Back to Journals » Infection and Drug Resistance » Volume 13
A Vaccine Against Group B Streptococcus: Recent Advances
Authors Carreras-Abad C ![]() , Ramkhelawon L
, Ramkhelawon L ![]() , Heath PT
, Heath PT ![]() , Le Doare K
, Le Doare K
Received 10 October 2019
Accepted for publication 10 March 2020
Published 29 April 2020 Volume 2020:13 Pages 1263—1272
DOI https://doi.org/10.2147/IDR.S203454
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Dr Sahil Khanna
Clara Carreras-Abad,1,2 Laxmee Ramkhelawon,1 Paul T Heath,1 Kirsty Le Doare1,3,4
1Paediatric Infectious Diseases Research Group and Vaccine Institute, Institute for Infection and Immunity, St George’s, University of London, London, UK; 2Department of Paediatrics, Obstetrics and Gynecology and Preventive Medicine and Public Health, Universitat Autònoma de Barcelona, Barcelona, Spain; 3Pathogen Immunity Group, Public Health England, Porton Down,UK; 4Medical Research Council/Uganda Virus Research Institute and London School of Hygiene and Tropical Medicine, Uganda Research Unit, Entebbe, Uganda
Correspondence: Clara Carreras-Abad
Paediatric Infectious Diseases Research Group, Institute for Infection and Immunity, St George’s, University of London, Jenner Wing, Level 2 Mailpoint J2C, Cranmer Terrace, London SW17 0RE, UK
Email [email protected]
Abstract: Group B streptococcus (GBS) causes a high burden of neonatal and infant disease globally. Implementing a vaccine for pregnant women is a promising strategy to prevent neonatal and infant GBS disease and has been identified as a priority by the World Health Organisation (WHO). GBS serotype-specific polysaccharide – protein conjugate vaccines are at advanced stages of development, but a large number of participants would be required to undertake Phase III clinical efficacy trials. Efforts are therefore currently focused on establishing serocorrelates of protection in natural immunity studies as an alternative pathway for licensure of a GBS vaccine, followed by Phase IV studies to evaluate safety and effectiveness. Protein vaccines are in earlier stages of development but are highly promising as they might confer protection irrespective of serotype. Further epidemiological, immunological and health economic studies are required to enable the vaccine to reach its target population as soon as possible.
Keywords: Group B streptococcus, Streptococcus agalactiae, maternal vaccines, maternal immunisation, neonatal sepsis, infant sepsis
Introduction
Group B streptococcus (GBS) is a leading cause of neonatal and infant sepsis and meningitis globally.1–3 GBS can also cause stillbirths, prematurity and disease in pregnant women, immunocompromised adults and the elderly but the highest incidence of disease is in neonates and young infants.4
A systematic review and meta-analysis conducted in 2017 estimated a global incidence of invasive infant GBS disease of 0.49 (95%Cl 0.43–0.56) per 1000 live births.5 In 2015, GBS was estimated to have caused 319,000 cases of invasive neonatal GBS disease globally, resulting in 90,000 deaths.3 Serotypes Ia, Ib, II, III and V account for 98% of all rectovaginal colonisation in pregnant women worldwide.6 The most frequent GBS serotype causing disease in infants is serotype III (61.5%) followed by Ia (19.1%), V (6.7%) and Ib (5.7%).5 However, the fulminating nature of disease during the first hours of life and the technical difficulties in making an etiological diagnosis in many low- and middle-income settings means that this might represent a significant underestimation of the true GBS disease burden.7 Epidemiological data on the burden of GBS disease, especially from African countries, where most infant deaths from all-cause sepsis occur, is urgently required.7 Infant mortality estimates are seven times higher in WHO African region (51 per 1000 live births) compared to WHO European region (7 per 1000 live births).8
Intrapartum Antibiotic Prophylaxis (IAP) has reduced the incidence of early onset disease (EOD, occurring from day 0 to 6 of life) in many countries using these strategies, especially those that screen all pregnant women for GBS rectovaginal colonisation during late pregnancy and give IAP to all GBS-colonised women regardless of presenting risk factors.9,10 However, IAP coverage is incomplete even in the best of settings,11 has no impact on late onset disease (LOD, occurring from day 7 to 90 of life), stillbirths and prematurity due to GBS, as well as a limited impact on disease in pregnant women.10,12 Widespread IAP use might also be an issue in the context of international efforts to control antimicrobial resistance. Furthermore, antibiotics might have an effect on the infant gut flora. Effects of early life events on the neonatal microbiome have been associated with increased rates of allergy, asthma and obesity.13–15
Novel Features of a Maternal Vaccine for GBS
A suitable vaccine against GBS given to pregnant women could provide effective protection to those forms of invasive disease that cannot be prevented with IAP or where IAP is not feasible or is incomplete. Furthermore, a vaccine would be more easily accessible than GBS culture in all settings and would avoid the need for antimicrobial administration, avoiding the potential negative consequences of IAP in the long term.
Maternal immunisation is already a successful tool to prevent tetanus,16 influenza17 and pertussis18 in young infants. The placental transfer of maternal antibodies from mother to infant reduces the window of susceptibility to infections during the first months of life.19 This same rationale has been used to investigate new vaccines against common infections, such as respiratory syncytial virus (RSV) and GBS.20 A major characteristic of these new vaccines is that they are being specifically designed for pregnant women.20
Vaccine Development: Overview of Current Efforts
During the 2015 World Health Organisation (WHO) Product Development for Vaccines Advisory Committee meeting, GBS was identified as a high priority for the development of a vaccine for maternal immunisation because of the major public health burden posed by GBS in low- and middle-income countries (LMIC), and the high technical feasibility for successful development.21 Recent estimates suggest that an effective GBS maternal vaccine (>80% efficacy), with high (90%) global coverage, could prevent 231,000 infant and maternal GBS cases, 41,000 stillbirths and 66,000 infant deaths annually.3
Evidence suggests that maternal immunisation with protein-conjugated GBS capsular polysaccharides may reduce the disease risk in neonates and young infants in a serotype-specific manner.22–24 In addition, as there are proteins that can be present in different serotypes, protein-based vaccines have the potential to provide protection across the serotype spectrum. These are also under evaluation.25,26 Table 1 summarizes the development status of current vaccine candidates.
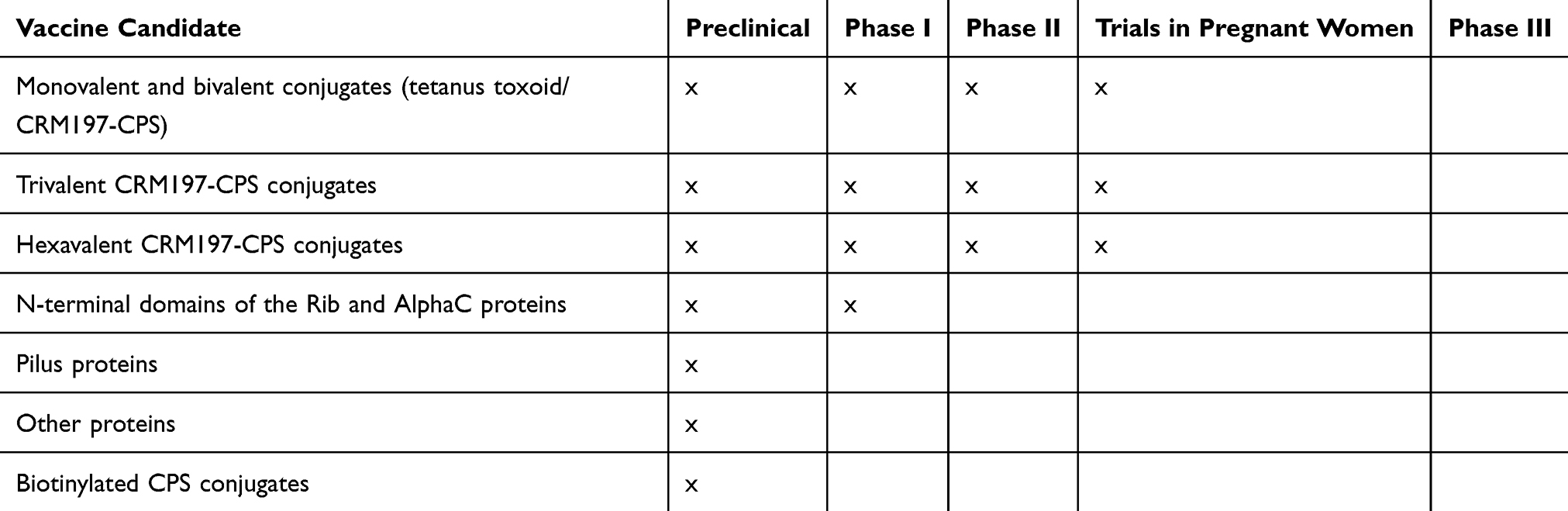 |
Table 1 Summary of Different Vaccine Candidates |
Capsular Polysaccharide Conjugate Vaccines
A number of virulence factors expressed by GBS are involved in colonisation, adherence, invasion and immune evasion27–29 and these could be used as potential vaccine candidates. One of the most well-studied virulence factors of GBS is its unique sialic acid-rich capsular polysaccharide (CPS) which inhibits complement deposition and protects the bacteria from opsonophagocytosis by immune cells, thus contributing to the evasion of host immune defense mechanisms.30,31 The CPS also enhances biofilm formation, inhibits the binding of antimicrobial peptides and neutrophil extracellular traps (NET) as well as disturbing bacterial adherence to the epithelium and mucus, thus increasing GBS invasiveness.32–38
GBS expresses 10 types of CPS (Ia, Ib, II–IX) that are structurally and antigenically different.39–41 Variously arranged monosaccharides and a sialic acid residue on the branching terminus of the repeating unit make up the CPS. According to recent meta-analyses, 97% of invasive isolates in all geographical regions are caused by five of the most common serotypes of GBS (Ia, Ib, II, III and V) with serotype III the most commonly found cause of disease in infants.5 Serotype IV is an emerging and increasing cause of invasive disease, especially in non-pregnant adults, with the potential to become an important cause of neonatal disease, with some cases already reported.42–45
As polysaccharides are T-cell independent antigens, the polysaccharide is conjugated to a protein carrier in order to trigger both a protective and a memory B-cell response. Earlier vaccines were conjugated to a tetanus toxoid, such vaccines might have particular value in LMIC where neonatal tetanus is still a concern.46,47 However, the main carrier protein currently used is CRM197, a nontoxic mutant of diphtheria toxin, which is highly immunogenic. Studies using either carrier protein demonstrated better immunogenicity with high levels of antibodies with CPS-conjugates compared to unconjugated vaccines.48,49
The first clinical trials were conducted with monovalent vaccines (Ia, Ib, II, III and V).48–52 However, single serotypes do not generally produce cross-reactive immunity against other serotypes, thus multivalent vaccines were developed. A phase I/II clinical trial (registered as NCT01193920 at the ClinicalTrials.gov database) of a trivalent (Ia, Ib and III) CRM197 conjugate vaccine in pregnant women reported higher levels of CPS-specific antibodies in infants at birth and no safety concerns .53 In 2017, a clinical trial of a GBS polysaccharide conjugate vaccine targeting serotypes Ia, Ib, II, III and V was started but, more recently, in view of the increase of disease caused by serotype IV, this serotype was added to create an hexavalent vaccine (Ia, Ib, II, III, IV, and V) with the aim to cover at least 98% of GBS isolates causing neonatal invasive disease (NCT03170609).5,54 In order to verify the clinical safety and immunogenicity of this hexavalent vaccine, further clinical trials will be required. Currently, a phase I/II clinical trial (NCT03765073) is being conducted in South Africa to evaluate the safety, tolerability and immunogenicity of a hexavalent vaccine in healthy non-pregnant and pregnant women.
It has been shown that opsonophagocytosis is the main mechanism for the host to clear GBS infection.24 A recent Phase II study (NCT01446289) demonstrated that maternal antibodies of pregnant women vaccinated with a trivalent glycoconjugated vaccine composed of CPS Ia, Ib and III result in opsono-phagocytic killing (OPK) titers against each GBS serotype. Analysis of cord sera revealed a strong positive correlation between IgG concentrations and OPK titers, which is predictive of functional activity against GBS infection.55
The role of anti-capsular antibodies in preventing GBS maternal colonisation, as well as ascending infection and neonatal transmission was recently evaluated in an animal model.56 Results show that systemic immunisation with a type III CRM197-glycoconjugate vaccine produces high levels of IgG that can reduce vaginal acquisition of serotype III during pregnancy.56 Studies in pregnant women showed the same association with natural immunity.57,58 Further studies will be needed to confirm the same results in vaccinated women.
Few studies have evaluated the number of doses that will be required per pregnancy for full immunity. In one study in healthy, non-pregnant women, no increase in antibody levels was shown after a second dose of a trivalent CRM197-glycoconjugate vaccine administered one month after the first dose.59 A recent study published in 2019 (NCT02690181) evaluated the safety and immunogenicity of a second dose of a trivalent (Ia, Ib and III) CRM197 conjugate vaccine in non-pregnant women over a long period of time (4 to 6 years) after the administration of the first dose. Antibody levels from previously GBS-vaccinated women increased ≥200-fold after a second dose. Women presenting with undetectable antibody levels after first dose also experienced an increase of anti-GBS concentrations after a second dose.60 These results suggest that further doses might be required in subsequent pregnancies.
Serocorrelates of Protection
Although several vaccine candidates are undergoing preclinical and clinical trials, in order to achieve licensure of a vaccine for GBS, phase III clinical trials may be required. These would need to be very large in order to demonstrate efficacy in countries able to conduct such trials.61 As there is a good correlation between immune response and clinical protection, licensure of a GBS vaccine could also be based on antibody measurement, if a specific antibody response can be correlated with protection.62–65 A review undertaken in 2019 synthesised the scientific evidence to define a serocorrelate of protection against GBS disease based on studies of natural infection.66 In such a scenario, a phase IV study will be required post-licensure to assess effectiveness. This is the same approach used for meningococcal B vaccine licensure in the United Kingdom.67
Assays for Antibody Quantification and Evaluation
The concentration of antibodies against serotype-specific CPS antigens can be quantified using standard immunogenicity assays (IA). However, there are several limitations to using current IA as the concentration measurement is very dependent on how well CPS is immobilized on an ELISA plate. There are other technical difficulties including inconsistent binding of immobilized CPS to the solid phase or a nonspecific serotype-independent binding, of antibodies with lower avidity.49 There has been much debate about the methods of CPS-binding, for example that used in pneumococcal assays (poly-L-lycine) or the novel biotin-streptavidin methods.66,68,69 The biotinylated method has the advantage of being able to use mass spectroscopy to determine the exact binding site of the biotin to the CPS, enabling the monitoring of any potential conformational changes to the CPS.69 However, whether this affects the performance of the assay is unknown.
The radio-antigen binding assay (RABA) had been used successfully to quantify levels of anti-GBS antibody as it measures antibody in its native state;70 however, as with most assays, there are several limitations, including low detection sensitivity, difficulties in obtaining and using radioisotopes and limited ability to quantify IgG isotopes. Therefore, it is imperative for techniques quantifying capsular serotype-specific antibodies in serum to be sensitive as well as serotype-specific.71 Multiplex immunoassays (MIAs) based on the Luminex technology are very useful in simultaneously quantifying the concentration of IgG antibodies against the capsule of multiple GBS serotypes. A Luminex-based direct immunoassay (dLIA) was recently developed for pneumococcal CPS.68 The latter could generate up to 143 test results in a single 96-well plate using similar principles to an ELISA assay for evaluating vaccines in clinical trials. MIAs quantifying IgG antibodies against the six most frequent GBS capsular variants (Ia, Ib, II, III, IV and V) would prove to be extremely useful in the standardisation of the assay used for GBS vaccine development.
The functionality of antibodies may also have a significant role in protection against GBS infections. ELISAs are limited in this aspect as they cannot distinguish between antibodies with low avidity and those with high avidity. Therefore, the opsonophagocytosis killing assay (OPkA) enables the measurement of antibody functionality.72 For the validation of the pneumococcal vaccine, the granulocytic cell line (HL60) was used, making the assay more reproducible.73 The assay was also multiplexed, which proved to be advantageous as it was less time-consuming and the amount of serum needed for the assay was reduced.74,75 Although a multiplexed OPkA for GBS (GBS-MOPA) has been developed for use in newborns, it only targets serotypes Ia, III and V.76 Therefore, a GBS-OPA targeting all possible vaccine serotypes is necessary for future GBS vaccine development and evaluation.
Protein Vaccines
Although CPS-conjugate vaccines have been demonstrated to induce good immunogenicity, there are still several limitations, including potential immune interference with other types of conjugate vaccines such as Haemophilus influenzae type b, meningococcal and pneumococcal conjugate vaccines.77,78 There is also the possibility of serotype replacement and switching post vaccination, as well as an increase in non-encapsulated GBS strains.79–81 Alternative vaccine candidates include structurally conserved protein antigens which can induce a strong immune response against most GBS strains. In order to develop a vaccine that can confer broad protection against GBS, several studies have identified proteins common to all GBS strains. Members of the Alp family, including AlphaC, Alp1 (Epsilon), Alp2, Alp3 (R28), Alp4 and Rib, are the most well-known and abundant surface proteins. These proteins are expressed by different serotypes (Table 2).82–85 Although there have been preclinical vaccine investigations using AlphaC, Alp3 and Rib proteins, the heterogeneity of the Alp sequence restricts the use of Alp proteins as potential vaccine candidates.86,87 Nonetheless, a protein vaccine based on the highly immunogenic N-terminal domains of AlphaC and Rib (GBS-NN) has been studied in a Phase I clinical trial (NCT02459262). The participants included 240 healthy women who were immunised with one or two doses of GBS-NN, generating an elevated level of GBS-NN specific antibodies by over-30 fold in both groups.82,88 GBS expresses either one of two-allelic serine-rich repeat 1 (Srr1) and serine rich-repeat 2(Srr2) proteins,89 both of which can bind to fibrinogen Aα chain through the “dock, lock and latch” mechanism, thus contributing to the pathogenesis of GBS meningitis and GBS colonisation of the vaginal surface.89–91 The antigenic latch domain consisting of 13 amino acids containing both Srr1 and Srr2, was shown to play a significant role in GBS pathogenesis. Murine models have exhibited serotype-independent protection against GBS infection after being vaccinated with the latch-peptide vaccine.92 C5a peptidase, which is a GBS virulence factor, was also considered as a universal protein vaccine or a carrier for GBS-CPS.93 C5a peptidase encapsulated within microspheres composed of lactic acid and glycolic acid co-polymer triggered systemic and mucosal immune responses in murine models, thus protecting them against multiple GBS serotypes.94,95 Another type of surface protein are the pilus proteins, which, in preclinical and human studies, were found to induce immune responses against different GBS serotypes.30,96,97
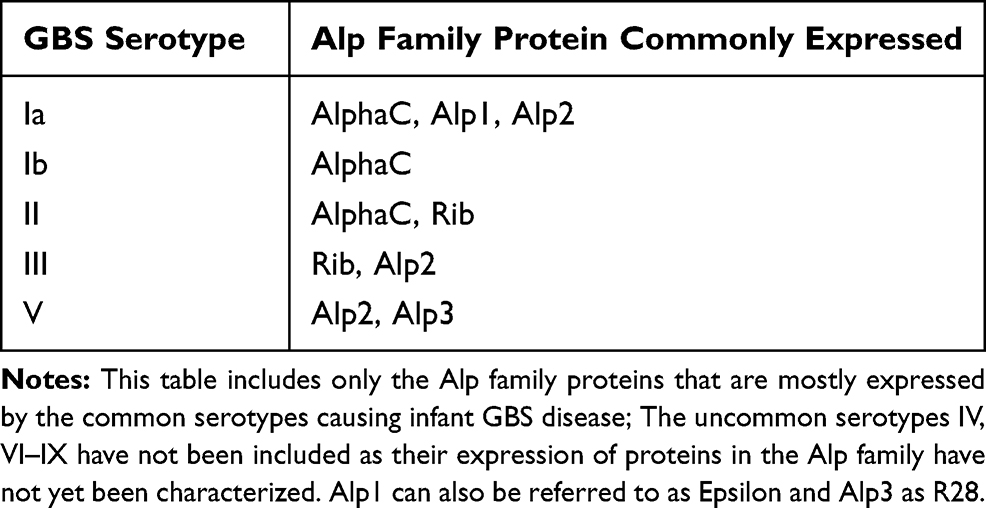 |
Table 2 Alp Family Proteins Commonly Expressed in Different GBS Serotypes |
Next Steps, New Perspectives
The most advanced vaccine candidates are hexavalent vaccines including serotypes Ia, Ib, II, III, IV, and V, which are now in phase II trials.54 Immunogenicity and safety of these candidates has been demonstrated in non-pregnant and, more recently, in pregnant women.22 Protein vaccines are in phase I trials including human studies in non-pregnant women.88,92,94-96
Several obstacles exist in moving the most advanced vaccines into phase III clinical trials. Given the relatively low incidence of GBS disease in Europe and the USA, large numbers of participants would be needed to determine vaccine efficacy.98 In addition, obstacles exist in determining what concentration of antibody is required to protect the infant for the duration of the period at risk (the 3 first months of life) as there are currently no internationally recognised correlates of protection with which to interpret individual study results.99 Therefore, a serocorrelate of protection against GBS is needed to accelerate the licensure of a vaccine. The standardisation of reagents to measure antibodies against GBS is crucial for the establishment of serological correlates of protection and for the development of GBS vaccines. The GASTON consortium was recently set up with this aim.66
On the other hand, efforts to identify common proteins to all GBS strains have been made in order to find a vaccine that confers protection against all GBS serotypes. Recent use of molecular techniques, such as multi-locus sequence type (MLST) and whole-genome sequencing (WGS), have allowed us to better characterise the GBS structure, as well as to identify the virulent lineages such as ST17 hypervirulent strain, strongly associated with serotype III. As it is important to understand the genetic lineages that are more likely to cause GBS disease in order to better define vaccine targets, a global genomic survey of GBS has been established (the JUNO project).100
Other Areas for Future Research
As mentioned above, it is important to establish rates of maternal colonisation and GBS disease worldwide, as well as to understand the relationship between colonisation and invasive infection, to assist assessments of vaccine efficacy. Regional serotype distribution is also required, especially from many LMIC were few data are currently available.
Once a vaccine is licensed, the number and timing of doses for optimum coverage during pregnancy and the number of doses required for full protection needs to be determined. There are other knowledge gaps remaining, including the placental transfer of vaccine-induced immune responses in special populations, such as women infected with HIV, malaria, syphilis and hepatitis B, among others. These infections, highly prevalent in LMIC, may alter the immune response to vaccines and impair antibody transfer across the placenta. A phase II trial using a GBS trivalent vaccine (Ia, Ib, III) undertaken in Malawi and South Africa among 270 pregnant women with or without HIV infection (NCT01412801) showed that the immune response to vaccines and serotype-specific antibody concentrations in infants at birth were lower in the HIV infected group.101 Maternal immunisation policies require understanding of the role of these endemic infections in generating immune responses that ensure adequate protection of infants in these challenging environments.102,103
Finally, cost-effectiveness evaluation is required. Cost-effectiveness studies have indicated that the predominant cost drivers are disease incidence, immunisation coverage and vaccine efficacy.104 In high income countries, where GBS disease is well characterized, it has been shown that a maternal vaccine would be more cost-effective compared to IAP and doing nothing.105 A population-based economic analysis in the USA concluded that vaccinating 80% of pregnant women with a vaccine that prevents 80% of cases among infants born at or after 34 weeks of gestation would prevent approximately 4100 neonatal cases annually with a net savings of 131 million USD.105 A study in South Africa also concluded that GBS maternal vaccination would be very cost-effective by WHO guidelines.106 This study reported that, assuming that vaccine efficacy varies from 50% to 90% with a 75% coverage, GBS immunisation alone, without IAP prevention, would prevent 30–54% of infant GBS cases compared to doing nothing. In contrast, risk factor based-IAP alone prevents 10% of infant GBS cases compared to doing nothing. Furthermore, at a vaccination cost between 10 and 30 USD, and mid-range efficacy, vaccine introduction costs range from 676 to 2390 USD per disability-adjusted life-year (DALY) averted, compared to doing nothing.106 A modeling study of different sub-Saharan African countries showed that maternal GBS immunisation could be a cost-effective intervention, with cost-effectiveness ratios similar to other recently introduced vaccines.107 37 African countries were clustered in four different groups according to their economic and health resources and public health outcomes. One country of each cluster was chosen as representative: Guinea-Bissau, Uganda, Nigeria and Ghana. At equal coverage to that of pregnant women that attend four or more antenatal visits and with vaccine efficacy of 70%, maternal vaccination would prevent one-third of GBS cases in Uganda and Nigeria, 42–43% in Guinea-Bissau and 55–57% in Ghana. For a vaccine price of 7 USD per dose, maternal vaccination would cost from 320 to 350 USD per DALY averted in Guinea-Bissau, Nigeria and Ghana, as well as 573 USD in Uganda. The vaccine would be less cost-effective in Uganda as neonatal mortality seems to be lower.107 A recent study by our group of cost-effectiveness of a potential hexavalent vaccine in the Gambia indicated that disease incidence was the key factor in determining cost-effectiveness in a low income setting as the cost of doing nothing is very inexpensive as infants would classically die at home without receiving treatment.108 These studies, together with more epidemiological data in LMIC, might raise the impact resulting from a vaccine prevention strategy.
Data provided in this review demonstrate that obtaining a vaccine for pregnant women is a promising strategy to prevent neonatal and infant GBS disease. Consensus among public health institutions and sponsors is now a priority to allow this breakthrough that will help reduce neonatal and infant mortality, especially in the most vulnerable populations.
Disclosure
Professor Paul T Heath and Dr Kirsty Le Doare are members of the WHO Scientific Advisory Group to provide inputs to the development of a value proposition for Group B Streptococcus (GBS) Vaccines. Dr Kirsty Le Doare has received funding from Pfizer in 2016 for an unrelated project. The authors report no other conflicts of interest in this work.
References
1. O’Sullivan CP, Lamagni T, Patel D, et al. Group B streptococcal disease in UK and Irish infants younger than 90 days, 2014–15: a prospective surveillance study. Lancet Infect Dis. 2019;19(1):83–90. doi:10.1016/S1473-3099(18)30555-3
2. Sigaúque B, Kobayashi M, Vubil D, et al. Invasive bacterial disease trends and characterization of group B streptococcal isolates among young infants in southern Mozambique, 2001–2015. PLoS One. 2018;13(1):e0191193. doi:10.1371/journal.pone.0191193
3. Seale AC, Bianchi-Jassir F, Russell NJ, et al. Estimates of the burden of Group B Streptococcal disease worldwide for pregnant women, stillbirths, and children. Clin Infect Dis. 2017;65:S200–19. doi:10.1093/cid/cix664
4. Raabe VN, Shane AL. Group B Streptococcus (Streptococcus agalactiae). Microbiol Spectr. 2019;7(2). doi:10.1128/microbiolspec.GPP3-0007-2018
5. Madrid L, Seale AC, Kohli-Lynch M, et al. Infant Group B streptococcal disease incidence and serotypes worldwide: systematic review and meta-analyses. J Infect Dis. 2017;65:S160–72. doi:10.1093/cid/cix656
6. Russell NJ, Seale AC, O’Driscoll M, et al. Maternal colonization with Group B Streptococcus and Serotype distribution worldwide: systematic review and meta-analyses. J Infect Dis. 2017;65:S100–11. doi:10.1093/cid/cix658
7. Lawn JE, Bianchi-Jassir F, Russell NJ, et al. Group B Streptococcal disease worldwide for pregnant women, stillbirths, and children: why, what, and how to undertake estimates? Clin Infect Dis. 2017;65:S89–99. doi:10.1093/cid/cix653
8. Unicef; World Health Organisation; The world Bank; United Nations. Levels and Trends in Child Mortality. Report 2019; 2019.
9. Verani JR, McGee L, Schrag SJ; Division of Bacterial Diseases, National Center for Immunization and Respiratory Diseases, Centers for Disease Control and Prevention (CDC). Prevention of perinatal group B streptococcal disease–revised guidelines from CDC, 2010. MMWR Morb Mortal Wkly Rep. 2010;59(RR–10):1–36.
10. Le Doare K, O’Driscoll M, Turner K, et al. Intrapartum antibiotic chemoprophylaxis policies for the prevention of Group B Streptococcal disease worldwide: systematic review. J Infect Dis. 2017;65:S143–51. doi:10.1093/cid/cix654
11. Schrag SJ, Verani JR. Intrapartum antibiotic prophylaxis for the prevention of perinatal group B streptococcal disease: experience in the United States and implications for a potential group B streptococcal vaccine. Vaccine. 2013;31:D20–D26. doi:10.1016/j.vaccine.2012.11.056
12. O’Connor KA. Group B Streptococcal disease in the era of intrapartum antibiotic prophylaxis. Clin Pediatr (Phila). 2007;40(6):361. doi:10.1177/000992280104000614
13. Castanys-Muñoz E, Martin MJ, Vazquez E. Building a beneficial microbiome from birth. Adv Nutr. 2016;7(2):323–330. doi:10.3945/an.115.010694
14. Kerr CA, Grice DM, Tran CD, et al. Early life events influence whole-of-life metabolic health via gut microflora and gut permeability. Crit Rev Microbiol. 2015;41:326–340. doi:10.3109/1040841X.2013.837863
15. Kumar M, Singh P, Murugesan S, et al. Microbiome as an immunological modifier. Methods Mol Biol. 2020;595–638.
16. Thwaites CL, Beeching NJ, Newton CR. Maternal and neonatal tetanus. Lancet. 2015;385(9965):362–370. doi:10.1016/S0140-6736(14)60236-1
17. Madhi SA, Cutland CL, Kuwanda L, et al. Influenza vaccination of pregnant women and protection of their infants. N Engl J Med. 2014;371(10):918–931. doi:10.1056/NEJMoa1401480
18. Amirthalingam G, Andrews N, Campbell H, et al. Effectiveness of maternal pertussis vaccination in England: an observational study. Lancet. 2014;384(9953):1521–1528. doi:10.1016/S0140-6736(14)60686-3
19. Davies HG, Carreras-Abad C, Le Doare K, Heath PT. Group B Streptococcus: trials and Tribulations. Pediatr Infect Dis J. 2019;38(6SSuppl 1):S72–6. doi:10.1097/INF.0000000000002328
20. Bardají A, MacDonald NE, Omer SB, Aguado T. Maternal immunization: a call to accelerate progress. Vaccine. 2019;37(22):2882–2883. doi:10.1016/j.vaccine.2019.04.031
21. Vekemans J, Moorthy V, Friede M, et al. Maternal immunization against Group B streptococcus: World Health Organization research and development technological roadmap and preferred product characteristics. Vaccine. 2018.
22. Heath PT. Status of vaccine research and development of vaccines for GBS. Vaccine. 2016;34(26):2876–2879. doi:10.1016/j.vaccine.2015.12.072
23. Dangor Z, Lala SG, Kwatra G, Madhi SA. Group B Streptococcus: developing a correlate of protection for a vaccine against neonatal infections. Curr Opin Infect Dis. 2016;29(3):262–267. doi:10.1097/QCO.0000000000000266
24. Fabbrini M, Rigat F, Rinaudo CD, et al. The protective value of maternal Group B Streptococcus antibodies: quantitative and functional analysis of naturally acquired responses to capsular polysaccharides and pilus proteins in European maternal sera. Clin Infect Dis. 2016;63(6):746–753. doi:10.1093/cid/ciw377
25. Dangor Z, Kwatra G, Izu A, et al. Association between maternal Group B Streptococcus surface-protein antibody concentrations and invasive disease in their infants. Expert Rev Vaccines. 2015;14(12):1651–1660. doi:10.1586/14760584.2015.1085307
26. World Health Organization (WHO). WHO preferred product characteristics for Group B Streptococcus vaccines. 2017;18. Available from: http://www.who.int/immunization/documents/en/.
27. Rajagopal L. Understanding the regulation of Group B Streptococcal virulence factors. Future Microbiol. 2009;4(2):201–221. doi:10.2217/17460913.4.2.201
28. Herbert MA, Beveridge CJE, Saunders NJ. Bacterial virulence factors in neonatal sepsis: group B streptococcus. Curr Opin Infect Dis. 2004;17:225–229. doi:10.1097/00001432-200406000-00009
29. Vornhagen J, Adams Waldorf KM, Rajagopal L. Perinatal Group B Streptococcal infections: virulence factors, immunity, and prevention strategies. Trends Microbiol. 2017;25:919–931. doi:10.1016/j.tim.2017.05.013
30. Nuccitelli A, Rinaudo CD, Maione D. Group B Streptococcus vaccine: state of the art. Ther Adv Vaccin. 2015;3(3):76–90. doi:10.1177/2051013615579869
31. Korir ML, Manning SD, Davies HD. Intrinsic maturational neonatal immune deficiencies and susceptibility to group B streptococcus infection. Clin Microbiol Rev. 2017;30:973–989. doi:10.1128/CMR.00019-17
32. Rosini R, Margarit I. Biofilm formation by Streptococcus agalactiae: influence of environmental conditions and implicated virulence factor. Front Cell Infect Microbiol. 2015;5. doi:10.3389/fcimb.2015.00006
33. Di Xia F, Mallet A, Caliot E, Gao C, Trieu-Cuot P, Dramsi S. Capsular polysaccharide of Group B Streptococcus mediates biofilm formation in the presence of human plasma. Microbes Infect. 2015;17(1):71–76. doi:10.1016/j.micinf.2014.10.007
34. D’Urzo N, Martinelli M, Pezzicoli A, et al. Acidic pH strongly enhances in vitro biofilm formation by a subset of hypervirulent ST-17 Streptococcus agalactiae strains. Appl Environ Microbiol. 2014;80(7):2176–2185. doi:10.1128/AEM.03627-13
35. Derré-Bobillot A, Cortes-Perez NG, Yamamoto Y, et al. Nuclease A (Gbs0661), an extracellular nuclease of Streptococcus agalactiae, attacks the neutrophil extracellular traps and is needed for full virulence. Mol Microbiol. 2013;89(3):518–531. doi:10.1111/mmi.2013.89.issue-3
36. Boldenow E, Gendrin C, Ngo L, et al. Group B Streptococcus circumvents neutrophils and neutrophil extracellular traps during amniotic cavity invasion and preterm labor. Sci Immunol. 2016;1(4). doi:10.1126/sciimmunol.aah6817.
37. Kothary V, Doster RS, Rogers LM, et al. Group B streptococcus induces neutrophil recruitment to gestational tissues and elaboration of extracellular traps and nutritional immunity. Front Cell Infect Microbiol. 2017;7. doi:10.3389/fcimb.2017.00019
38. Amulic B, Hayes G. Neutrophil extracellular traps. Curr Biol. 2011;21:R297–R298. doi:10.1016/j.cub.2011.03.021
39. Ferrieri P, Flores AE. Surface protein expression in Group B streptococcal invasive isolates. Adv Exp Med Biol. 1997;635–637.
40. Slotved HC, Kong F, Lambertsen L, Sauer S, Gilbert GL. Serotype IX, a proposed new Streptococcus agalactiae serotype. J Clin Microbiol. 2007;45(9):2929–2936. doi:10.1128/JCM.00117-07
41. Berti F, Campisi E, Toniolo C, et al. Structure of the type IX Group B Streptococcus capsular polysaccharide and its evolutionary relationship with types V and VII. J Biol Chem. 2014;289(34):23437–23448. doi:10.1074/jbc.M114.567974
42. Teatero S, Athey TBT, Van Caeseele P, et al. Emergence of serotype IV group B streptococcus adult invasive disease in Manitoba and saskatchewan, Canada, is driven by clonal sequence type 459 strains. J Clin Microbiol. 2015;53(9):2919–2926. doi:10.1128/JCM.01128-15
43. Ferrieri P, Lynfield R, Creti R, Flore AE. Serotype IV and invasive group B streptococcus disease in neonates, Minnesota, USA, 2000-20101. Emerg Infect Dis. 2013;19(4):553–558. doi:10.3201/eid1904.121572
44. Puopolo KM, Madoff LC. Type IV neonatal early-onset group B streptococcal disease in a United States hospital. J Clin Microbiol. 2007;45(4):1360–1362. doi:10.1128/JCM.02487-06
45. Martins ER, Pessanha MA, Ramirez M, et al. Analysis of group B streptococcal isolates from infants and pregnant women in Portugal revealing two lineages with enhanced invasiveness. J Clin Microbiol. 2007;45(10):3224–3229. doi:10.1128/JCM.01182-07
46. Kobayashi M, Vekemans J, Baker CJ, Ratner AJ, Le Doare K, Schrag SJ. Group B Streptococcus vaccine development: present status and future considerations, with emphasis on perspectives for low and middle income countries. F1000Research. 2016;5:2355. doi:10.12688/f1000research
47. Bröker M. Potential protective immunogenicity of tetanus toxoid, diphtheria toxoid and Cross Reacting Material 197 (CRM197) when used as carrier proteins in glycoconjugates. Hum Vaccin Immunother. 2016;12:664–667. doi:10.1080/21645515.2015.1086048
48. Baker CJ, Rench MA, McInnes P. Immunization of pregnant women with group B streptococcal type III capsular polysaccharide-tetanus toxoid conjugate vaccine. Vaccine. 2003;21:3468–3472. doi:10.1016/S0264-410X(03)00353-0
49. Baker CJ, Paoletti LC, Rench MA, Guttormsen H, Edwards MS, Kasper DL. Immune Response of Healthy Women to 2 Different Group B Streptococcal Type V Capsular Polysaccharide–Protein Conjugate Vaccines. J Infect Dis. 2004;189(6):1103–1112. doi:10.1086/jid.2004.189.issue-6
50. Baker CJ, Paoletti LC, Wessels MR, Guttormsen HK, Rench MA, Hickman ME. Safety and immunogenicity of capsular polysaccharide-tetanus toxoid conjugate vaccines for group B streptococcal types Ia and Ib. J Infect Dis. 1999;179(1):142–150. doi:10.1086/314574
51. Baker CJ, Paoletti LC, Rench MA, et al. Use of capsular polysaccharide–tetanus toxoid conjugate vaccine for Type II Group B Streptococcus in Healthy Women. J Infect Dis. 2000;182(4):1129–1138. doi:10.1086/jid.2000.182.issue-4
52. Kasper DL, Paoletti LC, Wessels MR, et al. Immune response to type III group B streptococcal polysaccharide- tetanus toxoid conjugate vaccine. J Clin Invest. 1996;98(10):2308–2314. doi:10.1172/JCI119042
53. Madhi SA, Koen A, Cutland CL, et al. Antibody kinetics and response to routine vaccinations in infants born to women who received an investigational trivalent Group B Streptococcus Polysaccharide CRM197 -Conjugate Vaccine during Pregnancy. Clin Infect Dis. 2017;65(11):1897–1904. doi:10.1093/cid/cix666
54. Buurman ET, Timofeyeva Y, Gu J, et al. A novel hexavalent capsular polysaccharide conjugate vaccine (GBS6) for the Prevention of Neonatal Group B Streptococcal Infections by Maternal Immunization. J Infect Dis. 2019;220(1):105–115. doi:10.1093/infdis/jiz062
55. Fabbrini M, Rigat F, Tuscano G, et al. Functional activity of maternal and cord antibodies elicited by an investigational group B Streptococcus trivalent glycoconjugate vaccine in pregnant women. J Infect. 2018;76(5):449–456. doi:10.1016/j.jinf.2018.01.006
56. Chiarot E, Spagnuolo A, Maccari S, et al. Protective effect of Group B Streptococcus type-III polysaccharide conjugates against maternal colonization, ascending infection and neonatal transmission in rodent models. Sci Rep. 2018;8(1). doi:10.1038/s41598-018-20609-5.
57. Kwatra G, Adrian PV, Shiri T, et al. Serotype-specific cell-mediated immunity associated with clearance of homotypic Group B Streptococcus Rectovaginal Colonization in Pregnant Women. J Infect Dis. 2016;213:1923–1926. doi:10.1093/infdis/jiw056
58. Le Doare K, Faal A, Jaiteh M, et al. Association between functional antibody against Group B Streptococcus and maternal and infant colonization in a Gambian cohort. Vaccine. 2017;35(22):2970–2978. doi:10.1016/j.vaccine.2017.04.013
59. Leroux-Roels G, Maes C, Willekens J, et al. A randomized, observer-blind Phase Ib study to identify formulations and vaccine schedules of a trivalent Group B Streptococcus vaccine for use in non-pregnant and pregnant women. Vaccine. 2016;34(15):1786–1791. doi:10.1016/j.vaccine.2016.02.044
60. Leroux-Roels G, Bebia Z, Maes C, et al. Safety and immunogenicity of a second dose of an investigational maternal trivalent Group B streptococcus vaccine in non-pregnant women 4–6 years after a first dose: results from a Phase 2 trial. Clin Infect Dis.
61. Kobayashi M, Schrag SJ, Alderson MR, et al. WHO consultation on group B Streptococcus vaccine development: report from a meeting held on 27–28 April 2016. Vaccine. 2016;37:7307–7314.
62. Lin FC, Philips JB
63. Lin FC, Weisman LE, Azimi PH, et al. Level of maternal IgG Anti–Group B Streptococcus Type III antibody correlated with protection of neonates against early‐onset disease caused by this pathogen. J Infect Dis. 2004;190(5):928–934. doi:10.1086/422756
64. Baker CJ, Carey VJ, Rench MA, et al. Maternal antibody at delivery protects neonates from early onset group B streptococcal disease. J Infect Dis. 2014;209(5):781–788. doi:10.1093/infdis/jit549
65. Carreras-Abad C, Cochet M, Hall T, et al. Developing a serocorrelate of protection against invasive group B streptococcus disease in pregnant women: a feasibility study. Health Technol Assess. 2019 Dec;23(67):1–40. doi:10.3310/hta23670
66. Le Doare K, Kampmann B, Vekemans J, et al. Serocorrelates of protection against infant group B streptococcus disease. Lancet Infect Dis. 2019;19(5):e162–71. doi:10.1016/S1473-3099(18)30659-5
67. Parikh SR, Andrews NJ, Beebeejaun K, et al. Effectiveness and impact of a reduced infant schedule of 4CMenB vaccine against group B meningococcal disease in England: a national observational cohort study. Lancet. 2016;388(10061):2775–2782. doi:10.1016/S0140-6736(16)31921-3
68. Pavliakova D, Giardina PC, Moghazeh S, et al. Development and validation of 13-plex luminex-based assay for measuring human serum antibodies to Streptococcus pneumoniae capsular polysaccharides. mSphere. 2018;3(4):e00128–18. doi:10.1128/mSphere.00128-18
69. Buffi G, Galletti B, Stella M, et al. Novel multiplex immunoassays for quantification of IgG against Group B Streptococcus Capsular Polysaccharides in Human Sera. mSphere. 2019;4(4). doi:10.1128/mSphere.00273-19.
70. Baker CJ, Kasper DL. Correlation of maternal antibody deficiency with susceptibility to neonatal Group B Streptococcal infection. N Engl J Med. 2010;294(14):753–756. doi:10.1056/NEJM197604012941404
71. Brigtsen AK, Kasper DL, Baker CJ, Jennings HJ, Guttormsen H. Induction of cross-reactive antibodies by immunization of healthy adults with Types Ia and Ib Group B Streptococcal Polysaccharide–Tetanus Toxoid Conjugate Vaccines. J Infect Dis. 2002;185(9):1277–1284. doi:10.1086/jid.2002.185.issue-9
72. Romero-Steiner S, Frasch CE, Carlone G, Fleck RA, Goldblatt D, Nahm MH. Use of opsonophagocytosis for serological evaluation of pneumococcal vaccines. Clin Vaccin Immunol. 2006;13:165–169. doi:10.1128/CVI.13.2.165-169.2006
73. Fleck RA, Romero-Steiner S, Nahm MH. Use of HL-60 cell line to measure opsonic capacity of pneumococcal antibodies. Clin Diagn Lab Immunol. 2005;12:19–27. doi:10.1128/CDLI.12.1.19-27.2005
74. Kim KH, Yu J, Nahm MH. Efficiency of a pneumococcal opsonophagocytic killing assay improved by multiplexing and by coloring colonies. Clin Diagn Lab Immunol. 2003;10(4):616–621. doi:10.1128/cdli.10.4.616-621.2003
75. Romero-Steiner S, Frasch C, Concepcion N, et al. Multilaboratory evaluation of a viability assay for measurement of opsonophagocytic antibodies specific to the capsular polysaccharides of Streptococcus pneumoniae. Clin Diagn Lab Immunol. 2003;10(6):1019–1024. doi:10.1128/cdli.10.6.1019-1024.2003
76. Choi MJ, Noh JY, Cheong HJ, et al. Development of a multiplexed opsonophagocytic killing assay (MOPA) for group B Streptococcus. Hum Vaccin Immunother. 2018;14(1):67–73. doi:10.1080/21645515.2017.1377379
77. Konini A, Kang M, Moghadas SM. Simulating immune interference on the effect of a bivalent glycoconjugate vaccine against Haemophilus influenzae Serotypes “a” and “b.”. Can J Infect Dis Med Microbiol. 2016;2016:1–8. doi:10.1155/2016/5486869
78. Borrow R, Dagan R, Zepp F, Hallander H, Poolman J. Glycoconjugate vaccines and immune interactions, and implications for vaccination schedules. Expert Rev Vaccin. 2011;10:1621–1631. doi:10.1586/erv.11.142
79. Flores AR, Galloway-Peña J, Sahasrabhojane P, et al. Sequence type 1 group B Streptococcus, an emerging cause of invasive disease in adults, evolves by small genetic changes. Proc Natl Acad Sci U S A. 2015;112(20):6431–6436. doi:10.1073/pnas.1504725112
80. Teatero S, Ferrieri P, Martin I, Demczuk W, CGeer A, Fittipaldi N. Serotype distribution, population structure, and antimicrobial resistance of group b streptococcus strains recovered from colonized pregnant women. J Clin Microbiol. 2017;55(2):412–422. doi:10.1128/JCM.01615-16
81. Bellais S, Six A, Fouet A, et al. Capsular switching in group B streptococcus CC17 hypervirulent clone: a future challenge for polysaccharide vaccine development. J Infect Dis. 2012;206(11):1745–1752. doi:10.1093/infdis/jis605
82. Areschoug T, Stålhammar-Carlemalm M, Larsson C, Lindahl G. Group B streptococcal surface proteins as targets for protective antibodies: identification of two novel proteins in strains of serotype V. Infect Immunol. 1999;67(12):6350–6357.
83. Furfaro LL, Chang BJ, Payne MS. Perinatal streptococcus agalactiae epidemiology and surveillance targets. Clin Microbiol Rev. 2018;31. doi:10.1128/CMR.00049-18
84. Lindahl G, Stålhammar-Carlemalm M, Areschoug T. Surface proteins of Streptococcus agalactiae and related proteins in other bacterial pathogens. Clin Microbiol Rev. 2005;18:102–127. doi:10.1128/CMR.18.1.102-127.2005
85. Smith TC, Roehl SA, Pillai P, Li S, Marrs CF, Foxman B. Distribution of novel and previously investigated virulence genes in colonizing and invasive isolates of Streptococcus agalactiae. Epidemiol Infect. 2007;135(6):1046–1054. doi:10.1017/S0950268806007515
86. Erdogan S, Fagan PK, Talay SR, et al. Molecular analysis of group B protective surface protein, a new cell surface protective antigen of group B streptococci. Infect Immunol. 2002;70(2):803–811. doi:10.1128/IAI.70.2.803-811.2002
87. Maeland JA, Afset JE, Lyng RV, Radtke A. Survey of immunological features of the alpha-like proteins of streptococcus agalactiae. Clin Vaccin Immunol. 2015;22:153–159. doi:10.1128/CVI.00643-14
88. Lin SM, Zhi Y, Ahn KB, Lim S, Seo HS. Status of group B streptococcal vaccine development. Clin Exp Vaccin Res. 2018;7(1):76–81. doi:10.7774/cevr.2018.7.1.76
89. Seo HS, Minasov G, Seepersaud R, et al. Characterization of fibrinogen binding by glycoproteins Srr1 and Srr2 of Streptococcus agalactiae. J Biol Chem. 2013;288(50):35982–35996. doi:10.1074/jbc.M113.513358
90. Seo HS, Mu R, Kim BJ, Doran KS, Sullam PM. Binding of Glycoprotein Srr1 of Streptococcus agalactiae to fibrinogen promotes attachment to brain endothelium and the development of meningitis. PLoS Pathog. 2012;8(10):e1002947. doi:10.1371/journal.ppat.1002947
91. Wang NY, Patras KA, Seo HS, et al. Group B streptococcal serine-rich repeat proteins promote interaction with fibrinogen and vaginal colonization. J Infect Dis. 2014;210:982–991. doi:10.1093/infdis/jiu151
92. Lin SM, Jang AY, Zhi Y, et al. Vaccination with a latch peptide provides serotypeindependent protection against group B streptococcus infection in mice. J Infect Dis. 2018;217(1):93–102. doi:10.1093/infdis/jix565
93. Bohnsack JF, Widjaja K, Ghazizadeh S, et al. A role for C5 and C5a-ase in the acute neutrophil response to Group B Streptococcal infections. J Infect Dis. 1997;175(4):847–855. doi:10.1086/jid.1997.175.issue-4
94. Santillan DA, Andracki ME, Hunter SK. Protective immunization in mice against group B streptococci using encapsulated C5a peptidase. Am J Obstet Gynecol. 2008;198(1):
95. Santillan DA, Rai KK, Santillan MK, Krishnamachari Y, Salem AK, Hunter SK. Efficacy of polymeric encapsulated C5a peptidasebased group B streptococcus vaccines in a murine model. Am J Obstet Gynecol. 2011;205:
96. Nuccitelli A, Cozzi R, Gourlay LJ, et al. Structure-based approach to rationally design a chimeric protein for an effective vaccine against Group B Streptococcus infections. Proc Natl Acad Sci U S A. 2011;108(25):10278–10283. doi:10.1073/pnas.1106590108
97. Edwards MS, Rench MA, Rinaudo CD, et al. Immune responses to invasive group B streptococcal disease in adults. Emerg Infect Dis. 2016;22(11):1877–1883. doi:10.3201/eid2211.160914
98. Madhi SA, Dangor Z, Heath PT, et al. Considerations for a phase-III trial to evaluate a group B Streptococcus polysaccharide-protein conjugate vaccine in pregnant women for the prevention of early- and late-onset invasive disease in young-infants. Vaccine. 2013;31:D52–D57. doi:10.1016/j.vaccine.2013.02.029
99. Heath PT, Culley FJ, Jones CE, et al. Group B streptococcus and respiratory syncytial virus immunisation during pregnancy: a landscape analysis. Lancet Infect Dis. 2017;17:e223–34. doi:10.1016/S1473-3099(17)30232-3
100. JUNO project [homepage on the Internet]. Available from: https://www.sanger.ac.uk/science/collaboration/juno-global-genomic-survey-streptococcus-agalactiae.
101. Heyderman RS, Madhi SA, French N, et al. Group B streptococcus vaccination in pregnant women with or without HIV in Africa: a non-randomised phase 2, open-label, multicentre trial. Lancet Infect Dis. 2016;16(5):546–555. doi:10.1016/S1473-3099(15)00484-3
102. Jones CE, Calvert A, Le Doare K. Vaccination in pregnancy - Recent developments. Pediatr Infect Dis J. 2018;37(2):191–193. doi:10.1097/INF.0000000000001822
103. Dauby N, Chamekh M, Melin P, Slogrove AL, Goetghebuer T. Increased risk of group B streptococcus invasive infection in HIV-exposed but uninfected infants: a review of the evidence and possible mechanisms. Front Immunol. 2016;7. doi:10.3389/fimmu.2016.00505
104. Wilder-Smith A, Longini I, Zuber PL, et al. The public health value of vaccines beyond efficacy: methods, measures and outcomes. BMC Med. 2017;15(1). doi:10.1186/s12916-017-0911-8.
105. Mohle Boetani JC, Schuchat A, Plikaytis BD, Smith JD, Broome CV. Comparison of prevention strategies for neonatal Group B streptococcal infection: a population-based economic analysis. JAMA J Am Med Assoc. 1993;270(12):1442–1448. doi:10.1001/jama.1993.03510120064032
106. Kim SY, Russell LB, Park J, et al. Cost-effectiveness of a potential group B streptococcal vaccine program for pregnant women in South Africa. Vaccine. 2014;32(17):1954–1963. doi:10.1016/j.vaccine.2014.01.062
107. Russell LB, Kim SY, Cosgriff B, et al. Cost-effectiveness of maternal GBS immunization in low-income sub-Saharan Africa. Vaccine. 2017;35(49):6905–6914. doi:10.1016/j.vaccine.2017.07.108
108. Ahmed N, Giorgakoudi K, Usuf E, et al. Potential cost-effectiveness of a maternal Group B streptococcal vaccine in The Gambia. Vaccine. 2020;38(15):3096–3104. doi:10.1016/j.vaccine.2020.02.071.
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.