Back to Journals » OncoTargets and Therapy » Volume 13
Analysis of Key Genes Regulating the Warburg Effect in Patients with Gastrointestinal Cancers and Selective Inhibition of This Metabolic Pathway in Liver Cancer Cells
Authors Zhang X, Guo J, Jabbarzadeh Kaboli P ![]() , Zhao Q, Xiang S, Shen J, Zhao Y, Du F, Wu X, Li M, Ji H, Yang X, Xiao Z
, Zhao Q, Xiang S, Shen J, Zhao Y, Du F, Wu X, Li M, Ji H, Yang X, Xiao Z ![]() , Wen Q
, Wen Q
Received 12 April 2020
Accepted for publication 3 July 2020
Published 27 July 2020 Volume 2020:13 Pages 7295—7304
DOI https://doi.org/10.2147/OTT.S257944
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sanjeev K. Srivastava
Xinyue Zhang,1– 3,* Jinan Guo,4,5,* Parham Jabbarzadeh Kaboli,2,6 Qijie Zhao,2 Shixin Xiang,2 Jing Shen,2,6 Yueshui Zhao,2,6 Fukuan Du,2,6 Xu Wu,2,6 Mingxing Li,2,6 Huijiao Ji,2,6 Xiao Yang,2 Zhangang Xiao,2,6 Qinglian Wen1,3
1Department of Oncology, The Affiliated Hospital of Southwest Medical University, Luzhou, Sichuan, People’s Republic of China; 2Laboratory of Molecular Pharmacology, Department of Pharmacology, School of Pharmacy, Southwest Medical University, Luzhou, Sichuan, People’s Republic of China; 3Academician (Expert) Workstation of Sichuan Province, Luzhou, Sichuan, People’s Republic of China; 4The Department of Urology, The Second Clinical Medical College of Jinan University (Shenzhen People’s Hospital), The First Affiliated Hospital of South University of Science and Technology of China, Shenzhen Urology Minimally Invasive Engineering Center, Shenzhen, Guangdong, People’s Republic of China; 5Shenzhen Public Service Platform on Tumor Precision Medicine and Molecular Diagnosis, Shenzhen, Guangdong, People’s Republic of China; 6South Sichuan Institute of Translational Medicine, Luzhou, Sichuan, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Zhangang Xiao
Laboratory of Molecular Pharmacology, Department of Pharmacology, School of Pharmacy, Southwest Medical University, Luzhou 646000, Sichuan, People’s Republic of China
Email [email protected]
Qinglian Wen
Department of Oncology, The Affiliated Hospital of Southwest Medical University, Luzhou, Sichuan 646000, People’s Republic of China
Email [email protected]
Objective: The Warburg effect, also known as aerobic glycolysis, plays a dominant role in the development of gastrointestinal (GI) cancers. In this study, we analyzed the expression of key genes involved in the Warburg effect in GI cancers and investigated the effect of suppressing the Warburg effect in vitro in liver cancer cell lines.
Methods: The Cancer Genome Atlas (TCGA) RNA-Seq data were used to determine gene expression levels, which were analyzed with GraphPad Prism 7.00. Genetic alterations were queried with cBioPortal. The influence of the Warburg effect on liver cancer cell viability, migration and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) activity was determined by means of MTT, transwell and GAPDH activity assays.
Results: The levels of expression of genes associated with the Warburg effect were increased in tumors. To our knowledge, this is the first report of upregulated expression of CUEDC2, HMGB2, PFKFB4, PFKP and SIX1 in liver cancer. Clinically, overexpression of these genes was associated with significantly worse overall survival of liver cancer patients. In vitro, selective inhibition of GADPH suppressed the growth and metastasis of Huh-7, Bel7404 and Hep3B hepatocellular carcinoma cell lines.
Conclusion: The Warburg effect may play an important role in GI cancers, especially in liver cancer.
Keywords: Warburg effect, gastrointestinal cancers, liver cancer, bioinformatics, GAPDH
Introduction
According to GLOBOCAN, liver cancer is the sixth most common form of cancer in the world. Among the estimated 9.6 million cancer deaths that occurred in 2018, liver cancer was the second cause of death in men and was among the top five cancer types resulting in death, with an estimated 8.2% mortality rate for both genders.1,2 Liver cancer is a highly heterogeneous disease with a wide range of causes, including HBV, HCV, fungal toxins, alcohol consumption, obesity, and diabetes.2
The Warburg effect is a metabolic phenotype commonly seen in tumors. Even in the presence of sufficient oxygen, cancer cells produce about 60% of their ATP through glycolysis instead of oxidative phosphorylation.3,4 The Warburg effect allows dividing cells to use intermediate glucose metabolites to double their biomass and suppress apoptosis.5 This phenomenon was first discovered by Otto Warburg in the 1920s and was called aerobic glycolysis. A number of studies have shown that hypoxia-inducible factor 1a (HIF1A) and PI3K/AKT signaling regulate key enzymes of aerobic glycolysis and therefore modulate the Warburg effect in different cancers.6 It has been shown that AKT1, in response to cellular stress and drug treatments such as lapatinib, activates Nrf2 and HIF1A signaling in breast cancer.7 Altered HIF1A increases the chance of recurrence in patients suffering from HCC.8 In addition, aerobic glycolysis is significantly increased in liver cancer stem cells (CSCs). Nicotinamide adenine dinucleotide (NAD+) is required for increased activity of mitochondria. Higher levels of ribosomal protein S5 (MRPS5) and NAD+ dependent deacetylase sirtuin-1 (SIRT1) effectively increase the expression of glycolytic proteins and of the Warburg effect in liver CSCs.9 Similarly, glycolysis inhibitors such as hexokinase (HK) inhibitors, suppress tumor xenograft progression.10 In addition to HK isoenzymes, pyruvate dehydrogenase E1α (PDHA1) is another key enzyme involved in triggering aerobic glycolysis in HCC.11
On the other hand, there are other enzymes which reportedly act as positive regulators of the Warburg effect in liver or other cancer cells, so their inhibition might be an effective means of treating liver cancer (Figure 1). For example, metastasis-associated in colon cancer protein 1 (MACC1) and EGFR-phosphorylated platelet isoform of phosphofructokinase 1 (PFKP) enhance glycolysis via PI3K/AKT-dependent positive feedback regulation.12,13 Liver cancer cell metastasis and motility are reduced by inhibition of MACC1 expression.14 Transcription factor sine homeobox 1 (SIX1) is also a key enzyme involved in the regulation of glucose uptake, lactate production, ATP generation, and increased oxygen consumption rate (OCR).15
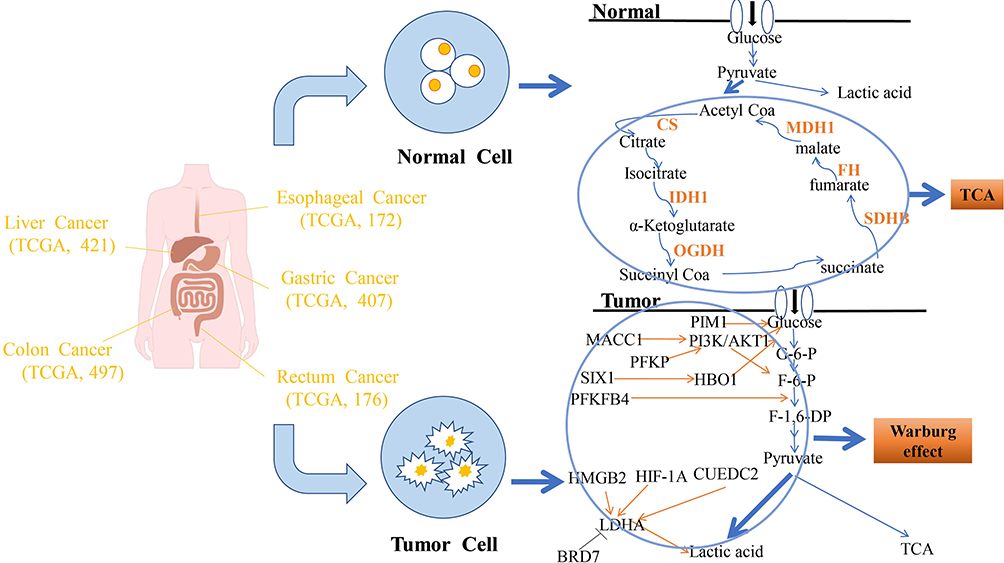 |
Figure 1 Schematic illustration of the roles played by the TCA cycle and the Warburg effect in gastrointestinal cancers and normal tissues. Normal tissues: The TCA cycle incorporates glucose metabolic products and transform them. Tumors: Under the influence of the Warburg effect, glucose metabolism is predominantly shunted towards pyruvate and lactic acid. PIM1, PI3K/AKT and HBO1 directly promote the conversion of glucose into G-6-P. PFKFB4 enhances the conversion of F-6-P into F-1, 6-DP. With the exception of BRD7, multiple factors can promote the production of lactic acid by LDHA. |
Cancer stem cells are responsible for drug resistance, so targeting cancer cell stemness is important to overcome drug-resistant phenotypes. Inhibition of SIX1 reduces stemness of HCC cells and therefore, sensitizes HCC cells to chemotherapy. SIX1 can bind to Sox2, which regulates stemness.16 In addition to Sox2, high-mobility-group protein 2 (HMGB2) is upregulated in liver cancer and is a key regulator of stem cell pluripotency.17 Proliferation of HCC cells depends on 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase-4 (PFKFB4) which is a key enzyme of glycolysis.18 Upregulation of Pim1 proto-oncogene (PIM1), a serine/threonine kinase, promotes glycolysis in HCC cells by enhancing Akt activation.19 CUE domain-containing 2 (CUEDC2) also plays a crucial role in promoting the Warburg effect by interacting with the glucocorticoid receptor and upregulating glucose transporter 3 (GLUT3) and lactate dehydrogenase A (LDHA), two key glycolytic proteins20; however, the effect of CUEDC2 on glycolysis in HCC cells has not been studied. Although aerobic glycolysis has been extensively studied, its precise details and mechanisms are not completely understood. The role of the Warburg effect in gastrointestinal cancers, especially in liver cancer, has not been systematically investigated. In addition to studying Warburg effect-related genes, we also compared the expression of genes involved in the tricarboxylic acid (TCA) cycle between normal and cancerous GI tissues.
Materials and Methods
Data and Samples
Clinical and gene expression data of patients with GI cancers were obtained from The Cancer Genome Atlas (TCGA), specifically from the TCGA-LIHC, TCGA-ESCA, TCGA-STAD, TCGA-READ and TCGA-COAD datasets, including information from both tumoral and normal tissues (number of samples: liver cancer, 421; esophageal cancer, 172; gastric cancer, 407; rectal cancer, 176; colon cancer, 497). Differential analysis of genes involved in the TCA (deep deletions, amplifications, and missense mutations) in GI cancer was performed by means of the cBioPortal for Cancer Genomics database (1700 samples). Protocols for data analysis were based on Zhao et al.21
Chemicals and Reagents
RPMI-1640, DMEM, FBS, antibiotics (penicillin and streptomycin, 0.25% w/v), trypsin, EDTA, and phosphate-buffered saline (PBS) were all obtained from Life Technologies (Grand Island, USA). Koningic acid (KA) was purchased from Cayman (Michigan, cat # 14,079). The GAPDH Activity Assay Kit was purchased from BioVision (Milpitas, cat # K680). 3-(4,5-Dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) was obtained from Tocris Bioscience (Bristol, cat # 5224/500).
Cell Lines and Cell Culture
HepG2, Hep3B, hepatoblastoma and HCC cell lines were purchased from the American Type Culture Collection (Manassas, USA). The Huh-7 HCC cell line was provided by Dr H. Nakabayashi (Hokkaido University School of Medicine, Japan).22 Bel740423 was obtained from the Cell Bank of the Chinese Academy of Sciences. MIHA-immortalized hepatocytes were obtained from Dr J.R. Chowdhury (Albert Einstein College of Medicine; New York, USA). The normal human liver cell line, LO2, was obtained from the cell bank of the Shanghai Institute of Cell Biology (Chinese Academy of Sciences; Shanghai, China). Use of these cell lines was approved by the ethics committee of Southwest Medical University. Cells were cultured in RPMI-1640 or DMEM with 10% fetal bovine serum (FBS) and antibiotics (100 U/mL streptomycin and 100 µg/mL penicillin) and incubated at 37°C in a humidified atmosphere with 5% CO2.
MTT Assay
Cell viability was analyzed by means of the MTT method. Briefly, after 12 hours of incubation to permit cell adherence, the experimental group was treated with 5 µM or 10 µM KA, whereas 0.01% dimethyl sulfoxide (DMSO) was added to the control group. After 24 hours, 10 µL MTT was added to each well. Following 4 hours of incubation, the solution was removed and DMSO was added. The absorbance at 490 nm was measured to calculate the average inhibition. The experiment was repeated three times with each cell line.
Transwell Migration Assay
Cell migration was measured by the transwell method, using 8-µm pore chambers (Corning, NY, cat # 3244). Briefly, different numbers of cells were seeded onto the upper chamber in serum-free medium. The lower chamber was filled with 600 µL of medium containing 1 µM KA (or not, in the control group). After 24 hours of incubation, the cells of the upper chamber were gently removed with cotton swabs, and the lower side of the membrane was dipped in 95% methanol and stained with crystal violet for 30 minutes. Five random fields were counted under a microscope.
GAPDH Activity Assay
Briefly, cells (106) were seeded onto 60mm dishes containing 1 µM KA or 0.01% DMSO (as control) and incubated for 24 hours. Trypsinized cells were subjected to the procedures described in the instructions of the GAPDH Activity Assay Kit and the protein content of the samples was determined. The absorbance was measured at 450 nm in kinetic mode for 30 minutes.
Statistical Analysis
TCGA data were first normalized and log2 transformed. Next, one-way ANOVA and the unpaired t-test were used to compare multiple groups and two groups, respectively, using SPSS 21.0 and GraphPad Prism 7.00 software programs. Overall survival was analyzed based on Kaplan–Meier curves using the Log rank test. P values <0.05 were considered statistically significant.
Results and Discussion
Expression of Genes Associated with the Warburg Effect
Cancer cells maintain high rates of glucose uptake, metabolism and fermentation to lactate.24 In this study, we conducted a systematic analysis of the expression of proteins, factors and enzymes associated with enhancement or inhibition of the Warburg effect in GI cancers. Our results indicate that the expression of genes associated with the Warburg effect are increased in tumors. To our knowledge, this is the first report of upregulated expression of CUEDC2, HMGB2, PFKFB4, PFKP, and SIX1 in liver cancer.
We analyzed the mRNA expression levels of genes associated with the Warburg effect in GI cancers using the TCGA database. The results confirmed a significant trend towards upregulation of all genes in tumors, especially in liver and esophageal cancer (Figure 2). In colon and gastric cancer, most genes showed similar results. In rectal cancer, expression of PFKFB4, SIX1 and MACC1 was significantly increased. It has recently been shown that SIX1 overexpression promotes HCC progression through downregulation of p53.25 SIX1 may increase resistance to chemotherapy in liver cancer.16,26
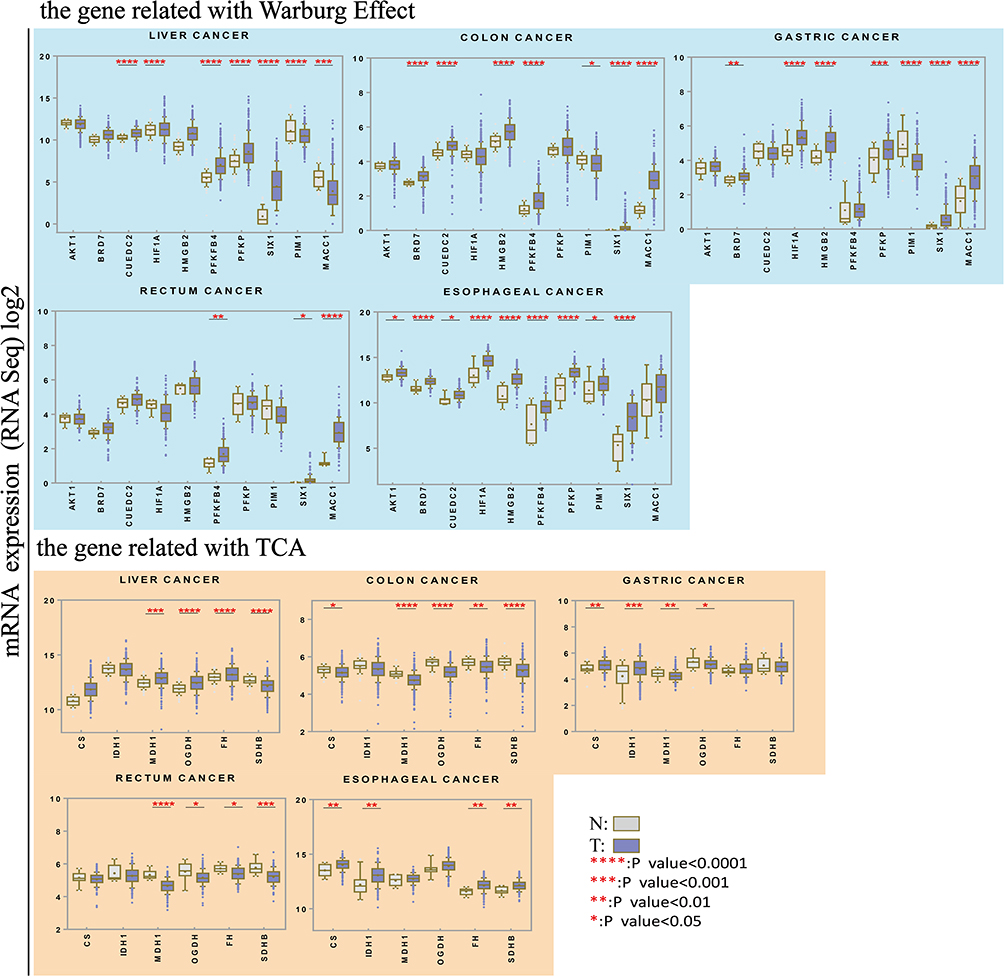 |
Figure 2 Expression of genes associated with the Warburg effect and the TCA cycle in GI cancers and normal tissues. Upper panel: Expression of genes associated with the Warburg effect. AKT1, BRD7, CUEDC2, HIF1A, HMGB2, PFKFB4, PFKP, PIM1, SIX1 and MACC1 expression in six different types of gastrointestinal cancers. TCGA RNA-Seq data was analyzed to compare expression between tumors (grey) and normal tissues (white). Lower panel: Expression of genes associated with the TCA cycle in tumoral and normal tissues. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.001. |
Under hypoxic conditions, PIM1 is upregulated, resulting in increased glucose uptake and facilitated glycolysis, which promotes tumor progression and metastasis.19 Downregulation of PIM1 suppressed tumor growth in HCC cells.27 We observed PIM1 expression was significantly decreased in gastric and liver cancer. It has been reported that inhibition of PIM kinases causes excessive mitochondrial fission or increased intracellular ROS production and apoptosis.28 Also, PIM1 knockdown significantly accelerated apoptosis in myoblasts.29 Therefore, PIM1 may have different roles in different cancer types.
Since BRD7 assists in the assembly of the p53 transcriptional complex, BRD7 is downregulated in several types of cancer. In contrast, we observed that BRD7 was upregulated in GI cancers, including liver cancer, which may be due to the wide range of mutations found in the GI cancers analyzed in this study. A previous study showed that poly-ubiquitinated HIF1A was upregulated in BRD7-overexpressing MCF-7 breast cancer cells, indicating that BRD7 promotes the degradation of HIF1A in an ubiquitination-dependent manner in breast cancer.30 Similarly, according to our data, HIF1A mRNA expression was upregulated, possibly revealing why BRD7 mRNA was highly expressed.
Furthermore, we found that MACC1, first reported to be a crucial biomarker of metastasis in colon cancer, was upregulated in GI cancers. In agreement with our findings, some studies have reported that MACC1 interacts with the ERK and AKT pathways in pancreatic cancer.31 It has also been reported that MACC1 is strongly associated with MET signaling in liver metastases of resected CRC and is involved in EMT in colorectal cancer.32,33 However, our analysis showed that it was downregulated in liver cancer. Until now, no studies have been conducted to try to understand the differential expression of MACC1 between primary and metastatic liver tumors.
Expression of Genes Related to the TCA Cycle
In addition to studying Warburg effect-associated genes, we also compared the expression of TCA-associated genes between normal and tumor tissues. As expected, expression of most genes was significantly higher in normal tissues than in GI tumor cells. Some genes showed high expression in colon and rectal cancers, but others showed an inverse pattern, including CS, SDHB, FH, and IDH1. Some studies have reported that CS activity is higher in human pancreatic ductal carcinoma than in adjacent, non-cancerous tissues.34 Next, we analyzed the genetic alterations found in the IDH1, FH and SDHB genes upregulated in GI cancers by means of cBioPortal (Supplementary Figure). Based on recent studies, mutations in the SDHB, IDH1, and FH genes are common and are associated with different kinds of tumors.35–37 Overall, these results probably explain why these genes are expressed at higher levels in tumors than in normal tissues.
Influence of the Warburg Effect on Clinical Progression
We analyzed the relationship between mRNA expression and clinical stages in GI cancer (Figure 3). The cancer stage codes were based on the American Joint Committee on Cancer definitions. Our results showed that high CUEDC2, HMGB2, MACC1, PFKFB4, and PFKP mRNA expression was associated with the worst stages of liver cancer. We also detected higher expression of MACC1, PFKFB4 and PIM1 in colon cancer patients with the worst stages. In contrast, expression of genes involved in the Warburg effect was not significantly associated with stages in other cancers. However, PIM1 expression was significantly increased in the more advanced stages of rectum cancer.
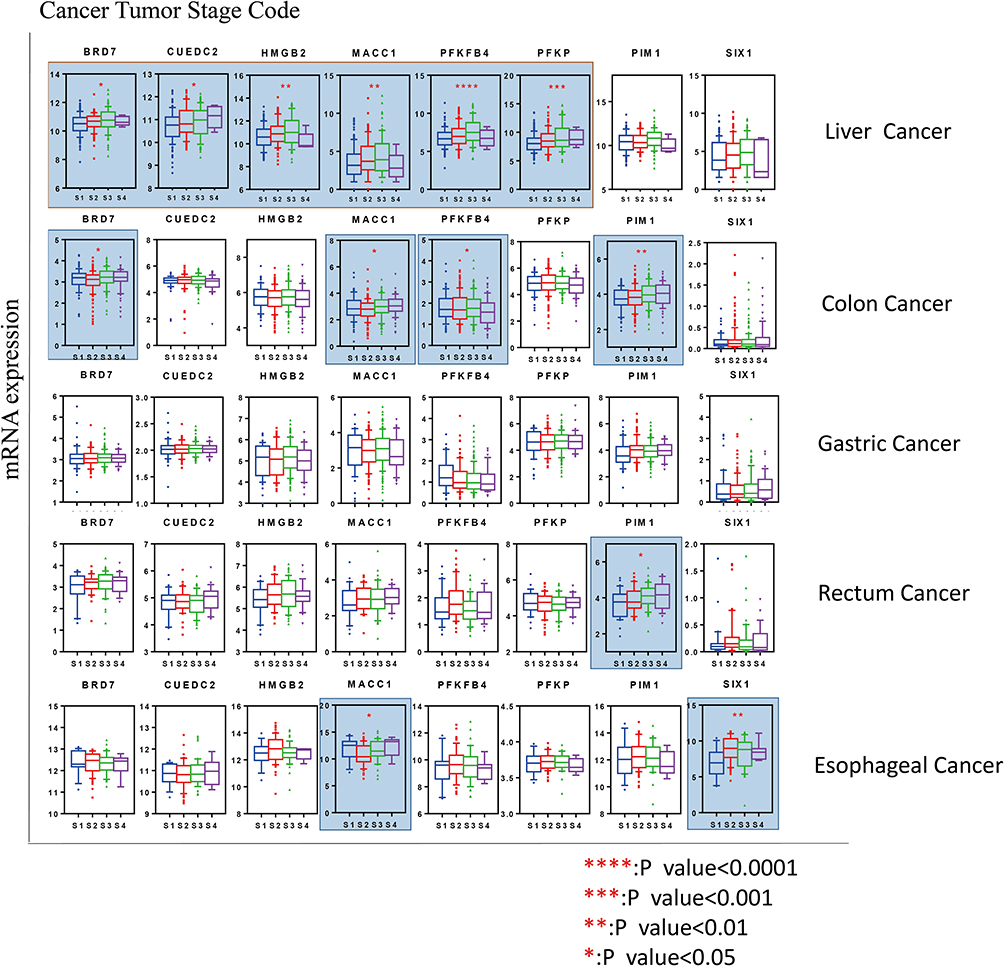 |
Figure 3 Association of genes linked to the Warburg effect with pathological stages. In cancers of the liver, esophagus, colon and rectum, expression of some genes was significantly associated with worse pathological stages. However, in gastric cancer, the correlation was not obvious. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.001. Abbreviations: S1, Stage-I; S2, Stage-II; S3, Stage-III; S4, Stage-IV. |
Furthermore, we analyzed the relationship between high or low expression of different genes in GI cancers and overall survival (OS) (Figure 4). Based on Kaplan–Meier curves and log-rank analysis, we found that high expression of CUEDC2, HMGB2, PFKFB4, PFKP, and SIX1 Warburg effect-associated genes was significantly associated with worse OS in patients with liver cancer. Collectively, these results suggest that the Warburg effect may play a role in the progression and outcome of GI and liver cancers. In agreement with our results, PFKP was previously reported as a marker of tumor progression and OS in liver cancer.38,39 The main problem encountered in the treatment of cancer is resistance to chemotherapy, which is crucially linked to cancer stem cells. We found that HMGB2 was strongly upregulated in all the cancers analyzed. Similarly, increased expression of HMGB2 has been reported in liver cancer.17 In addition, HMG members, including HMGB2, interact with hepatitis viruses (HBV and HCV) to modulate HCC progression.40,41
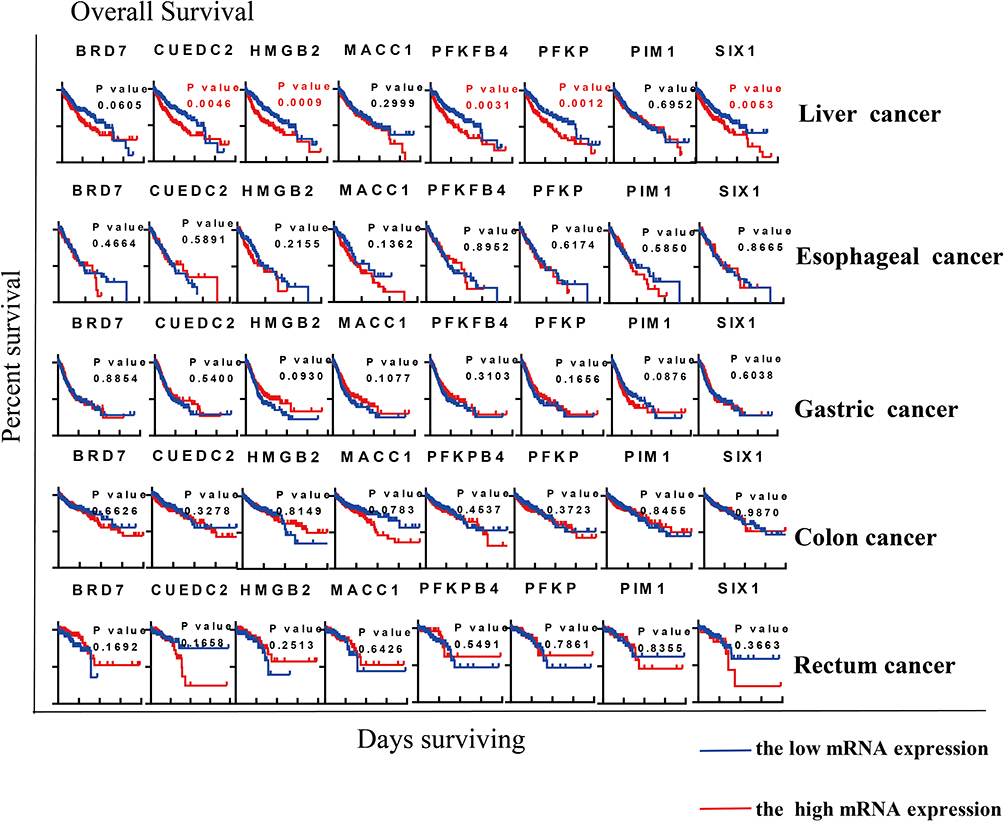 |
Figure 4 Association of genes linked to the Warburg effect with overall survival. The Warburg effect seemed to affect overall survival in liver cancer more than in any other type of cancer. High expression of genes was significantly associated with worse overall survival. High expression of CUEDC2, HMGB2, PFKFB4, PFKP and SIX1 was significantly associated with poor prognosis in liver cancer. Statistical analysis was carried out based on Kaplan–Meier curves. |
In addition to studying mRNA expression, we also investigated the influence of the Warburg effect on clinical parameters. The Warburg effect was identified as an essential factor associated with the proliferation, growth and metastasis of different tumors and with the tumor stage and OS. Patients with high expression of Warburg effect-associated genes showed more advanced stages and a poorer prognosis. This indicates that the Warburg effect may enhance tumor aggressiveness and worsen patient prognosis, particularly in liver cancer, and this result also has been demonstrated in other studies.42,43 Although the mechanism has not been investigated, increased expression of genes linked to the Warburg effect may be considered as a prognostic marker.
Inhibition of the Warburg Effect in Liver Cancer Cells
GAPDH promotes higher glycolysis in cells and KA has been shown to be an irreversible and selective inhibitor of GAPDH. Therefore, KA significantly reduces glycolysis and targets the Warburg effect in cancer cells44 (Figure 5A). Based on our bioinformatics results, we determined that liver cancer was more likely to be affected by key regulators of the Warburg effect than other GI cancers. In order to understand the influence of the Warburg effect in liver cancer cells, we used KA to suppress the Warburg effect by inhibiting GADPH. Based on the MTT assay, KA (5 µM and 10 µM) significantly inhibited the viability of Hep3B, Huh-7 and Bel7407 HCC cells (Figure 5B). Consistent with these results, migration of HepG2, Huh-7, Bel7407 and Hep3B cells was inhibited by treatment with 1 µM KA for 24 hours (Figure 5C). As shown in Figure 5D, GAPDH activity was strongly inhibited in liver cancer cells. We hypothesize that the explanation for these results is that KA inhibited the Warburg effect in liver cancer cells.
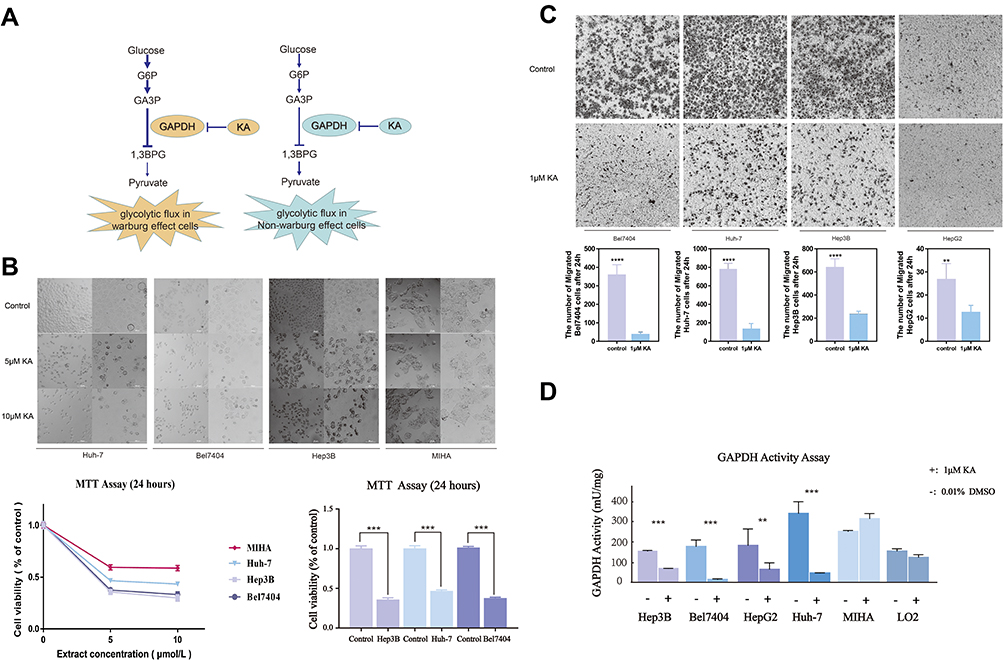 |
Figure 5 GAPDH inhibition suppresses the Warburg effect in HCC cells. (A) Diagram showing how KA-mediated inhibition of GAPDH interferes with glycolytic flux. (B) MIHA, Huh-7, Hep3B, and Bel7404 cell proliferation after GAPDH inhibition, measured with the MTT assay. Cellular viability in the treated groups (5 or 10 μmol/L KA for 24 h) was significantly decreased with respect to controls. (C) Representative microscopic fields showing migration of control cells and cells treated with 1μM KA. KA significantly suppressed migration of the four types of liver cancer cells. (D) GAPDH activity after treatment with 1μM KA or 0.01% DMSO (control). **P < 0.01, ***P < 0.001, ****P < 0.001. |
As expected, migration and growth of hepatocellular carcinoma cell lines were significantly inhibited after blocking the Warburg effect with the GAPDH inhibitor. According to some studies, the occurrence of liver cancer is indeed closely related to glycolysis.45,46 Since the liver is one of the three main metabolic centers in our body, the occurrence and development of malignant tumors in this organ may be closely linked to glucose metabolism. However, the influence of the Warburg effect in patients with liver cancer needs further investigation.
Conclusions
In conclusion, our analysis showed that genes associated with the Warburg effect were overexpressed in GI cancers. Our results also provided insight into the association between the Warburg effect and overall survival and clinical characteristics. Our study suggests that blocking the Warburg effect may be a promising approach to treat liver cancer.
Abbreviations
CS, citrate synthase; FH, fumarate hydratase; IDH1, isocitrate dehydrogenase (NADP (+))1, cytosolic; MDH1, malate dehydrogenase 1; OGDH, oxoglutarate dehydrogenase; SDHB, succinate dehydrogenase complex iron sulfur subunit B; ATK1, serine/threonine kinase 1; BRD7, bromodomain-containing 7; CUEDC2, CUE domain-containing protein 2; HIF1A, hypoxia-inducible factor 1 subunit alpha; HMGB2, high mobility group box protein 2; MACC1, metastasis associated in colon cancer 1; PFKFB4, 6-phosphofructo-2-kinase/fructose-2,6-biphosphatase 4; PFKP, platelet isoform of phosphofructokinase 1; PIM1, Pim-1 proto-oncogene, serine/threonine kinase; SIX1, sine oculis homeobox 1 transcription factor.
Acknowledgments
We would like to thank all of our colleagues at the Laboratory of Molecular Pharmacology, Department of Pharmacology, School of Pharmacy, Southwest Medical University, and at the Department of Oncology, Affiliated Hospital of Southwest Medical University for their assistance in this study.
Disclosure
The authors have no conflicts of interest to declare.
References
1. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre L, AJemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424. doi:10.3322/caac.21492
2. Grandhi MS, Kim AK, Ronnekleiv-Kelly SM, Kamel IR, Ghasebeh MA, Pawlik TM. Hepatocellular carcinoma: from diagnosis to treatment. Surg Oncol. 2016;25(2):74–85. doi:10.1016/j.suronc.2016.03.002
3. Gatenby R, AGillies RJ. Why do cancers have high aerobic glycolysis? Nat Rev Cancer. 2004;4(11):891–899. doi:10.1038/nrc1478
4. Lis P, Dylag M, Niedzwiecka K, Ko YH, Pedersen PL, Goffeau AUlaszewski S. The HK2 dependent “Warburg effect” and mitochondrial oxidative phosphorylation in cancer: targets for effective therapy with 3-bromopyruvate. Molecules. 2016;21(12):1730. doi:10.3390/molecules21121730
5. Jones RG, Thompson CB. Tumor suppressors and cell metabolism: a recipe for cancer growth. Genes Dev. 2009;23(5):537–548. doi:10.1101/gad.1756509
6. Courtnay R, Ngo DC, Malik N, Ververis K, Tortorella SM, Karagiannis TC. Cancer metabolism and the Warburg effect: the role of HIF-1 and PI3K. Mol Biol Rep. 2015;42(4):841–851. doi:10.1007/s11033-015-3858-x
7. Jabbarzadeh Kaboli P, Afzalipour Khoshkbejari M, Mohammadi M, et al. Targets and mechanisms of sulforaphane derivatives obtained from cruciferous plants with special focus on breast cancer - contradictory effects and future perspectives. Biomed Pharmacother. 2020;121:109635. doi:10.1016/j.biopha.2019.109635
8. Simon F, Bockhorn M, Praha C, et al. Deregulation of HIF1-alpha and hypoxia-regulated pathways in hepatocellular carcinoma and corresponding non-malignant liver tissue–influence of a modulated host stroma on the prognosis of HCC. Langenbecks Arch Surg. 2010;395(4):395–405. doi:10.1007/s00423-009-0590-9
9. Wei Z, Jia J, Heng G, et al. Sirtuin-1/mitochondrial ribosomal protein S5 axis enhances the metabolic flexibility of liver cancer stem cells. Hepatology. 2019;70(4):1197–1213. doi:10.1002/hep.30622
10. Xu SHerschman S, Herschman HR. A tumor agnostic therapeutic strategy for hexokinase 1–null/hexokinase 2–positive cancers. Cancer Res. 2019;79(23):5907–5914. doi:10.1158/0008-5472.CAN-19-1789
11. Sun J, Li J, Guo Z, et al. Overexpression of pyruvate dehydrogenase E1α subunit inhibits Warburg effect and induces cell apoptosis through mitochondria-mediated pathway in hepatocellular carcinoma. Oncol Res. 2019;27(4):407–414. doi:10.3727/096504018X15180451872087
12. Lee JH, Liu R, Li J, et al. EGFR-phosphorylated platelet isoform of phosphofructokinase 1 promotes PI3K activation. Mol Cell. 2018;70(2):197–210 e7. doi:10.1016/j.molcel.2018.03.018
13. Liu J, Pan C, Guo L, et al. A new mechanism of trastuzumab resistance in gastric cancer: MACC1 promotes the Warburg effect via activation of the PI3K/AKT signaling pathway. J Hematol Oncol. 2016;9(1):76. doi:10.1186/s13045-016-0302-1
14. Juneja M, Kobelt D, Walther W, et al. Statin and rottlerin small-molecule inhibitors restrict colon cancer progression and metastasis via MACC1. PLoS Biol. 2017;15(6):e2000784. doi:10.1371/journal.pbio.2000784
15. Yang X, Zhao H, Yang J, et al. MiR-150-5p regulates melanoma proliferation, invasion and metastasis via SIX1-mediated Warburg effect. Biochem Biophys Res Commun. 2019;515(1):85–91. doi:10.1016/j.bbrc.2019.05.111
16. Chen K, Wei H, Pan J, et al. Six1 is negatively correlated with poor prognosis and reduces 5-fluorouracil sensitivity via attenuating the stemness of hepatocellular carcinoma cells. Eur J Pharmacol. 2019;861:172599. doi:10.1016/j.ejphar.2019.172599
17. Zhao Y, Yang Z, Wu J, Wu R, Keshipeddy SK, Wright DWang L. High-mobility-group protein 2 regulated by microRNA-127 and small heterodimer partner modulates pluripotency of mouse embryonic stem cells and liver tumor initiating cells. Hepatol Commun. 2017;1(8):816–830. doi:10.1002/hep4.1086
18. Shu Y, Lu Y, Pang X, et al. Phosphorylation of PPARγ at Ser84 promotes glycolysis and cell proliferation in hepatocellular carcinoma by targeting PFKFB4. Oncotarget. 2016;7(47):76984–76994. doi:10.18632/oncotarget.12764
19. Leung CO, Wong CC, Fan DN, et al. PIM1 regulates glycolysis and promotes tumor progression in hepatocellular carcinoma. Oncotarget. 2015;6(13):10880–10892. doi:10.18632/oncotarget.3534
20. Zhong X, Tian S, Zhang X, et al. CUE domain-containing protein 2 promotes the Warburg effect and tumorigenesis. EMBO Rep. 2017;18(5):809–825. doi:10.15252/embr.201643617
21. Zhao Y, Zhao Q, Kaboli PJ, et al. m1A regulated genes modulate PI3K/AKT/mTOR and ErbB pathways in gastrointestinal cancer. Transl Oncol. 2019;12(10):1323–1333. doi:10.1016/j.tranon.2019.06.007
22. Nakabayashi H, Taketa K, Miyano K, Yamane TSato T, Sato J. Growth of human hepatoma cells lines with differentiated functions in chemically defined medium. Cancer Res. 1982;42(9):3858–3863.
23. Chen R, Zhu D, Ye X, Shen DLu D, Lu R. Establishment of three human liver carcinoma cell lines and some of their biological characteristics in vitro. Sci Sin. 1980;23(2):236–247.
24. Lu J. The Warburg metabolism fuels tumor metastasis. Cancer Metastasis Rev. 2019;38(1–2):157–164. doi:10.1007/s10555-019-09794-5
25. Cheng Q, Ning D, Chen J, Li X, Chen X, PJiang L. SIX1 and DACH1 influence the proliferation and apoptosis of hepatocellular carcinoma through regulating p53. Cancer Biol Ther. 2018;19(5):381–390. doi:10.1080/15384047.2018.1423920
26. Li B, Zhao S, Geng R, Huo ZZhang Z, Zhang H. The sineoculis homeobox Homolog 1 (SIX1) gene regulates paclitaxel resistance by affecting reactive oxygen species and autophagy in human hepatocellular carcinoma cell line HepG2. Med Sci Monit. 2018;24:2271–2279. doi:10.12659/MSM.906361
27. Pan XP, Wang HX, Tong DM, Li Y, Huang L, HWang C. miRNA-370 acts as a tumor suppressor via the downregulation of PIM1 in hepatocellular carcinoma. Eur Rev Med Pharmacol Sci. 2017;21(6):1254–1263.
28. Chauhan SS, Toth RK, Jensen CC, Casillas AL, Kashatus DF, Warfel NA. PIM kinases alter mitochondrial dynamics and chemosensitivity in lung cancer. Oncogene. 2020;39(12):2597–2611. doi:10.1038/s41388-020-1168-9
29. Liu Y, Shang Y, Yan Z, et al. Pim1 kinase positively regulates myoblast behaviors and skeletal muscle regeneration. Cell Death Dis. 2019;10(10):773. doi:10.1038/s41419-019-1993-3
30. Niu W, Luo Y, Wang X, et al. BRD7 inhibits the Warburg effect and tumor progression through inactivation of HIF1α/LDHA axis in breast cancer. Cell Death Dis. 2018;9(5):519. doi:10.1038/s41419-018-0536-7
31. Li Z, Yanfang W, Li J, et al. Tumor-released exosomal circular RNA PDE8A promotes invasive growth via the miR-338/MACC1/MET pathway in pancreatic cancer. Cancer Lett. 2018;432:237–250. doi:10.1016/j.canlet.2018.04.035
32. Wang Y, Zhao M, Zhao H, Cheng S, Bai RSong R, Song M. MicroRNA-940 restricts the expression of metastasis-associated gene MACC1 and enhances the antitumor effect of Anlotinib on colorectal cancer. Onco Targets Ther. 2019;12:2809–2822. doi:10.2147/OTT.S195364
33. Weidle UH, Birzele FKruger F, Krüger A. Molecular targets and pathways involved in liver metastasis of colorectal cancer. Clin Exp Metastasis. 2015;32(6):623–635. doi:10.1007/s10585-015-9732-3
34. Schlichtholz B, Turyn J, Goyke E, Biernacki M, Jaskiewicz K, Sledzinski ZSwierczynski J. Enhanced citrate synthase activity in human pancreatic cancer. Pancreas. 2005;30(2):99–104. doi:10.1097/01.mpa.0000153326.69816.7d
35. Valencia CA, López ML, Barreto AY, Rodríguez EG, Díaz CE. Hereditary leiomyomatosis and renal cell cancer syndrome: identification and clinical characterization of a novel mutation in the FH gene in a Colombian family. Fam Cancer. 2017;16(1):117–122. doi:10.1007/s10689-016-9922-4
36. Desideri E, Vegliante R, Ciriolo MR. Mitochondrial dysfunctions in cancer: genetic defects and oncogenic signaling impinging on TCA cycle activity. Cancer Lett. 2015;356(2 Pt A):217–223. doi:10.1016/j.canlet.2014.02.023
37. Raimundo N, Baysal BE, Shadel GS. Revisiting the TCA cycle: signaling to tumor formation. Trends Mol Med. 2011;17(11):641–649. doi:10.1016/j.molmed.2011.06.001
38. Cervantes-Anaya N, Ponciano-Gomez A, Lopez-Alvarez GS, et al. Downregulation of sorting nexin 10 is associated with overexpression of miR-30d during liver cancer progression in rats. Tumour Biol. 2017;39(4):1010428317695932. doi:10.1177/1010428317695932
39. Zhang Z, Li J, He T, et al. The competitive endogenous RNA regulatory network reveals potential prognostic biomarkers for overall survival in hepatocellular carcinoma. Cancer Sci. 2019;110(9):2905–2923. doi:10.1111/cas.14138
40. Dong ZZ, Yao M, Zheng WJ, Yao DF. [Clinical application of high mobility group box nuclear protein 1 subcomponent in the progression of liver cancer]. Zhonghua Gan Zang Bing Za Zhi. 2018;26(11):869–872. doi:10.3760/cma.j.issn.1007-3418.2018.11.014
41. Tang J, J X C, Chen L, et al. Metastasis associated in colon cancer 1 (MACC1) promotes growth and metastasis processes of colon cancer cells. Eur Rev Med Pharmacol Sci. 2016;20(13):2825–2834.
42. Kitamura K, Hatano E, Higashi T, et al. Proliferative activity in hepatocellular carcinoma is closely correlated with glucose metabolism but not angiogenesis. J Hepatol. 2011;55(4):846–857. doi:10.1016/j.jhep.2011.01.038
43. Beyoglu DIdle JR. The metabolomic window into hepatobiliary disease. J Hepatol. 2013;59(4):842–858. doi:10.1016/j.jhep.2013.05.030
44. Liberti MV, Dai Z, Wardell SE, et al. A predictive model for selective targeting of the Warburg effect through GAPDH inhibition with a natural product. Cell Metab. 2017;26(4):648–659 e8. doi:10.1016/j.cmet.2017.08.017
45. Beyoglu D, Imbeaud S, Maurhofer O, et al. Tissue metabolomics of hepatocellular carcinoma: tumor energy metabolism and the role of transcriptomic classification. Hepatology. 2013;58(1):229–238. doi:10.1002/hep.26350
46. Lee NCW, Carella MA, Papa SBubici C. High expression of glycolytic genes in cirrhosis correlates with the risk of developing liver cancer. Front Cell Dev Biol. 2018;6:138. doi:10.3389/fcell.2018.00138
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.