Back to Journals » Neuropsychiatric Disease and Treatment » Volume 16
ABCA7 Gene Expression and Genetic Association Study in Schizophrenia
Authors Yamazaki K, Yoshino Y, Kawabe K ![]() , Ibuki T, Ochi S
, Ibuki T, Ochi S ![]() , Mori Y, Ozaki Y, Numata S, Iga J
, Mori Y, Ozaki Y, Numata S, Iga J ![]() , Ohmori T
, Ohmori T ![]() , Ueno S
, Ueno S ![]()
Received 13 November 2019
Accepted for publication 25 January 2020
Published 13 February 2020 Volume 2020:16 Pages 441—446
DOI https://doi.org/10.2147/NDT.S238471
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Taro Kishi
Kiyohiro Yamazaki,1 Yuta Yoshino,1 Kentaro Kawabe,1 Tomomasa Ibuki,1 Shinichiro Ochi,1 Yoko Mori,1 Yuki Ozaki,1 Shusuke Numata,2 Jun-ichi Iga,1 Tetsuro Ohmori,2 Shu-ichi Ueno1
1Department of Neuropsychiatry, Molecules and Function, Ehime University Graduate School of Medicine, Toon, Ehime 791-0295, Japan; 2Department of Psychiatry, Course of Integrated Brain Sciences, Medical Informatics, Institute of Health Biosciences, the University of Tokushima Graduate School, Tokushima 770-8503, Japan
Correspondence: Jun-ichi Iga
Department of Neuropsychiatry, Molecules and Function, Ehime University Graduate School of Medicine, Shitsukawa, Toon, Ehime 791-0295, Japan
Tel +81-89-960-5315
Fax +81-89-960-5317
Email [email protected]
Introduction: Although ATP-binding cassette sub-family A member 7 gene (ABCA7) is known to be associated with Alzheimer’s disease, the relationship between ABCA7 and schizophrenia has been unknown.
Methods: Schizophrenia patients (n = 50; 24 males, 62.1 ± 0.50 years old) and age- and sex-matched healthy controls (n = 50) were recruited for the mRNA analysis. Additionally, a case-control study for the rs3764650 genotypes was performed with 1308 samples (control subjects; n = 527, schizophrenia patients; n = 781). All participants were Japanese, unrelated to each other, and living in the same area.
Results: The distributions of the rs3764650 genotypes in schizophrenia patients were not different from that of controls. However, the ABCA7 mRNA expression levels in schizophrenia patients were significantly higher than those in controls by a logistic regression analysis. Additionally, the ABCA7 mRNA expression levels in schizophrenia patients were correlated with the rs3764650 genotypes in a dose-dependent manner.
Discussion: The ABCA7 mRNA expression levels in peripheral blood with the rs3764650 genotypes may be related to pathological mechanisms in schizophrenia and may be a biological marker for schizophrenia.
Keywords: schizophrenia, ATP-binding cassette sub-family A member 7 gene, single nucleotide polymorphism, rs3764650, mRNA expression
Introduction
The exact cause of schizophrenia remains unknown although that had been tried to explain by dopamine hypothesis. Schizophrenia was thought to be one of endogenous mental illnesses, and brain changes in schizophrenia patients were difficult to be detected. However, in the recent studies, the pathophysiology of schizophrenia was often examined by imaging or genetic approaches. Several studies using MRI scanning reported that the grey-matter abnormalities were associated with psychotic symptoms,1 and the cortical volume loss in the right prefrontal region was associated with the onset of psychosis.2 The morphological and functional changes in the brain of schizophrenia are being revealed by these studies. In the genetic studies of schizophrenia the risk genes had been shown. The recent study reported that the RNA-sequencing data of prefrontal cortex in combination with the genotypes were analyzed and identified cis-acting splicing quantitative trait loci throughout the genome.3 This study observed significant enrichment of splicing quantitative trait loci among disease-associated loci by GWAS especially in schizophrenia. The finding can facilitate understanding of the genetic architecture of complex brain disorders such as schizophrenia.
The ABCA7 (ATP-binding cassette subfamily A member 7 gene) was identified as candidate genes for Alzheimer’s disease (AD) with large-scale genome-wide association studies.4–8 The ABCA7 encodes a member of the superfamily of ATP-binding cassette transporters expressed on both microglia and neurons in the human brain. The ABCA7 is involved in the transport of phospholipids and enhances phagocytosis by macrophages.9–13 Moreover, the ABCA7 expression is increased in the brain of AD, although a single nucleotide polymorphism (SNP) in the ABCA7 the most frequently studied, rs3764650 major allele T that decreases the risk of AD is associated with increased the ABCA7 expression.14 We reported that the ABCA7 mRNA expression level in peripheral blood of patients with AD was higher than that of normal subjects and was significantly correlated with cognitions by Mini Mental State Examination recall, the Alzheimer’s Disease Assessment Scale total score, and the Clinical Dementia Rating score regardless of rs3764650 and the methylation rate of its promoter.15 It is suggested that the ABCA7 expression may be a marker for the early stages of AD and disease progression.
Although there were dysfunctions of autoimmune system and elevated inflammatory cytokines in schizophrenia,16–18 there was only one study suggesting the defects in the phagocytotic pathway through ABCA7 may be a contributing factor for aberrant autoimmune system and elevated inflammatory cytokines in schizophrenia.19 This study reported the results of typing 11 and 5 SNPs for the ABCA1 and ABCA7 genes, and rs4149324 in ABCA1 was nominally associated with the disease in the Irish case–control study of schizophrenia sample and rs2242436 in ABCA7 was significantly associated with the disease in the Irish study of high-density schizophrenia families sample.19 These results suggested that the phagocytotic pathway including ABCA7 was involved in the development of schizophrenia.
However, this study has not reported the association among the ABCA7 expression, rs3764650 in the ABCA7 and the schizophrenia yet. Thus, the aim of this study was to examine the ABCA7 expression of peripheral blood and rs3764650 in the ABCA7 as a biological marker of schizophrenia.
Methods
Subjects
Demographic data and clinical characteristics of control subjects and schizophrenia patients are shown in Table 1. We enrolled 50 schizophrenia patients [24 males and 26 females, mean age ± standard deviation (SD) = 62.1 ± 0.50 years] who visited Ehime University Hospital and related community hospitals, Ehime, Japan, and control subjects [25 males and 25 females, mean age ± standard deviation (SD) = 61.8 ± 0.51 years] who were recruited from volunteers, staffs in the hospitals and company employees for the mRNA analysis. Schizophrenia patients were diagnosed according to the Diagnostic and Statistical Manual of Mental Disorders (DSM) – 5 criteria by at least two expert psychiatrists based on extensive clinical interviews and a review of medical records. Control subjects were documented to be free from psychiatric symptoms, and past histories of mental illness, and psychotropic medications. Schizophrenia patients were evaluated with the 18-item Brief Psychiatric Rating Scale (BPRS) and the Drug-Induced Extrapyramidal Symptoms Scale (DIEPSS). The medications of the schizophrenia patients were examined by the chlorpromazine equivalent dose.
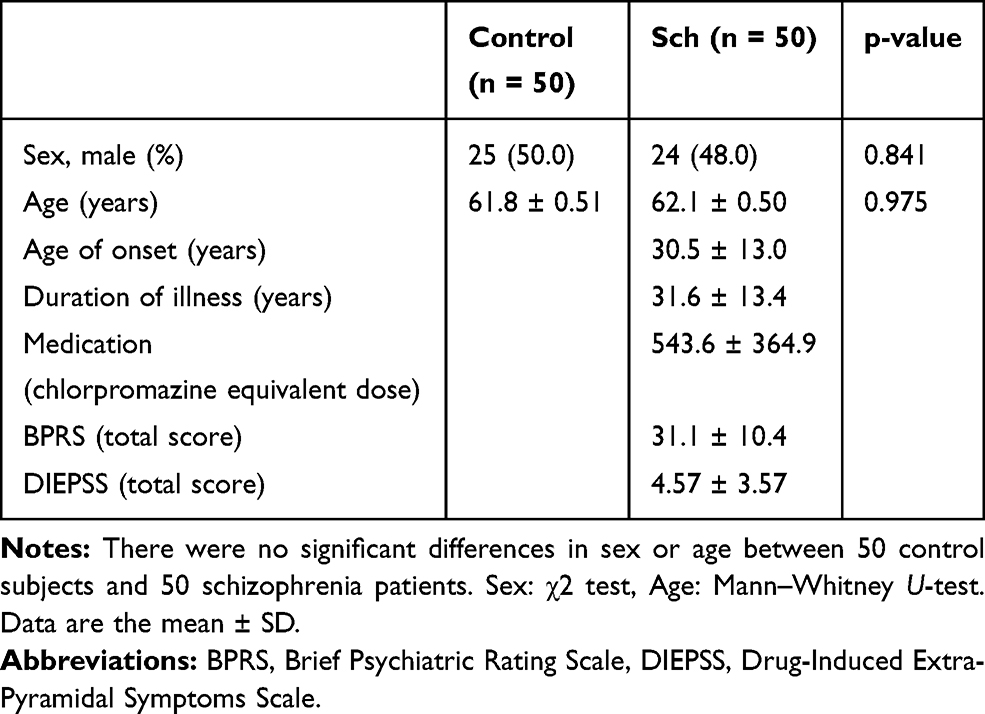 |
Table 1 Demographic Data and Clinical Characteristics of 50 Control Subjects and 50 Schizophrenia Patients |
Additionally, we enrolled 781 schizophrenia patients [451 males and 330 females, mean age ± standard deviation (SD) = 53.7 ± 14.5 years] and 527 control subjects [281 males and 246 females, mean age ± standard deviation (SD) = 37.2 ± 15.8 years] including 731 schizophrenia patients and 477 control subjects from Ehime or Tokushima University Hospital for a case–control genetic association study. Schizophrenia patients were diagnosed according to the Diagnostic and Statistical Manual of Mental Disorders (DSM) – 4 criteria by at least two expert psychiatrists based on extensive clinical interviews and a review of medical records. Control subjects were healthy volunteers without psychiatric signs, psychiatric family history, or history of mental disorders.
All participants were unrelated, of Japanese origin, and provided written informed consent forms that were approved by the institutional ethics committees of Ehime University and Tokushima University. This study was conducted in accordance with the Declaration of Helsinki.
Blood Sample Collections, Extraction of the Genomic DNA (gDNA), and Synthesis of the Complementary DNA (cDNA)
The gDNA was obtained from whole peripheral blood samples collected in potassium EDTA tubes and extracted using the QIAamp DNA Blood Mini Kit (Qiagen, Tokyo, Japan) according to the standard protocol. The total RNA was isolated from whole peripheral blood samples using PaxGene Blood RNA Systems tubes (BD, Tokyo, Japan) according to the standard protocol. The RNA concentration and purity were measured with a NanoDrop-1000 (Thermo Fisher Scientific, Yokohama, Japan), and the 260/280 ratio was between 1.8 and 2.0. RNA (1.0 μg) in a 40-μL total reaction volume per sample was used to synthesize the cDNA by using the High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA, USA).
Genotyping Analysis
The SNP analysis of rs3764650 was performed with real-time quantitative PCR. A specific TaqMan probe for the SNP analysis was selected for NM_019112.3; assay ID: C__27478162_10 (TaqMan Assays, Applied Biosystems). The minor allele frequency of rs3764650 was based on the current International HapMap project database (http://hapmap.ncbi.nlm.nih.gov/index.html.en). PCR was performed with 1 × TaqMan PCR Master Mix, 1 × TaqMan SNP genotyping assay, 10 ng gDNA, and ultrapure water in a final reaction volume of 6 μL in each well of an optical plate. Allelic discrimination was determined using the StepOnePlus Real-Time PCR System and analyzed using StepOnePlus software.
Expression Analysis
The mRNA expression analysis was performed by the quantitative reverse transcription-PCR using the StepOnePlus Real-Time PCR System (Applied Biosystems). The specific TaqMan probes were Hs01105117_m1 for ABCA7 and Hs99999905_m1 for GAPDH. We used GAPDH as a housekeeping gene because previous studies including ours consistently identified GAPDH as a suitable housekeeping gene for the peripheral blood gene expression analysis using the Paxgene blood RNA system.20–23 The final volume of the reaction was 10 μL including the TaqMan Universal Master Mix (Applied Biosystems). The expression levels were determined in triplicate. The ΔΔCt method was used to determine the relative expression levels using StepOne software (Applied Biosystems).24 To correct for observational errors, the PCR was performed in all plates with gDNA from the same control subjects in this study.
Statistical Analysis
Statistical analyses were performed using SPSS Statistics version 22.0 (IBM Corp., Tokyo, Japan) and EZR.25 The difference in sex or age between control subjects and schizophrenia patients was analyzed by χ2 test or Mann–Whitney U-test, respectively. For analysis of the effects of SNPs, all participants were divided into groups according to the presence or absence of the G allele of the rs3764650 as a previous study.14 Additionally, all participants were divided into three groups according to the number of the G allele of the rs3764650 for examining “dose-dependent” manner; T/T (dose = 0), G/T (dose = 1), and G/G (dose = 2). The ratios for each allele of the rs3764650 between control subjects and schizophrenia patients divided into the groups were compared by χ2 test in two sets that were 527 control subjects versus 781 schizophrenia patients. A logistic regression analysis was performed with the age, the sex, the rs3764650 genotypes, the expression levels between 50 schizophrenia patients and 50 control subjects. A linear regression analysis was performed among the expression and the sex, the age, the duration of illness, the dose of antipsychotics, the total score of the BPRS, that of the DIEPSS, the counts of the G allele of the rs3764650 in 50 schizophrenia patients. Although we have genomic DNA samples of 781 schizophrenia patients and 527 control subjects, we have only a small number (N=100) of samples with both mRNA and gDNA. The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
Results
Participant Characteristics
Demographic data and clinical characteristics of 50 control subjects and 50 schizophrenia patients are shown in Table 1. There were no differences of sex or age between 50 control subjects and 50 schizophrenia patients analyzed by χ2 test or Mann–Whitney U-test, respectively.
Genotyping and a Case–Control Study for the rs3764650
Distribution of genotypes and allele frequencies of the rs3764650 in 527 control subjects and 781 schizophrenia patients was shown in Table 2. There were no significant differences in genotypes and allele frequencies according to the χ2 test.
 |
Table 2 Genotype Distributions and Allele Frequencies of the Rs3764650 |
ABCA7 mRNA Expression Levels
The result of the logistic regression analysis between 50 schizophrenia patients and 50 control subjects revealed that significant change was only observed in expression levels and without the age, the sex, the rs3764650 genotypes [1.618 (95% CI: 1.763–14.415), p = 0.03]. The expression levels are shown in Figure 1.
 |
Figure 1 The ABCA7 mRNA expression levels of 50 schizophrenia patients and 50 control subjects. |
Relationships Among the ABCA7 mRNA Expression Levels, the rs3764650 Genotypes, and Each Parameter in Schizophrenia Patients
The multiple regression analysis including age, sex and genotype revealed a significant regression only between the ABCA7 mRNA expression level and the number of the G allele of the rs3764650 (Beta = 0.365, p = 0.01) [The regression equation: The ABCA7 mRNA expression level = 0.863 + 0.365 * number of G allele (0, 1, 2)].
Discussion
This study revealed three main findings.
First, the higher expression levels of the ABCA7 in schizophrenia patients compared to control subjects in the logistic regression analysis. This finding is consistent with the result of previous studies in the peripheral blood or the brain of the AD.14,15 The dysfunction of the ABCA7 could cause an abnormal autoimmune system and elevated inflammatory cytokines by the phagocytotic pathway. Several studies indicate the relationships between the schizophrenia and the peripheral inflammation markers. It was reported that in acute psychosis, the peripheral inflammation marker C-reactive protein (CRP) and neutrophil-to-lymphocyte ratio were significantly elevated relative to a normal cutoff, and CRP was significantly elevated in 43% of chronically ill patients with schizophrenia compared to 20% in controls.26 Additionally, schizophrenia patients had significantly higher interleukin-10 plasma levels compared to the controls in the recent study.27 Therefore, the higher expression levels of the ABCA7 in the peripheral blood may reflect immunological changes in schizophrenia as well as the AD. However, the expression levels of the ABCA7 in schizophrenia were not associated with the duration of illness in contrast to that of the AD.15 This finding could result from a difference in the pathological process in the two diseases. That means the difference between the gradual progression in the older age in AD and the relatively acute onset in the younger age in schizophrenia. Additionally, the schizophrenia patients in this study were in the chronic stage. The brain atrophy or neurodegenerative process in the imaging tests of schizophrenia is much milder than that of AD. Therefore, it is possible that the changes of the ABCA7 mRNA expression levels in the progression of schizophrenia could be smaller than that of AD. In the future, the prospective study for subjects in various ages is needed to verify this suggestion.
Second, there were no differences in genotype and allele frequencies between 527 control subjects and 781 schizophrenia patients. Our result was consistent with the recent study of the largest study to date of East Asian participants (22,778 schizophrenia cases and 35,362 controls).28 In this study, the rs3764650 was not associated with the development of schizophrenia in the same East Asian population.
Third, the ABCA7 mRNA expression levels were significantly associated with the number of the G allele of the rs3764650 in the multiple regression analysis. This result was not found in the study of AD.15 Although this finding supported relationships between the rs3764650 and gene expression, it is not consistent with this case–control study in 527 control subjects and 781 schizophrenia patients. Therefore, the rs3764650 may be related to the risk of schizophrenia through the ABCA7 mRNA expression levels, although the rs3764650 is not an independent risk factor of schizophrenia.
From these results, rs3764650 was related to the ABCA7 expression levels of schizophrenia, not the onset of schizophrenia. However, we considered that the ABCA7 played similar roles in schizophrenia and AD, because the ABCA7 expression levels of schizophrenia were higher than control subjects as well as AD.15 Therefore, we consider that the pathological conditions of schizophrenia are related to the dysfunctions of autoimmune system and elevated inflammations, and the higher ABCA7 expression levels of schizophrenia are compensatory mechanisms of these dysfunctions and inflammations. Because the higher ABCA7 expression levels of schizophrenia in the chronic stage were related to the risk alleles G of rs3764650, these compensatory mechanisms may work stronger in schizophrenia with the G alleles of rs3764650. However, this consideration should be confirmed by each stage of schizophrenia including the prodrome and early stages.
It should be noted that this study was cross-sectional. Because this notion was the limitation of this study and there were few reports for relationships between the ABCA7 and schizophrenia, we should recruit schizophrenia patients in various stages, the chronic and the acute stage of schizophrenia. Because this study included relatively old subjects, the possibility that the aging process has some effects on mRNA expression level could not be ruled out. Moreover, it should be concerned with the small sample sizes, especially the mRNA analysis. Due to the limited power, the negative findings in this study cannot be interpreted as no associations.
We concluded that the ABCA7 mRNA expression levels in peripheral blood associated with the rs3764650 genotypes may be related to pathological mechanisms in schizophrenia and may be a marker for schizophrenia.
Acknowledgments
We thank Ms. Chiemi Onishi for her technical assistance. This work was partially supported by a Health and Labor Science Research Grant from the Japanese Ministry of Health, Labor and Welfare and a Grant-in-Aid for Scientific Research from the Japanese Ministry of Education, Culture, Sports, Science and Technology (JSPS KAKENHI Grant Numbers 17K16381, 18K07564 and 18K15518).
Disclosure
Prof. Dr. Tetsuro Ohmori report grants and personal fees from Dainippon Sumitomo, Eizai, Otsuka, and Pfizer, grants from Novartis, Takeda, Tanabe Mitsubishi, Astellas, personal fees from Meiji, MSD, Jansen, Yoshitomi, Mochida, Kyowa, Elly Lilly, Daiichi Sankyo, outside the submitted work. The authors report no other conflicts of interest in this work.
References
1. Pantelis C, Velakoulis D, McGorry PD, et al. Neuroanatomical abnormalities before and after onset of psychosis: a cross-sectional and longitudinal MRI comparison. Lancet. 2003;361:281–288. doi:10.1016/S0140-6736(03)12323-9
2. Sun D, Phillips L, Velakoulis D, et al. Progressive brain structural changes mapped as psychosis develops in ‘at risk’ individuals. Schizophr Res. 2009;108:85–92. doi:10.1016/j.schres.2008.11.026
3. Takata A, Matsumoto N, Kato T. Genome-wide identification of splicing QTLs in the human brain and their enrichment among schizophrenia-associated loci. Nat Commun. 2017;8:14519. doi:10.1038/ncomms14519
4. Hollingworth P, Harold D, Sims R, et al. Common variants at ABCA7, MS4A6A/MS4A4E, EPHA1, CD33 and CD2AP are associated with Alzheimer’s disease. Nat Genet. 2011;43:429–435. doi:10.1038/ng.803
5. Lambert JC, Heath S, Even G, et al. Genome-wide association study identifies variants at CLU and CR1 associated with Alzheimer’s disease. Nat Genet. 2009;41:1094–1099. doi:10.1038/ng.439
6. Naj AC, Jun G, Reitz C, et al. Effects of multiple genetic loci on age at onset in late-onset Alzheimer disease: a genome-wide association study. JAMA Neurol. 2014;71:1394–1404. doi:10.1001/jamaneurol.2014.1491
7. Seshadri S, Fitzpatrick AL, Ikram MA, et al. Genome-wide analysis of genetic loci associated with Alzheimer disease. JAMA. 2010;303:1832–1840. doi:10.1001/jama.2010.574
8. Harold D, Abraham R, Hollingworth P, et al. Genome-wide association study identifies variants at CLU and PICALM associated with Alzheimer’s disease. Nat Genet. 2009;41:1088–1093. doi:10.1038/ng.440
9. Jehle AW, Gardai SJ, Li S, et al. ATP-binding cassette transporter A7 enhances phagocytosis of apoptotic cells and associated ERK signaling in macrophages. J Cell Biol. 2006;174:547–556. doi:10.1083/jcb.200601030
10. Kim WS, Guillemin GJ, Glaros EN, Lim CK, Garner B. Quantitation of ATP-binding cassette subfamily-A transporter gene expression in primary human brain cells. Neuroreport. 2006;17:891–896. doi:10.1097/01.wnr.0000221833.41340.cd
11. Kim WS, Weickert CS, Garner B. Role of ATP-binding cassette transporters in brain lipid transport and neurological disease. J Neurochem. 2008;104:1145–1166. doi:10.1111/jnc.2008.104.issue-5
12. Kim WS, Li H, Ruberu K, et al. Deletion of Abca7 increases cerebral amyloid-beta accumulation in the J20 mouse model of Alzheimer’s disease. J Neurosci. 2013;33:4387–4394.
13. Wang N, Lan D, Gerbod-Giannone M, et al. ATP-binding cassette transporter A7 (ABCA7) binds apolipoprotein A-I and mediates cellular phospholipid but not cholesterol efflux. J Biol Chem. 2003;278:42906–42912. doi:10.1074/jbc.M307831200
14. Vasquez JB, Fardo DW, Estus S. ABCA7 expression is associated with Alzheimer’s disease polymorphism and disease status. Neurosci Lett. 2013;556:58–62. doi:10.1016/j.neulet.2013.09.058
15. Yamazaki K, Yoshino Y, Mori T, et al. Gene expression and methylation analysis of ABCA7 in patients with Alzheimer’s disease. J Alzheimers Dis. 2017;57:171–181. doi:10.3233/JAD-161195
16. Jones AL, Mowry BJ, Pender MP, Greer JM. Immune dysregulation and self-reactivity in schizophrenia: do some cases of schizophrenia have an autoimmune basis? Immunol Cell Biol. 2005;83:9–17. doi:10.1111/imcb.2005.83.issue-1
17. Nunes SO, Matsuo T, Kaminami MS, Watanabe MA, Reiche EM, Itano EN. An autoimmune or an inflammatory process in patients with schizophrenia, schizoaffective disorder, and in their biological relatives. Schizophr Res. 2006;84:180–182. doi:10.1016/j.schres.2006.02.003
18. Potvin S, Stip E, Sepehry AA, Gendron A, Bah R, Kouassi E. Inflammatory cytokine alterations in schizophrenia: a systematic quantitative review. Biol Psychiatry. 2008;63:801–808. doi:10.1016/j.biopsych.2007.09.024
19. Chen X, Sun C, Chen Q, et al. Apoptotic engulfment pathway and schizophrenia. PLoS One. 2009;4:e6875. doi:10.1371/journal.pone.0006875
20. Carrol ED, Salway F, Pepper SD, et al. Successful downstream application of the Paxgene Blood RNA system from small blood samples in paediatric patients for quantitative PCR analysis. BMC Immunol. 2007;8:20. doi:10.1186/1471-2172-8-20
21. Hu N, Tan MS, Yu JT, et al. Increased expression of TREM2 in peripheral blood of Alzheimer’s disease patients. J Alzheimers Dis. 2014;38:497–501. doi:10.3233/JAD-130854
22. Watanabe S, Iga J, Nishi A, et al. Microarray analysis of global gene expression in leukocytes following lithium treatment. Hum Psychopharmacol. 2014;29:190–198. doi:10.1002/hup.v29.2
23. Watanabe SY, Iga J, Ishii K, et al. Biological tests for major depressive disorder that involve leukocyte gene expression assays. J Psychiatr Res. 2015;66–67:1–6. doi:10.1016/j.jpsychires.2015.03.004
24. Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods. 2001;25:402–408. doi:10.1006/meth.2001.1262
25. Kanda Y. Investigation of the freely available easy-to-use software ‘EZR’ for medical statistics. Bone Marrow Transplant. 2013;48:452–458. doi:10.1038/bmt.2012.244
26. Jacomb I, Stanton C, Vasudevan R, et al. C-reactive protein: higher during acute psychotic episodes and related to cortical thickness in schizophrenia and healthy controls. Front Immunol. 2018;9:2230. doi:10.3389/fimmu.2018.02230
27. Zhou Y, Peng W, Wang J, Zhou W, Zhou Y, Ying B. Plasma levels of IL-1Ra are associated with schizophrenia. Psychiatry Clin Neurosci. 2019;73:109–115. doi:10.1111/pcn.12794
28. Lam M, Chen CY, Li Z, et al. Comparative genetic architectures of schizophrenia in East Asian and European populations. Nat Genet. 2019;51:1670–1678. doi:10.1038/s41588-019-0512-x
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.