Back to Journals » Neuropsychiatric Disease and Treatment » Volume 22
Alcohol Cues Elicit Differences in Brain Activation Patterns of Abstinent Men and Women with Alcohol Use Disorders
Authors Sawyer KS ![]() , Oscar-Berman M
, Oscar-Berman M ![]() , Ruiz SM, Marinkovic K, Valmas MM, Harris GJ
, Ruiz SM, Marinkovic K, Valmas MM, Harris GJ
Received 12 September 2025
Accepted for publication 19 May 2026
Published 8 June 2026 Volume 2026:22 551536
DOI https://doi.org/10.2147/NDT.S551536
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Roger Pinder
Kayle S Sawyer,1– 4 Marlene Oscar-Berman,1– 3 Susan Mosher Ruiz,1– 3 Ksenija Marinkovic,5 Mary M Valmas,1– 3 Gordon J Harris3,6
1Psychology Research Service, VA Boston Healthcare System, Boston, MA, USA; 2Department of Anatomy & Neurobiology, Boston University Chobanian & Avedisian School of Medicine, Boston, MA, USA; 3Department of Radiology, Massachusetts General Hospital, Boston, MA, USA, Boston, MA, USA; 4Sawyer Scientific, LLC, Boston, MA, USA; 5Department of Psychology, San Diego State University, San Diego, CA, USA; 6Department of Radiology, Harvard Medical School, Boston, MA, USA
Correspondence: Kayle S Sawyer, Boston University Chobanian & Avedisian School of Medicine, 72 East Concord Street, Boston, MA, 02118, USA, Email [email protected]
Introduction: Cognitive, emotional, and behavioral abnormalities have been associated with alcohol use disorders (AUD). Because of their reward salience, photographs of alcoholic beverages can influence brain activation in individuals with AUD.
Methods: We employed fMRI to measure gender differences in brain activation elicited by alcohol cues in 84 men and women with and without a history of alcohol use disorders (ALC and NC, respectively). Brain activity was measured during performance on a delayed matching-to-sample (DMTS) working memory task; emotional faces served as the sample and match cues. During the delay period, intervening distractors were either reward-salient cues (alcoholic beverages) or neutral cues (nonalcoholic beverages or scrambled pictures).
Results: Region of interest analyses showed that the ALC group had significantly higher levels of activation throughout reward-related circuitry during alcohol distractor interference than during scrambled picture interference. Significant Group-by-Gender interactions were also observed: Compared to NC men (NCm), ALC men (ALCm) evidenced lower activation contrast between reward-salient cues and neutral cues in default mode network regions (including superior prefrontal and precuneus areas), while ALC women (ALCw) had more activation than NC women (NCw). Similar significant interactions were observed for task-regions (including superior parietal, lateral occipital, and prefrontal areas). Additionally, ALCw had higher DMTS accuracy scores than ALCm.
Discussion: The findings suggest that abstinent ALCm and ALCw differ in processing reward-salient cues, which can impact treatment and recovery.
Plain Language Summary: Brain reward regions activate highly when individuals with a history of alcohol use disorder view alcoholic beverages.
The brain regions identified subserve vision, memory, and judgment.
Opposite abnormalities in activation patterns appeared for men and women.
Keywords: alcoholism, reward, fMRI, gender, memory, emotion
Introduction
Alcohol use disorders (AUD) have been associated with deficits in cognitive and emotional functions.1 Because of their reward salience, alcohol cues such as pictures of alcoholic beverages elicit attentional bias and brain activation in individuals with AUD.2–4 Alcohol cues induce a hyperattentive state with attention drawn to the rewarding stimuli.5–8 Therefore, the cues selectively interfere with other cognitive abilities such as memory. Importantly, attentional bias toward alcohol-related stimuli also has been associated with level of craving, consumption, dependence, and physiological arousal.9–18
Functional magnetic resonance imaging (fMRI) studies of attentional biasing, and specifically cue reactivity, have often included either only men with and without a history of AUD, or groups of men and women with sample sizes too small to examine gender effects.4,19 However, gender impacts the ways in which alcohol affects the brain and behavior,20–29 due to interactions with physiological and social factors.30,31 In the present study with a cohort of men and women with and without AUD (ALCm, ALCw, NCm, and NCw), we examined gender differences using a delayed matching-to-sample (DMTS) task,32 which requires an attention-demanding kind of memory called working memory. The participants were required to remember photographs of emotional faces (the sample) while distracting pictures of alcoholic beverages, nonalcoholic beverages, or scrambled images intervened during the delay period. We chose faces as the sample stimuli, because in a prior study,33 we found that men with AUD had abnormally low brain activity in temporal limbic regions when viewing emotional face stimuli. Further, it has been shown that alcoholism is associated with impaired emotional perception, and specifically impaired emotional face decoding.22,34–37
It is important to note that two aspects of our DMTS task are relevant for differentiating cognitive and brain deficits in AUD, and these were analyzed separately: (1) attentional biasing, as indicated by brain responses to distractor cues, along with behavioral responses to probe faces, and (2) affective processing, as indicated by brain responses to the encoded faces.38,39 Therefore, we divided the results of the present project into two separate reports. One report40 described abnormalities in AUD-related brain activation to the encoded faces, and identified gender-related differences. For the present report, the data derived from the delay and match (the probe face) portions of the task allowed us to test the attentional biasing effect,41,42 wherein we expected alcohol cues, more than other cue types, to impair performance on memory for face identity. Thus, the present report contains the results pertaining only to the distractor and the probe face.
Functional MRI task contrasts reveal activation of multiple brain networks, and abnormalities in the default mode network (DMN) have been implicated in AUD and in psychiatric disorders.43,44 The DMN has been observed to be more active during storytelling, reading and memory tasks, imagining future scenarios, self-reference, rumination, and when the mind wanders while staring at a fixation cross during fMRI scanning.45–48 We refer to the DMN regions as fixation-regions because they are more active during the idle delay intervals when the fixation stimulus is presented between DMTS trials than during stimulus presentations. Cortical fixation-regions include: (1) an “anterior hub,” consisting of portions of the rostral anterior cingulate cortex (ACC), ventromedial prefrontal cortex, and medial superior frontal cortex; (2) a “posterior hub,” which includes portions of the posterior cingulate and precuneus, (3) the temporoparietal junction, which covers parts of the angular gyrus and inferior parietal lobule; and (4) the superior and middle temporal gyrus region.47,49 With functional connectivity analyses, these regions, as well as subcortical structures, including ventral tegmental area, nucleus accumbens, and caudate nucleus, also have been identified as part of the DMN.50
In addition to the fixation-regions, we examined task-regions, which are more engaged during the DMTS task than while looking at unengaging fixation crosses. Literature on distractor interference during working memory has suggested that task-regions involve a distributed network including prefrontal cortex, along with dorsal and ventral visual association cortex, which are necessary for attentional functioning51 and for inhibiting distracting visual stimuli.52,53 Additional task-regions involved in attention, working memory, and emotional processing, include the dorsal ACC and lateral prefrontal areas. The dorsal ACC in particular has been implicated in craving and attentional biasing.54
In advance of any analyses, we used prior literature to select ten a priori anatomically defined regions of interest (ROI) involved in cue exposure, distractor interference, craving, reward processing, salience, working memory, or emotional faces. The first nine ROI are the dorsolateral prefrontal cortex (DLPFC), ventrolateral prefrontal cortex (VLPFC), orbitofrontal cortex (OFC), insular cortex, parahippocampal gyrus, hippocampus, amygdala, fusiform, and ACC. Previous studies provide support for each of those nine a priori ROI and are detailed in the Appendix (Table A1).4,54–63 The tenth ROI is the “extended reward and oversight system” (EROS) as described and named in our previous papers.25,64 The EROS ROI is a single large but discontinuous composite ROI that had been created by combining 11 regions (seven of the ROI noted above, all but the VLPFC and fusiform), plus an additional four: nucleus accumbens, ventral diencephalon, subcallosal cortex, and temporal pole. Selection of these regions was based upon extant literature, and updated regionally-specific insight can be obtained from more recent reviews.65–68 For each of the ten ROI, we intended to confirm findings from each of the aforementioned studies that had identified abnormal activation by alcohol cues in AUD, and additionally to investigate differences between men and women. We expected lower brain activation in the ALC group in regions involved in facial identity and inhibition of distractor interference, but higher in regions responsible for reward salience.
In summary, we investigated brain activation for ten ROI, and for vertex-wise cortical analyses of fixation-regions and task-regions. We examined the accuracy of the participants’ memory for the face identities after exposure to attentionally salient pictures to test our hypothesis that alcohol cues would distract the ALC group more than the NC group. We determined how brain regions were activated by the distractor contrasts, how the contrasts differed for ALC and NC groups, and how those abnormalities varied by gender. We hypothesized that attentional biasing would be evident for the ALC group in the form of stronger brain activity contrasts (alcoholic beverage cues compared to nonalcoholic and scrambled stimuli). We conceptualize attentional biasing as the cognitive manifestation of cue reactivity; the diversion of working memory resources toward reward-salient distractors (alcoholic beverages) and away from maintaining task-relevant face identity. Thus, stronger brain activity contrasts for alcohol cues relative to neutral cues reflect both cue reactivity and attentional biasing within the task context.
Regarding gender differences, we made predictions based upon previously published work from our laboratory wherein we found that brain regions of ALCw (compared to NCw) were overactive in response to highly charged emotional stimuli.24 We hypothesized that the ALCw would evidence hyperactivation to alcohol stimuli (because they would be emotionally salient to the ALC group), whereas the activation contrasts for ALCm would be weaker. The emotional salience of alcoholic beverage stimuli is well established.55,69 We also expected to reinforce prior results24,33 showing diminished responses in ALCm than NCm during the processing of salient imagery, specifically for alcohol-related cues in regions making up the EROS ROI.
Methods
Participants
Participants in this study included 42 abstinent long-term ALC individuals (21 ALCw) and 42 NC controls (21 NCw), with comparable age, education, and IQ (see Table 1 in the Results). Participants were recruited through flyers placed in treatment and after-care facilities, the Boston VA Healthcare System facility, Massachusetts General Hospital, the Boston University School of Medicine, and in public places (eg, churches, stores), as well as through newspaper and internet advertisements This research was approved by the Boston University Chobanian & Avedisian School of Medicine Institutional Review Board (#H24686), VA Boston Healthcare System Institutional Review Board (#1017 and #1018), and the Partners Healthcare System Human Research Institutional Review Board (#2000P001891), and conforms with the principles of the Declaration of Helsinki. All participants gave written informed consent prior to participation, and they were compensated for their time.
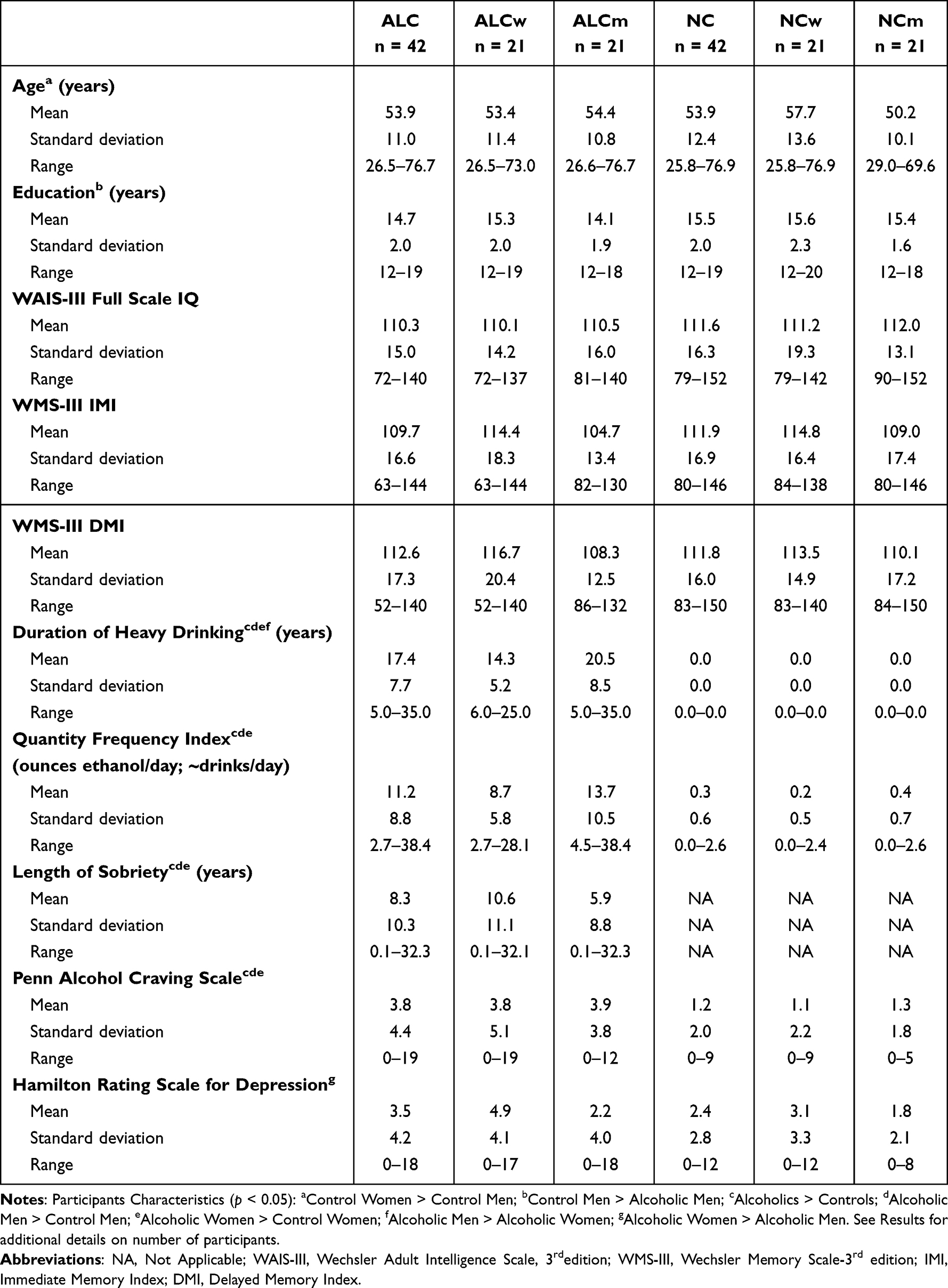 |
Table 1 Participant Characteristics |
Selection procedures began with a telephone interview to obtain age, education, health, alcohol and drug use history, including prescription drugs. Participants were right-handed, had normal or corrected-to-normal vision, and spoke English as their first language (or had acquired English as a second language by age five). Current drug use excepting nicotine was cause for exclusion, as were history of alcohol-related liver disease, epilepsy, head trauma resulting in loss of consciousness for 15 minutes or more, HIV, schizophrenia, or metal implants. Selection procedures were followed by self-reported psychological and behavioral assessments of participant characteristics.
Neuropsychological Assessment
Neuropsychological testing was conducted at the Department of Veterans Affairs (VA) Boston Healthcare System facility prior to scanning. Participants completed a medical history interview, vision test, handedness questionnaire,70 and a battery of tests as described below. All subjects were screened using the Hamilton Rating Scale for Depression71 and the Diagnostic Interview Schedule for the DSM-IV.72 The majority of participants also were administered the Wechsler Adult Intelligence Scale (WAIS-III) and the Wechsler Memory Scale (WMS-III).73 Four participants (two ALCw and two ALCm) received the WAIS-IV and WMS-IV,74 and WMS-III scores were not obtained from one ALCm. The scores for these participants were adjusted to account for differences in scoring outcomes relative to the earlier versions of the scales. Because craving for the rewarding effects of alcohol is known to serve as a trigger for relapse in those recovering from AUD,75 and alcohol cue exposure in particular is known to be related to relapse,76 all participants were administered the Penn Alcohol Craving Scale77 immediately before and approximately two weeks following the scan to assess any changes in alcohol craving patterns.
Alcohol Screening
The ALC participants met criteria for alcohol abuse or dependence, and consumed 21 or more alcoholic drinks per week for five or more years. Extent of alcohol use was assessed by calculating Quantity Frequency Index scores.78 The scores approximate the number of drinks consumed per day, and take into consideration the amount, type, and frequency of alcohol consumption either over the last six months (NC participants), or over the six months preceding cessation of drinking (ALC participants), and yields an estimate of ounces of ethanol per day. To remove the influence of current alcohol abuse, ALC participants must have been abstinent for at least four weeks before the scan date to be included. The ALC participants did not display symptoms of Korsakoff’s Syndrome nor dementia.79 Potential NC participants who had consumed 15–20 drinks per week for any length of time or who engaged in binge drinking were disqualified.
Functional Imaging Task
All participants were given a delayed matching-to-sample memory task32 in a magnetic resonance imaging (MRI) scanner, whereby they were asked to encode two faces that both had one of three emotional valences: positive, neutral, or negative (Figure 1). The face stimuli were shown in grayscale and were taken from a set of faces used in a previous study.33 These faces were displayed simultaneously for three seconds, followed by an asterisk (*) for one second. Subjects were asked to maintain these faces in memory while a colored distractor stimulus was shown. On different trials, the distractor stimulus was either a picture of an alcoholic beverage (alcbev; beer, wine, liquor, or mixed drink), a picture of a nonalcoholic beverage (nonalcbev; water, juice, milk, soda, coffee, tea, etc), or a scrambled nonsense picture (scrambled). Alcoholic and nonalcoholic beverage pictures were a combination of images used with permission from the Normative Appetitive Picture System (NAPS),80 and other previously published works on alcohol cues.60,61,81 Additional distractor images were modified from digital photographs taken at bars, liquor stores, and convenience stores. The scrambled images were created by inverting half the alcoholic and half the nonalcoholic beverage images and distorting them until they were not recognizable as any particular object, while preserving a match of primary visual characteristics. Each distractor picture was shown for three seconds, followed by an asterisk (*) for one second. Following the distractor picture, a single probe face was shown for two seconds, and the participants were instructed to report whether this face was one of the two faces they had just seen.
 |
Figure 1 Task presented during functional neuroimaging. The participants were shown two faces simultaneously for three seconds, followed by an asterisk (*) for one second. Next, a distractor was presented for three seconds. The probe face immediately followed, during which the participants had been trained to respond with a button press with either their index or middle finger to indicate whether the probe face matched the encoded face. During the inter-trial fixation interval, three crosses (+++) were shown on the screen for 2 to 30 seconds (mean 10 seconds). A total of 162 trials were presented. While the faces in this figure have been blurred and obscured in this publication to protect the identities of the individuals, the research participants saw the original unblurred and unobscured photographs. |
Each trial was 10 seconds in length, and was followed by a variable delay period (with a mean duration of 10 seconds, ranging from 2 to 22 seconds) during which the subject saw a set of crosshairs (+++) serving as a visual fixation. The task was divided into nine runs, each of which contained 18 trials. There were nine trial types made up of each combination of face valence and distractor type (eg, positive faces followed by alcohol distractor). Each emotion–distractor combination appeared twice per run. The stimulus order and variable inter-trial intervals were determined using optseq2 (http://surfer.nmr.mgh.harvard.edu/optseq), which optimizes statistical efficiency and hemodynamic response estimate accuracy for event-related experimental designs.82 In total, there were 54 trials for each distractor type (combined across facial expressions) and each face valence (combined across distractor types), for a total of 162 trials across the entire scan. The stimulus faces were balanced to contain 50% male and 50% female faces. Within a trial, the two encoded faces and probe face were matched on emotional expression and gender. This way, on match trials the probe facial image was identical to one of the encoded images, and on mismatch trials the facial identity changed but the emotional expression and gender did not.
The probe face matched one of the encoded faces on 50% of the trials, and match/mismatch trials appeared in a randomized order within each run. Responses were made by pressing one of two buttons with the index finger (match) or middle finger (mismatch) of the right hand. Participants were instructed to respond as quickly as possible without sacrificing accuracy. Additionally, participants could immediately correct a response by pressing the opposite button. To ensure the distractor images were viewed by all participants, they were told that it was necessary to pay attention to the pictures shown in between the faces on each trial, as they would be questioned about those images following the scan.
Behavioral Task Analyses
Responses to the face memory task were analyzed on the first level (individual subjects) using custom Excel templates. Trials were organized by Distractor type, facial Emotion, and Face Gender. Each trial was scored as correct, incorrect, or miss (ie, no response) When more than one response was made to a trial, the last response type (ie, yes/match or no/nonmatch) was accepted as the final answer, provided that the final response was at least 200 ms after the preceding response and no more than 10 s following the preceding response, When a single response was recorded and the reaction time was less than 200 ms, the trial was scored as a miss. For each participant, a mean overall reaction time (regardless of trial type) was calculated. Reaction times that exceeded three standard deviations from this mean were excluded from reaction time calculations by trial type. Participants’ patterns of responses were analyzed for consecutive misses to assure that they remained awake throughout the task. Three separate runs were identified, each in a different participant, wherein greater than five consecutive trials were missed; these runs were excluded from behavioral analyses.
Second level (group) effects on percent correct and reaction time (correct trials) were investigated using SPSS Version 17.0 (IBM, Chicago, IL, USA). Repeated-measures analyses of variance (ANOVA) were carried out with between-subjects factors of Group (ALC or NC) and Gender (female participant or male participant) and within-subjects factors of Distractor (alcbev, nonalcbev, or scrambled), Emotion (positive, negative, or neutral), and Face Gender (female face or male face). Repeated-measures ANOVA was selected for behavioral analyses given its robustness to moderate violations of normality assumptions with balanced sample sizes and continuous dependent variables such as percent correct and reaction times.
Image Acquisition
Imaging was conducted at the Massachusetts General Hospital’s Athinoula A. Martinos Center for Biomedical Imaging in Charlestown, MA. Data were acquired on a 3 Tesla Siemens (Erlangen, Germany) MAGNETOM Trio Tim MRI scanner with a 12-channel head coil. Sagittal T1-weighted MP-RAGE scans (TR = 2530 ms, TE = 3.39 ms, flip angle = 7°, FOV = 256 mm, slice thickness = 1.33 mm, slices = 128, matrix = 256 × 192) were collected for all subjects. For most participants, two such volumes were collected and averaged to aid in motion correction. An auto-align localizer was employed to adjust the acquired slices such that they ran parallel to an imaginary plane between the anterior and posterior commissures. Echo planar functional MRI blood oxygen level dependent (BOLD) scans were collected axially with 5 mm slice thickness and 3.125 × 3.125 mm in-plane resolution (64 × 64 matrix), allowing for whole brain coverage (32 interleaved slices, TR = 2 s, TE = 30 ms, flip angle = 90°). The event-related design included 18 trials per run with a total of nine runs. Within each six-minute run, 180 T2*-weighted volumes were collected. Functional volumes were auto-aligned to the anterior/posterior commissure line to ensure a similar slice prescription was employed across participants. Prospective Acquisition Correction (3D-PACE) was applied during collection of the functional volumes to minimize the influence of participants’ body motion.83 An IBM ThinkPad (Windows XP) running Presentation version 11.2 (NeuroBehavioral Systems, Albany, CA) software was used for visual presentation of the experimental stimuli and collection of participants’ responses. Stimuli were back projected onto a screen at the back of the scanner bore and were viewed by the participants through a mirror mounted to the head coil. All participants wore earplugs to attenuate scanner noise.
Structural Image Processing
Structural MPRAGE image analyses were performed for all participant data using the FreeSurfer (version 4.5.0) image analysis suite (http://surfer.nmr.mgh.harvard.edu). A multi-stage cortical surface reconstruction process was run on the two collected T1-weighted MP-RAGE scans,84 starting with motion correction, intensity normalization,85 Talairach registration,86 skull stripping,87 and segmentation88 of white matter, gray matter, and ventricles. Subsequently, boundaries were calculated delineating where gray and white matter meet, and where gray matter adjoins cerebrospinal fluid (“pial surfaces”) based on maximal shifts in image intensity between tissue types. These boundaries, as well as the subcortical segmentations, were visually inspected on each coronal slice for every subject, and manual interventions (eg, white matter volume corrections) were made when needed. The surface boundaries were used to generate computationally inflated two-dimensional cortical surface models, which allowed individual subjects to be registered to a spherical atlas by utilizing each subject’s cortical folding patterns. This registration was used to align the cortical geometry of all subjects within a group. Creation of these cortical surface models allowed improved data visualization as well as improved accuracy of within-group co-registration relative to an affine morph procedure.89 The cortical surface models were employed in an automated parcellation procedure that divides the surface into subregions based on gyral and sulcal anatomy. The Destrieux atlas parcellation for FreeSurfer90 was used to define anatomical ROI in the functional analyses for cortical areas, and the Desikan-Killiany FreeSurfer segmentation was used for subcortical areas.91
Functional Image Processing and Statistical Analyses
Effects of Group, Gender, Distractor, and Emotion on the BOLD signal were evaluated using both a whole-brain cluster analysis as well as ROI analyses. Processing of the functional data was performed using the FreeSurfer Functional Analysis Stream (FS-FAST), SPSS Version 17.0, and Matlab 7.4.0.
First-Level Functional Analyses
Preprocessing of the functional images for first level (individual subject) FS-FAST analysis included motion correction, intensity normalization,85 and spatial smoothing with a 5-mm Gaussian convolution kernel at full-width half-maximum. Trials were first combined across runs by distractor-emotional face valence pairs (ie, alcbev-positive, alcbev-negative, alcbev-neutral, nonalcbev-positive, nonalcbev-negative, nonalcbev-neutral, scrambled-positive, scrambled-negative, scrambled-neutral) and then collapsed across emotional valence. The BOLD response was estimated using a Finite Impulse Response (FIR) model, which allows for estimation of the time course of activity (percent signal change for a given condition) within a vertex or ROI for the entire trial period. For each condition, estimates of signal intensity were calculated for 2 pre-trial and 10 post-trial onset TRs, for a total analysis window of 24 seconds. Motion correction parameters calculated during alignment of the functional images were entered into the analysis as external regressors. Alignment of the T2*-weighted functional images with T1-weighted structural volumes was accomplished through an automated boundary-based registration procedure.92 These automated alignments were manually inspected to ensure accuracy.
Statistical maps were generated for each of the 84 individual subjects for contrasts between experimental conditions. Three contrasts were used to identify fixation-regions; they were made between distractor types and fixation: (1) alcbev vs. fixation, (2) nonalcbev vs. fixation, and (3) scrambled vs. fixation. Another three contrasts were used to assess cue responsivity: (1) alcbev vs. nonalcbev, (2) alcbev vs. scrambled, (3) nonalcbev vs. scrambled. Analyses of each of these contrasts included removal of prestimulus differences between the contrasted conditions by averaging the first three time points (two pre-trial onset and one post-trial onset) for each condition and subtracting this mean from each time point for that condition. Time points summed for inclusion in each contrast were chosen to reflect peak stimulus-related activity: Finite Impulse Response estimates of hemodynamic responses to the distractors were analyzed using a mean of the five TRs collected during the time period of 2–12 seconds post-distractor onset. Since the distractor is shown 4 seconds after the trial onset, the analysis window is 6–16 seconds following trial onset (time points 3 through 8).
Cortical Surface Cluster Analyses
We investigated cue-related brain activation in two separate cortical brain networks: (1) cue reactivity in fixation-regions, and (2) cue reactivity in task-regions. The brain network that was more active during presentation of the fixation cue than during the distractor images was used as our measure of fixation-regions. The network that was more active during the presentation of distractor images than during presentation of the fixation stimulus was used as our measure of the task-regions. In what follows, we first describe masking procedures and analyses we used to separate the networks.
The t-statistic maps for each condition vs. fixation were thresholded at p < 0.05 vertex-wise and were used to generate binary masks (Figure 2) separating fixation-regions and task-regions, thereby forming the six masks: alcbev greater than (or less than) fixation, nonalcbev greater than (or less than) fixation, scrambled greater than (or less than) fixation. These masks were used to separate the between-distractor analyses (described below).
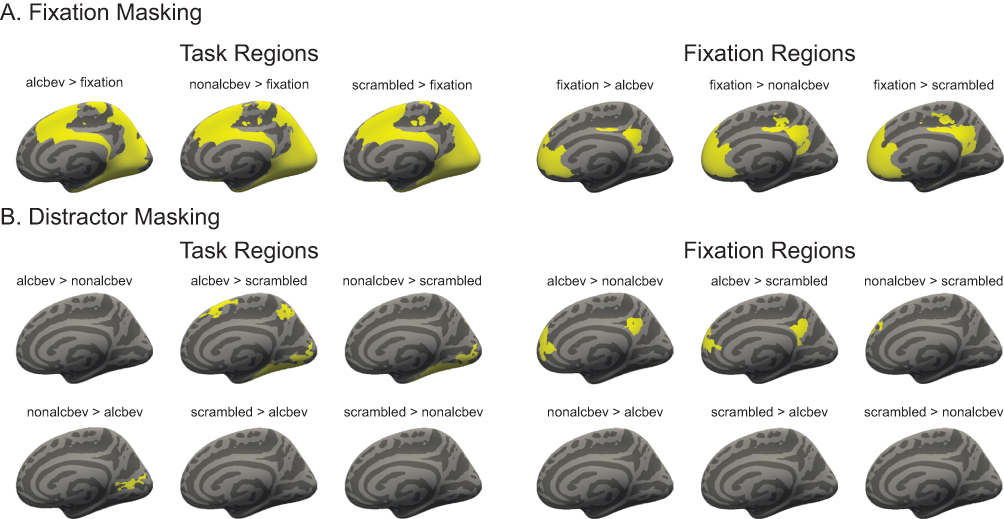 |
Figure 2 Analyses conducted with masked fMRI data for cortex. (A) The top set of brains represents analyses conducted for cortical regions in which network masking (for task and fixation-regions) were performed. (B) The bottom set of brains represents analyses conducted for cortical regions in which distractor masking (for task and fixation-regions) were performed. |
Second-level (group) analyses on cortical regions were accomplished using FS-FAST version 5.3, a surface-based morphing procedure for intersubject alignment and statistics.89 Group-averaged signal intensities during each experimental condition (alcbev, nonalcbev, scrambled) relative to fixation were calculated using the general linear model in spherical space for cortical regions, and were mapped onto the canonical cortical surface fsaverage, generating group-level weighted random-effects t-statistic maps masked to include only the cortex. Weighted random effects models were employed to reduce noise by considering individual subject variance. A 5 mm smoothing kernel (full-width half-maximum) was employed for all group and intergroup maps. Cluster correction on maps showing activity for each distractor condition vs. fixation was applied using FS-FAST Monte Carlo simulation with a clusterwise threshold of p < 0.05 corrected for two spaces (left hemisphere cortical and right hemisphere cortical), and additionally permutation testing with a vertex-wise threshold of p < 0.001 and a cluster-wise threshold of p < 0.05 corrected for two spaces (left hemisphere cortical and right hemisphere cortical). Cortical surface cluster regions were identified by the location of each cluster’s peak vertex on the cortical surface according to the Desikan-Killiany atlas.91
When we examined brain activation differences between the distractor types, we used three contrasts: alcbev vs. nonalcbev, alcbev vs. scrambled, and nonalcbev vs. scrambled. We investigated each direction of these contrasts separately. For example, brain regions with higher activation for alcbev than nonalcbev would be analyzed separately from those regions with higher activation for nonalcbev than alcbev.
Intergroup comparison t-statistic maps were generated using FS-FAST by comparing activation levels of all of the ALC participants with levels of all of the NC participants. Additionally, Group-by-Gender interaction maps for each contrast were calculated.
Region of Interest Analyses
The anatomically defined ROI for the distractor analyses included areas hypothesized a priori to be implicated in alcohol craving, distractor interference, and working memory for emotional faces, as described in the Introduction. Details, including atlas index numbers from the Destrieux90 parcellation (for cortical regions) and Desikan-Killiany91 segmentation (for subcortical regions), are provided in Table A1. The ROI were DLPFC, VLPFC, OFC, insular cortex, parahippocampal gyrus, hippocampus, amygdala, fusiform, ACC, and the multi-regional EROS.25,64 While not examined as a separate ROI, the PCC was included in the whole-brain cluster analysis, although recent research suggests the PCC is relevant to addictions.65 Left and right hemisphere regions were analyzed as separate ROI.
Statistical preprocessing and time course visualization of ROI data were performed using scripts written for Matlab version 7.4.0. Signal intensity for each region was averaged across all vertices (for surface-based ROI) or voxels (for volume-based ROI) included in the region for each condition on the individual participant level. To compute percent signal change for each participant within an ROI, signal estimate per condition and time point was divided by the average baseline activity for that participant. Time courses were normalized at the individual subject level for each condition by taking the mean of the first three time points (two pre-trial and one post-trial onset) and subtracting this mean from each time point. Group and Group-by-Gender averages of the normalized time courses were computed for each condition, and were visualized by plotting the percent signal change for each condition at each time point (ie, TR) of the trial.
For the distractor ROI analyses, percent signal changes of the BOLD signal within each ROI for the time window from 2 to 12 sec after distractor onset were entered as dependent variables into repeated-measures ANOVA models with between-group factors of Group (ALC or NC) and Gender (men or women) and within-subjects factor of distractor type (alcbev, nonalcbev, or scrambled).
Results
Research Participant Characteristics
Table 1 summarizes means, standard deviations, and ranges of participant demographics, drinking variables, and IQ and memory test scores.
The ALC and NC groups did not differ significantly by age. Although the NCw were older than the NCm, controls did not differ significantly from their respective ALC counterparts by age. ALCm had on average one year less education relative to NCm. Groups did not differ significantly on WAIS-III Full Scale IQ scores. While ALCw had higher Hamilton Rating Scale for Depression scores than ALCm, the average scores for all four subgroups (ALCm, ALCw, NCm, and NCw) were low. The mean scores for all subgroups were under 5, below the threshold for mild depression, which is 8. Therefore, depression likely contributed little to our observed gender differences.
By definition, the ALC group had longer durations of heavy drinking than the NC group. The ALCm on average drank heavily for six years more than did the ALCw. Five NCm and two NCw reported being lifetime abstainers. The ALC group reported higher levels of craving for alcohol than the NC group on the Penn Alcohol Craving Scale administered immediately prior to the scan; ALCm and ALCw did not differ on reported level of pre-scan alcohol craving. Eighty-one of the 84 participants were reached approximately two weeks after their scan date to be reassessed on alcohol craving level. One ALCw, one ALCm, and one NCw could not be reached for follow-up assessment on craving scores. Neither the ALC group nor the NC group displayed an increase in alcohol craving (ie, a significant change in craving scores) from the assessment on their scan date to their follow-up assessment.
Behavioral Results
Measures of participant performance on the face memory task were calculated for overall performance and for performance by each distractor type and facial emotion. Means, standard deviations, and ranges are reported for percent correct responses and reaction times in Tables A2 and A3, respectively.
A significant Group-by-Gender interaction was found for overall accuracy (F1,80 = 6.880, p = 0.01, Figure A1 and Table A2): ALCw had significantly higher accuracy scores than ALCm (mean 85.3% vs. 77.4%), and this gender difference was larger than that observed in the NC group. Accuracy and reaction times (Figure A2 and Table A3) did not vary significantly as a function of the Distractor, nor were there any significant interactions of Distractor with Group or Gender (all p > 0.05). The main effect of Emotion was significant for percent correct responses, wherein ALC and NC participants alike performed better on both positive and negative faces relative to neutral faces. Performance on positive and negative faces did not differ significantly. The effect of Emotion on percent correct responses did not vary as a function of Group or Gender (all p > 0.05). Regarding reaction time and Emotion, participants responded more quickly to positive face trials relative to neutral face trials; this effect did not vary by Group or by Gender. The main effect of Face Gender on percent correct responses was also significant (F1,80 = 6.80, p = 0.01), with the overall performance being better for male faces than for female faces. The effects of Group and Gender on percent correct responses and reaction times for Face Gender were not significant (all p > 0.05).
fMRI BOLD Effects
Effects of the distractors on the BOLD signal were assessed using group and intergroup cluster analyses for cerebral cortex, along with a-priori analyses of anatomical ROI that had been implicated by the literature. Below, we report group analyses of fixation contrasts and between-distractor conditions, followed by intergroup analyses of the same contrasts.
Cortical Cluster Analyses of Distractor Effects
Analyses of task contrasts revealed broadly similar activation patterns for the ALCw, ALCm, NCw, and NCm groups. During fixation, regions involved in the DMN were significantly more active than during the presentation of distractor images. As detailed in the Methods, these fixation-regions were masked and examined separately for subsequent analyses of contrasts between distractor types. Identical analyses were then performed for the task-regions. Significant clusters for between-distractor contrasts can be seen for fixation-regions first (Figures 3 and A3), and then for task-regions (Figures 4 and A4).
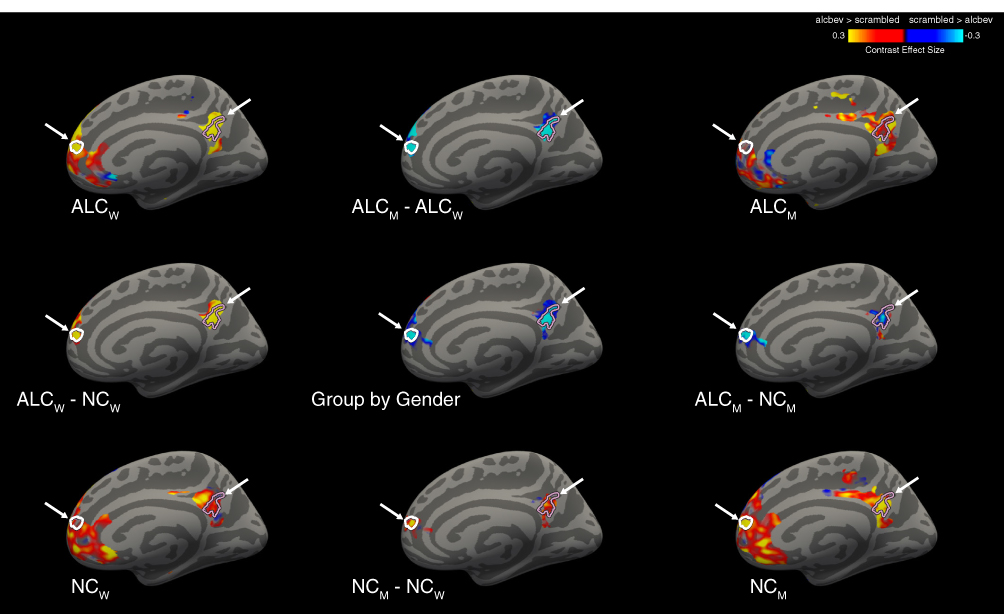 |
Figure 3 Cortical clusters significant for Group-by-Gender interactions for the alcbev vs. scrambled contrast within fixation-regions (right medial view). A significant Group-by-Gender interaction revealed several clusters (see Table A5), two of which are indicated by arrows on the medial surface of the right hemisphere, with cluster outlines overlaid on contrast values between alcbev and scrambled distractors. Group mean contrast values (for alcbev vs. scrambled within fixation-regions) are displayed in the four brain images located in the corners of the figure, and group comparisons are indicated by minus signs. Abbreviations: ALCm, Alcoholic men; ALCw, Alcoholic women; NCm, Nonalcoholic men; NCw, Nonalcoholic women. |
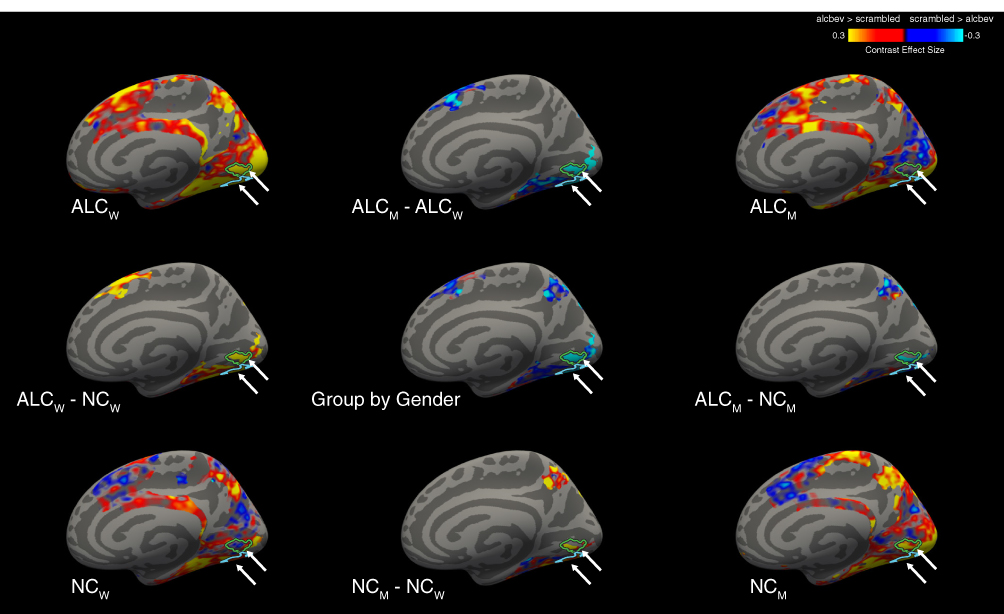 |
Figure 4 Cortical clusters significant for Group-by-Gender interactions, for the alcbev vs. scrambled contrast within task-regions (right medial view). A significant Group-by-Gender interaction revealed several clusters (see Table A5), two of which are indicated by arrows on the medial surface of the right hemisphere, with cluster outlines overlaid on contrast values between alcbev and scrambled distractors. Group mean contrast values (for alcbev vs. scrambled within task-regions) are displayed in the four brain images located in the corners of the figure, and group comparisons are indicated by minus signs. Abbreviations: ALCm, Alcoholic men; ALCw, Alcoholic women; NCm, Nonalcoholic men; NCw, Nonalcoholic women. |
All four groups had more brain activity (Table A4) in response to alcbev than scrambled distractors in the four main fixation-regions (the anterior and posterior medial hub regions, the temporal parietal junction, and the middle temporal gyrus), while the nonalcbev vs. scrambled contrast was less consistent. The alcbev vs. nonalcbev contrast generally indicated higher activation for the alcbev than nonalcbev. For task-regions, alcbev and nonalcbev elicited higher activation than scrambled in the occipital lobe and adjoining visual areas in temporal and parietal cortex (Table A4 and Figure A4).
Distractor Intergroup Cluster Analyses
The pattern of results indicated that ALCw and NCm had strong activation contrasts (beverages > scrambled) in visual areas and the medial fixation-regions, especially the posterior hub. The ALCw had greater activation contrast than NCw, while ALCm had lower activation contrast than NCm. Table 2 summarizes the regions where significant Group-by-Gender interactions were found for the distractor contrasts, ie, for alcbev vs nonalcbev, alcbev vs. scrambled, and nonalcbev vs. scrambled. Table A5 provides all significant Group-by-Gender interactions, and Figures 3, 4, A3, A4 illustrate alcbev vs. scrambled contrasts. In total, we observed 22 clusters where the Group-by-Gender interaction was statistically significant: Seven in fixation-regions and 15 in task-regions.
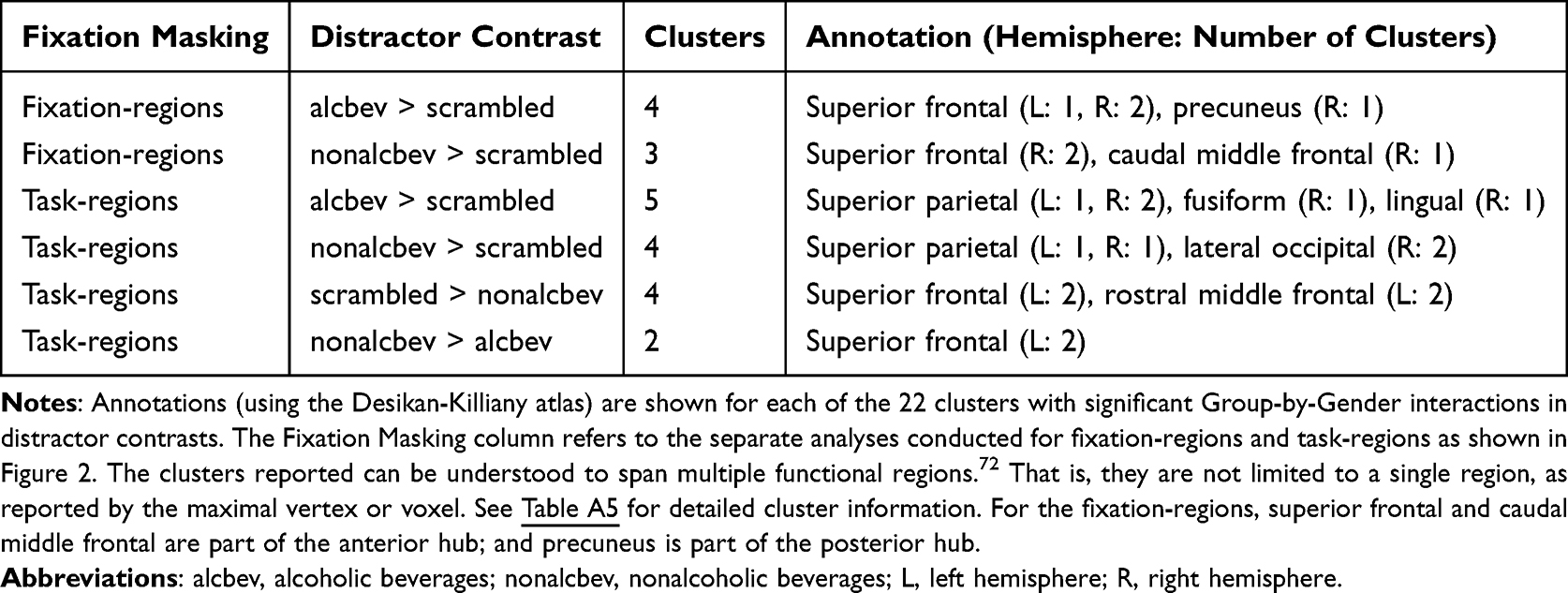 |
Table 2 Group-by-Gender Cortical Cluster Summary |
For the seven clusters in fixation-regions, six were in the anterior hub and one was in the posterior hub. For alcbev > scrambled, we observed one cluster in the posterior hub and three in the anterior hub. For three of these four clusters, ALCw and NCm had the strongest contrasts; in the fourth cluster, ALCw had the strongest contrast, whereas ALCm had the weakest. The remaining three clusters were found for nonalcbev > scrambled. As with the alcbev contrasts, ALCw and NCm had the strongest contrasts. All three clusters were in the right hemisphere of the anterior hub. Thus, the overall pattern consistent among the seven clusters was as follows: NCm had stronger contrast (beverage > scrambled) than ALCm, while muted or opposite direction comparison was observed for the women.
Of the 15 task-regions clusters, two were found for nonalcbev > alcbev (left superior frontal cortex). In both clusters, NCw had higher activation to nonalcoholic beverages, while NCm had higher activation to alcoholic beverages. In the other 13 clusters, the strongest contrasts were found for NCm and ALCw. Of those, the significant interactions were found in visual regions where nonalcbev > scrambled, while significant interactions in frontal regions were found for scrambled > nonalcbev.
Setting aside gender, analyses of activation levels comparing ALC and NC groups revealed 15 cortical clusters with significantly greater contrast levels for the ALC group than for the NC group (Table A6). Two of the clusters were in fixation-regions: one alcbev-region (temporoparietal junction), and one nonalcbev-region (posterior hub). The other 13, all alcbev-regions, were in task-regions located throughout the cortex: 7 frontal, 3 temporal, 2 parietal, and 1 occipital.
Distractor Region of Interest Analyses
Results of ANOVAs examining between-subjects effects of Group and Gender and within-subjects effects of Distractor type on BOLD percent signal change within each ROI are summarized in Table 3. Reported means and standard deviations represent the percent signal change across each ROI (unmasked) for the time period of 2 to 12 seconds post-distractor stimulus onset. These anatomically defined ROI included regions in EROS areas,25,64 as well as in regions associated with face memory maintenance and distractor interference, as described in the Introduction and Methods.
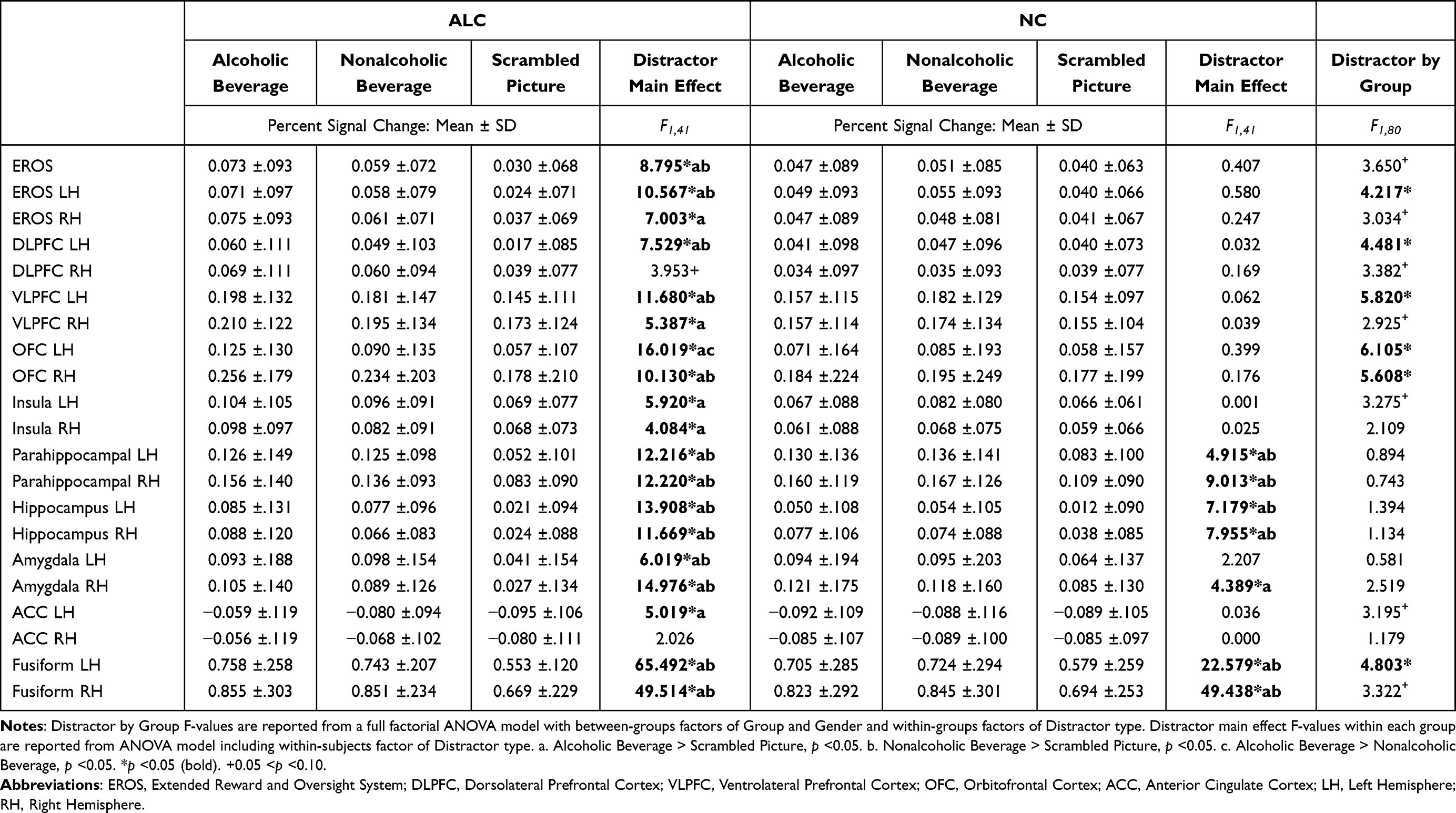 |
Table 3 Percent Signal Change for Each Distractor Type by Group, for a-Priori Regions |
Results from the ROI analyses of distractor effects indicated strong effects of distractor type on neural activation patterns. Specifically, among all participants, significantly higher responses were observed in several regions (EROS, left DLPFC, left VLPFC, right OFC, bilateral parahippocampal gyrus, bilateral hippocampus, bilateral amygdala, and bilateral fusiform) to both alcbev and nonalcbev relative to scrambled stimuli. The effect was significantly larger for the ALC group than the NC group in the left EROS and left fusiform regions, and in four prefrontal brain areas (left DLPFC, left VLPFC, left OFC, and right OFC). Additionally, in the alcbev vs. nonalcbev contrast, the ALC group showed significantly higher responses than the NC group in the left OFC ROI. Figure 5 shows percent signal change over time for the OFC, VLPFC, fusiform, and ACC activation, to demonstrate representative activity patterns. The left OFC shows the heightened activation for alcbev for the ALC group, the fusiform and VLPFC show higher activation in both groups to both beverage types, and the ACC demonstrates how activation is lower during distractor presentation than during fixation (for both groups and for all three distractor types).
 |
Figure 5 ROI - Percent signal changes in regions of interest for ALC and NC participants by distractor type. The regions of interest are: (A) Left OFC; (B) Left VLPFC; (C) Left ACC; and (D) Left Fusiform. The percent signal change represents brain activity during presentation of fixation and the task stimuli. Error bars represent the standard error of the mean. Time zero was set to the onset of the encoded faces, and signal zero was set to the average signal for the three initial time points (two pre-trial and one post-trial onset). The analysis window used to examine the distractor was 6 to 16 seconds following trial onset (2 to 12 seconds after distractor onset), as indicated by the thick line on the x axis. The ACC (C) is part of the anterior hub within the fixation-regions. The remaining areas are primarily task-regions. Abbreviations: OFC, orbitofrontal cortex; VLPFC, ventrolateral prefrontal cortex; ACC, anterior cingulate cortex. |
Applying a Bonferroni correction accounting for all 25 tests would set a critical value for statistical significance of main effects and Distractor, Group, and Gender interactions at p < 0.002. Using this threshold, main effects of Distractor would remain significant in left OFC, bilateral parahippocampal gyrus, bilateral hippocampus, right amygdala, and bilateral fusiform gyrus. All main effects of Group and Gender, as well as all interaction effects were p > 0.002.
Discussion
Behavioral Responses to Probe Face and Distractor Cues
In this study, ALC and NC participants alike were able to use emotional information to improve memory as assessed by a DMTS task, as has been previously reported in NC participants.32,93,94 This was evidenced by better memory performance (ie, higher accuracy) on positive and negative faces, and faster reaction times to positive faces, than neutral faces.
We had postulated that, compared to the NC group, the ALC group would make more recognition errors after alcohol distractors (relative to other distractor types). However, we found that regardless of the group, the distractor type did not substantially influence accuracy or reaction time. Although we also had expected performance by the ALC group to be impaired by alcoholic beverage cues, a significant interaction between group and distractor type was not evident. We did not find a behavioral attentional bias effect: The alcoholic beverage distractors (relative to nonalcoholic beverage or scrambled picture distractors) did not disproportionately decrease the number of correctly recalled faces among either the ALC or the NC group. As suggested in a review by Field and Cox,5 the strength of a bias effect among ALC groups may depend upon their drinking histories. Interestingly, Loeber et al 51 found that reduced attentional biasing to alcohol cues was associated with longer durations of heavy drinking, and our sample also had long durations. Further, participants with short vs. long lengths of abstinence might use different cognitive strategies, which could be reflected in both behavioral performance and brain activity. While the results were consistent with findings by Loeber and colleagues51 regarding chronicity, direct correlational analyses between drinking duration and attentional bias effects were not conducted in the present study, and findings would benefit from research with larger samples. Although combining participants with variable drinking histories might have masked the attentional biasing effect, measures of neural activity were more sensitive than behavioral measures. That is, the activation levels reflected attentional bias to beverage cues even when combining participants with differing patterns of drinking and abstinence.
Distractor fMRI Contrasts
The fMRI contrasts revealed broadly similar patterns of brain activity among the four subgroups, for both fixation-regions and task-regions. In both brain networks, the beverages (alcbev and nonalcbev) elicited higher activation than the scrambled distractors. This amounts to internal replications of our present fMRI results, with four independent samples (ALCm, ALCw, NCm, and NCw) revealing the same fixation-regions, same task-regions, and with mostly the same direction of effects for task contrasts within those regions. The results reflect the existence, location, and extent of the DMN,47,49 and also indicate that beverage pictures elicit higher activation than scrambled pictures in the DMN. Moreover, our results indicate that DMN regions are sensitive to the informational content of visual stimuli. In task-regions, the occipital lobe, along with adjoining visual areas in temporal and parietal cortex, were clearly more activated by beverage stimuli than by scrambled images. The results further suggest that content processing is not solely performed by those visual regions, because activation to beverage cues was identified in middle to posterior cingulate regions, which are involved in a multitude of cognitive functions.95,96
Gender Differences
The present research provides further evidence for the importance of considering gender when exploring effects of alcoholism on the brain.97,98 Many factors contribute to the observed differences in function for abnormalities identified comparing ALCm with NCm vs. ALCw with NCw.
Cortical group-level cluster analyses revealed significant Group-by-Gender interaction effects in 22 clusters. The general pattern of those findings indicated that ALCm had lower activation contrasts than NCm, while ALCw had higher activation contrasts than NCw. This pattern was observed primarily in contrasts between beverage and scrambled distractor conditions, and they were found in the two core medial DMN regions, as well as in visual association cortices. A similar pattern of results was found in a previous report24 in which emotional vs. neutral image contrasts were lower in ALCm than NCm, and stronger in ALCw than NCw. The lower brain reactivity for ALCm, and higher for ALCw, highlighted gender effects, suggesting possible differences in underlying bases for development of AUD. Of note, the results from other modalities also have indicated similar directions of the fMRI effects, with ALCw having larger reward regions than NCw, and higher fractional anisotropy than NCw, as compared to the smaller regions and lower fractional anisotropy found for ALCm than NCm.25,27
The gender-divergent abnormalities in the anterior and posterior hub regions of the DMN could be reflective of other gender differences observed in conjunction with AUD. The role of these regions in internal monitoring could relate to differences in pre-existing risk factors,30,31,99 or could represent differential consequences of alcohol abuse.100 The pattern of higher activation contrasts in ALCw versus ALCm localized to posterior DMN regions (precuneus) aligns with findings by Tschuemperlin and colleagues,101 who reported stronger implicit association-related activation localized to PCC and precuneus in female compared to male AUD patients. This convergence suggests that gender-divergent DMN responses to alcohol-related stimuli may reflect differential engagement of self-referential or memory-related processes during cue exposure. A similar pattern of group differences was identified in cortical regions associated with visual processing. That is, the results could represent a more fundamental impact that is not regionally specific. In the present study, effects of both increased activation in reward regions and decreased deactivation in DMN regions in response to alcohol pictures were strongest among ALCw in particular. One reason stronger alcohol cue-specific responses were observed among ALCw could be related to gender-based differences in physiological responses to alcohol cues,102 and with respect to gender-divergent responses to stress and craving.18 Aligned with this, larger responses to alcohol cues by female social drinkers relative to male social drinkers have been reported in superior and middle frontal gyri.103
Group Differences
In addition to significant gender interactions, we identified regions with differences between the ALC and NC groups. Cluster analyses showed that the ALC group had higher activation in two fixation-regions and 13 task-regions (Table A6). Differences in the posterior hub were identified in regions with stronger activation to nonalcoholic beverages than to scrambled images, while the other clusters had stronger activation to alcoholic beverages. The higher contrasts observed for the ALC group indicate a processing bias toward beverage cues across fixation- and task-regions. However, the stronger activation observed in the ALC group during alcohol distractors also may reflect engagement of executive control processes necessary to overcome attentional capture and maintain task performance.104,105 The absence of behavioral impairment despite elevated neural activation could indicate successful compensatory control processes, consistent with models suggesting enhanced executive control activation in addiction contexts to maintain behavioral regulation.65,68,106,107
Brain Responsivity in the Default Mode Network
Compared to the NC group, cluster analyses showed that the ALC group, and the ALCw in particular, had stronger contrasts in the anterior and posterior hubs, along with the temporoparietal junction. The results for ROI that include DMN regions also support the finding of contrast dampening in response to alcohol cues. Abnormal DMN functioning has been observed in other addictions and neuropsychiatric conditions.108–110 In AUD, abnormal functional connectivity among DMN regions has been reported.111,112 ALCw in particular had stronger contrasts for both the anterior and posterior hubs, an abnormality which could indicate a limitation in the level of detail processed, or the way in which it is integrated.113 Lower activation of the anterior hub specifically has been associated with dynamic attention allocation during task executions.114 This suggests that reduced deactivation of this region in AUD individuals when viewing alcohol pictures could be associated with failure to reallocate attention back to the task. That is, the ALC group could have decreased attention to the alcohol distractor cues. While the ALCw group’s abnormal DMN activation in conjunction with strong task performance may seem counter-intuitive, another explanation involves an interaction between task demands and DMN functions.115 In other words, in conjunction with equivalent or superior memory performance, our results suggest that, for ALCw especially, parts of the DMN might disengage, and show higher connectivity with control networks during highly demanding working memory tasks.
Brain Responsivity in Regions of Interest
The ROI analysis of the OFC provides evidence for reward-specific processing in the ALC group. In particular, reward-specific processing refers to their higher activation from the contrast between alcbev and nonalcbev. Several studies have reported enhanced OFC activation to alcohol cues,58,61,116,117 and research has established the role of the OFC in alcohol and drug addiction more generally.3 We had initially hypothesized lower activation for the ALC group in regions subserving facial identity maintenance and distractor inhibition. However, an alternative hypothesis also could be supported by our ROI findings in the OFC and EROS. That is, higher activation to alcoholic beverages in reward-salient regions may override or compete with task-related processes, thereby disrupting working memory maintenance via attentional capture rather than by diminished reward responsivity. The OFC activity may be particularly important for preoccupation and anticipation stages of the addiction cycle.118 Additionally, activity in this region has been shown to correlate with subjective craving ratings of viewed alcohol cues,60 and further correlated with relapse risk.119
In the regions where significant interactions of Group and Distractor type were identified, the distractor effect was found to be significant among the ALC group, but not among controls. In the EROS, DLPFC, VLPFC, OFC, and fusiform, the differences in activation levels among beverage distractor conditions (alcoholic and nonalcoholic) were larger relative to scrambled pictures in the ALC group than in the NC group. The higher responsivity of the ALC group to alcoholic beverages supports our hypothesis of greater attentional bias in the form of stronger alcbev vs. nonalcbev activity contrasts, but we did not expect the ALC group to have greater activation than the NC group to nonalcoholic beverages relative to scrambled cues. One explanation for this result is that many of the nonalcoholic beverages contain caffeine or sugar (eg, coffee, tea, soda), which, like alcoholic beverages, also stimulate reward-network activity120 As was suggested in an earlier meta-analysis,121 the attentional bias for caffeine-related cues may correlate more strongly with subjective craving than for alcohol-related cues. Moreover, craving for both caffeine and alcohol utilize similar neural circuits as are used for processing alcohol reward,122 as do the effects of sugar-related reward.123,124
Limitations
Although we observed a significant Group-by-Gender interaction in overall accuracy, we did not observe significant attentional bias (Distractor type) on behavior, despite significant findings of the bias effects in brain activation. This may reflect differences in sensitivity of the two types of measures, in the abstinence characteristics of our cohort, or in sample sizes too small to detect complex Group-by-Gender-by-Distractor interactions. Furthermore, behavioral measures may not be normally distributed, and violation of parametric assumptions may limit the interpretability of the repeated-measures ANOVA conducted. Those with long sobriety may have developed different cognitive strategies than AUD participants at an early stage of abstinence, especially strategies to deal with alcohol stimuli and to overcome craving. In that case, combining AUD participants with long and short LOS may have masked behavioral measures of attentional bias of accuracy and reaction time, while preserving the more sensitive and robust brain activation measures. Moreover, gender differences within the AUD group could be probed with sobriety as a covariate, or a subgroup analysis with narrow range could be conducted, although these analyses were beyond the scope of this report. Likewise, the variability in other participant characteristics such as IQ and age may diminish disclosure of AUD-related differences that additional research with larger samples could remedy.
It is not clear to what degree the abnormalities we observed result from, or predate, heavy drinking. In any case, our AUD cohort had drinking history values representative of the national population,125 thereby improving the generalizability of our results. The mean abstinence period for the ALC group was 8.3 years (range 4 weeks to 32 years). These values represent cross-sectional data, limiting conclusions regarding the trajectory of changes in brain activation or behavioral responses over the course of abstinence. Importantly, however, the processing deficits in AUD populations may vary by gender, and short- and long-term abstinence may have different paths of recovery for men and women.28
In conjunction with the multiple-comparison cluster correction procedures employed, the significance level we used (p < 0.05) has been shown to have higher false-positive rates than expected.126 Therefore, we conducted additional analyses to control false-positive errors with strict permutation testing (with a vertex-wise threshold of p < 0.001 and a cluster-wise threshold of p < 0.05), which has been shown to avoid the unexpectedly high false-positive rates identified for Gaussian field theory-based cluster-correction.126,127 However, the use of strict thresholds increases the chance of false-negative errors, and the stricter significance level may have resulted in us missing real gender effects; therefore, we reported the entirety of the group comparisons (with Tables A5 and A6 reflecting the main results, and Tables A7 and A8 reflecting the results using strict permutation testing). Although we report cluster labels by the location of the peak voxel or vertex, the clusters reported can be understood to span multiple functional regions.128 That is, they are not limited to a single region, as reported by the maximal vertex or voxel.
In the present study, there were 21 participants in each of the four subgroups. These cohort sizes may be considered as underpowered for examining sex differences. This is a persistent limitation faced by alcohol researchers studying sex differences, and we acknowledged that larger samples of male and female participants would strengthen our conclusions about sex differences.
Finally, our analyses did not include factors such as cigarette smoking, body mass index, and hormone therapy,1,129 which could possibly influence alcohol cue processing, reward, and DMN activity.
Conclusion
Compared to the NC group, the ALC group had stronger activation for the DMN, and overactivated reward regions during alcohol cue distraction. This suggests that attentional capture is not limited to reward regions, but also includes the DMN. If so, the DMN has a role in processing salient aspects of addictive substances.
The present study showed that alcohol cue distractors have powerful effects on reward-related regions of the brain, even in the absence of impaired performance when alcohol cues are employed as distracting stimuli. We also demonstrated that the increased responses in reward regions are accompanied by dampened DMN activity during the presentation of alcohol cues. Our results suggest that these effects are strongest among ALCw, and provide evidence for dimorphic patterns of responses to alcohol cues between ALCm and ALCw.
Abbreviations
ACC, anterior cingulate cortex; ALC, individuals with alcohol use disorders; ALCm, men with a history of alcohol use disorders; ALCw, women with a history of alcohol use disorders; ANOVAs, analyses of variance; AUD, alcohol use disorders; BOLD, blood oxygen level dependent; DIS, Diagnostic Interview Schedule; DMN, default mode network; DMTS, delayed matching-to-sample; DLPFC, dorsolateral prefrontal cortex; DMI, Delayed Memory Index; DSM-IV, Diagnostic Interview Schedule- 4th edition; EROS, extended reward and oversight system; fMRI, functional magnetic resonance imaging; FS-FAST, FreeSurfer Functional Analysis Stream; IMI, Immediate Memory Index; MRI, magnetic resonance imaging; NC, control participants without alcohol use disorders; NCm, women without a history of alcohol use disorders; NCw, men without a history of alcohol use disorders; OFC, orbitofrontal cortex; ROI, regions of interest; TR, repetition time; duration for collecting an entire fMRI volume; VLPFC, ventrolateral prefrontal cortex; WAIS-III, Wechsler Adult Intelligence Scale 3rd edition; WMS-III, Wechsler Memory Scale-3rd edition.
Data Sharing Statement
The original data contain protected health information and cannot be shared directly. Upon request to the corresponding author, specified portions may be able to be shared, depending upon institutional review board review.
Acknowledgments
This work was supported by funds from the US Department of Veterans Affairs Clinical Science Research and Development grant I01CX000326; National Institute on Alcohol Abuse and Alcoholism (NIAAA) of the National Institutes of Health US Department of Health and Human Services under Award Numbers R01AA007112, R01AA016624, K05AA00219, and K01AA13402; and shared instrumentation grants 1S10RR023401, 1S10RR019307, and 1S10RR023043 from the National Center for Research Resources (now National Center for Advancing Translational Sciences) at the Athinoula A. Martinos Center, Massachusetts General Hospital. Alcoholic and nonalcoholic beverage pictures were a combination of images used with permission from the Normative Appetitive Picture System (NAPS) (Stritzke et al, 2004), and other previously published works on alcohol cues (Wrase et al, 2002; Myrick et al, 2004). The authors thank Elinor Artsy, Sheeva Azma, Anne-Mette Guldberg, Zoe Gravitz, Doug Greve, Steve Lehar, Diane Merritt, George Papadimitriou, Alan Poey, Elizabeth Rickenbacher, Trinity Urban, and Robert Zondervan for assistance with consultation, manuscript preparation and recruitment, assessment, preparing testing materials, and neuroimaging of the research participants. We also greatly appreciate the helpful suggestions provided by the reviewers. Finally, we would like to acknowledge the role of the research participants for making this study possible. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health, the U.S. Department of Veterans Affairs, or the United States Government.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work. Primary contributors to study conception and design were MOB, SMR, KSS, KM, and GJH. The methodology, data curation, analyses, and figure creation were conducted primarily by SMR and KSS. Day-to-day operations were performed primarily by SMR, MMV, and KSS. The manuscript was written primarily by SMR, KSS, and MOB, and reviewed by all of the authors.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Oscar-Berman M, Valmas MM, Sawyer KS, Ruiz SM, Luhar RB, Gravitz ZR. Profiles of impaired, spared, and recovered neuropsychologic processes in alcoholism. Handb Clin Neurol. 2014;125:183–23.
2. Carter BL, Tiffany ST. Meta-analysis of cue-reactivity in addiction research. Addiction. 1999;94(3):327–340. doi:10.1046/j.1360-0443.1999.9433273.x
3. Goldstein RZ, Volkow ND. Drug addiction and its underlying neurobiological basis: neuroimaging evidence for the involvement of the frontal cortex. Am J Psychiatry. 2002;159(10):1642–1652. doi:10.1176/appi.ajp.159.10.1642
4. Schacht JP, Anton RF, Myrick H. Functional neuroimaging studies of alcohol cue reactivity: a quantitative meta-analysis and systematic review: alcohol cue imaging. Addict Biol. 2013;18(1):121–133. doi:10.1111/j.1369-1600.2012.00464.x
5. Field M, Cox W. Attentional bias in addictive behaviors: a review of its development, causes, and consequences. Drug Alcohol Depend. 2008;97(1–2):1–20. doi:10.1016/j.drugalcdep.2008.03.030
6. Franken IHA. Drug craving and addiction: integrating psychological and neuropsychopharmacological approaches. Prog Neuropsychopharmacol Biol Psychiatry. 2003;27(4):563–579. doi:10.1016/S0278-5846(03)00081-2
7. Townshend JM, Duka T. Attentional bias associated with alcohol cues: differences between heavy and occasional social drinkers. Psychopharmacology. 2001;157(1):67–74. doi:10.1007/s002130100764
8. Vollstädt-Klein S, Loeber S, Kirsch M, et al. Effects of cue-exposure treatment on neural cue reactivity in alcohol dependence: a randomized trial. Biol Psychiatry. 2011;69(11):1060–1066. doi:10.1016/j.biopsych.2010.12.016
9. Bordnick PS, Traylor A, Copp HL, et al. Assessing reactivity to virtual reality alcohol based cues. Addict Behav. 2008;33(6):743–756. doi:10.1016/j.addbeh.2007.12.010
10. Field M, Eastwood B. Experimental manipulation of attentional bias increases the motivation to drink alcohol. Psychopharmacology. 2005;183(3):350–357. doi:10.1007/s00213-005-0202-5
11. Field M, Mogg K, Zetteler J, Bradley BP. Attentional biases for alcohol cues in heavy and light social drinkers: the roles of initial orienting and maintained attention. Psychopharmacology. 2004;176(1):88–93. doi:10.1007/s00213-004-1855-1
12. Ryan F. Attentional bias and alcohol dependence: a controlled study using the modified stroop paradigm. Addict Behav. 2002;27(4):471–482. doi:10.1016/S0306-4603(01)00183-6
13. Sharma D, Albery IP, Cook C. Selective attentional bias to alcohol related stimuli in problem drinkers and non-problem drinkers. Addiction. 2001;96(2):285–295. doi:10.1046/j.1360-0443.2001.96228512.x
14. Sinha R, Fox HC, Hong KA, Bergquist K, Bhagwagar Z, Siedlarz KM. Enhanced negative emotion and alcohol craving, and altered physiological responses following stress and cue exposure in alcohol dependent individuals. Neuropsychopharmacology. 2009;34(5):1198–1208. doi:10.1038/npp.2008.78
15. Sawyer KS, Poey A, Ruiz SM, Marinkovic K, Oscar-Berman M. Measures of skin conductance and heart rate in alcoholic men and women during memory performance. PeerJ. 2015;3:e941.
16. Wiers CE, Stelzel C, Park SQ, et al. Neural correlates of alcohol-approach bias in alcohol addiction: the spirit is willing but the flesh is weak for spirits. Neuropsychopharmacology. 2014;39(3):688–697. doi:10.1038/npp.2013.252
17. Field M, Marhe R, Franken IHA. The clinical relevance of attentional bias in substance use disorders. CNS Spectr. 2014;19(3):225–230. doi:10.1017/S1092852913000321
18. Radoman M, Fogelman N, Lacadie C, Seo D, Sinha R. Neural correlates of stress and alcohol cue-induced alcohol craving and of future heavy drinking: evidence of sex differences. Am J Psychiatry. 2024;181(5):412–422. doi:10.1176/appi.ajp.20230849
19. Fryer SL, Jorgensen KW, Yetter EJ, et al. Differential brain response to alcohol cue distractors across stages of alcohol dependence. Biol Psychol. 2013;92(2):282–291. doi:10.1016/j.biopsycho.2012.10.004
20. Verplaetse TL, Cosgrove KP, Tanabe J, McKee SA. Sex/gender differences in brain function and structure in alcohol use: a narrative review of neuroimaging findings over the last 10 years. J Neurosci Res. 2021;99(1):309–323. doi:10.1002/jnr.24625
21. Becker JB, McClellan ML, Reed BG. Sex differences, gender and addiction. J Neurosci Res. 2017;95(1–2):136–147. doi:10.1002/jnr.23963
22. Hoffman LA, Lewis B, Nixon SJ. Neurophysiological and interpersonal correlates of emotional face processing in Alcohol Use Disorder. Alcohol Clin Exp Res. 2019;43(9):1928–1936. doi:10.1111/acer.14152
23. Rivas-Grajales AM, Sawyer KS, Karmacharya S, et al. Sexually dimorphic structural abnormalities in major connections of the medial forebrain bundle in alcoholism. NeuroImage Clin. 2018;19:98–105. doi:10.1016/j.nicl.2018.03.025
24. Sawyer KS, Maleki N, Urban T, et al. Alcoholism gender differences in brain responsivity to emotional stimuli. Elife. 2019;8. doi:10.7554/eLife.41723
25. Sawyer KS, Oscar-Berman M, Barthelemy OJ, Papadimitriou GM, Harris GJ, Makris N. Gender dimorphism of brain reward system volumes in alcoholism. Psychiatry Res Neuroimaging. 2017;263:15–25. doi:10.1016/j.pscychresns.2017.03.001
26. Seitz J, Sawyer KS, Papadimitriou G, et al. Alcoholism and sexual dimorphism in the middle longitudinal fascicle: a pilot study. Brain Imaging Behav. 2017;11(4):1006–1017. doi:10.1007/s11682-016-9579-5
27. Sawyer KS, Maleki N, Papadimitriou G, Makris N, Oscar-Berman M, Harris GJ. Cerebral white matter sex dimorphism in alcoholism: a diffusion tensor imaging study. Neuropsychopharmacology. 2018;43(9):1876–1883. doi:10.1038/s41386-018-0089-6
28. Fama R, Le Berre AP, Sullivan EV. Alcohol’s unique effects on cognition in women: a 2020 (Re)view to envision future research and treatment. Alcohol Res. 2020;40(2):03. doi:10.35946/arcr.v40.2.03
29. Kaag AM, Wiers RW, de Vries TJ, Pattij T, Goudriaan AE. Striatal alcohol cue-reactivity is stronger in male than female problem drinkers. Eur J Neurosci. 2019;50(3):2264–2273. doi:10.1111/ejn.13991
30. Ruiz SM, Oscar-Berman M. Gender and alcohol abuse: history and sociology. In: Martin SC, editor. Gender and Alcohol Abuse: History and Sociology. Vol 2. The SAGE Encyclopedia of Alcohol: Social, Cultural, and Historical Perspectives. Los Angeles: Sage Publications; 2015:586–591.
31. Mosher Ruiz S, Oscar-Berman M, Kemppainen MI, Valmas MM, Sawyer KS. Associations between personality and drinking motives among abstinent adult alcoholic men and women. Alcohol Alcohol. 2017;52(4):496–505. doi:10.1093/alcalc/agx016
32. Dolcos F, McCarthy G. Brain systems mediating cognitive interference by emotional distraction. J Neurosci. 2006;26(7):2072–2079. doi:10.1523/JNEUROSCI.5042-05.2006
33. Marinkovic K, Oscar-Berman M, Urban T, et al. Alcoholism and dampened temporal limbic activation to emotional faces. Alcohol Clin Exp Res. 2009;33(11):1880–1892. doi:10.1111/j.1530-0277.2009.01026.x
34. Oscar-Berman M, Hancock M, Mildworf B, Hutner N, Weber DA. Emotional perception and memory in alcoholism and aging. Alcohol Clin Exp Res. 1990;14(3):383–393. doi:10.1111/j.1530-0277.1990.tb00491.x
35. Philippot P, Kornreich C, Blairy S, et al. Alcoholics’ deficits in the decoding of emotional facial expression. Alcohol Clin Exp Res. 1999;23(6):1031–1038.
36. Clark US, Oscar-Berman M, Shagrin B, Pencina M. Alcoholism and judgments of affective stimuli. Neuropsychology. 2007;21(3):346–362. doi:10.1037/0894-4105.21.3.346
37. Lewis B, Price JL, Garcia CC, Nixon SJ. Emotional face processing among treatment-seeking individuals with alcohol use disorders: investigating sex differences and relationships with interpersonal functioning. Alcohol Alcohol. 2019;54(4):361–369. doi:10.1093/alcalc/agz010
38. Schweizer S, Satpute AB, Atzil S, et al. The impact of affective information on working memory: a pair of meta-analytic reviews of behavioral and neuroimaging evidence. Psychol Bull. 2019;145(6):566–609. doi:10.1037/bul0000193
39. Xu P, Peng S, Luo YJ, Gong G. Facial expression recognition: a meta-analytic review of theoretical models and neuroimaging evidence. Neurosci Biobehav Rev. 2021;127:820–836. doi:10.1016/j.neubiorev.2021.05.023
40. Oscar-Berman M, Ruiz SM, Marinkovic K, Valmas MM, Harris GJ, Sawyer KS. Brain responsivity to emotional faces differs in men and women with and without a history of alcohol use disorder. PLoS One. 2021;16(6):e0248831. doi:10.1371/journal.pone.0248831
41. Dowd EW, Pearson JM, Egner T. Decoding working memory content from attentional biases. Psychon Bull Rev. 2017;24(4):1252–1260. doi:10.3758/s13423-016-1204-5
42. Oscar-Berman M, Bonner RT. Matching- and delayed matching-to-sample performance as measures of visual processing, selective attention, and memory in aging and alcoholic individuals. Neuropsychologia. 1985;23(5):639–651. doi:10.1016/0028-3932(85)90065-X
43. Menon V. Large-scale brain networks and psychopathology: a unifying triple network model. Trends Cognit Sci. 2011;15(10):483–506. doi:10.1016/j.tics.2011.08.003
44. Zhang R, Volkow ND. Brain default-mode network dysfunction in addiction. Neuroimage. 2019;200:313–331. doi:10.1016/j.neuroimage.2019.06.036
45. Tops M, Boksem MAS, Quirin M, IJzerman H, Koole SL. Internally directed cognition and mindfulness: an integrative perspective derived from predictive and reactive control systems theory. Front Psychol. 2014;5:429. doi:10.3389/fpsyg.2014.00429
46. Beaty RE, Benedek M, Silvia PJ, Schacter DL. Creative cognition and brain network dynamics. Trends Cognit Sci. 2016;20(2):87–95. doi:10.1016/j.tics.2015.10.004
47. Buckner RL, DiNicola LM. The brain’s default network: updated anatomy, physiology and evolving insights. Nat Rev Neurosci. 2019;20(10):593–608. doi:10.1038/s41583-019-0212-7
48. Spreng RN, Mar RA, Kim ASN. The common neural basis of autobiographical memory, prospection, navigation, theory of mind, and the default mode: a quantitative meta-analysis. J Cogn Neurosci. 2009;21(3):489–510. doi:10.1162/jocn.2008.21029
49. Uddin LQ, Yeo BTT, Spreng RN. Towards a universal taxonomy of macro-scale functional human brain networks. Brain Topogr. 2019;32(6):926–942. doi:10.1007/s10548-019-00744-6
50. Alves PN, Foulon C, Karolis V, et al. An improved neuroanatomical model of the default-mode network reconciles previous neuroimaging and neuropathological findings. Commun Biol. 2019;2(1). doi:10.1038/s42003-019-0611-3
51. Loeber S, Vollstädt-Klein S, von der Goltz C, Flor H, Mann K, Kiefer F. Attentional bias in alcohol-dependent patients: the role of chronicity and executive functioning. Addict Biol. 2009;14(2):194–203. doi:10.1111/j.1369-1600.2009.00146.x
52. Clapp WC, Rubens MT, Gazzaley A. Mechanisms of working memory disruption by external interference. Cereb Cortex. 2010;20(4):859–872. doi:10.1093/cercor/bhp150
53. Jha AP, Fabian SA, Aguirre GK. The role of prefrontal cortex in resolving distractor interference. Cogn Affect Behav Neurosci. 2004;4(4):517–527. doi:10.3758/CABN.4.4.517
54. Goldstein RZ, Volkow ND. Dysfunction of the prefrontal cortex in addiction: neuroimaging findings and clinical implications. Nat Rev Neurosci. 2011;12(11):652–669.
55. Alba-Ferrara L, Müller-Oehring EM, Sullivan EV, Pfefferbaum A, Schulte T. Brain responses to emotional salience and reward in alcohol use disorder. Brain Imaging Behav. 2016;10(1):136–146. doi:10.1007/s11682-015-9374-8
56. George MS, Anton RF, Bloomer C, et al. Activation of prefrontal cortex and anterior thalamus in alcoholic subjects on exposure to alcohol-specific cues. Arch Gen Psychiatry. 2001;58(4):345–352. doi:10.1001/archpsyc.58.4.345
57. Heinz A, Wrase J, Kahnt T, et al. Brain activation elicited by affectively positive stimuli is associated with a lower risk of relapse in detoxified alcoholic subjects. Alcohol Clin Exp Res. 2007;31(7):1138–1147. doi:10.1111/j.1530-0277.2007.00406.x
58. Ray S, Hanson C, Hanson SJ, Bates ME. fMRI BOLD response in high-risk college students (Part 1): during exposure to alcohol, marijuana, polydrug and emotional picture cues. Alcohol Alcohol. 2010;45(5):437–443. doi:10.1093/alcalc/agq042
59. Sawyer KS, Adra N, Salz DM, et al. Hippocampal subfield volumes in abstinent men and women with a history of alcohol use disorder. PLoS One. 2020;15(8):e0236641. doi:10.1371/journal.pone.0236641
60. Myrick H, Anton RF, Li X, et al. Differential brain activity in alcoholics and social drinkers to alcohol cues: relationship to craving. Neuropsychopharmacology. 2004;29(2):393–402. doi:10.1038/sj.npp.1300295
61. Wrase J, Grüsser SM, Klein S, et al. Development of alcohol-associated cues and cue-induced brain activation in alcoholics. Eur Psychiatry. 2002;17(5):287–291. doi:10.1016/S0924-9338(02)00676-4
62. Tapert SF, Cheung EH, Brown GG, et al. Neural response to alcohol stimuli in adolescents with alcohol use disorder. Arch Gen Psychiatry. 2003;60(7):727–735. doi:10.1001/archpsyc.60.7.727
63. Ray S, Hanson C, Hanson SJ, Rahman RM, Bates ME. fMRI BOLD response of high-risk college students (Part 2): during memory priming of alcohol, marijuana and polydrug picture cues. Alcohol Alcohol. 2010;45(5):444–448. doi:10.1093/alcalc/agq043
64. Makris N, Oscar-Berman M, Jaffin SK, et al. Decreased volume of the brain reward system in alcoholism. Biol Psychiatry. 2008;64(3):192–202. doi:10.1016/j.biopsych.2008.01.018
65. Ceceli AO, Bradberry CW, Goldstein RZ. The neurobiology of drug addiction: cross-species insights into the dysfunction and recovery of the prefrontal cortex. Neuropsychopharmacology. 2022;47(1):276–291. doi:10.1038/s41386-021-01153-9
66. Fascher M, Nowaczynski S, Spindler C, Strobach T, Muehlhan M. Neural underpinnings of response inhibition in substance use disorders: weak meta-analytic evidence for a widely used construct. Psychopharmacology. 2024;241(1):1–17. doi:10.1007/s00213-023-06498-1
67. Zilverstand A, Huang AS, Alia-Klein N, Goldstein RZ. Neuroimaging impaired response inhibition and salience attribution in human drug addiction: a systematic review. Neuron. 2018;98(5):886–903. doi:10.1016/j.neuron.2018.03.048
68. Pfisterer M, Zimmermann S, Zaiser J, et al. Biomarkers in alcohol use disorder - The promise and pitfalls of neuroimaging drug cue reactivity. Biomark Neuropsych. 2025;12(100119):100119. doi:10.1016/j.bionps.2025.100119
69. Nguyen-Louie TT, Buckman JF, Ray S, Bates ME. Drinkers’ memory bias for alcohol picture cues in explicit and implicit memory tasks. Drug Alcohol Depend. 2016;160:90–96.
70. Briggs GG, Nebes RD. Patterns of hand preference in a student population. Cortex. 1975;11(3):230–238. doi:10.1016/S0010-9452(75)80005-0
71. Hamilton M. A rating scale for depression. J Neurol Neurosurg Psychiatry. 1960;23:56–62. doi:10.1136/jnnp.23.1.56
72. Robins LN, Cottler LB, Bucholz KK, Compton WM, North CS, Rourke K. Computerized Diagnostic Interview Schedule for the DSM-IV (C DIS-IV). NIMH/University of Florida; 2000.
73. Wechsler D. WAIS-III, Wechsler Adult Intelligence Scale, Third Edition: WMS-III, Wechsler Memory Scale, Third Edition: Technical Manual. Psychological Corporation; 1997:426.
74. Holdnack JA, Drozdick LW. CHAPTER 9 - Using WAIS-IV with WMS-IV. In: Weiss LG, Saklofske DH, Coalson DL, Raiford SE, editors. WAIS-IV Clinical Use and Interpretation. Academic Press; 2010:237–283.
75. Schneider F, Habel U, Wagner M, et al. Subcortical correlates of craving in recently abstinent alcoholic patients. Am J Psychiatry. 2001;158(7):1075–1083. doi:10.1176/appi.ajp.158.7.1075
76. Lubman DI. Addiction neuroscience and its relevance to clinical practice. Drug Alcohol Rev. 2007;26(1):1–2. doi:10.1080/09595230601036929
77. Flannery BA, Volpicelli JR, Pettinati HM. Psychometric properties of the penn alcohol craving scale. Alcohol Clin Exp Res. 1999;23(8):1289–1295. doi:10.1111/j.1530-0277.1999.tb04349.x
78. Cahalan D, Cisin IH, Crossley HM. American drinking practices: a national study of drinking behavior and attitudes. Monographs Rutgers Center Alcohol Studies. 1969;6:260.
79. Oscar-Berman M, Maleki N. Alcohol dementia, wernicke’s encephalopathy, and korsakoff’s syndrome. In: Alosco ML, Stern RA, editors. The Oxford Handbook of Adult Cognitive Disorders. Oxford University Press; 2019:743–758.
80. Stritzke WGK, Breiner MJ, Curtin JJ, Lang AR. Assessment of substance cue reactivity: advances in reliability, specificity, and validity. Psychol Addict Behav. 2004;18(2):148–159. doi:10.1037/0893-164X.18.2.148
81. Grüsser SM, Heinz A, Flor H. Standardized stimuli to assess drug craving and drug memory in addicts. J Neural Transm. 2000;107(6):715–720. doi:10.1007/s007020070072
82. Dale AM. Optimal experimental design for event-related fMRI. Hum Brain Mapp. 1999;8(2–3):109–114. doi:10.1002/(SICI)1097-0193(1999)8:2/3<109::AID-HBM7>3.0.CO;2-W
83. Thesen S, Heid O, Mueller E, Schad LR. Prospective acquisition correction for head motion with image-based tracking for real-time fMRI. Magn Reson Med. 2000;44(3):457–465.
84. Dale AM, Fischl B, Sereno MI. Cortical surface-based analysis. I. Segmentation and surface reconstruction. Neuroimage. 1999;9(2):179–194. doi:10.1006/nimg.1998.0395
85. Sled JG, Zijdenbos AP, Evans AC. A nonparametric method for automatic correction of intensity nonuniformity in MRI data. IEEE Transact Med Imag. 1998;17(1):87–97. doi:10.1109/42.668698
86. Talairach J, Tournoux P. Co-Planar Stereotaxic Atlas of the Human Brain: 3-Dimensional Proportional System: An Approach to Cerebral Imaging. G. Thieme; 1988.
87. Ségonne F, Dale AM, Busa E, et al. A hybrid approach to the skull stripping problem in MRI. NeuroImage. 2004;22(3):1060–1075. doi:10.1016/j.neuroimage.2004.03.032
88. Fischl B, Salat DH, Busa E, et al. Whole brain segmentation: automated labeling of neuroanatomical structures in the human brain. Neuron. 2002;33(3):341–355.
89. Fischl B, Sereno MI, Tootell RBH, Dale AM. High-resolution intersubject averaging and a coordinate system for the cortical surface. Human Brain Mapp. 1999;8(4):272–284. doi:10.1002/(SICI)1097-0193(1999)8:4<272::AID-HBM10>3.0.CO;2-4
90. Destrieux C, Fischl B, Dale A, Halgren E. Automatic parcellation of human cortical gyri and sulci using standard anatomical nomenclature. Neuroimage. 2010;53(1):1–15. doi:10.1016/j.neuroimage.2010.06.010
91. Desikan RS, Ségonne F, Fischl B, et al. An automated labeling system for subdividing the human cerebral cortex on MRI scans into gyral based regions of interest. Neuroimage. 2006;31(3):968–980.
92. Greve DN, Fischl B. Accurate and robust brain image alignment using boundary-based registration. Neuroimage. 2009;48(1):63–72. doi:10.1016/j.neuroimage.2009.06.060
93. LeDoux JE. The Emotional Brain: The Mysterious Underpinnings of Emotional Life. Simon & Schuster; 1996.
94. Dolcos F, LaBar KS, Cabeza R. Remembering one year later: role of the amygdala and the medial temporal lobe memory system in retrieving emotional memories. Proc Natl Acad Sci U S A. 2005;102(7):2626–2631. doi:10.1073/pnas.0409848102
95. Yeo BTT, Krienen FM, Eickhoff SB, et al. Functional specialization and flexibility in human association cortex. Cereb Cortex. 2016;26(1):465. doi:10.1093/cercor/bhv260
96. Heilbronner SR, Hayden BY. Dorsal anterior cingulate cortex: a bottom-up view. Annu Rev Neurosci. 2016;39:149–170. doi:10.1146/annurev-neuro-070815-013952
97. Ruiz SM, Oscar-Berman M, Sawyer KS, Valmas MM, Urban T, Harris GJ. Drinking history associations with regional white matter volumes in alcoholic men and women. Alcohol Clin Exp Res. 2013;37(1):110–122. doi:10.1111/j.1530-0277.2012.01862.x
98. Mann K, Ackermann K, Croissant B, Mundle G, Nakovics H, Diehl A. Neuroimaging of gender differences in alcohol dependence: are women more vulnerable? Alcohol Clin Exp Res. 2005;29(5):896–901. doi:10.1097/01.ALC.0000164376.69978.6B
99. Brighton R, Moxham L, Traynor V. Women and alcohol use disorders: factors that lead to harm. J Addict Nurs. 2016;27(3):205–213. doi:10.1097/JAN.0000000000000136
100. Merrill JE, Read JP. Motivational pathways to unique types of alcohol consequences. Psychol Addict Behav. 2010;24(4):705–711. doi:10.1037/a0020135
101. Tschuemperlin RM, Batschelet HM, Moggi F, et al. The neurophysiology of implicit alcohol associations in recently abstinent patients with alcohol use disorder: an event-related potential study considering gender effects. Alcohol Clin Exp Res. 2020;44(10):2031–2044. doi:10.1111/acer.14444
102. Rubio G, Martínez-Gras I, Ponce G, Quinto R, Jurado R, Jiménez-Arriero MÁ. Integration of self-guidance groups for relatives in a public program of alcoholism treatment. Adicciones. 2013;25(1):37–44. doi:10.20882/adicciones.70
103. Seo D, Jia Z, Lacadie CM, Tsou KA, Bergquist K, Sinha R. Sex differences in neural responses to stress and alcohol context cues. Human Brain Mapp. 2011;32(11):1998–2013. doi:10.1002/hbm.21165
104. Campanella S, Schroder E, Kajosch H, et al. Neurophysiological markers of cue reactivity and inhibition subtend a three-month period of complete alcohol abstinence. Clin Neurophysiol. 2020;131(2):555–565. doi:10.1016/j.clinph.2019.10.020
105. Grieder M, Soravia LM, Tschuemperlin RM, et al. Right inferior frontal activation during alcohol-specific inhibition increases with craving and predicts drinking outcome in alcohol use disorder. Front Psychiatry. 2022;13:909992. doi:10.3389/fpsyt.2022.909992
106. Boness CL, Witkiewitz K. Precision medicine in alcohol use disorder: mapping etiologic and maintenance mechanisms to mechanisms of behavior change to improve patient outcomes. Exp Clin Psychopharmacol. 2023;31(4):769–779. doi:10.1037/pha0000613
107. Ceceli AO, Parvaz MA, King S, et al. Altered prefrontal signaling during inhibitory control in a salient drug context in cocaine use disorder. Cereb Cortex. 2023;33(3):597–611. doi:10.1093/cercor/bhac087
108. Broyd SJ, Demanuele C, Debener S, Helps SK, James CJ, Sonuga-Barke EJS. Default-mode brain dysfunction in mental disorders: a systematic review. Neurosci Biobehav Rev. 2009;33(3):279–296. doi:10.1016/j.neubiorev.2008.09.002
109. Zhou HX, Chen X, Shen YQ, et al. Rumination and the default mode network: meta-analysis of brain imaging studies and implications for depression. Neuroimage. 2020;206:116287. doi:10.1016/j.neuroimage.2019.116287
110. Bednarski SR, Zhang S, Hong KI, Sinha R, Rounsaville BJ, Li CSR. Deficits in default mode network activity preceding error in cocaine dependent individuals. Drug Alcohol Depend. 2011;119(3):e51–e57. doi:10.1016/j.drugalcdep.2011.05.026
111. Chanraud S, Pitel AL, Pfefferbaum A, Sullivan EV. Disruption of functional connectivity of the default-mode network in alcoholism. Cereb Cortex. 2011;21(10):2272–2281. doi:10.1093/cercor/bhq297
112. Maleki N, Sawyer KS, Levy S, Harris GJ, Oscar-Berman M. Intrinsic brain functional connectivity patterns in alcohol use disorder. Brain Commun. 2022;4(6):fcac290. doi:10.1093/braincomms/fcac290
113. Sormaz M, Murphy C, Wang HT, et al. Default mode network can support the level of detail in experience during active task states. Proc Natl Acad Sci U S A. 2018;115(37):9318–9323. doi:10.1073/pnas.1721259115
114. Koshino H, Minamoto T, Ikeda T, Osaka M, Otsuka Y, Osaka N. Anterior medial prefrontal cortex exhibits activation during task preparation but deactivation during task execution. PLoS One. 2011;6(8):e22909. doi:10.1371/journal.pone.0022909
115. Leech R, Kamourieh S, Beckmann CF, Sharp DJ. Fractionating the default mode network: distinct contributions of the ventral and dorsal posterior cingulate cortex to cognitive control. J Neurosci. 2011;31(9):3217–3224. doi:10.1523/JNEUROSCI.5626-10.2011
116. Shields CN, Gremel CM. Review of orbitofrontal cortex in alcohol dependence: a disrupted cognitive map? Alcohol Clin Exp Res. 2020;44(10):1952–1964. doi:10.1111/acer.14441
117. Myrick H, Anton RF, Li X, Henderson S, Randall PK, Voronin K. Effect of naltrexone and ondansetron on alcohol cue-induced activation of the ventral striatum in alcohol-dependent people. Arch Gen Psychiatry. 2008;65(4):466–475. doi:10.1001/archpsyc.65.4.466
118. Koob GF, Volkow ND. Neurocircuitry of addiction. Neuropsychopharmacology. 2010;35(1):217–238. doi:10.1038/npp.2009.110
119. Reinhard I, Leménager T, Fauth-Bühler M, et al. A comparison of region-of-interest measures for extracting whole brain data using survival analysis in alcoholism as an example. J Neurosci Methods. 2015;242:58–64. doi:10.1016/j.jneumeth.2015.01.001
120. Garber AK, Lustig RH. Is fast food addictive? Curr Drug Abuse Rev. 2011;4(3):146–162. doi:10.2174/1874473711104030146
121. Field M, Munafò MR, Franken IHA. A meta-analytic investigation of the relationship between attentional bias and subjective craving in substance abuse. Psychol Bull. 2009;135(4):589–607. doi:10.1037/a0015843
122. Kunz S, Beblo T, Driessen M, Woermann F. fMRI of alcohol craving after individual cues: a follow-up case report. Neurocase. 2008;14(4):343–346. doi:10.1080/13554790802366020
123. Volkow ND, Wang GJ, Tomasi D, Baler RD. The addictive dimensionality of obesity. Biol. Psychiatry. 2013;73(9):811–818. doi:10.1016/j.biopsych.2012.12.020
124. Avena NM, Rada P, Hoebel BG. Evidence for sugar addiction: behavioral and neurochemical effects of intermittent, excessive sugar intake. Neurosci Biobehav Rev. 2008;32(1):20–39. doi:10.1016/j.neubiorev.2007.04.019
125. World Health Organization. Global Status Report on Alcohol and Health 2018. World Health Organization; 2019.
126. Eklund A, Nichols TE, Knutsson H. Cluster failure: why fMRI inferences for spatial extent have inflated false-positive rates. Proc Natl Acad Sci U S A. 2016;113(28):7900–7905. doi:10.1073/pnas.1602413113
127. Greve DN, Fischl B. False positive rates in surface-based anatomical analysis. Neuroimage. 2018;171:6–14. doi:10.1016/j.neuroimage.2017.12.072
128. Woo CW, Krishnan A, Wager TD. Cluster-extent based thresholding in fMRI analyses: pitfalls and recommendations. Neuroimage. 2014;91:412–419. doi:10.1016/j.neuroimage.2013.12.058
129. Luhar RB, Sawyer KS, Gravitz Z, Ruiz SM, Oscar-Berman M. Brain volumes and neuropsychological performance are related to current smoking and alcoholism history. Neuropsychiatr Dis Treat. 2013;9:1767–1784. doi:10.2147/NDT.S52298
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.