Back to Journals » Infection and Drug Resistance » Volume 15
Analysis of In Vitro Activity of Cefiderocol Against Carbapenem-Resistant Gram-Negative Bacilli by Broth Microdilution and Disk Diffusion Method: A Single-Center Study in Odisha, India
Authors Nayak G, Behera B ![]() , Mohanty S, Kar P, Jena J
, Mohanty S, Kar P, Jena J
Received 23 June 2022
Accepted for publication 14 September 2022
Published 10 October 2022 Volume 2022:15 Pages 5887—5897
DOI https://doi.org/10.2147/IDR.S378579
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Suresh Antony
Gayatree Nayak, Bijayini Behera, Srujana Mohanty, Punyatoya Kar, Jayanti Jena
Department of Microbiology, All India Institute of Medical Sciences [AIIMS], Bhubaneswar, Odisha, 751019, India
Correspondence: Bijayini Behera, Department of Microbiology, All India Institute of Medical Sciences [AIIMS], Bhubaneswar, Odisha, 751019, India, Tel +07750847562, Email [email protected]
Introduction: Cefiderocol (CFDC), a novel semi-synthetic siderophore cephalosporin has been developed to combat the menace of infections caused by carbapenem-resistant Gram-negative bacilli (CR-GNB) including Carbapenem-resistant Enterobacterales (CRE) and Carbapenem-resistant Nonfermenting Gram-negative bacilli (CR-NFGNB).
Methods: We determined the in vitro activity of CFDC against a contemporary collection of 503 CR-GNB isolates by the reference broth microdilution method (BMD) using Iron depleted cation adjusted Mueller-Hinton broth (ID-CAMHB). Performance of CFDC disk diffusion (DD) was evaluated against the reference BMD, as an alternative convenient testing method. Molecular characterization of carbapenemase in CR-GNB was performed by PCR targeting blaNDM-1, blaOXA-48like alleles, blaKPC, blaIMP, and, blaVIM. Minimum inhibitory concentration (MIC) distribution of CFDC in CR-GNB harbouring different carbapenemase enzymes was also analyzed.
Results: In our study, 81.7% (411/503) of CR-GNB isolates [81.3%, (278/342) CRE and 82.6% (133/161) CR-NFGNB] were susceptible to CFDC (p> 0.05). Categorical agreement (CA) of DD ranged from 79.8% to 87.5%, Minor error (mE) ranged from 0 to 14%, Major error (ME) ranged from 0 to 3.5%, and Very Major error (VME) ranged from 0 to 12.5% with variations by species tested. Overall CFDC MIC50 and MIC90 values of CR-GNB isolates without any carbapenemase genes were higher as compared to those with the presence of carbapenemase genes (4 μg/mL and 128 μg/mL versus 2 μg/mL and 16 μg/mL respectively).
Discussion: CFDC is not yet available for clinical use in India. Hence, multicentric studies are the need of the hour in India for standardization of CFDC susceptibility using disks and CAMHB from different manufacturers as well as understanding mechanisms of high MIC values.
Keywords: Cefiderocol, carbapenem-resistant Gram negative bacilli, broth microdilution, carbapenemases, disk diffusion, minimum inhibitory concentration
Introduction
The increasing prevalence of carbapenem-resistant Gram-negative bacilli (CR-GNB) is regarded as one of the greatest public health concerns across the globe with several CR-GNB, e.g., Carbapenem-resistant Enterobacterales (CRE), Carbapenem-resistant Pseudomonas aeruginosa (CRPA), and Carbapenem-resistant Acinetobacter baumannii (CRAB) being regarded as Priority I critical pathogens by the World Health Organization (WHO) due to existence of multiple resistance mechanisms and lack of effective antimicrobial agents against them.1
Cefiderocol (CFDC), a catechol substituted siderophore cephalosporin, has broad-spectrum activity against CR-GNB, including CRE, CRPA, and CRAB.2 The siderophore moiety of CFDC employs trojan horse like strategy allowing the entry of antimicrobial into the periplasmic space to reach the target site (PBPs) by active transport. CFDC displays structural stability to hydrolysis by both serine as well as various Metallo-β -lactamases.2 CFDC has completed Phase III clinical trial and is approved by the United States Food and Drug Administration (USFDA) for the treatment of complicated urinary tract infections (cUTIs), including pyelonephritis, Hospital-acquired pneumonia (HAP) or Ventilator-associated pneumonia (VAP), caused by designated susceptible GNB in adults and by European Medicines Agency (EMA) for the treatment of infections caused by aerobic GNB in adults with limited treatment options.3,4
Clinical and Laboratory Standards Institute (CLSI) had recommended broth microdilution (BMD) with minimum inhibitory concentration (MIC) determination as the only method for CFDC susceptibility testing in 2019 which requires iron-depleted culture media and subsequent cation adjustments to mimic the in vivo human plasma iron concentration.5 In the year 2020, CLSI also approved the disk diffusion (DD) method as the second method for the determination of CFDC susceptibility.6 In contrast to BMD, DD does not require iron depletion of standard Mueller-Hinton Agar (MHA) as iron is present in a bound state in the agar.6 Few studies have evaluated DD as an alternative suitable method to BMD for the determination of CFDC susceptibility.7,8 Moreover, there is no published literature regarding CFDC susceptibility from India. Hence, the present study was designed to determine CFDC susceptibility against a contemporary collection of CRE, CRPA, and CRAB isolates by BMD and DD. Simultaneously performance of the DD was also evaluated, taking BMD as the reference method. The MIC distribution of CFDC against CR-GNBs producing specific types of carbapenemase enzymes was analyzed.
Methods
Study Settings and Bacterial Isolates
This observational, cross-sectional study was conducted on 503 clinically significant, consecutive, non-duplicate CRE, CRPA, and CRAB isolates recovered from patients receiving medical care at a tertiary care, teaching, and referral hospital in Odisha. The isolates were collected prospectively for 2 years (Jan 2020–Dec 2021). GNB isolates testing resistant to any one of the Carbapenem group of antimicrobials (Ertapenem [only for CRE], Meropenem, Imipenem, Doripenem) were labelled as CR-GNB.9 CFDC susceptibility testing of the CR-GNB isolates was performed by both reference BMD as well as DD. The study was approved by the institutional ethics committee of All India Institute of Medical Sciences [AIIMS], Bhubaneswar, Odisha (Ref Number: IEC/AIIMS BBSR/PG-Thesis/2019-20/100 dated 14th December 2019).
MIC Determination, DD, and Interpretation
CFDC pure powder (CS-0016, CAS No: 122520S-94-5) was obtained from Chemscene India Pvt Ltd. CFDC stock solution (1000 μg/mL) was prepared in normal saline, and desired concentrations of working solutions (0.25–128 µg/mL) were made by twofold serial dilutions using normal saline as a diluent as per CLSI recommendations.5,6 Iron depleted Cation Adjusted Muller Hinton Broth (ID-CAMHB) was prepared as per CLSI recommendations.5,6 Briefly, 1g of iron-chelating agent Chelex® 100 sodium (Sigma Aldrich: C7901-25G) was added to 10 mL of 2X CAMHB (Sigma Aldrich:90922). The solution was continuously stirred with the help of a magnetic stirrer for 2 hrs at room temperature. Calcium (22.5 mg/L as Ca2+), magnesium (11.25 mg/L as Mg2+) and zinc (10 µM as ZnSO4) corresponding to 0.65 mg/L as Zn2+) were added to replenish the desired cations. The final concentrations of ions (iron, calcium, magnesium, and zinc) of the ID-CAMHB media were measured using atomic emission spectrometry (AAS) for iron and zinc and Inductive coupled plasma-optical emission spectrometry (ICP-OES) for calcium and magnesium. IDCAMHB after cation reconstitution was sterilized by 0.22µm hydrophilic polytetrafluoroethylene (PTFE) syringe filter and the final pH was adjusted to (7.2–7.4) with 1N hydrochloric acid. For BMD, to achieve 100 µL of volume in each well of the U bottom 96-well polystyrene plate (Tarson Micro Test Plate), 25 µL of drug, 25 µL inoculum of concentration (5 × 105 CFU/mL), and 50 µL of ID-CAMHB were added and incubated at 35 ± 2°C for 16 to 20 hours. MIC was read as the lowest concentration of CFDC that completely inhibited the growth of the organism in the wells as detected by the unaided eye. Trailing (multiple wells of tiny or faint growth relative to the growth control) was ignored while reading MIC. MIC50 and MIC90 values were calculated as per the standard method. DD was performed by using 30µg disk (FDC 30 µg, Liofilchem, s.r.l., Roseto Degli Abruzzi, Teramo, Italy) on unsupplemented standard Mueller-Hinton agar (MHA) (M1084 Hi-media Pvt Ltd, Mumbai, India). E. coli ATCC 25922 and P. aeruginosa ATCC 27853 (CFDC MIC range: 0.06–0.5µg/mL for both) were used as control strains for BMD and DD. CLSI breakpoints were used for the analysis of CFDC MICs and zone diameter.5,6
Agreement and Error Analysis
CFDC DD results were compared with MIC values for agreement and analysis of errors. CA was calculated as the percentage of isolates with results in the same category as the reference method, taking the total number of isolates tested as the denominator. Minor error (mE) was calculated as the percentage of isolates susceptible to CFDC by reference BMD method but in the intermediate category by the DD method or vice versa taking the total number of isolates tested as the denominator. Major error (ME) was calculated as the percentage of isolates showing MIC in the susceptible range by the reference BMD method and in the resistant range by DD method (false resistance by DD), taking total number of susceptible isolates by reference BMD method as the denominator. Very major error (VME) was calculated as the percentage of isolates showing MIC in the resistant range by reference BMD method and in the susceptible range by DD method (false susceptible by DD), taking total number of resistant isolates by BMD method as the denominator.
PCR for Carbapenemase Genes
Conventional uniplex PCR for five carbapenemase encoding genes (blaNDM-1, bla OXA−48like alleles, bla IMP, blaVIM, and blaKPC) were performed in 319 CR-GNB isolates. Standard published primers for five major carbapenemase encoding genes were used.10 CFDC MIC values of CR-GNB were stratified by the type of carbapenemase enzymes produced. Klebsiella pneumoniae ATCC BAA- 2146 and ATCC BAA-1705 were used as quality control strains for blaNDM and bla KPC respectively.
Results
Out of 503 CR-GNB isolates included in the present study, majority were CRE (342/503,68%) followed by CRAB (137/503, 27.2%) and CRPA (24/503,4.8%). Overall, E. coli was the most common isolate (183/503,36.4%) followed by K. pneumoniae (152/503,30.2%), A. baumannii complex (137/503, 27.2%), P. aeruginosa (24/503, 4.8%) and E. cloacae complex (7/503, 1.4%). Carbapenem-resistant E.coli isolates were recovered predominantly from urine (89/183, 48.6%) followed by blood samples (39/183, 21.3%) whereas Carbapenem-resistant K.pneumoniae were recovered predominantly from blood (68/152, 44.7%) followed by respiratory samples (39/152, 25.6%). The sample distribution of different genera of CR-GNB is depicted in Table 1.
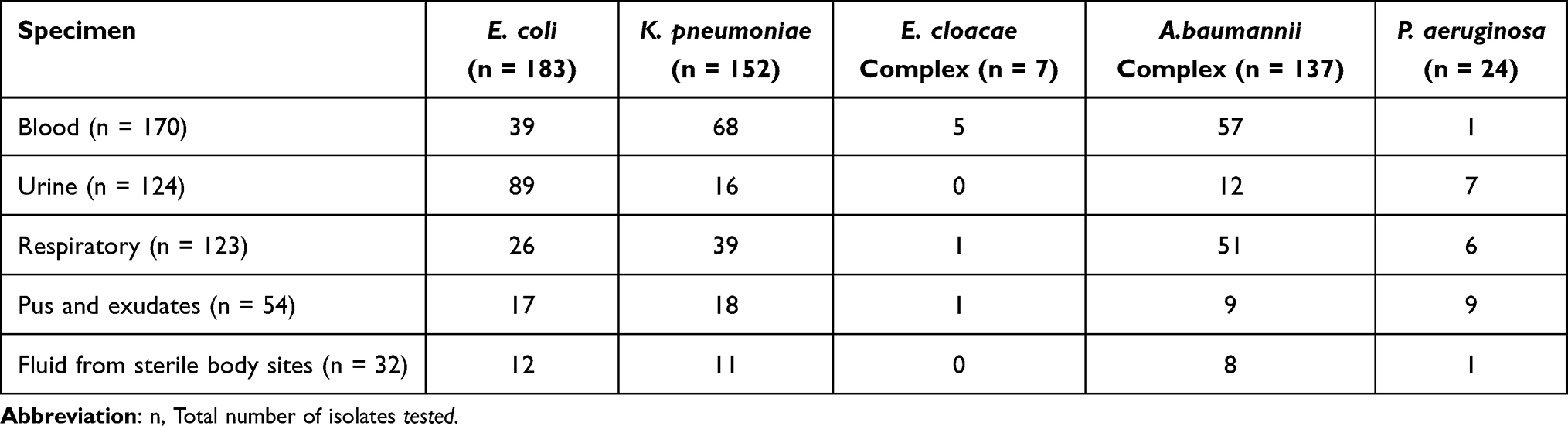 |
Table 1 Sample-Wise Distribution of Different Genera of Carbapenem-Resistant GNB (CR-GNB)(n=503) |
CFDC MIC Distribution of CR-GNB Isolates
CFDC MIC levels were highly variable depending on the CR- GNB species. The details of MIC distribution among 503 CR-GNB isolates are depicted in Table 2. 411/503 (81.7%) of CR-GNB isolates had CFDC MIC in the susceptible range, 48/503 (9.5%) had MICs in the intermediate range and 44 44/503 (8.7%) had MICs in the resistant category. At a susceptible breakpoint level of ≤ 4μg/mL, CFDC inhibited the growth of a higher percentage of NF-GNB (133/161, 82.6%) than Enterobacterales (278/342, 81. 3%), though the difference was not statistically significant (p = 0.69). Overall the most susceptible isolate was P. aeruginosa (21/24, 87.5%) and the least susceptible isolate was E. coli (144/183, 78.7%). Among CFDC susceptible Enterobacterales, K. pneumoniae was most susceptible (128/152, 84.2%, followed by E. coli (144/183, 78.7%) and among CFDC susceptible NF-GNB, P. aeruginosa (21/24, 87.5%) was more susceptible than A. baumannii complex (112/137, 81.7%). Out of the total 48 CR-GNB isolates in CFDC intermediate category, maximum isolates belonged to NF-GNB (18/161,11.2%) compared to Enterobacterales (30/342,8.8%). Out of a total of 44 CFDC-resistant CR-GNB, E. coli (23/44, 52.3%) was the highest in number followed by K. pneumoniae (10/44, 22.7%) and A. baumannii complex (8/44, 18.2%), whereas only (2/44, 4.5%) isolates of CRPA were resistant to CFDC. Overall CRE and CR-NFGNB had similar MIC50 and MIC90 values ie 2µg/mL and 8 µg/mL respectively. However, on genus-wise stratification of CRE, E. coli had MIC90 value of 32 µg/mL. MIC50 value P. aeruginosa (1 µg/mL) was lower than Acinetobacter baumannii complex (2 µg/mL).
 |
Table 2 CFDC MIC Distribution Against Different Genera of CR-GNB (n=503) |
CFDC DD of CR-GNB Isolates
In the present study, out of the 503 CR-GNB isolates tested by DD, 408 isolates (408/503, 81.1%) were susceptible, 51 isolates (51/503, 10.2%), were intermediate and 44 isolates (44/503, (8.7%) were resistant. CFDC susceptibility of NFGNB and CRE were (144/161,89.4%) and (264/342, 77.2%) respectively by DD. E. complex had 100% susceptibility to CFDC by DD, but the number of isolates were very less (7). On excluding E. complex, overall, CRAB (123/137, 89.8%) isolates were the most susceptible and CR E. coli (127/183, 69.4%) were the least susceptible to CFDC by DD. Across different genera which were resistant to CFDC, the highest number of resistant isolates belonged to E. coli (27/44, 61.4%) followed by K. pneumoniae (10/44, 22.7%) and A. baumannii complex (6/44, 13.6%). Only 1 isolate out of 24 CRPA and none of 7 E. cloacae complex was resistant to CFDC by DD. The distribution of CFDC MICs to zone diameters for Enterobacterales, A. baumannii complex, and P. aeruginosa are depicted in Figure 1.
 |
Figure 1 Distribution of CFDC MICs to zone diameters (A) for Enterobacterales, (B) for A. baumannii complex, (C), for P. aeruginosa). BMD MICs (µg/mL) are on the X axis, and zone diameters in (mm) are on the Y axis. The isolates highlighted in Green are in Categorical agreement (CA) by both the methods. Isolates showing VME are highlighted in red and ME are highlighted in brown color. Areas highlighted with grey contain isolates of Intermediate Category Isolates above (For BMD) and left (For DD) to Red bar are RESISTANT isolates. Isolates below (For BMD) and right (For DD) to Green bar are SUSCEPTIBLE isolates. |
Agreement and Error Analysis of CFDC BMD and DD
Agreement analysis of CFDC DD was done using BMD as the reference method. Categorical agreement (CA), mEs, MEs, and VMEs were assessed according to standard definitions. 90% of CA, 10% of mE, and 3% of ME and VME were considered acceptable based on CLSI guidelines. Two hundred eighty isolates of CRE (146 E. coli, 128 K. pneumoniae, and 6 E. cloacae complex) and 138 isolates of CR-NFGNB (117 A. baumannii complex, 21 P. aeruginosa) were in CA by both the methods. Collectively, CA ranged from 79.8% to 87.5%, mE ranged from 0 to 14%, ME ranged from 0 to 3.5%, and VME ranged from 0 to 12.5% (excluding E. cloacae complex), with variations by species tested. The number of isolates of E. cloacae complex was too low to be considered in the analysis. The CA of DD was higher in CR-NFGNB (85.7%) than CRE (81.9%). CA was lowest for CR E. coli (79.8%) and highest for CRPA (87.5%). However, for all the genera, CA of DD was below CLSI acceptable range (90%). mE of DD ranged from 0 to 13.6% and was almost similar for both CRE (12.6%) and CR-NFGNB (12.4%). The mE rate of DD for CRPA was 8.3% and in E. cloacae complex no mE was found. For all other organisms, the mE was beyond CLSI acceptable limit ie 10%. ME of DD ranged from 0–3.5% and was within CLSI acceptable range (<3%) for all genera, except for CR E. coli where ME was 3.5%. Six isolates of CRE showed ME, there were no ME in cases of CR-NFGNB. In contrast to mE, and ME, VME was higher in CR-NFGNB (10%) compared to CRE (8.8%). VME was lowest for CR E. coli (4.3%) Overall, VME was beyond CLSI acceptable range (<3%) for all CR-GNB, except for CRPA where there was no VME. Analysis of CFDC susceptibility results by BMD and DD, Agreement, and Error rates are summarised in Table 3.
 |
Table 3 Agreement and Error Rates of DD, Taking BMD as the Reference Method |
Genotypic Profile of Carbapenemases
Out of a total of 503 CR-GNB, genotypic analysis for five major carbapenemase encoding genes (blaNDM-1, bla OXA-48like allele, blaIMP, blaVIM, and blaKPC) could be performed in 319 isolates [201 CRE (85 E. coli, 110 K.pneumoniae, 6 E.cloacae complex), 103 CRAB and 15 CRPA]. Out of 319 isolates tested, 236 (236/319, 74%) CR-GNB isolates harbored one or more carbapenemase genes whereas 83(83/319, 26%) of CR-GNB isolates did not have any of the five tested carbapenemase genes. Among the carbapenemase producers, the highest number of isolates were NDM-1 producers (132/236, 55.9%) followed by combined NDM-1 and OXA48-like allele co-producers (76/236, 32.2%). None of the CR-GNB isolates harbored blaKPC, blaIMP, blaVIM. The majority of CR E. coli (60/85, 70.6%) and CRPA (8/15, 53.3%) were NDM-1 producers. However, 44.5% (49/110) of CRKP and 42.8% (3/7) of E.cloacae complex isolates harbored a combination of NDM-1 and OXA 48-like allele. In the majority of CRAB (47/103,71.2%) isolates, none of the five carbapenemase genes were detected.
CFDC Susceptibility by types of Carbapenemase
Among CRE, (22/27, 81.5%) of isolates harbouring blaOXA 48-like alleles were susceptible to CFDC, and (66/96, 68.7%) of isolates harbouring blaNDM-1 retained CFDC susceptibility. CRE isolates without the presence of any of the five carbapenemase genes had the least susceptibility (6/11, 54.5%) to CFDC. In case of CRAB, out of a total of 66 isolates that were not harboring any of the five carbapenemase genes, 47 (47/66,71.2%) isolates retained susceptibility to CFDC whereas out of the 37 isolates harboring one or more carbapenemase genes, 31 isolates (31/37,83.7%) were susceptible. On analysis of CFDC susceptibility based on the presence of a type of carbapenemase gene, CFDC susceptibility of CRAB isolates harboring blaNDM-1 was 85.7% (24/28) and the percentage susceptibility of isolates harboring blaNDM-1 and blaOXA 48-like allele was 75% (6/8). In case of CRPA, out of a total of 6 isolates that were not harboring any of the five carbapenemase genes, all (6/6100%) retained susceptibility to CFDC whereas out of the 9 isolates harboring one or more carbapenemase genes, 7 isolates were susceptible (7/9,77.8%). CFDC susceptibility of CRPA isolates harboring blaNDM-1 was 75%(6/8) and the percentage susceptibility of isolates harboring blaNDM-1 and blaOXA 48-like allelle was 100% (1/1). In cases of CRE isolates, out of a total of 190 isolates harboring any one or more carbapenemase gene/s 138 (138/190, 72.6%) isolates retained the susceptibility to CFDC whereas out of a total of 11 isolates that were not harbouring any of the five carbapenemase genes, 6 isolates were susceptible to CFDC (6/11,54.5%). CFDC susceptibility of CRE isolates harbouring blaNDM-1 was 68.7% (66/96), percentage susceptibility of isolates harbouring blaOXA 48-like alleles was 81.5% (22/27) and percentage susceptibility of isolates co-possessing blaNDM-1 and blaOXA 48-like alleles was 74.6% (50/67). In case of CFDC resistant CRE isolates (MIC ≥16 µg/mL), regardless of the species, isolates harbouring blaNDM-1 had a higher MIC distribution range, compared to CRE isolates co-producing blaNDM-1 and blaOXA 48-like alleles or blaOXA 48-like alleles alone. Furthermore, on analysing MIC50 and MIC90 values of specific groups of carbapenemase producers and carbapenemase non- producers it was found that MIC90 of Non -carbapenemase-producing CRE (Non-CP-CRE) (>128µg/mL) was 8-fold higher than carbapenemase-producing CRE (CP-CRE) (16µg/mL). Similarly, among CR-NFGNB, both MIC50 and MIC90 values of isolates (4 µg/mL,16µg/mL respectively) without any of the tested carbapenemase genes were two folds higher than isolates harbouring one or more carbapenemase encoding genes (2 µg/mL and 8 µg/mL respectively). [Table 4]
 |
Table 4 Distribution of MIC Across Different Carbapenemase Genes and Specific Groups of CR-GNB (n=319) |
Discussion
The present study demonstrates the performance of the CFDC susceptibility testing against CR-GNB isolates in an Indian tertiary care setting. In our study, 81.7% of CR-GNB isolates had CFDC MIC in the susceptible range. CFDC susceptibility of CRE isolates was 81.3% and of CR-NFGNB was 82.6%. In vitro susceptibility data of CFDC in three consecutive multinational surveillance studies (SIDERO-WT studies), covering over 28,000 Gram-negative bacteria [9205 clinical isolates in 2014–2015, 8954 in 2015–2016, and 10,470 in 2016–2017] had found CFDC susceptibility in >99% of isolates during each testing period.11–13 The SIDERO-WT studies included both carbapenem susceptible and non-susceptible isolates, whereas our study focussed only on CR-GNB isolates. Naas et al and Longshaw et al also investigated the in vitro activity of CFDC against GNB isolates from France and Europe respectively.14,15 In the study by Naas et al, overall, 99.0% of Enterobacterales and 99.7% of NF-GNB were CFDC susceptible.14 In the study by Longshaw et al, 81.6% of Enterobacterales, and 98.3% of P. aeruginosa were CFDC susceptible.15 Candel et al studied in vitro activity of CFDC in 11 European countries and the susceptibility rate of Carbapenem-resistant E. coli, Klebsiella spp., P. aeruginosa, Acinetobacter spp. to CFDC was 77.8%, 69.2%, 97.5% and 90.7% respectively.16 The CFDC susceptibility rate of CR-GNB isolates in our study is less than SIDERO-WT studies and matches the findings of Longshaw et al and Candel et al. These regional differences in CFDC susceptibility need further analysis.
In our study, among CRE isolates 85.7% (6/7) of Enterobacter cloacae complex, 84.2% (128/152) of K. pneumoniae, and 78.7% (144/183) of E. coli retained susceptibility to CFDC. However, in the studies by Longshaw et al and Candel et al Klebsiella spp. was the least susceptible isolate (82.8% and 69.2% respectively).15,16 In our study among CR- NFGNB, 87.5% (21/24) of P. aeruginosa and 81.7% (112/137) of A. baumannii complex were susceptible to CFDC. The percentage susceptibility rate of CRAB and CRPA isolates in our study is nearly similar to the findings of the study by Mushtaq et al in which 88.9% CRAB and 86.5% CRPA were susceptible to CFDC.17 In our study, out of 44 CFDC-resistant CR-GNB, E. coli (52.3%,23/44) was the highest in number followed by K. pneumoniae (22.7%,10/44). Similar to our findings, Iregui et al had described higher CFDC MICs in endemic E. coli and K. pneumoniae isolates.18 However, in the study by Ghebremedhin et al, the prevalence of CFDC resistance was reported to be more in CR-NFGNB (23.1% in CRAB and 18.9% in CRPA).19
In the present study CA of DD ranged from 79.8% to 87.5%, mE ranged from 0 to 14%,ME ranged from 0 to 3.5%, and VME ranged from 0 to 12.5% with variations by species tested. ME rates of all CR-GNB isolates were within the acceptable range of CLSI (3%) except for E.coli in which ME (3.5%) was beyond the acceptable range of CLSI. ME of CRE was 2.2% and no ME was observed in case of CR-NFGNB isolates. In our study, VME rates of all CR-GNB isolates were beyond the acceptable range of CLSI (3%) except for CRPA (no VME was found). Morris et al had compared the susceptibility results of CFDC DD against BMD in 58 CRE and 50 CRNF-GNB isolates using FDA-approved Hardy Disk and research-use-only (RUO) MASTDISCS and interpreted results by FDA, EUCAST, and investigational CLSI breakpoints (BPs).20 Agreement analysis and error analysis in their study varied between different disks, different species, and using different interpretative criteria. In a recent study by Bonnin et al, DD had 77% CA compared to BMD.21 The overall CA (83.1%) of DD in our study using Liofilchem RUO disk was lower than the overall CA (89%) reported by Morris et al but higher than the CA (77%) reported by Bonnin et al20,21 Our study used CAMHB and CFDC disk of single manufacturers. Further large-scale studies using disks and CA-MHB from different manufacturers are warranted to evaluate the utility of DD as an alternative to BMD for the determination of CFDC susceptibility.
In our study, overall CFDC MIC90 values of 503 CR-GNB isolates were 8 µg/mL. Among different genera of CRE, E. coli had the highest MIC90 value ie 32µg/mL, and MIC90 values of CRAB and CRPA were 8 µg/mL. These findings are nearly similar to the findings of the study by Hackel et al, which reported CFDC MIC90 values of CRE:4µg/mL, MDR A. baumannii:8µg/mL, MDR P. aeruginosa:1µg/mL, whereas the MIC90 values of isolates in the present study are higher than those described in SIDERO-WT studies.11–13,20 In SIDERO-WT studies, the MIC90 value of CFDC ranged from 0.25 to 1 μg/mL for Enterobacterales and 0.03 to 1 μg/mL for P. aeruginosa and 1 to 4 μg/mL for A. baumannii over three years, MIC90 values for Enterobacterales (0.5–1 μg/mL), P. aeruginosa (0.5 μg/mL) (0.25–0.5 μg/mL) did not witness any substantial change.11–13 However, for A. baumannii, the MIC90 values increased from 1 μg/mL to 4μg/mL in each consecutive year of the SIDERO-WT study.11–13 The reason for the high CFDC MIC of CR-GNB isolates in our study prior to drug exposure/usage needs additional investigation for the putative resistance mechanism.
In the present study, CFDC MIC50 and MIC90 values (4 and 128 µg/mL respectively) of CR-GNB isolates without five major carbapenemase genes were higher compared to MIC50 and MIC90 values (2 and 8µg/mL respectively) of CR-GNB isolates which harbored one or more carbapenemase gene. Among carbapenemase producers, blaNDM-1 harboring CR-GNB strains had two-fold higher MIC90 (16 µg/mL) than isolates bearing both NDM-1 and OXA48-like allele (8 µg/mL) or OXA48-like allele (8 µg/mL) alone. Additionally, MIC90 (>128µg/mL) of Non-CP-CRE was 8-fold higher than MIC90 (16µg/mL) of CP-CRE. Among NF-GNB, both MIC50 and MIC 90 values (4 µg/mL,16µg/mL respectively) of isolates harboring no carbapenemase encoding gene were two folds higher than MIC50 and MIC 90 values (2 µg/mL and 8 µg/mL respectively) of isolates harboring either one or more carbapenemase encoding genes. Morris et al also reported that the MIC90 value among non-carbapenemase-producing CRE (16 g/mL) was 4-fold higher than CP-CRE (4 g/mL).7 In another study by Kazmierczak et al, 35.7% of the NDM-positive isolates were not inhibited by ≤4 mg/L of CFDC.11 In SIDERO-WT and SIDERO-CR, MIC90 of blaNDM harboring CRE (8 µg/mL) was twofold higher than MIC90 (4 µg/mL) of CRE encoding other carbapenemase genes (bla OXA48, bla VIM, bla KPC).11–13,20 In non-carbapenemase producing E. coli and P. aeruginosa, mutations in outer membrane iron transporters (eg CirA and Fiu in E. coli and PiuA in P. aeruginosa) were associated with high CFDC MICs (≥16-fold increase).22,23 Additional mechanisms contributing to CFDC resistance in non-carbapenemase-producing GNB were not investigated in our study and thus cannot be commented upon.
Though a single-center study, this study is the first one to evaluate CFDC susceptibility in India, however, there are a few limitations. Only the five most common genes (bla NDM-1, bla OXA-48, bla KPC, blaIMP, blaVIM) encoding for Carbapenem resistance were studied in 319 of the 503 CR-GNB. Other mechanisms leading to Carbapenem resistance, e.g., porin loss and efflux pump mechanisms were not studied. We used RUO disk of CFDC and CAMHB from single manufacturers.
Conclusion
We found that 81.7% of CR-GNB isolates had CFDC MIC in the susceptible range which was lower than published international studies, where susceptibility rates were >90%. Higher CFDC MIC50 and MIC90 values were found in non-carbapenemase-producing isolates compared to carbapenemase-producing CR-GNB isolates. DD using RUO disk had 9.1% VME, 1.45% ME, and 12.5% mE. Further, a large-scale study with a greater number of isolates, disks, and CA-MHB from different manufacturers is required to evaluate DD as an alternative method of BMD. The genetic mechanism of CFDC resistance needed to be performed to elucidate the mechanisms leading to high MIC in CR-GNB isolates.
Acknowledgments
This work was partially supported by the Indian Council of Medical Research (ICMR) under MD thesis Grant [No.3/2/June-2020/PG-Thesis-HRD(33), dated 02.09.20. We sincerely thank Dr. Mamata Mohapatra, Principal Scientist,CSIR-IMMT for her support in the hydrometerological aspects of the study. We also acknowledge the technical assistance provided by Ms. Alaka Mahapatra and Miss Banani Dash.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Tacconelli E, Carrara E, Savoldi A, et al; WHO Pathogens Priority List Working Group. Discovery, research, and development of new antibiotics: the WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect Dis. 18;2018:318–327. doi:10.1016/S1473-3099(17)30753-3
2. Sato T, Cefiderocol: YK, Discovery C. In Vivo Profiles of a Novel Siderophore Cephalosporin. Clin Infect Dis. 2019;69:S538–43. doi:10.1093/cid/ciz826
3. Parsels KA, Mastro KA, Steele JM, Thomas SJ, Kufel WD. Cefiderocol: a novel siderophore cephalosporin for multidrug-resistant Gram-negative bacterial infections. J Antimicrob Chemother. 2021;76:1379–1391. doi:10.1093/jac/dkab015
4. Syed YY. Cefiderocol: a Review in Serious Gram-Negative Bacterial Infections. Drugs. 2021;81:1559–1571. doi:10.1007/s40265-021-01580-4
5. Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing; Twenty Ninth Informational Supplement. Wayne, PA: M100-S29. Clinical and Laboratory Standards Institute; 2019.
6. Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing; Thirtieth Informational Supplement. Wayne, PA: M100-S30. Clinical and Laboratory Standards Institute; 2020.
7. Morris CP, Bergman Y, Tekle T, Fissel J, Tamma PD, Simner PJ. Cefiderocol antimicrobial susceptibility testing against multidrug-resistant Gram-negative bacilli: a comparison of disk diffusion to broth microdilution. J Clin Microbiol. 2021;59:e01649–20.
8. Matuschek E, Longshaw C, Takemura M, Yamano Y, Kahlmeter G. Cefiderocol: EUCAST criteria for disc diffusion and broth microdilution for antimicrobial susceptibility testing. J Antimicrob Chemother. 2022;1:dkac080.
9. Kois AK, Nicolau DP, Kuti JL. Unresolved issues in the identification and treatment of carbapenem-resistant Gram-negative organisms. Curr Opin Infect Dis. 2020;33(6):482–494. doi:10.1097/QCO.0000000000000682
10. Veeraraghavan B, Shankar C, Karunasree S, et al. Carbapenem-resistant Klebsiella pneumoniae isolated from bloodstream infection: Indian experience. Pathog Glob Health. 2017;111:240–246. doi:10.1080/20477724.2017.1340128
11. Kazmierczak KM, Tsuji M, Wise MG, et al. In vitro activity of Cefiderocol, a siderophore cephalosporin, against a recent collection of clinically relevant carbapenem-non-susceptible Gram-negative bacilli, including serine carbapenemase- and metallo-β-lactamase-producing isolates (SIDERO-WT-2014 Study). Int J Antimicrob Agents. 2019;53:177–184. doi:10.1016/j.ijantimicag.2018.10.007
12. Hackel MA, Tsuji M, Yamano Y, et al. In Vitro Activity of the Siderophore Cephalosporin, Cefiderocol, against a Recent Collection of Clinically Relevant Gram-Negative Bacilli from North America and Europe, Including Carbapenem-Nonsusceptible Isolates (SIDERO-WT-2014 Study). Antimicrob Agents Chemother. 2017;61:e00093–17. doi:10.1128/AAC.00093-17
13. Karlowsky JA, Hackel MA, Tsuji M, et al. In Vitro Activity of Cefiderocol, a Siderophore Cephalosporin, Against Gram-Negative Bacilli Isolated by Clinical Laboratories in North America and Europe in 2015-2016: SIDERO-WT-2015. Int J Antimicrob Agents. 2019;53:456–466. doi:10.1016/j.ijantimicag.2018.11.007
14. Naas T, Lina G, Santerre Henriksen A, Longshaw C, Jehl F. In vitro activity of cefiderocol and comparators against isolates of Gram-negative pathogens from a range of infection sources: SIDERO-WT-2014-2018 studies in France. JAC Antimicrob Resist. 2021;3:dlab081. doi:10.1093/jacamr/dlab081
15. Longshaw C, Manissero D, Tsuji M, Echols R, Yamano Y. In vitro activity of the siderophore cephalosporin, Cefiderocol, against molecularly characterized, carbapenem-non-susceptible Gram-negative bacteria from Europe. JAC Antimicrob Resist. 2020;dlaa060. doi:10.1093/jacamr/dlaa060
16. Candel FJ, Santerre Henriksen A, Longshaw C, Yamano Y, Oliver A. In vitro activity of the novel siderophore cephalosporin, cefiderocol, in Gram-negative pathogens in Europe by site of infection. Clin Microbiol Infect. 2022;28(447):e1–447. doi:10.1016/j.cmi.2021.07.018
17. Mushtaq S, Sadouki Z, Vickers A, Livermore DM, Woodford N. In Vitro Activity of Cefiderocol, a Siderophore Cephalosporin, against Multidrug-Resistant Gram-Negative Bacteria. Antimicrob Agents Chemother. 2020;64:e01582–20. doi:10.1128/AAC.01582-20
18. Iregui A, Khan Z, Landman D, Quale J. Activity of Cefiderocol Against Enterobacterales, Pseudomonas aeruginosa, and Acinetobacter baumannii Endemic to Medical Centers in New York City. Microb Drug Resist. 2020;26:722–726. doi:10.1089/mdr.2019.0298
19. Ghebremedhin B, Ahmad-Nejad P. In-Vitro Efficacy of Cefiderocol in Carbapenem-Non-Susceptible Gram-Negative Bacilli of Different Genotypes in Sub-Region of North Rhine Westphalia, Germany. Pathogens. 2021;10:1258. doi:10.3390/pathogens10101258
20. Kohira N, West J, Ito A, Ito-Horiyama T, Nakamura R. In Vitro Antimicrobial Activity of a Siderophore Cephalosporin, S-649266, against Enterobacteriaceae Clinical Isolates, Including Carbapenem-Resistant Strains. Antimicrob Agents Chemother. 2015;60:729–734. doi:10.1128/AAC.01695-15
21. Bonnin RA, Emeraud C, Jousset AB, Naas T, Dortet L. Comparison of disk diffusion, MIC test strip and broth microdilution methods for cefiderocol susceptibility testing on carbapenem-resistant Enterobacterales. Clin Microbiol Infect. 2022;S1198–743. doi:10.1016/j.cmi.2022.04.013
22. Ito A, Nishikawa T, Matsumoto S, et al. Siderophore Cephalosporin Cefiderocol Utilizes Ferric Iron Transporter Systems for Antibacterial Activity against Pseudomonas aeruginosa. Antimicrob Agents Chemother. 2016;60:7396. doi:10.1128/AAC.01405-16
23. Ito A, Nishikawa T, Ota M, et al. Stability and low induction propensity of cefiderocol against chromosomal AmpC β-lactamases of Pseudomonas aeruginosa and Enterobacter cloacae. J Antimicrob Chemother. 2018;73:3049–3052. doi:10.1093/jac/dky317
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.