Back to Journals » OncoTargets and Therapy » Volume 13
Aloperine Exerts Antitumor Effects on Bladder Cancer in vitro
Authors Zhang L, Liang J, Liu X, Wu J, Tan D, Hu W
Received 28 April 2020
Accepted for publication 7 September 2020
Published 13 October 2020 Volume 2020:13 Pages 10351—10360
DOI https://doi.org/10.2147/OTT.S260215
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Nicola Silvestris
Lijun Zhang,1,* Jun Liang,1,* Xiaohua Liu,1 Jianhua Wu,1 Daqing Tan,1 Wei Hu2
1Department of Urology, Minda Hospital Affiliated to Hubei Minzu University, Enshi, Hubei Province, People’s Republic of China; 2Department of Urology, The First Affiliated Hospital of University of South of China, Hengyang, Hunan Province, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Lijun Zhang Tel +8615171021486
Email [email protected]
Background: Human bladder cancer is the most common malignant tumor of the urinary system and one of the 10 most common tumors of the whole body. Although most patients with bladder cancer exhibit a good prognosis with standard treatment, effective therapies for patients with a recurrent or advanced bladder cancer are unavailable. Therefore, highly effective drugs to treat such patients need to be developed. Aloperine (ALO), a natural compound isolated from Sophora alopecuroides, has antitumor properties. However, the role of ALO in human bladder cancer remains unclear.
Methods: In the present study, MTT was used to detect the cytotoxic effect of ALO on human BC cell line EJ and human urothelium cell line SV-HUC-1cells. Meanwhile, in order to investigate the effects of ALO on the proliferation, apoptosis, migration, and invasion of BC EJ cells and its mechanism by Cell Counting Kit-8 (CCK-8) assay, immunofluorescence, Hoechst 33342 staining, wound scratch assay, transwell migration and invasion assay, Western blot analysis.
Results: ALO can inhibit the proliferation and invasion of human bladder cancer EJ cells, and is low-toxic to human urothelium cells. Moreover, it can promote cellular apoptosis in vitro. Further analysis demonstrated the involvement of Caspase-dependent apoptosis following ALO treatment. ALO also downregulated the protein expression levels of Ras, p-Raf1 and p-Erk1/2.
Conclusion: ALO is a potential drug for human bladder cancer therapy.
Keywords: Aloperine, bladder cancer, apoptosis
Introduction
Bladder cancer (BC) is a malignant tumor on the mucous membrane of the bladder. It is the most common malignant tumor of the urinary system and one of the 10 most common tumors of the whole body.1 However, the pathogenesis and etiology of BC remain unclear. The incidence of urogenital tumors is the first in China and second only to prostate cancer in the west. The incidence of BCs is high in tumor registration areas, with 386,000 new cases worldwide per year and ranking ninth in the incidence of malignant tumors.2,3 BC can occur at any age, even in children. The incidence of the disease increases with age, with a high incidence among 50–70 year olds. BCs is three to four times more common in men than in women.4 In recent years, advances have been made in the prevention and treatment of BCs. However, the underlying mechanism and effective biomarkers for the pathological development of BCs require further investigations. During tumor progression, BC cells can proliferate and metastasize through several steps, including uncontrolled cell cycle regulation, signaling pathways, and apoptosis. Therefore, addressing the molecular events associated with the uncontrolled proliferation, migration, and invasion of BC cells is a key target for the development of new therapeutic drugs.
In general, the most effective treatment for cancer is chemotherapy drugs, including doxorubicin, mitomycin, and hydroxycamptothecin. However, most patients with BCs die because of drug resistance, side effects, and acidic environment.5,6 Therefore, finding plant-derived compounds with high antitumor activities and low toxic side effects have become the focus of research. In recent years, the application of compounds extracted from natural plants in the development of anti-tumor drugs has attracted wide attention because of their potential effectiveness and safety.7–9 Therefore, we investigated the efficacy of the phytochemical Aloperine (ALO), an alkaloid obtained from Sophora alopecuroides. S. alopecuroides is a traditional Chinese medicine with antipyretic, anti-inflammatory, and analgesic effects.10–13 ALO is a quinolizine alkaloid that has been isolated from S. alopecuroides extract (Figure 1A). It exhibits various pharmacological activities, including antibacterial, anti-allergic, antiviral, anti-renal injury, anti-nerve injury, anti-inflammatory, and anti-tumor effects.14–18 The antitumor effects of ALO have been demonstrated in several human cancer cells, including lung, colon, and multiple myeloma.18–21 However, few studies explored its anti-tumor effect on BCs and its related mechanism.
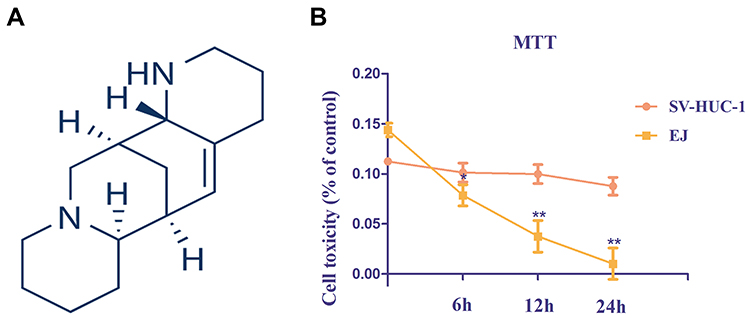 |
Figure 1 Structure of Aloperine and cytotoxicity assay. (A) Chemical structure of Aloperine. (B) Effects of Aloperine on cytotoxicity in EJ and SV-HUC-1 cells were determined by MTT assay. Experiments were performed at least three times with similar results. n=5; *P<0.05 and ** P<0.01 vs control group. |
Therefore, this study investigated the antitumor potential and the underlying molecular mechanisms of ALO on the BC cell line EJ in vitro. This study is expected to contribute to the therapy of BCs. We detected whether ALO plays a role in the viability, invasion, and apoptosis of BC EJ cells and elucidate the underlying mechanism. The results of this study suggest that ALO can be used to develop a potential therapeutic agent for BCs.
Materials and Methods
Reagents and Antibodies
Dulbecco’s Modified Eagle’s Medium (DMEM; high glucose), trypsin/EDTA solution, and fetal bovine serum (FBS) were purchased from Gibco® (Thermo Fisher Scientific, Waltham, MA, USA). Cell Counting Kit-8 (CCK)-8 and the Annexin V Apoptosis Detection kit I were acquired from MultiSciences Biotech Co., Ltd. (Hangzhou, Zhejiang, People’s Republic of China). ALO, PCNA, Bax, and Bcl-2 antibodies were purchased from Abcam, Inc. (Cambridge, UK). Caspase-3, Caspase-9, Ras, p-Raf1, Raf1, Erk1/2, p-Erk1/2, and GAPDH antibodies were purchased from Cell Signaling Technology, Inc. (Beverly, MA, USA). These antibodies were monoclonal and derived from rabbit. Hoechst 33,342 kits were obtained from Wuhan Boster Biological Technology Ltd. (Wuhan, China). The EZ-Cytox cell viability assay kit (MTT) was acquired from MultiSciences Biotech Co., Ltd. (Hangzhou, China). Trans-well chambers were obtained from Corning Co. (Corning, NY, USA). The protein assay kit was obtained from Bio-Rad Labs. (Hercules, CA, USA).
Cell Culture and Treatment
The human BC cell line EJ was purchased from the American Type Culture Collection (Manassas, VA, USA). The human urothelium cell line SV-HUC-1(normal control) was purchased from the Cell Bank of the Chinese Academy of Sciences (Shanghai, China), cultured in DMEM medium (containing 5.5 mmol/L D-glucose, normal glucose; NG), and supplemented with 10% FBS, 1% streptomycin, and penicillin in a humidified incubator with 95% air and 5% CO2 at 37°C. EJ cells were seeded in complete medium at 70-80% confluence in six-well plates. The function of ALO on the apoptosis and proliferation of EJ cells was determined. After culturing for 24 h, the medium was replaced with serum-free media. The cells were treated with different ALO concentrations (0, 25, 50, and 100 µmol/L) and incubated in a humidified incubator at 37°C for 12, 24, and 48 h. Then, the expression levels of related proteins and experiment were determined. All experiments were conducted in serum-free conditions and at least repeated thrice.
Cell Counting Kit-8 (CCK-8) Assay
The viabilities of EJ cells were measured by CCK-8 assay. In brief, cells treated with or without ALO were seeded into 96-well plates at a density of 2000 cells per well and then incubated for 24 h. Cell viability was measured using the CCK-8 method, in which 10 µL of the CCK-8 reagent was added and the cells were incubated at 37°C for 12, 24, and 48 h for proliferation measurements. Viability was detected using a PerkinElmer Victor3 1420 Multilabel Counter (PerkinElmer, Inc., Waltham, MA, USA).
Hoechst 33342 Staining
EJ cells at a density of 2×104 cells/well were seeded in 24-well plates. After incubation for 24 h, the cells were grown in fresh culture media containing ALO (0, 25, 50, and 100 µmol/L) and then incubated in a humidified incubator at 37°C for 24 h. Then, these cells were stained with Hoechst 33342 (10 mg/mL). After 20 min of culture at room temperature and in the dark, the cells were washed twice with PBS and observed immediately with a fluorescence microscope (original magnification, 400×; BX53; Olympus Corporation).
Cytotoxicity Assay
The MTT assay was performed to detect the survival of EJ and SV-HUC-1 cells. In the presence of an electronic-coupling reagent, mitochondrial dehydrogenase may be reduced to form orange formazan. Following treatment, the experimental cells were seeded into 96-well plates at a density of 6.0×103 cells/well. Subsequently, 10 µL MTT reagent was added to the plates for 45 min at 37°C, according to the manufacturer’s protocol. Cytotoxicity was measured at 6, 12 and 24 h (ALO concentrations 100 µmol/L). Optical density (OD) in each well was determined at 460 nm using a PerkinElmer VICTOR3 1420 Multilabel Counter (PerkinElmer, Inc., Waltham, MA, USA). The MTT assay was repeated three times.
Apoptosis Analysis
Cells apoptosis were performed using Annexin V Apoptosis Detection kit I. In brief, EJ cells at a density of 2×104 cells/well were treated with indicated doses of ALO (0, 25, 50, and 100 µmol/L) for 24 h. The treated cells were digested with trypsin and washed twice in with 1×PBS (4°C) in cold water. The cell pellet was resuspended with 500 µL of 1×binding buffer. Then, 5 µL of Annexin V-PE was added to the cell suspension in the dark at room temperature for 15 min in accordance with the manufacturer’s instructions. A 7-AAD solution (5 µL) was added in the cell suspension 5 min before flow cytometry analysis, and then 200 µL of 1×binding buffer was added for flow cytometry analysis. Stained cells were determined using a FACS Calibur system (BD, Franklin Lakes, NJ, USA) to examine apoptotic cells. Data analysis was performed using the ModFit software.
Wound Scratch Assay
EJ cells were cultured in fresh culture media to full confluence. A wound was created using a plastic scraper. After being washed twice with PBS, the medium was replaced with fresh culture media or fresh culture media containing ALO (0, 25, 50, and 100 µmol/L) and then incubated at 37°C for 24 h. Then, these cells were washed twice with PBS, and the wound was observed under a microscope (Olympus, Tokyo, Japan).
Transwell Migration and Invasion Assay
Cell migration and invasion assays were performed using. Transwell chamber (24-well, 8 µm pore-size membranes) was used in accordance with the manufacturer’s protocol. In invasion assay, after thawing at 4°C overnight, the Matrigel was diluted in serum-free medium, and 30 µL of the diluted Matrigel was evenly inoculated into the upper chamber to form a gel at 37°C. EJ cells (4×104 per well) were seeded in the upper compartments in 100 µL of serum-free medium, and 600 µL of medium with 20% FBS was added to the lower compartments as a chemoattractant at the same time. For trans-well migration and invasion assays, the EJ cells remaining at the upper surface of the membrane after incubating at 37°C for 24 h were removed with cotton swabs, and the cells on the lower surface of the membrane were the migrated cells. The cells that migrated or invaded to the lower surface of the membrane were fixed with 4% paraformaldehyde, stained in 10% crystal violet, and then counted under a microscope (Olympus, Tokyo, Japan).
Immunofluorescence
Indirect immunofluorescence was used on the control and ALO-treated EJ cells on coverslips. The cells were washed with PBS at 37°C, fixed in 4% paraformaldehyde for 10 min, and then permeabilized using 0.5% Triton X-100 buffer for 5 min at room temperature. Afterward, the cells were extensively washed thrice with PBS. These slides were blocked with 1% bovine serum albumin in PBS for 20 min at 4°C and then incubated with the primary antibodies as described above. The following polyclonal antibodies were used: anti-PCNA (1:100). The cells were stained with fluorescein isothiocyanate secondary antibodies (1:100) to visualize the primary antibodies. Stained cells were mounted using the antiquenching fluorescence mounting medium and viewed using an Olympus-BX51 fluorescence upright microscope (Olympus Corporation). Quantitation of immunofluorescence staining was carried out on coded cell coverslips as the integrated option density value.
Western Blot Analysis
EJ cells were harvested in RIPA lysis buffer containing protease inhibitor cocktail to measure protein concentrations using the BCA assay kit. Protein samples (30 µg) were loaded onto each lane, resolved using 10% SDS-PAGE, and then transferred onto activated polyvinylidene fluoride membranes. After blocking with 5% skim milk at room temperature for 1 h, the membranes were incubated with appropriate primary antibodies at 4°C overnight. After incubation with fluorescence-labeled secondary antibodies (IRDye700 and IRDye800, goat anti rabbit) for 1 h at 37°C, immunoreactive bands were visualized using an Odyssey infrared imaging system (LI-COR Biosciences, Lincoln, NE, USA).
Statistical Analysis
All statistical analyses were performed using SPSS 20.0 (IBM Corp., Armonk, NY, USA). Data are expressed as means ± standard deviation. One-way ANOVA, followed by the Student–Newman–Keuls test, was used for the quantitative data, whereas the Kruskal–Wallis test was used for non-normally distributed data. Statistical significance was considered at p <0.05.
Results
The Cytotoxic Effects of ALO on Human Urothelium Cells
EJ and SV-HUC-1 cells cytotoxicity were evaluated using the MTT assay. Low cytotoxicity was detected in the SV-HUC-1 groups. Conversely, ALO markedly enhanced EJ cytotoxicity, and peak at 24 h after application (Figure 1B).
ALO Inhibited the Proliferation of Human BC Cells
EJ cells were treated with ALO (0, 25, 50, and 100 µmol/L) for 12, 24, and 48 h to investigate the effect of ALO on the proliferation of human BC EJ cells. The results of CCK-8 assay showed that ALO suppressed cell proliferation in a dose-and time-dependent manner compared with the control (0 µmol/L) (Figure 2A). Compared with the control group, the groups treated with ALO at 25 and 50 µmol/L for 12 h showed no significant difference (p>0.05), but the difference was statistically significant when the concentration was 100 µmol/L (p<0.05). Compared with 0 h at the time point, the optimal concentration of ALO was 100 µmol/L at 12, 24, and 48 h with statistical significance (p<0.05). The difference at 24 h was the most obvious, indicating that 24 h is the best action time of ALO.
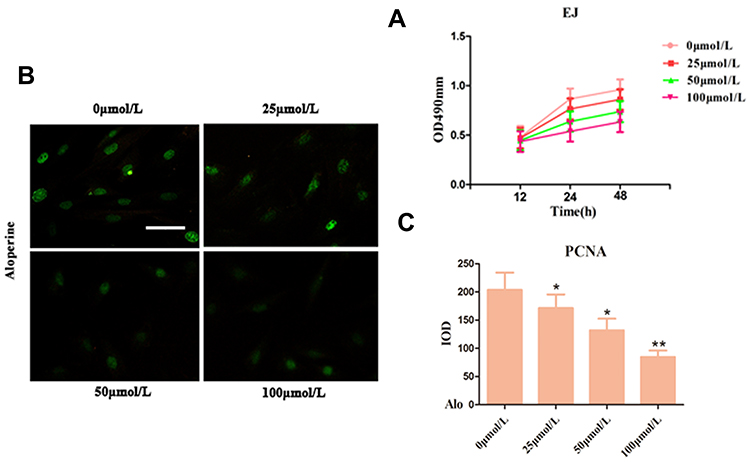 |
Figure 2 Effect of Alo inhibits human bladder cancer cell proliferation by CCK-8 and immunofluorescence analyses. (A) The EJ cells proliferation was evaluated was detected using the CCK-8 method at the indicated time points following treatment with Alo (0, 25, 50, and 100 µmol/L). (B) PCNA was detected following cultured with Alo 100µmol/L for 24 hours on immunofluorescence. (C) Quantification of IF staining-positive area by Image-Pro Plus 7.0 system. Immunofluorescent images of under the Olympus-BX51 fluorescence upright microscope were shown (scale bars, 40µm; magnification, ×400). The results were expressed as the mean ± standard deviation of three independent experiments and each was performed in triplicate. n=7; *P<0.05 and **P<0.01 vs non-Alo treated group (0µmol/L). Abbreviations: Alo, Aloperine; IOD, integrated optical density; PCNA, proliferating cell nuclear antigen; IF, immunofluorescent. |
Subsequently, immunofluorescence staining was performed after 24 h of treatment with ALO, and quantitative analysis of integrated option density values showed a significant reduction in PCNA compared with the control (Figure 2B and C). The results revealed that ALO reduced the PCNA of EJ cells compared with the untreated group.
ALO Induced the Apoptosis of Human BC Cells
Hoechst 33342 staining was used to determine whether ALO induces the apoptosis of human BC EJ cells on the basis of a series of morphological changes. The nuclei of EJ cells were uniformly stained in the untreated group, indicating these cells showed complete cell membrane morphology. However, the EJ cells treated with ALO (0, 25, 50, and 100 µmol/L) for 24 h clearly exhibited significant morphological changes compared with the untreated group, indicating that ALO significantly increased the apoptosis of EJ cells (Figure 3A). In addition, the effect of ALO on the apoptosis of EJ cells was analyzed by flow cytometry with Annexin V-FITC/PI staining. After treatment of EJ cells with ALO (0, 25, 50, and 100 µmol/L), the degree of apoptosis in the ALO group was significantly higher than that in the untreated group (Figure 3B and E). These findings suggest that ALO dramatically abated the proliferation of EJ cells in a dose-dependent manner possibly by increasing their apoptosis. Furthermore, the expression levels of apoptosis-related proteins, including Caspase-3, Caspase-9, Bax, and Bcl-2, were analyzed by Western blot analysis. ALO upregulated the expression levels of Caspase-3, Caspase-9, and Bax and downregulated the expression of Bcl-2 compared with the untreated group in EJ cells (Figure 3C and D).
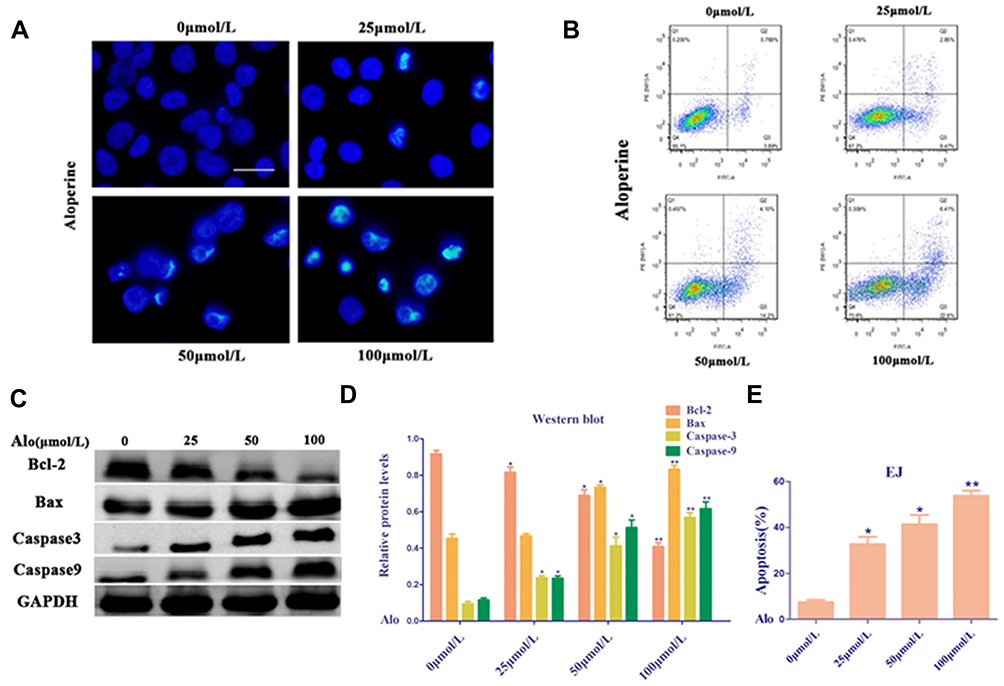 |
Figure 3 Alo induces bladder cancer cell apoptosis. (A) Images of apoptotic EJ cells incubated with vehicle or Alo (25,50,100µmol/L) for 24 h, as determined by Hoechst 33342 staining (scale bars, 40 µm). (B) EJ cells exposed to Alo (25,50,100µmol/L) for 24 h were harvested and analyzed by flow cytometry. (C) Western blotting was performed to analyze the expressions of Bax, Bcl-2 and Caspase-3 following Alo (25,50,100µmol/L) treatment for 24 h. Then the band intensity was quantified by ImageJ software. (D) The analysis of Western blotting. (E) The analysis of flow cytometry. The results were expressed as the mean ± standard deviation of three independent experiments and each was performed in triplicate. n = 5; *P<0.05 and **P<0.01 vs non-Alo treated group (0µmol/L). Abbreviation: Alo, Aloperine. |
ALO Suppressed the Migration and Invasion of Human BC Cells
Trans-well migration and wound healing assays were conducted to determine the effects of ALO on the migration and invasion of EJ cells. ALO dramatically suppressed the migration and invasion of EJ cells in a dose-dependent manner compared with the untreated group (Figure 4A and B). ALO treatment in EJ cells decreased the percent of invasion cells compared the untreated group (Figure 4C). These results indicated that ALO significantly depressed the migration and invasion of EJ cells.
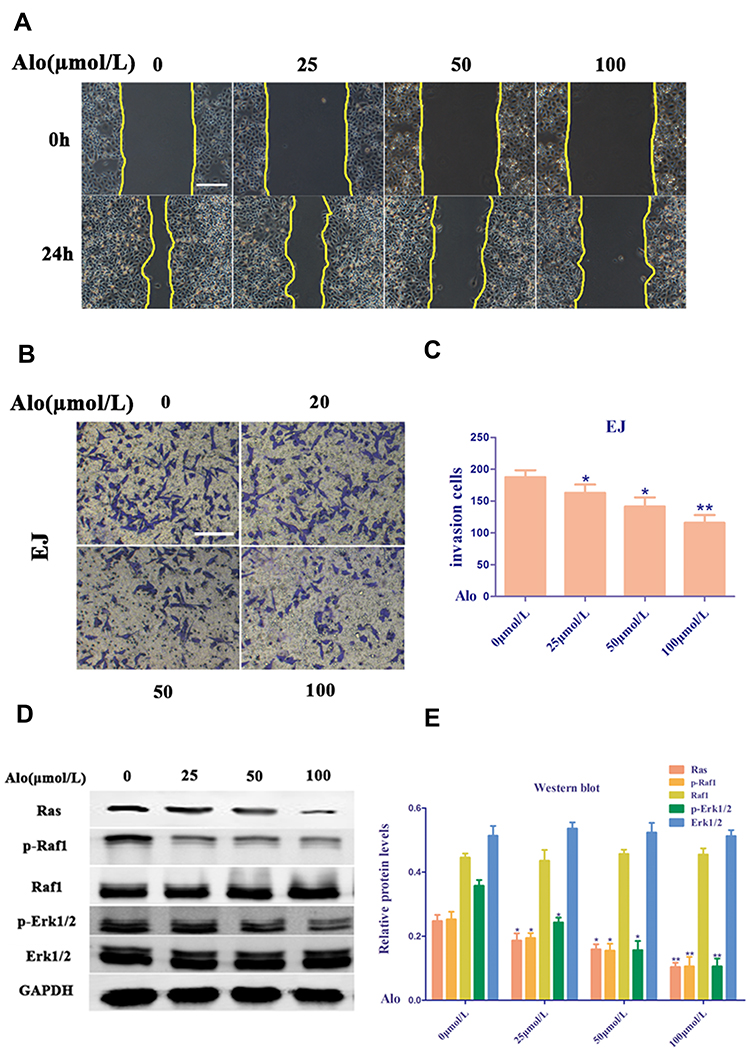 |
Figure 4 Alo suppresses human bladder cancer cell migration and invasion. (A) The EJ cells non-directional migration was detected by wound healing assay (scale bars, 160 µm; magnification, ×100). (B) The EJ cells directional migration was evaluated by Transwell assay (scale bars, 80µm; magnification, ×200). (C) The analysis of Transwell assay. (D) Western blotting was performed to analyze the expression of Ras, p-Raf1, Raf1, p-Erk1/2 and Erk1/2 following Alo (100µmol/L) treatment for 24 h. Then the band intensity was quantified by ImageJ software. (E) The analysis of Western blotting. The results were expressed as the mean ± standard deviation of three independent experiments. n=6; *P<0.05 and **P<0.01 vs non-Alo treated group (0µmol/L). Abbreviation: Alo, Aloperine. |
ALO Blocked the Ras Pathway in Human BC Cells
The Ras family is generally recognized as an oncogene. Three characteristic genes in the Ras family associated with human tumors are h-ras, k-ras, and n-ras, which are located on chromosomes 11, 12, and 1, respectively. In addition, the Ras/Raf-1/Erk1/2 pathway is downstream of the epidermal growth factor receptor, which is closely related to the occurrence and prognosis of various cancers. Erk1/2 is mainly activated by the phosphorylation of various growth factors, ion rays, and hydrogen peroxide, and enters the nucleus to act on transcription factors, such as Erk1/2, c-myc, c-fos, c-jun, ATF, NF-κB, and AP-1, promoting the transcription and expression of certain genes closely related to cell proliferation and differentiation.19 Thus, this study showed that ALO suppressed the proliferation, migration, and invasion and induced the apoptosis of EJ cells via these pathways. Treatment of human BC EJ cells with ALO (0, 25, 50, and 100 µmol/L) downregulated the level of Ras and decreased the phosphorylation levels of Raf1 and Erk1/2 (Figure 4D and E). These data suggest that ALO modulates the proliferation, migration, and invasion and induces the apoptosis of human BC EJ cells probably via the Ras/Raf1/Erk1/2 signaling pathway.
Discussion
BC is the second most common cancer of the genitourinary system, ranking 13th in mortality, although mortality rates are rising in most countries.6,22 Most are transitional epithelial cell carcinomas. The lateral and posterior walls of the bladder are the most common, followed by the trigonometric region and the apical region. Bladder tumors may be accompanied by renal pelvis, ureter, and urethra successively or simultaneously. In foreign countries, the incidence of bladder tumors is second only to prostate cancer in male genitourinary tumors. In China, it ranks first.23 The incidence rate in men is three to four times higher than that in women, and the age range is 50–70 years. Histological epithelial neoplasms account for 95% of this disease, among which more than 90% are transitional epithelial cell carcinomas.24,25 The EJ cell line is derived from malignant BC and is progressing to the muscle layer of the bladder wall, in contrast to cell lines derived from non-muscle-infiltrating BC, an early form of BC. At present, different treatment methods are available for BC depending on the pathological stage and grade.26 For the early initial bladder tumor, if the tumor is superficial and confined to the mucosa or lamina propria, transurethral bladder tumor resection, postoperative bladder perfusion therapy, and regular reexamination can be performed. Retaining the bladder may be life threatening in recurring and progressing BCs. For such patients, a radical cystectomy is usually required. Given the late progression of the disease, some patients may have a general diffusion or transfer. In addition, a patient with poor body conditions may not be able to accept radical bladder full cutting operation or chemical treatment. New immune therapy is recommended for such patients to provide a certain amount of care and relief.
The resistance of tumor cells and the side effects of chemotherapy drugs are other obstacles in tumor treatment.27 Therefore, the extraction of natural compounds from plants and animals has become the main research direction of anticancer drugs.7,28,29 At present, traditional Chinese medicine is still the mainstream of tumor prevention and treatment, and its characteristics of multiple components, links, and targets play an important role in the regulation of multiple genes, complex pathogenesis, and prevention and treatment of multiple tumors. Traditional Chinese medicine has good curative effect, high sensitivity to chemoradiotherapy, minimal toxic and side effects, and can prolong the survival time of patients. ALO is an alkaloid extracted from S. alopecuroides L. (Leguminosae), which is a type of traditional Chinese herb commonly used as an analgesic. Previous studies on ALO showed that it exhibits anti-inflammatory, antibacterial, and antiviral effects.30–32 Clinically, it has been widely used in the treatment of autoimmune diseases, such as rheumatism, lupus erythematosus, and eczema, in China, with remarkable efficacy.33 Recent studies have reported that ALO demonstrates anti-tumor effects on various tumor cells. As for its anti-cancer roles, ALO induces anti-tumor effects against several human cancers, including multiple myeloma, prostate cancer, breast cancer, thyroid cancer, osteosarcoma, and hepatocellular carcinoma.34 In addition, ALO can significantly induce apoptosis in colon cancer (SW480 and HCT116) cells.35 Although ALO induces cell cycle stagnation in the G2/M phase and simultaneously induces p21 and p53 expression to increase and decrease the expression of Cyclin D1 and B1, its specific target remains unclear. A clear understanding of the mechanism of ALO, particularly in patients with cancer, represents a matter of great interest for probable clinical practice in the future. However, whether ALO exhibits growth inhibitory effects on BC EJ cells and the underlying mechanisms remain unknown to date. Therefore, the present study investigated the effects of ALO on the proliferation, apoptosis, migration, and invasion of BC EJ cells and its mechanism.
Results showed that ALO dose- and time-dependently inhibited EJ cell proliferation, changed cell membrane morphology, and promoted EJ cell apoptosis. The Bcl-2 family includes pro-apoptotic and anti-apoptotic proteins. The pro-apoptotic Bcl-2 family protein Bax promotes apoptosis by activating Caspase and the release of mitochondrial cytochrome c, whereas the anti-apoptotic protein Bcl-2 inhibits apoptosis by blocking the release of cytochrome c. Our data shows that ALO can significantly lower the expression of Bcl-2. These results are similar to those of previous studies on the effects of ALO on other tumors.19,36,37 We assessed the effect of ALO on the activation of Caspase-3 and Caspase-9, and suggest that ALO induces apoptosis of EJ cells through a Caspase pathway. In addition, cytotoxicity tests suggested that ALO is low toxicity side effects to the human urothelium cells (normal control). In vitro experiments revealed that ALO inhibited the proliferation of human BC EJ cells. This result further confirmed that ALO has a positive inhibitory effect on the occurrence and progression of a variety of cancers, particularly BC. Another biological feature of tumor cells is invasion and metastasis, which are the leading causes of death in patients with BC. Thus, we performed wound healing, trans-well migration, and invasion assays on EJ cells to further confirm the effects of ALO on cell motility. Results showed that ALO can significantly suppress the migration and invasion of EJ cells.
Human tumors arise as a result of mutations in several genes that control the proliferation, differentiation, and apoptosis of normal cells, including activation of oncogenes and inactivation of tumor suppressor genes. About 30% of human tumors have mutations or activation of Ras genes and abnormal overexpression of Ras proteins.38,39 Therefore, the study of regulation of the Ras signal transduction pathway is important in the design of anti-tumor drugs targeting the cell signal transduction pathway. Ras protein is an important component of cell growth and proliferation signaling pathways.40,41 If the Ras protein is continuously activated, it can bind to the downstream effective protein and transmit the signal to the downstream signal element, which may lead to the abnormal proliferation of cells and the development of tumors. Therefore, we evaluated the role of ALO in Ras and its downstream signals. Results showed that ALO can significantly downregulate the Ras level and inhibit the phosphorylation levels of Raf1 and Erk1/2, indicating that ALO can block the Ras/Raf1/Erk1/2 signaling pathway. These results suggest that ALO exerts a good inhibitory effect on the occurrence and progression of human BC by blocking the Ras pathway.
Conclusion
In summary, compared with other alkaloids, ALO has the strongest effect on EJ cells in BC, and is low-toxic to human urothelium cells. ALO induces cell apoptosis and plays an anti-cancer role in human BC cells by inhibiting the proliferation, migration, and invasion of EJ cells. In addition, ALO-induced apoptosis may be involved in the inhibition of the Ras/Raf1/Erk1/2 signaling pathway. These results suggest that ALO as a chemotherapy-active drug in BC needs further study.
Abbreviations
ALO, Aloperine; ANOVA, analysis of variance; BC, bladder cancer; CCK-8, Cell Counting Kit-8; DMEM, Dulbecco minimum essential medium; MOI, multiplicity of infection; MTT, 3-(4,5-cimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide; PCNA, proliferating cell nuclear antigen.
Funding
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.
Disclosure
The authors report no conflicts of interest in this work. Lijun Zhang, Jun Liang contributed to the work equally and should be regarded as co-first authors.
References
1. Wang X, Ding Y, Wang J, Wu Y. Identification of the key factors related to bladder cancer by lncRNA-miRNA-mRNA three-layer network. Front Genet. 2020;10.
2. Xu Y, Wu G, Li J, et al. Screening and identification of key biomarkers for bladder cancer: a study based on TCGA and GEO data. Biomed Res Int. 2020;2020:1–20.
3. Wang K, Ma L, Tang J, et al. LncRNA00518 promotes cell proliferation through regulating miR-101 in bladder cancer. J Cancer. 2020;11(6):1468–1477. doi:10.7150/jca.35710
4. Efiloglu O, Basok BI, Turan T, et al. Role of serum and urine transforming growth factor beta 1, matrix metallopeptidase 9, tissue inhibitor of metalloproteinase 2, and nerve growth factor beta levels and serum neutrophil-to-lymphocyte ratio in predicting recurrence and progression risks in patients with primary non-muscle invasive bladder cancer. Turkish J Urol. 2020;1–7.
5. Hindupur SV, Schmid SC, Koch JA, et al. STAT3/5 inhibitors suppress proliferation in bladder cancer and enhance oncolytic adenovirus therapy. Int J Mol Sci. 2020;21:3. doi:10.3390/ijms21031106
6. D’Andrea D, Black PC, Zargar H, et al. Impact of sex on response to neoadjuvant chemotherapy in patients with bladder cancer. Urologic Oncol. 2020;38:
7. Wu P, Meng X, Zheng H, et al. Kaempferol attenuates ROS-induced hemolysis and the molecular mechanism of its induction of apoptosis on bladder cancer. Molecules. 2018;23:10. doi:10.3390/molecules23102592
8. Kim WT, Seo SP, Byun YJ, et al. Garlic extract in bladder cancer prevention: evidence from T24 bladder cancer cell xenograft model, tissue microarray, and gene network analysis. Int J Oncol. 2017;51(1):204–212. doi:10.3892/ijo.2017.3993
9. Rose A, El-Leithy T, Vom Dorp F, et al. Mistletoe plant extract in patients with nonmuscle invasive bladder cancer: results of a phase Ib/IIa single group dose escalation study. J Urol. 2015;194(4):939–943. doi:10.1016/j.juro.2015.04.073
10. Mao Q, Guo F, Liang X, Wu Y, Lu Y. Aloperine activates the pi3k/akt pathway and protects against coronary microembolisation-induced myocardial injury in rats. Pharmacology. 2019;104(12):90–97. doi:10.1159/000500761
11. Chang Z, Zhang P, Zhang M, et al. Aloperine suppresses human pulmonary vascular smooth muscle cell proliferation via inhibiting inflammatory response. Chin J Physiol. 2019;62(4):157–165. doi:10.4103/CJP.CJP_27_19
12. Zhao J, Zhang G, Li M, Luo Q, Leng Y, Liu X. Neuro-protective effects of aloperine in an Alzheimer’s disease cellular model. Biomed Pharmacother. 2018;108:137–143. doi:10.1016/j.biopha.2018.09.008
13. Yin W, Han J, Zhang Z, Han Z, Wang S. Aloperine protects mice against bleomycin-induced pulmonary fibrosis by attenuating fibroblast proliferation and differentiation. Sci Rep. 2018;8(1):6265. doi:10.1038/s41598-018-24565-y
14. Zhang X, Liu Q, Zhang N, et al. Discovery and evolution of aloperine derivatives as novel anti-filovirus agents through targeting entry stage. Eur J Med Chem. 2018;149:45–55. doi:10.1016/j.ejmech.2018.02.061
15. Wang C, Choi YH, Xian Z, Zheng M, Piao H, Yan G. Aloperine suppresses allergic airway inflammation through NF-κB, MAPK, and Nrf2/HO-1 signaling pathways in mice. Int Immunopharmacol. 2018;65:571–579. doi:10.1016/j.intimp.2018.11.003
16. Song S, Chen Y, Han F, et al. Aloperine activates the Nrf2‑ARE pathway when ameliorating early brain injury in a subarachnoid hemorrhage model. Exp Ther Med. 2018.
17. Ren D, Ma W, Guo B, Wang S. Aloperine attenuates hydrogen peroxide-induced injury via anti-apoptotic activity and suppression of the nuclear factor-κB signaling pathway. Exp Ther Med. 2017;13(1):315–320. doi:10.3892/etm.2016.3962
18. Hu S, Zhang Y, Zhang M, et al. Aloperine protects mice against ischemia-reperfusion (IR)-induced renal injury by regulating PI3K/AKT/mTOR signaling and AP-1 activity. Mol Med. 2016;21(1):912–923. doi:10.2119/molmed.2015.00056
19. Tian D, Li Y, Li X, Tian Z. Aloperine inhibits proliferation, migration and invasion and induces apoptosis by blocking the Ras signaling pathway in human breast cancer cells. Mol Med Rep. 2018;18(4):3699–3710.
20. Liu JS, Huo CY, Cao HH, et al. Aloperine induces apoptosis and G2/M cell cycle arrest in hepatocellular carcinoma cells through the PI3K/Akt signaling pathway. Phytomedicine. 2019;61:152843. doi:10.1016/j.phymed.2019.152843
21. Lee Y-R, Chen S-H, Lin C-Y, et al. In vitro antitumor activity of aloperine on human thyroid cancer cells through caspase-dependent apoptosis. Int J Mol Sci. 2018;19(1):312. doi:10.3390/ijms19010312
22. Chen J, Sun Y, Ou Z, et al. Androgen receptor-regulated circFNTA activates KRAS signaling to promote bladder cancer invasion. EMBO Rep. 2020;e48467.
23. He YT, Li DJ, Liang D, et al. [Incidence and mortality of bladder cancer in China, 2014]. Zhonghua Zhong Liu Za Zhi. 2018;40(9):647–652. Chinese.
24. Chen J, Li Y, Li Z, Cao L. LncRNA MST1P2/miR‐133b axis affects the chemoresistance of bladder cancer to cisplatin‐based therapy via Sirt1/p53 signaling. J Biochem Mol Toxicol. 2020;34. doi:10.1002/jbt.22452
25. Sun J, Zhang H, Tao D, et al. CircCDYL inhibits the expression of C-MYC to suppress cell growth and migration in bladder cancer. Artificial Cells, Nanomed Biotechnol. 2019;47(1):1349–1356. doi:10.1080/21691401.2019.1596941
26. Huang XX, Wang RX, Lin Q, et al. Inhibitory effects of 2-methoxyestradiol on cell growth and invasion in human bladder cancer T-24 cells. Pharmazie. 2017;72(2):87–90.
27. Li Y, Lv S, Ning H, et al. Down-regulation of CASC2 contributes to cisplatin resistance in gastric cancer by sponging miR-19a. Biomed Pharmacother. 2018;108:1775–1782. doi:10.1016/j.biopha.2018.09.181
28. Zhou L, Wei E, Zhou B, et al. Anti-proliferative benefit of curcumol on human bladder cancer cells via inactivating EZH2 effector. Biomed Pharmacother. 2018;104:798–805. doi:10.1016/j.biopha.2018.05.101
29. Shin -S-S, Park Y-J, Hwang B, et al. Triacanthine exerts antitumor effects on bladder cancer in vitro and in vivo. Phytomedicine. 2019;64:153069. doi:10.1016/j.phymed.2019.153069
30. Yu HI, Shen HC, Chen SH, et al. Autophagy modulation in human thyroid cancer cells following aloperine treatment. Int J Mol Sci. 2019;20:21. doi:10.3390/ijms20215315
31. Xu Z, Wang X, Chen X, et al. Identification of aloperine as an anti-apoptotic Bcl2 protein inhibitor in glioma cells. PeerJ. 2019;7:e7652. doi:10.7717/peerj.7652
32. Xu Y-Q, Jin S-J, Liu N, et al. Aloperine attenuated neuropathic pain induced by chronic constriction injury via anti-oxidation activity and suppression of the nuclear factor kappa B pathway. Biochem Biophys Res Commun. 2014;451(4):568–573. doi:10.1016/j.bbrc.2014.08.025
33. Wang H, Yang S, Zhou H, et al. Aloperine executes antitumor effects against multiple myeloma through dual apoptotic mechanisms. J Hematol Oncol. 2015;8:26. doi:10.1186/s13045-015-0120-x
34. Huang Y-X, Wang G, Zhu J-S, Zhang R, Zhang J. Traditional uses, phytochemistry, and pharmacological properties of Sophora alopecuroides L. Eur J Inflammation. 2016;14(2):128–132. doi:10.1177/1721727X16642779
35. Zhang L, Zheng Y, Deng H, Liang L, Peng J. Aloperine induces G2/M phase cell cycle arrest and apoptosis in HCT116 human colon cancer cells. Int J Mol Med. 2014;33(6):1613–1620. doi:10.3892/ijmm.2014.1718
36. Chen S, Jin Z, Dai L, et al. Aloperine induces apoptosis and inhibits invasion in MG-63 and U2OS human osteosarcoma cells. Biomed Pharmacother. 2018;97:45–52. doi:10.1016/j.biopha.2017.09.066
37. Muhammad T, Sakhawat A, Khan AA, et al. Aloperine in combination with therapeutic adenoviral vector synergistically suppressed the growth of non-small cell lung cancer. J Cancer Res Clin Oncol. 2020;146:861–874. doi:10.1007/s00432-020-03157-2
38. Guo YJ, Pan WW, Liu SB, Shen ZF, Xu Y, Hu LL. ERK/MAPK signalling pathway and tumorigenesis. Exp Ther Med. 2020;19(3):1997–2007.
39. Ta TV, Nguyen QN, Chu HH, Truong VL, Vuong LD. RAS/RAF mutations and their associations with epigenetic alterations for distinct pathways in Vietnamese colorectal cancer. Pathol Res Pract. 2020;216:152898. doi:10.1016/j.prp.2020.152898
40. Wiechmann S, Maisonneuve P, Grebbin BM, et al. Conformation-specific inhibitors of activated Ras GTPases reveal limited Ras dependency of patient-derived cancer organoids. J Biol Chem. 2020;295:4526–4540. doi:10.1074/jbc.RA119.011025
41. Ibanez Gaspar V, Catozzi S, Ternet C, Luthert PJ, Kiel C. Analysis of Ras-effector interaction competition in large intestine and colorectal cancer context. Small GTPases. 2020;1–17. doi:10.1080/21541248.2020.1724596
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.