Back to Journals » Journal of Inflammation Research » Volume 18
Anesthesia-Induced Ferroptosis: Bidirectional Regulation and Molecular Mechanisms in Cardio-Cerebral Injury
Authors Chen Y ![]() , Ouyang J
, Ouyang J ![]() , Zhao W, Cheng L, Ye X, Hu Y, Si Y, Niu Q
, Zhao W, Cheng L, Ye X, Hu Y, Si Y, Niu Q ![]() , Zhang H, Qiao Q, Zhang J
, Zhang H, Qiao Q, Zhang J ![]()
Received 11 July 2025
Accepted for publication 9 November 2025
Published 17 November 2025 Volume 2025:18 Pages 16023—16043
DOI https://doi.org/10.2147/JIR.S553229
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Qing Lin
Ying Chen,1,2,* Jie Ouyang,2,* Weili Zhao,2 Lu Cheng,3– 5 Xuerui Ye,3– 5 Yong Hu,3– 5 Yongyu Si,2 Qin Niu,2 Haoling Zhang,6 Qian Qiao,3– 5 Jingjing Zhang3– 5
1Department of Anesthesiology, Southwest Medical University, Luzhou, Sichuan Province, 646000, People’s Republic of China; 2Department of Anesthesiology, The Second Affiliated Hospital of Kunming Medical University, Kunming, Yunnan Province, 650000, People’s Republic of China; 3Department of Cardiovascular Medicine, Fuwai Yunnan Hospital, Chinese Academy of Medical Sciences, Affiliated Cardiovascular Hospital of Kunming Medical University, Kunming, 650000, People’s Republic of China; 4Yunnan Provincial Cardiovascular Clinical Medical Center, Kunming, 650000, People’s Republic of China; 5Yunnan Provincial Cardiovascular Clinical Medical Research Center, Kunming, 650000, People’s Republic of China; 6Department of Biomedical Science, Advanced Medical and Dental Institute, University Sains Malaysia, Penang, 13200, Malaysia
*These authors contributed equally to this work
Correspondence: Jingjing Zhang, Department of Cardiovascular Medicine, Fuwai Yunnan Hospital, Chinese Academy of Medical Sciences, Affiliated Cardiovascular Hospital of Kunming Medical University/Yunnan Provincial Cardiovascular Clinical Medical Center/Yunnan Provincial Cardiovascular Clinical Medical Research Center, Kunming, Yunnan Province, 650000, People’s Republic of China, Email [email protected] Haoling Zhang, Department of Biomedical Science, Advanced Medical and Dental Institute, University Sains Malaysia, Penang, 13200, Malaysia, Email [email protected]
Abstract: Perioperative use of common anesthetics—including sevoflurane, propofol, and dexmedetomidine—may induce cardio-cerebral injury via ferroptosis, an iron-dependent form of cell death. We introduce the “Molecular Switches”, proposing that these drugs act as tissue-specific switches regulating ferroptosis bidirectionally. Their effect (promote or inhibit) depends critically on local factors: receptor expression profiles, metabolic status, baseline redox tone, and post-translational modifications of key proteins like glutathione peroxidase 4 (GPX4). High-risk organs like heart and brain, characterized by elevated metabolic demands, polyunsaturated fatty acid (PUFA)-rich membranes, and stringent iron homeostasis, express unique molecular switch configurations explaining their susceptibility. During ischemia-reperfusion injury (IRI), leveraging this principle allows protective anesthetic strategies: targeting the nuclear factor erythroid 2–related factor 2 (Nrf2)/GPX4/solute carrier family 7 member 11 (SLC7A11) antioxidant axis enhances endogenous defenses, while inhibiting Acyl-CoA synthetase long-chain family member 4 (ACSL4)-mediated lipid peroxidation limits damage initiation. Crucially, effective myocardial protection prioritizes mitochondrial function recovery and iron efflux modulation, whereas cerebroprotection centers on preserving neuronal iron homeostasis and blood-brain barrier (BBB) integrity—distinct applications derived directly from understanding tissue-specific molecular switches. Addressing clinical translation challenges (limited drug specificity, complex polypharmacy effects, biomarker gaps), we advocate for developing personalized anesthetic protocols informed by molecular switch profiling, employing nanocarriers for targeted delivery across the BBB, and establishing AI-driven predictive models based on ferroptosis biomarkers. This framework provides novel insights for optimizing perioperative cardio-cerebral protection.
Keywords: anesthetics, ferroptosis, cardio-cerebral injury, molecular switches, ischemia-reperfusion injury, bidirectional regulation
Introduction
Ferroptosis, first described in 2012, is a distinct form of regulated cell death driven by intracellular iron overload and lipid peroxidation, with unique morphological and molecular features compared to apoptosis or necrosis.1 Its biochemical hallmarks include impaired glutathione peroxidase 4 (GPX4)/GSH-mediated lipid peroxide detoxification,2 Acyl-CoA synthetase long-chain family member 4 (ACSL4)/lysophosphatidylcholine acyltransferase 3 (LPCAT3)-driven incorporation of polyunsaturated fatty acids (PUFAs) into phospholipids,3 and iron-dependent Fenton reactions that exacerbate reactive oxygen species (ROS) accumulation and membrane damage.4
Early perioperative studies noted that anesthetic exposure—particularly sevoflurane and isoflurane—was associated with cognitive impairment and organ injury, though the mechanisms remained unclear.5 With the recognition of ferroptosis in 2012, research began linking anesthetic-induced organ dysfunction to iron overload and GPX4 suppression. Subsequent work demonstrated context-dependent effects: inhaled anesthetics promote ferroptosis in developing brain tissue,6,7 while intravenous agents such as propofol may either aggravate neuronal injury8,9 or provide protection in cerebral ischemia-reperfusion injury via nuclear factor erythroid 2–related factor 2 (Nrf2)/GPX4 activation.10 More recently, the “molecular switch” concept has been proposed, framing anesthetics as context-specific regulators of ferroptosis, modulated by receptor expression, metabolic state, and post-translational modifications of ferroptosis-related proteins.
The brain and heart are particularly susceptible to ferroptosis due to their high oxygen consumption, PUFA-rich membranes, and strict iron homeostasis requirements.11–14 Clinically, postoperative neurocognitive disorders affect up to 30–50% of elderly patients, while perioperative myocardial ischemia-reperfusion injury remains a leading cause of morbidity and mortality. However, current anesthetic strategies rarely integrate ferroptosis susceptibility into perioperative risk assessment, representing a translational gap between bench and bedside.
Despite extensive progress, inconsistencies remain. For example, while dexmedetomidine has been shown to inhibit ferroptosis and protect against ischemia-reperfusion injury in several studies,11,15,16 other findings suggest negligible effects under different experimental conditions. Such discrepancies may stem from variations in dosing, timing, animal models, or tissue-specific vulnerabilities, underscoring the need for critical comparative evaluation rather than uniform conclusions.
This review seeks not only to summarize anesthetic–ferroptosis interactions but also to provide a unifying framework: anesthetics as bidirectional modulators of ferroptosis through tissue-specific molecular switches. By integrating mechanistic studies, clinical epidemiology, and unresolved controversies, we aim to highlight the translational significance of ferroptosis in perioperative organ protection and identify priority directions for future research.
Bidirectional Mechanisms of Anesthetic Agents and Ferroptosis
Anesthetics exert complex, context-dependent effects on ferroptosis, acting as either promoters or inhibitors according to tissue type, exposure duration, and underlying pathology. This bidirectionality operates chiefly through concerted modulation of iron metabolism, lipid peroxidation, and antioxidant defenses, positioning anesthetics as potential molecular switches of ferroptosis. In practice, the directional outcome reflects (i) tissue-specific receptor landscapes and pathway activation states, (ii) differences in metabolic demand and substrate use (eg, high-energy neurons/cardiomyocytes vs metabolically reprogrammed tumors), (iii) baseline redox tone and iron homeostasis (eg, ischemia-reperfusion (I/R)-associated oxidative stress vs chronically stressed tumor microenvironments), and (iv) expression, post-translational modification, and localization of core regulators (GPX4, ACSL4, iron transporters). Consequently, any given anesthetic must be interpreted within its microenvironmental context; the combined state of these switches finally determines whether ferroptosis is promoted or inhibited.17 The bidirectional regulatory effects of various anesthetic agents on ferroptosis are summarized in Table 1.
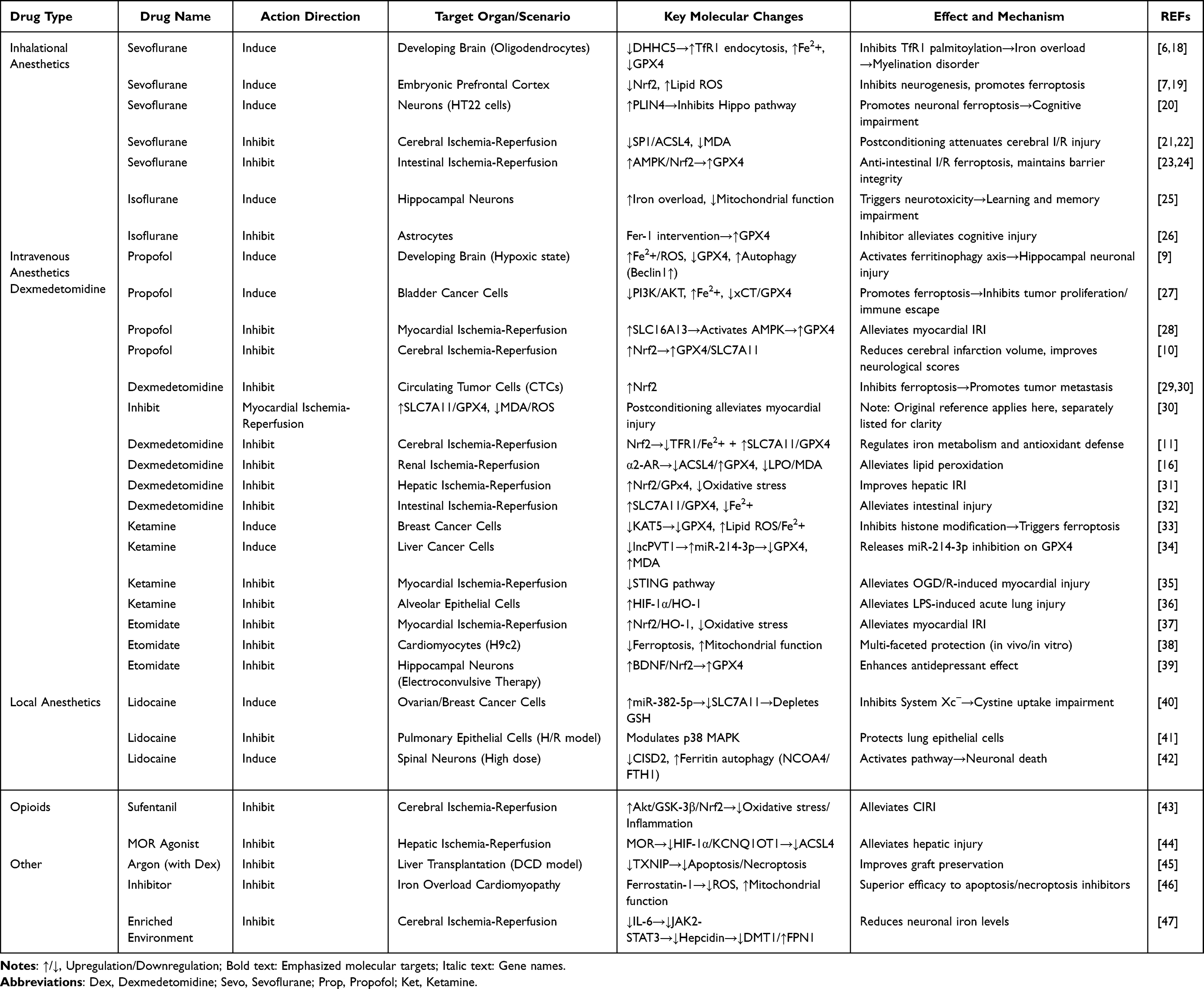 |
Table 1 Bidirectional Regulatory Effects of Anesthetic Agents on Ferroptosis |
Inhaled Anesthetics
Sevoflurane has been widely investigated for ferroptotic effects. In the developing brain, it disrupts iron homeostasis by inhibiting palmitoyltransferase ZDHHC5 (DHHC5)-mediated palmitoylation of transferrin receptor 1 (TfR1), causing abnormal iron uptake, intracellular iron overload, and oligodendrocyte ferroptosis.6 It also suppresses Nrf2 activity, weakening antioxidant defense and exacerbating lipid peroxidation in neuronal tissues.7 In hippocampal cells, sevoflurane upregulates perilipin 4 (PLIN4) and protein kinase C delta (PRKCD), inhibits Hippo signaling, and triggers neuronal injury consistent with ferroptosis.20,48,49 By contrast, in I/R models, sevoflurane postconditioning inhibits ACSL4 via the specificity protein 1 (SP1)/ACSL4 axis, reduces malondialdehyde (MDA), and restores GPX4, thereby protecting neurons;21 it also activates AMP-activated protein kinase (AMPK)/Nrf2/GPX4 signaling to maintain intestinal barrier integrity after I/R.23 Notably, dose and timing are critical, as higher concentrations can intensify oxidative stress and accelerate ferroptosis.20
Isoflurane shows a similar duality. In hippocampal neurons it increases iron uptake, induces mitochondrial dysfunction, and drives ferroptosis with cognitive sequelae; iron chelation mitigates this injury.25 In astrocytes, isoflurane-triggered ferroptosis is rescued by ferrostatin-1, underscoring the central role of GPX4.26 Collectively, inhaled anesthetics exemplify a context-dependent balance between pro- and anti-ferroptotic actions.
Intravenous Anesthetics
Propofol displays pronounced bidirectionality. In neonatal hippocampus under hypoxia, it promotes ferroptosis by upregulating TfR1/divalent metal transporter 1 (DMT1), downregulating ferroportin 1 (FPN1), and inhibiting GPX4/SLC7A11, causing iron overload and neuronal lipid peroxidation.8,9 It can concurrently activate autophagy (↑Beclin1, sequestosome 1 (SQSTM1) degradation), aggravating developmental neurotoxicity.37 In circulating tumor cells, propofol inhibits ferroptosis and may enhance metastatic survival potential.29 Conversely, in cerebral I/R, propofol activates Nrf2/GPX4, attenuates lipid peroxidation, and reduces infarct volume; GPX4 or Nrf2 inhibition (RAS-selective lethal 3 (RSL3), ML385) abolishes this protection.10 In cardiomyocytes, propofol preconditioning enhances SLC16A13–AMPK–GPX4 signaling, suppressing ferroptosis and limiting infarct size.28
Dexmedetomidine predominantly inhibits ferroptosis across organs. In myocardium, it activates the SLC7A11/GPX4 axis and regulates mitochondrial metabolism via pyruvate dehydrogenase kinase 4 (PDK4)/malate dehydrogenase 2 (MDH2) lactylation;50,51 in neurons, it downregulates the SRY-box transcription factor 9 (SOX9)/DMT1 axis and upregulates Nrf2, curbing iron uptake and oxidative injury.11,12 Protective effects in liver, kidney, and lung involve ACSL4 inhibition, ferritinophagy suppression, and broader antioxidant pathway activation.15,16,31,52
Ketamine demonstrates striking tissue specificity. In cancer models, it promotes ferroptosis by repressing GPX4 transcription through the lysine acetyltransferase 5 (KAT5)/histone H3 lysine 27 acetylation (H3K27ac) axis33 or via lncPVT1/miR-214-3p-mediated GPX4 suppression.34 In contrast, in cardiomyocytes, ketamine inhibits ferroptosis during I/R by downregulating stimulator of interferon genes (STING);35 under chronic stress, it increases GPX4 and ferritin heavy chain 1 (FTH1) while reducing TfR1 in habenular neurons, alleviating ferroptosis and improving function;53 and it protects against acute lung injury via hypoxia-inducible factor 1-alpha (HIF-1α)/heme oxygenase-1 (HO-1).36 In developing hippocampal neurons, ketamine can also activate the N-methyl-D-aspartate receptor (NMDA-R)/RASD1/DMT1 pathway, causing iron overload, GSH depletion, and cognitive consequences.54
Etomidate consistently inhibits ferroptosis: it activates Nrf2/HO-1 and suppresses lipid peroxidation in myocardial I/R,37 and upregulates brain-derived neurotrophic factor (BDNF)/Nrf2/GPX4 to mitigate ferroptosis in ECT-related hippocampal injury.39
Local Anesthetics
Lidocaine displays dose- and tissue-dependent effects. At low concentrations, it promotes ferroptosis in ovarian and breast cancer cells by upregulating miR-382-5p, which suppresses SLC7A11 and GPX4.40 At higher spinal doses, it induces neuronal ferroptosis via nuclear receptor coactivator 4 (NCOA4)-mediated ferritinophagy with concomitant FTH1/GPX4 suppression and CDGSH iron sulfur domain 2 (CISD2) downregulation.42 Conversely, in lung I/R, lidocaine activates p38 mitogen-activated protein kinase (p38 MAPK) and prevents ferroptosis in alveolar epithelial cells, conferring protection.41
Comparison of Conflicting Results Across Studies
Despite rapid progress, apparent inconsistencies persist. Propofol aggravates neuronal ferroptosis in neonatal hypoxia but protects mature brains during I/R.8–10 Dexmedetomidine is generally protective, though minimal effects have been reported depending on dose and timing.11,15,16 Such discrepancies stem from differences in developmental stage, organ context, anesthetic class/dose/exposure, and outcome readouts. Rather than classifying agents as uniformly pro- or anti-ferroptotic, anesthetics should be regarded as context-dependent molecular switches whose effects are set by iron metabolism, redox status, and microenvironmental factors. Recognizing these determinants is essential for reconciling findings and designing targeted perioperative strategies.
Key Molecular Pathways
Interference with Iron Metabolism
Common anesthetics (eg, halothane, isoflurane) upregulate TfR1 after administration, augmenting iron uptake and predisposing to intracellular overload.6 Enhanced ferritin degradation increases the labile iron pool, while dysfunction of iron-handling proteins (including transport and export components) further deranges iron homeostasis—changes linked to neuronal injury and cognitive dysfunction in the young and elderly.55,56 Disrupting iron-regulatory nodes (including transporter function) mitigates anesthesia-induced neuronal damage, underscoring iron metabolism as a pivotal target.57
Activation of Lipid Peroxidation
Iron accumulation perturbs lipid metabolism and fosters PUFA enrichment, sensitizing membranes to peroxidation.58 Increased lipoxygenase (LOX) activity and maladaptive phospholipid remodeling further accelerate lipid radical chain reactions and compromise membrane integrity—phenomena observed with anesthetic exposure and tightly coupled to ferroptotic death.26
Suppression of Antioxidant Defenses
Anesthetics can impair System Xc−, limiting cystine uptake, depleting GSH, and weakening ROS clearance.59 Sevoflurane-induced ferroptosis and oxidative stress are attenuated by restoring GSH/GPX4 levels.7,60 Diminished GPX4 activity permits lipid hydroperoxide accumulation and membrane breakdown, especially in developing/aging brain and cardiac tissue, aligning with postoperative cognitive and organ dysfunction.
Across these axes—iron dysregulation, lipid peroxidation, and antioxidant collapse—anesthetics form an integrated network that can precipitate or prevent ferroptosis depending on context. Targeting these nodes (eg, supporting GSH synthesis, boosting GPX4, modulating LOX or iron flux) offers avenues to mitigate anesthesia-associated neurotoxicity and cardiotoxicity, with precision interventions tailored to population-specific iron/redox states (eg, pediatric, elderly).
Mechanisms of Anesthesia-Induced Ferroptosis in Cardiac and Cerebral Injury
Anesthesia-induced ferroptosis contributes substantially to perioperative cardiac and cerebral injury. Both organs share features that predispose them to ferroptosis—high oxygen consumption, PUFA-enriched membranes, and strict iron regulation—yet they differ in vulnerability patterns and pathway emphasis. Below, we delineate organ-specific cascades linking anesthetic exposure to ferroptotic damage while maintaining continuity with preceding sections.
Cardiac Injury
Iron Overload and Transport Dysregulation
In cardiomyocytes, anesthetics disrupt iron homeostasis by upregulating TfR1 and DMT1 while downregulating the exporter ferroportin (FPN1). This shift enlarges the labile iron pool, enhances Fenton chemistry, and seeds ROS-driven membrane damage.13,54 In contrast, dexmedetomidine limits iron influx and promotes efflux by suppressing DMT1 and restoring FPN1 expression, thereby alleviating iron overload and early ferroptotic priming.11,12 In parallel, selective autophagic turnover of ferritin (ferritinophagy) can be engaged under anesthetic/reperfusion stress, further increasing cytosolic Fe2⁺ and reinforcing the overload state.
Lipid Peroxidation Cascade
Isoflurane and ketamine elevate ACSL4 and lLPCAT3, promoting the esterification and membrane incorporation of pro-ferroptotic PUFAs. This remodeling heightens susceptibility to peroxidation and supports lipid radical chain reactions.61,62 Loss or inhibition of GPX4 accelerates accumulation of lipid hydroperoxides, destabilizing sarcolemmal and mitochondrial membranes.
Antioxidant System Collapse
Propofol and lidocaine can depress System Xc− (SLC7A11) and GPX4, impairing cystine import, depleting GSH, and blunting detoxification of lipid peroxides.63 By contrast, etomidate and dexmedetomidine activate Nrf2 signaling, upregulate GPX4, and partially reconstitute redox homeostasis under ischemia-reperfusion stress.15,37
Mitochondrial Dysfunction
Mitochondria are primary targets of ferroptotic injury. Under hypoxia or reperfusion, anesthetics such as propofol amplify mitochondrial ROS, disrupt electron transport chain function, and depress adenosine triphosphate (ATP) generation. Dexmedetomidine counters this metabolic collapse through PDK4/MDH2 lactylation control, sustaining energetic balance and limiting downstream ferroptosis.51
Outcome: These converging axes—iron overload, PUFA-rich membrane remodeling, antioxidant failure, and mitochondrial ROS—culminate in cardiomyocyte ferroptosis, manifesting as contractile dysfunction, arrhythmias, and heightened susceptibility to ischemia-reperfusion injury.
Cerebral Injury
Iron Uptake Dysregulation
In neurons, inhaled anesthetics can engage the NMDA-R/RASD1/DMT1 axis, facilitating iron influx and neuronal iron overload.54 Propofol additionally suppresses FPN1, further favoring intracellular retention in hippocampal cells. Dexmedetomidine opposes this by inhibiting SOX9/DMT1 signaling, restoring iron balance and reducing ferroptotic drive.11
Blood–Brain Barrier (BBB) Disruption
Sevoflurane compromises BBB integrity by downregulating tight-junction proteins (occludin, claudin-5) and increasing matrix metalloproteinases (MMPs) and HIF-1α, which together enhance iron penetration and oxidative stress within the neurovascular unit.64 Isoflurane similarly promotes endothelial ferroptosis and microvascular leakage, amplifying parenchymal exposure to iron and inflammatory mediators.
ROS Accumulation and Lipid Peroxidation
Upregulated ACSL4/LPCAT3 and suppressed GPX4 favor excessive lipid peroxide buildup in neurons. Both inhaled and intravenous agents can depress Nrf2/SLC7A11, collapsing antioxidant defenses and accelerating ferroptosis. Mitochondrial ROS further amplify peroxidation kinetics, driving synaptic and dendritic injury.
Neuroinflammation and Cognitive Dysfunction
Ferroptotic neurons release damage-associated molecular patterns (DAMPs) that activate microglia and propagate inflammatory cascades, contributing to postoperative cognitive decline (POCD). Pharmacologic ferroptosis inhibition (eg, ferrostatin-1) or dexmedetomidine attenuates neuroinflammation and improves memory outcomes in experimental models.12,15
Outcome: Iron overload, BBB compromise, and ROS-driven lipid peroxidation converge on neuronal ferroptosis, culminating in cognitive impairment and perioperative neurotoxicity.
Comparative Insights
Cardiac vs Cerebral Tissue
Both rely on Nrf2/GPX4 defense, but cardiomyocytes are particularly vulnerable to mitochondrial metabolic collapse, whereas neurons are especially sensitive to iron-uptake dysregulation and BBB injury. Distinct susceptibilities of cardiac and cerebral tissues to ferroptosis and their anesthetic modulation are compared in Table 2.
 |
Table 2 Differences in Susceptibility to Ferroptosis Between Cardiac and Cerebral Organs and Anesthetic Impact |
Protective Anesthetics
Dexmedetomidine and etomidate consistently suppress ferroptosis and preserve organ function, while sevoflurane and propofol display context-dependent pro- or anti-ferroptotic actions shaped by developmental stage, ischemia-reperfusion status, and redox/iron baselines.
Clinical Context and Translational Evidence
Ferroptosis is now recognized as a major mechanistic axis of perioperative cardiac and cerebral injury; paradoxically, several anesthetics can also protect heart and brain by suppressing ferroptosis and normalizing iron/redox homeostasis. Table 3 summarizes classical and emerging assays for ferroptosis in experimental and clinical settings. Here, we connect the epidemiologic burden with biomarker/imaging readouts, reconcile heterogeneous findings, and embed the protective actions of anesthetics within a practical translational framework.
 |
Table 3 Mechanisms of Organ Protection by Anesthetic Agents via Ferroptosis Inhibition |
Epidemiology of Perioperative Cardio-Cerebral Complications
Postoperative neurocognitive disorders (PND) occur in ~30–50% of elderly surgical patients, with long-term decline in ~10–15%; risk is higher with prolonged anesthesia, major cardiovascular/orthopedic surgery, and comorbid cerebrovascular disease or diabetes.11–14 Perioperative myocardial injury (PMI) is detected in 8–19% of non-cardiac surgeries and remains a leading cause of perioperative morbidity and mortality, with I/R episodes amplifying cardiomyocyte ferroptosis. Other organ injuries (eg, acute kidney injury, hepatic ischemia) increasingly correlate with ferroptosis markers, underscoring the systemic perioperative footprint of this pathway.
Linking Ferroptosis Susceptibility to Clinical Outcomes
Biomarkers and imaging. Elevated lipid peroxidation products (MDA, 4-hydroxynonenal (4-HNE)) and reduced GPX4 activity associate with worse neurocognitive and cardiac outcomes. Systemic iron indices (ferritin, transferrin, Fe) reflect susceptibility, while circulating exosomal miRNAs (eg, miR-214-3p, miR-137) that regulate SLC7A11/GPX4 are emerging liquid-biopsy indicators. Iron-sensitive magnetic resonance imaging (MRI) (susceptibility-weighted brain imaging; cardiac T2*) identifies regional iron accumulation consistent with ferroptotic stress; functional metrics (lower heart-rate variability, higher troponin, impaired Mini-mental State Examination (MMSE)/montreal Cognitive Assessment (MoCA)) track with these biochemical profiles. Together, these data indicate that ferroptosis is clinically observable during the perioperative period and linked to adverse outcomes.
Protective actions of anesthetics—cardiac I/R. Myocardium is ferroptosis-prone due to high mitochondrial density and oxidative load. Multiple anesthetics counteract I/R-related ferroptosis by acting on iron export, lipid peroxidation, and GPX4-centered defenses:
- Sevoflurane upregulates growth differentiation factor 11 (GDF11) to activate phosphoinositide 3-kinase/Protein kinase B (PI3K/AKT), mitigating oxidative stress, inflammation, and apoptosis.79
- Propofol preconditioning suppresses ferroptosis via the SLC16A13–AMPK–GPX4 axis;28 combined propofol + dexmedetomidine synergistically engages Akt/mammalian target of rapamycin (mTOR) and Nrf2/GPX4, enhancing protection.83
- Etomidate activates Nrf2/HO-1 to inhibit ferroptosis37 and further limits mitochondrial damage while curbing ferroptotic death.27
- Dexmedetomidine activates SLC7A11/GPX4;30 relieves histone deacetylase 2 (HDAC2)-mediated repression of ferroportin to promote iron efflux;13 engages α2-receptors to drive Nrf2/SLC7A11/GPX4 and dampen ferroptosis/inflammation;50 increases nuclear receptor subfamily 3 group C member 1 (NR3C1) phosphorylation to inhibit PDK4, reduce lactate and MDH2 lactylation, and improve mitochondrial function;51 upregulates miR-141-3p to suppress lncRNA TUG1 and blunt hypoxia/reoxygenation-induced ferroptosis;80 and activates cyclic adenosine monophosphate (cAMP)/protein kinase A (PKA)/cAMP response element-binding protein (CREB) to raise GPX4 and reduce malondialdehyde (MDA)/ROS/Fe2+,81 as well as AMPK/glycogen synthase kinase-3 beta (GSK-3β)/Nrf2 to upregulate SLC7A11/GPX4.15
Protective actions of anesthetics—cerebral I/R. Brain susceptibility reflects tight BBB-dependent iron control, PUFA-rich neuronal membranes, and glial contributions to redox/inflammation:
- Sevoflurane postconditioning downregulates the SP1/ACSL4 axis, curbing lipid peroxidation and ferroptosis.21
- Dexmedetomidine activates Nrf2 to restrain transferrin receptor 1 (TFR1) and lower Fe2+ while upregulating SLC7A11/GPX4 and supporting GSH synthesis;11 in parallel, it limits neuronal iron entry via the SOX9/DMT1 axis.12
- Sufentanil engages Akt/GSK-3β/Nrf2, suppressing oxidative stress, inflammation, and ferroptosis to protect the brain.43
Protection strategies by organ focus. Although both organs share common targets—iron overload, lipid peroxidation, GPX4 inactivation—their protective emphases differ.
Cardio-protection prioritizes mitochondrial restoration (metabolic-state switch) via convergent Nrf2 and Akt/mTOR signaling and actively promotes iron efflux (eg, via ferroportin (FPN)) to relieve I/R-driven overload.
Neuroprotection emphasizes precise neuronal iron control (DMT1/FPN modulation), BBB integrity, and neuroinflammation restraint. Across organs, the Nrf2/GPX4/SLC7A11 axis is the dominant protective pathway, while ACSL4 inhibition broadly limits pro-ferroptotic lipid generation.
Adjunct and novel approaches. Beyond leveraging anesthetic pharmacology, direct ferroptosis targeting adds translational options. Iron chelators (eg, deferoxamine) and ferroptosis inhibitors (eg, ferrostatin-1) improve mitochondrial performance and systolic/diastolic function in iron-overload cardiac models, validating a causal role for ferroptosis and nominating it as a therapeutic axis.46 Non-pharmacologic modulation, such as enriched environment (EE), can reduce interleukin-6 (IL-6)–hepcidin signaling during cerebral I/R and rebalance neuronal iron transporters (↓DMT1, ↑FPN1), correcting overload and attenuating ferroptosis.47 Natural compounds (eg, astragaloside IV, luteolin) that recalibrate iron handling and oxidative stress further illustrate pathway tractability for cardio-cerebral protection.84,85
Reconciling Conflicting Results: Why Findings Differ
Heterogeneity across studies reflects patient factors (age, sex, baseline iron status such as anemia/hemochromatosis, comorbidities), anesthetic exposure (agent class, concentration, duration), adjunct medications (opioids, steroids, sedatives), and species/model differences that incompletely reproduce human perioperative physiology. Integrating these variables reconciles contradictions within the molecular-switch framework: anesthetics act as regulators whose net effects are set by organ context (mitochondrial density, BBB status), iron/redox baselines, and pathway activation states.
Translational significance. Recognizing ferroptosis susceptibility enables perioperative risk stratification and personalized anesthesia. Cardiac protection centers on mitochondrial restoration and iron efflux; neuroprotection prioritizes neuronal iron homeostasis, BBB preservation, and inflammation control. Incorporating ferroptosis biomarkers, iron-sensitive imaging, and patient-specific profiles into perioperative practice—alongside targeted modulators and rational anesthetic combinations—offers a path to reducing complications and improving long-term outcomes.
Therapeutic Strategies and Future Directions
Despite the growing recognition of ferroptosis as a driver of cardio-cerebral injury, translating these insights into perioperative benefit requires strategies that selectively suppress detrimental ferroptosis in vulnerable organs while, in certain contexts (eg, oncology), permitting or enhancing pro-ferroptotic activity. Below, we integrate pharmacological and anesthetic approaches with delivery innovations, biomarker development, and research priorities, while addressing key translational challenges raised above.
Pharmacological Modulation of Ferroptosis
Direct ferroptosis modulators hold promise for perioperative protection, yet insufficient drug specificity remains a central hurdle. Canonical inhibitors such as ferrostatin-1 and liproxstatin-1 block lipid peroxidation and protect neurons/cardiomyocytes in preclinical I/R models,10 and iron chelators (eg, deferoxamine) reduce the labile iron pool and attenuate anesthetic-induced ferroptosis.13 Antioxidants (N-acetylcysteine, coenzyme Q10) and mitochondria-targeted ROS scavengers restore redox balance and complement anesthetic regimens.63 However, unresolved issues include (i) uncertain causality in specific compartments (eg, whether endothelial ferroptosis is the primary cause of BBB breakdown under hypoxia);67 (ii) suboptimal pharmacokinetics in vivo (low bioavailability/rapid metabolism) that blunt efficacy;86 and (iii) broad-spectrum antioxidant effects that may intersect with apoptosis or pyroptosis, confounding interpretation and benefit.87,88 Moreover, most current agents preferentially counter GPX4-dependent ferroptosis and may inadequately address GPX4-independent variants linked to mitochondrial dysfunction, yielding incomplete protection.89 Thus, next-generation modulators should improve tissue selectivity, engage organ-relevant nodes (eg, ACSL4, iron exporters), and extend coverage to GPX4-independent subtypes.
Anesthetic Choice and Personalized Perioperative Management
Evidence supports precision anesthesia to align agent selection with ferroptosis susceptibility. Dexmedetomidine and etomidate consistently bolster Nrf2/GPX4 signaling and normalize iron handling, making them appealing in high-risk cardio-cerebral settings.11,12,15,37 By contrast, propofol and sevoflurane display context-dependent effects—protective in I/R, but potentially pro-ferroptotic under hypoxia or in the developing brain8–10,21—necessitating careful dosing/timing. Risk stratification using iron indices (serum ferritin, transferrin saturation), functional GPX4 readouts, and exosomal biomarkers can guide selection and perioperative titration.
Importantly, anesthetic combinations can act additively or antagonistically on ferroptosis, and their net effect hinges on drug class, ratio, developmental stage, and pathology. Synergistic protection has been reported for dexmedetomidine + low-dose propofol in cerebral I/R and for etomidate + dexmedetomidine in ECT-related hippocampal injury, whereas propofol combined with high-concentration sevoflurane exacerbates developmental neurotoxicity; ketamine + sevoflurane may amplify cognitive deficits. These patterns suggest favoring dexmedetomidine + propofol in aging/ischemic brains while avoiding sevoflurane/ketamine + propofol in the developing brain. Systematic evaluation in organoid co-cultures and in vivo multiparameter monitoring is warranted to map combination-specific ferroptosis trajectories across contexts.
Novel Delivery Systems and Off-Target Risk Mitigation
A major translational barrier is the risk of off-target effects. Systemic iron chelation can perturb global iron balance and precipitate organ toxicities,90 while broad ferroptosis inhibition might dampen other programmed cell death pathways, potentially undermining antitumor efficacy when cytotoxicity is desired.91 Nanocarriers and prodrug strategies that are activated by pathologic cues (low pH, high ROS) offer spatiotemporal control to confine activity to ischemic or inflamed territories, reducing collateral effects. Exosome-based vectors that deliver antioxidant cargo or GPX4 mRNA across the BBB exemplify organ-addressable approaches. Stimuli-responsive nanoparticles can further ensure on-demand release precisely where ferroptotic pressure is highest.
The Molecular Switch Hypothesis
The molecular switch framework reconciles heterogeneity: anesthetics can toggle ferroptosis ON (eg, tumor contexts where pro-ferroptosis may aid therapy) or OFF (eg, heart/brain where suppression preserves function). Practically, this means calibrating agent class, dose, and timing to the organ’s iron/redox baseline and microenvironmental constraints (mitochondrial density in myocardium; BBB integrity and PUFA-rich membranes in brain). Multi-omics and single-cell analyses should be used to chart switch states and identify actionable nodes (Nrf2/GPX4/SLC7A11; ACSL4/LPCAT3; iron import/export). The schematic overview of the ferroptosis pathways and anesthetic regulation is illustrated in Figure 1. It integrates the three major axes—iron metabolism dysregulation, lipid peroxidation activation, and antioxidant defense collapse—highlighting how commonly used anesthetics act as molecular switches to bidirectionally modulate ferroptosis.
 |
Figure 1 Core Molecular Mechanisms of Ferroptosis and Bidirectional Regulation by Anesthetic Agents. The schematic summarizes the three central pathways of ferroptosis and the regulatory effects of commonly used anesthetics: (1) Iron metabolism dysregulation: Anesthetics regulate iron uptake and efflux through the TfR1/DMT1–NCOA4–FPN1 axis. Sevoflurane (Sevo) enhances TfR1 endocytosis and promotes intracellular iron overload, while dexmedetomidine (Dex) facilitates iron efflux by inhibiting HDAC2-mediated repression of FPN1. (2) Lipid peroxidation activation: ACSL4, LPCAT3, and LOXs drive polyunsaturated fatty acid (PUFA) incorporation and phospholipid peroxidation, with mitochondrial ROS from the Fenton reaction amplifying damage. Sevoflurane aggravates lipid ROS production by suppressing the Hippo–YAP/TAZ pathway, whereas MOR agonists attenuate ACSL4 expression via the HIF-1α/KCNQ1OT1 axis. (3) Antioxidant defense collapse: The System Xc− antiporter (SLC7A11/SLC3A2) imports cystine for glutathione (GSH) synthesis, enabling GPX4 to detoxify lipid peroxides (L-OOH). Propofol (Prop) protects by activating the Nrf2/GPX4/SLC7A11 axis, while ketamine (Ket) promotes ferroptosis through KAT5-mediated suppression of GPX4 transcription. Overall, anesthetic agents function as molecular switches, exhibiting context-dependent bidirectional regulation of ferroptosis—either inducing neuronal and cardiac injury or providing organ protection depending on tissue type, pathological state, and drug dose. Symbols: (1) Red arrows (↑/↓): Upregulation or downregulation of molecular targets. (2) Dashed red lines: Pro-ferroptotic regulation by anesthetics. (3) Dashed green lines: Anti-ferroptotic regulation by anesthetics. (4) Solid black arrows: Core ferroptosis pathways (iron metabolism, lipid peroxidation, antioxidant defense). (5) Hexagon with drug name: Specific anesthetic agent intervention (eg, Sevo, Prop, Ket, Dex). Abbreviations: TfR1, Transferrin receptor 1; DMT1, Divalent metal transporter 1; NCOA4, Nuclear receptor coactivator 4; FPN1, Ferroportin 1; HDAC2, Histone deacetylase 2; Dex, Dexmedetomidine; Sevo, Sevoflurane; MOR, μ-opioid receptor; HIF-1α, Hypoxia-inducible factor 1 alpha; KCNQ1OT1, Potassium voltage-gated channel subfamily Q member 1 opposite strand/antisense transcript 1; ACSL4, Acyl-CoA synthetase long-chain family member 4; LPCAT3, Lysophosphatidylcholine acyltransferase 3; LOXs, Lipoxygenases; PUFA, Polyunsaturated fatty acid; ROS, Reactive oxygen species; GPX4, Glutathione peroxidase 4; System Xc−, Cystine/glutamate antiporter (SLC7A11/SLC3A2); SLC7A11/SLC3A2, Solute carrier family transporters, subunits of System Xc−; GSH, Reduced glutathione; GSSG, Oxidized glutathione; GR, Glutathione reductase; Nrf2, Nuclear factor erythroid 2-related factor 2; Prop, Propofol; Ket, Ketamine; KAT5, Lysine; acetyltransferase 5; L-OOH, Lipid hydroperoxides; L-OH, Reduced lipid alcohols; LIP, Labile iron pool; PL-OOH, Phospholipid hydroperoxides; OH, Hydroxyl radical. |
Biomarker Development and Monitoring Challenges
Current markers—MDA,92 4-HNE,93 serum iron/ferritin—lack tissue and death-mode specificity. MDA and 4-HNE rise in diverse injuries (apoptosis, autophagy), limiting etiologic precision, while systemic iron indices do not localize myocardial or neuronal ferroptosis. To improve specificity, sensitivity, and feasibility:
- Exosomal GPX4: Quantifying GPX4 protein/activity in cell-of-origin exosomes (neuronal, cardiomyocyte, endothelial) may better reflect compartmental antioxidant status and ferroptosis susceptibility.
- Iron-sensitive imaging: Brain susceptibility-weighted MRI and cardiac T2* can non-invasively map iron burden; however, iron overload alone is insufficient for ferroptosis and may be confounded by other iron-laden states.
- Lipid peroxidation fingerprints: Mass-spectrometric profiling of phospholipid hydroperoxides (eg, arachidonic acid–phosphatidylethanolamine hydroperoxide (AA-PE-OOH)/adrenic acid–phosphatidylethanolamine hydroperoxide (AdA-PE-OOH) in cerebrospinal fluid (CSF) or plasma) offers a ferroptosis-proximal signature superior to bulk MDA/4-HNE.
- Tissue expression panels: ACSL4, GPX4, SLC7A11, FSP1 provide mechanistic specificity but are limited by biopsy invasiveness perioperatively.
- Molecular probe imaging: Long-term goals include probes that selectively recognize oxidized phospholipids, inactivated GPX4, or specific iron pools, paired with positron emission tomography (PET)/single-photon emission computed tomography (SPECT)/MRI contrast to visualize ferroptosis in vivo and in real time.
The overarching requirement is a toolbox that combines high specificity (vs other cell deaths), early sensitivity, (minimally) non-invasive sampling, and dynamic readouts that track ferroptosis progression perioperatively.
Future Research Priorities
Prospective clinical trials should validate anesthetic–ferroptosis interactions and incorporate imaging/biomarker endpoints.13,14 Advanced models—patient-derived organoids and organ-on-chip—can recapitulate cardiac and cerebral ferroptosis under anesthetic exposure. Artificial intelligence integrating clinical variables, omics layers, and anesthetic dosing may enable real-time risk prediction and decision support. Finally, long-term outcomes must be assessed to determine whether perioperative ferroptosis modulation favorably impacts cognition and cardiovascular health.
Therapeutic strategy should evolve from generic antioxidant use to precision perioperative medicine, tailoring anesthetic choice, combinations, and adjuncts to ferroptosis susceptibility while minimizing off-target risks. The molecular-switch framework provides a testable paradigm to guide mechanism-anchored interventions, delivery innovations, and biomarker development—advancing safe, targeted cardio-cerebral protection. Comprehensive cardio-cerebral protection strategies targeting ferroptosis and related clinical challenges are outlined in Table 4.
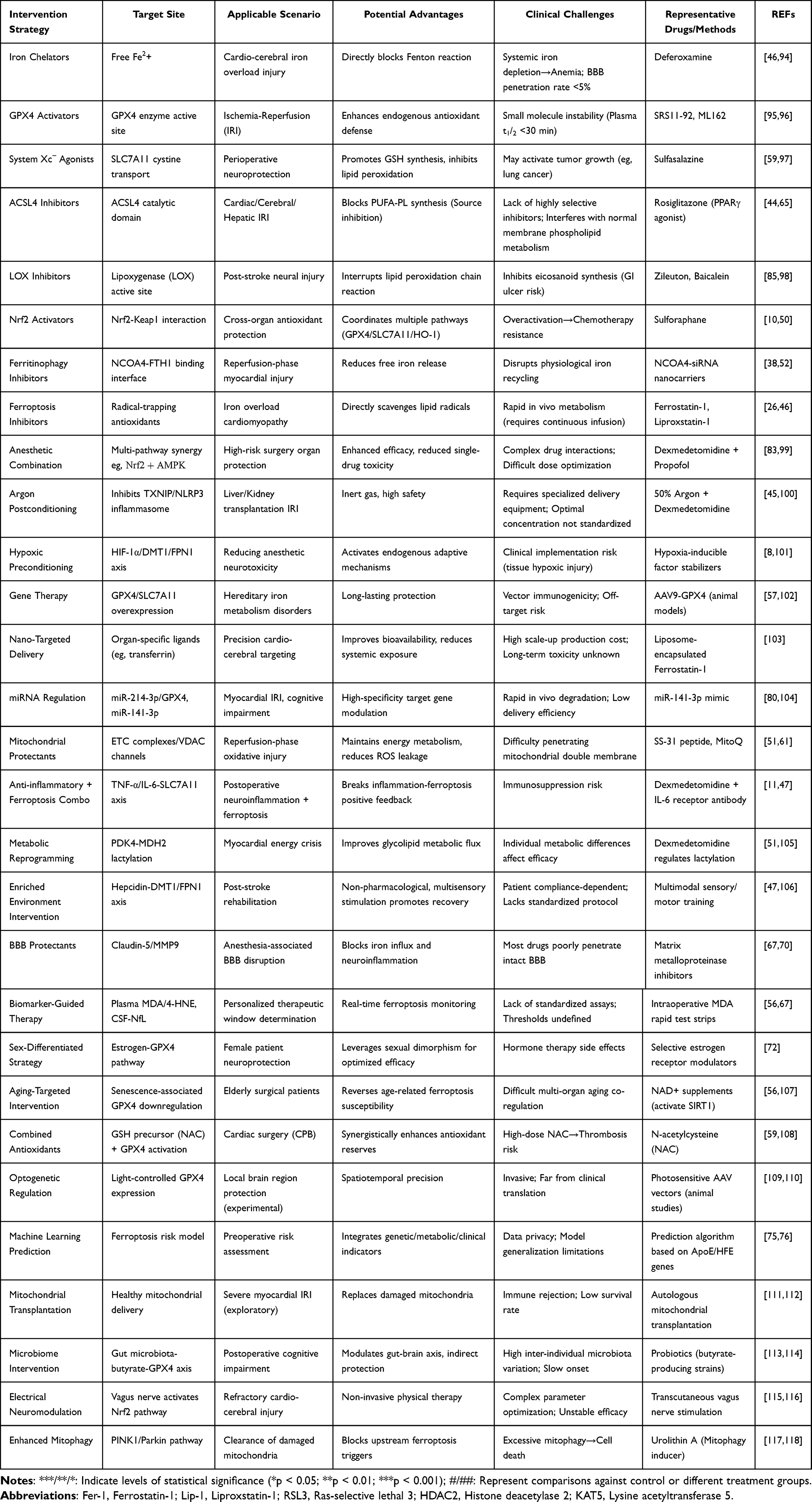 |
Table 4 Cardio-Cerebral Protection Strategies Targeting Ferroptosis and Clinical Challenges |
Conclusion
Ferroptosis has emerged as a pivotal pathway in perioperative organ injury, linking anesthetic exposure with neuronal and myocardial vulnerability. Over the last decade, studies have established that anesthetics can function as bidirectional modulators of ferroptosis, promoting or inhibiting it depending on developmental stage, tissue type, dose, and pathological context.
Three major conclusions can be drawn:
Organ-specific vulnerability: The brain and heart are especially prone to ferroptosis due to their high oxygen consumption, PUFA-rich membranes, and strict iron homeostasis requirements. These unique metabolic and structural features explain why anesthetic exposure can easily tip the balance toward ferroptosis-mediated injury.
Context-dependent anesthetic effects: Agents such as sevoflurane and propofol display dual activity—harmful in hypoxic or developmental states, yet protective in ischemia-reperfusion—whereas dexmedetomidine and etomidate consistently inhibit ferroptosis through the Nrf2/GPX4 and related antioxidant axes. This context-dependency reflects the influence of molecular “switches”, including receptor expression, metabolic status, and post-translational modifications.
Translational significance: Ferroptosis biomarkers (eg, GPX4, lipid peroxidation products, exosomal miRNAs) and imaging techniques (iron-sensitive MRI, cardiac T2*) now provide tangible bridges between preclinical discoveries and perioperative patient care. These advances open the door to biomarker-guided anesthesia strategies.
The molecular switch hypothesis offers a unifying framework: anesthetics toggle ferroptosis “on” in some contexts (eg, tumor tissues, where pro-ferroptotic effects may be beneficial) and “off” in others (eg, heart and brain, where ferroptosis suppression is protective). Understanding these switches at the molecular level—via iron metabolism, lipid peroxidation, and antioxidant signaling—will be critical for translating mechanistic insights into personalized anesthesia strategies.
Future efforts should focus on integrating multi-omics mapping, organoid modeling, and artificial intelligence (AI)-driven predictive platforms to capture ferroptosis dynamics across diverse perioperative scenarios. Prospective clinical trials are essential to validate ferroptosis-based perioperative risk stratification and to determine the efficacy of ferroptosis-targeting adjuvants such as iron chelators or GPX4 activators. Ultimately, incorporating ferroptosis regulation into anesthetic practice could shift perioperative medicine from descriptive observation to mechanism-driven, precision interventions, improving both short-term safety and long-term patient outcomes.
Funding
Supported by Science and Technology Department of Yunnan Province - Kunming Medical University, Kunming Medical joint special project - surface project, China, No. 202401AY070001-164; and Yunnan Provincial Department of Science and Technology Science and Technology Plan Project—Major Science and Technology Special Projects, China, No. 202405AJ310003; and the Yunnan Pan Xiangbin Expert Workstation under the Yunnan Provincial Project for Scientific and Technological Talents and Platforms Project, No. 202305AF150069.
Disclosure
Ying Chen and Jie Ouyang are co-first authors for this study. The authors report no conflicts of interest in this work.
References
1. Abudurousuli G, Xu S, Che J, Ding X, Gui B, Zhu L. Role of ferroptosis in effects of anesthetics on multiple organ diseases: a literature review. Heliyon. 2023;9:e20405. PMC10539942. doi:10.1016/j.heliyon.2023.e20405
2. Chen F, Kang R, Tang D, Liu J. Ferroptosis: principles and significance in health and disease. J Hematol Oncol. 2024;17:41. PMC11157757. doi:10.1186/s13045-024-01564-3
3. Costa I, Barbosa DJ, Benfeito S, et al. Molecular mechanisms of ferroptosis and their involvement in brain diseases. Pharmacol Ther. 2023;244:108373. doi:10.1016/j.pharmthera.2023.108373
4. Jiang X, Stockwell BR, Conrad M. Ferroptosis: mechanisms, biology and role in disease. Nat Rev Mol Cell Biol. 2021;22:266–282. PMC8142022. doi:10.1038/s41580-020-00324-8
5. Shu AH, Wang Q, Chen XB. Effect of different depths of anesthesia on postoperative cognitive function in laparoscopic patients: a randomized clinical trial. Curr Med Res Opin. 2015;31:1883–1887. doi:10.1185/03007995.2015.1075968
6. Liu H, Su B, Zhang Z, et al. Neonatal sevoflurane exposures inhibits DHHC5-mediated palmitoylation of TfR1 in oligodendrocytes, leading to hypomyelination and neurological impairments. J Adv Res. 2025. doi:10.1016/j.jare.2025.02.009
7. Song R, Wang R, Shen Z, Chu H. Sevoflurane diminishes neurogenesis and promotes ferroptosis in embryonic prefrontal cortex via inhibiting nuclear factor-erythroid 2-related factor 2 expression. Neuroreport. 2022;33:252–258. doi:10.1097/wnr.0000000000001775
8. Chen J, Xiao F, Chen L, et al. Role of ferroptosis in hypoxic preconditioning to reduce propofol neurotoxicity. Front Pharmacol. 2023;14:1121280. PMC9932196. doi:10.3389/fphar.2023.1121280
9. Liu L, Gao W, Yang S, et al. Ferritinophagy-mediated hippocampus ferroptosis is involved in cognitive impairment in immature rats induced by hypoxia combined with propofol. Neurochem Res. 2024;49:1703–1719. doi:10.1007/s11064-024-04128-6
10. Fan GB, Li Y, Xu GS, et al. Propofol inhibits ferroptotic cell death through the Nrf2/Gpx4 signaling pathway in the mouse model of cerebral ischemia-reperfusion injury. Neurochem Res. 2023;48:956–966. doi:10.1007/s11064-022-03822-7
11. Hu M, Men Y, Chen L, et al. Dexmedetomidine exerts its protective effect on cerebral ischemia reperfusion injury in mice by inhibiting ferroptosis. Zhong Nan Da Xue Xue Bao Yi Xue Ban. 2022;47:600–609. PMC10929910. doi:10.11817/j.issn.1672-7347.2022.210443
12. Liu Z, Liu Q, Zhang X, Li G. Dexmedetomidine inhibits ferroptosis by regulating the SRY-box transcription factor 9/divalent metal transporter-1 axis to alleviate cerebral ischemia/reperfusion injury. Chemical Biology & Drug Design. 2025;105:e70022. doi:10.1111/cbdd.70022
13. Fu Y, Wang Q, Wang D, Li Y. Dexmedetomidine inhibits ferroptosis to alleviate hypoxia/reoxygenation-induced cardiomyocyte injury by regulating the HDAC2/FPN pathway. Cardiovasc Drugs Ther. 2025. doi:10.1007/s10557-024-07664-z
14. Fang X, Ardehali H, Min J, Wang F. The molecular and metabolic landscape of iron and ferroptosis in cardiovascular disease. Nat Rev Cardiol. 2023;20:7–23. PMC9252571. doi:10.1038/s41569-022-00735-4
15. Wang Z, Yao M, Jiang L, et al. Dexmedetomidine attenuates myocardial ischemia/reperfusion-induced ferroptosis via AMPK/GSK-3β/Nrf2 axis. Biomed Pharmacother. 2022;154:113572. doi:10.1016/j.biopha.2022.113572
16. Tao WH, Shan XS, Zhang JX, et al. Dexmedetomidine attenuates ferroptosis-mediated renal ischemia/reperfusion injury and inflammation by inhibiting ACSL4 via α2-AR. Front Pharmacol. 2022;13:782466. PMC9307125. doi:10.3389/fphar.2022.782466
17. Dixon SJ, Lemberg KM, Lamprecht MR, et al. Ferroptosis: an iron-dependent form of nonapoptotic cell death. Cell. 2012;149:1060–1072. PMC3367386. doi:10.1016/j.cell.2012.03.042
18. Ma Y, Liu H, Ou Z, et al. DHHC5 facilitates oligodendrocyte development by palmitoylating and activating STAT3. Glia. 2022;70:379–392. doi:10.1002/glia.24113
19. Li J, Li J, Liu Y, et al. Nrf2 ameliorates sevoflurane-induced cognitive deficits in aged mice by inhibiting neuroinflammation in the hippocampus. Mol Neurobiol. 2025;62:8048–8064. doi:10.1007/s12035-025-04777-w
20. Zeng F, Zhou M, Li Q, Hu H, Chen C. Sevoflurane promotes neuronal ferroptosis via upregulation of PLIN4 to modulate the hippo signaling pathway. Neurotoxicology. 2024;105:1–9. doi:10.1016/j.neuro.2024.08.001
21. Lyu N, Li X. Sevoflurane postconditioning attenuates cerebral ischemia-reperfusion injury by inhibiting SP1/ACSL4-mediated ferroptosis. Hum Exp Toxicol. 2023;42:9603271231160477. doi:10.1177/09603271231160477
22. Cheng L, Zhu X, Liu Y, Zhu K, Lin K, Li F. ACSL4 contributes to sevoflurane-induced ferroptotic neuronal death in SH-SY5Y cells via the 5’ AMP-activated protein kinase/mammalian target of rapamycin pathway. Ann Transl Med. 2021;9:1454. PMC8506733. doi:10.21037/atm-21-4249
23. Zeng X, Jiang S, Wu Y, Zhong L, Liu X. Regulatory role of AMPK/Nrf2 signaling pathway in sevoflurane-enhanced intestinal protection against ischemia-reperfusion injury. Int J Biochem Cell Biol. 2024;177:106702. doi:10.1016/j.biocel.2024.106702
24. Cai Y, Wu Y, Guo Z, et al. Sevoflurane alleviates intestinal ischemia-reperfusion injury in aged mice. Med Gas Res. 2025;15:398–403. PMC12054681. doi:10.4103/mgr.MEDGASRES-D-24-00033
25. Liu P, Yuan J, Feng Y, Chen X, Wang G, Zhao L. Ferroptosis contributes to isoflurane-induced neurotoxicity and learning and memory impairment. Cell Death Discov. 2021;7:72. PMC8027876. doi:10.1038/s41420-021-00454-8
26. Zhang P, Shi X, He D, et al. Fer-1 protects against isoflurane-induced ferroptosis in astrocytes and cognitive impairment in neonatal mice. Neurotox Res. 2024;42:27. doi:10.1007/s12640-024-00706-2
27. Jin M, He K, Zhen S, et al. Asiaticoside enhances the effect of propofol on the invasion, ferroptosis and immune escape of bladder cancer. Drug Dev Res. 2024;85:e22242. doi:10.1002/ddr.22242
28. Sun F, He Y, Yang Z, et al. Propofol pretreatment inhibits ferroptosis and alleviates myocardial ischemia-reperfusion injury through the SLC16A13-AMPK-GPX4 pathway. Biomed Pharmacother. 2024;179:117345. doi:10.1016/j.biopha.2024.117345
29. Zhang B, Hou Q, Zhang X, et al. Anesthetic propofol inhibits ferroptosis and aggravates distant cancer metastasis via Nrf2 upregulation. Free Radic Biol Med. 2023;195:298–308. doi:10.1016/j.freeradbiomed.2022.12.092
30. Yu P, Zhang J, Ding Y, et al. Dexmedetomidine post-conditioning alleviates myocardial ischemia-reperfusion injury in rats by ferroptosis inhibition via SLC7A11/GPX4 axis activation. Hum Cell. 2022;35:836–848. doi:10.1007/s13577-022-00682-9
31. Zhang Y, Wei H, Wang M, et al. Dexmedetomidine alleviates ferroptosis following hepatic ischemia-reperfusion injury by upregulating Nrf2/GPx4-dependent antioxidant responses. Biomed Pharmacother. 2023;169:115915. doi:10.1016/j.biopha.2023.115915
32. z TX, Z L, H YX, G MX, W L, L YF. Effect of SLC7A11 in dexmedetomidine pretreatment induced reduction of ferroptosis caused by intestinal ischemia-reperfusion injury in mice. Chinese Journal of Clinical Pharmacology and Therapeutics. 2021;26:738–743. doi:10.12092/j.issn.1009-2501.2021.07.003
33. Li H, Liu W, Zhang X, Wu F, Sun D, Wang Z. Ketamine suppresses proliferation and induces ferroptosis and apoptosis of breast cancer cells by targeting KAT5/GPX4 axis. Biochem Biophys Res Commun. 2021;585:111–116. doi:10.1016/j.bbrc.2021.11.029
34. He GN, Bao NR, Wang S, Xi M, Zhang TH, Chen FS. Ketamine induces ferroptosis of liver cancer cells by targeting lncRNA PVT1/miR-214-3p/GPX4. Drug Des Devel Ther. 2021;15:3965–3978. PMC8458041. doi:10.2147/dddt.S332847
35. Ouyang J, Xiao W, Wu D, Bai M, Zhao Q, Li Y. Esketamine alleviated cardiomyocyte ferroptosis induced by oxygen-glucose deprivation/reoxygenation (OGD/R) via cyclic GMP-AMP synthase interactor. Cytotechnology. 2025;77:57. PMC11807036. doi:10.1007/s10616-025-00723-9
36. Shi J, Song S, Wang Y, et al. Esketamine alleviates ferroptosis-mediated acute lung injury by modulating the HIF-1α/HO-1 pathway. Int Immunopharmacol. 2024;142:113065. doi:10.1016/j.intimp.2024.113065
37. Lv Z, Wang F, Zhang X, Zhang X, Zhang J, Liu R. Etomidate attenuates the ferroptosis in myocardial ischemia/reperfusion rat model via Nrf2/HO-1 pathway. Shock. 2021;56:440–449. doi:10.1097/shk.0000000000001751
38. Chen L, Zhang LW, Pan XF, Liu X. Etomidate ameliorates ferroptosis and mitochondrial damage in myocardial ischemia/reperfusion injury. J Physiol Pharmacol. 2025;76. doi:10.26402/jpp.2025.1.01
39. Li X, Hu J, Zang X, et al. Etomidate improves the antidepressant effect of electroconvulsive therapy by suppressing hippocampal neuronal ferroptosis via upregulating BDNF/Nrf2. Mol Neurobiol. 2023;60:6584–6597. doi:10.1007/s12035-023-03499-1
40. Sun D, Li YC, Zhang XY. Lidocaine promoted ferroptosis by targeting miR-382-5p /SLC7A11 axis in ovarian and breast cancer. Front Pharmacol. 2021;12:681223. PMC8188239. doi:10.3389/fphar.2021.681223
41. Ma X, Yan W, He N. Lidocaine attenuates hypoxia/reoxygenation‑induced inflammation, apoptosis and ferroptosis in lung epithelial cells by regulating the p38 MAPK pathway. Mol Med Rep. 2022;25:150. doi:10.3892/mmr.2022.12666
42. Tan Y, Wang Q, Guo Y, et al. Dexmedetomidine mitigates lidocaine-induced spinal cord injury by repressing ferritinophagy-mediated ferroptosis by increasing CISD2 expression in rat models. J Bioenerg Biomembr. 2024;56:517–530. PMC11455791. doi:10.1007/s10863-024-10034-x
43. Zhu X, Han X, Wang J. Sufentanil-induced Nrf2 protein ameliorates cerebral ischemia-reperfusion injury through suppressing neural ferroptosis. Int J Biol Macromol. 2024;279:135109. doi:10.1016/j.ijbiomac.2024.135109
44. Zou S, Sun H, Peng Y, et al. Mu-opioid receptor alleviated ferroptosis in hepatic ischemia-reperfusion injury via the HIF-1α/KCNQ1OT1 axis. Am J Physiol Cell Physiol. 2023;324:C927–c40. doi:10.1152/ajpcell.00394.2022
45. Chen Q, Sun J, Liu X, et al. Dexmedetomidine and argon in combination against ferroptosis through tackling TXNIP-mediated oxidative stress in DCD porcine livers. Cell Death Discov. 2024;10:319. PMC11239900. doi:10.1038/s41420-024-02071-7
46. Kumfu S, Sripetchwandee J, Thonusin C, et al. Ferroptosis inhibitor improves cardiac function more effectively than inhibitors of apoptosis and necroptosis through cardiac mitochondrial protection in rats with iron-overloaded cardiomyopathy. Toxicol Appl Pharmacol. 2023;479:116727. doi:10.1016/j.taap.2023.116727
47. Luo Q, Zheng J, Fan B, Liu J, Liao W, Zhang X. Enriched environment attenuates ferroptosis after cerebral ischemia/reperfusion injury by regulating iron metabolism. Brain Res Bull. 2023;203:110778. doi:10.1016/j.brainresbull.2023.110778
48. Lin L, Ling X, Chen T, et al. Inhibition of hippocampal neuronal ferroptosis by liproxstatin-1 improves learning and memory function in aged mice with perioperative neurocognitive dysfunction. J Inflamm Res. 2025;18:2991–3007. PMC11880682. doi:10.2147/jir.S503784
49. Khan F, Verma M, Rautela I, Upadhye VJ, Pandey P, Kumar R. Correlation of neuroinflammation and therapeutic targets in perioperative neurocognitive disorders. Curr Pharm Biotechnol. 2024. doi:10.2174/0113892010315764240920064245
50. Wang M, Ren Z, Sun X, Li Y, Chen Z. Dexmedetomidine preconditioning attenuates ferroptosis in myocardial ischemia-reperfusion injury via α2 adrenergic receptor activation. Heliyon. 2024;10:e39697. PMC11544042. doi:10.1016/j.heliyon.2024.e39697
51. She H, Hu Y, Zhao G, et al. Dexmedetomidine ameliorates myocardial ischemia-reperfusion injury by inhibiting MDH2 lactylation via regulating metabolic reprogramming. Adv Sci (Weinh). 2024;11:e2409499. PMC11672254. doi:10.1002/advs.202409499
52. Qu L, Simayi A, Ma X, et al. Dexmedetomidine blocks NCOA4-dependent ferritinophagy to confer ferroptosis resistance in lung ischemia reperfusion injury via targeting NRF2. J Biochem Mol Toxicol. 2025;39:e70122. doi:10.1002/jbt.70122
53. Zhang M, Lyu D, Wang F, et al. Ketamine may exert rapid antidepressant effects through modulation of neuroplasticity, autophagy, and ferroptosis in the habenular nucleus. Neuroscience. 2022;506:29–37. doi:10.1016/j.neuroscience.2022.10.015
54. Wu J, Yang JJ, Cao Y, et al. Iron overload contributes to general anaesthesia-induced neurotoxicity and cognitive deficits. J Neuroinflammation. 2020;17:110. PMC7149901. doi:10.1186/s12974-020-01777-6
55. Zuo Y, Xie J, Zhang X, et al. Sevoflurane causes cognitive impairment by inducing iron deficiency and inhibiting the proliferation of neural precursor cells in infant mice. CNS Neuroscience & Therapeutics. 2024;30:e14612. doi:10.1111/cns.14612
56. Zhao L, Gong H, Huang H, Tuerhong G, Xia H. Participation of mind bomb-2 in sevoflurane anesthesia induces cognitive impairment in aged mice via modulating ferroptosis. ACS Chem Neurosci. 2021;12:2399–2408. doi:10.1021/acschemneuro.1c00131
57. Wang S, Wu Z, Bu X, et al. MEF2C alleviates postoperative cognitive dysfunction by repressing ferroptosis. CNS Neuroscience & Therapeutics. 2024;30:e70066. doi:10.1111/cns.70066
58. Miao M, Han Y, Wang Y, et al. Dysregulation of iron homeostasis and ferroptosis in sevoflurane and isoflurane associated perioperative neurocognitive disorders. CNS Neurosci Ther. 2024;30:e14553. PMC10853900. doi:10.1111/cns.14553
59. Soria FN, Pérez-Samartín A, Martin A, et al. Extrasynaptic glutamate release through cystine/glutamate antiporter contributes to ischemic damage. J Clin Invest. 2014;124:3645–3655. PMC4109556. doi:10.1172/jci71886
60. Liu H, Yan L, Niu H, Miao Z. Effects of glutathione tablets on ferroptosis pathway and oxidative stress-related indexes in serum of patients undergoing sevoflurane inhalation general anesthesia and its clinical significance. Altern Ther Health Med. 2024;30:249–255.
61. Takashi Y, Tomita K, Kuwahara Y, et al. Mitochondrial dysfunction promotes aquaporin expression that controls hydrogen peroxide permeability and ferroptosis. Free Radic Biol Med. 2020;161:60–70. PMC7530583. doi:10.1016/j.freeradbiomed.2020.09.027
62. Li S, Lei Z, Yang X, et al. Propofol protects myocardium from ischemia/reperfusion injury by inhibiting ferroptosis through the AKT/p53 signaling pathway. Front Pharmacol. 2022;13:841410. PMC8966655. doi:10.3389/fphar.2022.841410
63. Chen B, Fan P, Song X, Duan M. The role and possible mechanism of the ferroptosis-related SLC7A11/GSH/GPX4 pathway in myocardial ischemia-reperfusion injury. BMC Cardiovasc Disord. 2024;24:531. PMC11445876. doi:10.1186/s12872-024-04220-3
64. Qiu Y, Mo C, Xu S, et al. Research progress on perioperative blood-brain barrier damage and its potential mechanism. Front Cell Dev Biol. 2023;11:1174043. PMC10124715. doi:10.3389/fcell.2023.1174043
65. Gao R, Ou F, Lin J, Chen J, Wu Z. ACSL4 regulates LPS-induced ferroptosis in cardiomyocytes through FASN. Ann Clin Lab Sci. 2024;54:756–764.
66. Jing F, Wang Q, Xu Y, Dong J, Huang H, Li Y. MAZ regulates ferroptosis, apoptosis and differentiation of oligodendrocyte precursor cells. Brain Res. 2025;1849:149349. doi:10.1016/j.brainres.2024.149349
67. Liu Q, Song T, Chen B, Zhang J, Li W. Ferroptosis of brain microvascular endothelial cells contributes to hypoxia-induced blood-brain barrier injury. FASEB J. 2023;37:e22874. doi:10.1096/fj.202201765R
68. Ji M, Sun Q, Zhang G, et al. Microglia-derived TNF-α mediates Müller cell activation by activating the TNFR1-NF-κB pathway. Exp Eye Res. 2022;214:108852. doi:10.1016/j.exer.2021.108852
69. Liao S, Apaijai N, Luo Y, et al. Cell death inhibitors protect against brain damage caused by cardiac ischemia/reperfusion injury. Cell Death Discov. 2021;7:312. PMC8542034. doi:10.1038/s41420-021-00698-4
70. Längrich T, Bork K, Horstkorte R, et al. Disturbance of key cellular subproteomes upon propofol treatment is associated with increased permeability of the blood-brain barrier. Proteomes. 2022;10. PMC9397097. doi:10.3390/proteomes10030028
71. Chen SH, Yuan KC, Lee YC, et al. Iron and advanced glycation end products: emerging role of iron in androgen deficiency in obesity. Antioxidants (Basel). 2020;9. PMC7139764. doi:10.3390/antiox9030261
72. Shang Y, Cao T, Ma X, et al. Estrogen-induced FXR1 promotes endocrine resistance and bone metastasis in breast cancer via BCL2 and GPX4. Front Cell Dev Biol. 2025;13:1563353. PMC11973456. doi:10.3389/fcell.2025.1563353
73. Seo AY, Xu J, Servais S, et al. Mitochondrial iron accumulation with age and functional consequences. Aging Cell. 2008;7:706–716. PMC3849824. doi:10.1111/j.1474-9726.2008.00418.x
74. Jiang J, Li Y, Chen Y, Wu Q, Ding S. Ambient fine particulate matter induces cardiac fibrosis through triggering ferroptosis by heme degradation induced-iron overload. Ecotoxicol Environ Saf. 2025;297:118227. doi:10.1016/j.ecoenv.2025.118227
75. Katsarou MS, Latsi R, Papasavva M, et al. Population-based analysis of the frequency of HFE gene polymorphisms: correlation with the susceptibility to develop hereditary hemochromatosis. Mol Med Rep. 2016;14:630–636. PMC4918619. doi:10.3892/mmr.2016.5317
76. Cooper JG, Ghodsi M, Stukas S, Leach S, Brooks-Wilson A, Wellington CL. APOE ε4 carrier status modifies plasma p-tau181 concentrations in cognitively healthy super-seniors. Alzheimers Dement. 2024;20:4373–4380. PMC11180846. doi:10.1002/alz.13804
77. Chen W, Zhang Y, Wang Z, et al. Dapagliflozin alleviates myocardial ischemia/reperfusion injury by reducing ferroptosis via MAPK signaling inhibition. Front Pharmacol. 2023;14:1078205. PMC9986553. doi:10.3389/fphar.2023.1078205
78. Fang X, Cai Z, Wang H, et al. Loss of cardiac ferritin H facilitates cardiomyopathy via Slc7a11-mediated ferroptosis. Circ Res. 2020;127:486–501. doi:10.1161/circresaha.120.316509
79. Zhou RS, Xue XH, Bi Y, et al. Sevoflurane activates PI3K/AKT signaling pathway by upregulating GDF11 expression to attenuate ischemia/reperfusion injury in cardiomyocytes. Discov Med. 2024;36:2071–2078. doi:10.24976/Discov.Med.202436189.191
80. Zhu M, Yuan Z, Wen C, Wei X. DEX inhibits H/R-induced cardiomyocyte ferroptosis by the miR-141-3p/lncRNA TUG1 axis. Thorac Cardiovasc Surg. 2024. doi:10.1055/s-0044-1787691
81. Ma X, Xu J, Gao N, Tian J, Song T. Dexmedetomidine attenuates myocardial ischemia-reperfusion injury via inhibiting ferroptosis by the cAMP/PKA/CREB pathway. Mol Cell Probes. 2023;68:101899. doi:10.1016/j.mcp.2023.101899
82. Yang T, Feng X, Zhao Y, et al. Dexmedetomidine enhances autophagy via α2-AR/AMPK/mTOR pathway to inhibit the activation of NLRP3 inflammasome and subsequently alleviates lipopolysaccharide-induced acute kidney injury. Front Pharmacol. 2020;11:790. PMC7326938. doi:10.3389/fphar.2020.00790
83. Yang K, Kong X, Xie C, et al. Combined administration of dexmedetomidine and propofol mitigates myocardial ischemia/reperfusion injury by modulating the Akt/mTOR/Nrf2 axis to suppress ferroptosis. Eur J Pharmacol. 2025;997:177599. doi:10.1016/j.ejphar.2025.177599
84. Chen YH, Lin H, Wang Q, Hou JW, Mao ZJ, Li YG. Protective role of silibinin against myocardial ischemia/reperfusion injury-induced cardiac dysfunction. Int J Biol Sci. 2020;16:1972–1988. PMC7211181. doi:10.7150/ijbs.39259
85. Liu H, Zhao Z, Yan M, Zhang Q, Jiang T, Xue J. Calycosin decreases cerebral ischemia/reperfusion injury by suppressing ACSL4-dependent ferroptosis. Arch Biochem Biophys. 2023;734:109488. doi:10.1016/j.abb.2022.109488
86. Yu Z, Mo Z, Qiu Y, Lu H, Zheng B, Liu L. Emerging therapeutic approaches targeting ferroptosis in cancer: focus on immunotherapy and nanotechnology. Curr Pharm Biotechnol. 2024;25:2012–2021. doi:10.2174/0113892010276664231228124157
87. Wang X, Li W, Dong Y, et al. Ferrostatin-1 mitigates ionizing radiation-induced intestinal injuries by inhibiting apoptosis and ferroptosis: an in vitro and in vivo study. Int J Radiat Biol. 2023;99:1607–1618. doi:10.1080/09553002.2023.2194399
88. Dong HQ, Liang SJ, Xu YL, et al. Liproxstatin-1 induces cell cycle arrest, apoptosis, and caspase--3/GSDME-dependent secondary pyroptosis in K562 cells. Int J Oncol. 2022;61. PMC9450809. doi:10.3892/ijo.2022.5409
89. Zilka O, Shah R, Li B, et al. On the mechanism of cytoprotection by Ferrostatin-1 and Liproxstatin-1 and the role of lipid peroxidation in ferroptotic cell death. ACS Cent Sci. 2017;3:232–243. PMC5364454. doi:10.1021/acscentsci.7b00028
90. Nunes LLA, Dos Reis LM, Osorio R, et al. High ferritin is associated with liver and bone marrow iron accumulation: effects of 1-year deferoxamine treatment in hemodialysis-associated iron overload. PLoS One. 2024;19:e0306255. PMC11315289. doi:10.1371/journal.pone.0306255
91. Yang S, Xu Y, Ahmad T, et al. Mdivi-1 alleviates ferroptosis induced by hypoxia combined with propofol in HT22 cells by inhibiting excessive mitophagy. Mol Biol Rep. 2024;51:870. doi:10.1007/s11033-024-09812-y
92. Tsikas D. Assessment of lipid peroxidation by measuring malondialdehyde (MDA) and relatives in biological samples: analytical and biological challenges. Anal Biochem. 2017;524:13–30. doi:10.1016/j.ab.2016.10.021
93. Eskelinen M, Saimanen I, Koskela R, Holopainen A, Selander T, Eskelinen M. Plasma concentration of the lipid peroxidation (LP) biomarker 4-ηydroxynonenal (4-HNE) in benign and cancer patients. Vivo. 2022;36:773–779. PMC8931927. doi:10.21873/invivo.12764
94. Franklin ME, Grant JL, Lee GM, et al. Effects of iron accumulation and its chelation on oxidative stress in intracortical implants. Acta Biomater. 2025;200:703–723. PMC12183001. doi:10.1016/j.actbio.2025.05.026
95. Cheff DM, Huang C, Scholzen KC, et al. The ferroptosis inducing compounds RSL3 and ML162 are not direct inhibitors of GPX4 but of TXNRD1. Redox Biol. 2023;62:102703. PMC10149367. doi:10.1016/j.redox.2023.102703
96. Chen Y, He W, Wei H, et al. Srs11-92, a ferrostatin-1 analog, improves oxidative stress and neuroinflammation via Nrf2 signal following cerebral ischemia/reperfusion injury. CNS Neurosci Ther. 2023;29:1667–1677. PMC10173707. doi:10.1111/cns.14130
97. Sun S, Guo C, Gao T, et al. Hypoxia enhances glioma resistance to sulfasalazine-induced ferroptosis by upregulating SLC7A11 via PI3K/AKT/HIF-1α axis. Oxid Med Cell Longev. 2022;2022:7862430. PMC9699746. doi:10.1155/2022/7862430
98. Li M, Meng Z, Yu S, et al. Baicalein ameliorates cerebral ischemia-reperfusion injury by inhibiting ferroptosis via regulating GPX4/ACSL4/ACSL3 axis. Chem Biol Interact. 2022;366:110137. doi:10.1016/j.cbi.2022.110137
99. Zhou Y, Yang Y, Yi L, Pan M, Tang W, Duan H. Propofol and dexmedetomidine ameliorate endotoxemia-associated encephalopathy via inhibiting ferroptosis. Drug Des Devel Ther. 2024;18:1349–1368. PMC11055548. doi:10.2147/dddt.S458013
100. Schmitz SM, Dohmeier H, Stoppe C, et al. Inhaled argon impedes hepatic regeneration after ischemia/reperfusion injury in rats. Int J Mol Sci. 2020;21. PMC7432339. doi:10.3390/ijms21155457
101. Lv J, Liang Y, Tu Y, Chen J, Xie Y. Hypoxic preconditioning reduces propofol-induced neuroapoptosis via regulation of Bcl-2 and Bax and downregulation of activated caspase-3 in the hippocampus of neonatal rats. Neurol Res. 2018;40:767–773. doi:10.1080/01616412.2018.1477545
102. Weber-Adrian D, Kofoed RH, Silburt J, et al. Systemic AAV6-synapsin-GFP administration results in lower liver biodistribution, compared to AAV1&2 and AAV9, with neuronal expression following ultrasound-mediated brain delivery. Sci Rep. 2021;11:1934. PMC7820310. doi:10.1038/s41598-021-81046-5
103. Carlos A, Mendes M, Cruz MT, Pais A, Vitorino C. Ferroptosis driven by nanoparticles for tackling glioblastoma. Cancer Lett. 2024;611:217392. doi:10.1016/j.canlet.2024.217392
104. Liu F, Jiang LJ, Zhang YX, et al. Inhibition of miR-214-3p attenuates ferroptosis in myocardial infarction via regulating ME2. Biochem Biophys Res Commun. 2023;661:64–74. doi:10.1016/j.bbrc.2023.04.031
105. Zhu J, Zhang Y. Dexmedetomidine inhibits the migration, invasion, and glycolysis of glioblastoma cells by lactylation of c-myc. Neurol Res. 2024;46:1105–1112. doi:10.1080/01616412.2024.2395069
106. Deng YH, Dong LL, Zhang YJ, Zhao XM, He HY. Enriched environment boosts the post-stroke recovery of neurological function by promoting autophagy. Neural Regen Res. 2021;16:813–819. PMC8178758. doi:10.4103/1673-5374.297084
107. Bloomer SA. Hepatic macrophage abundance and phenotype in aging and liver iron accumulation. Int J Mol Sci. 2022;23. PMC9223835. doi:10.3390/ijms23126502
108. Schmitt B, Vicenzi M, Garrel C, Denis FM. Effects of N-acetylcysteine, oral glutathione (GSH) and a novel sublingual form of GSH on oxidative stress markers: a comparative crossover study. Redox Biol. 2015;6:198–205. PMC4536296. doi:10.1016/j.redox.2015.07.012
109. Taylor NK, Guggenbiller MJ, Mistry PP, King OD, Harper SQ. A self-complementary AAV proviral plasmid that reduces cross-packaging and ITR promoter activity in AAV vector preparations. Mol Ther Methods Clin Dev. 2024;32:101295. PMC11320455. doi:10.1016/j.omtm.2024.101295
110. Boyd RF, Sledge DG, Boye SL, et al. Photoreceptor-targeted gene delivery using intravitreally administered AAV vectors in dogs. Gene Ther. 2016;23:223–230. PMC4840844. doi:10.1038/gt.2015.96
111. Guariento A, Doulamis IP, Duignan T, et al. Mitochondrial transplantation for myocardial protection in ex-situ‒perfused hearts donated after circulatory death. J Heart Lung Transplant. 2020;39:1279–1288. doi:10.1016/j.healun.2020.06.023
112. Sun X, Gao R, Li W, et al. Alda-1 treatment promotes the therapeutic effect of mitochondrial transplantation for myocardial ischemia-reperfusion injury. Bioact Mater. 2021;6:2058–2069. PMC7809100. doi:10.1016/j.bioactmat.2020.12.024
113. Chen Y, Ho CT, Zhang X. The regulatory mechanism of intermittent fasting and probiotics on cognitive function by the microbiota-gut-brain axis. J Food Sci. 2025;90:e70132. doi:10.1111/1750-3841.70132
114. Rajanala K, Kumar N, Chamallamudi MR. Modulation of gut-brain axis by probiotics: a promising anti-depressant approach. Curr Neuropharmacol. 2021;19:990–1006. PMC8686316. doi:10.2174/1570159x19666201215142520
115. Li S, Stampas A, Frontera J, Davis M, Li S. Combined transcranial direct current stimulation and breathing-controlled electrical stimulation for management of neuropathic pain after spinal cord injury. J Rehabil Med. 2018;50:814–820. doi:10.2340/16501977-2379
116. Wang LC, Wei WY, Ho PC. Short-term cortical electrical stimulation during the acute stage of traumatic brain injury improves functional recovery. Biomedicines. 2022;10. PMC9405844. doi:10.3390/biomedicines10081965
117. Hou Y, Chu X, Park JH, et al. Urolithin A improves Alzheimer’s disease cognition and restores mitophagy and lysosomal functions. Alzheimers Dement. 2024;20:4212–4233. PMC11180933. doi:10.1002/alz.13847
118. Moradi N, Champsi S, Hood DA. Sulforaphane, Urolithin A, and ZLN005 induce time-dependent alterations in antioxidant capacity, mitophagy, and mitochondrial biogenesis in muscle cells. Sports Med Health Sci. 2025;7:16–27. PMC11624366. doi:10.1016/j.smhs.2024.03.011
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.