Back to Journals » Clinical, Cosmetic and Investigational Dentistry » Volume 16
A Review of a Breakdown in the Barrier: Tight Junction Dysfunction in Dental Diseases
Authors Do TT ![]() , Nguyen VT
, Nguyen VT ![]() , Nguyen NTN
, Nguyen NTN ![]() , Duong KTT
, Duong KTT ![]() , Nguyen TTM
, Nguyen TTM ![]() , Le DNT
, Le DNT ![]() , Nguyen TH
, Nguyen TH ![]()
Received 19 August 2024
Accepted for publication 15 November 2024
Published 30 December 2024 Volume 2024:16 Pages 513—531
DOI https://doi.org/10.2147/CCIDE.S492107
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Christopher E. Okunseri
Thao Thi Do,1,* Vy Thuy Nguyen,1,* Ngoc Tran Nhu Nguyen,1 Kim Tran Thien Duong,1 Tri Ta Minh Nguyen,1 Duong Nguyen Thuy Le,1 Tin Hoang Nguyen2
1Oral Diagnosis and Periodontology Department, Faculty of Odonto-Stomatology, Can Tho University of Medicine and Pharmacy, Can Tho City, 900000, Vietnam; 2Department of Physiology, Faculty of Medicine, Can Tho University of Medicine and Pharmacy, Can Tho City, 900000, Vietnam
*These authors contributed equally to this work
Correspondence: Tin Hoang Nguyen, Department of Physiology, Faculty of Medicine, Can Tho University of Medicine and Pharmacy, 179 Nguyen Van Cu Street, An Khanh Ward, Ninh Kieu District, Can Tho City, 900000, Vietnam, Tel +84344895522, Email [email protected]
Abstract: The tight junction (TJ), a type of cell-cell junction, regulates the permeability of solutes across epithelial and endothelial cellular sheets and is believed to maintain cell polarity. However, recent studies have provided conflicting views on the roles of TJs in epithelial polarity. Membrane proteins, including occludin, claudin, and the junction adhesion molecule, have been identified as TJ components. TJs are predominantly found at the stratum granulosum and stratum corneum. Although it remains unclear whether the disruption of TJs is the cause or consequence of certain dental diseases, evidence suggests that TJ dysfunction may be a crucial factor in gingival epithelial barrier impairment and the progression of oral diseases. Bacterial infection is among the most specific factors we found that may contribute to the breakdown of the epithelial barrier formed by TJs in dental diseases. Bacteria and their products may weaken the epithelial barrier by directly destroying intercellular junctions or altering the expression of junctional proteins. Additionally, they may induce the production of inflammatory cytokines, which could lead to the downregulation of TJ proteins and, consequently, impair the epithelial barrier. This review introduces a novel perspective by exploring, for the first time, the role of TJs dysfunction in the breakdown of the oral epithelial barrier and its potential link to the progression of dental diseases such as gingivitis, periodontitis, Sjӧgren syndrome, and oral squamous cell carcinoma.
Keywords: epithelial barrier, gingival inflammation, Sjӧgren syndrome, oral squamous cell carcinoma, periodontitis
Introduction
The epithelium functions as a frontline protective barrier, separating underlying tissues from the external environment and guarding against physical, chemical, and microbial threats. This protection is ensured by various types of cell-cell junctions, including tight junctions (TJs).1 TJs seal the paracellular space within the apical regions of epithelial and endothelial cells2 and are found mostly in the granular and cornified layer (and in lower concentration in other layers) of the oral epithelium.3 TJs have been indicated to play a crucial role in various dental conditions, including gingivitis,4–7 periodontitis,8–10 Sjӧgren syndrome (SS),11 oral squamous cell carcinoma (OSCC),12–14 abnormal tooth formation,15 and Immunoglobulin G4-related sialadenitis (IgG4-RS).16,17
Disruption of the TJs in the gingival barrier is closely related to inflammatory agents that facilitate the spread of bacteria and inflammation. When TJs are disrupted, bacterial products and their toxins penetrate tissues more easily.8 The integrity of the epithelial barrier can be compromised when bacteria invade the gingival tissues through the intercellular spaces in the junctional epithelium (JE) or sulci, which are maintained by TJs.18 Alternatively, changes in the expression of junctional proteins can weaken this barrier.19 Gingivitis and periodontitis are two prevalent forms of periodontal disease resulting from the disruption of this delicate balance. SS is a long-term autoimmune disorder marked by inflammation and the dysfunction of moisture-producing glands. Modifications in the production of TJ proteins alter the expression of these glands.11 OSCC is a common form of oral cancer characterized by the malignant transformation of squamous cells in the oral cavity.20 Some TJ proteins are strongly expressed in tumor cells with keratinization21 and are reported to be involved in the pathophysiology of OSCC.22
The role of TJs is well-recognized in the medical community, with their function and importance extensively documented in organs such as the brain,23,24 kidney,25,26 skin,27,28 and gut.29 For example, the absence of zonula occludens-1 (ZO-1) proteins and occludin in the brain can contribute to the breakdown of the blood-brain barrier, potentially leading to neurological disorders such as multiple sclerosis.30 Additionally, this research indicates that the ischemia associated with stroke is linked to the relocation of specific TJ proteins, such as claudin (CLDN)-1, ZO-1, and ZO-2, from the cell’s outer membrane to the cytoplasm.30 Within the kidney, the significance of CLDN in kidney-related ailments is emphasized. Malfunctions in CLDN function can cause various kidney conditions, including hypomagnesemia, hypercalciuria, the formation of kidney stones, and hypertension.26 However, research on TJs in gingival pathology is limited.
A comprehensive understanding of TJs and their relevance in dental diseases is crucial for the early detection, prevention, and treatment of these conditions.31–33 This review aims to clarify the molecular structure and function of TJs under normal physiological conditions and analyze their relevance in periodontitis, OSCC,34,35 and SS.36
Tight Junctions: Structural and Functional Overview
Epithelial or endothelial cells form four types of intercellular junction complexes, including TJs, adherens junctions (AJs), hemidesmosomes, and gap junctions (GJs)37 (Figure 1A). AJs and desmosomes create robust mechanical bonds between neighboring cells, ensuring their cohesion, while GJs facilitate distinctive cellular communication pathways.38,39 TJs function as protein-sealing structures that encircle neighboring cells, creating a belt-like arrangement on the apical side of the lateral membrane. By occluding the intercellular space, they effectively separate the lumen from the interstitial space, functioning as a barrier and a fence.40,41
 |
Figure 1 Cell-cell junctions and structure of tight junctions in epithelium. (A) Some types of cell-cell junctions. (B) Structure and proteins of tight junctions between epithelial cells. Claudin forms the tight junction strand that interacts with ZO. No sequence similarity exists between claudin and occludin. ZO proteins are tight junction scaffold proteins that link actin filaments to tight junctions. This figure is the authors’ graphical work drawn using Inkscape 1.3.2. Abbreviations: PM, Plasma membrane; AJ, Adherens junction; GJ, Gap junction; HDS, Hemidesmosome; BL, Basal lamina; ZO, Zonula occludens; JAM, Junctional adhesion molecule. |
Junction Molecular Composition
TJs are located at the most apical region of the lateral membrane.2 The TJ strand, a fibril-like polymer comprising CLDN molecules within the plasma membrane, constitutes the functional unit of TJs.42 A TJ is a reticular structure formed by more than 40 proteins that are either transmembrane (CLDNs, occludin, the junctional adhesion molecule [JAM], etc.) or cytosolic scaffolding proteins (ZO, cingulin, etc.) and actin cytoskeleton38 (Figure 1). Each TJ strand binds transversely to another, establishing connections between adjacent cells and forming a structure that resembles a “zippering up” mechanism from the lateral to apical direction.43 Moreover, cytoplasmic proteins function as scaffold proteins, linking TJ strands with cytoskeletal proteins to sustain TJ functionality.2 Overall, unraveling the specific protein interactions within TJs of living cells, and even in cell-free systems, has proven challenging.44
Transmembrane Proteins
CLDN: The CLDN family was identified in 199845 and has emerged as pivotal in forming the backbone structure of the TJ strand when polymerized on the cell membrane. The family has at least 27 members, each exhibiting a specific distribution across organs and tissues.41 CLDNs possess four transmembrane domains, two extracellular loops (the first extracellular segment [ECS1] and the second extracellular segment [ECS2]), and two cytoplasmic termini (C-terminus and N-terminus). ECS1 and ECS2 have distinct amino acid sequences. ECS1 mainly contributes to the paracellular barrier or selective channel attributes, whereas ECS2 helps maintain the opposing lateral cell membranes in close contact.46 CLDNs interact with the ZO family of scaffolding proteins via their cytoplasmic region,32 which in turn are essential for TJ assembly.47–49 CLDN protein polymerization to create strands in epithelial cells relies on the expression of ZO family proteins.50 The C-terminal postsynaptic density protein (PDZ)-binding motifs facilitate direct interactions with intracellular proteins, which include ZO-1, ZO-2, and ZO-3.43,51
Occludin: Myelin/lymphocyte and related proteins for vesicle trafficking and membrane link (MARVEL) D1, also called occludin, together with MARVEL D2 (tricellulin) and MARVEL D3 protein, belongs to the TJ-associated MARVEL protein family.52–54 Occludin, the first transmembrane protein of the TJ to be found in 1993,52 characterized as a tetra-transmembrane protein,55 comprises four transmembrane domains, two extracellular loops, and two cytoplasmic termini. However, no structural or sequence similarity exists between occludin and CLDNs.56 Occludin is believed to interact, whether directly or indirectly, with the co-expression of CLDN-1 and CLDN-2.57 The role of occludin in barrier formation has remained controversial. A study has shown that the inclusion of this protein in the TJ strands led to an increase in transepithelial electrical resistance and paracellular permeability, suggesting its role in forming selective pores.58
Junctional adhesion molecules: JAMs, belonging to the immunoglobulin superfamily, are identified as transmembrane proteins that undergo glycosylation.43 JAM proteins comprise three primary subtypes: JAM-A, JAM-B, and JAM-C (also referred to as JAM-1, −2, and −3), which exhibit comparable sequences and structures.56 The common structure of JAMs includes a single transmembrane domain with its C-terminus in the cytoplasm and N-terminus outside the cell.41 These transmembrane domain proteins are crucial for maintaining the integrity of epithelial barriers,59,60 playing an essential role in the early stages of TJ formation and regulation. Specifically, JAMs contribute to the regulation of TJ assembly in epithelial cells.41 When cell-cell junctions are disrupted, inhibiting JAMs impairs the recovery of barrier function, leading to the loss of occludin and JAMs, while ZO-1 remains unaffected.61 JAMs are not only present in cells forming TJs but are also involved in various cellular functions, such as the movement of leukocytes and endothelial and epithelial cells, as well as modulating paracellular permeability.62
Cytosolic Scaffolding Proteins
ZO proteins: The ZO family, including ZO-1, ZO-2, and ZO-3, are TJ scaffold proteins.56,63 ZO functions as a scaffolding protein that connects transmembrane and cytoplasmic proteins and potentially links AJs to TJs.64 ZO-1 and ZO-2 regulate TJ assembly44,47 and can also determine where the polymerization of CLDNs occurs, indicating their essential role in TJ formation.49 These proteins have multiple important functions, such as interacting with nuclear and dual residency proteins and participating in regulating cell growth.1 Each ZO protein has three PDZ domains: one Src homology 3 domain, one guanylate kinase domain, and one proline-rich region. ZO proteins can associate with several proteins through PDZ domains, such as JAM-A, CLDNs, occludin, and F-actin.41 Studies have demonstrated that ZO-165 and ZO-266 bind to occludin directly.
Cingulin: Cingulin is situated within the cytoplasm, forming a parallel dimer comprising two subunits, each weighing 140–160 kDa. These subunits comprise a globular head, a coiled-coil rod, and a globular tail. Myosin and ZO-3 can bind to the NH2-terminus and COOH-terminus of cingulin, whereas ZO-1 and ZO-2 specifically bind to the NH2-terminus.67 Essentially, cingulin functions as a scaffold protein, facilitating the connection and stabilization of the TJ structure by interacting with ZO family proteins and cytoskeletal elements. Additionally, cingulin is involved in signal transduction pathways, including the regulation of Ras homolog family member A signaling to govern epithelial cell proliferation.68,69
Actin Cytoskeleton
The cytoskeleton comprises microtubules, microfilaments, and intermediate filaments. It was proposed that the actin cytoskeleton is linked to TJs by ZO proteins64 (Figure 1B). ZO-170 and ZO-271 with proline-rich C-terminals were shown to bind F-actin, while ZO-2 does not bind actin.72 The cytoskeleton maintains cell morphology and bears external forces; it also helps maintain the cell’s internal structure, which restricts the mobility and distribution of polarized membrane molecules. This role is fulfilled by not only the actomyosin cytoskeleton but also the spectrin/fodrin cytoskeleton.64 Microfilaments are cytoskeletal components that enable the opening and closing of TJs and mainly comprise actin.73 It has been hypothesized that any occurrence leading to a decrease or elimination of the interaction between the TJ plaque and the actomyosin cytoskeleton would enhance the epithelium’s barrier function. The binding to F-actin occurs through the actin-binding region. ZO-1 binds to actin via a 28-residue actin-binding site (ABS) at the TJ. The ABS of ZO-1 is shown to have a weaker affinity toward actin than the α-catenin of the AJ.74 This study also indicates that increased affinity to the ABS of ZO-1 leads to a decreased epithelial barrier function, and a weaker link between ZO-1 and actin causes junction elongation.
Roles of Tight Junctions
Several studies have acknowledged that TJs have two main functions: firstly, they separate the apical from the basolateral cell surface domains to maintain cell polarity (the fence function); secondly, they regulate ions, water, and various macromolecules through the paracellular space (the barrier function).75 However, recent studies have provided conflicting views on the roles of TJs in epithelial polarity.
Barrier Functions
A major function of TJs is to determine the selective permeability in leaky epithelia and form a permeability barrier in tight and leaky epithelia.2,44,76 The paracellular barrier regulates selective permeability between cells, enabling passive transport of molecules across the tissue and between distinct compartments of the body.2 The function of tight epithelia is the creation of a diffusion barrier by tight control of the transcellular and paracellular pathways.77 The junctional gate is a semipermeable diffusion barrier that discriminates solutes based on size and charge. Solutes cross the junctional paracellular pathway along two routes.2 It is hypothesized that the flux in the leak pathway results from the breaking and annealing of CLDN strands, potentially influenced by intermolecular associations among CLDNs, occludin, ZO-1, and the actin cytoskeleton.8,29 The charge-selective permeation pathway is believed to comprise pores across serially arranged barriers, enabling the diffusion of ions and small uncharged molecules. The estimated diameter of these pores is ~4–8 Å and depends on the tissue and molecule analyzed. The size-selective pathway enables the diffusion of larger solutes and macromolecules up to a size limit of ~30–60 Å.2 In leaky epithelia, this barrier functions as a low-capacity leak pathway, facilitating the permeation of larger macromolecules but lacking ion selectivity.
The Fence Function and Epithelial Polarity: Traditional Views and New Evidence
The interactions among TJ proteins not only establish paracellular barriers; due to the TJ’s apical circumferential structure, it has also long been accepted that they segregate membrane proteins and lipids between apical and basolateral regions and are essential in epithelial polarity through maintaining the asymmetric distribution of membrane proteins and lipid functions.78,79 However, recent studies have challenged this function of TJs. Numerous models have been proposed to explain how TJs form a membrane fence, including lipid micelle, size-dependent exclusion, molecular crowding, and lipid microdomain models.45 However, these models are still subject to refutation, and a lack of concrete and unified evidence remains. It was reported that in an epithelial cell line lacking ZO proteins, lipids from the apical and basolateral membranes remained segregated. This finding led to the conclusion that TJs are not essential for maintaining lipid polarity in epithelial cells.80 Conversely, in ZO-1 and ZO-2 double knockout cells, epithelial polarity was found to be disorganized.47 Recent studies have presented conflicting perspectives on the roles of TJs in epithelial polarity; the absence of TJs did not affect apicobasal polarization,47 so this function is unclear.
Tight Junctions in Oral Mucosa
The oral mucosa comprises three layers: a surface squamous stratified epithelium, known as the oral epithelium, under which is connective tissue termed the lamina propria and submucosa. The oral mucosa is categorized into three types, the lining, masticatory, and specialized mucosae, and has regional differences in keratinization, tissue thickness, and cell turnover time. The keratinized or parakeratinized stratified squamous epithelium covers the surface of the gingiva and hard palate mucosa, known as masticatory mucosa.81,82 Like the epidermis, the main cell type in the oral mucosa is the keratinocyte, and these cells are arranged as four classified sublayers, starting from the deepest: the stratum basale, followed by the stratum spinosum, stratum granulosum, and stratum corneum.33 The oral mucosa acts as a barrier to protect the deeper tissues from mechanical damage and prevent the entry of pathogens and exogenous harmful substances, maintaining tissue integrity and homeostasis. It comprises physical (cell-cell and cell-extracellular matrix junctions, especially TJs), microbiological, and immune barriers.83
Although limited research has been conducted on stratified squamous epithelial tissues, particularly the oral mucosa, it is believed that the unique integrity of this first line of defense is maintained by the interconnection of highly specialized junctional complexes, including TJs.33
Epithelial Barrier of the Gingiva
The establishment of the oral mucosal attachment around teeth is specific to the oral cavity. The gingival tissues comprise free, attached gingiva and interdental papillae (Figure 2A). Free gingiva forms a short cuff around the teeth without attachment, while attached gingiva is connected to the tooth or bone through junctional epithelial attachment and collagen fibers. Interdental papillae are located between teeth, comprising keratinized epithelium over connective tissue84 (Figure 2A). The free gingiva comprises three elements: (1) the oral gingival epithelium (OGE), oriented toward the oral cavity; (2) sulcular epithelium (SE), facing the tooth but without direct contact; and (3) JE, in contact with the tooth enamel.85–87 The OGE is a keratinizing type, serving as an effective barrier against microbial invasion into the gingival connective tissue. Conversely, the SE and JE are primarily non-keratinized,88 making them semipermeable and enabling the passage of large substances from the gingival sulcus into the connective tissue beneath. The JE cells maintain the junction between the enamel and epithelium by attaching to the basal lamina on the enamel through hemidesmosomes, a process vital for tooth emergence into the oral cavity.18
 |
Figure 2 Structure of periodontium and demonstration of tight junction location in oral epithelium. (A) Anatomy and histology of the gingival unit. (B) Localization of tight junctions in gingival stratified squamous epithelium. This figure is the authors’ graphical work drawn using Inkscape 1.3.2. Abbreviations: OGE, Oral gingival epithelium; SE, Sulcular epithelium; JE, Junctional epithelium; GS, Gingival sulcus; CT, Connective tissue; PL, Periodontal ligament; TJs, Tight junctions, cell-cell junctions (TJ, AJ, GJ, Desmosome). |
Gingival tissues are specialized to function as a defense mechanism for the body’s peripheral regions.88 The gingival connective tissue comprises a dense network of collagen fibers that provide firmness and anchor the gingiva to the cementum and alveolar bone (Figure 2A). It also contains abundant lymphatic and blood vessels, nerves, and cells related to immune-inflammatory and vascular functions.89 The gingival epithelial barrier is critical in protecting against pathogens, foreign substances, and mechanical stress.90 Dysbiosis involves an imbalance in bacterial species, which can disrupt host-microbe interactions, leading to inflammation and bone loss.91,92
The primary mechanisms supporting resistance to mechanical injury include (1) the presence of plentiful intercellular junctions, primarily desmosomes, which tightly bind the cells; (2) undulations in the epithelium and connective tissue (known as rete ridges); and (3) the stratum corneum. Additionally, the stratum corneum, coupled with relatively narrow intercellular spaces, contributes to lower permeability compared to the non-keratinized lining mucosa of the JE and SE.90
The initial layer of the gingival barrier is formed by gingival crevicular fluid (GCF), a highly specific oral cavity fluid representing periodontal health. It can be a useful predictor or diagnosis marker of periodontal diseases such as gingivitis and periodontitis (chronic and aggressive):93–97 (I) proteolytic and hydrolytic enzyme biomarkers: alkaline phosphatase,98 aspartate aminotransferase,99 cathepsin-B;100 (II) bone-related biomarker: osteocalcin,101 osteopontin,101 osteonectin;102 (III) cell death and tissue breakdown products: chondroitin-4-sulfate,103 and fibrinogen.102,104
The amount of GCF in healthy sulcus is typically low.93 The ongoing supply of GCF forms a layer-like protective barrier, defending TJs against lipopolysaccharides (LPSs) and various pathogen-associated molecular patterns by cleansing the epithelial surface.8,105 GCF is essential for initiating an immune response and preventing microbes from invading tissues. In periodontal disease, the inflammatory response leads to changes in the constituents of GCF and selective upregulation of certain cytokines.93 A wide range of cytokines disrupts the barrier function of epithelial and endothelial cells by affecting the structure and functionality of TJs.106 Qiurong Li et al indicated that tumor necrosis factor-α (TNF-α) and interferon-γ (IFN-γ) can disrupt TJs at the epithelial barrier by changing the lipid composition and fatty acyl substitutions of phospholipids in membrane microdomains of TJs.107
The neutrophil extracellular traps (NETs) constitute the second stratum of the gingival barrier.108–111 NETs, structures evolved within the innate immune system, are web-like meshworks comprising chromatin. Activated neutrophils generate these structures primarily in response to pathogen challenges. NETs eliminate, damage, and entrap bacteria, preventing their spread.108 The final two properties of NETs imitate the primary characteristics of the mucus layer. The third and final layer of the gingival barrier is the epithelial lining. The primary role of the JE is to establish a barrier to the external environment, encompassing physical, biochemical, and pathogenic aspects.8
Tight Junctions and Oral Health
Similar to their positioning in the epidermis, where functional TJs are located in the second layer of the stratum granulosum,112,113 TJs in gingival tissue are located exclusively in the granular and cornified layers (Figure 2B). Unlike cultured gingival keratinocytes in vitro, which exhibit an extensive network of TJ strands, TJs in gingival tissue do not form complex strand structures.3 Indirect immunofluorescence analysis of healthy gingiva revealed that CLDN-1 and ZO-1 were present in the stratum granulosum but absent from the stratum corneum and stratum basale, while occludin was mainly expressed in the stratum granulosum.7 Extremely low JAM-A levels were detected in the granular/keratinized layers of the SE.114 In multilayered gingival epithelial cultures, gene expression analysis revealed that CLDN-1, −4, −12, −17, −25, JAM-A, and occludin are the most highly expressed.115
The epithelium functions as a frontline protective barrier, separating underlying tissues from the external environment and guarding against physical, chemical, and microbial threats. Maintaining the integrity of this barrier is essential for oral as well as overall health. With their semipermeable structure, TJs are crucial in regulating the paracellular transport of ions, water, and solutes.116 Occludin has been identified in the surface layer of the gingival epithelium, while CLDN-1 is present in the uppermost layer.117,118 CLDNs exhibit barrier properties that directly control the gate function of paracellular TJ channels. It is evident that the optimal expression of TJ proteins is crucial for upholding normal physiological function, and any imbalance could result in pathological consequences.33
Several TJ proteins have been reported to be expressed in salivary glands, such as CLDNs-1, −2, −3, −4, and −5; occludin; JAM-A; and ZO-1.33,119 These proteins have been detected particularly in acinar and ductal cells and in endothelial cells surrounding the salivary epithelium.119,120 The structure and function of TJs must vary between serous, mucous, and mixed acini, as well as between intercalated, striated, and excretory ducts. While the functional importance of TJs in salivary gland epithelium remains inadequately understood, it is suggested that the presence of TJs within the salivary epithelium is both necessary and an indicator of dysfunction.120
The generation of saliva is crucial for fundamental oral cavity functions, including swallowing and digestion, while also demonstrating antimicrobial properties. Essential elements of saliva, such as ions, electrolytes (sodium and chloride), proteins, and lipids, are produced in the acinar cells of major salivary glands and undergo alterations within the salivary ducts.121 The movement of K+ and Cl− along their gradients creates a potential difference across the epithelium, causing Na+ and water to diffuse through the TJs.122 The molecular composition of TJs influences the permeability of paracellular pathways during water secretion.123,124 Xin Cong et al indicated that ZO-1, CLDN-3, and −11 are important in the secretion of the submandibular gland under physiological and pathophysiological conditions.124 A recent study found that the concentration of electrolytes affects barrier function and protein localization in bovine mammary epithelium.125 This suggests that TJ protein interactions may change in response to ion secretion in salivary glands. Additionally, proinflammatory cytokines, TNF-α and IFN-γ, reduce CLDN-1 expression, indicating that CLDN-1 may contribute to TJ integrity in salivary epithelium.120,126 Changes in the expression and organization of TJs have been observed in salivary glands affected by SS, highlighting their crucial role in understanding salivary gland dysfunction.11
The Breakage of Epithelial Barrier Tight Junctions in Gingiva
It is imperative to uphold the integrity of the epithelial barrier for oral and overall health.18 Bacteria can compromise this barrier by invading gingival tissues, which are bound by TJs, through the intercellular spaces in the junctional and sulcular/pocket epithelium.
One mechanism of paracellular bacterial invasion involves the direct destruction of intercellular junctions. Human gingival epithelial cells express JAM-A,115 a TJ protein essential for regulating selective permeability based on the molecular weight of target molecules. Porphyromonas gingivalis proteases, called gingipains, mechanically degrade the mature form of JAM-A on the plasma membrane, increasing the penetration of 40 kDa dextran, LPS, peptidoglycan (PGN), and gingipains into subepithelial tissues, which regulates the gingival epithelial barrier function.127 Additionally, gingipains are implicated in increasing the permeability of the human cerebral endothelial cell monolayer through the degradation of TJ proteins, including ZO-1 and occludin.24
Another means to weaken the epithelial barrier is by altering the expression of junctional proteins. Research has shown that cell infection with P. gingivalis promoted the expression of the TJ genes encoding occludin, CLDN-1, and CLDN-4, which enhances barrier function and decreases cell-cell permeability.128 In a rat model of periodontal disease, chronic LPS exposure reduced CLDN-1 expression in the JE and impaired the epithelial barrier function. These findings suggest that decreased CLDN-1 may contribute to the onset of periodontal disease.126 Grainyhead-like 2 (GRHL2), an epithelial-specific transcription factor, regulates the expression of junction proteins. In a study using a GRHL2 conditional knockout (KO) mouse model, W. Chen et al suggested that P. gingivalis bacteria may enhance paracellular penetration through the oral mucosa, partially by targeting the expression of GRHL2 in the oral epithelial cells.129 This impairs the epithelial barrier by inhibiting junction protein expression. Immunohistochemical assessment of the periodontal lesions confirmed decreased protein levels of ZO-1, β-cat, and CLDN-3 in the JE of GRHL2KO mice. Consequently, alveolar tissue destruction and systemic bacteremia increased. These results suggested that TJ destruction may be a crucial step in gingival barrier function failure and oral diseases.
T-helper 17 (Th17) cells, enriched in the epithelial barrier, contribute to maintaining the integrity of the oral epithelial barrier by regulating TJ proteins such as CLDN and occludin.11 These interleukin (IL)-17-producing CD4+ T cells have been implicated in the pathogenesis of multiple autoimmune and inflammatory disorders.95 Th17 cells produce IL-17, IL-17F, and IL-22, inducing a massive tissue reaction due to the broad distribution of the IL-17 and IL-22 receptors.105 The application of IFNs and Th17 cytokines has shown significant disruption of the epithelial barrier, evidenced by discontinuous ZO-1 immunolocalization.90 L. Abusleme and N. M. Moutsopoulo provided evidence for IL-17A-dependent regulation of occludin during epithelial injury that limits excessive permeability and maintains barrier integrity. This study also indicated that the absence of IL-17A results in increased epithelial injury and compromised barrier function after dextran sulfate sodium (DSS) treatment, with IL-17A signaling through Act-1 supporting barrier function by maintaining occludin localization at TJs during DSS injury.16 Furthermore, physical-mechanical mastication can damage and promote Th17 cell development in gingival tissue.43 Aggor FEY et al showed that oral candidiasis involved the cooperative activities of IL-22 and IL-17.96 The interaction between Th17, IL-17, and TJs is expressed in not only gingival tissue but also the salivary gland. Zhang et al demonstrated that IL-17 derived from infiltrating lymphocytes impairs the integrity of the TJ barrier through the Nuclear factor kappa B signaling pathway, potentially contributing to salivary gland dysfunction in SS.94
The Critical Role of Tight Junctions in Some Dental Diseases
The Interplay Between Gingivitis, Periodontitis, and Tight Junctions
Pathophysiology of Gingivitis and Periodontitis
Gingivitis is an inflammatory condition limited to the surface periodontal tissue, including the outer epithelial and adjacent connective tissue layers.130,131 Gingivitis begins with bacterial components such as LPS, PGNs, lipoteichoic acids, proteases, and toxins present in the biofilm on tooth surfaces, triggering inflammatory reactions. These inflammatory factors, part of the host’s response, involve the activation and recruitment of inflammatory cells through toll-like receptors (TLRs) on host cells that recognize bacterial antigens and products such as LPS and PGNs found in the biofilm on tooth surfaces132,133 (Figure 3). This initiates an inflammatory response, attracting lymphocytes and macrophages to the tissue (Figure 3B), leading to the breakdown of 60–70% of the collagen in the gingival connective tissue.133,134 However, the bone remains unaffected, enabling the repair and remodeling of gingival tissues without permanent damage. When bacterial antigens are encountered by antigen-presenting cells such as dendritic cells, macrophages, and B cells, naïve CD4+ T helper cells (Th0) differentiate into various subsets, including Th1, Th2, Th17, and regulatory T cells, based on the cytokines they produce. Cytokines and chemokines such as IL-1, IL-6, IL-12, IL-17, IL-18, IL-21, TNF-α, and IFN-γ in the GCF are significant in periodontal inflammation development.133 Many inflammatory factors, such as a surplus of LPS, oncostatin M, neutrophil proteases, bacterial proteases, and toxins, deteriorate gingival TJs. The observation that transient bacteremia commonly occurs in inflamed gingiva during activities such as chewing, tooth brushing, and dental procedures implies that inflammation-weakened TJs rupture when exposed to physiological mechanical strains. This phenomenon, known as an epithelial barrier breach, enables viable bacteria to enter the blood.8 The inflammatory response is a natural defense mechanism designed to avoid bacterial infections reaching deeper tissues such as bones. However, if this inflammation becomes uncontrolled and chronic, it can cause permanent damage to the periodontal tissues (Figure 3C–D). This can result in periodontitis and its associated symptoms, including periodontal pockets, loss of attachment, receding gingiva, loose teeth, tooth movement, and ultimately tooth loss.135,136 Figures 4 and 5 illustrate the progression of gingivitis and its severe consequences on dental health. Figure 4 shows gingivitis with visible signs of inflammation, including red, swollen, and irritated gingival tissues, particularly along the gingival margin. Across Panels A, B, and C, the gingiva appears tender and prone to bleeding, which are hallmark features of gingivitis, typically resulting from plaque accumulation and bacterial infection. In a related condition, Figure 5 shows a different patient with periodontitis, where Panel A illustrates advanced enamel erosion and inflamed gingiva in the upper arch, Panel B displays similar conditions in the lower arch with carious lesions, and Panels C, D, and E provide multiple angles of the deteriorated dental structures with pronounced gingival inflammation, gingival recession, and plaque accumulation, indicative of extensive periodontal disease.
 |
Figure 3 An overview of gingivitis and periodontitis. The process begins with the accumulation of dental plaque (A), a complex biofilm formed by the colonization of bacteria enclosed by a protective matrix. (B) If this biofilm is not disrupted and continues to accumulate, plaque bacteria produce toxins that penetrate the gingival tissues. The host responds to this bacterial infection with the classical innate immune response, where pathogens trigger leukocytes to release proinflammatory mediators, such as cytokines and chemokines. This cascade of reactions activates adaptive immunity, involving specific T and B lymphocytes. Neutrophils and macrophages, activated by these lymphocytes, release proinflammatory cytokines that increase vascular permeability. These immune cells migrate to the gingival tissue through the bloodstream, causing the gingival tissues to become red, swollen, tender, and more prone to bleeding. (C and D) If untreated, gingivitis can lead to early periodontitis, where the gingiva pulls away from teeth, forming pockets and causing attachment and bone loss. This figure is the authors’ graphical work drawn using Inkscape 1.3.2. Abbreviations: PT, Periodontal tissue; TJs, Tight junctions; RBC, Red blood cell. |
 |
Figure 4 Clinical case: Gingivitis. (A) Frontal view. (B) Right buccal view. (C) Left buccal view. This figure is from Faculty of Odonto-Stomatology, Can Tho University of Medicine and Pharmacy in Vietnam. The patient provided consent to publish the images. |
 |
Figure 5 Clinical case: Periodontitis. (A) Occlusal view of the upper arch. (B) Occlusal view of the lower arch. (C) Right buccal view. (D) Frontal view. (E) Left buccal view. This figure is from Faculty of Odonto-Stomatology, Can Tho University of Medicine and Pharmacy in Vietnam. The patient provided consent to publish the images. |
Impact of Inflammation on Tight Junctions
TJs create a physical barrier to inhibit the entry of bacteria and their products. Disruption of the TJs in the gingival barrier is closely related to inflammatory agents that facilitate the spread of bacteria and inflammation. When TJs are disrupted, bacterial products and their toxins penetrate tissues more easily.8 As inflammation increases, edema and structural changes appear in the epithelium, manifesting as widened intercellular spaces. If this widening continues, only a few isolated TJs remain, with no intermediate junctions.137 During the inflammatory response, cell junctions are affected by and affect the inflammatory cytokines.138 Downregulation of TJs by proinflammatory cytokines (TNF, LIGHT, IL-1) can occur within hours without altering TJ protein expression.139 Occludin and ZO-1 are downregulated, whereas CLDN-1 and CLDN-4 are overexpressed15 (Table 1). TNF-α is a key mediator of inflammation89 and is markedly increased in individuals with inflammatory conditions.96 IL-1β has been shown to enhance the integrity of the gingival keratinocyte barrier, demonstrated by an increase in transepithelial electrical resistance, upregulation of TJ proteins, and an increase in the formation of TJ strands. This suggests that reinforcing the physical barrier is crucial in the initial defense mechanisms of gingival epithelial cells.7 Furthermore, the inflammatory cytokine IL-6 enhances endothelial permeability by causing changes in the redistribution of the tight junctional protein ZO-1.5 Güney et al found a positive correlation between ZO-1 and TNF-α, concluding that the presence of free ZO-1 in the environment increases with periodontal inflammation.10 TNF-α is known to be a key mediator of inflammation6 and is markedly increased in individuals with inflammatory conditions.96 In periodontitis, an intriguing nexus within this intricate landscape of bone resorption is illuminated by the role of the CLDN-18 protein.9
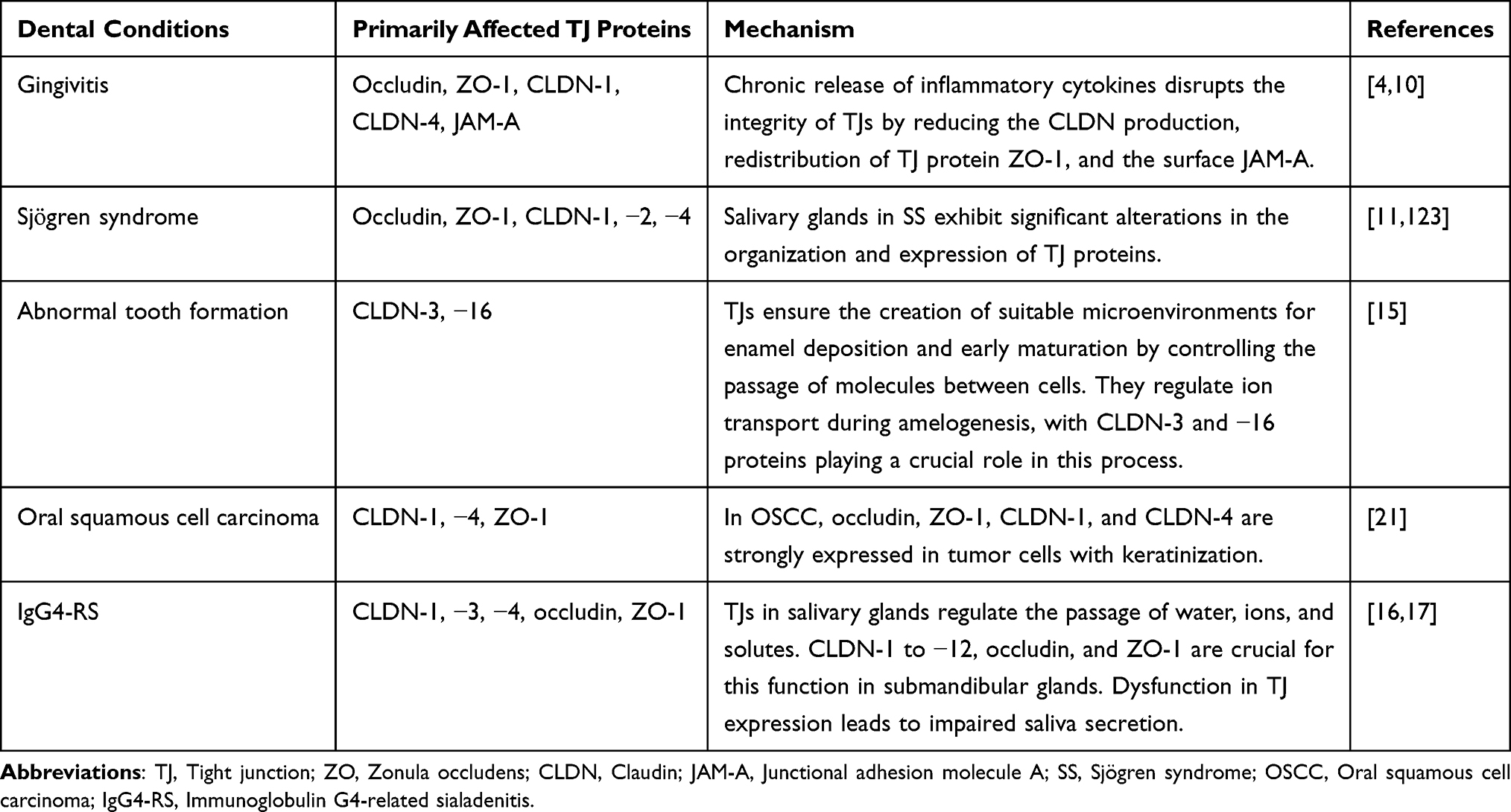 |
Table 1 Correlation Between Dental Conditions and Tight Junction Proteins |
Studies have shown an association between smoking and gingivitis, mediated through TJs.140 Smoking promotes the penetration of LPS and PGN into the gingival epithelium by reducing the surface expression of JAM-A. Conversely, overexpression of JAM-A, such as that induced by vitamin C, has been shown to inhibit the penetration of LPS and PGN caused by cigarette smoke extract.10,141 Additionally, smoking can lead to abnormal tyrosine phosphorylation of TJ proteins, altering their distribution in the cell membrane and disrupting junction formation.10
The Interplay Between Oral Cancer and Tight Junctions
Oral cancer includes malignancies arising in the oral cavity and oropharynx. The etiology of oral cancer is complex, involving a mixture of genetic factors, environmental influences, and changes at the molecular level in the tissues. Recent years have seen growing recognition of the importance of the tumor microenvironment, including the role of the epithelial barrier function in the context of oral carcinogenesis. Emerging evidence suggests that TJs play a nuanced role in the pathophysiology of various cancers and tumorigenesis, including OSCC.12 TJs, positioned as the most apical structure connecting epithelial and endothelial cells, are widely recognized for regulating cell and tissue homeostasis. Any disruption in this cohesion can result in cancer cell invasion and subsequent metastasis.22 During the metastatic process, TJs are the first structure impeding the path to successful metastasis of the cancer cell. For the tumor cell to proceed effectively, TJ structures must be disturbed and dismantled to enable cancer cell penetration.22
CLDN-1 has been reported to play a role in various parts of the oral cavity (including the tongue, gingiva, mouth, and palate) in OSCC142 and is frequently altered in several cancers13 (Table 1), with overexpression associated with stage progression in OSCC.12 CLDN-1 shows a high probability (96.2%) of being overexpressed at messenger ribonucleic acid and protein levels in OSCCs with perineural and angiolymphatic invasion. Thus, it is suggested that CLDN-1 overexpression is associated with early steps of invasion into the underlying tissue, metastasis, and aggressive tumor behavior in OSCC.12,13 However, a study examining healthy oral control tissue from the same individuals reported that, conversely, CLDN-1 expression in OSCC did not differ from that of the intraindividual control tissue and showed no significant association with clinicopathological features.143 While CLDN-1 is expressed throughout the epithelium, occludin, ZO-1, and CLDN-4 predominate at the cell-cell boundary in the granular layer. In OSCC, occludin, ZO-1, CLDN-1, and CLDN-4 are strongly expressed in tumor cells with keratinization, such as cancer pearls14 (Table 1).
The loss of CLDN-7 expression is a frequent event in OSCC,144 with reduced CLDN-7 expression potentially associated with OSCC development and an unfavorable prognosis. A lack of CLDN-7 expression in the tumor center may be used to identify patients at increased risk for regional recurrence.145 Conversely, elevated levels of CLDN-7 correlate with longer survival times, suggesting a positive prognosis.146,147 CLDN-7 has been suggested to function as a tumor suppressor in oral cancer, highlighting its potential as a therapeutic target in oral cancer treatment.31 Findings also suggest that CLDN-4 could be deemed a key immunohistochemical marker for excluding the diagnosis of mesothelioma.13
Matrix metallopeptidase (MMP)-2 and −9 have been demonstrated to regulate the invasion ability of OSCC cells.148 It is shown that MMP-9 cleaves cell-cell junctions, and apical MMP-9 treatment significantly increases transepithelial conductance and alters the immunolocalization of TJ proteins, CLDN-1, and occludin.149 Collectively, these findings suggest that MMP-2 and −9 may influence the integrity of TJs, leading to OSCC cell invasion. Kim et al found that hypoxia enhanced the invasion ability of OSCC cells by increasing MMP-2 and MMP-9 activities, disrupted TJs, and promoted the metastasis of OSCC via the loss of partitioning-defective protein 3.150
Disruption of these junctional proteins of TJs in OSCC increases the permeability of the epithelial barrier. This may facilitate the invasion of cancer cells and surrounding tissue and promote metastasis.151,152 OSCC is characterized by the occurrence of an epithelial-mesenchymal transition (EMT), during which cancer cells become more invasive and migratory. During an EMT, TJs may be disrupted. Different signaling pathways involved in OSCC development (such as Wingless-related integration site, Notch, and Mitogen-activated protein kinase pathways) have all been reported to impact TJs.153 Activation of these pathways can influence the expression and localization of TJs.
The Interplay Between Sjӧgren Syndrome and Tight Junctions
The salivary glands include several secretory units that connect to the oral cavity via a duct system.154 Each unit comprises a cluster of cells arranged in secretory acini. Three pairs of major salivary glands (parotid, submandibular, and sublingual) and many minor salivary glands are scattered throughout the oral cavity within the lamina propria of the oral mucosa.155
SS is a chronic autoimmune disorder characterized by the inflammation and dysfunction of moisture-producing glands, such as the salivary and lacrimal glands. This condition primarily affects the exocrine glands, resulting in a progressive decline in the secretion of saliva and tears and hence dryness in the mouth and eyes.156 Salivary hyposecretion leads to difficulties in chewing and swallowing dry food, along with an elevated susceptibility to dental caries and oral candidiasis.157 Dry eye syndrome triggers ocular symptoms such as irritation, blurry vision, and vision fluctuations, which can progress to severe corneal ulceration, posing a serious risk to vision. Furthermore, patients experience fatigue, depression, and inflammation in non-glandular tissues such as the brain, lungs, and gastrointestinal tract.157
Focal lymphocytic sialadenitis is a diagnostic hallmark of SS and appears as an accumulation of mononuclear cells, including lymphocytes, macrophages, follicular dendritic cells, and dendritic cells.158,159 Initially, T cells primarily populate the infiltrate around ducts in the early stages, whereas in the later stages, B cells become predominant, and the infiltrate expands to also involve acini.160 The pathogenesis of SS underscores the central role played by activated B cells. This activation prompts the generation of autoantibodies and an increase in IgG levels, a condition known as hypergammaglobulinemia, which is evident in certain SS patients.161 In patients with SS, B cells activated by TLRs exhibit altered responsiveness and heightened cytokine production compared to healthy control subjects.161
The destruction of acinar cells, extensive infiltration by lymphocytes, and localized release of inflammatory cytokines, including IL-6, IL-1β, IL-10, IFN-γ, and TNF-α, are characteristic features of SS.162 Studies indicate that IFN-γ and TNF-α compromise the barrier function of TJs, reducing CLDN-1 production. In SS, salivary glands exhibit significant alterations in the organization and expression of TJ proteins. Notably, occludin and ZO-1 undergo downregulation, while CLDN-1 and CLDN-4 are overexpressed (Table 1). Moreover, these CLDNs transition from the apical to the basolateral side of acinar cells in the minor salivary glands.11 The integrity of TJs in SS is intricately linked to the secretion of inflammatory mediators.120
Conclusion
TJs, a type of intercellular junction, are vital for ensuring the structural and functional integrity of the gingival barrier, regulating selective permeability, homeostasis, and the immune response. Besides their expression in the epithelium, TJs are observed in ductal structures such as the salivary glands. In SS, inflammation in the salivary glands disrupts TJs by affecting protein production. Additionally, TJs are involved in the disease progression of OSCC. Changes in the expression of CLDN-1, −4, and −7 could reflect the disruption of the epithelial barrier function and may provide insights into the mechanisms underlying OSCC pathogenesis, offering potential targets for diagnosis and therapeutic intervention. In gingivitis and periodontitis, inflammation can compromise TJ integrity due to the action of inflammatory cytokines. This disruption leads to increased permeability of the epithelial barriers, enabling pathogens and immune cells to infiltrate tissues more easily, exacerbating inflammation. Future research could focus on developing non-invasive diagnostic tests to detect biomarkers of TJ dysfunction in saliva or GCF, which would significantly enhance the early detection and management of dental diseases.
Abbreviations
ABS, Actin-binding site; AJ, Adherens junction; CLDN, Claudin; DSS, Dextran sulfate sodium; ECS, Extracellular segment; EMT, Epithelial-mesenchymal transition; GCF, Gingival crevicular fluid; GJ, Gap junction; GRHL, Grainyhead-like; IFN, Interferon; IgG4-RS, Immunoglobulin G4-related sialadenitis; IL, Interleukin; JAM, Junctional adhesion molecule; JE, Junctional epithelium; KO, Knockout; LPS, Lipopolysaccharide; MARVEL, Myelin/lymphocyte and related proteins for vesicle trafficking and membrane link; MMP, Matrix metallopeptidase; NETs, Neutrophil extracellular traps; OGE, Oral gingival epithelium; OSCC, Oral squamous cell carcinoma; PGN, Peptidoglycan; PDZ, Postsynaptic density protein; SE, Sulcular epithelium; SS, Sjӧgren syndrome; Th, T-helper; TJ, Tight junction; TLR, Toll-like receptor; TNF, Tumor necrosis factor; ZO, Zonula occludens.
Acknowledgments
We express our gratitude to Can Tho University of Medicine and Pharmacy for the time and effort they devoted to the study. We also extend our heartfelt thanks to our mentors, Professor Yoshihiro Kubo and Professor Miko Furuse from the National Institute for Physiological Sciences in Myodaiji, Okazaki 444-8585, Japan, for their valuable guidance throughout our research.
Author Contributions
All authors made a significant contribution to the work reported, whether in its conception, study design, execution, data acquisition, analysis, interpretation, or all these areas; participated in drafting, revising, or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
There is no funding to report.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Samiei M, Ahmadian E, Eftekhari A, Eghbal MA, Rezaie F, Vinken M. Cell junctions and oral health. EXCLI J. 2019;18:317–330. doi:10.17179/excli2019-1370
2. Zihni C, Mills C, Matter K, Balda M. Tight junctions: from simple barriers to multifunctional molecular gates. Nat Rev Mol Cell Biol. 2016;17(9):564–580. doi:10.1038/nrm.2016.80
3. Groeger S, Doman E, Chakraborty T, Meyle J. Effects of Porphyromonas gingivalis infection on human gingival epithelial barrier function in vitro. Eur J Oral Sci. 2010;118(6):582–589. doi:10.1111/j.1600-0722.2010.00782.x
4. Shashikanth N, France MM, Xiao R, et al. Tight junction channel regulation by interclaudin interference. Nat Commun. 2022;13(1):3780. doi:10.1038/s41467-022-31587-8
5. Desai TR, Leeper NJ, Hynes KL, Gewertz BL. Interleukin-6 causes endothelial barrier dysfunction via the protein kinase C pathway. J Surg Res. 2002;104(2):118–123. doi:10.1006/jsre.2002.6415
6. Bradley J. TNF‐mediated inflammatory disease. J Pathol. 2008;214(2):149–160. doi:10.1002/path.2287
7. Stolte KN, Pelz C, Yapto CV, Raguse J-D, Dommisch H, Danker K. IL-1β strengthens the physical barrier in gingival epithelial cells. Tissue Barriers. 2020;8(3):1804249. doi:10.1080/21688370.2020.1804249
8. Vitkov L, Singh J, Schauer C, et al. Breaking the gingival barrier in periodontitis. Int J Mol Sci. 2023;24(5):4544. doi:10.3390/ijms24054544
9. Alshbool FZ, Mohan S. Emerging multifunctional roles of claudin tight junction proteins in bone. Endocrinology. 2014;155(7):2363–2376. doi:10.1210/en.2014-1173
10. Güney Z, Kurgan Ş, Önder C, et al. Expression of tight junction proteins in smokers and non‐smokers with generalized Stage III periodontitis. J Periodontal Res. 2023;58(6):1281–1289. doi:10.1111/jre.13184
11. Ewert P, Aguilera S, Alliende C, et al. Disruption of tight junction structure in salivary glands from Sjögren’s syndrome patients is linked to proinflammatory cytokine exposure. Arthritis Rheum. 2010;62(5):1280–1289. doi:10.1002/art.27362
12. Dos Reis PP, Bharadwaj RR, Machado J, et al. Claudin 1 overexpression increases invasion and is associated with aggressive histological features in oral squamous cell carcinoma. Cancer. 2008;113(11):3169–3180. doi:10.1002/cncr.23934
13. Ouban A, Ahmed A. Claudins in human cancer, A review. Histol Histopathol. 2010;25(1):83–90. doi:10.14670/HH-25.83
14. Morita K, Tsukita S, Miyachi Y. Tight junction-associated proteins (occludin, ZO-1, claudin-1, claudin-4) in squamous cell carcinoma and bowen’s disease. Br J Dermatol. 2004;151(2):328–334. doi:10.1111/j.1365-2133.2004.06029.x
15. Bardet C, Courson F, Wu Y, et al. Claudin-16 deficiency impairs tight junction function in ameloblasts, leading to abnormal enamel formation. J Bone Miner Res. 2016;31(3):498–513. doi:10.1002/jbmr.2726
16. Min S-N, Wu -L-L, Zhang -Y-Y, Zhu W-X, Cong X, Yu G-Y. Disruption of tight junction structure contributes to secretory dysfunction in IgG4-related sialadenitis. J Mol Histol. 2020;51(1):33–46. doi:10.1007/s10735-019-09854-8
17. Zhang XM, Huang Y, Zhang K, et al. Expression patterns of tight junction proteins in porcine major salivary glands: a comparison study with human and murine glands. J Anatomy. 2018;233(2):167–176. doi:10.1111/joa.12833
18. Takahashi N, Sulijaya B, Yamada-Hara M, Tsuzuno T, Tabeta K, Yamazaki K. Gingival epithelial barrier: regulation by beneficial and harmful microbes. Tissue Barriers. 2019;7(3):e1651158. doi:10.1080/21688370.2019.1651158
19. Jiang Q, Yu Y, Ruan H, Luo Y, Guo X. Morphological and functional characteristics of human gingival junctional epithelium. BMC Oral Health. 2014;14(1):30. doi:10.1186/1472-6831-14-30
20. Do TT, Bui LNV, Nguyen L, Le LN, Tran DTP. Clinical and pathological features of oral cancer in a high-risk community in Vietnam. J Maxillofac Oral Surg. 2023; 2023:1–5. doi:10.1007/s12663-023-01997-2.
21. Babkair H, Yamazaki M, Uddin MS, et al. Aberrant expression of the tight junction molecules claudin-1 and zonula occludens-1 mediates cell growth and invasion in oral squamous cell carcinoma. Human Pathol. 2016;57:51–60. doi:10.1016/j.humpath.2016.07.001
22. Martin TA, Jiang WG. Loss of tight junction barrier function and its role in cancer metastasis. Biochimica et Biophysica Acta. 2009;1788(4):872–891. doi:10.1016/j.bbamem.2008.11.005
23. Hsieh JT, Rathore APS, Soundarajan G, St John AL. Japanese encephalitis virus neuropenetrance is driven by mast cell chymase. Nat Commun. 2019;10(1):706. doi:10.1038/s41467-019-08641-z
24. Nonaka S, Kadowaki T, Nakanishi H. Secreted gingipains from Porphyromonas gingivalis increase permeability in human cerebral microvascular endothelial cells through intracellular degradation of tight junction proteins. Neurochem Int. 2022;154:105282. doi:10.1016/j.neuint.2022.105282
25. Pei L, Solis G, Nguyen MTX, et al. Paracellular epithelial sodium transport maximizes energy efficiency in the kidney. J Clin Invest. 2016;126(7):2509–2518. doi:10.1172/JCI83942
26. Hou J. The kidney tight junction (Review). IntJ Mol Med. 2014;34(6):1451–1457. doi:10.3892/ijmm.2014.1955
27. Giri S, Takada A. Tight junction proteins, claudin and occludin in pathological conditions and aging of skin and oral mucosa: a review. 北海道医療大学歯学雑誌. 2019;38(2):17–27.
28. Bäsler K, Brandner JM. Tight junctions in skin inflammation. Pflugers Arch Eur J Physiol. 2017;469(1):3–14. doi:10.1007/s00424-016-1903-9
29. Turner JR, Buschmann MM, Romero-Calvo I, Sailer A, Shen L. The role of molecular remodeling in differential regulation of tight junction permeability. Semin Cell Dev Biol. 2014;36:204–212. doi:10.1016/j.semcdb.2014.09.022
30. Bednarczyk J, Lukasiuk K. Tight junctions in neurological diseases. Acta Neurobiol Exp. 2011;71(4):393–408. doi:10.55782/ane-2011-1861
31. Xu Y-N, Deng M-S, Liu Y-F, Yao J, Xiao Z-Y. Tight junction protein CLDN17 serves as a tumor suppressor to reduce the invasion and migration of oral cancer cells by inhibiting epithelial-mesenchymal transition. Arch Oral Biol. 2022;133:105301. doi:10.1016/j.archoralbio.2021.105301
32. Ben Lagha A, Grenier D. Black tea theaflavins attenuate Porphyromonas gingivalis virulence properties, modulate gingival keratinocyte tight junction integrity and exert anti‐inflammatory activity. J Periodontal Res. 2017;52(3):458–470. doi:10.1111/jre.12411
33. Wan H, Gadmor H, Brown L. Cell-cell interactions in the oral mucosa: tight junctions and gap junctions. In: Bergmeier LA, editor. Oral Mucosa in Health and Disease. Springer International Publishing; 2018:19–30. doi: 10.1007/978-3-319-56065-6_2
34. Feller L, Lemmer J. Oral squamous cell carcinoma: epidemiology, clinical presentation and treatment. J Cancer Ther. 2012;03(04):263–268. doi:10.4236/jct.2012.34037
35. Tan Y, Wang Z, Xu M, et al. Oral squamous cell carcinomas: state of the field and emerging directions. Int J Oral Sci. 2023;15(1):44. doi:10.1038/s41368-023-00249-w
36. André F, Böckle BC. Sjögren’s syndrome. JDDG. 2022;20(7):980–1002. doi:10.1111/ddg.14823
37. Dejana E, Corada M, Lampugnani MG. Endothelial cell‐to‐cell junctions. FASEB J. 1995;9(10):910–918. doi:10.1096/fasebj.9.10.7615160
38. Campbell HK, Maiers JL, DeMali KA. Interplay between tight junctions and adherens junctions. Exp Cell Res. 2017;358(1):39–44. doi:10.1016/j.yexcr.2017.03.061
39. Holthöfer B, Windoffer R, Troyanovsky S, Leube RE. Structure and function of desmosomes. Int Rev Cytol. 2007;65–163. doi:10.1016/S0074-7696(07)64003-0
40. Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P. Molecular Biology of the Cell.
41. Zhang GH, Wu LL, Yu GY. Tight junctions and paracellular fluid and ion transport in salivary glands. Chin J Dent Res. 2013;16(1):13–46.
42. Furuse M, Takai Y. Recent advances in understanding tight junctions. Fac Rev. 2021;10. doi:10.12703/r/10-18
43. Chiba H, Osanai M, Murata M, Kojima T, Sawada N. Transmembrane proteins of tight junctions. Biochimica et Biophysica Acta. 2008;1778(3):588–600. doi:10.1016/j.bbamem.2007.08.017
44. Shen L, Weber CR, Raleigh DR, Yu D, Turner JR. Tight junction pore and leak pathways: a dynamic duo. Annu Rev Physiol. 2011;73:283–309. doi:10.1146/annurev-physiol-012110-142150
45. Otani T, Furuse M. Tight junction structure and function revisited. Trends Cell Biol. 2020;30(10):805–817. doi:10.1016/j.tcb.2020.08.004
46. Tsukita S, Tanaka H, Tamura A. The claudins: from tight junctions to biological systems. Trends Biochem Sci. 2019;44(2):141–152. doi:10.1016/j.tibs.2018.09.008
47. Otani T, Nguyen TP, Tokuda S, et al. Claudins and JAM-A coordinately regulate tight junction formation and epithelial polarity. J Cell Biol. 2019;218(10):3372–3396. doi:10.1083/jcb.201812157
48. Phua DCY, Xu J, Ali SM, Boey A, Gounko NV, Hunziker W. ZO-1 and ZO-2 are required for extra-embryonic endoderm integrity, primitive ectoderm survival and normal cavitation in embryoid bodies derived from mouse embryonic stem cells. PLoS One. 2014;9(6):e99532. doi:10.1371/journal.pone.0099532
49. Umeda K, Ikenouchi J, Katahira-Tayama S, et al. ZO-1 and ZO-2 independently determine where claudins are polymerized in tight-junction strand formation. Cell. 2006;126(4):741–754. doi:10.1016/j.cell.2006.06.043
50. Furuse M, Sasaki H, Fujimoto K, Tsukita S. A single gene product, claudin-1 or-2, reconstitutes tight junction strands and recruits occludin in fibroblasts. J Cell Biol. 1998;143(2):391–401. doi:10.1083/jcb.143.2.391
51. Stache C, Hölsken A, Fahlbusch R, et al. Tight junction protein claudin-1 is differentially expressed in craniopharyngioma subtypes and indicates invasive tumor growth. Neuro-Oncology. 2014;16(2):256–264. doi:10.1093/neuonc/not195
52. Furuse M, Hirase T, Itoh M, et al. Occludin: a novel integral membrane protein localizing at tight junctions. J Cell Biol. 1993;123(6):1777–1788. doi:10.1083/jcb.123.6.1777
53. Ikenouchi J, Furuse M, Furuse K, Sasaki H, Tsukita S, Tsukita S. Tricellulin constitutes a novel barrier at tricellular contacts of epithelial cells. J Cell Biol. 2005;171(6):939–945. doi:10.1083/jcb.200510043
54. Steed E, Rodrigues NT, Balda MS, Matter K. Identification of MarvelD3 as a tight junction-associated transmembrane protein of the occludin family. BMC Cell Biol. 2009;10:1–14. doi:10.1186/1471-2121-10-95
55. Lu R-Y, Yang W-X, Hu Y-J. The role of epithelial tight junctions involved in pathogen infections. Mol Biol Rep. 2014;41(10):6591–6610. doi:10.1007/s11033-014-3543-5
56. Zhao X, Zeng H, Lei L, et al. Tight junctions and their regulation by non-coding RNAs. Int J Biol Sci. 2021;17(3):712–727. doi:10.7150/ijbs.45885
57. Förster C. Tight junctions and the modulation of barrier function in disease. Histochem Cell Biol. 2008;130(1):55–70. doi:10.1007/s00418-008-0424-9
58. Balda MS, Whitney JA, Flores C, González S, Cereijido M, Matter K. Functional dissociation of paracellular permeability and transepithelial electrical resistance and disruption of the apical-basolateral intramembrane diffusion barrier by expression of a mutant tight junction membrane protein. J Cell Biol. 1996;134(4):1031–1049. doi:10.1083/jcb.134.4.1031
59. Monteiro AC, Sumagin R, Rankin CR, et al. JAM-A associates with ZO-2, afadin, and PDZ-GEF1 to activate Rap2c and regulate epithelial barrier function. Mol Biol Cell. 2013;24(18):2849–2860. doi:10.1091/mbc.e13-06-0298
60. Mitchell LA, Ward C, Kwon M, et al. Junctional adhesion molecule A promotes epithelial tight junction assembly to augment lung barrier function. Am J Pathol. 2015;185(2):372–386. doi:10.1016/j.ajpath.2014.10.010
61. Liu Y, Nusrat A, Schnell FJ, et al. Human junction adhesion molecule regulates tight junction resealing in epithelia. J Cell Sci. 2000;113(13):2363–2374. doi:10.1242/jcs.113.13.2363
62. Bazzoni G. The JAM family of junctional adhesion molecules. Curr Opin Cell Biol. 2003;15(5):525–530. doi:10.1016/S0955-0674(03)00104-2
63. Bauer H, Zweimueller-Mayer J, Steinbacher P, Lametschwandtner A, Bauer HC. The dual role of Zonula Occludens (ZO) proteins. J Biomed Biotechnol. 2010;2010:1–11. doi:10.1155/2010/402593
64. Hartsock A, Nelson WJ. Adherens and tight junctions: structure, function and connections to the actin cytoskeleton. Biochimica et Biophysica Acta. 2008;1778(3):660–669. doi:10.1016/j.bbamem.2007.07.012
65. Itoh M, Nagafuchi A, Moroi S, Tsukita S. Involvement of ZO-1 in cadherin-based cell adhesion through its direct binding to α catenin and actin filaments. J Cell Biol. 1997;138(1):181–192. doi:10.1083/jcb.138.1.181
66. Itoh M, Morita K, Tsukita S. Characterization of ZO-2 as a MAGUK family member associated with tight as well as adherens junctions with a binding affinity to occludin and α catenin. J Biol Chem. 1999;274(9):5981–5986. doi:10.1074/jbc.274.9.5981
67. Cordenonsi M, D’Atri F, Hammar E, et al. Cingulin contains globular and coiled-coil domains and interacts with Zo-1, Zo-2, Zo-3, and Myosin. J Cell Biol. 1999;147(7):1569–1582. doi:10.1083/jcb.147.7.1569
68. Citi S, Paschoud S, Pulimeno P, et al. The tight junction protein cingulin regulates gene expression and RhoA signaling. Ann NY Acad Sci. 2009;1165(1):88–98. doi:10.1111/j.1749-6632.2009.04053.x
69. Van Itallie CM, Anderson JM. Architecture of tight junctions and principles of molecular composition. Semin Cell Dev Biol. 2014;36:157–165. doi:10.1016/j.semcdb.2014.08.011
70. Fanning AS, Jameson BJ, Jesaitis LA, Anderson JM. The tight junction protein ZO-1 establishes a link between the transmembrane protein occludin and the actin cytoskeleton. J Biol Chem. 1998;273(45):29745–29753. doi:10.1074/jbc.273.45.29745
71. Wittchen ES, Haskins J, Stevenson BR. Exogenous expression of the amino-terminal half of the tight junction protein ZO-3 perturbs junctional complex assembly. J Cell Biol. 2000;151(4):825–836. doi:10.1083/jcb.151.4.825
72. Wittchen ES, Haskins J, Stevenson BR. Protein interactions at the tight junction: actin has multiple binding partners, and ZO-1 forms independent complexes with ZO-2 and ZO-3. J Biol Chem. 1999;274(49):35179–35185. doi:10.1074/jbc.274.49.35179
73. Ratheesh A, Yap AS. A bigger picture: classical cadherins and the dynamic actin cytoskeleton. Nat Rev Mol Cell Biol. 2012;13(10):673–679. doi:10.1038/nrm3431
74. Belardi B, Hamkins-Indik T, Harris AR, Kim J, Xu K, Fletcher DA. A weak link with actin organizes tight junctions to control epithelial permeability. Dev. Cell. 2020;54(6):792–804.e7. doi:10.1016/j.devcel.2020.07.022
75. Sawada N. Tight junction‐related human diseases. Pathol Int. 2013;63(1):1–12. doi:10.1111/pin.12021
76. Anderson JM, Van Itallie CM. Physiology and function of the tight junction. Cold Spring Harb Perspect Biol. 2009;1(2):a002584. doi:10.1101/cshperspect.a002584
77. Krug SM, Fromm M. Special issue on “the tight junction and its proteins: more than just a barrier”. Int J Mol Sci. 2020;21(13):4612. doi:10.3390/ijms21134612
78. Schneeberger EE, Lynch RD. Structure, function, and regulation of cellular tight junctions. Am J Physiol Lung Cell Mol Physiol. 1992;262(6):L647–L661. doi:10.1152/ajplung.1992.262.6.L647
79. van Meer G, Simons K. The function of tight junctions in maintaining differences in lipid composition between the apical and the basolateral cell surface domains of MDCK cells. EMBO J. 1986;5(7):1455–1464. doi:10.1002/j.1460-2075.1986.tb04382.x
80. Ikenouchi J, Suzuki M, Umeda K, et al. Lipid polarity is maintained in absence of tight junctions. J Biol Chem. 2012;287(12):9525–9533. doi:10.1074/jbc.M111.327064
81. Qin R, Steel A, Fazel N. Oral mucosa biology and salivary biomarkers. Clin Dermatol. 2017;35(5):477–483. doi:10.1016/j.clindermatol.2017.06.005
82. Brizuela M, Winters R. Histology, oral mucosa. In: StatPearls. StatPearls Publishing; 2024.
83. Şenel S. An overview of physical, microbiological and immune barriers of oral mucosa. Int J Mol Sci. 2021;22(15):7821. doi:10.3390/ijms22157821
84. Mendenhall WM, Foote RL, Sandow PL, Fernandes RP. Chapter 32 - oral cavity. In: Gunderson LL, Tepper JE, editors. Clinical Radiation Oncology.
85. Bosshardt DD, Lang NP. The junctional epithelium: from health to disease. J Dent Res. 2005;84(1):9–20. doi:10.1177/154405910508400102
86. Presland RB, Dale BA. Epithelial structural proteins of the skin and oral cavity: function in health and disease. Crit Rev Oral Biol Med. 2000;11(4):383–408. doi:10.1177/10454411000110040101
87. Nakamura M. Histological and immunological characteristics of the junctional epithelium. Japan Dent Sci Rev. 2018;54(2):59–65. doi:10.1016/j.jdsr.2017.11.004
88. Schroeder HE, Listgarten MA. The gingival tissues: the architecture of periodontal protection. Periodontology 2000. 1997;13(1):91–120. doi:10.1111/j.1600-0757.1997.tb00097.x
89. Egelberg J, Badersten A, Claffey N. Odontoscience. In: Periodontal Examination. OdontoScience; 1997.
90. Presland RB, Jurevic RJ. Making sense of the epithelial barrier: what molecular biology and genetics tell us about the functions of oral mucosal and epidermal tissues. J Dent Educ. 2002;66(4):564–574. doi:10.1002/j.0022-0337.2002.66.4.tb03536.x
91. Abusleme L, Dupuy AK, Dutzan N, et al. The subgingival microbiome in health and periodontitis and its relationship with community biomass and inflammation. ISME J. 2013;7(5):1016–1025. doi:10.1038/ismej.2012.174
92. Hajishengallis G, Darveau RP, Curtis MA. The keystone-pathogen hypothesis. Nat Rev Microbiol. 2012;10(10):717–725. doi:10.1038/nrmicro2873
93. Bibi T, Khurshid Z, Rehman A, Imran E, Srivastava KC, Shrivastava D. Gingival Crevicular Fluid (GCF): a diagnostic tool for the detection of periodontal health and diseases. Molecules. 2021;26(5):1208. doi:10.3390/molecules26051208
94. Lamster IB, Ahlo JK. Analysis of gingival crevicular fluid as applied to the diagnosis of oral and systemic diseases. Ann NY Acad Sci. 2007;1098(1):216–229. doi:10.1196/annals.1384.027
95. Khurshid Z, Mali M, Naseem M, Najeeb S, Zafar M. Human Gingival Crevicular Fluids (GCF) Proteomics: an Overview. Dent J. 2017;5(1):12. doi:10.3390/dj5010012
96. Zhang Q, Chen B, Zhu D, Yan F. Biomarker levels in gingival crevicular fluid of subjects with different periodontal conditions: a cross-sectional study. Arch Oral Biol. 2016;72:92–98. doi:10.1016/j.archoralbio.2016.08.020
97. Pei J, Li F, Xie Y, Liu J, Yu T, Feng X. Microbial and metabolomic analysis of gingival crevicular fluid in general chronic periodontitis patients: lessons for a predictive, preventive, and personalized medical approach. EPMA Journal. 2020;11:197–215. doi:10.1007/s13167-020-00202-5
98. Bezerra Júnior AA, Pallos D, Cortelli JR, Saraceni CHC, Queiroz CS. Evaluation of organic and inorganic compounds in the saliva of patients with chronic periodontal disease. Revista odonto ciência. 2010;25:234–238. doi:10.1590/S1980-65232010000300003
99. Sheth TS, Verma SJ. Analysis of aspartate aminotransferase in gingival crevicular fluid: a study with initial therapy. J Indian Soc Periodontol. 2011;15(3):235–239. doi:10.4103/0972-124X.85666
100. Eley B, Cox S. The relationship between gingival crevicular fluid cathepsin B activity and periodontal attachment loss in chronic periodontitis patients: a 2‐year longitudinal study. J Periodontal Res. 1996;31(6):381–392. doi:10.1111/j.1600-0765.1996.tb00507.x
101. Fassbender WJ, Steinhauer B, Stracke H, Schumm-Draeger P-M, Usadel K-H. Validation of a new automated immunoassay for measurement of intact osteocalcin. Clin Lab. 2002;48(1–2):31–38.
102. Khiste SV, Ranganath V, Nichani AS, Rajani V. Critical analysis of biomarkers in the current periodontal practice. J Indian Soc Periodontol. 2011;15(2):104–110. doi:10.4103/0972-124X.84376
103. Airila‐Månsson S, Söder B, Kari K, Meurman JH. Influence of combinations of bacteria on the levels of prostaglandin E2, interleukin‐1β, and granulocyte elastase in gingival crevicular fluid and on the severity of periodontal disease. J Periodontol. 2006;77(6):1025–1031. doi:10.1902/jop.2006.050208
104. Huynh QN, Wang S, Tafolla E, et al. Specific fibronectin fragments as markers of periodontal disease status. J Periodontol. 2002;73(10):1101–1110. doi:10.1902/jop.2002.73.10.1101
105. Subbarao KC, Nattuthurai GS, Sundararajan SK, Sujith I, Joseph J, Syedshah YP. Gingival crevicular fluid: an overview. J Pharm Bioallied Sci. 2019;11(Suppl 2):S135–S139. doi:10.4103/JPBS.JPBS_56_19
106. Capaldo CT, Nusrat A. Cytokine regulation of tight junctions. Biochim Biophys Acta. 2009;1788(4):864–871. doi:10.1016/j.bbamem.2008.08.027
107. Li Q, Zhang Q, Wang M, et al. Interferon-γ and tumor necrosis factor-α disrupt epithelial barrier function by altering lipid composition in membrane microdomains of tight junction. Clin Immunol. 2008;126(1):67–80. doi:10.1016/j.clim.2007.08.017
108. Vitkov L, Klappacher M, Hannig M, Krautgartner WD. Extracellular neutrophil traps in periodontitis. J Periodontal Res. 2009;44(5):664–672. doi:10.1111/j.1600-0765.2008.01175.x
109. Vitkov L, Klappacher M, Hannig M, Krautgartner WD. Neutrophil fate in gingival crevicular fluid. Ultrastruct Pathol. 2010;34(1):25–30. doi:10.3109/01913120903419989
110. Vitkov L, Minnich B, Knopf J, Schauer C, Hannig M, Herrmann M. NETs are double-edged swords with the potential to aggravate or resolve periodontal inflammation. Cells. 2020;9(12):2614. doi:10.3390/cells9122614
111. Vitkov L, Muñoz LE, Schoen J, et al. Neutrophils orchestrate the periodontal pocket. Front Immunol. 2021;12. doi:10.3389/fimmu.2021.788766
112. Matsui T, Amagai M. Dissecting the formation, structure and barrier function of the stratum corneum. Int Immunol. 2015;27(6):269–280. doi:10.1093/intimm/dxv013
113. Yoshida K, Yokouchi M, Nagao K, Ishii K, Amagai M, Kubo A. Functional tight junction barrier localizes in the second layer of the stratum granulosum of human epidermis. J Dermatological Sci. 2013;71(2):89–99. doi:10.1016/j.jdermsci.2013.04.021
114. Ko YK, Hong S, Kim HM, et al. Characterization of junctional structures in the gingival epithelium as barriers against bacterial invasion. J Periodontal Res. 2022;57(4):799–810. doi:10.1111/jre.13003
115. Belibasakis GN, Kast JI, Thurnheer T, Akdis CA, Bostanci N. The expression of gingival epithelial junctions in response to subgingival biofilms. Virulence. 2015;6(7):704–709. doi:10.1080/21505594.2015.1081731
116. Dejana E. Endothelial cell–cell junctions: happy together. Nat Rev Mol Cell Biol. 2004;5(4):261–270. doi:10.1038/nrm1357
117. Padilla C, Lobos O, Hubert E, et al. Periodontal pathogens in atheromatous plaques isolated from patients with chronic periodontitis. J Periodontal Res. 2006;41(4):350–353. doi:10.1111/j.1600-0765.2006.00882.x
118. Hatakeyama S, Ishida K, Takeda Y. Changes in cell characteristics due to retinoic acid; specifically, a decrease in the expression of claudin‐1 and increase in claudin‐4 within tight junctions in stratified oral keratinocytes. J Periodontal Res. 2010;45(2):207–215. doi:10.1111/j.1600-0765.2009.01219.x
119. Maria OM, Kim J-WM, Gerstenhaber JA, Baum BJ, Tran SD. Distribution of tight junction proteins in adult human salivary glands. J Histochem Cytochem. 2008;56(12):1093–1098. doi:10.1369/jhc.2008.951780
120. Baker OJ. Current trends in salivary gland tight junctions. Tissue Barriers. 2016;4(3):e1162348. doi:10.1080/21688370.2016.1162348
121. Carpenter GH. The secretion, components, and properties of saliva. Annu Rev Food Sci Technol. 2013;4(1):267–276. doi:10.1146/annurev-food-030212-182700
122. Powell D. Barrier function of epithelia. Am J Physiol Gastrointest Liver Physiol. 1981;241(4):G275–G288. doi:10.1152/ajpgi.1981.241.4.G275
123. Rosenthal R, Milatz S, Krug SM, et al. Claudin-2, a component of the tight junction, forms a paracellular water channel. J Cell Sci. 2010;123(11):1913–1921. doi:10.1242/jcs.060665
124. Cong X, Zhang Y, Shi L, et al. Activation of transient receptor potential vanilloid subtype 1 increases expression and permeability of tight junction in normal and hyposecretory submandibular gland. Lab Invest. 2012;92(5):753–768. doi:10.1038/labinvest.2012.12
125. Quesnell RR, Erickson J, Schultz BD. Apical electrolyte concentration modulates barrier function and tight junction protein localization in bovine mammary epithelium. Am J Physiol Cell Physiol. 2007;292(1):C305–C318. doi:10.1152/ajpcell.00567.2005
126. Baker OJ, Camden JM, Redman RS, et al. Proinflammatory cytokines tumor necrosis factor-α and interferon-γ alter tight junction structure and function in the rat parotid gland Par-C10 cell line. Am J Physiol Cell Physiol. 2008;295(5):C1191–C1201. doi:10.1152/ajpcell.00144.2008
127. Takeuchi H, Sasaki N, Yamaga S, Kuboniwa M, Matsusaki M, Amano A. Porphyromonas gingivalis induces penetration of lipopolysaccharide and peptidoglycan through the gingival epithelium via degradation of junctional adhesion molecule 1. PLoS Pathog. 2019;15(11):e1008124. doi:10.1371/journal.ppat.1008124
128. Guo W, Wang P, Liu Z-H, Ye P. Analysis of differential expression of tight junction proteins in cultured oral epithelial cells altered by Porphyromonas gingivalis, Porphyromonas gingivalis lipopolysaccharide, and extracellular adenosine triphosphate. Int J Oral Sci. 2018;10(1):e8–e8. doi:10.1038/ijos.2017.51
129. Chen W, Alshaikh A, Kim S, et al. Porphyromonas gingivalis impairs oral epithelial barrier through targeting GRHL2. J Dent Res. 2019;98(10):1150–1158. doi:10.1177/0022034519865184
130. Martínez-García M, Hernández-Lemus E. Periodontal inflammation and systemic diseases: an overview. Front Physiol. 2021;12:709438. doi:10.3389/fphys.2021.709438
131. Graves D. Cytokines that promote periodontal tissue destruction. J Periodontol. 2008;79(8S):1585–1591. doi:10.1902/jop.2008.080183
132. Madianos PN, Bobetsis YA, Kinane DF. Generation of inflammatory stimuli: how bacteria set up inflammatory responses in the gingiva. J Clin Periodontol. 2005;32(s6):57–71. doi:10.1111/j.1600-051X.2005.00821.x
133. Yucel-Lindberg T, Båge T. Inflammatory mediators in the pathogenesis of periodontitis. Expert Rev Mol Med. 2013;15:e7. doi:10.1017/erm.2013.8
134. Ohlrich E, Cullinan M, Seymour G. The immunopathogenesis of periodontal disease. Aust Dent J. 2009;54:S2–S10. doi:10.1111/j.1834-7819.2009.01139.x
135. Panagakos F, Scannapieco F. Periodontal inflammation: from gingivitis to systemic disease. Gingival dis. 2011;2011:155–168. doi:10.5772/37923
136. Hernández-Monjaraz B, Santiago-Osorio E, Monroy-García A, Ledesma-Martínez E, Mendoza-Núñez VM. Mesenchymal stem cells of dental origin for inducing tissue regeneration in periodontitis: a mini-review. Int J Mol Sci. 2018;19(4):944. doi:10.3390/ijms19040944
137. Thilander H. Epithelial changes in gingivitis: an electron microscopic study. J Periodontal Res. 1968;3(4):303–312. doi:10.1111/j.1600-0765.1968.tb01936.x
138. Puzhankara L, Rajagopal A, Kedlaya MN, Karmakar S, Nayak N, Shanmugasundaram S. Cell junctions in periodontal health and disease: an insight. Eur J Dent. 2024;18(2):448–457. doi:10.1055/s-0043-1775726
139. Shen L. Tight junctions on the move: molecular mechanisms for epithelial barrier regulation. Ann NY Acad Sci. 2012;1258(1):9–18. doi:10.1111/j.1749-6632.2012.06613.x
140. Pindborg JJ. Tobacco and gingivitis. J Dent Res. 1947;26(3):261. doi:10.1177/00220345470260030901
141. Yamaga S, Tanigaki K, Nakamura E, et al. Cigarette smoke extract impairs gingival epithelial barrier function. Sci Rep. 2023;13(1):9228. doi:10.1038/s41598-023-36366-z
142. Ouban A, Ahmed A. Analysis of the distribution and expression of claudin-1 tight junction protein in the oral cavity. Appl Immunohistochem Mol Morphol. 2015;23(6):444–448. doi:10.1097/PAI.0000000000000104
143. Zejc T, Piontek J, Schulzke J-D, Fromm M, Ervens J, Rosenthal R. Clinical significance of claudin expression in oral squamous cell carcinoma. Int J Mol Sci. 2022;23(19):11234. doi:10.3390/ijms231911234
144. Lourenço SV, Coutinho‐Camillo CM, Buim MEC, et al. Claudin‐7 down‐regulation is an important feature in oral squamous cell carcinoma. Histopathology. 2010;57(5):689–698. doi:10.1111/j.1365-2559.2010.03685.x
145. Melchers L, De Bruin LB, Schnell U, et al. Lack of claudin-7 is a strong predictor of regional recurrence in oral and oropharyngeal squamous cell carcinoma. Oral Oncol. 2013;49(10):998–1005. doi:10.1016/j.oraloncology.2013.07.008
146. Yoshizawa K, Nozaki S, Kato A, et al. Loss of claudin-7 is a negative prognostic factor for invasion and metastasis in oral squamous cell carcinoma. Oncol Rep. 2013;29(2):445–450. doi:10.3892/or.2012.2161
147. Phattarataratip E, Sappayatosok K. Expression of claudin-5, claudin-7 and occludin in oral squamous cell carcinoma and their clinico-pathological significance. J Clin Exp Dent. 2016;8(3):e299–306. doi:10.4317/jced.52801
148. Lee SS, Choi JH, Lim SM, Kim GJ, Lee SK, Jeon YK. Alteration of pituitary tumor transforming gene 1 by MicroRNA-186 and 655 regulates invasion ability of human oral squamous cell carcinoma. Int J Mol Sci. 2021;22(3):1021. doi:10.3390/ijms22031021
149. Vermeer PD, Denker J, Estin M, et al. MMP9 modulates tight junction integrity and cell viability in human airway epithelia. Am J Physiol Lung Cell Mol Physiol. 2009;296(5):L751–L762. doi:10.1152/ajplung.90578.2008
150. Kim S, Park S, Moon E-H, Kim GJ, Choi J. Hypoxia disrupt tight junctions and promote metastasis of oral squamous cell carcinoma via loss of par3. Cancer Cell Int. 2023;23(1):79. doi:10.1186/s12935-023-02924-8
151. Sobel G, Páska C, Szabó I, Kiss A, Kádár A, Schaff Z. Increased expression of claudins in cervical squamous intraepithelial neoplasia and invasive carcinoma. Human Pathol. 2005;36(2):162–169. doi:10.1016/j.humpath.2004.12.001
152. Kominsky SL, Argani P, Korz D, et al. Loss of the tight junction protein claudin-7 correlates with histological grade in both ductal carcinoma in situ and invasive ductal carcinoma of the breast. Oncogene. 2003;22(13):2021–2033. doi:10.1038/sj.onc.1206199
153. Kyuno D, Takasawa A, Kikuchi S, Takemasa I, Osanai M, Kojima T. Role of tight junctions in the epithelial-to-mesenchymal transition of cancer cells. Biochimica et Biophysica Acta. 2021;1863(3):183503. doi:10.1016/j.bbamem.2020.183503
154. Tandler B. Structure of the duct system in mammalian major salivary glands. Microsc Res Tech. 1993;26(1):57–74. doi:10.1002/jemt.1070260107
155. Tandler B. Introduction to mammalian salivary glands. Microsc Res Tech. 1993;26(1):1–4. doi:10.1002/jemt.1070260102
156. Maślińska M, Przygodzka M, Kwiatkowska B, Sikorska-Siudek K. Sjögren’s syndrome: still not fully understood disease. Rheumatol Int. 2015;35(2):233–241. doi:10.1007/s00296-014-3072-5
157. De Paiva CS, Jones DB, Stern ME, et al. Altered mucosal microbiome diversity and disease severity in sjögren syndrome. Sci Rep. 2016;6(1):23561. doi:10.1038/srep23561
158. Adamson TC, Fox RI, Frisman DM, Howell FV. Immunohistologic analysis of lymphoid infiltrates in primary Sjogren’s syndrome using monoclonal antibodies. J Immunol. 1983;130(1):203–208. doi:10.4049/jimmunol.130.1.203
159. Shiboski SC, Shiboski CH, Criswell LA, et al. American college of rheumatology classification criteria for sjögren’s syndrome: a data‐driven, expert consensus approach in the Sjögren’s international collaborative clinical alliance cohort. Arthritis Care Res. 2012;64(4):475–487. doi:10.1002/acr.21591
160. Verstappen GM, Pringle S, Bootsma H, Kroese FG. Epithelial–immune cell interplay in primary Sjögren syndrome salivary gland pathogenesis. Nat Rev Rheumatol. 2021;17(6):333–348. doi:10.1038/s41584-021-00605-2
161. Nikitakis N, Papaioannou W, Sakkas L, Kousvelari E. The autoimmunity–oral microbiome connection. Oral Dis. 2017;23(7):828–839. doi:10.1111/odi.12589
162. Fox RI, Kang HI, Ando D, Abrams J, Pisa E. Cytokine mRNA expression in salivary gland biopsies of Sjögren’s syndrome. J Immunol. 1994;152(11):5532–5539. doi:10.4049/jimmunol.152.11.5532
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.