Back to Journals » International Journal of Nanomedicine » Volume 20
Advancing Epigenetic Combination Therapy in Oncology: Multifunctional Nano-Drug Delivery Systems for Synergistic Efficacy and Precision Modulation
Authors Gao T ![]() , Fu S, Quan X, Sun J, Jiang M, Li J
, Fu S, Quan X, Sun J, Jiang M, Li J
Received 8 September 2025
Accepted for publication 2 December 2025
Published 11 December 2025 Volume 2025:20 Pages 14853—14883
DOI https://doi.org/10.2147/IJN.S566173
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 5
Editor who approved publication: Professor Dong Wang
Tong Gao,1,* Shunli Fu,2,* Xianghua Quan,1 Jialin Sun,1 Man Jiang,1 Jing Li1
1Department of Pharmacy, the Affiliated Hospital of Qingdao University, Qingdao, 266000, People’s Republic of China; 2Tumor Immunology and Cytotherapy of Medical Research Center, Shandong Provincial Key Laboratory of Clinical Research for Pancreatic Diseases, the Affiliated Hospital of Qingdao University, Qingdao University, Qingdao, 266000, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Jing Li, Department of Pharmacy, The Affiliated Hospital of Qingdao University, Qingdao, 266000, People’s Republic of China, Email [email protected]
Abstract: Epigenetic modifications regulate gene expression at the transcriptional level, contributing to tumorigenesis and progression. While epigenetic-targeted combination therapies have gained prominence in oncology treatment management, their clinical efficacy remains constrained by differences in pharmacokinetics and biodistribution among combined agents. Nano-drug delivery systems (NDDS) demonstrate unique potential through co-delivery of therapeutic agents and optimization of their pharmacokinetic profiles. Furthermore, the development of multifunctional NDDS opens new possibilities for precision modulation in cancer treatment, offering valuable insights for clinical translation. Here, this review first outlined the intervention mechanisms of epigenetic dysregulation and analyzed the applications of epigenetic combination approaches. Subsequently, we highlight the transformative potential of NDDS in epigenetic combination therapy, with particular emphasis on how multifunctional NDDS design enables precise therapeutic regulation. This comprehensive analysis aims to advance the clinical translation of epigenetic-based combination strategies through innovative drug delivery solutions. In the future, with the continuous development of AI-driven NDDS design, biomimetic carriers, and dynamic epigenetic editing tools, it will be possible to overcome the clinical challenges of NDDS, enabling truly personalized cancer treatment.
Keywords: epigenetics, cancer combination therapy, nano-drug delivery systems, multifunctional drug delivery strategy
Introduction
Epigenetics refers to a dynamic and reversible regulatory mechanism that modulates gene expression through chemical modification or alteration of chromosome structure without altering the underlying DNA sequence.1,2 This sophisticated regulation system operates through coordinated cycles of covalent modifications to DNA and histones, primarily mediated by chromatin-modifying enzymes working in a tightly regulated and synergistic manner.3,4 These biochemical alterations ultimately lead to structural reorganization of chromatin, thereby influencing transcriptional accessibility.5 The epigenetic regulatory machinery comprises three functionally distinct enzyme categories: “writers”, “erasers” and “readers”.1,6,7 Among them, writers catalyze the addition of epigenetic marks, including DNA methyltransferases (DNMTs), histone lysine methyltransferases (KMTs) and histone acetyltransferases (HATs).8 Erasers mediate removal of post-translational modifications, including histone lysine demethylases (KDMs) and histone deacetylases (HDACs).9,10 Readers are specialized protein domains that recognize specific epigenetic signatures, including bromodomains and chromodomains.11,12 Emerging evidence has established the crucial role of epigenetic dysregulation in cancer pathogenesis, with particular emphasis on the aberrant expression and functional alterations of epigenetic enzymes.13,14 This mechanistic understanding has spurred significant therapeutic interest, positioning epigenetic modulators as promising targets for novel anticancer strategies.15,16
Several epigenetic-targeted agents have received FDA approval for hematologic malignancies, demonstrating the clinical potential of epigenetic modulation in cancer treatment.14,16,17 However, treatment resistance remains a significant therapeutic challenge, with some patients showing limited responses to monotherapy.18,19 To address this limitation, combinatorial approaches integrating epigenetic drugs with conventional therapies (chemotherapy, radiotherapy) and emerging modalities (targeted therapy, immunotherapy) are being actively investigated.20–22 Despite this promise, the clinical translation of epigenetic combination therapies faces several persistent hurdles. The epigenetic-targeted agents often act broadly on global gene expression patterns and therapeutically promising, also raise concerns regarding off-target effects and systemic toxicity. Specifically, these include the poor bioavailability and rapid clearance of some epigenetic drugs, dose-limiting off-target toxicities due to non-specific biodistribution, and the eventual emergence of multi-drug resistance. While previous reviews have extensively cataloged various combinatorial strategies, they often overlook the central pharmacological barrier: the divergent pharmacokinetic profiles between epigenetic drugs and their partner agents, suboptimal tumor accumulation, dose-limiting systemic toxicity from non-specific biodistribution, which prevents truly synergistic efficacy. Nano-drug delivery systems (NDDS) offer a promising platform to overcome these limitations through co-encapsulation of multiple therapeutics with tunable ratios, enhanced physicochemical stability and prolonged circulation, tumor-selective accumulation and controlled release.23 Furthermore, the complexity of epigenetic regulation extends beyond nuclear modifications, encompassing organelle-specific enzyme localization (eg, mitochondrial DNMTs, lysosomal HDACs) and microenvironmental reprogramming of tumor-associated immune/stromal cells.24–26 This multilayered regulatory network requires the development of intelligent NDDS with multi-functionality, including spatiotemporal coordination, biological navigation, and subcellular precision, etc. The development of NDDS for precision epigenetic engineering has emerged as a transformative frontier in oncology therapeutics.
Based on this, this review provides a critical analysis of epigenetic-based combination therapies in oncology. We first outline the role of epigenetic dysregulation in cancer pathogenesis and analyze the therapeutic applications of epigenetic combination approaches. Subsequently, we highlight the transformative potential of NDDS in epigenetic combination therapy, with particular emphasis on how multifunctional NDDS design enables precise therapeutic regulation. This comprehensive analysis aims to advance the clinical translation of epigenetic-based combination strategies through innovative drug delivery solutions.
Epigenetic and Drugs in Clinical
Epigenetics is a widely occurring phenomenon in tumors, and the study of epigenetics is of great significance for the individualized treatment of tumors. The main epigenetic mechanisms in tumors involve DNA methylation, histone modification, non-coding RNA regulation, and chromatin remodeling. Targeting these regulatory processes provides a promising approach for developing novel cancer therapies. This section discusses how epigenetic alterations contribute to tumor progression and reviews currently available epigenetic-based drugs used in clinical practice (Figure 1).
 |
Figure 1 Different epigenetic modifications regulating gene expression and representative epigenetic drugs in clinical. |
Epigenetic in Cancer Development
DNA Methylation
DNA methylation is one of the most central epigenetic modifications that regulates gene expression through structural alterations in chromatin, DNA conformation stability, and protein–DNA interactions while preserving genomic integrity. This heritable chemical modification modulates critical biological processes including cell cycle regulation, apoptosis, and embryonic development without altering nucleotide sequences.27–29 Catalyzed by DNA methyltransferases (DNMTs) utilizing S-adenosylmethionine as methyl donor, the process converts cytosine to 5-methylcytosine at CpG islands, particularly in promoter regions where it induces chromatin compaction and transcriptional silencing.30,31 The mammalian DNMT family comprises five isoforms with distinct functions: DNMT1 maintains methylation patterns during DNA replication, DNMT3a/b establish de novo methylation critical for embryogenesis and oncogenesis, DNMT3L facilitates structural coordination of DNMT3 complexes, while DNMT2 primarily methylates tRNA.32–34 Tumorigenesis involves characteristic methylation aberrations featuring genome-wide hypomethylation that destabilizes chromatin and activates proto-oncogenes, coupled with localized CpG island hypermethylation that silences tumor suppressor genes. These dual epigenetic perturbations synergistically drive malignant transformation through disrupted genomic surveillance and dysregulated oncogenic pathways.35,36
Histone Modifications
Histone modifications represent a fundamental epigenetic mechanism through which dynamic post-translational alterations regulate chromatin architecture and gene expression in eukaryotes. Comprising five highly conserved core proteins (H1, H2A, H2B, H3, H4), histones assemble with DNA into nucleosomes, with their N-terminal tails undergoing reversible covalent modifications including methylation, acetylation, phosphorylation, and ubiquitination.37,38 These chemical alterations modulate chromatin compaction by altering electrostatic interactions between positively charged histones and negatively charged DNA, thereby controlling transcriptional accessibility.39,40 The tightly regulated balance of histone methyltransferases (HMTs) and demethylases (HDMs) governs lysine/arginine methylation patterns that maintain heterochromatin integrity, mediate genomic imprinting, and influence DNA repair processes, with dysregulation implicated in oncogenesis through aberrant silencing of tumor suppressors.41,42 Similarly, histone acetylation dynamics controlled by histone acetyltransferases (HATs) and deacetylases (HDACs) regulate chromatin relaxation through neutralization of histone charges, facilitating transcription factor binding.43,44 Notably, these enzymatic regulators exhibit broad substrate specificity, modifying non-histone targets including p53, Myc, and β-catenin to influence protein stability and oncogenic signaling. Bromodomain-containing proteins (BRD2/3/4) further propagate acetylation signals by recognizing acetylated lysines, with their frequent dysregulation in cancers underscoring the therapeutic potential of targeting this chromatin recognition system.45,46 The tumor-specific redistribution of histone modification patterns, particularly H3/H4 hyperacetylation and site-specific methylation aberrations, establishes these epigenetic marks as both diagnostic biomarkers and actionable therapeutic targets in cancer biology.47,48
Non-Coding RNA Modification
Non-coding RNA (ncRNA) modifications constitute an emerging epigenetic regulatory layer where chemically altered RNA molecules, though non-protein-coding, exert profound control over gene expression by modulating chromatin architecture and DNA topology through interactions with nucleic acids and proteins.49,50 Major ncRNA subtypes including miRNAs, lncRNAs, and circRNAs undergo dynamic post-transcriptional modifications such as methylation, acetylation, and ribosylation, with N6-methyladenosine (m6A) representing the most abundant and well-characterized modification involving methyl group addition to adenine residues.51,52 This reversible process is orchestrated by coordinated actions of writer complexes (m6A methyltransferases like METTL3/METTL14), erasers (demethylases such as FTO/ALKBH5), and reader proteins that decode methylation signals.53,54 Spatial enrichment of m6A near stop codons and within long exons enables its multifaceted regulation of tumor biology, governing cancer stem cell plasticity, therapeutic resistance mechanisms, and tumor microenvironment interactions.55,56 The epitranscriptomic machinery’s capacity to influence oncogenic pathways through both direct RNA processing and indirect modulation of tumor metabolism/immune evasion highlights its diagnostic and therapeutic potential in oncology, with dysregulated m6A dynamics now recognized as critical drivers of malignant transformation and cancer progression.57,58
Chromatin Remodeling
Chromatin remodeling serves as a dynamic epigenetic regulatory mechanism that orchestrates gene expression by modulating nucleosome architecture through two principal pathways: histone-tail covalent modifications mediated by protein complexes like Polycomb group (PcG) proteins, and ATP-dependent structural reorganization driven by chromatin remodeling complexes that utilize hydrolysis energy to reposition nucleosomes.14,59 Operating on the fundamental nucleosome unit composed of DNA wrapped around histone octamers, this process enables cells to adapt gene activity to environmental stimuli through three-dimensional chromatin reorganization mechanisms – nucleosome repositioning, loop formation via chromatin cyclases, and chromatin decompaction through depolymerase-mediated relaxation.60,61 These structural transformations regulate transcriptional accessibility either by altering histone-DNA binding interfaces through post-translational modifications or by physically restructuring nucleosome arrays to expose regulatory DNA elements.62,63 The resultant chromatin state transitions, ranging from tightly packed heterochromatin to transcriptionally permissive euchromatin, critically influence developmental processes and genome stability while serving as pivotal drivers of oncogenesis when dysregulated.64,65 Aberrant chromatin remodeling in cancer manifests through pathological gene silencing (tumor suppressor inactivation) or illegitimate oncogene activation, often mediated through malfunctioning chromatin-modifying enzymes that disrupt the delicate balance between chromatin condensation and decompaction states essential for proper transcriptional control.66,67
Epigenetic Drugs in Clinical
DNA Methyltransferase Inhibitors (DNMT Inhibitors)
DNMT inhibitors exert therapeutic effects by inhibiting DNMT activity, thereby blocking the transfer of methyl groups from S-adenosylmethionine to cytosine residues and suppressing abnormal DNA methylation, which reactivates silenced tumor suppressor genes for cancer treatment. Unlike conventional cytotoxic chemotherapies, these inhibitors primarily induce epigenetic reprogramming rather than immediate cell death. Clinically approved nucleoside analogs including Azacitidine and Decitabine incorporate into DNA during replication, where they are recognized by DNMTs and subsequently inactivate the enzymes through covalent binding to cysteine sulfhydryl groups in their catalytic sites. Both agents are currently approved for myelodysplastic syndrome treatment, with several other DNMT inhibitors undergoing clinical evaluation.68–70
Isocitrate Dehydrogenase Inhibitors (IDH Inhibitors)
IDH inhibitors target tumor-associated IDH mutations that disrupt normal enzymatic activity, converting α-ketoglutarate (α-KG) into the oncogenic metabolite 2-hydroxyglutarate (2HG), which accumulates in mutated cells and induces epigenetic dysregulation through DNA/histone hypermethylation. Clinically approved agents including Enasidenib and Ivosidenib suppress 2HG production by competitively inhibiting mutant IDH enzymes, thereby restoring normal methylation patterns and impeding tumor progression. Enasidenib specifically treats relapsed/refractory acute myeloid leukemia (AML) with IDH2 mutations, while Ivosidenib is indicated for IDH1-mutated relapsed/refractory myelodysplastic syndromes.71–74
Histone Methyltransferase Inhibitors (HMT Inhibitors)
HMT inhibitors have emerged as targeted therapies with clinical progress in recent years, particularly against key enzymes like lysine methyltransferase EZH2, DOT1L, and arginine methyltransferases (PRMTs). EZH2 hyperactivation, implicated in tumorigenesis and stem-like transcriptional reprogramming across malignancies including breast cancer, castration-resistant prostate cancer, small cell lung cancer, and neuroblastoma, represents the most clinically advanced target, exemplified by the FDA-approved EZH2 inhibitor Tazemetostat for epithelioid sarcoma and relapsed/refractory follicular lymphoma. Concurrently, the DOT1L inhibitor pinometostat (EPZ-5676) has completed Phase 1 evaluation in leukemia patients, demonstrating the expanding therapeutic landscape of HMT inhibition in oncology.75–78
Lysine-Specific Demethylase Inhibitors (LSD Inhibitors)
LSD inhibitors target LSD1, a flavin adenine dinucleotide-dependent enzyme whose functional outcomes vary depending on associated cofactors and substrates. This demethylase demonstrates critical regulatory roles in hematological malignancies like acute myeloid leukemia (AML), while its dysregulation in solid tumors correlates with histological grading, malignant progression, and clinical prognosis. Although no LSD1-targeting drugs have received global approval, multiple candidates including TCP, ORY1001, IMG-7289, SP2577, and CC-90011 have entered clinical evaluation, predominantly in early-phase trials focusing on small cell lung cancer (SCLC) and AML indications.79–81
Bromodomain and Extra-Terminal Inhibitors (BET Inhibitors)
BET inhibitors target the BET protein family comprising four members (BRD2, BRD3, BRD4, BRDT) that are frequently overexpressed in malignancies and regulate oncogenic transcriptional programs. As an emerging class of anticancer agents including JQ1, OTX-015, and OPN-0610, these inhibitors exert therapeutic effects by disrupting BET protein–chromatin interactions to suppress oncogene transcription. Their potent antitumor activity has been demonstrated across multiple cancer types, with particular efficacy observed in hematological malignancies such as leukemia and lymphoma.82–85
Histone Deacetylase Inhibitors (HDAC Inhibitors)
HDAC inhibitors target 18 human HDAC isoforms categorized into four classes (I, IIa/IIb, III, IV) based on structural features and catalytic mechanisms. Representing the most extensively developed epigenetic therapeutics, these agents are structurally classified by their Zn2⁺-binding groups: hydroxamic acids (vorinostat, panobinostat), cyclic tetrapeptides (romidepsin), investigational short-chain fatty acids, and benzamides (belinostat, chidamide). The first three categories broadly inhibit class I/II HDACs as pan-inhibitors, while benzamide derivatives exhibit selective suppression of class I and specific class IIb isoforms without affecting class IIa HDAC activity.86–89
Epigenetic-Based Cancer Combination Therapy
The integration of epigenetic drugs with other treatments, including standard chemotherapy, targeted therapy, and immunotherapy, has evolved into an attractive alternative for oncology treatment. A rational combination therapy regimen can overcome the limitations of single-agent epigenetic therapy, thereby enhancing antitumor effects and reducing drug resistance. This section presents an overview of clinically investigated epigenetic combination regimens, evaluating their pharmacological rationale and therapeutic outcomes in contemporary oncology practice.
Combined with Epigenetic Therapy
The complexity of epigenetic regulation in tumorigenesis has driven the development of combination therapies targeting key enzymes such as DNA methyltransferases (DNMT) and histone deacetylases (HDAC), with preclinical evidence demonstrating that concurrent DNMT-HDAC inhibition suppresses MYC proto-oncogene expression, counteracts immune evasion, and enhances cytotoxic T-cell recruitment to exert antitumor immunity.90–93 Early clinical validation emerged from a Phase I/II trial evaluating azacytidine (DNMT inhibitor) and entinostat (HDAC inhibitor) in non-small cell lung cancer, where all 12 enrolled patients exhibited clinical benefit: one achieved 14-month complete response, one 8-month partial response, two sustained 18- and 14-month disease stabilization, and eight maintained stability ≥12 weeks.94 Biomarker analysis revealed circulating tumor DNA methylation status as predictive of outcomes, with methylated-profile patients showing superior response rates and survival compared to unmethylated counterparts. Concurrently, dual-action HDAC/DNMT inhibitors under development demonstrate multifactorial antitumor effects through tumor suppressor reactivation coupled with suppression of proliferation, metastasis, and survival pathways in malignant cells.91,95 However, the clinical implementation of such combinations faces significant challenges. The non-specific action of epigenetic drugs may cause widespread transcriptomic alterations, leading to unpredictable off-target effects. Furthermore, the dynamic and reversible nature of epigenetic states necessitates precise timing and duration of drug administration, which is difficult to control precisely in clinical practice. Determining the optimal dosing combination is also particularly challenging, as the synergistic window between different epigenetic drugs is often narrow.
Combined with Chemotherapy
The synergistic potential of combining DNMT/HDAC inhibitors with chemotherapy is supported by preclinical and clinical evidence demonstrating that epigenetic priming enhances tumor cell chemosensitivity while reversing acquired resistance mechanisms.96–98 This strategy leverages epigenetic drugs to augment DNA-damaging chemotherapeutics, as exemplified in a Phase II trial where azacitidine and chidamide combined with GemOx achieved favorable tolerability and efficacy in relapsed/refractory peripheral T-cell lymphoma.99 The low-dose DNMT inhibitor decitabine has been demonstrated to resensitise chemotherapy resistant diffuse large B cell lymphoma (DLBCL) cells to doxorubicin without causing significant damage.100 Similar to DNMT inhibitors, HDAC inhibitors have demonstrated the ability to reverse chemoresistance by reprogramming malignant cells to regain sensitivity to cytotoxic agents. For instance, panobinostat attenuates hypoxia-induced cisplatin resistance in non-small cell lung cancer (NSCLC) cells via HIF-1α destabilization. Mechanistically, HIF-1α activation has been shown to confer multidrug resistance through transcriptional regulation of chemoprotective pathways.101,102 Despite this promise, combining epigenetic drugs with chemotherapy presents important limitations. Epigenetic priming may simultaneously enhance chemotherapy toxicity to normal tissues, resulting in a narrowed therapeutic window. The complex pharmacokinetic interactions between chemotherapeutic agents and epigenetic modulators also complicate dose optimization. Moreover, tumor heterogeneity may lead to inconsistent responses to epigenetic sensitization across different patient subsets, necessitating the development of reliable predictive biomarkers to guide patient selection.
Combined with Targeted Therapy
Epigenetic therapy demonstrates potential in reversing resistance mechanisms driven by genetic alterations and transcriptional reprogramming, particularly in models devoid of defined genetic drivers.103–105 HDAC inhibition has been reported to overcome resistance to various kinase inhibitors. For instance, MPT0E028, a relatively novel oral histone deacetylase inhibitor, was shown to enhance apoptosis induced by first-line epidermal growth factor receptor (EGFR) tyrosine kinase inhibitor (TKI) erlotinib in EGFR-TKI-resistant non-small cell lung cancer (NSCLC) cells.106 Another preliminary study demonstrated that the combination of EGFR-TKIs with vorinostat reversed acquired resistance to EGFR-TKIs and promoted apoptotic cell death in NSCLC models.107 In hematologic malignancies, azacytidine combined with the BCL-2 inhibitor venetoclax synergistically enhances apoptosis through MCL-1 suppression, overcoming therapeutic resistance.108,109 This azacytidine-venetoclax regimen has received FDA breakthrough therapy designation for untreated AML patients ineligible for intensive chemotherapy, with ongoing evaluations in MDS and AML clinical trials.110,111 Major obstacles to implementing epigenetic-targeted therapy combinations include managing the risk of overlapping toxicities and the heterogeneity of resistance mechanisms. Epigenetic reprogramming may trigger compensatory signaling pathway activation, undermining the efficacy of targeted therapies. Simultaneously, the impact of epigenetic drugs on the pharmacokinetics of targeted therapies is not fully understood, potentially altering drug exposure levels and effectiveness. The lack of reliable biomarkers to identify patient populations most likely to benefit from combination therapy also limits its clinical application.
Combined with Endocrine Therapy
Therapeutic targeting of lineage-specific transcriptional dependencies, exemplified by estrogen receptor (ER) signaling in breast cancer and androgen receptor (AR) pathways in prostate cancer, is profoundly influenced by epigenetic mechanisms that sustain oncogenic transcription in therapy-resistant malignancies.112–115 This interplay is demonstrated through KAT6A, a MYST-family histone acetyltransferase amplified in 10–15% of breast cancers where its genetic ablation reduces ERα expression, prompting clinical evaluation of the first-in-class inhibitor PF-07248144 as monotherapy and combined with fulvestrant or letrozole/palbociclib in advanced ER+ breast cancer.116–118 Parallel epigenetic vulnerabilities emerge in prostate cancer progression, where EZH2 overexpression correlates with castration resistance and neuroendocrine differentiation, while preclinical models reveal EZH2 inhibition not only suppresses malignant transformation but also enhances CD8+ T-cell cytotoxicity and IFN-γ production to restore androgen therapy sensitivity.119 Clinical trials are exploring EZH2 inhibitors combined with enzalutamide or androgen deprivation therapies, underscoring the strategic convergence of epigenetic modulation and hormonal targeting in overcoming transcriptional addiction across cancer subtypes.120,121 Challenges for such combination strategies lie in the complexity and plasticity of epigenetic mechanisms in endocrine therapy resistance. Epigenetic modulators may interfere with the normal physiological functions of hormone receptors, leading to endocrine imbalances. The potential impact of long-term epigenetic intervention on hormone-dependent tissues (such as bone and cardiovascular systems) remains unclear. Furthermore, the optimal timing and sequencing strategy for epigenetic–endocrine combination therapy requires further exploration to maximize clinical benefit.
Combined with Immunotherapy
Epigenetic modifications drive oncogenesis by dysregulating tumor suppressor pathways and oncogenic networks across diverse malignancies while modulating immune cell activation and effector functions, establishing bidirectional crosstalk between tumor epigenetics and immunology.122,123 DNMT inhibitors exert dual immunostimulatory effects through transposable element reactivation-induced double-stranded RNA production (TLR3-mediated type I interferon response) and enhanced MHC-I/antigen presentation machinery, thereby overcoming limitations of checkpoint inhibitors like PD-1/PD-L1 or CTLA-4 antibodies that frequently fail due to inadequate T-cell priming.122,124–126 Clinical validation emerges from trials in Hodgkin lymphoma where decitabine combined with pembrolizumab improved complete response rates (71% vs 32%) and progression-free survival (100% vs 76%) versus monotherapy.127 Parallel findings in SMARCB1-deficient tumors demonstrate tazemetostat-mediated EZH2 inhibition increases tumor-infiltrating CD8+ T cells/Tregs while upregulating PD-1/LAG-3 expression, illustrating how epigenetic modulation primes checkpoint-responsive tumor microenvironments.128 These mechanistic and clinical insights position epigenetic-checkpoint inhibitor combinations as transformative strategies for enhancing cancer immunotherapy efficacy. However, epigenetic-immunotherapy combinations face unique challenges. The immune activation by epigenetic drugs may trigger autoimmune-like toxicities, potentially synergizing with immune-related adverse events from immune checkpoint inhibitors. The impact of epigenetic reprogramming on T-cell function is context-dependent and may in some circumstances promote T-cell exhaustion. The dynamic nature of the immune microenvironment also demands precise timing and sequencing of epigenetic modulation relative to immunotherapy administration to maximize synergistic antitumor immunity.
Combined with Radiotherapy
Radiotherapy induces tumor cell DNA double-strand breaks via ionizing radiation, yet therapeutic efficacy is often compromised by aberrant DNA repair mechanisms underlying radiation resistance.129 Chromatin compaction states, predominantly regulated through epigenetic modifications, critically determine both radiation-induced DNA damage extent and repair capacity, establishing epigenetic targets as radiosensitization strategies.130,131 Notably, epigenetic regulators operate within interconnected networks coordinating radiation response, as exemplified by ionizing radiation-induced DNA-PK complex phosphorylation of fumarase at threonine 236, which mediates its interaction with histone variant H2A.X at damage sites. Local fumarate accumulation inhibits histone demethylase KDM2B, elevating H3K36me2 levels that promote DNA-PK complex recruitment to facilitate error-prone non-homologous end joining (NHEJ) repair and survival.132–134 This multilayered epigenetic crosstalk underscores the therapeutic rationale for combining epigenetic modulators with radiotherapy, enabling dose reduction to mitigate toxicity while overcoming acquired resistance through synergistic pathway targeting.135 Though epigenetic-radiotherapy combinations remain exploratory, advances in epigenomic profiling and real-time damage detection technologies are accelerating the development of precision combinatorial regimens. Key limitations to implementing epigenetic-radiotherapy combinations include the potential for increased normal tissue toxicity and the lack of radiosensitization biomarkers. Epigenetic drugs may enhance radiation damage to normal tissues, leading to exacerbated treatment-related toxicities. Significant heterogeneity exists in how different tumor types respond to epigenetic radiosensitization, requiring the development of predictive biomarkers to guide patient stratification. Furthermore, the optimal timing and administration schedule for combining epigenetic modulation with radiotherapy still need further optimization in clinical trials to balance efficacy with safety.
NDDS for Epigenetic-Based Cancer Combination Therapy
The integration of epigenetic therapeutics with conventional modalities (chemotherapy, radiotherapy, targeted therapy, immunotherapy) has demonstrated preclinical and clinical promise in overcoming therapeutic resistance, though conventional multidrug regimens face challenges in maintaining precise drug ratios due to divergent physicochemical properties, pharmacokinetic profiles, and biodistribution patterns that compromise tumor-specific delivery and exacerbate systemic toxicity. Emerging nanomedicine platforms—including liposomes, polymeric nanoparticles, micellar systems, dendrimers, and inorganic nanocarriers—address these limitations through coordinated co-delivery of therapeutic payloads, enhancing drug stability while optimizing spatiotemporal release kinetics to achieve synergistic dosing ratios at tumor sites. These nano-drug delivery systems (NDDS) exhibit transformative potential for epigenetic-based combination therapies by engineering precise pharmacological coordination that maximizes antitumor efficacy while minimizing off-target effects (Figure 2). The advantages and limitations of various NDDS were summarized in Table 1.
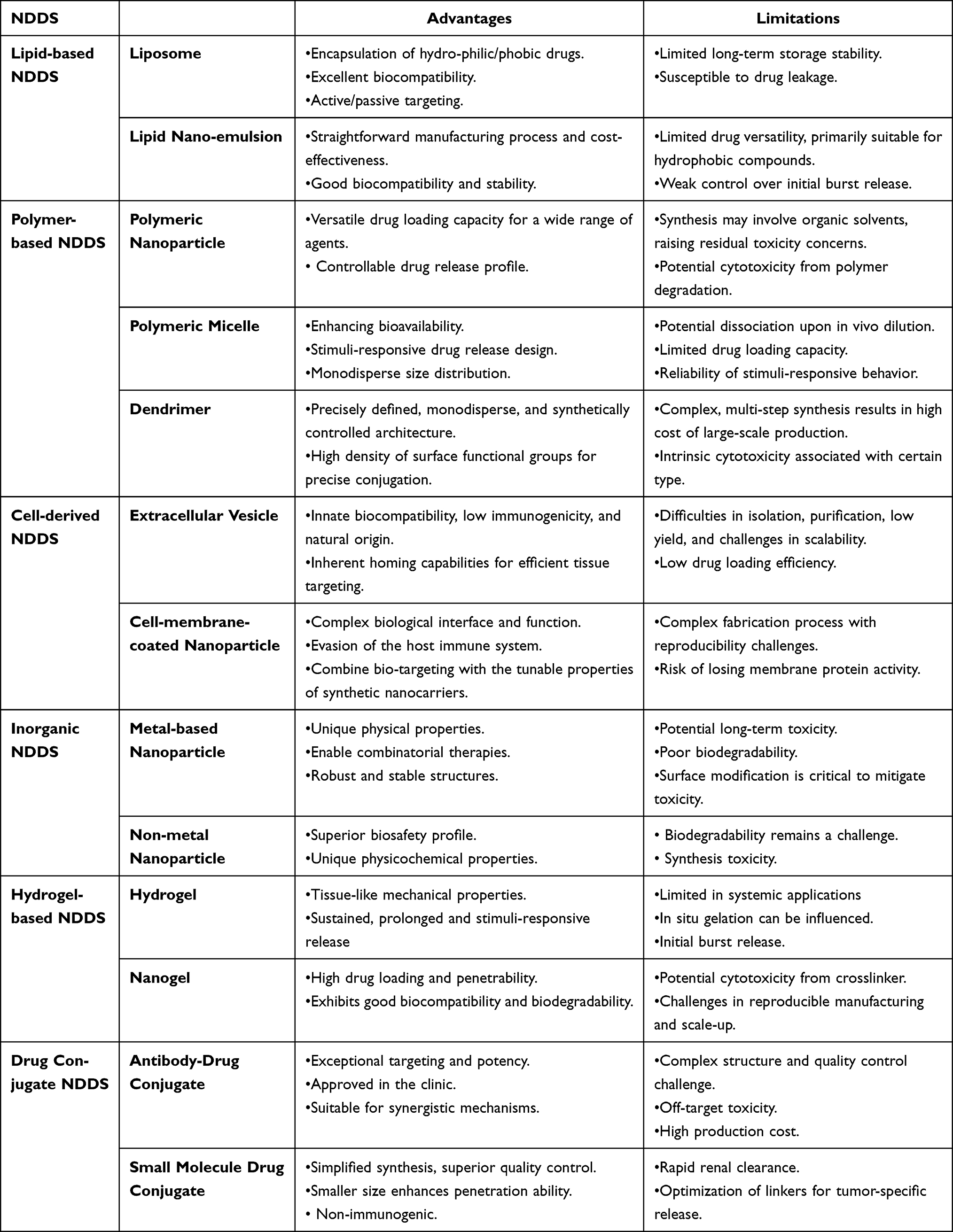 |
Table 1 Summary of Advantages and Limitations of Various Nanocarrier Systems for Drug Delivery |
 |
Figure 2 Summary of NDDS for epigenetic-based cancer combination therapy. |
Lipid-Based Nanoparticles
Liposomes
Liposomes, closed spherical vesicles formed by phospholipid-cholesterol bilayers, represent a versatile drug delivery platform capable of encapsulating both hydrophilic and lipophilic therapeutics to enhance solubility, bioavailability, and targeted accumulation while minimizing systemic exposure.136 Their structural versatility enables surface functionalization with polymers, targeting ligands, and other bioactive molecules, allowing precise modulation of biodistribution patterns and therapeutic payload release kinetics. By concentrating drug delivery to pathological sites through passive/active targeting mechanisms, these nanocarriers significantly improve therapeutic indices while reducing off-target toxicity, with engineered modifications further amplifying their pharmacological value across diverse therapeutic applications.137,138
Liposomes demonstrate versatile applications in epigenetic combination therapies through co-delivery of epigenetic modulators with chemotherapeutic agents, immunotherapeutics, and photosensitizers, effectively addressing drug resistance while enhancing tumor-specific targeting. Tang et al engineered liposomes co-encapsulating the DNMT inhibitor zebularine and gemcitabine, which reversed pancreatic cancer chemoresistance by upregulating drug-sensitivity markers, achieving 4-fold intracellular gemcitabine accumulation and 22-fold plasma half-life extension.139 Similarly, Lin et al’s liposomal co-delivery of panobinostat (HDAC inhibitor) and osimertinib suppressed glycolytic metabolism and epithelial–mesenchymal transition in NSCLC, overcoming therapeutic resistance. At the end of the experiment, the tumor inhibition rate was calculated. The results showed that the inhibition rate of the combined drug co-loaded in liposomes was 82%, while that of the combined drug free group was 51%, and the group using osimertinib alone had an inhibition rate of 45%.140 Immuno-epigenetic strategies include He et al’s dual-loaded liposomes with panobinostat and JQ1 (BET inhibitor) that reprogrammed tumor-associated macrophages and enhanced CD8+ T-cell function,141 while Tu et al’s chidamide/BMS-202 liposomes blocked PD-L1 and amplified immunogenicity to inhibit primary/metastatic growth and metastatic tumors with an inhibitory rate of metastasis of up to 96%.142 Liang et al further optimized this approach using cationic liposomes co-loaded with zebularine, JQ1, and CpG to elevate tumor antigen presentation and T-cell infiltration alongside PD-L1 downregulation.143 Multimodal systems exemplified by Sun et al’s thermosensitive liposomes incorporating copper sulfide, CpG, and JQ1 enabled laser-triggered immunogenic cell death and precision release of immunomodulators, stimulating dendritic cell maturation and cytotoxic lymphocyte recruitment for amplified antitumor immunity.144
Lipid nanoemulsions
Lipid nanoemulsions, nanoscale oil-in-water systems (20–200 nm diameter), serve as efficient carriers for hydrophobic therapeutics due to manufacturing simplicity, biocompatibility, and structural stability, with soybean oil-based formulations demonstrating enhanced tumor targeting via the enhanced permeability and retention effect.145,146 Their sub-200nm size facilitates enzymatic/hydrolytic degradation resistance, prolonging drug bioavailability while concentrating therapeutic payloads in malignant tissues.147 Functionalization with tumor-specific ligands further improves precision, as exemplified by Kim et al’s lysophosphatidic acid receptor 1 (LPAR1)-targeted nanoemulsions co-encapsulating decitabine and panobinostat, which reduced systemic toxicity while maintaining drug stability in triple-negative breast cancer models.148 Cellular uptake and biodistribution correlated with LPAR1 expression levels, enabling tumor-selective epigenetic reprogramming through decitabine-mediated DNA hypomethylation and panobinostat-induced chromatin remodeling, collectively reactivating tumor suppressor genes while suppressing oncogenic pathways. The results showed that the combined effect of the drug-loaded nanoemulsion significantly reduced the expression levels of FOXM1 mRNA and protein by up to 80%.
The review on the application of lipid-based nanoparticles for epigenetic-based cancer therapy was summarized as follows: Lipid-based nanocarriers have emerged as dominant platforms in combinatorial drug delivery, representing both the most clinically advanced and commercially viable nanomedicine systems to date. As the fastest-growing category of NDDS, their structural versatility enables continuous innovation in targeting precision through novel ligand engineering and stimuli-responsive designs while maintaining unparalleled translational potential for pharmaceutical development.
Polymer-Based Nanoparticles
Polymer Nanoparticles
Polymer nanoparticles, solid colloidal carriers (10–1000 nm) engineered from natural biopolymers like chitosan and alginate derivatives or synthetic polymers including PLGA, PEG, and polystyrene, function as versatile drug delivery platforms through either nanosphere matrices that uniformly disperse therapeutic payloads or nanocapsule structures that encapsulate drugs within protective cores.149 These systems demonstrate exceptional cargo versatility, accommodating hydrophobic/hydrophilic agents across molecular weight spectrums from small molecules to biologics, proteins, and genetic therapeutics, while precise modulation of nanoparticle composition, surface charge, and degradation kinetics enables controlled drug loading efficiency and programmable release profiles.150,151
Polymer nanoparticles demonstrate significant potential in epigenetic combination therapy through multifunctional engineering approaches, as evidenced by Li et al’s PLGA-based system co-delivering JQ1 (BET inhibitor) and icariin, which effectively suppressed breast cancer progression and bone metastasis through optimized biodistribution. The tumor weight data showed that PLGA-based co-loaded system suppressed 65.8% of tumor growth, which was 2.0-fold higher than that of the free drug groups.152 Ledezma et al further expanded therapeutic modalities by encapsulating indocyanine green (photosensitizer) with nexturastat (HDAC6 inhibitor) in PLGA nanoparticles, achieving NIR-responsive photothermal ablation combined with histone deacetylase inhibition to prolong melanoma survival outcomes.153 Hu et al engineered PEG-PCL nanoparticles conjugated with nivolumab (PD-1 antibody) and loaded with decitabine (DNMT inhibitor), enhancing DNA hypomethylation effects while restoring PD1+CD8+ T-cell infiltration for improved checkpoint blockade efficacy.154 Advanced hybrid systems developed by Su et al combined lipid-coated PLGA nanoparticles co-encapsulating decitabine and doxorubicin, demonstrating synergistic apoptosis induction through epigenetic chemosensitization mechanisms.155 These case studies collectively validate polymer nanoplatforms’ capacity to integrate epigenetic modulation with diverse treatment modalities while addressing pharmacological challenges through structural engineering.
Polymeric Micelles
Polymeric micelles, self-assembled nanoscale structures formed by amphiphilic block copolymers in aqueous solutions, consist of hydrophilic polyethylene glycol (PEG) segments and hydrophobic domains such as polylactic acid (PLA), polycaprolactone (PCL), or phospholipid derivatives.156,157 These supramolecular assemblies demonstrate versatile drug-loading capacities for diverse therapeutic agents—from small molecules to proteins—enhancing drug stability, solubility, and bioavailability while exhibiting tumor-targeting potential through passive accumulation or ligand-mediated active targeting.158 Their structural adaptability enables rational design of stimuli-responsive systems (eg, pH-, redox-, or enzyme-sensitive) by engineering copolymer compositions to achieve spatiotemporally controlled drug release aligned with pathological microenvironments, thereby optimizing therapeutic precision and minimizing off-target effects.159
Polymeric micelle architectures enable innovative epigenetic combination strategies through tailored therapeutic delivery mechanisms, as exemplified by Naz et al’s PLGA-PEG micelles stabilizing azacytidine for pH-responsive release in breast cancer cells, bypassing nucleoside transporter limitations while resisting drug efflux to enhance antiproliferative effects.160 Xiong et al engineered redox-responsive prodrug micelles releasing γ-oryzanol (DNMT inhibitor) and α,β-methylene adenosine 5’-diphosphate (adenosine deaminase inhibitor) upon glutathione activation, synergistically inducing pyroptosis and reversing immunosuppressive microenvironments. The mean survival time of the combined group (17.8 days) was significantly prolonged compared to that of the control group (11 days), the group receiving α,β-methylene adenosine 5’-diphosphate therapy alone (12.7 days) and the group reviving the γ-oryzanol therapy alone group (14.2 days). In addition, 1/6 of mice in the combined group survived longer than 25 days.161 Advanced multimodal systems include Ding et al’s dual-micelle platform combining TBE photosensitizer with decitabine for light-controlled activation of cGAS-STING pathways and pyroptosis-mediated immunogenic cell death, which promoted antigen-presenting cell maturation and T-cell antitumor immunity. Finally, dual-micelle with laser treatment exhibited the most robust tumor eradication with a regression rate of 70.9%.162 Further innovation is demonstrated by Zhao et al’s DSPE-PEG2000-stabilized micelles co-assembling chlorin e6 (Ce6) and mocetinostat through electrostatic interactions, achieving tumor-specific accumulation and deep tissue penetration for photo-enhanced epigenetic regulation that suppressed both primary and metastatic tumor progression through optimized pharmacokinetics and immunomodulation. More importantly, combinative administration of micelles and α-PD-L1 upon light exposure was found to significantly inhibit the distant tumor growth. It was calculated that the primary and distant tumor inhibitory rates were about 92.5% and 82.2%, respectively, after the photodynamic amplified ICB therapy.163
Dendrimers
Dendrimers represent a class of monodisperse macromolecules characterized by radially branched architectures comprising a core, linear polymeric backbones, and hierarchically organized dendritic side chains, enabling precise control over their mass, three-dimensional geometry, and surface functionality.164,165 These structurally defined polymers possess exterior-active functional groups for covalent conjugation of biomolecules or imaging agents, while their internal cavities accommodate therapeutic payloads such as nucleic acids or small molecules. Commonly employed dendritic scaffolds include poly(amidoamine) (PAMAM), polypropylene imine (PPI), polyethylene imine (PEI), polylysine (PLL), and polyaryl ether derivatives, with their programmable surface chemistry and encapsulation capacity making them versatile platforms for controlled drug delivery and biomedical applications.166,167
Dendrimers demonstrate multifaceted potential in epigenetic combination therapy by enabling co-delivery of small molecule drugs and genetic therapeutics through programmable molecular architectures, as exemplified by Zong et al’s tumor-targeted dendrimer-HDAC inhibitor conjugates, where hydroxamic acid-linked HDACi remains pharmacologically inert until tumor-specific esterase cleavage reactivates its epigenetic modulation capacity.168 Kaundal et al further optimized gene targeting using polyethylenimine (PEI)-EZH2 siRNA complexes that enhanced systemic stability and hematopoietic compatibility, achieving potent EZH2 silencing in acute myeloid leukemia (AML) xenograft models with significant reductions in peripheral/blood marrow AML cell counts. The PEI-EZH2 siRNA complexes in AML-bearing animals significantly reduced CD45+/CD11b+ AML populations to 31% in peripheral circulation.169 These studies collectively highlight dendrimers’ capacity to overcome pharmacological barriers through structural precision—enhancing payload stability, tissue specificity, and therapeutic efficacy in epigenetic regulation.
The review on the application of polymer-based nanoparticles for epigenetic-based cancer therapy was summarized as follows: Polymer-based nanoparticles hold significant innovation potential through systematic architectural engineering that progresses in tandem with advancements in synthetic chemistry, while their translation into functionalized delivery systems necessitates concurrent optimization of industrial-scale production safety and organic reagent biocompatibility profiles.
Cell-Derived Nanoparticles
Extracellular Vesicles (EVs)
EVs—encompassing micrometer-scale microvesicles and 30–100 nm exosomes—are naturally secreted lipid-bilayer nanostructures that mediate intercellular communication through targeted cargo delivery, functioning in immune regulation, angiogenesis, and inflammatory processes while exhibiting inherent biocompatibility, prolonged circulation, and low immunogenicity.170,171 These endogenous nanocarriers enable precision drug delivery via membrane fusion-mediated cytoplasmic transfer of therapeutic payloads, as evidenced by studies demonstrating their capacity to bypass complex cellular uptake pathways through efficient membrane integration.172 Recent advances leverage EVs’ intrinsic targeting specificity and biosafety profiles to engineer advanced drug delivery systems, with research confirming their translational potential in constructing clinically viable therapeutic platforms through optimized cargo encapsulation and cell-specific homing mechanisms.173
EVs demonstrate multifaceted applications in epigenetic combination therapy through engineered delivery systems, as evidenced by You et al’s EV-based platform targeting m6A modification via YTHDF1 siRNA delivery, which suppressed gastric cancer progression through m6A-dependent transcriptional regulation. The tumor-bearing mice treated with EV-based platform showed the longest survival time with an overall survival rate of 66.7% compared to the other groups.174 Simultaneously, Zhai et al developed PD-1-enriched T-cell membrane vesicles encapsulating the LSD1 inhibitor ORY-1001, achieving tumor-specific recognition that amplified intratumoral IFN-γ/MHC-I signaling, resulting in 8-fold and 29-fold increases in cytotoxic/activated T-cell infiltration with potent xenograft suppression.175 Further extending this paradigm, Du et al’s mesenchymal stem cell-derived EVs co-delivered YTHDF1-targeting siRNA and docetaxel (DTX), synergizing epigenetic modulation with chemotherapy through enhanced cellular uptake and nucleic acid protection.176 Complementing these approaches, Lin et al engineered artificial exosomes incorporating tumor membrane proteins to co-encapsulate vorinostat (HDAC inhibitor) and doxorubicin, demonstrating sustained tumor retention and synergistic NSCLC suppression without significant toxicity. Artificial exosome containing vorinostat and doxorubicin almost completely suppressed the growth of tumor volume and weight, compared with artificial exosomes containing a single drug presenting an inhibition of ~58%.177 Collectively, these EV-based strategies exemplify precision epigenetic-chemotherapy integration through endogenous targeting mechanisms and payload stabilization.
Cell Membrane Carrier
Cell membrane-coated carriers leverage the inherent biological advantages of native cellular structures, where isolated membranes from macrophages, erythrocytes, dendritic cells, or tumor cells are fused with synthetic nanoparticles through extrusion techniques to create biomimetic nanomaterials that minimize host immune responses while preserving source cell functionality.178 These hybrid systems combine the physicochemical characteristics of engineered nanoparticles with the biological recognition capabilities of natural cell membranes, resulting in enhanced biocompatibility and optimized drug delivery kinetics. The fabrication process involves sequential co-extrusion of cell membrane fragments and core nanoparticles through polycarbonate membranes, generating carriers that retain both the targeting properties of biological membranes and the controlled release profiles of synthetic nanoplatforms, thereby achieving precise therapeutic delivery through dual biological-physical optimization mechanisms.179
Cell membrane coating technology significantly enhances epigenetic combination therapy by optimizing drug biodistribution and therapeutic precision, as demonstrated through multiple innovative approaches. Gao et al engineered macrophage membrane-coated nanoinducers co-loaded with paclitaxel (PTX) and decitabine, which reversed triple-negative breast cancer chemoresistance by inducing tumor tissue demethylation, restoring T-cell functionality, and sensitizing malignancies to chemotherapy through inherent tumor-homing capabilities. This nanoinducers, when combined with PD-1 checkpoint blockade, can achieve a tumor suppression rate of 90.04%.180 Simultaneously, Zhang et al developed lung cancer cell membrane-disguised nanoparticles carrying BRD4-targeting proteolysis-targeting chimeras (PROTACs), enabling dual targeting of lung cancer cells and tumor-associated macrophages to synergistically induce apoptosis in subcutaneous and orthotopic tumor models through epigenetic reader protein degradation. About 90% mice survived over 18 days initially from the treatment with drug-loaded nanoparticles, when all of the mice were nearly dead in the control groups.181 Furthermore, Wang et al’s glioblastoma multiforme (GBM) therapeutic platform utilized tumor cell membrane-wrapped nanoparticles containing ultrasmall Cu2ₓSe, indoximod, and JQ1, which crossed the blood–brain barrier to remodel the tumor immune microenvironment (TIME) by activating CD8⁺ T-cell infiltration, elevating splenic memory T cells, and preventing tumor recurrence through coordinated epigenetic modulation and immune checkpoint regulation.182 These membrane-engineered systems collectively demonstrate enhanced tumor targeting, payload protection, and multimodal therapeutic synergy in epigenetic–immune combination regimens.
The review on the application of cell-derived nanoparticles for epigenetic-based cancer therapy was summarized as follows: Overall, EVs and cell membrane-based carriers demonstrate precise tumor-targeting capabilities and combinatorial drug delivery potential in murine models, while clinical translation remains constrained by suboptimal exosome isolation yields and challenges in scalable manufacturing, necessitating breakthroughs in bioprocessing technologies to bridge preclinical efficacy with therapeutic applications.
Inorganic Nanoparticles
Metallic-Based Nanoparticles
Epigenetic-radiotherapy combination strategies are being revolutionized by metal-based nanoparticles composed of high-atomic-number elements that enhance treatment efficacy through dual photon-interactive mechanisms—inducing ROS generation via low-energy electron emission while functioning as co-delivery platforms for epigenetic therapeutics. These nanoplatforms achieve tumor-specific accumulation through surface functionalization with targeting ligands, enabling precise radiosensitization and combinatorial payload release.183,184 Preclinical validation encompasses diverse metallic systems including gold, platinum, rare-earth, and semiconductor nanoparticles engineered for concurrent radiation enhancement and epigenetic drug transport, with their multifunctional architecture optimizing spatiotemporal coordination between DNA damage potentiation and epigenetic modulation.185,186
Epigenetic-radiotherapy combinations are being advanced through innovative nanoplatforms, as demonstrated by Kaundal et al’s hypericin-conjugated gold nanoparticles targeting mitochondrial IDH2-EZH1/2 interactions in glioblastoma, achieving multimodal tumor ablation via laser activation in xenograft models. The results show that the tumor volume of the hypericin-conjugated gold nanoparticles treatment group is almost 10-fold more suppressed than the PBS control group, and 3-fold with the hypericin group187 Simultaneously, Song et al engineered leukemia stem cell membrane-coated bimetallic Mn2⁺/Fe3⁺ MOFs delivering DNA hypomethylation agents and autophagy-inducing peptides, which restored MHC-I expression and interferon signaling to enhance T-cell immunity against leukemic progenitors. The results showed that the low dose of the above-mentioned preparation (50 µg/mL) inhibited tumor growth by 67.6% compared to 31.7% in the AZA alone group, and achieved 66.7% survival rate over a period of 60 days.On the other hand, the high dose of the above-mentioned preparation (100 µg/mL) achieved almost complete tumor elimination, and 100% of the treated mice survived for 60 days post-injection.188 Addressing MYC-mediated radioresistance, Wang et al engineered tungsten-based nanosystems co-loaded with 5-Aza (DNMTi) and ITF-2357 (HDACi) that suppress MYC-driven immunosuppression while enhancing dendritic cell antigen presentation and cytotoxic T-cell infiltration through radiotherapy-potentiated immunogenic effects.189 Wang et al’s EGCG-W6⁺ polyphenol complexes induce caspase-3-mediated gasdermin E cleavage during irradiation, triggering pyroptosis while counteracting radiotherapy-induced Treg elevation through coordinated epigenetic modulation and radiosensitization.190 These multifunctional platforms demonstrate enhanced immunogenic cell death pathways surpassing conventional radiotherapy through precision epigenetic-immune crosstalk regulation.
Non-Metallic Nanoparticles
Non-metallic nanoparticles—a critical subset of nanomaterials distinguished by their composition of non-metallic elements—exhibit unique physicochemical properties including quantum confinement effects, enhanced surface-to-volume ratios, and tunable optoelectronic behaviors that enable superior adsorption capacities and catalytic reactivities.191,192 While demonstrating improved biosafety profiles compared to metallic counterparts, these systems face challenges in biodegradation that require material engineering solutions. Their structural precision facilitates targeted drug delivery through size-dependent cellular interactions and controlled release kinetics, minimizing off-target effects while optimizing therapeutic payload delivery. Representative systems such as mesoporous silica nanoparticles (MSNs) and carbon-based nanostructures exemplify this category’s translational potential, leveraging ordered porosity and electronic configurations to achieve spatiotemporal control over drug loading/release processes in biomedical applications.193,194
Epigenetic combination therapies are being optimized through advanced nanocarrier systems, as demonstrated by Gao et al’s mesoporous silica nanoparticles (MSNs) co-encapsulating paclitaxel (PTX) and decitabine with surface biofunctionalization, which achieved tumor-specific demethylation to reverse T-cell exhaustion while enhancing chemosensitivity in a precisely controlled drug ratio.180 Concurrently, Yuan et al engineered CD44-targeted MSNs integrating pH/redox-responsive β-cyclodextrin gatekeepers for co-delivery of doxorubicin (DOX) and siGCN5, suppressing multidrug-resistant breast tumor growth through dual inhibition of P-glycoprotein efflux and histone acetyltransferase GCN5, with hyaluronic acid coating enhancing tumor-specific internalization. Of note, in an MDR breast tumor model, DOX and siGCN5 co-delivered MSNs inhibits MDR tumor growth by 77%, abolishes P-gp-mediated drug resistance, and eliminates DOX’s systemic toxicity.195 Complementing these approaches, Gu et al addressed the aqueous insolubility of the EHMT2 inhibitor UNC0646 by developing physisorbed nanodiamond complexes (ND-UNC0646) that enabled pH-responsive drug release and improved dispersibility, demonstrating enhanced anti-tumor efficacy in orthotopic liver cancer models through intravenous delivery while maintaining dosing precision.196 These multifunctional platforms exemplify the integration of epigenetic modulation with chemotherapy and targeted delivery to overcome pharmacological barriers.
The review on the application of inorganic nanoparticles for epigenetic-based cancer therapy was summarized as follows: In conclusion, inorganic nanomaterials demonstrate distinct advantages in combinatorial drug delivery systems, offering precise tumor-targeting capabilities and extensive therapeutic applicability through their tunable physicochemical properties. However, the clinical translation of these nanoplatforms necessitates rigorous pharmacological optimization to reconcile therapeutic efficacy with biocompatibility profiles, requiring systematic engineering approaches that address biodistribution control, biodegradation kinetics, and long-term biosafety parameters to fully harness their diagnostic and therapeutic potential in oncology applications.
Hydrogel Delivery Systems
Hydrogels
Hydrogels represent three-dimensional crosslinked polymer networks synthesized through covalent or non-covalent interactions between hydrophilic monomers, exhibiting unique water-retention capacities through swelling behavior that enables absorption of substantial aqueous volumes while maintaining structural integrity without dissolution.197,198 Characterized by tissue-mimetic elasticity derived from their high water content, these biomimetic materials demonstrate exceptional potential in therapeutic agent encapsulation and controlled release, facilitated by tunable mesh sizes and stimuli-responsive release mechanisms including diffusion-controlled kinetics, pH/thermal-triggered swelling, and enzymatic/redox-mediated degradation.199 This unique physicochemical profile enables sustained local drug depot formation with prolonged high-concentration payload maintenance, while their programmable responsiveness to physiological or external stimuli allows spatiotemporal precision in therapeutic delivery across biomedical applications.200,201
Hydrogel-based epigenetic drug delivery systems demonstrate transformative therapeutic potential through engineered responsiveness to tumor microenvironments, as exemplified by Ruan et al’s pH/calcium carbonate nanoparticle-embedded hydrogel co-encapsulating anti-PD-1 antibodies and zebularine—this dual-delivery platform enhanced tumor-associated antigen presentation while reversing PD-L1-mediated immunosuppression, achieving tumor growth inhibition and survival extension in murine models through coordinated checkpoint blockade and DNA demethylation mechanisms.202 Ji et al developed ROS-responsive hydrogels releasing GSK-LSD1 (LSD1 inhibitor) and 5-fluorouracil (5-FU) that reduced cancer stemness through H3K4me2 epigenetic remodeling while increasing MHC-I expression, converting chemotherapy-resistant triple-negative breast tumors into treatment-sensitive phenotypes with suppression of postoperative recurrence and reduction in metastatic burden. By the day 20 after hydrogel injection, the 5-FU@Gel showed little effect on tumor growth, while the iLSD1@Gel reduced the average tumor size by 38.0 ± 8.5%. Furthermore, iLSD1+5-FU@Gel achieved remarkable efficacy, and most tumors ceased proliferation in the first 20 d and exhibited slow growth velocities afterward. The median survival time of mice after PBS mock treatment was 26 days, which was slightly increased to 30 and 34 d by the 5-FU@Gel and iLSD1@Gel, respectively. In contrast, the iLSD1+5-FU@Gel significantly extended this value to 60 d.203 These intelligent hydrogel platforms exemplify precision epigenetic reprogramming through spatiotemporal control of therapeutic payloads, leveraging microenvironmental triggers (pH/ROS) to overcome multidrug resistance while establishing sustained local drug reservoirs that maintain therapeutic concentrations for 14–21 days post-implantation.
Nanogels
Nanogels—nanoscale three-dimensional networks formed through physical or chemical crosslinking of polymeric macromolecules such as acrylics, polyvinylpyrrolidone, or polysaccharides—combine the structural advantages of hydrogels with nanoparticle functionality, exhibiting high drug-loading capacities, sustained-release kinetics, and enhanced tissue permeability through sub-200 nm particle sizes.204 Their swollen hydrogel architecture protects encapsulated therapeutics from degradation while providing modifiable surfaces for targeted delivery, with biodegradability and biocompatibility profiles enabling clinical translation.205 Vijayaraghavalu et al demonstrated this potential through disulfide-crosslinked N-isopropylacrylamide/vinylpyrrolidone nanogels co-loading decitabine (DAC) and vorinostat (SAHA), which overcame multidrug resistance via epigenetic reprogramming, achieving tumor growth suppression at subtherapeutic doxorubicin doses through sustained chromatin remodeling effects.206 This mechanistic paradigm—attributed to prolonged epigenetic modulator release from nanogel matrices—provides a universal strategy against therapy-resistant malignancies driven by aberrant DNA methylation or histone modification, with tunable crosslinking methods (noncovalent interactions vs covalent bonding) allowing precise control over drug release profiles tailored to specific tumor microenvironments.
The review on the application of hydrogel delivery systems for epigenetic-based cancer therapy was summarized as follows: Hydrogel-based drug delivery systems demonstrate transformative potential in contemporary therapeutics, with researchers actively engineering combinatorial platforms that integrate epigenetic modulation capabilities through hydrogel matrices while advancing multifunctional hydrogel architectures to synergize with radiotherapy, immunotherapy, and targeted molecular interventions. This dual-axis development strategy—enhancing epigenetic payload precision through stimuli-responsive release mechanisms and optimizing viscoelastic properties for tissue-specific integration—addresses critical challenges in spatiotemporal drug control, enabling sustained therapeutic agent delivery with programmable degradation kinetics that maintain bioactive concentrations across treatment cycles.
Conjugated Drug
Antibody-Drug Conjugates (ADCs)
ADCs, which combine tumor-targeting monoclonal antibodies with cytotoxic payloads via specialized linkers, represent a transformative therapeutic modality that merges the precision of biologics with the potency of chemotherapy.207,208 With nearly 10 ADCs now FDA-approved and over 100 in clinical trials, this platform’s success has catalyzed exploration of next-generation constructs integrating epigenetic modulators—where antibody-mediated delivery of chromatin-remodeling agents could synergize with cytotoxic payloads through mechanisms like DNA damage response potentiation and immune microenvironment reprogramming, potentially overcoming limitations of conventional combination therapies through spatiotemporal coordination of epigenetic priming and tumor cell elimination.209,210
Emerging epigenetic antibody-drug conjugates (ADCs) are expanding the therapeutic scope beyond traditional cytotoxic payloads, as exemplified by Cini et al’s cetuximab-based ADC incorporating the HDAC inhibitor ST7612AA1 through lysine amide conjugation—this EGFR-targeting construct demonstrated tumor-specific internalization and potent antitumor efficacy in solid tumor models while circumventing the systemic toxicity associated with conventional microtubule-inhibitor ADCs, representing the first clinical-stage ADC with epigenetic modulation capabilities applicable to both oncologic and non-oncologic indications.211 Complementing this approach, Milazzo et al engineered trastuzumab-derived ADC ST8176AA1 through TCEP-mediated disulfide reduction and maleimide-thiol conjugation of activated HDAC inhibitor ST7464AA1, which maintained ErbB2 binding specificity while exhibiting enhanced tumor growth inhibition in ovarian/colon cancer xenografts and pancreatic PDX models compared to parental antibody therapy. ST8176AA1 induced a median survival time of 84 days compared to 43 and 74 days of vehicle- and trastuzumab-treated groups, respectively. These results are consistent with the previous study confirming that ST8176AA1 is statistically significant more effective than trastuzumab against ovary carcinoma.212 Mechanistic studies confirmed the enhanced efficacy stemmed from HDAC-mediated chromatin remodeling rather than cytotoxic mechanisms, with pharmacokinetic analyses revealing 14-day sustained HDAC inhibition in tumor tissues versus <48h activity with free inhibitors, validating ADCs as precision delivery platforms for epigenetic therapeutics.
Small Molecule-Drug Conjugates (SMDCs)
SMDCs, an emerging class of targeted therapeutics, share structural homology with ADCs through their tripartite architecture comprising targeting ligands, chemical linkers, and effector payloads (eg, cytotoxic agents or E3 ligase recruiters), yet fundamentally diverge by utilizing low-molecular-weight targeting moieties instead of monoclonal antibodies.213 This substitution confers distinct pharmacological advantages: simplified synthesis with enhanced process control and reduced manufacturing costs compared to biologics, inherent non-immunogenicity facilitating improved safety profiles, and superior tumor penetration enabled by sub-10 nm hydrodynamic diameters, particularly advantageous in solid malignancies where extracellular matrix density limits macromolecule diffusion.214,215 The small molecule targeting approach preserves precise drug localization while eliminating antibody-mediated immunogenic risks, with preclinical evaluations demonstrating stable pharmacokinetic behavior and tumor-selective payload release through microenvironment-responsive linker systems.216
Small molecule-drug conjugates are advancing epigenetic combination therapies through innovative nanostructural designs, as exemplified by Xu et al’s cisplatin-vorinostat supramolecular nanoparticles that achieve 99% tumor suppression in drug-resistant A549 models through synchronized drug release and prolonged circulation, overcoming the negligible efficacy of free drug combinations.217 Concurrently, Wang et al’s zinc-mediated self-assembled nanofibers co-delivering 5-azacytidine and vorinostat demonstrate dual epigenetic modulation by coordinating DNA demethylation with HDAC inhibition, inducing tumor-selective apoptosis in gastric cancer through sustained intratumoral retention. It is clearly found that compared with free vorinostat or the 5-azacytidine treatment, the drug mixture treatment exhibited a better antitumor effect, which may be explained by the synergistic actions. Surprisingly, mice treated with the nanofibers exhibited the smallest tumor size and the highest inhibitory rate, suggesting the best antitumor efficiency of nanofibers.218 Complementing these approaches, Ye et al engineered glutathione-responsive prodrug nanoparticles (NJ) cloaked with pH-sensitive polymers, where tumor-acidified dissociation enables intracellular release of JQ1 (BET inhibitor) and NLG919 (IDO1 inhibitor), concurrently blocking PD-L1 upregulation and tryptophan catabolism to reverse interferon-γ-induced immune evasion—a strategy amplified when combined with photodynamic therapy to eradicate multiple immunosuppressive pathways through spatiotemporal control of epigenetic-immune crosstalk.219
The review on the application of conjugated drug for epigenetic-based cancer therapy was summarized as follows: Conjugated drugs represent a transformative paradigm in oncology therapeutics, pioneering next-generation precision medicine through their capacity to integrate tumor-targeting specificity with multimodal therapeutic payloads—a breakthrough underscored by the global biopharmaceutical sector’s accelerated investment. This therapeutic class is poised to redefine cancer treatment boundaries through enhanced tumor selectivity, payload potency amplification, and resistance mechanism circumvention.
Others
Recent advances in nanocarrier design are revolutionizing epigenetic combination therapies through innovative delivery strategies, as exemplified by Luo et al’s folic acid receptor-targeted nanoparticles co-delivering azacitidine and PARP inhibitors, which achieved tumor-selective enrichment through protein corona engineering while enabling synergistic antitumor effects with PD-1 inhibitors in lung cancer models.220 Concurrently, Kaundal et al’s albumin-based nanoparticles demonstrated enhanced nuclear localization of EZH2 inhibitor EPZ011989 in AML cells through optimized non-covalent interactions, improving therapeutic index compared to free drug administration.221 Yang et al further advanced pH-responsive delivery using apolipoprotein-mimetic liposomes that synchronized 5-azacitidine and HDAC inhibitor release, inducing cell cycle arrest and apoptosis via Bim/caspase-3 activation while reactivating tumor suppressor genes in breast cancer models.222 Complementing these approaches, Tian et al engineered photothermally responsive polydopamine nanoparticles that coupled JQ1-mediated BET inhibition with localized hyperthermia, generating cytotoxic T-cell responses that prevented tumor rechallenge in 85% of treated mice through sustained epigenetic-immune reprogramming.223 These multimodal platforms collectively demonstrate how nanoscale engineering optimizes spatiotemporal control of epigenetic payloads while overcoming biological barriers through material-specific targeting mechanisms.
The review on the application of other NDDS for epigenetic-based cancer therapy was summarized as follows: The exploration of non-canonical nanocarrier architectures offers innovative solutions to overcome the limitations of conventional drug delivery systems through tailored physicochemical properties and programmable targeting strategies.
Delivery Strategies for Epigenetic-Based Cancer Combination Therapy
The NDDS demonstrates significant potential and promising development prospects in epigenetic-based combination therapy due to its capacity for co-delivering multiple therapeutic agents. The complexity of epigenetic modifications, characterized by diverse molecular targets, is exemplified in the tumor microenvironment where epigenetic reprogramming of immune cells, stromal cells, and other components influences cancer progression. Within cellular contexts, epigenetic enzymes may serve as therapeutic targets localized in specific organelles. While these systems must achieve effective tumor accumulation, challenges arise when addressing temporally dependent therapeutic targets requiring sequential or programmed drug release. Current limitations include the intricate temporal control of combination therapies and compromised patient compliance caused by complex administration regimens. To address these issues, advanced NDDS should integrate multifunctional capabilities beyond mere drug co-loading, encompassing tumor tissue targeting, multi-cellular delivery precision, and spatiotemporal regulation at subcellular levels (Figure 3).
 |
Figure 3 Delivery strategies for epigenetic-based cancer combination therapy. |
Passive Targeting
Passive targeting leverages the physiological and pathological characteristics of tumor sites by modulating nanoparticle properties such as size, morphology, structure, and surface characteristics to achieve tumor accumulation without relying on molecular recognition mechanisms.224,225 This approach capitalizes on the defective vascular architecture and impaired lymphatic drainage in rapidly proliferating tumors, collectively termed the enhanced permeability and retention (EPR) effect. These pathological features allow macromolecules and nanoparticles to extravasate into tumor tissues while hindering their systemic clearance, enabling conventional nano-drug delivery systems to passively concentrate therapeutic agents at tumor sites.226,227 The simplicity of designing such systems, coupled with their scalable production processes, has made them commercially attractive to pharmaceutical companies, as evidenced by all currently marketed anti-tumor nanomedicines belonging to this category. Beyond passive accumulation, emerging research demonstrates that precisely engineered nanoparticle morphologies and surface modifications can further enhance targeting efficacy, particularly in epigenetic drug delivery applications.228,229 These advanced design strategies have garnered significant scientific attention in recent years due to their potential to overcome limitations of conventional EPR-dependent systems while expanding therapeutic possibilities in tumor microenvironment modulation.
The particle size of nanomedicine delivery systems critically governs their tumor penetration efficacy and intratumoral retention capacity. Experimental evidence demonstrates an inverse correlation between nanoparticle dimensions and solid tumor penetration within defined size thresholds – smaller systems (eg, 12.3 nm PARP inhibitor/5-azacytidine co-delivery nanoparticles, PAZA) exhibit superior tumor-targeting and interstitial penetration compared to larger counterparts (eg, 100 nm variants, PVD).220 Conversely, tumor retention capacity shows positive size dependence, with smaller nanoparticles demonstrating significantly reduced intratumoral persistence. This fundamental trade-off between penetration depth and retention duration has driven the development of stimuli-responsive nanoplatforms capable of dynamic size modulation. Current innovative designs employ subtherapeutic-sized nanoparticles that undergo tumor microenvironment-triggered structural reorganization (via self-assembly, electrostatic interactions, or phase transitions) to achieve size transformation from sub-20 nm penetration-optimized states to retention-enhanced configurations exceeding 100 nm. This spatiotemporal size adaptation strategy synergistically improves both deep tumor permeation and prolonged drug retention, significantly amplifying therapeutic outcomes.230,231 Such intelligent nanosystems provide valuable insights for advancing epigenetic combination therapies, where coordinated delivery of multiple agents requires precise spatiotemporal control over drug distribution and release kinetics.
The surface charge of NDDS critically modulates their circulation longevity and cellular internalization efficiency. Positively charged NDDS exhibit heightened plasma protein binding affinity, leading to accelerated clearance by the mononuclear phagocyte system (MPS), whereas negatively charged counterparts demonstrate reduced opsonization and extended plasma half-lives, explaining the predominant use of anionic surface modifications in clinical nanomedicines.232,233 This charge-dependent biointeraction creates a therapeutic paradox: while cationic systems facilitate cellular uptake through electrostatic attraction to negatively charged cell membranes, their rapid systemic elimination compromises therapeutic efficacy. To resolve this dichotomy, advanced nanoplatforms with dynamically tunable surface properties have been engineered. A representative example involves cationic liposomes co-encapsulating Zebularine (DNA methyltransferase inhibitor) and JQ1 (BET protein inhibitor), which are electrostatically shielded with carboxymethyl-chitosan (CG-J/ZL).143 This design maintains a stealthy anionic surface during systemic circulation, minimizing MPS recognition, yet undergoes pH-responsive charge reversal in the acidic tumor microenvironment. The resultant switch to positive surface potential enhances both tumor tissue penetration depth and cancer cell internalization rates. Such intelligent charge-transition systems exemplify the strategic integration of prolonged circulation stability with tumor-specific activation, providing a versatile blueprint for optimizing epigenetic combination therapies that require spatiotemporal coordination of drug bioavailability and target engagement.
Active Targeting
Active targeting mechanisms operate through precise ligand-receptor interactions, where nanoparticles functionalized with targeting moieties (eg, antibodies, peptides, or small-molecule ligands) via bioconjugation techniques achieve selective binding to overexpressed cellular receptors or tumor-associated biomarkers.234 This molecular recognition process facilitates receptor-mediated endocytosis, enabling intracellular delivery of therapeutic payloads to predetermined cell populations.235 The advancement of cellular biology and pathological understanding has transformed active targeting from theoretical concept to clinical reality, allowing spatiotemporal control over drug biodistribution while mitigating systemic toxicity – particularly critical for epigenetic combination therapies requiring cell-type-specific delivery to both malignant cells and tumor microenvironment components.
Tumor neovascularization constitutes a critical pathophysiological hallmark supporting malignant progression, characterized by distinct molecular signatures on endothelial cells including upregulated integrins, aminopeptidase N, and transmembrane glycoproteins.236,237 These angiogenic biomarkers enable targeted drug delivery through ligand-receptor recognition systems, as exemplified by NGR peptide-modified cationic liposomes engineered for CD13 receptor binding on tumor vasculature. In the 4T1 murine breast cancer model, Liu et al demonstrated that NGR-functionalized liposomes co-encapsulating Zebularine (DNA methyltransferase inhibitor) and JQ1 (BET bromodomain inhibitor) achieved significantly higher tumor accumulation versus non-targeted counterparts.143 Beyond synthetic targeting strategies, bioinspired approaches leverage endogenous cellular tropism – macrophage-derived exosome membrane-camouflaged nanoparticles loaded with suberoylanilide hydroxamic acid (SAHA) exploit the α4β1 integrin/VCAM-1 axis to infiltrate invasive tumor niches. This biomimetic design preserves natural immune-evasion properties while inheriting macrophage-specific tumor homing capabilities, resulting in significantly reduction in metastatic burden compared to conventional nanoparticles in preclinical trials.238 The convergence of synthetic biology and nanomedicine thus establishes new paradigms for overcoming biological barriers in cancer treatment, particularly for the spatial complexity of epigenetic combination therapies that require coordinated delivery to both neovasculature and tumor parenchyma.
Tumor cells constitute the principal cellular components of malignancies, making their selective elimination a cornerstone of cancer therapeutics. The accelerated proliferation of tumor cells drives the overexpression of surface receptors—including transferrin receptors, folate receptors, low-density lipoprotein receptors, and glucose transporters—compared to normal cells. These molecular signatures enable targeted drug delivery through receptor-specific ligands, as exemplified by lactoferrin-modified dual-drug liposomes co-loaded with Panobinostat and JQ1, which achieve active targeting via binding to low-density lipoprotein receptor-associated protein 1 (LRP1) on tumor surfaces.141 Similarly, hyaluronic acid-functionalized solid lipid nanoparticles (SLNs) exploit CD44 receptor overexpression for tumor-specific delivery of the histone deacetylase inhibitor Vorinostat.239 Beyond tumor cells, epigenetic modulation involves multiple cellular components within the tumor microenvironment. Tumor-associated lymphocytes, particularly T cells and macrophages, critically promote tumor progression and metastasis, necessitating targeted therapeutic strategies. Nanoparticles conjugated with anti-programmed death-1 monoclonal antibodies (eg, nivolumab) and loaded with decitabine (DAC) demonstrate dual functionality: they selectively target PD-1-overexpressing tumor-infiltrating lymphocytes (TILs), inhibit DNA methylation to reverse PD1+CD8+ TIL exhaustion, and synergistically enhance immune checkpoint blockade efficacy through T cell reinvigoration.154 The integration of molecular recognition precision with epigenetic modulators’ pleiotropic mechanisms establishes a transformative framework for next-generation combination therapies, particularly in immune-cold tumors refractory to conventional treatments.
Responsive Release
Although nanomedicine delivery systems with passive or active targeting can deliver epigenetic drugs to tumor sites and inhibit tumor development by regulating specific cellular epigenetic mechanisms, the TME poses a significant obstacle to antitumor drug delivery. The TME, characterized by complex tumor localization, consists of various cells and their secretions, including peripheral blood vessels, extracellular matrix, tumor cells, and cytokines.240 As tumors develop, the TME undergoes substantial physicochemical changes at cellular and tissue levels, manifesting as high cell density, elevated interstitial pressure, micro-acidity, hypoxia, and abnormal expression of extracellular matrix components and enzymes.241,242 While characteristics such as high cell density and interstitial pressure limit nanoparticle penetration and distribution within tumors, other features like hypoxia, micro-acidity, and overexpression of specific enzymes are exploited as endogenous stimuli in smart responsive drug delivery systems. These systems utilize these conditions to mediate targeted molecular activation, structural transformations, and controlled drug release, thereby enhancing tumor-targeted delivery efficiency.243,244 Such responsive nano-drug delivery systems demonstrate significant potential for clinical translation of epigenetic therapies. Specifically, their intelligent responsiveness enables carrier depolymerization within tumor tissues followed by precise cellular targeting, which could substantially improve the efficacy of epigenetic treatments.
Throughout tumorigenesis and progression, the TME acquires pathological features including hypoxia, acidosis, elevated oxidative stress, supraphysiological glutathione (GSH) levels, and upregulated enzymatic activity. These distinctive features enable selective therapeutic strategies for nanomedicine delivery systems, particularly hypoxia-responsive, pH-responsive, reduction-oxidation (redox)-responsive, ATP-responsive, and enzyme-responsive platforms specifically designed for TME modulation.245,246 A representative paradigm involves core-shell nanoparticles loaded with Zebularine, JQ1, and CpG, surface-functionalized with carboxymethyl chitosan (CMCS). This pH-sensitive coating undergoes degradation in acidic TME, triggering sequential drug release: Zebularine induces tumor-associated antigen (TAA) upregulation, potentiating tumor immunogenicity. JQ1 suppresses PD-L1 expression to abrogate immune checkpoint signaling. CpG activates dendritic cell maturation while reprogramming tumor-associated macrophages (TAMs) toward tumoricidal phenotypes.143 This multimodal approach demonstrates significant potential for revitalizing anti-tumor immunity through coordinated epigenetic modulation and immune microenvironment remodeling.
The strategic integration of multiplexed TME stimuli with rationally engineered cascade-responsive nanoplatforms enables a hierarchical therapeutic paradigm encompassing tumor-preferential drug accumulation, subcellular targeting precision, and spatiotemporally controlled release kinetics, which synergistically amplifies antitumor epigenetic therapy efficacy while minimizing systemic toxicity. Illustrating this concept, Ruan et al developed a dual-responsive hydrogel platform capable of sequentially sensing acidic pH and reactive oxygen species (ROS) gradients to co-deliver anti-PD1 antibodies (aPD1) and the hypomethylating agent Zebularine (Zeb). Mechanistically, pH-sensitive calcium carbonate nanoparticles (CaCO3 NPs) were utilized to nanoformulate aPD1, which were subsequently co-encapsulated with Zeb within a ROS-labile hydrogel matrix (Zeb/aPD1-NPs-Gel). Preclinical evaluation in B16F10 melanoma models demonstrated concomitant immunogenic cell death induction and immunosuppressive network reprogramming, culminating in significant tumor growth suppression and extended median survival.202 This multimodal therapeutic strategy, integrating TME-responsive cascade delivery with epigenetic-immune crosstalk modulation, establishes a blueprint for precision oncology by orchestrating spatiotemporal drug release dynamics to concurrently potentiate tumor immunogenicity and dismantle immunosuppressive barriers, ultimately achieving therapeutic synergy with minimized off-target effects.
Intracellular Regulation
The complexity of epigenetic modification lies in its diverse cellular targets, including organelle-localized epigenetic enzymes that require precise intracellular delivery. Nanoparticle intracellular trafficking critically determines therapeutic efficacy through a multistage transport process: upon reaching the plasma membrane, nanoparticles undergo endocytic uptake via membrane invagination, forming endocytic vesicles that mature into early endosomes. These vesicular carriers subsequently engage the intracellular sorting network—a coordinated system involving the Golgi apparatus, endoplasmic reticulum (ER), and lysosomes—to determine nanoparticle fate. While most nanoparticles remain confined within the endolysosomal pathway, some achieve cytoplasmic escape prior to lysosomal fusion through membrane-disruptive mechanisms, thereby evading enzymatic degradation.247–249 Current research focuses on engineering nanoparticles with lysosomal escape capabilities using pH-responsive biomaterials or autophagy-inducing components, strategies particularly relevant for epigenetic drug delivery where nuclear access or mitochondrial targeting often requires intact cytoplasmic release. This spatial control over intracellular trafficking addresses the dual challenge of protecting epigenetic modulators from lysosomal inactivation while ensuring precise subcellular localization for optimal therapeutic intervention.
The endosomal-lysosomal pathway represents a critical barrier for therapeutic nanoparticles due to its degradative microenvironment characterized by acidic pH and enzymatic activity, which compromises drug bioavailability. To circumvent this limitation, nanoparticle engineering focuses on developing endosomal escape mechanisms involving membrane disruption strategies such as the proton sponge effect, lipid bilayer destabilization, and pH-dependent fusogenic properties.250 A representative paradigm employs vorinostat-loaded nanoparticles constructed from pH-responsive poly(lactic-co-glycolic acid) (PLGA) and 1,2-dioleoyl-3-trimethylammonium-propane (DOTAP). Upon endocytosis, the acidic endosomal environment triggers protonation of DOTAP’s tertiary amine groups, inducing osmotic swelling through the proton sponge effect. This physicochemical transition destabilizes endosomal membranes, facilitating cytoplasmic release of intact therapeutic payloads while bypassing lysosomal degradation.251 Such intelligent nanoparticle architectures demonstrate the potential to overcome intracellular trafficking barriers critical for epigenetic drug delivery, where maintaining drug structural integrity and achieving nuclear localization are essential for restoring normal chromatin remodeling processes.
Autophagic processes critically influence nanoparticle intracellular trafficking through their dual role as a degradation pathway and therapeutic modulator. Distinct from classical endocytic pathways, autophagy mediates lysosomal degradation of cytoplasmic components through autophagosome-lysosome fusion, posing a potential risk for recapturing cytoplasmically escaped nanoparticles.252,253 While this lysosomal re-encapsulation compromises drug bioavailability, emerging strategies exploit autophagy’s regulatory mechanisms for controlled drug release. A proof-of-concept design employs azacitidine-loaded biomimetic metal-organic frameworks (MOFs) functionalized with autophagy-related protein Beclin-1. Upon cellular internalization, Beclin-1 mediates selective binding to Golgi-associated plant pathogenesis-related protein 1 (GAPR-1), inducing autophagosome accumulation at Golgi apparatus membranes. This targeted autophagy activation achieves pH-responsive release of azacitidine while bypassing lysosomal degradation, enhancing DNA methyltransferase inhibition efficacy in leukemia models.188 Such autophagy-mediated delivery systems present a paradigm-shifting approach for epigenetic therapeutics, leveraging cellular quality-control mechanisms to achieve spatiotemporal drug release. These advancements highlight the necessity of comprehensive autophagy profiling in nanomedicine develocularly for epigenetic drugs requiring sustained nuclear bioavailability.
Challenges from the Laboratory to the Clinic
Despite the significant potential demonstrated by NDDS-based epigenetic combination therapies in preclinical studies, their translation from laboratory to clinical applications remains great challenges. These challenges span critical areas including scalable manufacturing, complex in vivo behaviors, and long-term safety.
A primary challenge is the transition from laboratory-scale production to large-scale manufacturing compliant with Good Manufacturing Practice (GMP) guidelines. The intricate architecture of NDDSs-such as the co-loading of multiple epigenetic drugs with diverse physicochemical properties, surface modification with targeting ligands, or incorporation of environmentally responsive polymers-poses significant challenges during process scaling. These challenges including batch-to-batch variability in critical quality attributes, fluctuations in drug loading efficiency, and high production costs severely constrain their clinical translation feasibility.
Furthermore, the pharmacokinetic (PK) profiles and immunogenicity risks of engineered nanocarriers in humans present another major challenge. The favorable PK characteristics observed in preclinical models may significantly diverge in human subjects due to inter-individual physiological variations, the formation of plasma protein coronas, and recognition by the mononuclear phagocyte system. This variability leads to unpredictable biodistribution, clearance rates, and tumor targeting efficiency. More critically, the introduction of nanocarriers may inadvertently activate both innate and adaptive immune responses. Such immunogenicity not only accelerates carrier clearance and compromises therapeutic efficacy but could also trigger severe inflammatory reactions, introducing unpredictable risks to treatment regimens.
Particularly noteworthy are the nanotoxicological and long-term safety concerns inherent in NDDS-mediated epigenetic combination therapies, which represent a core issue requiring meticulous evaluation in clinical translation. On the one hand, the mechanisms of action of epigenetic drugs themselves are non-specific. Off-target delivery of drugs or premature leakage from nanocarriers could result in unintended editing of the epigenome in healthy tissues. Such non-specific epigenetic perturbations—for instance, aberrant alterations in DNA methylation or widespread dysregulation of histone modifications—may induce permanent functional impairment in non-cancerous cells and potentially even drive the development of novel pathological states, the long-term consequences of which remain difficult to fully assess. On the other hand, the intrinsic biosafety of the carrier materials is paramount. For example, certain metal-based nanoparticles (such as those composed of gold, silver, or iron oxide) or metal-organic frameworks (MOFs) may undergo incomplete biodegradation and clearance in vivo, leading to their long-term accumulation in the reticuloendothelial system (eg, liver and spleen). This accumulation not only potentially induces organ-specific oxidative stress and inflammatory damage but also presents a significant regulatory blind spot due to the lack of long-term follow-up data regarding their chronic toxicity risks.
Finally, the complexity of multifunctional nano-platforms introduces unprecedented challenges in regulatory science. When a single formulation integrates multiple functionalities—such as targeted delivery, controlled release, imaging capabilities, and even immunomodulation—it transcends the definitional boundaries of conventional pharmaceuticals. Regulatory agencies are consequently faced with the challenges of how to classify such complex products, establish matching quality control standards, and design rational clinical trials that can simultaneously validate the efficacy of their multiple functions alongside their overall safety profile.
In summary, the translational pathway of NDDS-based epigenetic combination therapies from the laboratory to the clinic constitutes a process of addressing systematically interconnected challenges. These challenges permeate the entire continuum-from scalable manufacturing, through in vivo evaluation, to long-term safety-and ultimately converge at the level of regulatory science, demanding the establishment of novel evaluation frameworks for these highly complex therapeutic products. Only through deep interdisciplinary integration, building solid bridges between materials science, epigenetics, nanotoxicology, and clinical medicine, and by systematically overcoming these bottlenecks one by one, can the full potential of these therapies in precision oncology be ultimately realized.
Conclusion
The pivotal role of epigenetic dysregulation in oncogenesis and tumor progression has been increasingly elucidated, with distinct mechanisms of gene activation/silencing through DNA methylation, histone modification, and chromatin remodeling driving therapeutic development. While epigenetic drugs targeting these pathways demonstrate clinical potential, monotherapy limitations necessitate combinatorial approaches requiring nanomedicine-enabled precision delivery. Current challenges in multi-drug regimens—including differential pharmacokinetics, spatiotemporal release coordination, and tumor microenvironment-specific targeting—are being addressed through advanced nano-architectures. Engineered carriers such as liposomes, polymeric nanoparticles, and inorganic frameworks now achieve programmable drug release via stimuli-responsive designs, allowing sequential activation of epigenetic modifiers while maintaining therapeutic payload integrity.
Translating these nanoplatforms into clinical applications still faces significant hurdles. At the industrial level, critical parameters such as nanoparticle stability, drug loading efficiency, and scalability for mass production require substantial optimization. In clinical translation, it is imperative to establish reliable patient stratification strategies and address complex regulatory approval requirements. Notably, recent preclinical studies demonstrating breakthroughs in overcoming tumor heterogeneity and immune evasion underscore the value of interdisciplinary collaboration integrating nanotechnology, epigenetics, and clinical medicine. Furthermore, integrating multi-omics technologies such as proteomics, transcriptomics, and single-cell epigenomics into the development of NDDS combined therapy provides powerful tools for understanding tumor epigenetic heterogeneity and optimizing treatment strategies. These high-throughput technologies can reveal the molecular networks of drug action, identify biomarkers related to drug resistance, and guide the personalized design of nanocarriers, thereby maximizing the precision of treatment.
Looking ahead, emerging technologies are poised to substantially advance epigenetic combination therapies. AI-guided carrier design leverages deep learning of multi-omics data and nanomaterial properties to enable inverse design of NDDS and dynamic optimization of drug combinations. Bioinspired smart delivery platforms, by mimicking biological sensing–response mechanisms and intercellular communication, equip nanocarriers with spatiotemporally precise drug release capabilities and tumor microenvironment adaptivity. Meanwhile, personalized epigenetic strategies integrate patient-specific epigenetic profiles to construct precision treatment platforms. The deep integration of these cutting-edge directions will propel epigenetic therapy toward truly personalized cancer treatment in the clinic.
Accordingly, a systematic research agenda must be pursued with the following priorities: First, it is essential to develop nanocarrier systems that balance functionality with scalable manufacturing, establishing standardized preparation processes. Second, an AI platform integrating multi-dimensional data should be constructed to achieve precise matching of therapeutic strategies. Concurrently, environmentally adaptive intelligent delivery systems must be designed to enable accurate spatiotemporal control over drug release. Furthermore, an epigenetic biomarker-based patient stratification framework needs to be established, alongside collaborative efforts with regulatory agencies to develop corresponding evaluation standards. Ultimately, through the deep integration of materials science, computational science, and clinical medicine, a complete innovation chain—from fundamental discovery to clinical application—should be built. The implementation of this systematic research agenda will strongly propel epigenetic nanomedicine toward realizing its revolutionary potential and pave the way for new approaches in precision cancer therapy.
Acknowledgments
This work was supported by Natural Science Foundation of Shandong Province (ZR2024QH183 and ZR2025QC1717).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Allis CD, Jenuwein T. The molecular hallmarks of epigenetic control. Nat Rev Genet. 2016;17(8):487–500. doi:10.1038/nrg.2016.59
2. Cavalli G, Heard E. Advances in epigenetics link genetics to the environment and disease. Nature. 2019;571(7766):489–499. doi:10.1038/s41586-019-1411-0
3. Bannister AJ, Kouzarides T. Regulation of chromatin by histone modifications. Cell Res. 2011;21(3):381–395. doi:10.1038/cr.2011.22
4. Marmorstein R, Zhou MM. Writers and readers of histone acetylation: structure, mechanism, and inhibition. Cold Spring Harbor Perspectives Biol. 2014;6(7):a018762. doi:10.1101/cshperspect.a018762
5. Zhou VW, Goren A, Bernstein BE. Charting histone modifications and the functional organization of mammalian genomes. Nat Rev Genet. 2011;12(1):7–18. doi:10.1038/nrg2905
6. Arrowsmith CH, Bountra C, Fish PV, Lee K, Schapira M. Epigenetic protein families: a new frontier for drug discovery. Nat Rev Drug Discov. 2012;11(5):384–400. doi:10.1038/nrd3674
7. Jeltsch A. Molecular enzymology of mammalian DNA methyltransferases. Curr Top Microbiol Immunol. 2006;301:203–225. doi:10.1007/3-540-31390-7_7
8. Greer EL, Shi Y. Histone methylation: a dynamic mark in health, disease and inheritance. Nat Rev Genet. 2012;13(5):343–357. doi:10.1038/nrg3173
9. Shi YG, Tsukada Y. The discovery of histone demethylases. Cold Spring Harbor Perspectives Biol. 2013;5(9). doi:10.1101/cshperspect.a017947
10. Bolden JE, Peart MJ, Johnstone RW. Anticancer activities of histone deacetylase inhibitors. Nat Rev Drug Discov. 2006;5(9):769–784. doi:10.1038/nrd2133
11. Filippakopoulos P, Knapp S. Targeting bromodomains: epigenetic readers of lysine acetylation. Nat Rev Drug Discov. 2014;13(5):337–356. doi:10.1038/nrd4286
12. Eissenberg JC, Elgin SC. The HP1 protein family: getting a grip on chromatin. Curr Opin Genet Dev. 2000;10(2):204–210. doi:10.1016/s0959-437x(00)00058-7
13. Terekhanova NV, Karpova A, Liang WW, et al. Epigenetic regulation during cancer transitions across 11 tumour types. Nature. 2023;623(7986):432–441. doi:10.1038/s41586-023-06682-5
14. Dawson MA, Kouzarides T. Cancer epigenetics: from mechanism to therapy. Cell. 2012;150(1):12–27. doi:10.1016/j.cell.2012.06.013
15. Wang N, Ma T, Yu B. Targeting epigenetic regulators to overcome drug resistance in cancers. Signal Transduct Target Ther. 2023;8(1):69. doi:10.1038/s41392-023-01341-7
16. Jones PA, Issa JP, Baylin S. Targeting the cancer epigenome for therapy. Nat Rev Genet. 2016;17(10):630–641. doi:10.1038/nrg.2016.93
17. Szyf M. Targeting DNA methylation in cancer. Ageing Res. Rev. 2003;2(3):299–328.
18. Azad N, Zahnow CA, Rudin CM, Baylin SB. The future of epigenetic therapy in solid tumours--lessons from the past. Nat Rev Clin Oncol. 2013;10(5):256–266. doi:10.1038/nrclinonc.2013.42
19. Lu J, Xue S, Wang Y, et al. Venetoclax and decitabine vs intensive chemotherapy as induction for young patients with newly diagnosed AML. Blood. 2025. doi:10.1182/blood.2024027217
20. DiNardo CD, Jonas BA, Pullarkat V, et al. Azacitidine and venetoclax in previously untreated acute myeloid leukemia. New Engl J Med. 2020;383(7):617–629. doi:10.1056/NEJMoa2012971
21. Azad NS, Gray RJ, Overman MJ, et al. Nivolumab is effective in mismatch repair-deficient noncolorectal cancers: results from arm Z1D-A subprotocol of the NCI-MATCH (EAY131) Study. J Clin Oncol. 2020;38(3):214–222. doi:10.1200/jco.19.00818
22. Shen H, Laird PW. Interplay between the cancer genome and epigenome. Cell. 2013;153(1):38–55. doi:10.1016/j.cell.2013.03.008
23. Mitchell MJ, Billingsley MM, Haley RM, Wechsler ME, Peppas NA, Langer R. Engineering precision nanoparticles for drug delivery. Nat Rev Drug Discov. 2021;20(2):101–124. doi:10.1038/s41573-020-0090-8
24. Hogg SJ, Beavis PA, Dawson MA, Johnstone RW. Targeting the epigenetic regulation of antitumour immunity. Nat Rev Drug Discov. 2020;19(11):776–800. doi:10.1038/s41573-020-0077-5
25. Bellizzi D, D’Aquila P, Scafone T, et al. The control region of mitochondrial DNA shows an unusual CpG and non-CpG methylation pattern. DNA Res. 2013;20(6):537–547. doi:10.1093/dnares/dst029
26. Lawrence RE, Zoncu R. The lysosome as a cellular centre for signalling, metabolism and quality control. Nat Cell Biol. 2019;21(2):133–142. doi:10.1038/s41556-018-0244-7
27. Mattei AL, Bailly N, Meissner A. DNA methylation: a historical perspective. Trends Genet. 2022;38(7):676–707. doi:10.1016/j.tig.2022.03.010
28. Moore LD, Le T, Fan G. DNA methylation and its basic function. Neuropsychopharmacology. 2013;38(1):23–38. doi:10.1038/npp.2012.112
29. Meng H, Cao Y, Qin J, et al. DNA methylation, its mediators and genome integrity. Int J Biol Sci. 2015;11(5):604–617. doi:10.7150/ijbs.11218
30. Martisova A, Holcakova J, Izadi N, Sebuyoya R, Hrstka R, Bartosik M. DNA methylation in solid tumors: functions and methods of detection. Int J Mol Sci. 2021;22(8). doi:10.3390/ijms22084247
31. Lee AV, Nestler KA, Chiappinelli KB. Therapeutic targeting of DNA methylation alterations in cancer. Pharmacol Therapeut. 2024;258:108640. doi:10.1016/j.pharmthera.2024.108640
32. Jansz N. DNA methylation dynamics at transposable elements in mammals. Essays Biochem. 2019;63(6):677–689. doi:10.1042/ebc20190039
33. Klutstein M, Nejman D, Greenfield R, Cedar H. DNA methylation in cancer and aging. Cancer Res. 2016;76(12):3446–3450. doi:10.1158/0008-5472.Can-15-3278
34. Nishiyama A, Nakanishi M. Navigating the DNA methylation landscape of cancer. Trends Genet. 2021;37(11):1012–1027. doi:10.1016/j.tig.2021.05.002
35. Liu J, Huang B, Ding F, Li Y. Environment factors, DNA methylation, and cancer. Environ Geochem Health. 2023;45(11):7543–7568. doi:10.1007/s10653-023-01749-8
36. Bhootra S, Jill N, Shanmugam G, Rakshit S, Sarkar K. DNA methylation and cancer: transcriptional regulation, prognostic, and therapeutic perspective. Med Oncol. 2023;40(2):71. doi:10.1007/s12032-022-01943-1
37. Zaib S, Rana N, Khan I. Histone modifications and their role in epigenetics of cancer. Current Medi Chem. 2022;29(14):2399–2411. doi:10.2174/0929867328666211108105214
38. Audia JE, Campbell RM. Histone modifications and cancer. Cold Spring Harbor Perspectives Biol. 2016;8(4):a019521. doi:10.1101/cshperspect.a019521
39. Neganova ME, Klochkov SG, Aleksandrova YR, Aliev G. Histone modifications in epigenetic regulation of cancer: perspectives and achieved progress. Seminars Cancer Biol. 2022;83:452–471. doi:10.1016/j.semcancer.2020.07.015
40. Yao W, Hu X, Wang X. Crossing epigenetic frontiers: the intersection of novel histone modifications and diseases. Signal Transduction Targeted Ther. 2024;9(1):232. doi:10.1038/s41392-024-01918-w
41. Andrés M, García-Gomis D, Ponte I, Suau P, Roque A. Histone H1 post-translational modifications: update and future perspectives. Int J Mol Sci. 2020;21(16). doi:10.3390/ijms21165941
42. Zhang Y, Sun Z, Jia J, et al. Overview of Histone Modification. Adv Exp Med Biol. 2021;1283:1–16. doi:10.1007/978-981-15-8104-5_1
43. Wang Z, Liu Z, Lv M, Luan Z, Li T, Hu J. Novel histone modifications and liver cancer: emerging frontiers in epigenetic regulation. Clinical Epigenetics. 2025;17(1):30. doi:10.1186/s13148-025-01838-8
44. Hyun K, Jeon J, Park K, Kim J. Writing, erasing and reading histone lysine methylations. Exp Mol Med. 2017;49(4):e324. doi:10.1038/emm.2017.11
45. Sawan C, Herceg Z. Histone modifications and cancer. Adv Genetics. 2010;70:57–85. doi:10.1016/b978-0-12-380866-0.60003-4
46. Nowacka-Zawisza M, Wiśnik E. DNA methylation and histone modifications as epigenetic regulation in prostate cancer (Review). Oncol Rep. 2017;38(5):2587–2596. doi:10.3892/or.2017.5972
47. Taylor EM, Koss B, Davis LE, Tackett AJ. Histone modifications as biomarkers for immunotherapy. Methods Mol. Biol. 2020;2055:213–228. doi:10.1007/978-1-4939-9773-2_10
48. Markouli M, Strepkos D, Piperi C. Impact of histone modifications and their therapeutic targeting in hematological malignancies. Int. J. Mol. Sci. 2022;23(21). doi:10.3390/ijms232113657
49. Shaath H, Vishnubalaji R, Elango R, et al. Long non-coding RNA and RNA-binding protein interactions in cancer: experimental and machine learning approaches. Sem. Cancer Biol. 2022;86(Pt 3):325–345. doi:10.1016/j.semcancer.2022.05.013
50. Anastasiadou E, Jacob LS, Slack FJ. Non-coding RNA networks in cancer. Nat Rev Cancer. 2018;18(1):5–18. doi:10.1038/nrc.2017.99
51. Huang H, Weng H, Chen J. m(6)A modification in coding and non-coding RNAs: roles and therapeutic implications in cancer. Cancer Cell. 2020;37(3):270–288. doi:10.1016/j.ccell.2020.02.004
52. Yao ZT, Yang YM, Sun MM, et al. New insights into the interplay between long non-coding RNAs and RNA-binding proteins in cancer. Cancer Commun. 2022;42(2):117–140. doi:10.1002/cac2.12254
53. An Y, Duan H. The role of m6A RNA methylation in cancer metabolism. Mol Cancer. 2022;21(1):14. doi:10.1186/s12943-022-01500-4
54. Ge Y, Liu T, Wang C, et al. N6-methyladenosine RNA modification and its interaction with regulatory non-coding RNAs in colorectal cancer. RNA Biology. 2021;18(sup2):551–561. doi:10.1080/15476286.2021.1974749
55. Ma S, Chen C, Ji X, et al. The interplay between m6A RNA methylation and noncoding RNA in cancer. J. Hematol. Oncol. 2019;12(1):121. doi:10.1186/s13045-019-0805-7
56. Li J, Wang X, Wang H. RNA modifications in long non-coding RNAs and their implications in cancer biology. Bioorg. Med. Chem. 2024;113:117922. doi:10.1016/j.bmc.2024.117922
57. Wang X, Lu Z, Gomez A, et al. N6-methyladenosine-dependent regulation of messenger RNA stability. Nature. 2014;505(7481):117–120. doi:10.1038/nature12730
58. Qin S, Mao Y, Wang H, Duan Y, Zhao L. The interplay between m6A modification and non-coding RNA in cancer stemness modulation: mechanisms, signaling pathways, and clinical implications. Int J Bio Sci. 2021;17(11):2718–2736. doi:10.7150/ijbs.60641
59. Kadoch C. Diverse compositions and functions of chromatin remodeling machines in cancer. Sci Transl Med. 2019;11(501). doi:10.1126/scitranslmed.aay1018
60. Zhuang K, Wang L, Lu C, et al. Assessment of SWI/SNF chromatin remodeling complex related genes as potential biomarkers and therapeutic targets in pan-cancer. Mol Cancer. 2024;23(1):176. doi:10.1186/s12943-024-02015-w
61. Nebbioso A, Tambaro FP, Dell’Aversana C, Altucci L. Cancer epigenetics: moving forward. PLoS Genetics. 2018;14(6):e1007362. doi:10.1371/journal.pgen.1007362
62. Malone HA, Roberts CWM. Chromatin remodellers as therapeutic targets. Nat Rev Drug Discov. 2024;23(9):661–681. doi:10.1038/s41573-024-00978-5
63. Kaur J, Daoud A, Eblen ST. Targeting chromatin remodeling for cancer therapy. Curr Mol Pharmacol. 2019;12(3):215–229. doi:10.2174/1874467212666190215112915
64. Hao F, Zhang Y, Hou J, Zhao B. Chromatin remodeling and cancer: the critical influence of the SWI/SNF complex. Epigenetics & Chromatin. 2025;18(1):22. doi:10.1186/s13072-025-00590-w
65. Marazzi I, Greenbaum BD, Low DHP, Guccione E. Chromatin dependencies in cancer and inflammation. Nat Rev Mol Cell Biol. 2018;19(4):245–261. doi:10.1038/nrm.2017.113
66. Xiang K, Wang E, Mantyh J, et al. Chromatin remodeling in patient-derived colorectal cancer models. Adv Sci. 2024;11(16):e2303379. doi:10.1002/advs.202303379
67. Zhang FL, Li DQ. Targeting chromatin-remodeling factors in cancer cells: promising molecules in cancer therapy. Int. J. Mol. Sci. 2022;23(21). doi:10.3390/ijms232112815
68. Zhou Z, Li HQ, Liu F. DNA methyltransferase inhibitors and their therapeutic potential. Curr. Top. Med. Chem. 2018;18(28):2448–2457. doi:10.2174/1568026619666181120150122
69. Hu C, Liu X, Zeng Y, Liu J, Wu F. DNA methyltransferase inhibitors combination therapy for the treatment of solid tumor: mechanism and clinical application. Clin Epigenetics. 2021;13(1):166. doi:10.1186/s13148-021-01154-x
70. Lopez M, Gilbert J, Contreras J, Halby L, Arimondo PB. Inhibitors of DNA methylation. Adv Exp Med Biol. 2022;1389:471–513. doi:10.1007/978-3-031-11454-0_17
71. Huang F, Tian T, Wu Y, Che J, Yang H, Dong X. Isocitrate dehydrogenase 2 inhibitors for the treatment of hematologic malignancies: advances and future opportunities. Mini Rev Med Chem. 2021;21(9):1113–1122. doi:10.2174/1389557520666201130102724
72. Mellinghoff IK, Ellingson BM, Touat M, et al. Ivosidenib in isocitrate dehydrogenase 1-mutated advanced glioma. J Clin Oncol. 2020;38(29):3398–3406. doi:10.1200/jco.19.03327
73. Norman M, Yamartino K, Gerstein R, et al. A review of the isocitrate dehydrogenase inhibitors in management of adult patients with AML and MDS. Exp Rev hematol. 2024;17(11):755–767. doi:10.1080/17474086.2024.2422554
74. Abaza Y, Patel AA. Novel therapies in myelodysplastic syndrome: where do venetoclax and isocitrate dehydrogenase inhibitors fit in? Cancer J. 2023;29(3):188–194. doi:10.1097/ppo.0000000000000657
75. Zhao Y, Jiang B, Gu Z, et al. Discovery of cysteine-targeting covalent histone methyltransferase inhibitors. Eur. J. Med. Chem. 2023;246:115028. doi:10.1016/j.ejmech.2022.115028
76. Li B, Xia Y, Mei S, et al. Histone H3K27 methyltransferase EZH2 regulates apoptotic and inflammatory responses in sepsis-induced AKI. Theranostics. 2023;13(6):1860–1875. doi:10.7150/thno.83353
77. Marzochi LL, Cuzziol CI, Nascimento Filho C, et al. Use of histone methyltransferase inhibitors in cancer treatment: a systematic review. Eur J Pharmacol. 2023;944:175590. doi:10.1016/j.ejphar.2023.175590
78. Rugo HS, Jacobs I, Sharma S, et al. The promise for histone methyltransferase inhibitors for epigenetic therapy in clinical oncology: a narrative review. Adv Ther. 2020;37(7):3059–3082. doi:10.1007/s12325-020-01379-x
79. Baby S, Shinde SD, Kulkarni N, Sahu B. Lysine-specific demethylase 1 (LSD1) inhibitors: peptides as an emerging class of therapeutics. ACS Chem Biol. 2023;18(10):2144–2155. doi:10.1021/acschembio.3c00386
80. Li ZR, Gu MZ, Xu X, Zhang JH, Zhang HL, Han C. Promising natural lysine specific demethylase 1 inhibitors for cancer treatment: advances and outlooks. Chin J Nat Med. 2022;20(4):241–257. doi:10.1016/s1875-5364(22)60141-9
81. Song Y, Zhang H, Yang X, Shi Y, Yu B. Annual review of lysine-specific demethylase 1 (LSD1/KDM1A) inhibitors in 2021. Eur. J. Med. Chem. 2022;228:114042. doi:10.1016/j.ejmech.2021.114042
82. Bertoni F, Stathis A. Bromodomain and Extra-Terminal domain inhibitors for lymphoid malignancies. Curr Opin Hematol. 2019;26(4):273–280. doi:10.1097/moh.0000000000000519
83. Nguyen MV, Loof L, Falchook GS. Bromodomain and extra-terminal (BET) domain protein inhibitors for solid tumor cancers. J. Immunother. Precis. Oncol. 2020;3(1):16–22. doi:10.4103/jipo.Jipo_2_20
84. Kulikowski E, Rakai BD, Wong NCW. Inhibitors of bromodomain and extra-terminal proteins for treating multiple human diseases. Med Res Rev. 2021;41(1):223–245. doi:10.1002/med.21730
85. Feng L, Wang G, Chen Y, et al. Dual-target inhibitors of bromodomain and extra-terminal proteins in cancer: a review from medicinal chemistry perspectives. Med Res Rev. 2022;42(2):710–743. doi:10.1002/med.21859
86. Zhao LM, Zhang JH. Histone deacetylase inhibitors in tumor immunotherapy. Curr Med Chem. 2019;26(17):2990–3008. doi:10.2174/0929867324666170801102124
87. Zhao C, Dong H, Xu Q, Zhang Y. Histone deacetylase (HDAC) inhibitors in cancer: a patent review (2017-present). Expert Opin. Ther. Pat. 2020;30(4):263–274. doi:10.1080/13543776.2020.1725470
88. Sonthalia M, Roy BS, Chandrawanshi D, et al. Histone deacetylase inhibitors as antidiabetic agents: advances and opportunities. Eur J Pharmacol. 2022;935:175328. doi:10.1016/j.ejphar.2022.175328
89. Shanmukha KD, Paluvai H, Lomada SK, Gokara M, Kalangi SK. Histone deacetylase (HDACs) inhibitors: clinical applications. Prog. Mol. Biol. Transl. Sci. 2023;198:119–152. doi:10.1016/bs.pmbts.2023.02.011
90. Thurn KT, Thomas S, Moore A, Munster PN. Rational therapeutic combinations with histone deacetylase inhibitors for the treatment of cancer. Future Oncology. 2011;7(2):263–283. doi:10.2217/fon.11.2
91. Huang W, Zhu Q, Shi Z, et al. Dual inhibitors of DNMT and HDAC induce viral mimicry to induce antitumour immunity in breast cancer. Cell Death Discov. 2024;10(1):143. doi:10.1038/s41420-024-01895-7
92. Topper MJ, Vaz M, Chiappinelli KB, et al. Epigenetic therapy ties MYC depletion to reversing immune evasion and treating lung cancer. Cell. 2017;171(6):1284–1300.e21. doi:10.1016/j.cell.2017.10.022
93. Zhu WG, Otterson GA. The interaction of histone deacetylase inhibitors and DNA methyltransferase inhibitors in the treatment of human cancer cells. Curr Med Chem Anticancer Agents. 2003;3(3):187–199. doi:10.2174/1568011033482440
94. Juergens RA, Wrangle J, Vendetti FP, et al. Combination epigenetic therapy has efficacy in patients with refractory advanced non-small cell lung cancer. Cancer Discov. 2011;1(7):598–607. doi:10.1158/2159-8290.Cd-11-0214
95. Chang Y, Guo H, Li X, et al. Development of a first-in-class DNMT1/HDAC inhibitor with improved therapeutic potential and potentiated antitumor immunity. J Med Chem. 2024;67(18):16480–16504. doi:10.1021/acs.jmedchem.4c01310
96. Bauer K, Hauswirth A, Gleixner KV, et al. BRD4 degraders may effectively counteract therapeutic resistance of leukemic stem cells in AML and ALL. Am J Hematol. 2024;99(9):1721–1731. doi:10.1002/ajh.27385
97. Zhang J, Wu L, Wang C, Xie X, Han Y. Research progress of long non-coding RNA in tumor drug resistance: a new paradigm. Drug Des Devel Ther. 2024;18:1385–1398. doi:10.2147/dddt.S448707
98. Solta A, Boettiger K, Kovács I, et al. Entinostat enhances the efficacy of chemotherapy in small cell lung cancer through s-phase arrest and decreased base excision repair. Clin Cancer Res. 2023;29(22):4644–4659. doi:10.1158/1078-0432.Ccr-23-1795
99. Ding K, Liu H, Yang H, et al. A prospective Phase 2 study of combination epigenetic therapy against relapsed/refractory peripheral T cell lymphoma. Med. 2024;5(11):1393–1401.e2. doi:10.1016/j.medj.2024.07.007
100. Clozel T, Yang S, Elstrom RL, et al. Mechanism-based epigenetic chemosensitization therapy of diffuse large B-cell lymphoma. Cancer Discov. 2013;3(9):1002–1019. doi:10.1158/2159-8290.Cd-13-0117
101. Fischer C, Leithner K, Wohlkoenig C, et al. Panobinostat reduces hypoxia-induced cisplatin resistance of non-small cell lung carcinoma cells via HIF-1α destabilization. Mol Cancer. 2015;14:4. doi:10.1186/1476-4598-14-4
102. Xue K, Gu JJ, Zhang Q, et al. Vorinostat, a histone deacetylase (HDAC) inhibitor, promotes cell cycle arrest and re-sensitizes rituximab- and chemo-resistant lymphoma cells to chemotherapy agents. J. Cancer Res. Clin. Oncol. 2016;142(2):379–387. doi:10.1007/s00432-015-2026-y
103. Levitzki A, Klein S. My journey from tyrosine phosphorylation inhibitors to targeted immune therapy as strategies to combat cancer. Proc. Natl. Acad. Sci. USA. 2019;116(24):11579–11586. doi:10.1073/pnas.1816012116
104. Zhong L, Li Y, Xiong L, et al. Small molecules in targeted cancer therapy: advances, challenges, and future perspectives. Signal Transduct Target Therpy. 2021;6(1):201. doi:10.1038/s41392-021-00572-w
105. Vander Velde R, Yoon N, Marusyk V, et al. Resistance to targeted therapies as a multifactorial, gradual adaptation to inhibitor specific selective pressures. Nature Comm. 2020;11(1):2393. doi:10.1038/s41467-020-16212-w
106. Chen MC, Chen CH, Wang JC, et al. The HDAC inhibitor, MPT0E028, enhances erlotinib-induced cell death in EGFR-TKI-resistant NSCLC cells. Cell Death & Dis. 2013;4(9):e810. doi:10.1038/cddis.2013.330
107. Lee TG, Jeong EH, Kim SY, Kim HR, Kim CH. The combination of irreversible EGFR TKIs and SAHA induces apoptosis and autophagy-mediated cell death to overcome acquired resistance in EGFR T790M-mutated lung cancer. Int J Cancer. 2015;136(11):2717–2729. doi:10.1002/ijc.29320
108. Konopleva M, Pollyea DA, Potluri J, et al. Efficacy and biological correlates of response in a phase II study of venetoclax monotherapy in patients with acute myelogenous leukemia. Cancer Discov. 2016;6(10):1106–1117. doi:10.1158/2159-8290.Cd-16-0313
109. Bose P, Gandhi V, Konopleva M. Pathways and mechanisms of venetoclax resistance. Leukemia & Lymphoma. 2017;58(9):1–17. doi:10.1080/10428194.2017.1283032
110. Tsao T, Shi Y, Kornblau S, et al. Concomitant inhibition of DNA methyltransferase and BCL-2 protein function synergistically induce mitochondrial apoptosis in acute myelogenous leukemia cells. Annals of Hematol. 2012;91(12):1861–1870. doi:10.1007/s00277-012-1537-8
111. DiNardo CD, Pratz K, Pullarkat V, et al. Venetoclax combined with decitabine or azacitidine in treatment-naive, elderly patients with acute myeloid leukemia. Blood. 2019;133(1):7–17. doi:10.1182/blood-2018-08-868752
112. Heath H, Mogol AN, Santaliz Casiano A, Zuo Q, Madak-Erdogan Z. Targeting systemic and gut microbial metabolism in ER(+) breast cancer. Trends Endocrinol Metab. 2024;35(4):321–330. doi:10.1016/j.tem.2023.12.008
113. Llinas-Bertran A, Bellet-Ezquerra M, Seoane JA. Epigenetic control of cancer cell dormancy and awakening in endocrine therapy resistance. Cancer Discov. 2024;14(5):704–706. doi:10.1158/2159-8290.Cd-24-0282
114. Culig Z, Puhr M. Androgen receptor-interacting proteins in prostate cancer development and therapy resistance. Am J Pathol. 2024;194(3):324–334. doi:10.1016/j.ajpath.2023.12.003
115. Fischetti I, Botti L, Sulsenti R, et al. Combined therapy targeting AR and EZH2 curbs castration-resistant prostate cancer enhancing anti-tumor T-cell response. Epigenomics. 2024;16(9):653–670. doi:10.2217/epi-2023-0374
116. Baell JB, Leaver DJ, Hermans SJ, et al. Inhibitors of histone acetyltransferases KAT6A/B induce senescence and arrest tumour growth. Nature. 2018;560(7717):253–257. doi:10.1038/s41586-018-0387-5
117. Yan F, Li J, Milosevic J, et al. KAT6A and ENL form an epigenetic transcriptional control module to drive critical leukemogenic gene-expression programs. Cancer Discovery. 2022;12(3):792–811. doi:10.1158/2159-8290.Cd-20-1459
118. Mukohara T, Park YH, Sommerhalder D, et al. Inhibition of lysine acetyltransferase KAT6 in ER(+)HER2(-) metastatic breast cancer: a phase 1 trial. Nature Medicine. 2024;30(8):2242–2250. doi:10.1038/s41591-024-03060-0
119. Fioravanti R, Stazi G, Zwergel C, Valente S, Mai A. Six years (2012-2018) of researches on catalytic EZH2 inhibitors: the boom of the 2-pyridone compounds. Chemical Record (New York, NY). 2018;18(12):1818–1832. doi:10.1002/tcr.201800091
120. Morel KL, Sheahan AV, Burkhart DL, et al. EZH2 inhibition activates a dsRNA-STING-interferon stress axis that potentiates response to PD-1 checkpoint blockade in prostate cancer. Nature Cancer. 2021;2(4):444–456. doi:10.1038/s43018-021-00185-w
121. Ge R, Wang Z, Montironi R, et al. Epigenetic modulations and lineage plasticity in advanced prostate cancer. Annals of Oncology: Official Journal of the European Society for Medical Oncology. 2020;31(4):470–479. doi:10.1016/j.annonc.2020.02.002
122. Topper MJ, Vaz M, Marrone KA, Brahmer JR, Baylin SB. The emerging role of epigenetic therapeutics in immuno-oncology. Nature Reviews Clinical Oncology. 2020;17(2):75–90. doi:10.1038/s41571-019-0266-5
123. Jones PA, Ohtani H, Chakravarthy A, De Carvalho DD. Epigenetic therapy in immune-oncology. Nature Reviews Cancer. 2019;19(3):151–161. doi:10.1038/s41568-019-0109-9
124. Cañadas I, Thummalapalli R, Kim JW, et al. Tumor innate immunity primed by specific interferon-stimulated endogenous retroviruses. Nature Medicine. 2018;24(8):1143–1150. doi:10.1038/s41591-018-0116-5
125. Liu Z, Ren Y, Weng S, Xu H, Li L, Han X. A new trend in cancer treatment: the combination of epigenetics and immunotherapy. Frontiers in Immunology. 2022;13:809761. doi:10.3389/fimmu.2022.809761
126. Aspeslagh S, Morel D, Soria JC, Postel-Vinay S. Epigenetic modifiers as new immunomodulatory therapies in solid tumours. Annals of Oncology: Official Journal of the European Society for Medical Oncology. 2018;29(4):812–824. doi:10.1093/annonc/mdy050
127. Liu Y, Wang C, Li X, et al. Improved clinical outcome in a randomized phase II study of anti-PD-1 camrelizumab plus decitabine in relapsed/refractory Hodgkin lymphoma. Journal for Immunotherapy of Cancer. 2021;9(4). doi:10.1136/jitc-2021-002347
128. Gounder MM, Zhu G, Roshal L, et al. Immunologic correlates of the abscopal effect in a SMARCB1/INI1-negative poorly differentiated chordoma after EZH2 inhibition and radiotherapy. Clinical Cancer Research: an Official Journal of the American Association for Cancer Research. 2019;25(7):2064–2071. doi:10.1158/1078-0432.Ccr-18-3133
129. Larsen BD, Benada J, Yung PYK, et al. Cancer cells use self-inflicted DNA breaks to evade growth limits imposed by genotoxic stress. Science (New York, NY). 2022;376(6592):476–483. doi:10.1126/science.abi6378
130. Wang L, Dou X, Chen S, et al. YTHDF2 inhibition potentiates radiotherapy antitumor efficacy. Cancer Cell. 2023;41(7):1294–1308.e8. doi:10.1016/j.ccell.2023.04.019
131. Graham K, Unger E. Overcoming tumor hypoxia as a barrier to radiotherapy, chemotherapy and immunotherapy in cancer treatment. International Journal of Nanomedicine. 2018;13:6049–6058. doi:10.2147/ijn.S140462
132. Das C, Adhikari S, Bhattacharya A, et al. Epigenetic-metabolic interplay in the DNA damage response and therapeutic resistance of breast cancer. Cancer Research. 2023;83(5):657–666. doi:10.1158/0008-5472.Can-22-3015
133. Zhao H, Gao S, Han Y, et al. Conversion of Ku80 K568 crotonylation to SUMOylation facilitates DNA non-homologous end joining and cancer radioresistance. Signal Transduction and Targeted Therapy. 2025;10(1):127. doi:10.1038/s41392-025-02210-1
134. Jiang Y, Qian X, Shen J, et al. Local generation of fumarate promotes DNA repair through inhibition of histone H3 demethylation. Nature Cell Biology. 2015;17(9):1158–1168. doi:10.1038/ncb3209
135. Wang EC, Min Y, Palm RC, et al. Nanoparticle formulations of histone deacetylase inhibitors for effective chemoradiotherapy in solid tumors. Biomaterials. 2015;51:208–215. doi:10.1016/j.biomaterials.2015.02.015
136. Shah S, Dhawan V, Holm R, Nagarsenker MS, Perrie Y. Liposomes: advancements and innovation in the manufacturing process. Advanced Drug Delivery Reviews. 2020;154-155:102–122. doi:10.1016/j.addr.2020.07.002
137. Guimarães D, Cavaco-Paulo A, Nogueira E. Design of liposomes as drug delivery system for therapeutic applications. International Journal of Pharmaceutics. 2021;601:120571. doi:10.1016/j.ijpharm.2021.120571
138. Liu G, Hou S, Tong P, Li J. Liposomes: preparation, characteristics, and application strategies in analytical chemistry. Critical Reviews in Analytical Chemistry. 2022;52(2):392–412. doi:10.1080/10408347.2020.1805293
139. Tang M, Lozano Hernandez L, Reginald-Opara JN, et al. Zebularine suppressed gemcitabine-induced senescence and improved the cellular and plasma pharmacokinetics of gemcitabine, augmented by liposomal co-delivery. International Journal of Pharmaceutics. 2021;602:120659. doi:10.1016/j.ijpharm.2021.120659
140. Lin TT, Xiong W, Chen GH, et al. Epigenetic-based combination therapy and liposomal codelivery overcomes osimertinib-resistant NSCLC via repolarizing tumor-associated macrophages. Acta Pharmacologica Sinica. 2024;45(4):867–878. doi:10.1038/s41401-023-01205-4
141. He Y, Fang Y, Zhang M, et al. Remodeling “cold” tumor immune microenvironment via epigenetic-based therapy using targeted liposomes with in situ formed albumin Corona. Acta Pharmaceutica Sinica B. 2022;12(4):2057–2073. doi:10.1016/j.apsb.2021.09.022
142. Tu K, Yu Y, Wang Y, et al. Combination of chidamide-mediated epigenetic modulation with immunotherapy: boosting tumor immunogenicity and response to PD-1/PD-L1 blockade. ACS Applied Materials & Interfaces. 2021;13(33):39003–39017. doi:10.1021/acsami.1c08290
143. Liang S, Liu M, Mu W, et al. Nano-regulator inhibits tumor immune escape via the “two-way regulation” epigenetic therapy strategy. Advanced Science. 2024;11(9):e2305275. doi:10.1002/advs.202305275
144. Sun H, Yu T, Li X, et al. Second near-infrared photothermal-amplified immunotherapy using photoactivatable composite nanostimulators. Journal of Nanobiotechnology. 2021;19(1):433. doi:10.1186/s12951-021-01197-5
145. Banasaz S, Morozova K, Ferrentino G, Scampicchio M. Encapsulation of lipid-soluble bioactives by nanoemulsions. Molecules. 2020;25(17). doi:10.3390/molecules25173966
146. Cunha S, Costa CP, Moreira JN, Sousa Lobo JM, Silva AC. Using the quality by design (QbD) approach to optimize formulations of lipid nanoparticles and nanoemulsions: a review. Nanomedicine: Nanotechnology, Biology, and Medicine. 2020;28:102206. doi:10.1016/j.nano.2020.102206
147. Hörmann K, Zimmer A. Drug delivery and drug targeting with parenteral lipid nanoemulsions - A review. Journal of Controlled Release: Official Journal of the Controlled Release Society. 2016;223:85–98. doi:10.1016/j.jconrel.2015.12.016
148. Kim B, Pena CD, Auguste DT. Targeted lipid nanoemulsions encapsulating epigenetic drugs exhibit selective cytotoxicity on CDH1(-)/FOXM1(+) triple negative breast cancer cells. Molecular Pharmaceutics. 2019;16(5):1813–1826. doi:10.1021/acs.molpharmaceut.8b01065
149. Nicolas J, Couvreur P. [Polymer nanoparticles for the delivery of anticancer drug]. Medecine Sciences: M/S. 2017;33(1):11–17. doi:10.1051/medsci/20173301003 Les nanoparticules polymères pour la délivrance de principes actifs anticancéreux
150. Deirram N, Zhang C, Kermaniyan SS, Johnston APR, Such GK. pH-responsive polymer nanoparticles for drug delivery. Macromolecular Rapid Communications. 2019;40(10):e1800917. doi:10.1002/marc.201800917
151. Ferreira Soares DC, Domingues SC, Viana DB, Tebaldi ML. Polymer-hybrid nanoparticles: current advances in biomedical applications. Biomedicine & Pharmacotherapy = Biomedecine & Pharmacotherapie. 2020;131:110695. doi:10.1016/j.biopha.2020.110695
152. Li Z, Liu P, Chen W, et al. Hypoxia-cleavable and specific targeted nanomedicine delivers epigenetic drugs for enhanced treatment of breast cancer and bone metastasis. Journal of Nanobiotechnology. 2023;21(1):221. doi:10.1186/s12951-023-01939-7
153. Ledezma DK, Balakrishnan PB, Cano-Mejia J, et al. Indocyanine green-nexturastat A-PLGA nanoparticles combine photothermal and epigenetic therapy for melanoma. Nanomaterials. 2020;10(1). doi:10.3390/nano10010161
154. Hu N, Li W, Hong Y, et al. A PD1 targeted nano-delivery system based on epigenetic alterations of T cell responses in the treatment of gastric cancer. Molecular Therapy Oncolytics. 2022;24:148–159. doi:10.1016/j.omto.2021.12.006
155. Su X, Wang Z, Li L, et al. Lipid-polymer nanoparticles encapsulating doxorubicin and 2’-deoxy-5-azacytidine enhance the sensitivity of cancer cells to chemical therapeutics. Molecular Pharmaceutics. 2013;10(5):1901–1909. doi:10.1021/mp300675c
156. Ghosh B, Biswas S. Polymeric micelles in cancer therapy: state of the art. Journal of Controlled Release: Official Journal of the Controlled Release Society. 2021;332:127–147. doi:10.1016/j.jconrel.2021.02.016
157. Deshmukh AS, Chauhan PN, Noolvi MN, et al. Polymeric micelles: basic research to clinical practice. International Journal of Pharmaceutics. 2017;532(1):249–268. doi:10.1016/j.ijpharm.2017.09.005
158. Yokoyama M. Polymeric micelles as drug carriers: their lights and shadows. Journal of Drug Targeting. 2014;22(7):576–583. doi:10.3109/1061186x.2014.934688
159. Jain A, Bhardwaj K, Bansal M. Polymeric micelles as drug delivery system: recent advances, approaches, applications and patents. Current Drug Safety. 2024;19(2):163–171. doi:10.2174/1574886318666230605120433
160. Naz A, Cui Y, Collins CJ, Thompson DH, Irudayaraj J. PLGA-PEG nano-delivery system for epigenetic therapy. Biomedicine & Pharmacotherapy = Biomedecine & Pharmacotherapie. 2017;90:586–597. doi:10.1016/j.biopha.2017.03.093
161. Xiong H, Ma X, Wang X, et al. Inspired epigenetic modulation synergy with adenosine inhibition elicits pyroptosis and potentiates cancer immunotherapy. Advanced Functional Materials. 2021;31(20). doi:10.1002/adfm.202100007
162. Ding F, Liu J, Ai K, et al. Simultaneous activation of pyroptosis and cGAS-STING pathway with epigenetic/ photodynamic nanotheranostic for enhanced tumor photoimmunotherapy. Advanced materials (Deerfield Beach, Fla). 2024;36(7):e2306419. doi:10.1002/adma.202306419
163. Zhao L, Huang C, Zheng R. Photodynamic amplified immune checkpoint-blockade therapy of self-delivery bioregulator via epigenetic reprogramming. Chemical Engineering Journal. 2023;453(1). doi:10.1016/j.cej.2022.139729, et.
164. da Silva Santos S, Igne Ferreira E, Giarolla J. Dendrimer Prodrugs. Molecules. 2016;21(6). doi:10.3390/molecules21060686
165. Rodríguez-Acosta GL, Hernández-Montalbán C, Vega-Razo MFS, Castillo-Rodríguez IO, Martínez-García M. Polymer-dendrimer hybrids as carriers of anticancer agents. Current Drug Targets. 2022;23(4):373–392. doi:10.2174/1389450122666210906121803
166. Sharma AK, Gothwal A, Kesharwani P, Alsaab H, Iyer AK, Gupta U. Dendrimer nanoarchitectures for cancer diagnosis and anticancer drug delivery. Drug Discovery Today. 2017;22(2):314–326. doi:10.1016/j.drudis.2016.09.013
167. Kim Y, Park EJ, Na DH. Recent progress in dendrimer-based nanomedicine development. Archives of Pharmacal Research. 2018;41(6):571–582. doi:10.1007/s12272-018-1008-4
168. Zong H, Shah D, Selwa K, et al. Design and evaluation of tumor-specific dendrimer epigenetic therapeutics. ChemistryOpen. 2015;4(3):335–341. doi:10.1002/open.201402141
169. Kaundal B, Kushwaha AC, Srivastava AK, Karmakar S, Choudhury SR. A non-viral nano-delivery system targeting epigenetic methyltransferase EZH2 for precise acute myeloid leukemia therapy. Journal of Materials Chemistry B. 2020;8(37):8658–8670. doi:10.1039/d0tb01177k
170. van Niel G, Raposo G, D’Angelo G. Shedding light on the cell biology of extracellular vesicles. Nature Reviews Molecular Cell Biology. 2018;19(4):213–228. doi:10.1038/nrm.2017.125
171. Gupta D, Zickler AM, El Andaloussi S. Dosing extracellular vesicles. Advanced Drug Delivery Reviews. 2021;178:113961. doi:10.1016/j.addr.2021.113961
172. Marar C, Starich B, Wirtz D. Extracellular vesicles in immunomodulation and tumor progression. Nature Immunology. 2021;22(5):560–570. doi:10.1038/s41590-021-00899-0
173. Nowak M, Górczyńska J, Kołodzińska K, Rubin J, Choromańska A. Extracellular vesicles as drug transporters. International Journal of Molecular Sciences. 2023;24(12). doi:10.3390/ijms241210267
174. You Q, Wang F, Du R, et al. m(6) A reader YTHDF1-Targeting engineered small extracellular vesicles for gastric cancer therapy via epigenetic and immune regulation. Advanced Materials (Deerfield Beach, Fla). 2023;35(8):e2204910. doi:10.1002/adma.202204910
175. Zhai Y, Wang J, Lang T, et al. T lymphocyte membrane-decorated epigenetic nanoinducer of interferons for cancer immunotherapy. Nature Nanotechnology. 2021;16(11):1271–1280. doi:10.1038/s41565-021-00972-7
176. Du R, You Q, Liu J, Wang C, Zhu L, Yang Y. Dual-functional extracellular vesicles enable synergistic treatment via m6A reader YTHDF1-targeting epigenetic regulation and chemotherapy. 2023;16(12):13309–13321. Nano Research. doi:10.1007/s12274-023-6105-0.
177. Lin Y, Li S, Xiao Z, et al. Epigenetic inhibition assisted chemotherapeutic treatment of lung cancer based on artificial exosomes. Pharmacological Research. 2021;171:105787. doi:10.1016/j.phrs.2021.105787
178. Luk BT, Zhang L. Cell membrane-camouflaged nanoparticles for drug delivery. Journal of Controlled Release: Official Journal of the Controlled Release Society. 2015;220(Pt B):600–607. doi:10.1016/j.jconrel.2015.07.019
179. Smith D, Artursson P, Avdeef A, et al. Passive lipoidal diffusion and carrier-mediated cell uptake are both important mechanisms of membrane permeation in drug disposition. Molecular Pharmaceutics. 2014;11(6):1727–1738. doi:10.1021/mp400713v
180. Gao T, Sang X, Huang X, et al. Macrophage-camouflaged epigenetic nanoinducers enhance chemoimmunotherapy in triple negative breast cancer. Acta Pharmaceutica Sinica B. 2023;13(10):4305–4317. doi:10.1016/j.apsb.2022.11.018
181. Zhang HT, Peng R, Chen S, et al. Versatile nano-PROTAC-induced epigenetic reader degradation for efficient lung cancer therapy. Advanced Science. 2022;9(29):e2202039. doi:10.1002/advs.202202039
182. Wang T, Zhang H, Qiu W, Han Y, Liu H, Li Z. Biomimetic nanoparticles directly remodel immunosuppressive microenvironment for boosting glioblastoma immunotherapy. Bioactive Materials. 2022;16:418–432. doi:10.1016/j.bioactmat.2021.12.029
183. Jiang X, He C, Lin W. Supramolecular metal-based nanoparticles for drug delivery and cancer therapy. Current Opinion in Chemical Biology. 2021;61:143–153. doi:10.1016/j.cbpa.2021.01.005
184. Zhang C, Yan L, Gu Z, Zhao Y. Strategies based on metal-based nanoparticles for hypoxic-tumor radiotherapy. Chemical Science. 2019;10(29):6932–6943. doi:10.1039/c9sc02107h
185. Liu Y, Zhang P, Li F, et al. Metal-based NanoEnhancers for Future Radiotherapy: radiosensitizing and Synergistic Effects on Tumor Cells. Theranostics. 2018;8(7):1824–1849. doi:10.7150/thno.22172
186. Klębowski B, Depciuch J, Parlińska-Wojtan M, Baran J. Applications of noble metal-based nanoparticles in medicine. International Journal of Molecular Sciences. 2018;19(12). doi:10.3390/ijms19124031
187. Kaundal B, Karmakar S, Roy Choudhury S. Mitochondria-targeting nano therapy altering IDH2-mediated EZH2/EZH1 interaction as precise epigenetic regulation in glioblastoma. Biomaterials Science. 2022;10(18):5301–5317. doi:10.1039/d1bm02006d
188. Song Y, Zhang L, Wang Y, et al. A bimetallic metal-organic-framework-based biomimetic nanoplatform enhances anti-leukemia immunity via synchronizing DNA demethylation and RNA hypermethylation. Advanced Materials (Deerfield Beach, Fla). 2023;35(16):e2210895. doi:10.1002/adma.202210895
189. Wang G, Yan J, Tian H, et al. Dual-epigenetically relieving the MYC-correlated immunosuppression via an advanced nano-radiosensitizer potentiates cancer immuno-radiotherapy. Advanced Materials (Deerfield Beach, Fla). 2024;36(19):e2312588. doi:10.1002/adma.202312588
190. Wang G, Li B, Tian H, et al. A metal–phenolic nanocoordinator launches radiotherapeutic cancer pyroptosis through an epigenetic mechanism. Advanced Functional Materials. 2023;33(23). doi:10.1002/adfm.202213425
191. Chen YC, Huang XC, Luo YL, Chang YC, Hsieh YZ, Hsu HY. Non-metallic nanomaterials in cancer theranostics: a review of silica- and carbon-based drug delivery systems. Science and Technology of Advanced Materials. 2013;14(4):044407. doi:10.1088/1468-6996/14/4/044407
192. Yanar F, Carugo D, Zhang X. Hybrid nanoplatforms comprising organic nanocompartments encapsulating inorganic nanoparticles for enhanced drug delivery and bioimaging applications. Molecules. 2023;28(15). doi:10.3390/molecules28155694
193. Mattoussi H, Rotello VM. Inorganic nanoparticles in drug delivery. Advanced Drug Delivery Reviews. 2013;65(5):605–606. doi:10.1016/j.addr.2013.04.012
194. Tang F, Li L, Chen D. Mesoporous silica nanoparticles: synthesis, biocompatibility and drug delivery. Advanced Materials (Deerfield Beach, Fla). 2012;24(12):1504–1534. doi:10.1002/adma.201104763
195. Yuan Y, Liu J, Yu X, et al. Tumor-targeting pH/redox dual-responsive nanosystem epigenetically reverses cancer drug resistance by co-delivering doxorubicin and GCN5 siRNA. Acta Biomaterialia. 2021;135:556–566. doi:10.1016/j.actbio.2021.09.002
196. Gu M, Toh TB, Hooi L, Lim JJ, Zhang X, Chow EK. Nanodiamond-mediated delivery of a g9a inhibitor for hepatocellular carcinoma therapy. ACS Applied Materials & Interfaces. 2019;11(49):45427–45441. doi:10.1021/acsami.9b16323
197. Gutierrez AM, Frazar EM, XK MV, Paul P, Hilt JZ. Hydrogels and hydrogel nanocomposites: enhancing healthcare through human and environmental treatment. Adv Healthcare Mat. 2022;11(7):e2101820. doi:10.1002/adhm.202101820
198. Cheng Q, Hao A, Xing P. Stimulus-responsive luminescent hydrogels: design and applications. Adv Colloid Interface Sci. 2020;286:102301. doi:10.1016/j.cis.2020.102301
199. Ho TC, Chang CC, Chan HP, et al. Hydrogels: properties and applications in biomedicine. Molecules. 2022;27(9). doi:10.3390/molecules27092902
200. Narayanaswamy R, Torchilin VP. Hydrogels and their applications in targeted drug delivery. Molecules. 2019;24(3). doi:10.3390/molecules24030603
201. Khan F, Atif M, Haseen M, et al. Synthesis, classification and properties of hydrogels: their applications in drug delivery and agriculture. J Mat Chem B. 2022;10(2):170–203. doi:10.1039/d1tb01345a
202. Ruan H, Hu Q, Wen D, et al. A dual-bioresponsive drug-delivery depot for combination of epigenetic modulation and immune checkpoint blockade. Adv Mat. 2019;31(17):1806957.
203. Ji X, Guo D, Ma J, et al. Epigenetic remodeling hydrogel patches for multidrug-resistant triple-negative breast cancer. Adv Mat. 2021;33(18):e2100949. doi:10.1002/adma.202100949
204. Ashwani PV, Gopika G, Arun Krishna KV, Jose J, John F, George J. Stimuli-responsive and multifunctional nanogels in drug delivery. Chem Biodiversity. 2023;20(11):e202301009. doi:10.1002/cbdv.202301009
205. Vashist A, Perez Alvarez G, Andion Camargo V, et al. Recent advances in nanogels for drug delivery and biomedical applications. Biomat Sci. 2024;12(23):6006–6018. doi:10.1039/d4bm00224e
206. Vijayaraghavalu S, Labhasetwar V. Nanogel-mediated delivery of a cocktail of epigenetic drugs plus doxorubicin overcomes drug resistance in breast cancer cells. Drug Delivery Transl Res. 2018;8(5):1289–1299. doi:10.1007/s13346-018-0556-y
207. Dumontet C, Reichert JM, Senter PD, Lambert JM, Beck A. Antibody-drug conjugates come of age in oncology. Nat Rev Drug Discovery. 2023;22(8):641–661. doi:10.1038/s41573-023-00709-2
208. Colombo R, Tarantino P, Rich JR, LoRusso PM, de Vries EGE. The journey of antibody-drug conjugates: lessons learned from 40 years of development. Cancer Discovery. 2024;14(11):2089–2108. doi:10.1158/2159-8290.Cd-24-0708
209. Drago JZ, Modi S, Chandarlapaty S. Unlocking the potential of antibody-drug conjugates for cancer therapy. Nat Rev Clin Oncol. 2021;18(6):327–344. doi:10.1038/s41571-021-00470-8
210. Tarantino P, Carmagnani Pestana R, Corti C, et al. Antibody-drug conjugates: smart chemotherapy delivery across tumor histologies. CA. 2022;72(2):165–182. doi:10.3322/caac.21705
211. Cini E, Faltoni V, Petricci E, et al. Antibody drug conjugates (ADCs) charged with HDAC inhibitor for targeted epigenetic modulation. Chem Sci. 2018;9(31):6490–6496. doi:10.1039/c7sc05266a
212. Milazzo FM, Vesci L, Anastasi AM, et al. ErbB2 targeted epigenetic modulation: anti-tumor efficacy of the ADC trastuzumab-HDACi ST8176AA1. Front Oncol. 2019;9:1534. doi:10.3389/fonc.2019.01534
213. Wang T, Li M, Wei R, et al. Small molecule-drug conjugates emerge as a new promising approach for cancer treatment. Mol Pharmaceut. 2024;21(3):1038–1055. doi:10.1021/acs.molpharmaceut.3c01049
214. Zhuang C, Guan X, Ma H, Cong H, Zhang W, Miao Z. Small molecule-drug conjugates: a novel strategy for cancer-targeted treatment. Eur J Med Chem. 2019;163:883–895. doi:10.1016/j.ejmech.2018.12.035
215. Abbasi YF, Bera H, Cun D, Yang M. Recent advances in pH/enzyme-responsive polysaccharide-small-molecule drug conjugates as nanotherapeutics. Carbohydrate Polymers. 2023;312:120797. doi:10.1016/j.carbpol.2023.120797
216. Zhang J, Hu F, Aras O, Chai Y, An F. Small molecule-drug conjugates: opportunities for the development of targeted anticancer drugs. ChemMedChem. 2024;19(11):e202300720. doi:10.1002/cmdc.202300720
217. Xu S, Zhu X, Huang W, Zhou Y, Yan D. Supramolecular cisplatin-vorinostat nanodrug for overcoming drug resistance in cancer synergistic therapy. J Controlled Release. 2017;266:36–46. doi:10.1016/j.jconrel.2017.09.007
218. Wang L, Zhang C, Hong Y, et al. Integrating epigenetic modulators in nanofibers for synergistic gastric cancer therapy via epigenetic reprogramming. Nano Lett. 2021;21(1):298–307. doi:10.1021/acs.nanolett.0c03665
219. Ye J, Hou B, Chen F, et al. Bispecific prodrug nanoparticles circumventing multiple immune resistance mechanisms for promoting cancer immunotherapy. Acta Pharmaceutica Sinica B. 2022;12(6):2695–2709. doi:10.1016/j.apsb.2021.09.021
220. Luo Z, Wan Z, Ren P, et al. In situ formation of fibronectin-enriched protein corona on epigenetic nanocarrier for enhanced synthetic lethal therapy. Adv Sci. 2024;11(19):e2307940. doi:10.1002/advs.202307940
221. Kaundal B, Srivastava AK, Dev A, Mohanbhai SJ, Karmakar S, Roy Choudhury S. Nanoformulation of EPZ011989 attenuates EZH2-c-Myb epigenetic interaction by proteasomal degradation in acute myeloid leukemia. Mol Pharmaceut. 2020;17(2):604–621. doi:10.1021/acs.molpharmaceut.9b01071
222. Yang CL, Chao YJ, Wang HC, et al. Local ablation of gastric cancer by reconstituted apolipoprotein B lipoparticles carrying epigenetic drugs. Nanomedicine. 2021;37:102450. doi:10.1016/j.nano.2021.102450
223. Tian Y, Wang X, Zhao S, et al. JQ1-loaded polydopamine nanoplatform inhibits c-MYC/programmed cell death ligand 1 to enhance photothermal therapy for triple-negative breast cancer. ACS Appl Mat Interfaces. 2019;11(50):46626–46636. doi:10.1021/acsami.9b18730
224. Alavi M, Hamidi M. Passive and active targeting in cancer therapy by liposomes and lipid nanoparticles. Drug Metab Personalized Ther. 2019;34(1). doi:10.1515/dmpt-2018-0032
225. Bertrand N, Wu J, Xu X, Kamaly N, Farokhzad OC. Cancer nanotechnology: the impact of passive and active targeting in the era of modern cancer biology. Adv Drug Delivery Rev. 2014;66:2–25. doi:10.1016/j.addr.2013.11.009
226. Shinde VR, Revi N, Murugappan S, Singh SP, Rengan AK. Enhanced permeability and retention effect: a key facilitator for solid tumor targeting by nanoparticles. Photodiagn Photodynamic Ther. 2022;39:102915. doi:10.1016/j.pdpdt.2022.102915
227. Subhan MA, Yalamarty SSK, Filipczak N, Parveen F, Torchilin VP. Recent advances in tumor targeting via EPR effect for cancer treatment. J Personalized Med. 2021;11(6). doi:10.3390/jpm11060571
228. Siafaka PI, Üstündağ Okur N, Karavas E, Bikiaris DN. Surface modified multifunctional and stimuli responsive nanoparticles for drug targeting: current status and uses. Int J Mol Sci. 2016;17(9). doi:10.3390/ijms17091440
229. Huynh NT, Roger E, Lautram N, Benoît JP, Passirani C. The rise and rise of stealth nanocarriers for cancer therapy: passive versus active targeting. Nanomedicine. 2010;5(9):1415–1433. doi:10.2217/nnm.10.113
230. Xu M, Qi Y, Liu G, Song Y, Jiang X, Du B. Size-dependent in vivo transport of nanoparticles: implications for delivery, targeting, and clearance. ACS Nano. 2023;17(21):20825–20849. doi:10.1021/acsnano.3c05853
231. Yu W, Liu R, Zhou Y, Gao H. Size-tunable strategies for a tumor targeted drug delivery system. ACS Central Sci. 2020;6(2):100–116. doi:10.1021/acscentsci.9b01139
232. Liu X, Li H, Jin Q, Ji J. Surface tailoring of nanoparticles via mixed-charge monolayers and their biomedical applications. Small. 2014;10(21):4230–4242. doi:10.1002/smll.201401440
233. Albanese A, Tang PS, Chan WC. The effect of nanoparticle size, shape, and surface chemistry on biological systems. Annual Rev Biomed Eng. 2012;14:1–16. doi:10.1146/annurev-bioeng-071811-150124
234. Raj S, Khurana S, Choudhari R, et al. Specific targeting cancer cells with nanoparticles and drug delivery in cancer therapy. Seminars Cancer Biol. 2021;69:166–177. doi:10.1016/j.semcancer.2019.11.002
235. Bazak R, Houri M, El Achy S, Kamel S, Refaat T. Cancer active targeting by nanoparticles: a comprehensive review of literature. J Cancer Res Clin Oncol. 2015;141(5):769–784. doi:10.1007/s00432-014-1767-3
236. Vyas D, Patel M, Wairkar S. Strategies for active tumor targeting-an update. Eur J Pharmacol. 2022;915:174512. doi:10.1016/j.ejphar.2021.174512
237. Lugano R, Ramachandran M, Dimberg A. Tumor angiogenesis: causes, consequences, challenges and opportunities. Cell Mol Life Sci. 2020;77(9):1745–1770. doi:10.1007/s00018-019-03351-7
238. Li H, Li S, Lin Y, et al. Artificial exosomes mediated spatiotemporal-resolved and targeted delivery of epigenetic inhibitors. J Nanobiotechnol. 2021;19(1):364. doi:10.1186/s12951-021-01107-9
239. Tran TH, Choi JY, Ramasamy T, et al. Hyaluronic acid-coated solid lipid nanoparticles for targeted delivery of vorinostat to CD44 overexpressing cancer cells. Carbohydrate Polymers. 2014;114:407–415. doi:10.1016/j.carbpol.2014.08.026
240. Overchuk M, Zheng G. Overcoming obstacles in the tumor microenvironment: recent advancements in nanoparticle delivery for cancer theranostics. Biomaterials. 2018;156:217–237. doi:10.1016/j.biomaterials.2017.10.024
241. Estrella V, Chen T, Lloyd M, et al. Acidity generated by the tumor microenvironment drives local invasion. Cancer Res. 2013;73(5):1524–1535. doi:10.1158/0008-5472.Can-12-2796
242. Swartz MA, Lund AW. Lymphatic and interstitial flow in the tumour microenvironment: linking mechanobiology with immunity. Nat Rev Cancer. 2012;12(3):210–219. doi:10.1038/nrc3186
243. Jiang W, Von Roemeling CA, Chen Y, et al. Designing nanomedicine for immuno-oncology. Nat Biomed Eng. 2017;1(2):0029.
244. Lou J, Zhang L, GJAT Z. Advancing cancer immunotherapies with nanotechnology. Adv Therapeut. 2019;2(4):1800128.
245. Kanamala M, Wilson WR, Yang M, Palmer BD, Wu Z. Mechanisms and biomaterials in pH-responsive tumour targeted drug delivery: a review. Biomaterials. 2016;85:152–167. doi:10.1016/j.biomaterials.2016.01.061
246. Wang Y, Shim MS, Levinson NS, Sung HW, Xia Y. Stimuli-responsive materials for controlled release of theranostic agents. Adv Funct Mater. 2014;24(27):4206–4220. doi:10.1002/adfm.201400279
247. Smith SA, Selby LI, Johnston APR, Such GK. The endosomal escape of nanoparticles: toward more efficient cellular delivery. Bioconjugate Chem. 2019;30(2):263–272. doi:10.1021/acs.bioconjchem.8b00732
248. Mrksich K, Padilla MS, Mitchell MJ. Breaking the final barrier: evolution of cationic and ionizable lipid structure in lipid nanoparticles to escape the endosome. Adv Drug Delivery Rev. 2024;214:115446. doi:10.1016/j.addr.2024.115446
249. Panyam J, Zhou WZ, Prabha S, Sahoo SK, Labhasetwar V. Rapid endo-lysosomal escape of poly(DL-lactide-co-glycolide) nanoparticles: implications for drug and gene delivery. Faseb J. 2002;16(10):1217–1226. doi:10.1096/fj.02-0088com
250. Cupic KI, Rennick JJ, Johnston AP, Such GK. Controlling endosomal escape using nanoparticle composition: current progress and future perspectives. Nanomedicine. 2019;14(2):215–223. doi:10.2217/nnm-2018-0326
251. Peng Q, Li H, Deng Q, et al. Hybrid artificial cell-mediated epigenetic inhibition in metastatic lung cancer. J Colloid Interface Sci. 2021;603:319–332. doi:10.1016/j.jcis.2021.06.066
252. Mohammadinejad R, Moosavi MA, Tavakol S, et al. Necrotic, apoptotic and autophagic cell fates triggered by nanoparticles. Autophagy. 2019;15(1):4–33. doi:10.1080/15548627.2018.1509171
253. Zeng J, Shirihai OS, Grinstaff MW. Degradable nanoparticles restore lysosomal ph and autophagic flux in lipotoxic pancreatic beta cells. Adv Healthcare Mat. 2019;8(12):e1801511. doi:10.1002/adhm.201801511
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.