Back to Journals » International Journal of Women's Health » Volume 17
Altered Expression of CXCL16/CXCR6 and Its Correlation with Decidual Macrophage Polarization in Preeclampsia
Authors Huang S ![]() , Cao X, Xu Y
, Cao X, Xu Y ![]() , Zhu J
, Zhu J ![]() , Su M
, Su M
Received 17 September 2025
Accepted for publication 9 December 2025
Published 13 December 2025 Volume 2025:17 Pages 5411—5423
DOI https://doi.org/10.2147/IJWH.S567928
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Vinay Kumar
Sijia Huang,1,* Xueyan Cao,1,* Yuling Xu,1 Juan Zhu,1 Min Su2
1Department of Obstetrics and Gynecology, Affiliated Hospital of Nantong University, Medical School of Nantong University, Nantong, 226001, People’s Republic of China; 2Department of Obstetrics and Gynecology, Affiliated Hospital of Nantong University, Nantong, 226001, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Min Su, Email [email protected]
Objective: The CXCL16/CXCR6 signaling pathway is implicated in macrophage polarization. The correlation between CXCL16 and CXCR6 expressions in decidual tissues from patients with preeclampsia (PE) and macrophage polarization remains unexplored. This study aimed to investigate the correlation between the chemokine axis CXCL16/CXCR6 and macrophage polarization at the maternal-fetal interface in PE.
Methods: In this study, macrophage polarization status in the decidua was assessed using Western blotting (WB), immunohistochemistry (IHC), and immunofluorescence (IF) staining. For quantitative characterization, primary decidual macrophages were isolated and subjected to flow cytometry analysis to further evaluate macrophage polarization. Expression of CXCL16 and CXCR6 in decidual tissues was evaluated by WB and IHC. Additionally, the expressions of CXCL16 and CXCR6 in decidual macrophages were assessed by immunofluorescence double staining.
Results: IHC and WB demonstrated a significant upregulation of CD86 (P < 0.05), alongside a downregulation of CXCL16, CXCR6, and CD206 (P < 0.01) in PE decidual tissues. IF double staining revealed an increased abundance of CD68⁺CD80⁺/CD86⁺ cells in the decidua of patients with PE, while CD68⁺CD163⁺/CD206⁺ and CD68⁺CXCL16⁺/CXCR6⁺ cells was decreased. Flow cytometry analysis of primary decidual macrophages isolated from PE patients confirmed an increase in the proportions of CD68⁺CD80⁺ (P < 0.01) and CD68⁺CD86⁺ (P < 0.0001), and a marked decrease in CD68⁺CD163⁺ (P< 0.001) and CD68⁺CD206⁺ (P < 0.0001). These results indicate dysregulated decidual macrophage polarization in PE, characterized by an M1 bias that may be mediated by impaired CXCL16/CXCR6 signaling at the maternal-fetal interface.
Keywords: CXCL16, CXCR6, preeclampsia, decidua, macrophage polarization
Introduction
Preeclampsia (PE) is a pregnancy-specific syndrome marked by progressive clinical deterioration. It is clinically defined by the new onset of hypertension after 20 weeks of gestation in a previously normotensive woman, accompanied by proteinuria, maternal organ dysfunction, and/or placental insufficiency. Globally, PE accounts for more than 70,000 maternal deaths and 500,000 fetal/neonatal deaths annually, with the highest incidence of maternal mortality observed in developing countries.1 Recent studies have demonstrated that functional impairments in local placental immune cells, including macrophages, uterine natural killer cells, and regulatory T cells, and the resulting disruption of immune tolerance, provide a critical pathological basis for PE development.2
Macrophages are widely distributed throughout body tissues and differentiate from monocytes that migrate from the bloodstream. These cells can be polarized into the M1 phenotype by the lipopolysaccharide alone or in combination with Th1 cytokines, including interferon-gamma. M1 macrophages secrete pro-inflammatory cytokines and are involved in early pathogen clearance and inflammatory tissue damage.3 Th2 cytokines, including interleukin-4 (IL-4) and IL-13, can induce macrophage polarization toward the M2 phenotype. M2 macrophages are essential in mitigating inflammation, promoting angiogenesis, remodeling the extracellular matrix, and restoring tissue homeostasis.4 Decidual macrophages, the second largest immune cell population at the maternal-fetal interface and accounting for 20–30% of all leukocytes, persist during gestation and dynamically engage in several biological processes.5,6 Macrophage polarization refers to the acquisition of specific phenotypes and functions in response to microenvironmental stimuli and signals in their respective tissue.7,8 Macrophages (CD68+) can polarize into two distinct phenotypes: classically activated (M1) or pro-inflammatory macrophages (CD86+) and alternatively activated (M2) or anti-inflammatory macrophages (CD206+) based on two antagonistic pathways of arginine metabolism.7,9 Therefore, the phenotype of decidual macrophages undergoes alterations in response to changes within the maternal-fetal interface microenvironment. Decidual macrophages protect the semi-allogeneic fetus from maternal immune rejection and critically mediate implantation, trophoblast invasion, spiral artery remodeling, angiogenesis, and pathogen clearance. Dysregulation of decidual macrophages results in pregnancy complications including PE, recurrent pregnancy loss, and preterm birth.10
Chemokines are a superfamily of chemotactic cytokines primarily characterized by their ability to guide cell migration along concentration gradients.11 Chemokine ligand 16 (CXCL16), a constituent of the chemokine superfamily, exclusively binds CXC motif receptor 6 (CXCR6). CXCL16 exists dually as a transmembrane protein that acts as a scavenger receptor for oxidized low-density lipoprotein (oxLDL) and an adhesion molecule for CXCR6⁺cells, and a soluble chemokine that induces chemotaxis in CXCR6-expressing cells.12 CXCL16 is highly expressed at the maternal-fetal interface and is essential in sustaining normal pregnancy. Previous studies have demonstrated that CXCL16 secreted by trophoblasts inhibits granzyme B secretion from decidual γδ T cells while upregulating the expression of the anti-apoptotic marker Bcl-xL in trophoblasts, thereby maintaining normal pregnancy.13 Wang et al demonstrated that trophoblast-secreted CXCL16 binds the macrophage surface receptor CXCR6, driving monocyte differentiation into M2 macrophages, which suppress natural killer (NK) cell activity by inhibiting cytotoxicity, thereby establishing and maintaining normal pregnancy.14 Arutyunyan et al revealed that first-trimester dM1 and dM2 macrophages co-express CXCL16, with concomitant upregulation of its receptor CXCR6 in extravillous trophoblasts (EVTs), where these cell types are proximally localized.15 Their study further demonstrates CXCL16-mediated upregulation of placenta-specific genes, endothelial integrity regulators, cytokeratins, actin-binding molecules, and lectin family members, thereby facilitating trophoblast motility and invasion.15 Notably, elevated CXCL16 levels are observed in the maternal circulation of pregnancies affected by PE and gestational diabetes mellitus (GDM),16 while other studies have demonstrated lower expression levels of CXCL16 in the placental tissue of preeclamptic patients compared to normal pregnant women.17 Nevertheless, the role of CXCL16 and CXCR6 expressions in decidual macrophages, particularly their polarization dynamics in PE pathogenesis remains unexplored.
This study employed immunohistochemistry (IHC), immunofluorescence (IF), and primary decidual macrophage isolation to investigate macrophage polarization status in PE decidua. We analyzed CXCL16/CXCR6 expression and macrophage co-localization in decidua of preeclamptic patients and normal pregnant women, elucidating CXCL16-CXCR6 axis dysregulation in decidual macrophages and its pathogenic relevance to PE, thereby providing novel mechanistic insights into PE pathogenesis.
Materials and Methods
Patients and Sample Collection
This study enrolled 10 patients with PE and 14 women with normal pregnancy who underwent elective cesarean delivery at the Department of Obstetrics and Gynecology, Affiliated Hospital of Nantong University between November 2024 and May 2025.
PE is defined as systolic/diastolic blood pressure ≥ 140/90 mmHg after 20 weeks of gestation with random urine protein 2+ or 24-h urinary protein ≥ 300 mg. Normal pregnant women without complications who underwent cesarean section in the third trimester due to uterine scarring or other indications. The exclusion criteria included patients with chronic hypertension, diabetes mellitus, autoimmune disorders, uterine rupture, renal diseases, multifetal gestation, conception via assisted reproductive technology, maternal infections (including histologic or clinical chorioamnionitis), and those with history of smoking or alcohol abuse.
Decidual tissues from 14 healthy women who underwent elective cesarean delivery were collected as the control group, and the decidual tissues of 10 women who underwent cesarean delivery with PE were collected as the experimental group. After placental delivery, decidual samples were obtained by scraping the placental bed at the uterine fundus. Tissues were partially transported in pre-chilled phosphate-buffered saline (PBS) for immediate primary macrophage isolation, partially fixed in 10% neutral buffered formalin for IHC and IF, partially washed with physiological saline and stored at –80 °C or in liquid nitrogen for subsequent protein/RNA extraction.
Decidual Macrophage Isolation
Decidual tissues obtained post-cesarean were promptly transferred to the laboratory. The tissues were minced with ophthalmic scissors under sterile conditions and repeatedly washed with ice-cold PBS to remove maternal blood. The tissue fragments were suspended in RPMI-1640 medium (MeilunBio, Dalian, China) and subsequently digested for 60 min at 37 °C in a shaking incubator with 0.5 mg/mL collagenase IV (Solaibio, Beijing, China) and 150 U/mL DNase I (Beyotime, Shanghai, China). The suspension was sequentially filtered through 150 and 300-μm mesh filters to eliminate undigested debris. Filtered cells were pelleted by centrifugation, treated with red blood cell lysis buffer, and recentrifuged. The pellet was resuspended in RPMI-1640 and layered onto a Percoll density gradient (Cytiva, Marlborough, MA). Decidual immune cells were collected from the 40–60% density interface.
FCM Analysis
Isolated immune cells were stained with fluorochrome-conjugated monoclonal antibodies under light-protected conditions following the manufacturer’s protocols. Cells were subsequently washed thrice with ice-cold PBS. The following monoclonal antibodies were obtained from eBioscience (San Diego, CA, USA). The cells were incubated with PE-conjugated anti-CD80 (1:50), PE-conjugated anti-CD86 (1:50), PE-Cyanine7-conjugated anti-CD163 (1:50), PE-Cyanine7-conjugated anti-CD206 (1:50) in the dark for 30 minutes, fixed and permeabilized with BD Fixation and Permabilization for 20 minutes, and washed with 5 mL of BD Prem Wash. The cells were incubated with FITC-conjugated anti-CD68 (1:50) in the dark for 30 minutes, after which the CD68+CD80+, CD68+CD86+, CD68+CD163+, CD68+CD206+ cells were detected via flow cytometry. Flow cytometry was performed using a FACScalibur system (BD Biosciences, San Jose, CA, USA), and data analysis was performed using FlowJo software (TreeStar, San Carlos, CA, USA).
Western Blot (WB) Analysis
Total protein was extracted from decidual tissues using RIPA buffer supplemented with protease inhibitors (Beyotime, Shanghai, China). The protein concentration was measured using a bicinchoninic acid assay kit (Beyotime, Shanghai, China). We isolated 30 µg of protein using SDS-PAGE and subsequently transferred it to polyvinylidene fluoride membranes. The proteins were separated by electrophoresis and transferred onto a membrane according to their molecular weights. Membranes were blocked for 15 min at room temperature with rapid blocking buffer (NCM Biotech, Suzhou, China), and subsequently incubated with the following primary antibodies: mouse anti-human GAPDH (1:100000, Proteintech, Wuhan, China), rabbit anti-human CD86 (1:1000, Proteintech, Wuhan, China), rabbit anti-human CD206 (1:1000, Proteintech, Wuhan, China), rabbit anti-human CXCL16 (1:1000, Thermo Fisher Scientific, Shanghai, China), and rabbit anti-human CXCR6 (1:1000, Proteintech, Wuhan, China). After overnight incubation at 4 °C with gentle agitation, retrieve the primary antibody solution, the membrane was then washed three times with TBST, for 10 minutes each.The membranes were incubated with species-specific secondary antibodies (1:30,000, Cell Signaling Technology, Danvers, MA, USA) for 2 h at room temperature. The secondary antibody solution was retrieved, and the membrane was washed again with TBST. Protein bands were visualized utilizing the Odyssey imaging system (LI-COR, Lincoln, NE, USA), and quantitative analysis was performed using ImageJ software (NIH, Bethesda, MD, USA).
Reverse Transcription-Polymerase Chain Reaction (RT-PCR) Analysis
Total RNA was isolated from decidual tissues utilizing TRIzol reagent (Vazyme, Nanjing, China) according to the manufacturer’s instructions. For reverse transcription, 1 μg of total RNA was reverse-transcribed in a 20 μL reaction volume utilizing HiScript Q RT SuperMix for quantitative polymerase chain reaction (PCR; Vazyme), which contains a gDNA wiper to eliminate genomic DNA contamination. Additionally, quantitative real-time PCR was performed with 10 μL cDNA template using ChamQ SYBR qPCR Master Mix (Vazyme) on a real-time PCR system. The expression of the target gene was calculated using the 2-ΔΔCt method. All primers were developed by Sangon Biotech (Shanghai, China). Table 1 presents the primer sequences utilized for amplification.
 |
Table 1 The Primers Used for PCR Analysis |
Immunohistochemistry (IHC)
Fresh decidual tissues were fixed in formaldehyde, dehydrated, embedded in paraffin, and sectioned at 4 μm thickness. All sections were simultaneously subjected to deparaffinization by immersing them in fresh xylene twice for 30 minutes each. Subsequently, they were sequentially hydrated through a graded ethanol series: absolute ethanol for 10 minutes, 95% ethanol for 10 minutes, 80% ethanol for 10 minutes, and 75% ethanol for 5 minutes. The sections were then washed three times for 5 minutes each in PBS on a shaking platform. After drying gently, the sections were placed in a humidified chamber. Freshly prepared 3% H2O2 solution was applied to the tissue areas and incubated at room temperature for 30 minutes in the dark to quench endogenous peroxidase activity. Following this, the sections were washed three times with PBS for 5 minutes each. Microwave-assisted antigen retrieval was performed in a 10 mmol/L citrate buffer (pH 6.0) for 3–4 min. The sections were then allowed to cool down to room temperature naturally. After cooling, they were washed three times with PBS for 5 minutes each. Finally, the tissue sections were circled with a hydrophobic pen, followed by the application of blocking serum and incubation at 37°C for 30 minutes. Sections were incubated with following primary antibodies: CD68 antibody,1:200, Proteintech, Wuhan, China), CD86 antibody (1:200, Proteintech, Wuhan, China), CD206 antibody (1:2000, Proteintech, Wuhan, China) at 4 °C overnight. Biotinylated goat anti-mouse or goat anti-rabbit secondary antibodies were administered for 30 min at room temperature the following day. Immunostaining was performed using chromogenic substrates, and the findings were examined using light microscopy. Whole-slide images were captured at 200× magnification and analyzed using Image J software (NIH, Bethesda, MD, USA).
Immunofluorescence (IF)
Paraffin-embedded sections were deparaffinized using xylene and rehydrated through an ethanol gradient, followed by PBS washes. Microwave-assisted antigen retrieval was performed in a 10 mmol/L citrate buffer (pH 6.0) for 3–4 min. Endogenous peroxidase activity was inhibited using 3% H2O2, and nonspecific binding was obstructed by incubating tissues with 5% bovine serum albumin for 1 h at room temperature. The initial procedures were performed as described previously for the IHC protocol. Primary antibody cocktails were administered to sections and incubated overnight at 4 °C in a humidified chamber. On the following day, the sections were washed three times with PBS for 5 minutes each. After blotting dry around the tissues, the sections were incubated with species-appropriate secondary antibodies (corresponding to the host of the primary antibodies) for 2 hours at room temperature in the dark. Following incubation, the sections were washed three times with PBS for 5 minutes each. After blotting dry again, the nuclei were counterstained by applying DAPI solution within the circled area and incubating for 10 minutes in the dark. Finally, the sections were washed with PBS as described in the previous step, blotted dry, and mounted with an anti-fade mounting medium. Imaging was performed utilizing an inverted fluorescence microscope, with co-localization analysis conducted in Image J software (NIH, Bethesda, MD, USA). The primary antibodies utilized in this experiment included: CD68 antibody (1:200, Proteintech, Wuhan, China), CD80 antibody (1:200, Abcam, UK), CD86 antibody (1:200, Proteintech, Wuhan, China), CD163 antibody (1:200, Proteintech, Wuhan, China), CD206 antibody (1:400, Proteintech, Wuhan, China).
Statistical Analysis
All experiments were conducted at least three times, and the data were presented as the mean ± standard deviation. The data conforming to normal distribution and homogeneity of variance between the two groups were analyzed using Student’s t-test. P < 0.05 was considered statistically significant. The statistical software GraphPad Prism software (version 9) was used for the statistical analysis.
Results
Abnormal Macrophage Polarization in Patients with PE
To examine the polarization of decidual macrophages in PE, we initially performed IHC and WB analyses. IHC results revealed that CD68, CD86, and CD206 were expressed in the membranous and cytoplasmic compartments of decidua (Figure 1A). Compared to those in normal pregnant women, the expressions of CD68 and CD86 were higher in the decidual tissues of patients with PE, while CD206 expression was lower (Figure 1B). WB analyses revealed that the protein levels of CD86 were increased and those of CD206 were decreased in the PE group compared with the normal pregnancy group (Figure 1C and D). Simultaneously, the mRNA levels of the inflammatory cytokines TNF-α and IL-6 were higher in the decidual tissues of the PE group than those in the normal pregnancy group (Figure 1E).
 |
Figure 1 Expression of CD68, CD86 and CD206 in decidual tissues of normal pregnancy women and patients with PE. (A) The expression levels of CD68, CD86 and CD206 in decidual tissues were assessed by immunohistochemistry (×200). (B) Semiquantitative analysis of (A). (C) The relative protein levels of CD86 and CD206 were detected via Western blotting. GAPDH was used as an endogenous control. (D) Semiquantitative analysis of (C). (E) The relative mRNA levels of TNF-α and IL-6 were detected via q-PCR. Student’s t-test was used for the statistical analysis, and *p<0.05; **p < 0.01. |
Subsequently, we examined the decidual tissues using IF co-localization staining. We utilized CD68 as a pan-macrophage marker, CD80 and CD86 as M1 macrophage markers, CD163 and CD206 as M2 macrophage markers. Fluorescence microscopy analysis revealed that most CD68+ decidual macrophages in women with PE brightly stained CD80 and CD86 (Figure 2A and B), whereas most CD68+ decidual macrophages in women with normal pregnancy stained positive for CD163 and CD206 (Figure 2C and D).
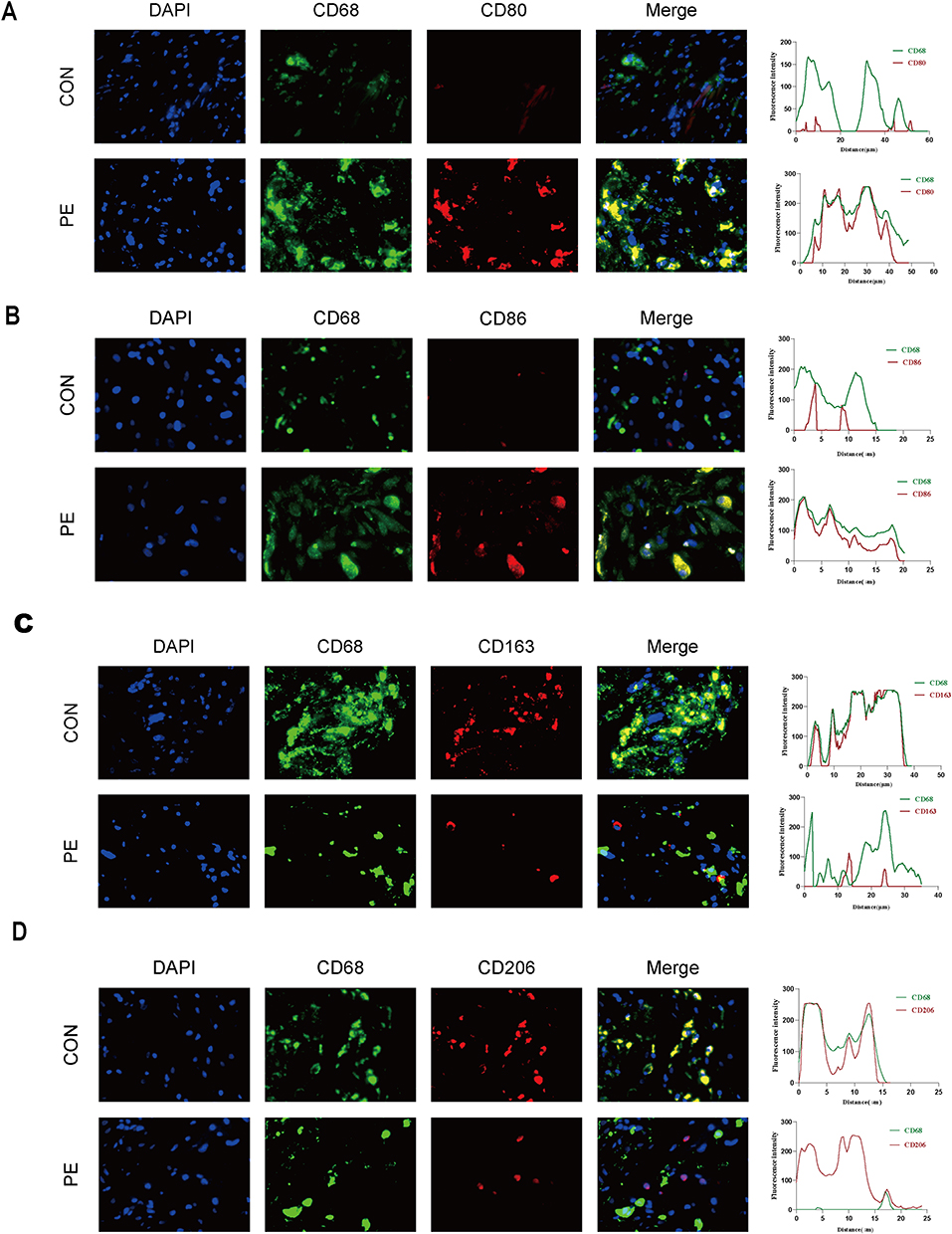 |
Figure 2 Immunofluorescence staining for M1 and M2 in normal pregnancy and patients with PE. CD68 is the maker for pan-macrophages. CD80 and CD86 are markers for M1, CD163 and CD206 are markers for M2 macrophages. Scale bars: 100 µm. (A and B) The CD68+CD80+and CD68+CD86+ cells in decidual tissues were detected by immunofluorescence double staining. (C and D) The CD68+CD163+ and CD68+CD206+ cells in decidual tissues were detected by immunofluorescence double staining. |
We accurately isolated and extracted primary macrophage from decidual tissues, followed by phenotypic characterization and analysis using flow cytometry. We classified CD68+CD80+ and CD68+CD86+ cells as M1 macrophages and CD68+CD163+ and CD68+CD206+ cells as M2 macrophages. Flow cytometry results revealed that compared with normal pregnancy, the proportion of CD68+CD80+ and CD68+CD86+ M1 macrophages was increased in patients with PE, while the proportion of CD68+CD206+ and CD68+CD163+ M2 macrophages was decreased (Figure 3A and B). Collectively, these results revealed abnormal polarization of decidual macrophages in PE, with a skew toward the M1 phenotype.
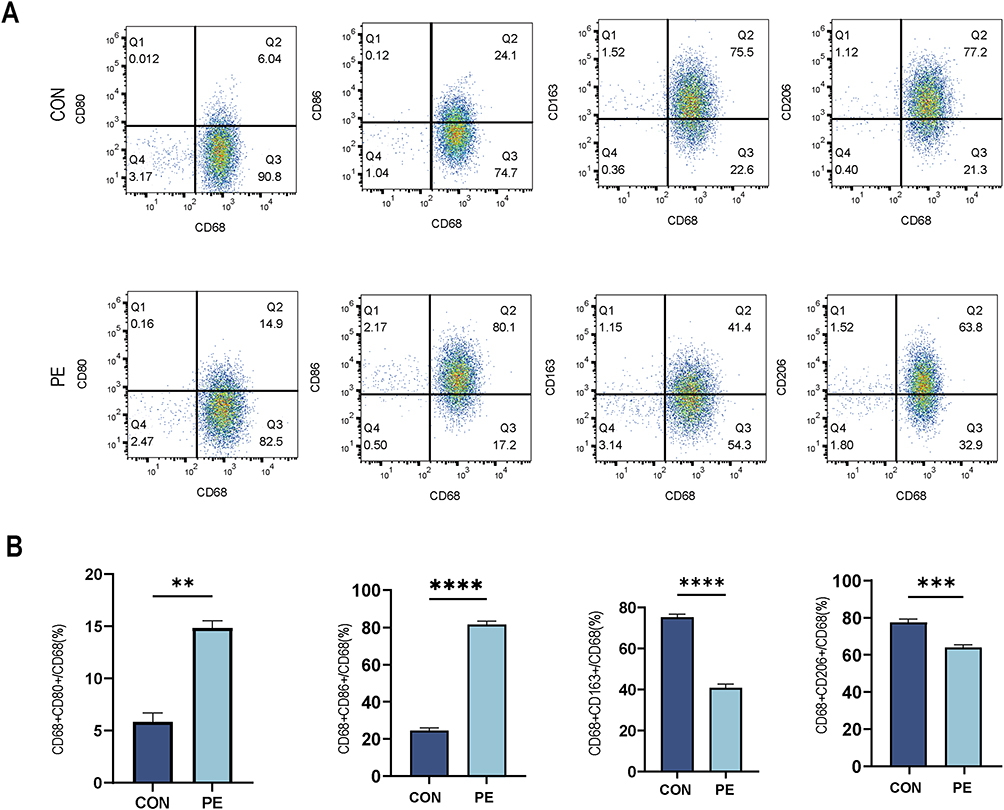 |
Figure 3 Decidual macrophages polarization of normal pregnant women and patients with PE. (A) Flow cytometric analysis to determine the proportion of M1 (CD68+CD80+ or CD68+CD86+) and M2 (CD68+CD163+ or CD68+CD206+) in normal pregnancy and patients with PE. (B) Semiquantitative analysis of (A). Student’s t-test was used for the statistical analysis, and **p<0.01; ***p < 0.001; ****p < 0.0001. |
Expressions of CXCL16 and CXCR6 Decreased in Decidual Tissues of PE Patients
We utilized IHC and WB to detect CXCL16 and CXCR6 expressions in decidual tissues. The results indicated that CXCL16 and CXCR6 were predominantly localized to the cell membrane (Figure 4A). Compared to pregnant women, patients with PE exhibited lower expressions of CXCL16 and CXCR6 in decidual tissues (Figure 4B). WB analysis revealed significantly decreased CXCL16 and CXCR6 expression levels in the decidual tissues of patients with PE (Figure 4C and D). Furthermore, qPCR results confirmed reduced mRNA expressions of CXCL16 and CXCR6 in decidual tissues in PE (Figure 4E).
 |
Figure 4 Expression of CXCL16 and CXCR6 in decidual tissues and decidual macrophages of normal pregnant women and patients with PE. (A) The expression levels of CXCL16 and CXCR6 in decidual tissues were assessed by immunohistochemistry (×200). (B) Semiquantitative analysis of (A). (C) The relative protein levels of CXCL16 and CXCR6 in decidual tissues were detected via Western blotting. (D) Semiquantitative analysis of (C). (E) The relative mRNA levels of CXCL16 and CXCR6 were detected via q-PCR. (F) The CD68+CXCL16+ cells in decidual tissues were detected by immunofluorescence double staining. (G) The CD68+CXCR6+ cells in the decidual tissues were detected by immunofluorescence double staining. Scale bars: 100 µm. Student’s t-test was used for the statistical analysis, and *p < 0.05; **p < 0.01. |
The proportions of CD68+CXCL16+ and CD68+CXCR6+ double-positive cells were assessed via IF double-staining to evaluate CXCL16 and CXCR6 expressions in decidual macrophages. In decidual tissues from patients with PE, the proportions of CD68+CXCL16+ cells and CD68+CXCR6+ cells were lower than those in normal pregnant women (Figure 4F and G). These findings indicated that CXCL16 and CXCR6 were expressed in decidual macrophages, and their expressions were reduced in both decidual tissues and macrophages from patients with PE.
Discussion
Preeclampsia is a highly complex immunological process involving diverse pathophysiological mechanisms, etiologies, and risk factors. Although its exact etiology remains unclear, endothelial dysfunction, abnormal angiogenesis, insufficient trophoblast invasion, and impaired remodeling of maternal spiral arteries are considered primary contributing factors.18 PE has been initially classified into two distinct subgroups: early-onset (< 34 weeks of gestation) and late-onset (≥ 34 weeks of gestation) disease, stratified according to gestational age at diagnosis or delivery based on histological, clinical, biological, hemodynamic, and epidemiological criteria.2 Impaired early placental development and subsequent fetal growth restriction are commonly associated with early-onset PE, whereas late-onset PE (LOPE) is predominantly related to maternal endothelial dysfunction.18 Macrophages are remarkably adaptable and heterogeneous immune cells whose phenotype and functional polarization—dictated by the local microenvironment—enable multifunctional capacities, including (1) phagocytosis of pathogens, infected cells, debris, and apoptotic cells; (2) antigen presentation through processed peptides on MHC molecules; (3) secretion of cytokines (IL-1β, IL-6, and TNF-α).7 Decidual macrophages represent the primary antigen-presenting cells within the decidua.2 Their high-density perivascular accumulation surrounding invading trophoblasts and spiral arteries highlights their crucial role in modulating the maternal-fetal interface.19 Although the etiology of PE remains incompletely understood, the polarization status of decidual macrophages—toward pro-inflammatory (M1) or anti-inflammatory (M2) phenotypes—is essential in sustaining placental vascular remodeling and maternal-fetal immunological tolerance. An imbalance in macrophage polarization induces a pro-inflammatory shift at the maternal-fetal interface, suppressing angiogenesis and resulting in vascular endothelial injury, inadequate trophoblast invasion, and compromised remodeling of uterine spiral arteries. These pathological cascades collectively facilitate the onset and progression of PE.20
Decidual macrophages exhibit stage-specific phenotypic and functional adaptations during gestation. The M1 phenotype dominates prior to embryo implantation; however, extravillous trophoblast (EVT) invasion triggers a transition in decidual macrophages toward a persistent mixed M1-M2 repertoire essential for placental development and spiral artery remodeling, culminating in M2 macrophage predominance that sustains maternal-fetal tolerance throughout gestation.21 Macrophages are distinguished by their capacity to remodel many tissues and secrete several growth factors and cytokines. Consequently, decidual macrophages likely exert critical functions in regulating embryonic implantation, placental development, trophoblast behavior, and decidual homeostasis19 encompassing the establishment of permissive local immune balance, facilitating EVT invasion, and coordinating remodeling of uterine smooth muscle, glands, and spiral arteries.22,23 Macrophage functionality is closely associated with their appropriate polarization status, which is essential for maintaining a physiological immune microenvironment at the maternal-fetal interface.24 Under physiological conditions, M1 and M2 macrophages coexist in a balanced state9 with a predominant M2 phenotype in decidual macrophages. Disruption of this homeostasis induces immunological dysregulation, resulting in obstetric complications, including PE.
Herein, we identified and analyzed pan-macrophage, M1-, and M2-polarization markers in preeclamptic decidual tissues using IHC and WB, supplemented by IF co-localization staining to evaluate macrophage distribution within the decidual microenvironment. We performed IHC and WB to analyze pan-macrophage, M1-, and M2-polarization markers in preeclamptic decidual tissues, supplemented by IF co-localization staining to assess spatial macrophage distribution. Furthermore, we isolated primary decidual macrophages for flow cytometric analysis of M1 and M2 markers. Our findings revealed that patients with PE exhibit upregulated expression of the pan-macrophage marker CD68, alongside elevated levels of M1 markers (CD80 and CD86) and decreased levels of M2 markers (CD163 and CD206) in decidual macrophages. These observations align with established literature. These findings indicate a phenotypic shift from M2 to M1 polarization in decidual macrophages during PE, indicating that dysregulated macrophage polarization contributes to disease pathogenesis.
Moving beyond the established paradigm of macrophage imbalance, our study sought to position this shift within a specific molecular context. The predominant pathophysiological model of PE centers on an imbalance of placental angiogenic factors, most notably the elevated sFlt-1/PlGF ratio, which serves as a robust clinical predictor of systemic endothelial dysfunction.25,26 Our findings, which highlight the downregulation of the CXCL16/CXCR6 axis, illuminate a distinct and potentially complementary pathological pathway: the dysregulation of local immune communication at the maternal-fetal interface.
Chemokines are a superfamily of low-molecular-weight chemotactic proteins that facilitate leukocyte migration by binding to G protein-coupled receptors expressed on target cells.27 The secretion of appropriate chemokine signals by decidual cells promotes the recruitment of predominantly anti-inflammatory leukocyte subsets, which is essential for sustaining pregnancy.12 Decidual cells dynamically remodel their chemokine secretion profiles to facilitate various stages of embryo implantation, while chemokine expression regulates immune polarization responses.27,28 CXCL16, as a member of the chemokine family, is expressed and secreted by various cell types, including macrophages, dendritic cells, and cancer cells.16 CXCL16 exhibits context-dependent roles across pathologies: it facilitates atherosclerosis through T-cell activation and inflammatory cascade induction while contributing to cancer cell proliferation and migration.29 Lee et al revealed that adipose tissue macrophages polarize toward M1 or M2 phenotypes through the modulation of the CXCL16/CXCR6 axis, exhibiting immunosuppressive effects by regulating pro- or anti-inflammatory pathways.30 Hong et al demonstrated that macrophages facilitate ovarian cancer cell migration and invasion through the regulation of the CXCL16/CXCR6 signaling pathway.31 Chemokines are also essential in gestational maintenance. Dysregulated chemokine expression is associated with pathological conditions such as recurrent pregnancy loss (RPL) and preeclampsia.32 CXCL16/CXCR6 are highly expressed at the maternal-fetal interface. Current evidence indicates that trophoblast-derived CXCL16 binds to CXCR6 receptors on decidual immune cells, thereby facilitating immunomodulatory effects essential for maintaining fetomaternal immune tolerance.11,30 Huang et al demonstrated that during the first trimester, CXCL16 localizes to cytoplasmic and membranous regions of cytotrophoblasts, syncytiotrophoblasts, and EVT, whereas CXCR6 is predominantly expressed in decidual immune cells.12,33 Sun et al reported significantly reduced CXCL16/CXCR6 expression in chorionic-decidual tissues from patients experiencing recurrent miscarriage compared to normal pregnancy tissues.12 Impaired remodeling of uterine spiral arteries is recognized as a principal pathomechanism in PE, while studies have demonstrated that CXCL16 facilitates HUVEC proliferation, migration, and tube formation in in vitro angiogenesis models, proposing CXCL16 as a novel pro-angiogenic factor.11,34 However, research on the expression profile of CXCL16/CXCR6 in the maternal-fetal interface (specifically within decidual tissue) and its association with PE remains limited. Herein, we examined and analyzed CXCL16 and CXCR6 expressions in decidual tissues from patients with PE using IHC and WB. Furthermore, we performed co-localization analysis via IF staining to assess CXCL16 and CXCR6 expressions within decidual macrophages. Through the methods mentioned above, we found that CXCL16 and CXCR6 expressions were lower in the decidual tissues of patients with PE compared to healthy pregnant women. Furthermore, IF co-localization analysis revealed reduced CD68+CXCL16+ and CD68+CXCR6+ expressions within the decidua of patients with PE compared to those of healthy pregnant women.
Our discovery of reduced CXCL16/CXCR6 expression in PE decidua adds a novel dimension to the chemokine dysregulation observed in pregnancy complications. While this exploratory study, limited by its sample size, cannot definitively assess the diagnostic specificity and sensitivity of CXCL16/CXCR6 as standalone biomarkers, its value lies in mechanistic insight. The correlation between CXCL16/CXCR6 downregulation and M1 macrophage polarization provides a plausible explanation for the local inflammatory milieu in PE. This mechanism is distinct from, but may interact with, the well-characterized angiogenic imbalance.
This study demonstrated that in the decidual tissues of patients with PE, the number of macrophages is increased, and these macrophages exhibit polarization toward the M1 phenotype. Furthermore, CXCL16 and CXCR6 expressions are decreased in the decidual tissues and decidual macrophages of patients with PE. However, PE is a disease with multiple etiologies and mechanisms. This study has some limitations, including a small sample size and verification of the results only at the protein level, based on cellular phenotype. The statistical power may be influenced by the small sample size, warranting larger-scale studies in the future. The specific mechanisms by which the CXCL16/CXCR6 signaling pathway regulates macrophage polarization and its correlation with PE warrant further investigation.
Conclusion
We initially assessed the expression of CD68, CD86, CD206, CXCL16, and CXCR6 in decidual tissues using IHC and WB. IHC analysis indicated that the expression of CD68 and CD86 was significantly increased in the decidua of patients with PE (P < 0.01 and P < 0.05, respectively). In contrast, the expression levels of CD206, CXCL16, and CXCR6 were significantly downregulated (P < 0.05, P < 0.01, and P < 0.05). These findings were consistent with the WB results, which showed an increased protein level of CD86 (P < 0.01) and decreased protein levels of CD206, CXCL16, and CXCR6 (P < 0.01, P < 0.05, and P < 0.05) in the PE group. IF double staining further revealed an increased abundance of CD68⁺CD80⁺/CD86⁺ cells, alongside a decreased presence of CD68⁺CD163⁺/CD206⁺ and CD68⁺CXCL16⁺/CXCR6⁺ cells in the PE decidua. To obtain precise quantification, flow cytometric analysis was performed on primary decidual macrophages isolated from PE patients. The results confirmed a significant increase in the proportions of CD68⁺CD80⁺ (P < 0.01) and CD68⁺CD86⁺ (P < 0.0001) cells, and a marked decrease in CD68⁺CD163⁺ (P < 0.001) and CD68⁺CD206⁺ (P < 0.0001) cells.
Our findings indicate that an imbalance in decidual macrophage polarization is associated with the pathogenesis of preeclampsia. Additionally, we demonstrated a relationship between the CXCL16/CXCR6 signaling pathway and PE, indicating a potential association between CXCL16/CXCR6 and decidual macrophage polarization in PE. During PE, the number of macrophages at the maternal-fetal interface increases, and these cells polarize toward the M1 phenotype. Furthermore, the decreased expressions of CXCL16 and CXCR6 observed in the decidual tissues and decidual macrophages of patients with PE may contribute to altered macrophage polarization, thereby facilitating the onset and progression of PE. Despite the limitations of this study, it provides novel insights into PE pathogenesis.
Abbreviations
PE, Preeclampsia; CXCL16, C-X-C Motif Chemokine Ligand 16; CXCR6, C-X-C Motif receptor 6; IL-1β, Interleukin-1β; IL-6, Interleukin-6; TNF-α, Tumor necrosis factor-α; WB, Western Blot; RT-qPCR, Real-time quantitative polymerase chain reaction; IHC, Immunohistochemistry; IF, Immunofluorescence.
Ethics Approval and Consent to Participate
This study was conducted in accordance with the Declaration of Helsinki. The studies involving human participants were reviewed and approved by the ethical committee of Affiliated Hospital of Nantong University (Approval No. 2023-K210) on 24 December 2023. The patients/participants provided their written informed consent to participate in this study.
Acknowledgments
The author thanks the Ethics Committee of the Affiliated Hospital of Nantong University. We would like to thank all tutors for their guidance in scientific research and all participants for their support and encouragement in research data collection.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by Scientific research project of Nantong Health Commission (MS2023018), Scientific research project of Jiangsu Provincial Health Commission (H2023001), Nantong Science and Technology Bureau Plan Project (MSZ2024160), and Research Project of Nantong Maternal and Child Health Specialist Alliance (TFM202502).
Disclosure
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Li Y, Sang Y, Chang Y. et al. A Galectin-9–Driven CD11c high decidual macrophage subset suppresses uterine vascular remodeling in preeclampsia. Circulation. 2024;149(21):1670–1688. doi:10.1161/CIRCULATIONAHA.123.064391
2. Boulanger H, Bounan S, Mahdhi A, et al. Immunologic aspects of preeclampsia. AJOG Global Rep. 2024;4(1):100321–100338. doi:10.1016/j.xagr.2024.100321
3. Gao X, Lu C, Miao Y, Ren J, Cai X. Role of macrophage polarisation in skin wound healing. Int Wound J. 2023;20(7):2551–2562. doi:10.1111/iwj.14119
4. Kiseleva V, Vishnyakova P, Elchaninov A, Fatkhudinov T, Sukhikh G. Biochemical and molecular inducers and modulators of M2 macrophage polarization in clinical perspective. Int Immunopharmacol. 2023;122:110583–110592. doi:10.1016/j.intimp.2023.110583
5. Socha MW, Malinowski B, Puk O, et al. The role of NF-κB in uterine spiral arteries remodeling, insight into the cornerstone of preeclampsia. Int J Mol Sci. 2021;22(2):704–717. doi:10.3390/ijms22020704
6. Yin T, Li X, Li Y, Zang X, Liu L, Du M. Macrophage plasticity and function in cancer and pregnancy. Front Immunol. 2024;14:17. doi:10.3389/fimmu.2023.1333549
7. Shapouri‐Moghaddam A, Mohammadian S, Vazini H, et al. Macrophage plasticity, polarization, and function in health and disease. J Cell Physiol. 2018;233(9):6425–6440. doi:10.1002/jcp.26429
8. Yunna C, Mengru H, Lei W, Weidong C. Macrophage M1/M2 polarization. Eur J Pharmacol. 2020;877:173090–173098. doi:10.1016/j.ejphar.2020.173090
9. Luo M, Zhao F, Cheng H, Su M, Wang Y. Macrophage polarization: an important role in inflammatory diseases. Front Immunol. 2024;15:1352946.
10. Wang L-L, Li Z-H, Wang H, Kwak-Kim J, Liao A-H. Cutting edge: the regulatory mechanisms of macrophage polarization and function during pregnancy. J Reprod Immunol. 2022;151:103627–103636. doi:10.1016/j.jri.2022.103627
11. Shi JW, Yang HL, Fan DX, et al. The role of CXC chemokine ligand 16 in physiological and pathological pregnancies. Am J Reprod Immunol. 2020;83(4):e13223–13250. doi:10.1111/aji.13223
12. Mei J, Yan Y, Li S-Y, et al. CXCL16/CXCR6 interaction promotes endometrial decidualization via the PI3K/AKT pathway. Reproduction. 2019;157(3):273–282. doi:10.1530/REP-18-0417
13. Fan D-X, Zhou W-J, Jin L-P, Li M-Q, Xu X-H, Xu C-J. Trophoblast-derived CXCL16 decreased granzyme B production of decidual γδ T cells and promoted Bcl-xL expression of trophoblasts. Reprod Sci. 2019;26(4):532–542. doi:10.1177/1933719118777638
14. Wang X-Q, Zhou W-J, Hou -X-X, Fu Q, Li D-J. Trophoblast-derived CXCL16 induces M2 macrophage polarization that in turn inactivates NK cells at the maternal–fetal interface. Cell Mol Immunol. 2018;15(12):1038–1046. doi:10.1038/s41423-018-0019-x
15. Arutyunyan A, Roberts K, Troulé K, et al. Spatial multiomics map of trophoblast development in early pregnancy. Nature. 2023;616(7955):143–151. doi:10.1038/s41586-023-05869-0
16. Lekva T, Michelsen AE, Aukrust P, et al. CXC chemokine ligand 16 is increased in gestational diabetes mellitus and preeclampsia and associated with lipoproteins in gestational diabetes mellitus at 5 years follow-up. Diabetes Vasc Dis Res. 2017;14(6):525–533. doi:10.1177/1479164117728011
17. Dunk CE, van Dijk M, Choudhury R, et al. Functional evaluation of STOX1 (STORKHEAD-BOX PROTEIN 1) in placentation, preeclampsia, and preterm birth. Hypertension. 2021;77(2):475–490. doi:10.1161/HYPERTENSIONAHA.120.15619
18. Aneman I, Pienaar D, Suvakov S, Simic TP, Garovic VD, McClements L. Mechanisms of key innate immune cells in early- and late-onset preeclampsia. Front Immunol. 2020;11:19. doi:10.3389/fimmu.2020.01864
19. Ning F, Liu H, Lash GE. The role of decidual macrophages during normal and pathological pregnancy. Am J Reprod Immunol. 2016;75(3):298–309. doi:10.1111/aji.12477
20. Rodríguez-Morales P, Franklin RA. Macrophage phenotypes and functions: resolving inflammation and restoring homeostasis. Trends Immunol. 2023;44(12):986–998. doi:10.1016/j.it.2023.10.004
21. Sun F, Wang S, Du M. Functional regulation of decidual macrophages during pregnancy. J Reprod Immunol. 2021;143:103264. doi:10.1016/j.jri.2020.103264
22. Krop J, Tian X, van der Hoorn M-L, Eikmans M. The Mac is back: the role of macrophages in human healthy and complicated pregnancies. Int J Mol Sci. 2023;24(6):5300. doi:10.3390/ijms24065300
23. Du X, Liu H, Shi J, Yang P, Gu Y, Meng J. The PD-1 /PD-L1 signaling pathway regulates decidual macrophage polarization and may participate in preeclampsia. J Reprod Immunol. 2024;164:104258–104266. doi:10.1016/j.jri.2024.104258
24. Brown MB, von Chamier M, Allam AB, Reyes L. M1/M2 macrophage polarity in normal and complicated pregnancy. Front Immunol. 2014;5. doi:10.3389/fimmu.2014.00606
25. Stepan H, Galindo A, Hund M, et al. Clinical utility of sFlt‐1 and PlGF in screening, prediction, diagnosis and monitoring of pre‐eclampsia and fetal growth restriction. Ultrasound Obstet Gynecol. 2023;61(2):168–180. doi:10.1002/uog.26032
26. MacDonald TM, Walker SP, Hannan NJ, Tong S, Kaitu’u-Lino Tu J. Clinical tools and biomarkers to predict preeclampsia. eBioMedicine. 2022;75:103780. doi:10.1016/j.ebiom.2021.103780
27. He Y-Y, He X-J, Guo P-F, et al. The decidual stromal cells-secreted CCL2 induces and maintains decidual leukocytes into Th2 bias in human early pregnancy. Clin Immunol. 2012;145(2):161–173. doi:10.1016/j.clim.2012.07.017
28. Guzeloglu-Kayisli O, Kayisli U, Taylor H. The role of growth factors and cytokines during implantation: endocrine and paracrine interactions. Semin Reprod Med. 2009;27(01):062–079. doi:10.1055/s-0028-1108011
29. Deng L, Chen N, Li Y, Zheng H, Lei Q. CXCR6/CXCL16 functions as a regulator in metastasis and progression of cancer. Biochim Biophys Acta Rev Cancer. 2010;1806(1):42–49. doi:10.1016/j.bbcan.2010.01.004
30. Lee S-C, Lee Y-J, Choi I, Kim M, Sung J-S. CXCL16/CXCR6 axis in adipocytes differentiated from human adipose derived mesenchymal stem cells regulates macrophage polarization. Cells. 2021;10(12):3410. doi:10.3390/cells10123410
31. Hong L, Wang S, Li W, Wu D, Chen W. Tumor-associated macrophages promote the metastasis of ovarian carcinoma cells by enhancing CXCL16/CXCR6 expression. Pathol Res Pract. 2018;214(9):1345–1351. doi:10.1016/j.prp.2018.07.009
32. Nancy P, Tagliani E, Tay C-S, Asp P, Levy DE, Erlebacher A. Chemokine gene silencing in decidual stromal cells limits T cell access to the maternal-fetal interface. Science. 2012;336(6086):1317–1321. doi:10.1126/science.1220030
33. Huang Y, Zhu X-Y, Du M-R, Li D-J. Human trophoblasts recruited T lymphocytes and monocytes into decidua by secretion of chemokine CXCL16 and interaction with CXCR6 in the first-trimester pregnancy. J Immunol. 2008;180(4):2367–2375. doi:10.4049/jimmunol.180.4.2367
34. Yu X, Zhao R, Lin S, et al. CXCL16 induces angiogenesis in autocrine signaling pathway involving hypoxia-inducible factor 1α in human umbilical vein endothelial cells. Oncol Rep. 2016;35(3):1557–1565. doi:10.3892/or.2015.4520
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
CXCL16 Promotes the Development of Chronic Atrophic Gastritis by Regulating M1 Macrophage Polarisation
Ma X, Liu N, Cui W, Mai P
Journal of Inflammation Research 2026, 19:551832
Published Date: 19 January 2026