Back to Journals » International Journal of Nanomedicine » Volume 20
Advancing Cancer Immunotherapy Using Lipid Nanoparticle-Based Approaches
Authors Prazeres PHDM ![]() , Costa da Silva GH, Azevedo GV
, Costa da Silva GH, Azevedo GV ![]() , Alves da Silva NJ
, Alves da Silva NJ ![]() , Carvalho Costa PA, Da Silva WN, Lobo AO
, Carvalho Costa PA, Da Silva WN, Lobo AO ![]() , Guimaraes PPG
, Guimaraes PPG
Received 17 May 2025
Accepted for publication 27 September 2025
Published 8 October 2025 Volume 2025:20 Pages 12283—12305
DOI https://doi.org/10.2147/IJN.S541061
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Prof. Dr. RDK Misra
Pedro Henrique Dias Moura Prazeres,1,* Gabriel Henrique Costa da Silva,1,* Gabriel Vieira Azevedo,1 Natalia Jordana Alves da Silva,1 Pedro Augusto Carvalho Costa,1 Walison Nunes Da Silva,1 Anderson Oliveira Lobo,2 Pedro Pires Goulart Guimaraes1
1Department of Physiology and Biophysics, Institute of Biological Sciences, Federal University of Minas Gerais, Belo Horizonte, MG, 31270-901, Brazil; 2Department of Materials Engineering, Federal University of Piauí, Teresina, PI, 64049-550, Brazil
*These authors contributed equally to this work
Correspondence: Pedro Pires Goulart Guimaraes, Department of Physiology and Biophysics, Federal University of Minas Gerais, Belo Horizonte, MG, Brazil, Email [email protected]
Abstract: Cancer immunotherapy, including adoptive cell therapies, cancer vaccines, and cytokine-based therapies, have revolutionized targeted approaches in the treatment of different tumors. However, the broader application of immunotherapies, such as for engineered T cells expressing a chimeric antigen receptor (CAR-T cells), remains limited by challenges in production, systemic toxicity, and inefficient delivery, especially in solid tumors. Recent advances in nucleic acid delivery technologies, notably ionizable lipid nanoparticles (LNP), offer promising solutions to overcome these barriers. LNPs have shown potential in delivering messenger RNA (mRNA), and DNA for the generation of CAR-T cells, cancer vaccines, bispecific antibodies, and cytokine-based immunotherapies. The clinical success of LNP-based platforms in mRNA COVID-19 vaccines and interference RNA therapies for genetic disorders further validates their effectiveness in gene delivery, highlighting LNPs as versatile carriers for therapeutic nucleic acids. Furthermore, LNPs can be optimized for off-the-shelf formulations, enabling personalized treatments targeting specific patient needs. In this review, we highlight the role of LNP platforms in advancing mRNA and DNA delivery for cancer immunotherapy. We explore their potential to improve CAR-T cell production, advance cancer vaccines, and support the development of bispecific antibody- and cytokine-based therapies, ultimately paving the way for more effective, scalable, and accessible immunotherapeutic strategies.
Keywords: cancer immunotherapy, adoptive cell therapy, CAR-T cells, cancer vaccines, lipid nanoparticles, gene delivery
Introduction
Cancer in its various forms remains the second major cause of mortality, also representing a great challenge to the public healthcare systems worldwide.1–3 The increase in incidence and mortality of this disease relates to contemporary diets and lifestyle as well as environmental changes.1 Taken together, this outlines the need for investment in basic and clinical research that focus on developing preventive and therapeutic approaches against this disease.
Different immunotherapies have emerged as effective alternatives to conventional treatments like chemotherapy, offering more targeted and enhanced responses across various cancers.4,5 Among these, chimeric antigen receptor (CAR)-T cell therapy has revolutionized the treatment of hematological tumors by harnessing engineered T cells to selectively target tumor antigens.6 However, the broader application of CAR-T cells, particularly in solid tumors, is limited by challenges in manufacturing, antigen selection, and the immunosuppressive tumor microenvironment.7 Clinical efforts are ongoing to extend CAR-T therapy to solid tumors such as hepatocarcinoma, pediatric tumors, and neuroblastoma.6,8
In parallel, other immunotherapy strategies, such as cytokine therapies, cancer vaccines, and bispecific antibodies (BsAbs), seek to activate the patient’s immune system without the need for cell transfer. These approaches aim to stimulate immune effector cells, reverse T cell exhaustion, and promote tumor cell killing. While promising, their clinical efficacy is often hindered by systemic toxicity, short half-life, and poor biodistribution, which limit their ability to effectively reach and act within the tumor microenvironment. Overcoming these delivery barriers remains critical to unlocking the full potential of these immunotherapeutic modalities.
To overcome the limitations faced by current immunotherapeutic strategies, lipid nanoparticles (LNP) can be applied as versatile gene delivery systems in multiple contexts.9,10 These platforms enable a fast and effective delivery of nucleic acids to cells and tissues, both in vivo and ex vivo without affecting cell viability. Compared to viral vectors currently used in CAR-T cell production, LNPs do not integrate the genetic material into the host genome, thereby reducing safety concerns and significantly shortening the engineering timeline. Moreover, LNPs can be tailored to deliver different nucleic acid cargos, offering alternatives to protein-based delivery of agents like cytokines, and enabling the generation of potent cancer vaccines and bsAbs directly in the body. These features make LNP platforms attractive for both off-the-shelf therapies targeting different tumor antigens and personalized approaches based on individual patient profiles.11–13 In this review, we explore the expanding role of LNP platforms in cancer immunotherapy, with a focus on their application in messenger RNA (mRNA), and DNA-based delivery strategies to enhance their efficacy, specificity, and accessibility of treatments such as CAR-T cells, cancer vaccines, cytokines, and bsAbs.
Overview of Lipid Nanoparticles
Lipid Nanoparticles
Nanoparticles or nanomaterials are defined as materials with external dimensions ranging from 1 to 200nm and have been employed in antitumor therapy for several years.14 These materials can serve as drug delivery systems, increasing permeability in the tumor microenvironment and circulation time.15 The importance of these nanoparticles is substantial, attracting investments and indicating a significant advancement in the treatment of various diseases.16
LNPs are included in the class of nanomaterials, among other types of nanoparticles, which are characterized by a spherical shape containing at least one lipid bilayer surrounding an internal aqueous compartment.16 These LNPs are widely used as delivery systems because of their low-complexity production process, are biocompatible, have good biodistribution17,18 and are easy to optimize depending on the load to be delivered.19 Additionally, LNPs have the capacity to accommodate large payloads within them, justifying their use in therapies already approved by the US regulatory agency (FDA).20 Recently, in response to the COVID-19 pandemic, LNPs platforms have been extensively employed by companies such as Moderna and Pfizer in the production of mRNA vaccines.17,21
LNPs are formed from a multicomponent lipid system, composed of an ionizable lipid, a phospholipid, cholesterol, and a polyethylene glycol (PEG)-lipid conjugate13 (Figure 1A). Ionizable cationic lipids are typically defined by the presence of a tertiary amine that remains unprotonated under neutral conditions and becomes positively charged in acidic pH environments.22 These lipids are primarily responsible for the encapsulation of nucleic acids within LNPs. During LNP formation, ionizable lipids are positively charged, enabling electrostatic interactions with the negatively charged phosphate groups of nucleic acids, which facilitates their complexation within the nanoparticle.23 These lipids also mediate endosomal escape and the subsequent release of the nucleic acid cargo into the cell. Endosomal acidification leads to lipid protonation, promoting interaction with endogenous lipids and destabilization of the endosomal membrane, thereby releasing the genetic material into the cytoplasm.22 Although the exact mechanism has not been fully elucidated, this remains a widely accepted hypothesis.
 |
Figure 1 Development and use of LNPs. (A) LNPs are prepared using a microfluidic mixing method that combines an aqueous phase containing the nucleic acid and an ethanolic phase containing the lipid mix. (B) LNPs loaded with one of various types of nucleic acids protect their cargo from nuclease degradation, evade immune detection and efficiently deliver it to the cells. LNPs are endocytosed by the cells, leading to the formation of an endosome containing the LNP within the cytoplasm. Following endosomal escape and LNP destabilization, the nucleic acid is released into the cytoplasm. Once released, it may be directly translated (mRNA), processed for gene silencing (siRNA), or transported to the nucleus for transcription (DNA). |
Phospholipids are helper lipids that enhance the stability of the system by contributing to the structure of the lipid bilayer. They assist in the efficient encapsulation of nucleic acids and some of those possess fusogenic properties, which improve the transfection efficiency of LNPs.24 Phospholipids such as 1,2-distearoyl-sn-glycero-3-phosphocholine (DSPC) and 1,2-dioleoyl-sn-glycero-3-phosphoethanolamine (DOPE) have been widely used in lipid formulations for years. DSPC, in particular, has been clinically used for siRNA delivery in the drug Onpattro®,25 as well as in mRNA vaccines against SARS-CoV-2 (Moderna mRNA-1273 and Pfizer/BioNTech BNT162b).26
Cholesterol is another helper lipid, primarily responsible for providing structural integrity and directly influencing the membrane fluidity of LNPs.27 It stabilizes the LNP structure by filling the gaps between phospholipids, which also contributes to enhanced stability in the presence of serum proteins.28 This stabilization is critical to ensuring nanoparticle integrity during systemic circulation.
The final component of lipid nanoparticles is PEG-lipids, which influence several characteristics of LNPs, including particle size and population homogeneity.29 They prevent particle aggregation, impact circulation and biodistribution profiles in vivo,30,31 and also play an important role in modulating the immune response.32,33 These lipids create a hydrophilic steric barrier on the nanoparticle surface, contributing to the stability.34 It has been demonstrated that nanoparticles lacking PEG-lipids are unstable and polydisperse.
Therefore, each component plays a critical role in the architecture and performance of LNPs, making them a versatile platform with a high potential for modulation. This versatility helps explain their success in both clinical and preclinical settings, particularly in gene therapy, vaccines, and cancer immunotherapy. Given their complexity, detailed characterization of LNPs is essential to ensure reproducibility and safety.
Characterization and Manufacturing
Lipid nanoparticles can be produced through various methods, among which microfluidic mixing stands out as the most commonly employed approach.35 This method has gained prominence due to its scalability, reproducibility, and precise control over operational parameters.36 Microfluidic mixing is based on the simultaneous injection of an ethanolic phase containing the lipids and an aqueous buffered phase containing the nucleic acids (Figure 1A). These phases meet within microchannels where chaotic mixing occurs, promoting the self-assembly and complexation of components to form LNPs.37 Several factors influence the characteristics of LNPs, including the buffer pH, lipid-to-nucleic acid ratio, and flow rate parameters during mixing.36–38
The characterization of LNPs is essential to ensure reproducibility and to predict their performance.39 Key parameters include particle size and polydispersity index (PDI), typically measured by dynamic light scattering (DLS) or nanoparticle tracking analysis (NTA).40,41 Zeta potential is used to estimate the colloidal stability of the formulations.42 Encapsulation efficiency and nucleic acid content are usually assessed through fluorescence-based assays, such as RiboGreen (for RNA) and PicoGreen (for DNA).43,44 Meanwhile, transmission electron microscopy (TEM) or cryo-TEM are employed to analyze the morphology and internal structure of LNPs.13,45 An important aspect of LNP quality control is their stability under different storage conditions, which can affect particle size, structural integrity, nucleic acid retention, and transfection capability.46 Sterility control and endotoxin levels are also critical to prevent adverse immune responses.47,48 These parameters are crucial for ensuring the clinical translation and therapeutic application of LNP formulations.
Recent advances in LNP manufacturing have increasingly leveraged systematic optimization strategies, notably Design of Experiments (DOE). DOE enables the simultaneous evaluation of multiple variables, avoiding the inefficiency of one-factor-at-a-time approaches.49 This framework has been applied to modulate lipid molar ratios for high-throughput LNP library generation,50 to investigate structural variants of specific components—such as PEG-lipids—and assess their in vivo performance,51 and to fine-tune process parameters, including flow rate and pH, for control over key physicochemical attributes.49 The resulting statistical models facilitate the concurrent optimization of hydrodynamic diameter, polydispersity index, and encapsulation efficiency.39 By reducing experimental burden while maximizing information yield, DOE expedites formulation development and enhances predictive control over LNP performance.52
In parallel, machine learning (ML) is emerging as a powerful tool for LNP design, enabling the extraction of predictive relationships from large datasets.53 ML approaches have been used to forecast mRNA vaccine efficacy from compositional data, revealing critical substructural features of ionizable lipids that influence immunogenicity.54,55 The integration of DOE with ML holds particular promise, coupling statistically rigorous experimentation with adaptive predictive modeling. Together, these computational and statistical tools are poised to become foundational to the next generation of rational, application-tailored LNP development, reducing both cost and development timelines.56
Overall, stringent quality parameters and accelerated development timelines are essential for advancing LNP technologies, particularly in the context of nucleic acid delivery for next-generation immunotherapies.
LNP Pharmacokinetics and Pharmacodynamics
Understanding the in vivo biodistribution and metabolism of LNPs is determinant for the development of optimized and safe therapies. In cancer, a major challenge is to engineer delivery systems that can efficiently reach the tumor microenvironment (TME) and perform their therapeutic function.57 Additionally, reducing toxicity to healthy tissues and adverse effects caused by immunotherapies is a central motivation to employ LNPs,10 as they can be finely tuned to accumulate in tumor sites, while reducing off-target delivery to the organs.16 To achieve these goals, some strategies can be explored, including rigorous studies of LNP pharmacokinetics, through variations in administration routes and tailoring lipid composition and molar ratios.
In the context of cancer vaccines, intramuscular and subcutaneous administration routes are the most commonly used.58 These routes aim to deliver LNPs to the lymph nodes and spleen and, consequently, increase the transfection in antigen-presenting cells (APCs).59,60 This is critical, as antigen presentation by APCs promotes strong induced anti-tumor immunity.61 On the other hand, intravenous injection is commonly used for systemic drug delivery, although this approach promotes a great accumulation of LNPs in the liver.62 This occurs mainly due to the interaction of LNPs with plasma proteins, such as apolipoprotein E (ApoE), which directs them to hepatocytes expressing low-density lipoprotein receptors and facilitates their internalization.63 Notably, tailoring LNP composition modifies the protein corona, changing the biodistribution of formulations.64 To overcome hepatic tropism, high-throughput screening methods have been used to identify LNPs with enhanced extrahepatic delivery.65 For example, DNA barcode technology has been applied to quantify the nucleic acid delivery of large LNP libraries and find those with differential biodistribution.66 Most recently, this method was used to screen 94 unique LNPs, enabling the identification of formulations that preferentially deliver mRNA to head and neck squamous cell carcinoma, by systematically varying the lipidic composition and molar ratios.67
For many years, passive accumulation of nanoparticles in tumors was attributed to the enhanced permeability and retention (EPR) effect, caused by abnormal growth of endothelial tissue, containing intercellular gaps in blood vessels, and reduced lymphatic drainage.16,57 However, new discoveries demonstrate that just a small fraction of nanoparticles reach the TME through passive transport. Instead, active trans-endothelial transport is the main mechanism for their accumulation.68 Understanding these mechanisms provides valuable insights for rational development of effective nanosystems. In this context, active targeting approaches, such as LNP surface functionalization with antibodies69,70 or peptides,71 have gained significant attention and will be further discussed in subsequent sections.
Beyond active and passive targeting, alternative administration routes have also been investigated. Intratumoral injection provides localized therapy that can reduce systemic toxicity, while triggering systemic anti-tumor immune responses capable of reducing tumor burden at distant sites.72–74 Similarly, intranasal and inhalable administration have emerged as promising strategies for targeting primary or metastatic tumors in the lungs.75 Early pre-clinical studies using nebulized nanoparticles for intranasal delivery demonstrated regression of lung metastases in an MC38 cell model.23
Once absorbed and distributed, LNPs enter cells through endocytosis. This process can involve different pathways, including protein-mediated mechanisms, such as clathrin and claveolin-dependent endocytosis, as well as independent pathways, like macropinocytosis.76 Initially, LNPs are maintained in early endosomes, which subsequently mature and acidify.77 The decrease of endosomal pH protonates the ionizable cationic lipids which interact with negatively charged lipids in bilayer, destabilizing the membrane and promoting nucleic acid release into the cytoplasm.78 This process, known as endosomal escape, must occur before endosomes mature into lysosomes, where both LNPs and their cargo would be degraded. Importantly, the apparent pKa of the LNP is a key physicochemical parameter for this process, with several studies reporting that values between 6.0 and 6.5 are optimal for efficient endosomal escape and enhanced transfection.79
After overcoming all barriers to efficient cargo delivery, metabolism and excretion are the last pharmacokinetic processes that influence LNP function. Metabolism plays a fundamental role to guarantee the safe elimination of LNP-based therapies. However, if it occurs too fast, therapeutic efficacy may be compromised.80 Thus, developing LNPs with an optimal balance between safe metabolism and efficiency is essential. One factor that strongly influences this balance is the presence of PEG-lipids. While non-PEGylated LNPs are rapidly cleared from circulation, PEGylated LNPs exhibit reduced clearance, resulting in longer circulation times, extended half-life and improved chances of reaching target organs or TME.31,81 It is important to emphasize that most lipids used in clinical formulations are designed to be biodegradable. For example, the ionizable lipids used in the Pfizer and Moderna vaccines (ALC-0315 and SM-102, respectively) contain ester groups that undergo hydrolysis, allowing efficient elimination.82
In conclusion, the efficacy of LNP-based immunotherapies is multifactorial and demands robust basic research. Advances in the understanding of cell biology and the mechanisms of LNP interactions with biological systems are critical for the design of optimized formulations. Moreover, the ability to modulate the biodistribution of LNPs has revolutionized the field of drug delivery and continues to open new opportunities for safer and more effective therapies.
mRNA vs DNA Delivery
The choice of the genetic cargo to be encapsulated within LNPs should take into account the specific properties and limitations of each type of molecule, depending on the intended therapeutic application.83 For immunotherapies to be effective, nucleic acids must overcome multiple biological barriers, both extracellular and intracellular.16 When administered in vivo, nucleic acids (specially mRNA) must be protected from degradation by endonucleases present in physiological fluids.26 Then, LNPs must reach the target tissue, be internalized by the target cells, and subsequently escape from the endosome.84 Once in the cytoplasm, mRNA is immediately available for translation, whereas plasmid DNA (pDNA) must first overcome an additional barrier by crossing the nuclear membrane85 (Figure 1B).
mRNA offers rapid and direct expression kinetics, as translation begins as soon as the molecule is delivered into the cytoplasm, typically peaking between 6 to 24 hours.86 This expression is short-lived and transient, decreasing progressively as the mRNA is degraded,87 which is desirable to control the effects of cellular therapies such as CAR-T, and limit systemic effects of cytokines.70
pDNA, on the other hand, has slower expression kinetics—it must enter the nucleus to be transcribed into mRNA before translation can occur in the cytoplasm, with peak expression generally occurring 24 to 72 hours after delivery.86 pDNA enables prolonged antigen expression, making it an excellent option for vaccines, as extended exposure can promote a stronger and more durable immune response.88
In terms of stability, pDNA exhibits greater chemical and structural stability compared to mRNA. This allows for longer-term storage at milder temperatures, or even at room temperature.29,46 mRNA is more susceptible to hydrolysis and requires stricter storage conditions to preserve its integrity, typically at −20°C to −80°C.89 This difference significantly impacts the logistics of distribution, storage, and the overall cost of nucleic acid-based therapies, directly affecting their accessibility and global reach. Insights gained from comparing mRNA and pDNA delivery serve as a foundation for evaluating the broader landscape of gene delivery, particularly the advantages and limitations of LNPs relative to other platforms (Table 1).90–96
 |
Table 1 RNA vs DNA Delivery |
LNPs Vs Other Gene Delivery Systems
Viral vectors, electroporation, and LNPs are the most commonly used approaches to gene delivery. Viral vectors, such as lentiviruses and adenoviruses, are widely employed due to their high transduction efficiency and potential for long-term expression.59 However, viral vectors provide high transduction efficiency and prolonged expression —permanent in the case of lentiviruses due to genomic integration, and as long-lasting episomes with adenoviruses— but carry risks of insertional mutagenesis, increased immunogenicity, limitations on repeat dosing, and high production costs.97–100 In contrast, LNPs do not pose a risk of genomic integration, have a lower probability of triggering undesired immune responses, offer greater scalability, and involve lower production costs,26,101 however, LNP-mediated transgene expression is inherently transient—reaching maximal protein output 24 hours post-administration and declining to subtherapeutic levels within one week—thus necessitating repeated dosing or sustained-release formulations to sustain therapeutic efficacy.87,102,103
Electroporation is a frequently used method for ex vivo cell engineering, such as the generation of CAR-T cells.104,105 Although it demonstrates high efficacy, it is often accompanied by significant cytotoxicity and reduced cell viability,106 which limits its broader clinical application. In comparison, LNPs exhibit very low toxicity, making them suitable for both ex vivo and in vivo strategies.70,106–108
Cationic polymers, such as polyethyleneimine (PEI), are also used as non-viral vectors,108 offering advantages like low production costs and ease of optimization.109 However, they are often associated with high toxicity and only moderate in vivo performance,109,110 limitations that are effectively addressed by LNPs. While polymeric nanoparticles offer advantages in formulation stability and customizability, they generally show lower gene delivery efficiency and more variable in vivo outcomes. In terms of immunogenicity, LNPs elicit strong innate immune responses that can be beneficial in vaccine settings, whereas polymeric systems may provoke unpredictable immune activation depending on their charge and composition.111,112
In summary, when comparing different platforms for genetic material delivery, LNPs emerge as a highly promising solution, combining safety, adaptability, and efficiency. This positions them as one of the main alternatives to viral vectors, even overcoming other non-viral methods such as electroporation and polymer-based platforms, especially for emerging applications in cancer immunotherapy. The use of nanoparticles has increasingly stood out as an effective and safe approach. Moreover, this methodology offers remarkable flexibility, allowing for application in both in vivo and ex vivo delivery strategies. This enables greater personalization of treatments and broadens the potential uses of the technique.
Advantages of in vivo Delivery Over ex vivo Manipulation
The use of lipid nanoparticles for in vivo delivery of nucleic acids offers a novel perspective compared to traditional ex vivo cell manipulation, which is used in the production of CAR-T cells.104,105,107 In this approach, mononuclear cells must be harvested from the peripheral blood of a patient or a donor, followed by T cell isolation, commonly using anti CD3 microbeads, and activation. After that, cells are genetically modified through viral (lentivirus or retrovirus) or non-viral (vectors, such as electroporation or LNPs) systems. The engineered T cells are then expanded in vitro using media supplemented with cytokines such as IL-2, IL-7 and IL-15 to promote clonal proliferation and maturation. Throughout this workflow, rigorous quality-control measures (including flow cytometric assessment of CAR expression, cell viability assays, sterility testing, and functional cytotoxicity assays) are performed to ensure both potency and safety.107,113 Finally, the modified cells are reinfused into the patient (Figure 2A). Although effective, this process is costly, labor-intensive, time-consuming, and requires sophisticated infrastructure and strict closed-system protocols under Good Manufacturing Practices (GMP) conditions.114
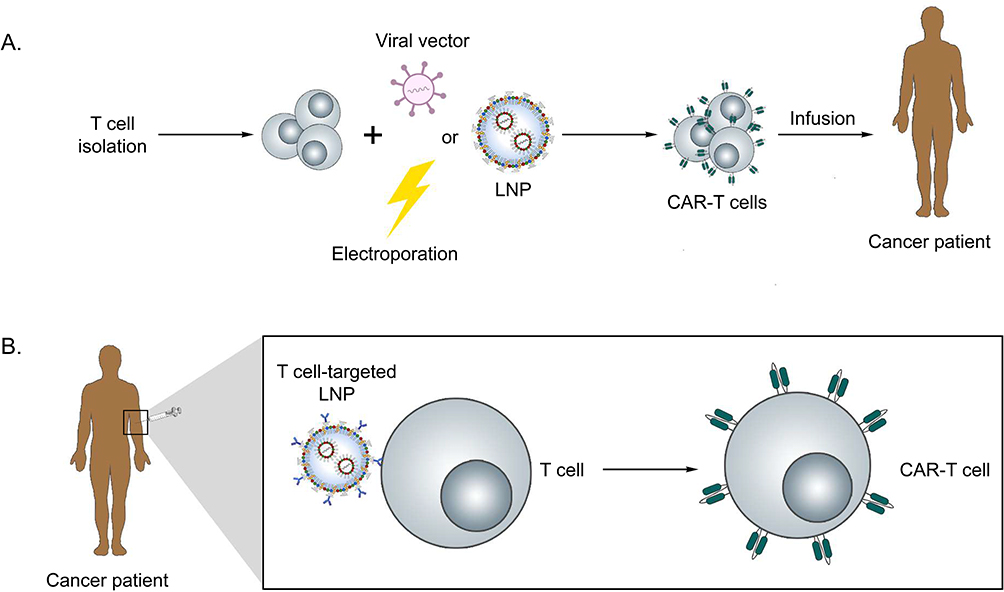 |
Figure 2 CAR-T cell therapy production cycle. (A) Ex vivo CAR-T cell engineering: The patient’s peripheral blood is collected and lymphocytes are isolated. T cells are genetically modified ex vivo using various methods, such as LNPs, viral vectors, or electroporation. Following genetic modification, CAR-T cells are expanded in vitro, and finally infused intravenously into the patient. (B) In vivo CAR-T engineering: LNPs are administered intravenously, enabling the transfection of T cells to induce CAR expression in vivo. |
In contrast, in vivo delivery bypasses the need for cell isolation and manipulation (Figure 2B). Lipid nanoparticles carrying mRNA or pDNA can be directly injected into the patient, overcoming biological barriers and allowing the genetic engineering process to occur within the body.70,115 These LNPs can be surface-functionalized with anti-CD3 antibodies or T cell–specific ligands, thereby enabling preferential targeting of circulating T lymphocytes following systemic administration.116 Upon receptor-mediated endocytosis, the particles deliver mRNA into the cytosol, leading to CAR expression directly in endogenous T cells.117,118 Current efforts are increasingly directed toward refining targeting ligands, optimizing lipid composition, and incorporating controlled mRNA release systems, all of which represent active avenues of investigation aimed at enhancing therapeutic efficacy and prolonging the durability of responses.119–121 This approach drastically reduces processing time, overall cost, and the need for large-scale infrastructure, enabling more affordable and scalable therapies and expanding access across different socioeconomic groups. Finally, it is important to emphasize that ex vivo culture conditions differ significantly from the homeostatic in vivo environment. Therefore, it is crucial to investigate how each step of the manufacturing process impacts T cell quality, including potential induction of cellular exhaustion.122,123
As previously mentioned, the use of lipid nanoparticles has profoundly transformed the field of molecular delivery, providing innovative solutions across a wide range of therapeutic applications. LNPs constitute a highly versatile and clinically validated platform capable of efficiently encapsulating and delivering diverse nucleic acids, with substantial translational potential. Beyond serving as general delivery vehicles, they act as key enablers of next-generation therapies by directly addressing major challenges in immune modulation, targeted gene transfer, and in vivo cell engineering. Collectively, these properties position LNPs as a pivotal link between nucleic acid delivery technologies and the rapidly evolving field of immunotherapy, where their ability to reprogram immune responses can be harnessed to advance cancer treatment and expand therapeutic accessibility. In the following sections, we will discuss recent advances in immunotherapies and highlight the applications of LNPs in these strategies.
Cancer Immunotherapies Overview
Immunotherapies against cancer have become a hallmark in the treatment of solid and hematological tumors. These therapies aim at inducing or improving immune cell responses against tumor cells. The first line of immunotherapy was based on cytokines, such as interferon alpha (IFN-α) and interleukin 2 (IL-2), which resulted in severe adverse effects, despite their success against leukemia and melanoma.124–126 In the late 1990s, monoclonal antibodies emerged as a new line of treatment, promising a more targeted approach against lymphoma (Rituximab) and lymphocytic leukemia (Ibrutinib).35,127 These advances led to the development of immune checkpoint blockers (ICBs), which revolutionized the treatment of metastatic melanoma and other solid tumors by targeting CTLA-4 (Ipilimumab) and PD-1 (Pembrolizumab).128–130 The use of ICBs facilitates the effective recognition and elimination of tumors, and promotes the activation of immune cells, especially T cells.129,130
Over time, targeted approaches have been developed to render immunotherapies more effective. In this context, an “acquired immunity” approach known as adoptive cell therapy (ACT) was developed.131 ACT comprises three forms of adoptive transfer of T cells, which include tumor infiltrating lymphocytes (TILs), T cells engineered to express a T cell receptor (TCR) or a CAR6 (Figure 3). These different types of cellular immunotherapy have shown significant results in the clinic against leukemias and other neoplasms, and their applications keep increasing with new technologies.132,133
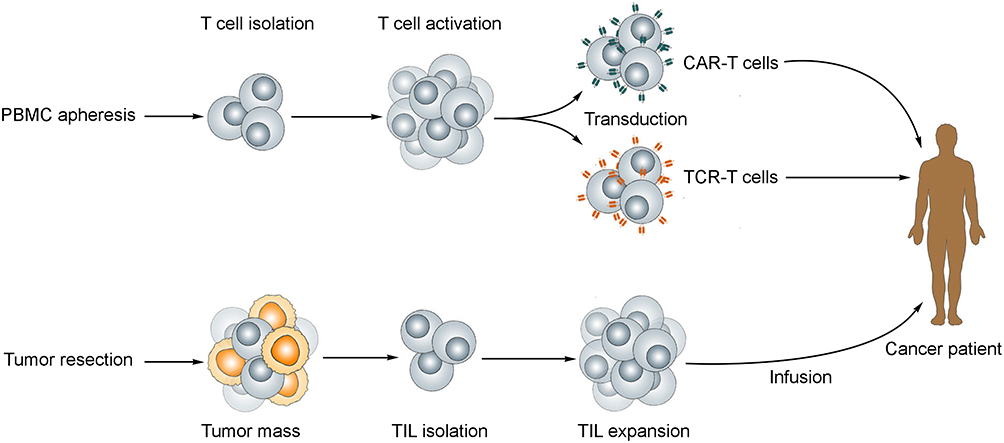 |
Figure 3 Adoptive T cell therapy. For the production of CAR-T and TCR-T cells (top), T lymphocytes are collected from the patient’s peripheral blood and transduced to express the desired receptor. TILs (bottom) are isolated from a resected tumor mass, and infused back to the patients following ex vivo activation and expansion. |
Another approach to activate the immune cells against specific tumor targets is the use of therapeutic vaccines. This category of immunotherapy overcomes the necessity of ex vivo manipulation of T cells, as well as their transplantation,134 which opens the way for faster and more personalized treatments. These vaccines rely on the presentation of tumor antigens to T cells by APCs, and may be one of the most personalized forms of immunotherapy when allied with targeted delivery by LNPs.
In summary, the field of immunotherapy has thrived in recent years. However, hurdles in the production process, biodistribution and adverse effects that arise due to autoimmunity still prevent a broader use of these therapies. In this context, LNPs provide a means of facilitating the production process of engineered T cells, and rapid application of gene delivery technologies that would favor therapeutic vaccines and other necessities for targeted delivery of genetic cargo.10 CAR-T cell therapy is a significant methodology that has been applied in immunotherapy and it means a strong candidate to revolutionize immunotherapy, and even autoimmune diseases.
CAR-T Cell Therapy
The concept of chimeric receptors was elucidated in the late 1980s as a tool to induce the recognition of peptides independently of the interaction of T-cell receptor (TCR) and major histocompatibility complex (MHC).135 This marked a significant improvement to TILs or TCR T cells, as tumor cells commonly employ MHC masking as an immune evasion mechanism.136 The CAR is a membrane-bound protein composed by extracellular, transmembrane and intracellular domains.137 These domains have been optimized for clinical use against different tumors, and, in summary, lead to the activation of the engineered T cell upon antigen engagement.138 The development of new generations of CARs led to better survival and function of the T cells, in addition to a reduction in exhaustion and improved tumor infiltration, which are important features in the context of solid tumors.139 Approved products against hematological tumors commonly target CD19 and B cell maturation antigen (BCMA), although clinical studies have investigated alternative tumor associated markers for hematological tumors and for solid tumors.140,141 Novel CAR constructs have been developed over the years, with additional features that allow the control of antigen engagement as well as the control of cytokine release.142,143 However, T cells engineered with new designs as well as clinically approved ones rely on ex vivo manipulation, and the introduction of exogenous genetic material in these cells.7 Alternatively, non-viral vectors have been applied in the manufacture of CAR-T cells. Some investigations have explored electroporation using transposon-based systems, which require less time for ex vivo manipulation than viral transduction.144,145
Integrative viral vectors increase both the price and manufacture time of CAR-T cells products. Combined, these factors become impeditive to a broader application of this therapy, while also affecting the function of the final product.146 A common issue of using integrative approaches, either viral or non-viral, is the constitutive expression of the receptor in the engineered cell. This has been associated with the immunogenicity provoked by the therapy.147 These long-term adverse effects, such as B cell aplasia and immune effector cell-associated neurotoxicity syndrome (ICANS) arise from CAR-T cell antigen engagement on healthy cells.148 Potentially, these adverse effects could be avoided by applying technologies that employ a transient expression of CAR on T cells in the manufacturing process.107
Alternative production methods that allow such transient expression could be applied in the production of CAR-T cells. This would help mitigate the emergence of adverse effects, and significantly reduce the cost and manufacture time, thus broadening the accessibility of CAR-T cell therapy.7 LNPs can be used as tools for RNA-based T cell engineering and, by screening novel ionizable lipids, their transfection can be enhanced as described by Billingsley et al.149,150 Interestingly, by varying the molecular ratios of the components of LNPs, our group has shown improved transfection of T cells using pDNA. Another strategy, which in addition to varying lipids, is the incorporation of DNA-condensing agents, such as the P3000 reagent, during LNP synthesis has also been investigated as a strategy to enhance nucleic acid delivery for CAR-T therapy. This approach improves the efficiency of LNPs following multiple administrations in Jurkat cells, enabling sustained transient expression while preserving cell viability, even under repeated pDNA delivery at 48 h intervals.151 This study provided valuable insights into a non-integrative method of cell engineering, while maintaining cell viability and improved CAR expression when compared to conventional methods such as electroporation and commercial transfection reagents.107
New possibilities of T cell engineering also arise from LNPs. Some platforms have been optimized for increased delivery ex vivo, while concomitantly being used to activate transformed cells. This may lead to significant reduction in the manufacture time, since cell expansion and activation are major bottlenecks of this process.150 An innovative application of LNPs in CAR-T cell engineering is the in vivo delivery of mRNA, which overcomes the need of isolating and manipulating the cells ex vivo.70 Recently, it was demonstrated that LNPs encapsulating CAR mRNA, targeted to the spleen and functionalized with anti-CD3 antibodies, enable the in situ engineering of T cells to express both CAR and IL-7.121 This strategy generated CAR-T cells that effectively infiltrated and eradicated melanoma in vivo with high efficacy and safety, without inducing cytokine release syndrome.121 This technology could improve CAR-T cell function by co-delivering mRNA for CAR expression and siRNA to downregulate exhaustion molecules such as PD-1.152 Interestingly, recent advances on co-delivery have successfully generated CAR-T cells using a transposon-transposase system, which consolidates the applications of LNP platforms in ACT.116
In vivo co-delivery of minicircle DNA (mcDNA) encoding CAR together with transposase mRNA within functionalized nanoparticles bearing anti-CD3 and anti-CD7 demonstrated significant efficacy in leukemia models following a single administration.153 These findings highlight the versatility of LNPs in enabling the efficient co-delivery of multiple genetic cargos while supporting transposase-mediated genomic integration.153 Collectively, this approach offers a promising platform to democratize CAR-T therapies by reducing manufacturing costs, simplifying logistics, and expanding large-scale accessibility.
CAR-T cell therapies represent a highly personalized and potent strategy by reprogramming T cells to recognize and eliminate tumor cells, supported by significant advances in vector design, gene editing technologies, and manufacturing platforms that have expanded their clinical applicability and safety profile. Similarly, cancer vaccines aim to selectively activate these same T cells in vivo, leveraging tumor-associated antigens to elicit a robust and targeted antitumor immune response.
Cancer Vaccines
Another way of leveraging the immune system to target tumor cells is to present peptides processed by APCs and activate antigen-specific cytotoxic T cells.10 This form of immunotherapy, namely cancer vaccines, have been used in the form of cell-based platforms, such as Sipuleucel-T (a dendritic cell vaccine) with significant results against castration-resistant prostate cancer.154 However, other cell-based candidates have shown limited efficacy in clinical trials.154 As a vaccine platform, cancer vaccines could greatly benefit from novel delivery methods, such as LNPs, which emerge as alternatives to traditional delivery methods, and do not rely on cell or virus-based treatments.155,156 With these platforms, nucleic acid delivery can be targeted towards APCs, triggering a more effective activation of antigen-specific cytotoxic cells.155–157
Furthermore, these LNPs can be chemically optimized or functionalized to improve their uptake by APCs. This has been investigated by BioNTech in a Phase I/II clinical trial.158 In this approach, a lipoplex encapsulating an RNA for Claudin 6, used as tumor associated antigen, is given in combination with CAR-T cells targeting the same antigen.159 Preclinical data suggested that this drives the expansion of CAR-T cells and opens an alternative to the treatment of relapsed solid tumors.158 The use of LNPs to enhance cancer treatment have been widely used as described in works using the HPV model of cancer, including the oropharyngeal form. In this way, it was described the success of delivering RNA and saRNA resulting in the decreasing of tumor volume.160,161 Also was possible to observe the reduction in tumor volume of B16F10-OVA mice model when vaccinating the animals with OVA even in mRNA or with the protein associated to the LNP.60,162 In summary, vaccines have been shown to be versatile tools when efficient antigen presentation is necessary. The delivery of nucleic acids (both DNA and RNA) by LNPs is also a proven tool for the fast development of vaccines, as evidenced by the strategies employed in the COVID-19 vaccines.13,17,163 Therefore, LNP platforms may also be a practical way to tailor personalized cancer vaccines, which can be designed using sequencing data from the patient’s own biopsies and trigger efficient T cell responses against antigens that would otherwise be difficult to target.
Furthermore, cytokines are described as key molecules that orchestrate the entire immune system and are intrinsically linked to the body’s response to cancer. As such, they play a fundamental role in cancer treatment and can be used to enhance the immune response against cancer cells. In this context, the use of cytokines encapsulated in LNPs has emerged as a promising and widely explored strategy in cancer therapy, as we will discuss next.
Cytokine-Based Immunotherapy
Cytokines have been fundamental in cancer immunotherapy due to their potent immunomodulatory effects. The first cytokine to be approved by the FDA for cancer treatment was IFN-α in 1986, for hairy cell leukemia.164 This milestone was followed by the approval of IL-2 for metastatic renal cell carcinoma and advanced melanoma in the early 1990s.165 Despite these achievements, the clinical application of cytokines has been hindered by their short half-life and systemic toxicity.166 The necessity for high doses to achieve therapeutic concentrations within the TME has limited their efficacy and highlighted the need for innovative delivery systems that can localize cytokine activity to tumors while minimizing systemic exposure.167,168
The challenges associated with cytokine therapy stem from their mode of action and pharmacokinetics. Cytokines act in a paracrine or autocrine manner and have a relatively short half-life, necessitating large quantities to be systemically administered to achieve a sufficient concentration within the TME.169 This often results in severe toxicities, limiting the dose that can be safely administered. For instance, IL-2 therapy is associated with capillary leak syndrome, a potentially life-threatening condition characterized by hypotension, edema, and organ dysfunction.170 Similarly, IFN-α can cause flu-like symptoms, fatigue, and depression. The use of cytokines in cancer therapy has also been limited by their lack of specificity, leading to off-target effects.171 Consequently, developing localized therapy that directly delivers immune-stimulatory cytokines to tumors is a promising strategy to overcome this dilemma.168 Localized delivery can enhance the therapeutic index by maximizing the concentration of cytokines within the tumor while minimizing systemic exposure.172 The administration of IL-21 and IL-7 mRNA in LNP was described as increasing the CD8+ infiltrated in mouse model of colorectal cancer and breast cancer.74 Using the same mouse model of colorectal cancer, when treating the animals with IL-2 mRNA in different LNPs, a reduction was observed in the size of tumor mass.173 In addition, a recent study described the treatment of Balb/c animals injected with 4T1 cells and treated with IFNα2 mRNA LNPs. They observed as a result, an increase of CXCL9 and CXCL10 chemokines, besides reduction on primary tumor.174 Also, there is described an increase in CD8+ T cells response after injection of IL-12 mRNA in animal model of melanoma.
Furthermore, the TME is a complex and immunosuppressive environment that can limit the efficacy of cytokine therapy.175 Tumors often develop mechanisms to evade immune surveillance, including the production of immunosuppressive cytokines, such as transforming Growth Factor β (TGF-β), and the recruitment of immune suppressive cells, such as regulatory T cells.176 Therefore, effective cytokine therapy must not only deliver immune-stimulatory cytokines to the tumor but also overcome the immunosuppressive TME.177 Recent advances in our understanding of the TME have led to the development of novel strategies to modulate the immune response and enhance the efficacy of cytokine therapy.178
Genetic engineering and gene editing have enabled the development of novel cytokine-based therapies with improved specificity and reduced toxicity. For example, the use of CAR-T cells, which are engineered to also express specific cytokine receptors, have shown promise in enhancing the efficacy and specificity of cytokine therapy.179 In addition, the combination of cytokine therapy with other immunotherapies, such as checkpoint inhibitors, has shown potential in enhancing anti-tumor immune responses.168 The use of cytokines to modulate the TME and enhance the efficacy of checkpoint inhibitors is an area of active research. Overall, the development of novel cytokine-based therapies with improved specificity and reduced toxicity is crucial for realizing the full potential of cytokine therapy in cancer treatment.180
Therefore, mRNA-based approaches enable the in-situ production of therapeutic proteins, including cytokines, directly within target tissues.181 In line with this, LNPs have been shown to effectively deliver mRNA to a range of tissues, including the liver, spleen, and tumors.182 The use of LNPs to deliver mRNA-encoded cytokines, such as IL-12, IL-15 and IL-10, has shown potential in enhancing anti-tumor immune responses and promoting tumor regression.73,183
Moreover, tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) is a cytokine that has received significant interest in oncology due to its remarkable ability to selectively induce apoptosis in cancer cells while sparing healthy tissues.184 This selective cytotoxicity is primarily mediated through TRAIL’s binding to death receptors, particularly DR4 and DR5, which are overexpressed on the surface of many malignant cells.185 Upon receptor engagement, TRAIL initiates a well-characterized intracellular signaling cascade that culminates in the activation of caspases and the execution of programmed cell death184,185 (Figure 4).
 |
Figure 4 mRNA-TRAIL delivery mediated by LNPs. After endocytosis, the mRNA is released into the cytoplasm and translated into TRAIL protein, which is secreted and can act in both autocrine and paracrine manners by binding to death receptors on cancer cells, triggering apoptosis through caspase-8 and caspase-3 activation. |
Several investigations have demonstrated that mRNA-encoded TRAIL, when delivered using LNP platforms, retains its tumoricidal activity and promotes meaningful tumor regression.186–188 Notably, a study employing LNP-mediated delivery of TRAIL mRNA in a murine model of colon cancer reported significant tumor reduction, confirming both the functionality of the expressed protein and the effectiveness of the delivery system.189 These findings underscore the therapeutic promise of this approach and highlight its potential to revitalize interest in TRAIL-based therapies. Ongoing research aimed at optimizing LNP design, improving tumor-targeting efficiency, and enhancing the stability and safety of mRNA formulations will be crucial for the successful clinical translation of this promising therapeutic platform.
Although cytokines hold great promise as anticancer agents, their clinical application as recombinant proteins has been hampered by poor pharmacokinetics, including rapid clearance, short half-life, and limited stability, necessitating frequent high-dose administrations that increase the risk of systemic toxicity.182,190 To overcome these limitations, mRNA–LNP platforms have emerged as a powerful alternative, enabling endogenous production of cytokines directly within target cells and ensuring sustained, localized expression in the TME.186 This strategy bypasses the intrinsic instability of recombinant proteins while allowing more precise control of dosage and expression kinetics. Preclinical studies have demonstrated that LNP-mediated cytokine delivery can preserve biological activity and promote meaningful antitumor responses, underscoring the therapeutic potential of this approach.73,191
Clinical trials are exploring the use of mRNA-LNPs encoding cytokines for cancer treatment. For example, mRNA-2752, an LNP formulation encoding OX40L, IL-23, and IL-36γ, is being evaluated for intratumoral injection in patients with advanced malignancies (NCT03739931). Preliminary results indicate that such localized delivery can stimulate robust anti-tumor immune responses with manageable safety profiles.189 These studies underscore the potential of mRNA-LNP platforms to deliver potent immunostimulatory agents directly to the TME, offering a promising approach for cancer therapy.189
The clinical development of mRNA-LNPs encoding cytokines is an active area of research, with multiple ongoing trials evaluating their safety and efficacy in various cancer types.192 These trials are designed to assess the therapeutic potential of mRNA-LNPs in different cancer settings, including as monotherapy or in combination with other therapies, such as checkpoint inhibitors or chemotherapy.193 The results of these trials will be critical in determining the future of mRNA-LNP therapeutics in cancer therapy. As our understanding of the immune system continues to evolve, it is likely that cytokine therapy will remain a vital component of cancer treatment, offering new hope for patients with advanced malignancies.194
In summary, the importance of cytokines in orchestrating immune responses is well established, and this remains true in the context of cancer. The administration of these molecules via LNPs has shown promise in enhancing their immunomodulatory effects, contributing meaningfully to cancer therapy. Given the central role of the patient’s immune system in therapeutic success, other approaches have also aimed to amplify this response, making treatment more multifactorial. Bispecific antibodies align with this strategy by promoting precise targeting of immune cells toward tumor cells. This technology is highly promising, and in the next section, we will explore how LNPs and bispecific antibodies can be combined to deliver targeted and innovative therapeutics.
Bispecific Antibodies and LNP Functionalization
BsAbs represent a groundbreaking advancement in therapeutic antibody engineering, offering unique capabilities in targeting and treating a variety of diseases. BsAbs are a promising drug immunotherapy with significant potential to address a wide range of diseases, such as cancer and autoimmune disorders.195,196 Unlike traditional monoclonal antibodies that bind to a single antigen, BsAbs are designed to simultaneously bind to two different antigens or epitopes. This dual binding capability enables them to bridge interactions between cells, providing enhanced therapeutic precision and efficacy.197
One of the primary applications of bispecific antibodies is in cancer treatment. BsAbs bind to tumor-associated antigens and CD3 subunits on T cells simultaneously. This binding recruits T cells to the tumor and T cells become activated and release cytolytic proteins that destroy the tumor cells.198,199, BsAbs can enhance tumor specificity and immune system involvement, minimize collateral damage to healthy tissues and when combined with other therapies, such as chemotherapy, can promote anti-cancer activity.200,201 BsAbs have been used to treat hematological malignancies and select solid cancers.197 The clinical success of therapies like Blinatumomab, a BsAb approved for treating acute lymphoblastic leukemia, highlights the potential of this approach. Beyond oncology, bispecific antibodies are being explored for use in treating autoimmune diseases, infectious diseases, and even neurological disorders.202–204 Their adaptability and modularity make them highly versatile tools in addressing complex disease mechanisms. Successfully developing BsAbs requires addressing the challenges of managing toxicities while understanding their binding kinetics, stability, and pharmacokinetics.197
Functionalization plays a crucial role in the design and optimization of BsAbs, enabling precise modulation of their biological activity and therapeutic performance by introducing chemical or molecular modifications. This can involve techniques like covalent or non-covalent attachment of dyes, nanoparticles, or other molecules to the antibody’s surface.205 Advances in antibody engineering have made it possible to tailor BsAbs with improved stability, reduced immunogenicity, and enhanced pharmacokinetics. Functionalization techniques ensure that the dual-binding properties of these antibodies are precisely tuned for specific therapeutic goals.206
One approach in which BsAbs can improve LNP delivery is the functionalization of nanoparticles. This process aims to tailor LNPs for specific therapeutic applications, such as targeted drug delivery to tumors or enhancing mRNA delivery to the brain71,207 (Figure 5). Molecules such as antibodies and peptides can be attached to the LNP surface in order to enhance targeted delivery to diseased cells, improve efficacy, reduce off-target effects, and increase payload delivery. To address issues with circulation time and nonspecific uptake, BsAbs have been used as bridges to target cell surface proteins like PD-L1, CD4, and GRP78. Embedding hemagglutinin antigen (HA) epitopes in LNPs enables BsAbs to enhance transfection efficiency, with notable in vivo and ex vivo improvements in cancer cells and human T cells.208–210 This approach also demonstrates tumor-specific drug delivery with robust binding and therapeutic expression. BsAbs further facilitate targeted mRNA delivery to specific proteins like EGFR and PSMA, showcasing an adaptable, scalable strategy for advancing precision medicine and mRNA therapeutics.210
 |
Figure 5 Antibody-functionalized LNPs improve targeted delivery. Conjugation of antibodies to the LNP surface enhances delivery to specific cells (eg, tumor cells) while minimizing nonspecific uptake by non-target organs such as the liver, reducing off-target accumulation. |
In summary, bispecific antibodies are a transformative class of immunotherapeutics, combining precision and versatility. When combined with innovative functionalization strategies using LNPs, their potential to reshape the treatment landscape across diverse diseases is vast, making them a centerpiece of modern biomedical research and clinical development.
Outlook and Challenges
LNPs have established themselves as a highly promising platform for nucleic acid delivery, opening new horizons for vaccine development, cell therapies, and immunotherapy. Despite recent successes, as evidenced by mRNA-based COVID-19 vaccines, several challenges still limit their full potential. One major obstacle is the difficulty in targeting the LNPs to specific cell types and tissues, to reduce off-target effects and liver accumulation. In terms of manufacturing, although LNPs offer a more cost-effective alternative to viral vectors, issues related to reproducibility persist.
In immunotherapy, LNPs offer new strategies for in vivo engineering of immune cells, cancer vaccination, cytokine delivery, and the development of T cell engagers and bispecific antibodies. The future of LNP technology is moving toward the development of more biodegradable and less immunogenic lipids, the incorporation of targeting ligands for greater specificity, the integration of machine learning and artificial intelligence tools to optimize formulations. Advances in these areas will be crucial to expanding the application of LNPs not only in cancer treatment but also in broader immunotherapeutic approaches, enabling more personalized, potent, and safer therapies.
Future research must focus on optimizing the design of these LNPs, refining their interaction with immune cells, and targeting the immunosuppressive TME to enable more effective therapies. By addressing these issues, the integration of LNP-based platforms into cancer immunotherapy holds the promise of creating more accessible, efficient, and personalized treatments for a wide range of cancers. Ultimately, continued innovations in both the design of delivery technologies and the fundamental understanding of immune system dynamics will pave the way for the next generation of cancer immunotherapies, offering new hope for patients worldwide.
Conclusion
LNPs platforms have emerged as a transformative tool for enhancing the delivery of nucleic acids in cancer immunotherapy. Their ability to deliver mRNA and DNA safely and efficiently enables new possibilities for improving the precision, scalability, and accessibility of immunotherapeutic strategies. By supporting the development of engineered CAR-T cells, cancer vaccines, cytokine therapies, and bsAbs, LNPs offer a versatile and non-viral alternative capable of addressing key challenges such as systemic toxicity, poor biodistribution, and manufacturing complexity. Continued innovation in LNP design and functionalization will be critical to advancing both off-the-shelf and personalized immunotherapies, paving the way for more effective treatments across a wider range of cancers.
Acknowledgement
Research supported by the 2024 AACR Maximizing Opportunity for New Advancements in Research in Cancer (MONARCA) Grant for Latin America, Grant Number 24-15-01-PIRE; National Council for Scientific and Technological Development-CNPq (444429/2024-7, 406266/2024-7, 305932/2022-5; 422002/2023-2; 408482/2022-2; 465425/2014-3), CAPES-FAPEMIG (38/2022 Programa de desenvolvimento da pós-graduação parcerias estratégicas nos estados III), FAPEMIG (APQ-00826-21; APQ-02402-23; RED-00202-22 Rede de Pesquisa em Imunobiológicos e Biofármacos para terapias avançadas e inovadoras; RED-00081-23 Rede de Pesquisa e Inovação para Bioengenharia de Nanosistemas).
Disclosure
The authors declare no potential conflicts of interest in this work.
References
1. Abdeen SK, Mastandrea I, Stinchcombe N, Puschhof J, Elinav E. Diet-microbiome interactions in cancer. Cancer Cell. 2025;43(4):680–707. doi:10.1016/j.ccell.2025.03.013.
2. Cena H, Calder PC. Defining a healthy diet: evidence for the role of contemporary dietary patterns in health and disease. Nutrients. 2020;12(2). doi:10.3390/nu12020334
3. Sung H, Ferlay J, Siegel RL, et al. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin. 2021;71(3):209–249. doi:10.3322/caac.21660
4. Hallek M, Shanafelt TD, Eichhorst B. Chronic lymphocytic leukaemia. Lancet. 2018;391(10129):1524–1537. doi:10.1016/S0140-6736(18)30422-7
5. Mellman I, Coukos G, Dranoff G. Cancer immunotherapy comes of age. Nature. 2011;480(7378):480–489. doi:10.1038/nature10673
6. June CH, O’Connor RS, Kawalekar OU, Ghassemi S, Milone MC. CAR T cell immunotherapy for human cancer. Science. 2018;359(6382):1361–1365. doi:10.1126/science.aar6711
7. Seow Y, Wood MJ. Biological Gene Delivery Vehicles: beyond Viral Vectors. Mol Ther. 2009;17(5):767–777. doi:10.1038/mt.2009.41
8. Guzman G, Reed MR, Bielamowicz K, Koss B, Rodriguez A. CAR-T Therapies in Solid Tumors: opportunities and Challenges. Curr Oncol Rep. 2023;25(5):479–489. doi:10.1007/s11912-023-01380-x
9. Riedl SAB, Kaiser P, Raup A, Synatschke CV, Jérôme V, Freitag R. Non-Viral Transfection of Human T Lymphocytes. Processes. 2018;6(10):188.
10. Riley RS, June CH, Langer R, Mitchell MJ. Delivery technologies for cancer immunotherapy. Nat Rev Drug Discov. 2019;18(3):175–196. doi:10.1038/s41573-018-0006-z
11. Guimarães PPG, Figueroa-Espada CG, Riley RS, et al. In vivo bone marrow microenvironment siRNA delivery using lipid–polymer nanoparticles for multiple myeloma therapy. Proc Natl Acad Sci. 2023;120(25):e2215711120. doi:10.1073/pnas.2215711120
12. Costa PAC, da Silva WN, Moura Prazeres PHD, et al. siRNA lipid nanoparticles for CXCL12 silencing modulate brain immune response during Zika infection. Biomed Pharmacother. 2024:170. doi;10.1016/j.biopha.2023.115981.
13. Guimaraes LC, Costa PAC, Scalzo Junior SRA, et al. Nanoparticle-based DNA vaccine protects against SARS-CoV-2 variants in female preclinical models. Nat Commun. 2024;15(1):590. doi:10.1038/s41467-024-44830-1
14. Shi J, Kantoff PW, Wooster R, Farokhzad OC. Cancer nanomedicine: progress, challenges and opportunities. Nat Rev Cancer. 2017;17(1):20–37. doi:10.1038/nrc.2016.108
15. Gong N, Sheppard NC, Billingsley MM, June CH, Mitchell MJ. Nanomaterials for T-cell cancer immunotherapy. Nat Nanotechnol. 2021;16(1):25–36. doi:10.1038/s41565-020-00822-y
16. Mitchell MJ, Billingsley MM, Haley RM, Wechsler ME, Peppas NA, Langer R. Engineering precision nanoparticles for drug delivery. Nat Rev Drug Discov. 2021;20(2):101–124. doi:10.1038/s41573-020-0090-8
17. Swingle KL, Hamilton AG, Mitchell MJ. Lipid Nanoparticle-Mediated Delivery of mRNA Therapeutics and Vaccines. Trends Mol Med. 2021;27(6):616–617. doi:10.1016/j.molmed.2021.03.003
18. Huang X, Kon E, Han X, et al. Nanotechnology-based strategies against SARS-CoV-2 variants. Nat Nanotechnol. 2022;2022:1. doi:10.1038/s41565-022-01174-5
19. Edeline J, Houot R, Marabelle A, Alcantara M. CAR-T cells and BiTEs in solid tumors: challenges and perspectives. J Hematol Oncol. 2021;14(1):65. doi:10.1186/s13045-021-01067-5
20. Anselmo AC, Mitragotri S. Nanoparticles in the clinic: an update. Bioeng Transl Med. 2019;4(3):e10143–e10143. doi:10.1002/btm2.10143
21. Huang X, Kong N, Zhang X, Cao Y, Langer R, Tao W. The landscape of mRNA nanomedicine. Nat Med. 2022;28(11):2273–2287. doi:10.1038/s41591-022-02061-1
22. Semple SC, Akinc A, Chen J, et al. Rational design of cationic lipids for siRNA delivery. Nat Biotechnol. 2010;28(2):172–176. doi:10.1038/nbt.1602
23. Cullis PR, Hope MJ. Lipid Nanoparticle Systems for Enabling Gene Therapies. Mol Ther. 2017;25(7):1467–1475. doi:10.1016/j.ymthe.2017.03.013
24. Kim BK, Hwang GB, Seu YB, Choi JS, Jin KS, Doh KO. DOTAP/DOPE ratio and cell type determine transfection efficiency with DOTAP-liposomes. Biochim Biophys Acta. 2015;1848(10):1996–2001. doi:10.1016/j.bbamem.2015.06.020
25. Akinc A, Maier MA, Manoharan M, et al. The Onpattro story and the clinical translation of nanomedicines containing nucleic acid-based drugs. Nat Nanotechnol. 2019;14(12):1084–1087. doi:10.1038/s41565-019-0591-y
26. Hou X, Zaks T, Langer R, Dong Y. Lipid nanoparticles for mRNA delivery. Nat Rev Mater. 2021;6(12):1078–1094. doi:10.1038/s41578-021-00358-0
27. Patel S, Ashwanikumar N, Robinson E, et al. Naturally-occurring cholesterol analogues in lipid nanoparticles induce polymorphic shape and enhance intracellular delivery of mRNA. Nat Commun. 2020;11(1):983. doi:10.1038/s41467-020-14527-2
28. Sakurai F, Nishioka T, Yamashita F, Takakura Y, Hashida M. Effects of erythrocytes and serum proteins on lung accumulation of lipoplexes containing cholesterol or DOPE as a helper lipid in the single-pass rat lung perfusion system. Eur J Pharm Biopharm. 2001;52(2):165–172. doi:10.1016/S0939-6411(01)00165-5
29. Mui BL, Tam YK, Jayaraman M, et al. Influence of Polyethylene Glycol Lipid Desorption Rates on Pharmacokinetics and Pharmacodynamics of siRNA Lipid Nanoparticles. Mol Ther Nucleic Acids. 2013;2:e139. doi:10.1038/mtna.2013.66
30. Zhang L, Seow BYL, Bae KH, et al. Role of PEGylated lipid in lipid nanoparticle formulation for in vitro and in vivo delivery of mRNA vaccines. J Control Release. 2025;380:108–124. doi:10.1016/j.jconrel.2025.01.071
31. Suzuki T, Suzuki Y, Hihara T, et al. PEG shedding-rate-dependent blood clearance of PEGylated lipid nanoparticles in mice: faster PEG shedding attenuates anti-PEG IgM production. Int J Pharm. 2020:588. doi;10.1016/j.ijpharm.2020.119792.
32. Wang H, Wang Y, Yuan C, et al. Polyethylene glycol (PEG)-associated immune responses triggered by clinically relevant lipid nanoparticles in rats. NPJ Vaccines. 2023;8(1):1–13. doi:10.1038/s41541-023-00766-z
33. Tenchov R, Sasso JM, Zhou QA. PEGylated Lipid Nanoparticle Formulations: immunological Safety and Efficiency Perspective. Bioconjug Chem. 2023;34(6):941–960. doi:10.1021/ACS.BIOCONJCHEM.3C00174/
34. Kurimoto S, Yoshinaga N, Igarashi K, Matsumoto Y, Cabral H, Uchida S. PEG-OligoRNA Hybridization of mRNA for Developing Sterically Stable Lipid Nanoparticles toward In Vivo Administration. Molecules. 2019;24(7):1303. doi:10.3390/molecules24071303
35. Grillo-López AJ, White CA, Dallaire BK, et al. Rituximab: the first monoclonal antibody approved for the treatment of lymphoma. Curr Pharm Biotechnol. 2000;1(1):1–9. doi:10.2174/1389201003379059
36. Maeki M, Fujishima Y, Sato Y, et al. Understanding the formation mechanism of lipid nanoparticles in microfluidic devices with chaotic micromixers. PLoS One. 2017;12(11):e0187962. doi:10.1371/journal.pone.0187962
37. Kimura N, Maeki M, Ishida A, Tani H, Tokeshi M. One-Step Production Using a Microfluidic Device of Highly Biocompatible Size-Controlled Noncationic Exosome-like Nanoparticles for RNA Delivery. ACS Appl Bio Mater. 2021;4(2):1783–1793. doi:10.1021/acsabm.0c01519
38. Roces CB, Lou G, Jain N, et al. Manufacturing Considerations for the Development of Lipid Nanoparticles Using Microfluidics. Pharmaceutics. 2020;12(11):1095. doi:10.3390/pharmaceutics12111095
39. Terada T, Kulkarni JA, Huynh A, et al. Characterization of Lipid Nanoparticles Containing Ionizable Cationic Lipids Using Design-of-Experiments Approach. Langmuir. 2021;37(3):1120–1128. doi:10.1021/acs.langmuir.0c03039
40. Wang X, Liu S, Sun Y, et al. Preparation of selective organ-targeting (SORT) lipid nanoparticles (LNPs) using multiple technical methods for tissue-specific mRNA delivery. Nat Protoc. 2023;18(1):265–291. doi:10.1038/s41596-022-00755-x
41. Berger M, Degey M, Leblond Chain J, et al. Effect of PEG Anchor and Serum on Lipid Nanoparticles: development of a Nanoparticles Tracking Method. Pharmaceutics. 2023;15(2):597. doi:10.3390/pharmaceutics15020597
42. Lunardi CN, Gomes AJ, Rocha FS, De Tommaso J, Patience GS. Experimental methods in chemical engineering: zeta potential. Can J Chem Eng. 2021;99(3):627–639. doi:10.1002/cjce.23914
43. Vinales I, Silva-Espinoza JC, Medina BA, et al. Selective Transfection of a Transferrin Receptor-Expressing Cell Line with DNA–Lipid Nanoparticles. ACS Omega. 2024;9(38):39533–39545. doi:10.1021/acsomega.4c03541
44. Schober GB, Story S, Arya DP. A careful look at lipid nanoparticle characterization: analysis of benchmark formulations for encapsulation of RNA cargo size gradient. Sci Rep. 2024;14(1). doi:10.1038/s41598-024-52685-1
45. Scalzo S, Santos AK, Ferreira HAS, et al. Ionizable Lipid Nanoparticle-Mediated Delivery of Plasmid DNA in Cardiomyocytes. Int J Nanomed. 2022;17:2865–2881.
46. Athaydes Seabra Ferreira H, Ricardo Aluotto Scalzo Júnior S, Kelton Santos de Faria K, et al. Cryoprotectant optimization for enhanced stability and transfection efficiency of pDNA-loaded ionizable lipid nanoparticles. Int J Pharm. 2024;665:124696. doi:10.1016/j.ijpharm.2024.124696
47. Oostingh GJ, Casals E, Italiani P, et al. Problems and challenges in the development and validation of human cell-based assays to determine nanoparticle-induced immunomodulatory effects. Part Fibre Toxicol. 2011;8(1):8. doi:10.1186/1743-8977-8-8
48. Li Y, Boraschi D. Endotoxin Contamination: a Key Element in the Interpretation of Nanosafety Studies. Nanomedicine. 2015;2015:269–287. doi:10.2217/nnm.15.196
49. Nag K, Sarker MEH, Kumar S, et al. DoE-derived continuous and robust process for manufacturing of pharmaceutical-grade wide-range LNPs for RNA-vaccine/drug delivery. Sci Rep. 2022;12(1). doi:10.1038/s41598-022-12100-z
50. Hamilton AG, Swingle KL, Thatte AS, et al. High-Throughput In Vivo Screening Identifies Differential Influences on mRNA Lipid Nanoparticle Immune Cell Delivery by Administration Route. ACS Nano. 2024;18(25):16151–16165. doi:10.1021/acsnano.4c01171
51. Liu L, Kim JH, Li Z, et al. PEGylated lipid screening, composition optimization, and structure-activity relationship determination for lipid nanoparticle-mediated mRNA delivery. Nanoscale. 2025;17(18):11329–11344. doi:10.1039/d5nr00433k
52. Qin Y, Walters AA, Rouatbi N, Wang JTW, Abdel-Bar HM, Al-Jamal KT. Evaluation of a DoE based approach for comprehensive modelling of the effect of lipid nanoparticle composition on nucleic acid delivery. Biomaterials. 2023;299. doi:10.1016/j.biomaterials.2023.122158
53. Li B, Raji IO, Gordon AGR, et al. Accelerating ionizable lipid discovery for mRNA delivery using machine learning and combinatorial chemistry. Nat Mater. 2024;23(7):1002–1008. doi:10.1038/s41563-024-01867-3
54. Wang W, Feng S, Ye Z, Gao H, Lin J, Ouyang D. Prediction of lipid nanoparticles for mRNA vaccines by the machine learning algorithm. Acta Pharm Sin B. 2022;12(6):2950–2962. doi:10.1016/j.apsb.2021.11.021
55. Bae SH, Choi H, Lee J, et al. Rational Design of Lipid Nanoparticles for Enhanced mRNA Vaccine Delivery via Machine Learning. Small. 2025;21(8). doi:10.1002/smll.202405618
56. Maharjan R, Hada S, Lee JE, et al. Comparative study of lipid nanoparticle-based mRNA vaccine bioprocess with machine learning and combinatorial artificial neural network-design of experiment approach. Int J Pharm. 2023:640. doi;10.1016/j.ijpharm.2023.123012.
57. Wilhelm S, Tavares AJ, Dai Q, et al. Analysis of nanoparticle delivery to tumours. Nature Rev Mater. 2016;1(5):1–12. doi:10.1038/natrevmats.2016.14
58. Fan T, Xu C, Wu J, et al. Lipopolyplex-formulated mRNA cancer vaccine elicits strong neoantigen-specific T cell responses and antitumor activity. Sci Adv. 2024;10(41). doi:10.1126/SCIADV.ADN9961
59. Oberli MA, Reichmuth AM, Dorkin JR, et al. Lipid Nanoparticle Assisted mRNA Delivery for Potent Cancer Immunotherapy. Nano Lett. 2017;17(3):1326–1335. doi:10.1021/ACS.NANOLETT.6B03329
60. Chen J, Ye Z, Huang C, et al. Lipid nanoparticle-mediated lymph node–targeting delivery of mRNA cancer vaccine elicits robust CD8 + T cell response. Proc Natl Acad Sci. 2022;119(34). doi:10.1073/pnas.2207841119
61. Trumpfheller C, Longhi MP, Caskey M, et al. Dendritic cell-targeted protein vaccines: a novel approach to induce T-cell immunity. J Intern Med. 2012;271(2):183–192. doi:10.1111/J.1365-2796.2011.02496.X
62. Mitchell MJ, Billingsley MM, Haley RM, Wechsler ME, Peppas NA, Langer R. Engineering precision nanoparticles for drug delivery. Nat Rev Drug Discov. 2020;20(2):101–124. doi:10.1038/s41573-020-0090-8
63. Akinc A, Querbes W, De S, et al. Targeted delivery of RNAi therapeutics with endogenous and exogenous ligand-based mechanisms. Mol Ther. 2010;18(7):1357–1364. doi:10.1038/mt.2010.85
64. Dilliard SA, Sun Y, Brown MO, et al. The interplay of quaternary ammonium lipid structure and protein Corona on lung-specific mRNA delivery by selective organ targeting (SORT) nanoparticles. J Control Release. 2023;361:361–372. doi:10.1016/J.JCONREL.2023.07.058
65. Zhang Y, Gao Z, Yang X, Xu Q, Lu Y. Leveraging high-throughput screening technologies in targeted mRNA delivery. Mater Today Bio. 2024;26:101101. doi:10.1016/J.MTBIO.2024.101101
66. Dahlman JE, Kauffman KJ, Xing Y, et al. Barcoded nanoparticles for high throughput in vivo discovery of targeted therapeutics. Proc Natl Acad Sci U S A. 2017;114(8):2060–2065. doi:10.1073/PNAS.1620874114/-/DCSUPPLEMENTAL
67. Huayamares SG, Lokugamage MP, Rab R, et al. High-throughput screens identify a lipid nanoparticle that preferentially delivers mRNA to human tumors in vivo. J Control Release. 2023;357:394–403. doi:10.1016/J.JCONREL.2023.04.005
68. Sindhwani S, Syed AM, Ngai J, et al. The entry of nanoparticles into solid tumours. Nat Mater. 2020;19(5):566–575. doi:10.1038/s41563-019-0566-2
69. Palanki R, Riley JS, Bose SK, et al. In utero delivery of targeted ionizable lipid nanoparticles facilitates in vivo gene editing of hematopoietic stem cells. Proc Natl Acad Sci U S A. 2024;121(32):e2400783121. doi:10.1073/PNAS.2400783121/SUPPL_FILE/PNAS.2400783121.SAPP.PDF
70. Billingsley MM, Gong N, Mukalel AJ, et al. In Vivo mRNA CAR T Cell Engineering via Targeted Ionizable Lipid Nanoparticles with Extrahepatic Tropism. Small. 2023:2304378. doi;10.1002/smll.202304378.
71. Han EL, Tang S, Kim D, et al. Peptide-Functionalized Lipid Nanoparticles for Targeted Systemic mRNA Delivery to the Brain. Nano Lett. 2024;25(2). doi:10.1021/ACS.NANOLETT.4C05186
72. Hotz C, Wagenaar TR, Gieseke F, et al. Local delivery of mRNA-encoding cytokines promotes antitumor immunity and tumor eradication across multiple preclinical tumor models. Sci Transl Med. 2021;13(610). doi:10.1126/SCITRANSLMED.ABC7804/SUPPL_FILE/SCITRANSLMED.ABC7804_DATA_FILE_S1.ZIP
73. Liu JQ, Zhang C, Zhang X, et al. Intratumoral delivery of IL-12 and IL-27 mRNA using lipid nanoparticles for cancer immunotherapy. J Control Release. 2022;345:306–313. doi:10.1016/j.jconrel.2022.03.021
74. Hamouda AEI, Filtjens J, Brabants E, et al. Intratumoral delivery of lipid nanoparticle-formulated mRNA encoding IL-21, IL-7, and 4-1BBL induces systemic anti-tumor immunity. Nat Commun. 2024;15(1):1–20. doi:10.1038/s41467-024-54877-9
75. Chow MYT, Qiu Y, Lam JKW. Inhaled RNA Therapy: from Promise to Reality. Trends Pharmacol Sci. 2020;41(10):715–729. doi:10.1016/J.TIPS.2020.08.002/ASSET/1D172246-03EA-4345-989C-CBF5DC60762F/MAIN.ASSETS/GR2.JPG
76. Sahay G, Alakhova DY, Kabanov AV. Endocytosis of nanomedicines. J Control Release. 2010;145(3):182–195. doi:10.1016/J.JCONREL.2010.01.036
77. Huotari J, Helenius A. Endosome maturation. EMBO J. 2011;30(17):3481–3500. doi:10.1038/EMBOJ.2011.286/ASSET/B5610303-F3DC-4963-B0C2-F2EB4A5C7FDA/ASSETS/GRAPHIC/EMBJ2011286-FIG-0007-M.JPG
78. Chatterjee S, Kon E, Sharma P, Peer D. Endosomal escape: a bottleneck for LNP-mediated therapeutics. Proc Natl Acad Sci U S A. 2024;121(11):e2307800120. doi:10.1073/PNAS.2307800120/ASSET/753B2757-1D43-4A2A-AC8D-8BD68E7140F8/ASSETS/IMAGES/LARGE/PNAS.2307800120FIG03.JPG
79. Jayaraman M, Ansell SM, Mui BL, et al. Maximizing the potency of siRNA lipid nanoparticles for hepatic gene silencing in vivo. Angew Chem. 2012;124(34):8657–8661.
80. Knaggs KLM, Sun Y, Walz BA, Pang J, Khan OF. The role of excipients in lipid nanoparticle metabolism: implications for enhanced therapeutic effect. Ther Deliv. 2025;16(7):687–700. doi:10.1080/20415990.2025.2506977;CTYPE:STRING:JOURNAL
81. Suk JS, Xu Q, Kim N, Hanes J, Ensign LM. PEGylation as a strategy for improving nanoparticle-based drug and gene delivery. Adv Drug Deliv Rev. 2016;99:28–51. doi:10.1016/J.ADDR.2015.09.012
82. Jörgensen AM, Wibel R, Bernkop-Schnürch A. Biodegradable Cationic and Ionizable Cationic Lipids: a Roadmap for Safer Pharmaceutical Excipients. Small. 2023;19(17):2206968. doi:10.1002/SMLL.202206968
83. Ho W, Gao M, Li F, Li X, Zhang XQ, Xu X. Next-Generation Vaccines: nanoparticle-Mediated DNA and mRNA Delivery. Adv Healthc Mater. 2021;10(8):2001812. doi:10.1002/adhm.202001812
84. Sebastiani F, Yanez Arteta M, Lerche M, et al. Apolipoprotein E Binding Drives Structural and Compositional Rearrangement of mRNA-Containing Lipid Nanoparticles. ACS Nano. 2021;15(4):6709–6722. doi:10.1021/acsnano.0c10064
85. Kulkarni JA, Myhre JL, Chen S, et al. Design of lipid nanoparticles for in vitro and in vivo delivery of plasmid DNA. Nanomedicine. 2017;13(4):1377–1387. doi:10.1016/j.nano.2016.12.014
86. Nasr SS, Paul P, Loretz B, Lehr CM. Realizing time-staggered expression of nucleic acid-encoded proteins by co-delivery of messenger RNA and plasmid DNA on a single nanocarrier. Drug Deliv Transl Res. 2024;14(12):3339–3353. doi:10.1007/s13346-024-01668-w
87. Sahin U, Karikó K, Ö T. mRNA-based therapeutics — developing a new class of drugs. Nat Rev Drug Discov. 2014;13(10):759–780. doi:10.1038/nrd4278
88. Liao HC, Shen KY, Yang CH, et al. Lipid nanoparticle-encapsulated DNA vaccine robustly induce superior immune responses to the mRNA vaccine in Syrian hamsters. Mol Ther Methods Clin Dev. 2024;32(1). doi:10.1016/j.omtm.2023.101169
89. Kim J, Eygeris Y, Gupta M, Sahay G. Self-assembled mRNA vaccines. Adv Drug Deliv Rev. 2021;170:83–112. doi:10.1016/j.addr.2020.12.014
90. Hao Y, Ji Z, Zhou H, et al. Lipid‐based nanoparticles as drug delivery systems for cancer immunotherapy. MedComm. 2023;4(4). doi:10.1002/mco2.339
91. Bhuyan PK, Dallas M, Kraynyak K, et al. Durability of response to VGX-3100 treatment of HPV16/18 positive cervical HSIL. Hum Vaccin Immunother. 2021;17(5):1288–1293. doi:10.1080/21645515.2020.1823778
92. Algazi A, Bhatia S, Agarwala S, et al. Intratumoral delivery of tavokinogene telseplasmid yields systemic immune responses in metastatic melanoma patients. Ann Oncol. 2020;31(4):532–540. doi:10.1016/j.annonc.2019.12.008
93. Tugues S, Burkhard SH, Ohs I, et al. New insights into IL-12-mediated tumor suppression. Cell Death Differ. 2015;22(2):237–246. doi:10.1038/cdd.2014.134
94. Zuckerman JE, Gritli I, Tolcher A, et al. Correlating animal and human phase Ia/Ib clinical data with CALAA-01, a targeted, polymer-based nanoparticle containing siRNA. Proc Natl Acad Sci. 2014;111(31):11449–11454. doi:10.1073/pnas.1411393111
95. Weber JS, Carlino MS, Khattak A, et al. Individualised neoantigen therapy mRNA-4157 (V940) plus pembrolizumab versus pembrolizumab monotherapy in resected melanoma (KEYNOTE-942): a randomised, phase 2b study. Lancet. 2024;403(10427):632–644. doi:10.1016/S0140-6736(23)02268-7
96. Sahin U, Oehm P, Derhovanessian E, et al. An RNA vaccine drives immunity in checkpoint-inhibitor-treated melanoma. Nature. 2020;585(7823):107–112. doi:10.1038/s41586-020-2537-9
97. Pichard V, Guilbaud M, Devaux M, et al. Incomplete elimination of viral genomes is associated with chronic inflammation in nonhuman primate livers after AAV-mediated gene transfer. Gene Ther. 2025;32(3):287–298. doi:10.1038/s41434-025-00514-z
98. Shao L, Shi R, Zhao Y, et al. Genome-wide profiling of retroviral DNA integration and its effect on clinical pre-infusion CAR T-cell products. J Transl Med. 2022;20(1). doi:10.1186/s12967-022-03729-5
99. PhD MM. Assays Improve Gene Therapy Production. GEN. 2025. https://www.genengnews.com/topics/bioprocessing/assays-improve-gene-therapy-production/.
100. Staff GEN. Vector Production Processes Play Key Role in High Gene Therapy Prices. GEN. 2022. https://www.genengnews.com/insights/vector-production-processes-play-key-role-in-high-gene-therapy-prices/.
101. Jürgens DC, Deßloch L, Porras-Gonzalez D, et al. Lab-scale siRNA and mRNA LNP manufacturing by various microfluidic mixing techniques – an evaluation of particle properties and efficiency. OpenNano. 2023;12:100161. doi:10.1016/j.onano.2023.100161
102. Rothgangl T, Tálas A, Ioannidi EI, et al. Treatment of a metabolic liver disease in mice with a transient prime editing approach. Nat Biomed Eng. 2025. doi:10.1038/s41551-025-01399-4
103. Loughrey D, Paunovska K, Echeverri ES, Tiegreen KE, Dahlman JE. The time course of in vivo cellular responses to LNPs. Chem Commun. 2025;61(23):4535–4538. doi:10.1039/D4CC06659F
104. Shah PD, Huang AC, Xu X, et al. Phase I Trial of Autologous RNA-electroporated cMET-directed CAR T Cells Administered Intravenously in Patients with Melanoma and Breast Carcinoma. Canc Res Commun. 2023;3(5):821–829. doi:10.1158/2767-9764.CRC-22-0486
105. Beatty GL, Haas AR, Maus MV, et al. Mesothelin-Specific Chimeric Antigen Receptor mRNA-Engineered T Cells Induce Antitumor Activity in Solid Malignancies. Cancer Immunol Res. 2014;2(2):112–120. doi:10.1158/2326-6066.CIR-13-0170
106. Kitte R, Rabel M, Geczy R, et al. Lipid nanoparticles outperform electroporation in mRNA-based CAR T cell engineering. Mol Ther Methods Clin Dev. 2023:31. doi;10.1016/j.omtm.2023.101139.
107. Prazeres PHDM, Ferreira H, Costa PAC, et al. Delivery of Plasmid DNA by Ionizable Lipid Nanoparticles to Induce CAR Expression in T Cells. Int J Nanomed. 2023;18(null):5891–5904. doi:10.2147/IJN.S424723
108. Taranejoo S, Liu J, Verma P, Hourigan K. A review of the developments of characteristics of PEI derivatives for gene delivery applications. J Appl Polym Sci. 2015;132(25). doi:10.1002/app.42096
109. Liu C, Xie Y, Li X, et al. Folic Acid/Peptides Modified PLGA–PEI–PEG Polymeric Vectors as Efficient Gene Delivery Vehicles: synthesis, Characterization and Their Biological Performance. Mol Biotechnol. 2021;63(1):63–79. doi:10.1007/s12033-020-00285-5
110. Lv H, Zhang S, Wang B, Cui S, Yan J. Toxicity of cationic lipids and cationic polymers in gene delivery. J Control Release. 2006;114(1):100–109. doi:10.1016/j.jconrel.2006.04.014
111. Bisso PW, Gaglione S, Guimarães PPG, Mitchell MJ, Langer R. Nanomaterial Interactions with Human Neutrophils. ACS Biomater Sci Eng. 2018;4(12):4255–4265. doi:10.1021/acsbiomaterials.8b01062
112. Cronin JG, Jones N, Thornton CA, Jenkins GJS, Doak SH, Clift MJD. Nanomaterials and Innate Immunity: a Perspective of the Current Status in Nanosafety. Chem Res Toxicol. 2020;33(5):1061–1073. doi:10.1021/acs.chemrestox.0c00051
113. Carrillo MA, Zhen A, Mu W, et al. Stem cell-derived CAR T cells show greater persistence, trafficking, and viral control compared to ex vivo transduced CAR T cells. Mol Ther. 2024;32(4):1000–1015. doi:10.1016/j.ymthe.2024.02.026
114. Watanabe N, Mo F, McKenna MK. Impact of Manufacturing Procedures on CAR T Cell Functionality. Front Immunol. 2022;13:876339. doi:10.3389/fimmu.2022.876339
115. Acevedo O. Capstan Therapeutics Presents Preclinical Data on Lead In Vivo CAR-T Candidate, CPTX2309, at American College of Rheumatology (ACR) Convergence 2024. Capstan Therapeutics. 2024. https://www.capstantx.com/press-releases/capstan-therapeutics-presents-preclinical-data-on-lead-in-vivo-car-t-candidate/.
116. Hunter TL, Bao Y, Zhang Y, et al. In vivo CAR T cell generation to treat cancer and autoimmune disease. Science. 2025;388(6753):1311–1317. doi:10.1126/science.ads8473
117. Tombácz I, Laczkó D, Shahnawaz H, et al. Highly efficient CD4+ T cell targeting and genetic recombination using engineered CD4+ cell-homing mRNA-LNPs. Mol Ther. 2021;29(11):3293–3304. doi:10.1016/j.ymthe.2021.06.004
118. Kheirolomoom A, Kare AJ, Ingham ES, et al. In situ T-cell transfection by anti-CD3-conjugated lipid nanoparticles leads to T-cell activation, migration, and phenotypic shift. Biomaterials. 2022:281. doi;10.1016/j.biomaterials.2021.121339.
119. Fu R, Li H, Li R, McGrath K, Dotti G, Gu Z. Delivery Techniques for Enhancing CAR T Cell Therapy against Solid Tumors. Adv Funct Mater. 2021;31(44). doi:10.1002/adfm.202009489
120. Smith TT, Stephan SB, Moffett HF, et al. In situ programming of leukaemia-specific t cells using synthetic DNA nanocarriers. Nat Nanotechnol. 2017;12(8):813–822. doi:10.1038/NNANO.2017.57
121. Li Y, Tian Y, Li C, et al. In situ engineering of mRNA-CAR T cells using spleen-targeted ionizable lipid nanoparticles to eliminate cancer cells. Nano Today. 2024;59:102518. doi:10.1016/j.nantod.2024.102518
122. Engels B, Zhu X, Yang J, et al. Preservation of T-Cell Stemness with a Novel Expansionless CAR-T Manufacturing Process, Which Reduces Manufacturing Time to Less Than Two Days, Drives Enhanced CAR-T Cell Efficacy. Blood. 2021;138(Supplement 1):2848. doi:10.1182/blood-2021-146246
123. Ghassemi S, Nunez-Cruz S, O’Connor RS, et al. Reducing Ex Vivo Culture Improves the Antileukemic Activity of Chimeric Antigen Receptor (CAR) T Cells. Cancer Immunol Res. 2018;6(9):1100–1109. doi:10.1158/2326-6066.Cir-17-0405
124. Ahmed S, Rai KR. Interferon in the treatment of hairy-cell leukemia. Best Pract Res Clin Haematol. 2003;16(1):69–81. doi:10.1016/s1521-6926(02)00084-1
125. Alwan LM, Grossmann K, Sageser D, Van Atta J, Agarwal N, Gilreath JA. Comparison of acute toxicity and mortality after two different dosing regimens of high-dose interleukin-2 for patients with metastatic melanoma. Target Oncol. 2014;9(1):63–71. doi:10.1007/s11523-013-0276-7
126. Rosenberg SA. IL-2: the first effective immunotherapy for human cancer. J Immunol. 2014;192(12):5451–5458. doi:10.4049/jimmunol.1490019
127. Burger JA. Treatment of Chronic Lymphocytic Leukemia. N Engl J Med. 2020;383(5):460–473. doi:10.1056/NEJMra1908213
128. Hodi FS, O’Day SJ, McDermott DF, et al. Improved survival with ipilimumab in patients with metastatic melanoma. N Engl J Med. 2010;363(8):711–723. doi:10.1056/NEJMoa1003466
129. Alsaab HO, Sau S, Alzhrani R, et al. PD-1 and PD-L1 Checkpoint Signaling Inhibition for Cancer Immunotherapy: mechanism, Combinations, and Clinical Outcome. Front Pharmacol. 2017;8:561. doi:10.3389/fphar.2017.00561
130. Webb ES, Liu P, Baleeiro R, Lemoine NR, Yuan M, Wang YH. Immune checkpoint inhibitors in cancer therapy. J Biomed Res. 2018;32(5):317–326. doi:10.7555/jbr.31.20160168
131. Billingham RE, Brent L, Medawar PB. Quantitative studies on tissue transplantation immunity. II. The origin, strength and duration of actively and adoptively acquired immunity. Proc R Soc Lond B Biol Sci. 1954;143(910):58–80. doi:10.1098/rspb.1954.0054
132. Ma H, Padmanabhan Iyer S, Parmar S, Gong Y. Adoptive cell therapy for acute myeloid leukemia. Leuk Lymphoma. 2019;60(6):1370–1380. doi:10.1080/10428194.2018.1553300
133. Zhang R, Zhang Z, Liu Z, et al. Adoptive cell transfer therapy for hepatocellular carcinoma. Front Med. 2019;13(1):3–11.
134. Rurik JG, Tombácz I, Yadegari A, et al. CAR T cells produced in vivo to treat cardiac injury. Science. 2022;375(6576):91–96. doi:10.1126/science.abm0594
135. Kuwana Y, Asakura Y, Utsunomiya N, et al. Expression of chimeric receptor composed of immunoglobulin-derived V regions and T-cell receptor-derived C regions. Biochem Biophys Res Commun. 1987;149(3):960–968. doi:10.1016/0006-291X(87)90502-X
136. Garrido F, Aptsiauri N, Doorduijn EM, Garcia Lora AM, van Hall T. The urgent need to recover MHC class I in cancers for effective immunotherapy. Curr Opin Immunol. 2016;39:44–51. doi:10.1016/j.coi.2015.12.007
137. Castellarin M, Sands C, Da T, et al. A rational mouse model to detect on-target, off-tumor CAR T cell toxicity. JCI Insight. 2020;5(14). doi:10.1172/jci.insight.136012
138. Chailyan A, Marcatili P, Tramontano A. The association of heavy and light chain variable domains in antibodies: implications for antigen specificity. Febs J. 2011;278(16):2858–2866. doi:10.1111/j.1742-4658.2011.08207.x
139. Cappell KM, Kochenderfer JN. A comparison of chimeric antigen receptors containing CD28 versus 4-1BB costimulatory domains. Nat Rev Clin Oncol. 2021;18(11):715–727. doi:10.1038/s41571-021-00530-z
140. Ahmad A, Uddin S, Steinhoff M. CAR-T Cell Therapies: an Overview of Clinical Studies Supporting Their Approved Use against Acute Lymphoblastic Leukemia and Large B-Cell Lymphomas. Int J Mol Sci. 2020;21(11). doi:10.3390/ijms21113906
141. Wei J, Guo Y, Wang Y, et al. Clinical development of CAR T cell therapy in China: 2020 update. Cell Mol Immunol. 2021;18(4):792–804. doi:10.1038/s41423-020-00555-x
142. Giordano-Attianese G, Gainza P, Gray-Gaillard E, et al. A computationally designed chimeric antigen receptor provides a small-molecule safety switch for T-cell therapy. Nat Biotechnol. 2020;38(4):426–432. doi:10.1038/s41587-019-0403-9
143. Reichenbach P, Giordano Attianese GMP, Ouchen K, et al. A lentiviral vector for the production of T cells with an inducible transgene and a constitutively expressed tumour-targeting receptor. Nat Biomed Eng. 2023;7(9):1063–1080. doi:10.1038/s41551-023-01013-5
144. Chicaybam L, Abdo L, Viegas M, et al. Transposon-mediated generation of CAR-T cells shows efficient anti B-cell leukemia response after ex vivo expansion. Gene Ther. 2020;27(1–2):85–95. doi:10.1038/s41434-020-0121-4
145. de Macedo Abdo L, Barros LRC, Saldanha Viegas M, et al. Development of CAR-T cell therapy for B-ALL using a point-of-care approach. Oncoimmunology. 2020;9(1):1752592. doi:10.1080/2162402x.2020.1752592
146. Bulliard Y, Andersson BS, Baysal MA, Damiano J, Tsimberidou AM. Reprogramming T cell differentiation and exhaustion in CAR-T cell therapy. J Hematol Oncol. 2023;16(1):108. doi:10.1186/s13045-023-01504-7
147. Wagner DL, Fritsche E, Pulsipher MA, et al. Immunogenicity of CAR T cells in cancer therapy. Nat Rev Clin Oncol. 2021;2021:1–15.
148. Parker KR, Migliorini D, Perkey E, et al. Single-Cell Analyses Identify Brain Mural Cells Expressing CD19 as Potential Off-Tumor Targets for CAR-T Immunotherapies. Cell. 2020;183(1):126–142.e17. doi:10.1016/j.cell.2020.08.022
149. Billingsley MM, Hamilton AG, Mai D, et al. Orthogonal Design of Experiments for Optimization of Lipid Nanoparticles for mRNA Engineering of CAR T Cells. Nano Lett. 2022;22(1):533–542. doi:10.1021/acs.nanolett.1c02503
150. Billingsley MM, Singh N, Ravikumar P, Zhang R, June CH, Mitchell MJ. Ionizable lipid nanoparticle-mediated mRNA delivery for human CAR T cell engineering. Nano Lett. 2020;20(3):1578–1589.
151. Pirrottina A, Renzi S, Digiacomo L, et al. Precondensed Plasmid DNA Enhances CAR-T Cell Generation via Lipid Nanoparticles. ACS Omega. 2025;10(27):29804–29814. doi:10.1021/acsomega.5c04308
152. Hamilton AG, Swingle KL, Joseph RA, et al. Ionizable Lipid Nanoparticles with Integrated Immune Checkpoint Inhibition for mRNA CAR T Cell Engineering. Adv Healthc Mater. 2023;12(30). doi:10.1002/adhm.202301515
153. Bimbo JF, van Diest E, Murphy DE, et al. T cell-specific non-viral DNA delivery and in vivo CAR-T generation using targeted lipid nanoparticles. J Immunother Cancer. 2025;13(7):e011759. doi:10.1136/jitc-2025-011759
154. Kantoff PW, Higano CS, Shore ND, et al. Sipuleucel-T Immunotherapy for Castration-Resistant Prostate Cancer. N Engl J Med. 2010;363(5):411–422. doi:10.1056/NEJMoa1001294
155. Li L, Goedegebuure SP, Gillanders WE. Preclinical and clinical development of neoantigen vaccines. Ann Oncol. 2017;28(suppl_12):xii11–xii17. doi:10.1093/annonc/mdx681
156. Pardi N, Hogan MJ, Porter FW, Weissman D. mRNA vaccines - a new era in vaccinology. Nat Rev Drug Discov. 2018;17(4):261–279. doi:10.1038/nrd.2017.243
157. Chen D, Love KT, Chen Y, et al. Rapid Discovery of Potent siRNA-Containing Lipid Nanoparticles Enabled by Controlled Microfluidic Formulation. J Am Chem Soc. 2012;134(16):6948–6951. doi:10.1021/ja301621z
158. Mackensen A, Haanen JBAG, Koenecke C, et al. CLDN6-specific CAR-T cells plus amplifying RNA vaccine in relapsed or refractory solid tumors: the phase 1 BNT211-01 trial. Nat Med. 2023;29(11):2844–2853. doi:10.1038/s41591-023-02612-0
159. Reinhard K, Rengstl B, Oehm P, et al. An RNA vaccine drives expansion and efficacy of claudin-CAR-T cells against solid tumors. Science. 2020;367(6476):446–453. doi:10.1126/science.aay5967
160. Qiu K, Duan X, Mao M, et al. mRNA-LNP vaccination-based immunotherapy augments CD8+ T cell responses against HPV-positive oropharyngeal cancer. NPJ Vaccines. 2023;8(1). doi:10.1038/s41541-023-00733-8
161. Ramos da Silva J, Bitencourt Rodrigues K, Formoso Pelegrin G, et al. Single immunizations of self-amplifying or non-replicating mRNA-LNP vaccines control HPV-associated tumors in mice. Sci Transl Med. 2023;15(686). doi:10.1126/scitranslmed.abn3464
162. Chen J, Qiu M, Ye Z, et al. In Situ Cancer Vaccination Using Lipidoid Nanoparticles. 2021;Vol. 7. Available from https://www.science.org.
163. Liu MA. A Comparison of Plasmid DNA and mRNA as Vaccine Technologies. Vaccines. 2019;7(2):37.
164. Borden EC. Interferons α and β in cancer: therapeutic opportunities from new insights. Nat Rev Drug Discov. 2019;18(3):219–234. doi:10.1038/s41573-018-0011-2
165. Mullard A. Restoring IL-2 to its cancer immunotherapy glory. Nat Rev Drug Discov. 2021;20(3):163–165. doi:10.1038/d41573-021-00034-6
166. Waldmann TA. Cytokines in Cancer Immunotherapy. Cold Spring Harb Perspect Biol. 2018;10(12):a028472. doi:10.1101/cshperspect.a028472
167. Ryffel B. Interleukin-12: role of Interferon-γ in IL-12 Adverse Effects. Clin Immunol Immunopathol. 1997;83(1):18–20. doi:10.1006/clin.1996.4306
168. Yi M, Li T, Niu M, et al. Targeting cytokine and chemokine signaling pathways for cancer therapy. Signal Transduct Target Ther. 2024;9(1):176. doi:10.1038/s41392-024-01868-3
169. Colombo MP, Trinchieri G. Interleukin-12 in anti-tumor immunity and immunotherapy. Cytokine Growth Factor Rev. 2002;13(2):155–168. doi:10.1016/S1359-6101(01)00032-6
170. Siddall E, Khatri M, Radhakrishnan J. Capillary leak syndrome: etiologies, pathophysiology, and management. Kidney Int. 2017;92(1):37–46. doi:10.1016/j.kint.2016.11.029
171. Conlon KC, Miljkovic MD, Waldmann TA. Cytokines in the Treatment of Cancer. Journal of Interferon & Cytokine. Research. 2019;39(1):6–21. doi:10.1089/jir.2018.0019
172. Dholakia J, Cohen AC, Leath CA, Evans ET, Alvarez RD, Thaker PH. Development of Delivery Systems for Local Administration of Cytokines/Cytokine Gene-Directed Therapeutics: modern Oncologic Implications. Curr Oncol Rep. 2022;24(4):389–397. doi:10.1007/s11912-022-01221-3
173. Jiang Y, Zhang Y, Liu C, et al. Tumor-activated IL-2 mRNA delivered by lipid nanoparticles for cancer immunotherapy. J Control Release. 2024;368:663–675. doi:10.1016/j.jconrel.2024.03.016
174. Fick K, Kerns N, Zhao Y, et al. Lipid nanoparticle-delivered IFNα2 activates Cxcl9 to increase T cell tumor recruitment to suppress lung metastasis. J Immunother Cancer. 2025;13(7):e011415. doi:10.1136/jitc-2024-011415
175. de Visser KE, Joyce JA. The evolving tumor microenvironment: from cancer initiation to metastatic outgrowth. Cancer Cell. 2023;41(3):374–403. doi:10.1016/j.ccell.2023.02.016
176. Wu B, Zhang B, Li B, Wu H, Jiang M. Cold and hot tumors: from molecular mechanisms to targeted therapy. Signal Transduct Target Ther. 2024;9(1):274. doi:10.1038/s41392-024-01979-x
177. Giri S, Lamichhane G, Pandey J, et al. Immune Modulation and Immunotherapy in Solid Tumors: mechanisms of Resistance and Potential Therapeutic Strategies. Int J Mol Sci. 2025;26(7):2923. doi:10.3390/ijms26072923
178. Chen J, Yang L, Ma Y, Zhang Y. Recent advances in understanding the immune microenvironment in ovarian cancer. Front Immunol. 2024;15. doi:10.3389/fimmu.2024.1412328
179. Atsavapranee ES, Billingsley MM, Mitchell MJ. Delivery technologies for T cell gene editing: applications in cancer immunotherapy. EBioMedicine. 2021;67:103354. doi:10.1016/j.ebiom.2021.103354
180. Muhammad S, Fan T, Hai Y, Gao Y, He J. Reigniting hope in cancer treatment: the promise and pitfalls of IL-2 and IL-2R targeting strategies. Mol Cancer. 2023;22(1):121. doi:10.1186/s12943-023-01826-7
181. Qin S, Tang X, Chen Y, et al. mRNA-based therapeutics: powerful and versatile tools to combat diseases. Signal Transduct Target Ther. 2022;7(1):166. doi:10.1038/s41392-022-01007-w
182. Zhang T, Yin H, Li Y, et al. Optimized lipid nanoparticles (LNPs) for organ-selective nucleic acids delivery in vivo. iScience. 2024;27(6):109804. doi:10.1016/j.isci.2024.109804
183. Liu C, Huang X, Chen KS, et al. Systemic reprogramming of tumour immunity via IL-10-mRNA nanoparticles. Nat Nanotechnol. 2025. doi:10.1038/s41565-025-01980-7
184. Montinaro A, Walczak H. Harnessing TRAIL-induced cell death for cancer therapy: a long walk with thrilling discoveries. Cell Death Differ. 2023;30(2):237–249. doi:10.1038/s41418-022-01059-z
185. Bertheloot D, Latz E, Franklin BS. Necroptosis, pyroptosis and apoptosis: an intricate game of cell death. Cell Mol Immunol. 2021;18(5):1106–1121. doi:10.1038/s41423-020-00630-3
186. Guimarães PPG, Gaglione S, Sewastianik T, Carrasco RD, Langer R, Mitchell MJ. Nanoparticles for Immune Cytokine TRAIL-Based Cancer Therapy. ACS Nano. 2018;12(2):912–931. doi:10.1021/acsnano.7b05876
187. Gu T, Wang M, Fu X, et al. Intratumoural delivery of TRAIL mRNA induces colon cancer cell apoptosis. Biomed Pharmacother. 2024:174. doi;10.1016/j.biopha.2024.116603.
188. Riera-Domingo C, Leite-Gomes E, Charatsidou I, et al. Breast Tumors Interfere with Endothelial TRAIL at the Premetastatic Niche to Promote Cancer Cell Seeding. 2023. https://www.science.org.
189. da Silva W, Carvalho Costa PA, Scalzo Júnior SR, et al. Ionizable Lipid Nanoparticle-Mediated TRAIL mRNA Delivery in the Tumor Microenvironment to Inhibit Colon Cancer Progression. Int J Nanomed. 2024;19:2655–2673. doi:10.2147/ijn.s452896
190. Stuckey DW, Shah K. TRAIL on trial: preclinical advances in cancer therapy. Trends Mol Med. 2013;19(11):685–694. doi:10.1016/j.molmed.2013.08.007
191. Aunins EA, Phan AT, Alameh MG, et al. An Il12 mRNA-LNP adjuvant enhances mRNA vaccine-induced CD8 T cell responses. Sci Immunol. 2025;10(108). doi:10.1126/sciimmunol.ads1328
192. Zhong Y, Du S, Dong Y. mRNA delivery in cancer immunotherapy. Acta Pharm Sin B. 2023;13(4):1348–1357. doi:10.1016/j.apsb.2023.03.001
193. Bidram M, Zhao Y, Shebardina NG, et al. mRNA-Based Cancer Vaccines: a Therapeutic Strategy for the Treatment of Melanoma Patients. Vaccines. 2021;9(10):1060. doi:10.3390/vaccines9101060
194. Ghemrawi R, Abuamer L, Kremesh S, et al. Revolutionizing Cancer Treatment: recent Advances in Immunotherapy. Biomedicines. 2024;12(9):2158. doi:10.3390/biomedicines12092158
195. Baeuerle PA, Reinhardt C. Bispecific T-cell engaging antibodies for cancer therapy. Cancer Res. 2009;69(12):4941–4944. doi:10.1158/0008-5472.CAN-09-0547
196. Labrijn AF, Janmaat ML, Reichert JM, PWHI P. Bispecific antibodies: a mechanistic review of the pipeline. Nat Rev Drug Discov. 2019;18(8):585–608. doi:10.1038/S41573-019-0028-1
197. Herrera M, Pretelli G, Desai J, et al. Bispecific antibodies: advancing precision oncology. Trends Cancer. 2024;10(10):893–919. doi:10.1016/J.TRECAN.2024.07.002
198. Qin X, Ning W, Luo W, Xia N. Stepping forward: t-cell redirecting bispecific antibodies in cancer therapy. Acta Pharm Sin B. 2024;14(6):2361–2377. doi:10.1016/J.APSB.2024.03.027
199. van de Donk NWCJ, Zweegman S. T-cell-engaging bispecific antibodies in cancer. Lancet. 2023;402(10396):142–158. doi:10.1016/S0140-6736(23)00521-4
200. Kim HU, Kim YK. Bispecific antibodies and CLEM: an analytical approach to advanced cell imaging for therapeutic strategies. Appl Microsc. 2025;55(1):1. doi:10.1186/S42649-024-00106-Y
201. Lan HR, Chen M, Yao SY, Chen JX, Jin KT. Bispecific antibodies revolutionizing breast cancer treatment: a comprehensive overview. Front Immunol. 2023;14:1266450. doi:10.3389/FIMMU.2023.1266450
202. Wagner EK, Maynard JA. Engineering therapeutic antibodies to combat infectious diseases. Curr Opin Chem Eng. 2018;19:131. doi:10.1016/J.COCHE.2018.01.007
203. Yu YJ, Atwal JK, Zhang Y, et al. Therapeutic bispecific antibodies cross the blood-brain barrier in nonhuman primates. Sci Transl Med. 2014;6(261). doi:10.1126/SCITRANSLMED.3009835/SUPPL_FILE/6-261RA154_SM.PDF
204. Zhao Q. Bispecific Antibodies for Autoimmune and Inflammatory Diseases: clinical Progress to Date. BioDrugs. 2020;34(2):111–119. doi:10.1007/S40259-019-00400-2
205. Eloy JO, Petrilli R, Trevizan LNF, Chorilli M. Immunoliposomes: a review on functionalization strategies and targets for drug delivery. Colloids Surf B Biointerfaces. 2017;159:454–467. doi:10.1016/J.COLSURFB.2017.07.085
206. Amash A, Volkers G, Farber P, et al. Developability considerations for bispecific and multispecific antibodies. MAbs. 2024;16(1):2394229. doi:10.1080/19420862.2024.2394229
207. Marques AC, Costa PC, Velho S, Amaral MH. Lipid Nanoparticles Functionalized with Antibodies for Anticancer Drug Therapy. Pharmaceutics. 2023;15(1):216. doi:10.3390/PHARMACEUTICS15010216
208. Amabile A, Phelan M, Yu Z, et al. Bispecific antibody targeting of lipid nanoparticles. bioRxiv. 2024. doi:10.1101/2024.12.20.629467
209. Zhang Y, Zhang Z. The history and advances in cancer immunotherapy: understanding the characteristics of tumor-infiltrating immune cells and their therapeutic implications. Cell Mol Immunol. 2020;17(8):807–821. doi:10.1038/s41423-020-0488-6
210. Dietmair B, Humphries J, Mercer TR, Thurecht KJ, Howard CB, Cheetham SW. Targeted mRNA delivery with bispecific antibodies that tether LNPs to cell surface markers. Mol Ther Nucleic Acids. 2025;36(2). doi:10.1016/j.omtn.2025.102520
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Hydrogel-Based Vaccines: A Promising Approach for Cancer Immunotherapy
Zhang W, Zhang Q, Cai J, He J, Xu Z, Chen X, Chen G
International Journal of Nanomedicine 2025, 20:11389-11415
Published Date: 18 September 2025