTEN-null glioma. Cancer Cell. 2019;35(6):868–884e6. doi:10.1016/j.ccell.2019.05.003
20. Aringer M, Costenbader K, Daikh D, et al. 2019 European league against rheumatism/American college of rheumatology classification criteria for systemic lupus erythematosus. Arthritis Rheumatol. 2019;71(9):1400–1412. doi:10.1002/art.40930
21. Uribe AG, Vila LM, McGwin G, et al. The systemic lupus activity measure-revised, the Mexican Systemic Lupus Erythematosus Disease Activity Index (SLEDAI), and a modified SLEDAI-2K are adequate instruments to measure disease activity in systemic lupus erythematosus. J Rheumatol. 2004;31(10):1934–1940.
22. Yee CS, Farewell VT, Isenberg DA, et al. The use of systemic lupus erythematosus disease activity index-2000 to define active disease and minimal clinically meaningful change based on data from a large cohort of systemic lupus erythematosus patients. Rheumatology. 2011;50(5):982–988. doi:10.1093/rheumatology/keq376
23. Hoi A, Igel T, Mok CC, Arnaud L. Systemic lupus erythematosus. Lancet. 2024;403(10441):2326–2338. doi:10.1016/S0140-6736(24)00398-2
24. Ma K, Du W, Xiao F, et al. IL-17 sustains the plasma cell response via p38-mediated Bcl-xL RNA stability in lupus pathogenesis. Cell Mol Immunol. 2021;18(7):1739–1750. doi:10.1038/s41423-020-00540-4
25. Ma K, Li J, Wang X, et al. TLR4(+)CXCR4(+) plasma cells drive nephritis development in systemic lupus erythematosus. Ann Rheum Dis. 2018;77(10):1498–1506. doi:10.1136/annrheumdis-2018-213615
26. Akerstrom B, Brodin T, Reis K, Bjorck L. Protein G: a powerful tool for binding and detection of monoclonal and polyclonal antibodies. J Immunol. 1985;135(4):2589–2592. doi:10.4049/jimmunol.135.4.2589
27. Aringer M. Inflammatory markers in systemic lupus erythematosus. J Autoimmun. 2020;110:102374. doi:10.1016/j.jaut.2019.102374
28. Tsutsumi A, Hayashi T, Chino Y, et al. Significance of antiprothrombin antibodies in patients with systemic lupus erythematosus: clinical evaluation of the antiprothrombin assay and the antiphosphatidylserine/prothrombin assay, and comparison with other antiphospholipid antibody assays. Mod Rheumatol. 2006;16(3):158–164. doi:10.3109/s10165-006-0481-7
29. Urrego-Callejas T, Hernandez A, Giraldo SR, et al. Relevance of non-criteria antiphospholipid antibodies titers among patients with antiphospholipid syndrome and patients with systemic lupus erythematosus. Reumatol Clin. 2023;19(4):197–203. doi:10.1016/j.reuma.2022.07.003
30. Sammaritano LR. Antiphospholipid syndrome. Best Pract Res Clin Rheumatol. 2020;34(1):101463. doi:10.1016/j.berh.2019.101463
31. Domingues V, Chock EY, Dufrost V, et al. Increased risk of acute and chronic microvascular renal lesions associated with antiphospholipid antibodies in patients with systemic lupus erythematosus: a systematic review and meta-analysis. Autoimmun Rev. 2022;21(10):103158. doi:10.1016/j.autrev.2022.103158
32. Qin R, Wu H, Guan H, et al. Anti-phospholipid autoantibodies in human diseases. Clin Immunol. 2023;256:109803. doi:10.1016/j.clim.2023.109803
33. Olaru F, Dobel T, Lonsdorf AS, et al. Intracapillary immune complexes recruit and activate slan-expressing CD16+ monocytes in human lupus nephritis. JCI Insight. 2018;3(11). doi:10.1172/jci.insight.96492
34. Li Y, Lee PY, Reeves WH. Monocyte and macrophage abnormalities in systemic lupus erythematosus. Arch Immunol Ther Exp. 2010;58(5):355–364. doi:10.1007/s00005-010-0093-y
35. Tas SW, Quartier P, Botto M, Fossati-Jimack L. Macrophages from patients with SLE and rheumatoid arthritis have defective adhesion in vitro, while only SLE macrophages have impaired uptake of apoptotic cells. Ann Rheum Dis. 2006;65(2):216–221. doi:10.1136/ard.2005.037143
36. Kavai M, Szegedi G. Immune complex clearance by monocytes and macrophages in systemic lupus erythematosus. Autoimmun Rev. 2007;6(7):497–502. doi:10.1016/j.autrev.2007.01.017
37. Richoz N, Tuong ZK, Loudon KW, et al. Distinct pathogenic roles for resident and monocyte-derived macrophages in lupus nephritis. JCI Insight. 2022;7(21). doi:10.1172/jci.insight.159751
38. Jing C, Castro-Dopico T, Richoz N, et al. Macrophage metabolic reprogramming presents a therapeutic target in lupus nephritis. Proc Natl Acad Sci U S A. 2020;117(26):15160–15171. doi:10.1073/pnas.2000943117
39. Martinez J, Malireddi RK, Lu Q, et al. Molecular characterization of LC3-associated phagocytosis reveals distinct roles for Rubicon, NOX2 and autophagy proteins. Nat Cell Biol. 2015;17(7):893–906. doi:10.1038/ncb3192
40. Yang S, Zhao M, Jia S. Macrophage: key player in the pathogenesis of autoimmune diseases. Front Immunol. 2023;14:1080310. doi:10.3389/fimmu.2023.1080310
41. Tan HY, Wang N, Li S, et al. The reactive oxygen species in macrophage polarization: reflecting its dual role in progression and treatment of human diseases. Oxid Med Cell Longev. 2016;2016:2795090. doi:10.1155/2016/2795090
42. Lai ZW, Hanczko R, Bonilla E, et al. N-acetylcysteine reduces disease activity by blocking mammalian target of rapamycin in T cells from systemic lupus erythematosus patients: a randomized, double-blind, placebo-controlled trial. Arthritis Rheum. 2012;64(9):2937–2946. doi:10.1002/art.34502
43. Pannu N, Bhatnagar A. Oxidative stress and immune complexes: pathogenic mechanisms in pristane induced murine model of lupus. Immunobiology. 2020;225(1):151871. doi:10.1016/j.imbio.2019.11.006
44. Jiang T, Xiao H, Li B, et al. LOX overexpression programming mediates the osteoclast mechanism of low peak bone mass in female offspring rats caused by pregnant dexamethasone exposure. Cell Commun Signal. 2023;21(1):84. doi:10.1186/s12964-023-01115-2
45. Ji LN, Wu S, Fu DQ, et al. Jieduquyuziyin Prescription alleviates hepatic gluconeogenesis via PI3K/Akt/PGC-1alpha pathway in glucocorticoid-induced MRL/lpr mice. J Ethnopharmacol. 2022;284:114815. doi:10.1016/j.jep.2021.114815
46. McGaha TL, Madaio MP. Lupus nephritis: animal modeling of a complex disease syndrome pathology. Drug Discov Today Dis Models. 2014;11:13–18. doi:10.1016/j.ddmod.2014.08.002
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.