")
Back to Journals » International Journal of Nanomedicine » Volume 18
A Powerful Tool in the Treatment of Myocardial Ischemia-Reperfusion Injury: Natural and Nanoscale Modified Small Extracellular Vesicles Derived from Mesenchymal Stem Cells
Authors Zhou Z, Zhang X, Wang S, Wang X, Mao J
Received 8 October 2023
Accepted for publication 12 December 2023
Published 28 December 2023 Volume 2023:18 Pages 8099—8112
DOI https://doi.org/10.2147/IJN.S443716
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Lijie Grace Zhang
Zhou Zhou,1,2,* Xuan Zhang,1,* Shuai Wang,1 Xianliang Wang,1 Jingyuan Mao1
1Cardiovascular Department, First Teaching Hospital of Tianjin University of Traditional Chinese Medicine/National Clinical Research Center for Chinese Medicine Acupuncture and Moxibustion, Tianjin, 300381, People’s Republic of China; 2Graduate School, Tianjin University of Traditional Chinese Medicine, Tianjin, 301617, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Xianliang Wang; Jingyuan Mao, Email [email protected]; [email protected]
Abstract: Myocardial ischemia-reperfusion injury (MI/RI) constitutes a pivotal determinant impacting the long-term prognosis of individuals afflicted by ischemic cardiomyopathy subsequent to reperfusion therapy. Stem cells have garnered extensive application within the realm of MI/RI investigation, yielding tangible outcomes. Stem cell therapy encounters certain challenges in its application owing to the complexities associated with stem cell acquisition, a diminished homing rate, and a brief in vivo lifespan. Small extracellular vesicles (sEV) originating from mesenchymal stem cells (MSCs) have been demonstrated to possess the benefits of abundant availability, reduced immunogenicity, and a diminished tumorigenic incidence. They can exert their effects on damaged organs, improving injuries by transporting a lot of constituents, including proteins, RNA, lipid droplets, and more. This phenomenon has garnered substantial attention in the context of MI/RI treatment. Simultaneously, MSC-derived sEV (MSC-sEV) can exhibit enhanced therapeutic advantages through bioengineering modifications, biomaterial incorporation, and natural drug interventions. Within this discourse, we shall appraise the utilization of MSC-sEV and their derivatives in the context of MI/RI treatment, aiming to offer valuable insights for future research endeavors related to MI/RI.
Keywords: myocardial ischemia-reperfusion injury, mesenchymal stem cells, small extracellular vesicles, nanoscale modification, natural drug
Introduction
As medical technology and management strategies continue to advance, the rate of reperfusion therapy for individuals afflicted by ischemic cardiomyopathy has seen perpetual enhancement. Consequently, the short-term mortality rate among patients has been significantly ameliorated.1,2 Nonetheless, the Global Burden of Disease Report for the year 2022 underscores that on a global scale, fatalities attributed to ischemic cardiomyopathy maintain their preeminent position as the foremost cause of death.3 The escalating mortality rate among individuals afflicted by ischemic cardiomyopathy is primarily associated with cardiac insufficiency stemming from myocardial infarction. Notably, myocardial ischemia-reperfusion injury (MI/RI), induced by the reperfusion therapy itself, assumes a pivotal role in precipitating the deterioration of cardiac function.4,5 Henceforth, the prevention of MI/RI, along with the revival of myocardial stunning and hibernation instigated by reperfusion injury, emerges as efficacious stratagems aimed at augmenting the enduring prognosis of individuals grappling with ischemic cardiomyopathy.6,7
Since the advent of the 21st century, regenerative therapy involving stem cells has achieved remarkable milestones in the treatment of myocardial injury, particularly ischemic damage.8–10 Among these, mesenchymal stem cells (MSCs) have distinguished themselves amidst the diverse array of stem cell types, owing to their innate attributes of self-renewal, versatile differentiation capabilities, and substantial autocrine and paracrine capacities.11,12 A notable instance of their clinical application can be found in the RIMECARD trial, wherein the intravenous administration of mesenchymal stem cells derived from umbilical cord tissue (UCMSCs) significantly ameliorated patients’ cardiac function and overall quality of life.13 Furthermore, the HUC-HEART study demonstrated that patients who underwent coronary artery bypass grafting (CABG) in conjunction with UCMSC administration exhibited notably superior cardiac function one year post-surgery compared to those who received CABG alone.14,15 These outcomes inspired our faith in the potential of stem cell therapies. Nonetheless, the direct utilization of stem cells in the context of MI/RI therapy is not without challenges, including issues of low survival rates, abbreviated retention periods, tumorigenicity, and complexities surrounding standardized production.16–18 Furthermore, there remains a contentious debate regarding the potential for stem cell transplantation to precipitate ventricular arrhythmia.19 Small extracellular vesicles (sEV) represent lipid bilayered vesicles, secreted by cells, with a diameter less than 200nm, facilitating the transfer of cytoplasmic cargo and mediating intercellular transport.20 Small extracellular vesicles, particularly those derived from Mesenchymal Stem Cells (MSC-sEV), are abundant in a plethora of functional proteins, RNA, and lipids, yielding myocardial protective effects akin to their parent cells.21 Simultaneously, cell-free therapeutic approaches relying on sEV circumvent issues such as immune responses, tumorigenicity, and infusion-related toxicity often associated with stem cell-based interventions, while enabling large-scale production.22,23
Regarding the pathogenesis, MI/RI refers to the secondary damage to myocardial tissue following vascular reconstruction due to mechanisms like metabolic changes, mitochondrial dysfunction, inflammatory response, relaxation of autophagy regulation, and excessive production of reactive oxygen species.24 These mechanisms lead to alterations in microvascular endothelial permeability, increased extravasation of fluids, immune cell infiltration, ultimately resulting in secondary damage to myocardial tissue. At present, clinical treatments for MI/RI primarily involve strengthening antiplatelet therapies, anticoagulation, and vasodilation of microvessels.25–27 However, the effectiveness of these strategies still requires further clarification, and they might also increase the risk of bleeding for patients.28 Research indicates that MSC-sEV can mitigate myocardial damage induced by ischemia-reperfusion by modulating various pathological processes in cardiomyocytes, such as apoptosis, autophagy, and inflammatory responses.29,30 This cell-free therapy presents a new approach for treating MI/RI.
Consequently, the development of biomedical agents associated with MSC-sEV has emerged as a pivotal breakthrough in the realm of MI/RI treatment. It is imperative to highlight that a consensus regarding the specific markers for sEV subtypes remains elusive. Therefore, in this discourse, we align ourselves with the standpoint set forth by the International Society for Extracellular Vesicles (MISEV2018) and refrain from employing nomenclature such as “exosomes”, “particles”, or “microvesicles”, as has been customary in numerous studies, opting instead for the term “small Extracellular vesicles (sEV)” as a precise specification.31,32 Concurrently, this review encompasses a diverse array of recent investigations into the application of MSC-sEV for the enhancement of MI/RI outcomes, delving into areas of nanoscale modification and natural drug intervention within the context of MSC-sEV. As research continues to advance, interdisciplinary collaboration is poised to exert a formidable influence in propelling the evolution of MI/RI therapeutics.
MSCs in MI/RI Regulation
Mesenchymal stem cells (MSCs) can be procured from a diverse array of tissues, encompassing bone marrow, adipose tissue, umbilical cord, placenta, synovial membrane, and dental pulp.33–38 These MSCs have garnered significant attention within the sphere of regenerative medicine research owing to their multifaceted abilities, encompassing the facilitation of wound healing, immune modulation, angiogenesis, anti-inflammatory effects, and antioxidant properties.39 Presently, the MSCs employed in studies pertaining to MI/RI therapy predominantly originate from bone marrow, adipose tissue, umbilical cord, and heart-resident mesenchymal stem cells. While these MSCs share commonalities in surface marker expression, certain investigations have elucidated variances in marker profiles between them.40–47 Functionally, it has been posited that bone marrow mesenchymal stem cells (BMSCs) may exhibit a superior capacity for osteogenesis and angiogenesis in comparison to adipose-derived mesenchymal stem cells (ADMSCs) and UCMSCs.48,49 BMSCs, however, display limitations in terms of their extraction and proliferation capabilities, whereas UCMSCs demonstrate a heightened propensity for proliferation relative to ADMSCs.50,51 Furthermore, UCMSCs and ADMSCs are characterized by their ample presence and robust anti-aging attributes, rendering them increasingly prominent candidates for therapeutic endeavors.52,53 While investigations comparing the functional attributes of cardiac mesenchymal stem cells (CMSCs) with those of other stem cell types are relatively scarce, prior studies have indicated that CMSCs exert a regulatory influence over numerous physiological and pathological processes within the cardiac domain.54 The distinctive attributes of these aforementioned MSCs are summarized in Table 1.
 |
Table 1 Characteristics of Each Mesenchymal Stem Cell |
The Regulation of Natural MSC-sEV on Multiple Pathological Pathways in MI/RI
Apoptosis
Apoptosis, being the classical form of programmed cell demise, represents the predominant mode of cellular attrition in response to ischemic or oxidative stimuli.55 Under duress, the Protein kinase B/Nuclear factor kappa-B (AKT/NFκB) signaling cascade is activated, subsequently up-regulating the downstream classical apoptotic pathway encompassing Bcl-2/Bax/Caspase-3, thereby instigating the commencement of apoptosis.56,57 In the mouse model of MI/RI, wherein the left anterior descending branch was ligated and subsequently recanalized, sEV derived from ADMSCs, referred to as ADMSC-sEV, conveyed miR-221/222 to cardiomyocytes intracellularly. This internalization event mitigated the down-regulation of myocardial miR-221/222 induced by MI/RI, thereby inhibiting apoptosis.58 A dual fluorescein reporter gene assay confirmed that the p53 up-regulated modulator of apoptosis (PUMA) and ETS proto-oncogene 1 (ETS-1) are the direct targets of miR-221/222, both of which can be silenced by miR-221/222. In vitro studies substantiated that ADMSC-sEV inhibited apoptosis in a hypoxia/reoxygenation (H/R) cardiomyocyte model by regulating the PUMA/AKT/NFκB pathway via miR-221/222 and curbed cardiomyocyte hypertrophy by restraining ETS-1 expression via miR-221/222. ETS-1 has been established as a pivotal gene in the provocation of cardiomyocyte hypertrophy. Although cell hypertrophy is commonly regarded as a compensatory response by surviving cells to systolic wall tension subsequent to cardiomyocyte depletion, hypertrophic cardiomyocytes also contribute significantly to abnormal wall motion, decreased compliance, and eventual heart failure.59,60 miR-486-5p, harbored within bone marrow-derived mesenchymal stem cell-secreted extracellular vesicles (BMSC-sEV), has been shown to down-regulate the expression of Phosphatase and tensin homolog (PTEN) in the myocardium of rat models afflicted with MI/RI. In vitro investigations have demonstrated that BMSC-sEV mitigate PTEN expression in cardiomyocytes via miR-486-5p, thereby activating the Phosphoinositide 3-kinase (PI3K)/AKT signaling pathway. This activation in turn regulates the downstream Bcl-2/Bax/CASP3 pathway, repressing the apoptotic cascade in cardiomyocytes under H/R conditions.61 Consequently, the modulation of cardiomyocyte apoptosis by MSC-sEV stands as a pivotal therapeutic approach for mitigating myocardial MI/RI injury.
Pyroptosis
The classical pathway of pyroptosis is mediated through the NLRP3 inflammasome/Caspase-1 (CASP1) pathway, with the NLRP3 inflammasome composed of Nod-like receptor protein 3 (NLRP3), apoptosis-associated speck-like protein containing a CARD (ASC), and CASP1. This pathway becomes activated under ischemia/reperfusion (I/R) conditions, resulting in the cleavage of the pyroptosis executive protein Gasdermin-D (GSDMD) by activated CASP1 within the NLRP3 inflammasome. Subsequently, the N-terminal fragment of GSDMD (GSDMD-N) undergoes oligomerization and integrates into the cell membrane, eliciting cardiomyocyte swelling, membrane rupture, and the release of inflammatory mediators, culminating in inflammatory cell death.62 Research has substantiated that NLRP3 is a direct target of miR-320b. In cell models subjected to H/R, BMSC-sEV delivered miR-320b, effectively suppressing the process of pyroptosis in cardiomyocytes by directly inhibiting the NLRP3/CASP1 pathway.63 Furthermore, beyond its direct inhibition of NLRP3 inflammation, the regulation of the pyroptosis-effector protein GSDMD represents a promising anti-MI/RI strategy.64 In both mouse MI/RI models and cell models exposed to H/R, miR-182-5p within BMSC-sEV attenuated the expression of GSDMD and inflammatory-associated proteins ASC and CASP1, thereby mitigating cell pyroptosis. Importantly, miR-182-5p exhibited no influence on NLRP3 expression.65 This underscores the potential of BMSC-sEV in inhibiting MI/RI by concurrently targeting multiple facets of pyroptosis through the diverse constituents present in BMSC-sEV.
Ferroptosis
Ferroptosis stands as an oxidative cell demise pathway reliant upon iron ions, distinguishing itself from cell necrosis, apoptosis, and autophagy through its unique cellular death mechanisms.66 Ferroptosis ensues when the intracellular glutathione (GSH)-dependent antioxidant system is disrupted, such as down-regulation of glutathione peroxidase GPX4, leading to the consequent up-regulation of lipid-activating enzymes, including Acyl-CoA synthetase (ACSL4). This results in the accumulation of lipid-reactive oxygen species and the induction of oxidative cell death.67 Evidently, ferroptosis plays a significant role in myocardial I/R injury, where oxidative stress is a prominent pathological factor.68 Zhang et al69 harnessed BMSC-sEV to deliver the LncRNA miR9-3 host gene (miR9-3hg) to mouse cardiomyocytes subjected to H/R conditions, effectively suppressing the ferroptotic process. miR9-3hg inhibits ferroptosis by targeting Pumilio RNA binding family member 2 (PUM2) protein. Protein co-immunoprecipitation assays unveiled a protein interaction between PUM2 and Peroxiredoxin 6 (PRDX6), with PRDX6 emerging as a pivotal player in safeguarding cells against ferroptosis.70 Thus, BMSC-sEV orchestrates the PUM2/PRDX6 signaling pathway within cardiomyocytes through miR9-3hg, averting ferroptosis induced by H/R and ameliorating reperfusion injury. Additionally, Song et al reported the substantial up-regulation of Divalent Metal Transporter 1 (DMT1) in mouse cardiomyocytes subjected to H/R conditions. DMT1 overexpression significantly promoted the ferroptotic process in cardiomyocytes. Luciferase gene reporter experiments elucidated that miR-23a-3p directly targeted DMT1. UCMSC-sEV intervention in H/R cardiomyocytes markedly repressed DMT1 expression in cardiomyocytes, thereby dampening the ferroptotic process. Mechanistic studies have suggested that miR-23a-3p, harbored within UCMSC-sEV, curtails DMT1 expression in cardiomyocytes.71
Autophagy
Autophagy, a highly conserved intracellular program, encompasses three primary forms, namely macroautophagy, microautophagy, and chaperone-mediated autophagy. Among these, macroautophagy has been most extensively and comprehensively studied in the context of disease.72 Autophagy permits cells to eliminate aged and damaged cellular components, including proteins, lipids, and dysfunctional organelles, thereby promoting cellular longevity. Nevertheless, under certain unmitigated lethal stimuli, autophagy can precipitate cell death, referred to as autophagic death.73–75 The equilibrium of autophagy is pivotal in determining the ultimate fate of cardiomyocytes in ischemic cardiomyopathy. Chen et al revealed the heightened expression of miR-143-3p within BMSC-sEV. BMSC-sEV effectively curtailed apoptosis and autophagy in H9C2 cardiomyocytes exposed to H/R conditions. miR-143-3p played a key role in inhibiting the cardiomyocyte Checkpoint Kinase 2 (CHK2) through targeted repression.76 CHK2 is instrumental in activating the CHK2/Beclin1 signaling cascade, which in turn promotes autophagy. Thus, miR-143-3p inhibition of CHK2/Beclin1 pathway safeguards cardiomyocytes from excessive autophagy-induced damage during reperfusion.77 In the realm of MI/RI treatment, beyond safeguarding cardiomyocytes to enhance cardiac function, the preservation of cardiac microvascular endothelial cells (CMECs) to ensure adequate blood supply to ischemic regions has emerged as a crucial therapeutic strategy.78 Research has delineated that heightened miR-143-3p expression under MI/RI conditions hinders the Bcl-2/Beclin1 signaling axis, fostering apoptosis and autophagy in CMECs. Within this context, Urothelial carcinoma-associated 1 (UCA1) contained within UCMSC-sEV competes for miR-143-3p binding in CMEC cells, enhancing cell viability.79 This underscores the significance of maintaining autophagic balance for cell survival, and MSC-sEV emerges as a potent regulator of autophagic equilibrium within diverse cardiac microenvironmental cell types, promising substantial applicability in the prevention and management of MI/RI.
Macrophage Polarization
The MI/RI process is invariably accompanied by an inflammatory cascade within the cardiac microenvironment, with Macrophages (Mφ) playing a pivotal role in orchestrating the inflammatory environment.80 In the initial stages of injury, Mφ predominantly undertake the task of debris clearance, manifesting the pro-inflammatory M1 phenotype. Subsequently, Mφ gradually polarize towards the anti-inflammatory M2 phenotype, secreting not only anti-inflammatory mediators but also a multitude of growth factors, crucial for wound healing and scar tissue formation.81,82 Shen et al demonstrated that myocardial infusion of BMSCs ameliorated cardiac function in MI/RI mouse models by modulating M2 polarization of macrophages, and this effect was compromised upon the inhibition of miR-21-5p expression within BMSCs. This underscores the pivotal role of miR-21-5p in regulating the M1/M2 equilibrium within myocardial macrophages orchestrated by BMSC-sEV.83 An additional study observed that BMSC-sEV, apart from significantly enhancing cardiac function and reducing myocardial infarction size in MI/RI mice, effectively mitigated inflammatory infiltration in myocardial tissue. BMSC-sEV were internalized by macrophages, promoting M2 polarization contingent on miR-182.84 Mechanistic studies unveiled that miR-182 within BMSC-sEV inhibited Toll-like receptor 4 (TLR4) expression, resulting in the down-regulation of the Myeloid differentiation primary response protein MyD88 (MyD88)/NF-κB signaling pathway and the concurrent up-regulation of the PI3K/Akt signaling pathway, thus fostering M2 macrophage polarization. This underscores the multifaceted regulation of BMSC-sEV on the M1/M2 balance within macrophages, thereby modulating the extent of MI/RI. Therefore, a comprehensive perspective is imperative for exploiting BMSC-sEV in regulating macrophages and ameliorating MI/RI pathology. The collective regulation of MSC-sEV on various pathological facets of MI/RI is illustrated in Figure 1.
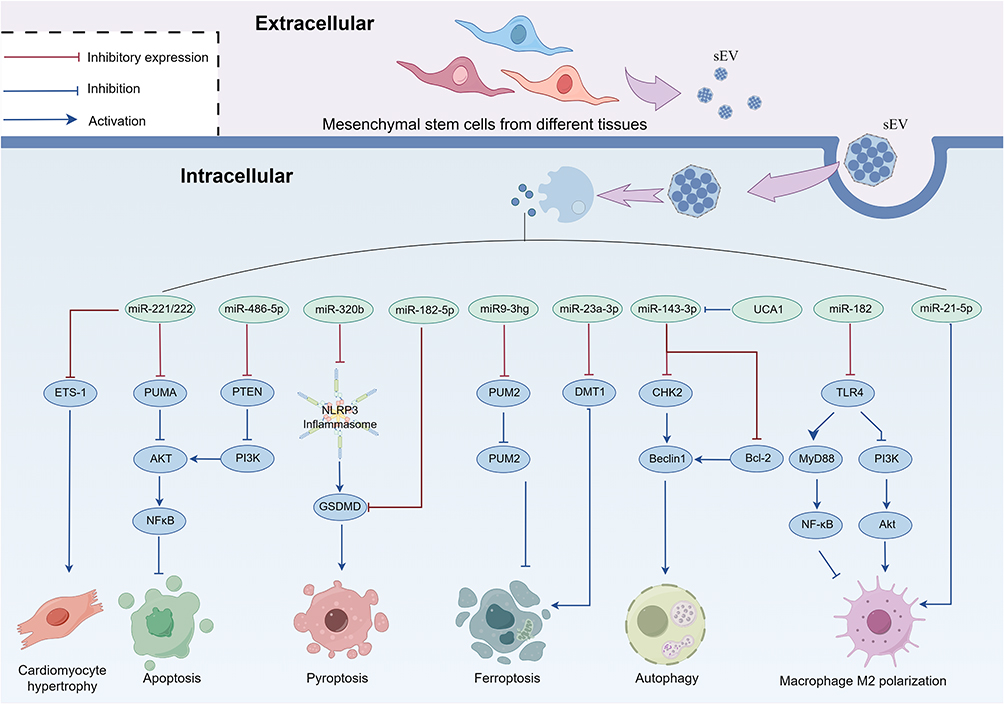 |
Figure 1 The regulation of natural MSC-sEV on multiple pathological pathways in MI/RI (By FigDraw). |
The Regulation of Nanoscale Modified MSC-sEV in MI/RI
As a therapeutic modality, sEV also encounters impediments, such as the deficiency in its targeting prowess towards specific organs and its swift clearance by mononuclear macrophages. These limitations constrict its broader application. Unaltered sEV tends to amass predominantly within the liver, kidney, spleen, and other organs subsequent to intravenous administration, a disposition unfavorable for its utilization in cardiovascular maladies.85 Concurrently, the propensity of phagocytic cells to efficiently engulf sEV surpasses that of non-phagocytic cells, thus impeding the effective delivery of sEV to afflicted target organs.86,87 Other investigations have additionally revealed that intramuscular administration of myospherogenic sEV can mitigate scar formation and enhance myocardial ejection fraction in myocardial ischemia model pigs, while intracoronary administration of the same sEV yields negligible therapeutic impact.88 This phenomenon may be attributed to the innate deficiency in homing ability exhibited by most naturally occurring sEV. Consequently, the modification of MSC-sEV to attain immune evasion, precise organ targeting, and prolonged organ residence emerges as a viable strategy to augment the efficacy of MSC-sEV in the context of MI/RI.
The surfaces of nanovesicles originating from different cells often bear membrane proteins specific to their parent cells. By merging sEV with nanovesicles or fusing nanovesicles among themselves, nanoparticles possessing multiple cellular characteristics can be acquired. Such nanoparticles are widely employed in drug delivery.89 Platelets and monocytes conspicuously accumulate within injured blood vessels and myocardial tissue subsequent to MI/RI. Platelets adhere to damaged vasculature via binding interactions with von Willebrand factor (vWF) secreted by endothelial cells, facilitated by platelet glycoprotein Ib alpha chain (GPIbα) on their surface. This adhesion is further fortified through binding between platelet surface glycoproteins GPIa/IIa and GPVI to sub-endothelial collagen, and GPIIb/IIIa to fibrin.90,91 Chemokine receptors present on monocyte surfaces become activated in response to upregulated chemokines induced by MI/RI injury, thereby altering the configuration of cell surface adhesion proteins, generating binding sites for endothelial ligands, and ultimately culminating in adhesion, migration, differentiation into macrophages, and localization within ischemic and peripheral regions.92,93 These macrophages play a pivotal role in the clearance of deceased cell debris and the facilitation of cardiac restoration. However, research has indicated that the recruitment of certain monocyte subtypes within cardiac tissue can exacerbate myocardial damage, casting uncertainty on whether monocytes foster cardiac function recovery or exacerbate injury following cardiac trauma.94,95 Nevertheless, this tendency of accumulation within damaged tissues remains a subject of considerable interest. Additionally, platelet membrane surface protein P-selectin can engage with monocyte surface protein PSGL1, while platelet membrane protein GPIb binds to Macrophage receptor 1 (MAC1), thus stabilizing intercellular adhesion. Subsequently, monocytes, as carrier cells, transport platelets into the myocardium via their own chemotactic attributes, facilitating the delivery of platelets to the injured myocardium.96 Therefore, employing nano modification to endow MSC-sEV with adhesion and chemotactic capabilities from platelets and monocytes may prove to be an efficacious approach in facilitating targeted therapy of injured tissues.
Nanovesicles modified with platelet membrane proteins (P-sEV), obtained by co-squeezing BMSC-sEV with nanoscale platelet membrane vesicles of platelet origin through fusion, has endothelium tropism reliant on platelet membrane proteins. P-sEV exhibit heightened internalization by endothelial cells distressed by hypoxia. Furthermore, P-sEV enhances endothelial cell vitality and angiogenesis capacity, and inhibiting endothelial cell apoptosis. Moreover, the potential of P-sEV to target endothelial cell injury, inhibit endothelial cell apoptosis, and stimulate angiogenesis has been corroborated in mouse models of MI/RI.97 Notably, platelet membrane proteins GPIbα and P-selectin adorn the surface of P-sEV, and these P-sEV enter injured tissues in concert with monocytes through adhesion. M1-polarized macrophages (of circulating monocyte origin) absorb P-sEV, avoiding lysosomal degradation via an endosomal escape mechanism, and subsequently release their contents into the cytoplasm of M1 macrophages. This process modulates the immune milieu within the damaged myocardium, fostering the transition of macrophages from a pro-inflammatory phenotype (M1) to an anti-inflammatory phenotype (M2).98 In addition to harnessing platelet membrane properties, the modification of MSC-sEV with monocytes membranes holds promising applications. Zhang et al employed an extrusion technique to fuse isolated MSC-sEV with monocytes membrane (Mons), resulting in monocyte-mimicking bioinspired MSC-sEV (Mon-sEV). Mon-sEV conserves the characteristics of MSC-sEV in terms of characterization and biological functionality, with monocytes membrane-derived adhesion proteins gracing its surface. Mon-sEV demonstrates a pronounced ability to chemotax endothelial cells and cardiomyocytes. Moreover, Mon-sEV exhibits significantly superior anterior ventricular wall enrichment and mitigation of myocardial injury in comparison to MSC-sEV in mouse models of myocardial MI/RI injury. This effect stems from the heightened expression of ICAM1 on the surfaces of cardiac endothelial cells and cardiomyocytes under MI/RI injury conditions, inducing homing of Mon-sEV containing adhesion proteins on their surfaces.99 These investigations validate that nano modification aimed at enhancing targeting proficiency of MSC-sEV towards damaged tissues holds considerable promise.
In addition to bolstering the capacity to target damaged tissues, the extension of MSC-sEV’s residence time within the organism constitutes a pivotal strategy to ensure in vivo efficacy.100 In vivo, sEV clearance primarily occurs via macrophage-mediated processes.87 However, certain tumor cells can robustly express the surface protein CD47, which belongs to the immunoglobulin superfamily, thereby achieving immune evasion through CD47 binding to signal-regulatory protein alpha (SIRRPα) on macrophages, releasing a “do not eat me” signal and inhibiting phagocytosis.101 Wei et al harnessed CD47 functionality to fabricate sEV (CD47-sEV) derived from CD47-overexpressing MSCs, effectively prolonging sEV retention time in vivo. Simultaneously, electrotransfection was employed to “load” miR-21 into the sEV, resulting in stable miR-21-enriched anti-phagocytic sEV (miR21-CD47-sEV).102 Further research indicates that these nanoscale modified sEV can target and downregulate the expression of PTEN in MI/RI mouse myocardial cells via miR-21. This suppression inhibits cell apoptosis, promotes the release of anti-inflammatory factors in myocardial tissue, and fosters angiogenesis, thereby ameliorating the cardiac functional damage induced by MI/RI.
Beyond the modulation of the heart’s intrinsic process of remodeling and repair, the therapeutic approach of implanting cardiac tissue engineering materials into the afflicted region through surgical or minimally invasive techniques has garnered significant attention.103,104 The biological hydrogel is a hydrophilic polymer nanostructure with high water content and rapid diffusion rates. When in its liquid state and injected into the injured area, it undergoes gelation and integrates with host tissues induced by factors such as light, chemicals, ions, temperature, and pH. This aims to provide mechanical support for the infarcted heart, reduce ventricular wall stress, compensate for contractile function, and inhibit ventricular remodeling.103,105 AT-EHBPE and HA-SH form a pair of nano-hydrogel precursors with excellent conductivity and low cytotoxicity. Zou et al designed a gel system, AT-EHBPE/HA-SH, capable of anchoring UCMSC-sEV (Gel@sEV). This composite system possesses attributes such as controllable gel kinetics, shear-thinning injectability, electrical conductivity harmonizing with native myocardium, pliability, dynamic stability congruent with heartbeat, and commendable cellular compatibility. Importantly, Gel@sEV administration, in comparison to sole UCMSC-sEV injection, significantly prolongs sEV retention time in rats.106 Furthermore, Gel@sEV exhibits enhanced efficacy in terms of mitigating myocardial fibrosis and promoting angiogenesis compared to either sEV or hydrogel administered individually. The distinct characteristics of various nanomodified MSC-sEV are showcased in Table 2.
 |
Table 2 Nanoscale Modified MSC-sEV |
MSC-sEV Pretreated with Natural Drugs
While there exists a substantial body of evidence supporting the effectiveness of natural remedies in direct disease treatment, this approach has encountered challenges such as the limited oral bioavailability of certain constituents, a low cellular uptake rate, and the instability of active ingredients over time.107–109 These hurdles constrain the standardized application and exploration of natural pharmaceutical preparations.110 A promising solution to surmount these issues involves harnessing nanocarriers, including small extracellular vesicles, for the extraction and conveyance of the active elements found in natural medicines.111
In China, the utilization of Suxiao Jiuxin pills (SJP) to ameliorate postoperative symptoms in patients suffering from acute coronary syndromes (ACS) following percutaneous coronary intervention (PCI) has garnered clinical validation.112 Optimizing the administration route of SJP could unearth additional pharmacological benefits. Ruan et al conducted an experiment in which mouse CMSCs were pretreated with SJP, and the secretion of sEV from CMSCs was assessed. The findings demonstrated that SJP could augment sEV release from CMSCs via a GTP-dependent pathway, a more pronounced effect compared to the individual treatment of CMSCs with the core constituents of SJP (tetramethylpyrazine, borneol, etc.). This underscores the harmonizing influence inherent to Chinese herbal medicine.113 In addition to augmenting sEV secretion, the intervention of natural remedies can also enhance the efficacy of sEV in the context of MI/RI. In another investigation, Ruan et al employed sEV (SJP-sEV) obtained from SJP-pretreated CMSCs to intervene in mouse cardiomyocytes. The results exhibited that SJP could inhibit the expression of Histone demethylase UTX (UTX) and elevate the levels of histone H3K27 trimethylated protein (H3K27me3) within the cells.114 Histone-dependent chromatin remodeling mediated by H3K27me3 plays an indispensable role in cell regeneration, survival, and proliferation.115 SJP-sEV stimulates cardiomyocyte proliferation under conditions of oxidative stress by modulating the UTX/H3K27me3 pathway. This research introduces a novel avenue for Traditional Chinese Medicine (TCM) treatment of MI/RI.
Oridonin, an isolated compound derived from oridonin leaves, belongs to the ent-kaurane tetracyclic diterpene class and possesses anti-inflammatory, antibacterial, and cell-regulatory properties. Nevertheless, the clinical development of Oridonin is hampered by its limited aqueous solubility and poor bioavailability.116,117 Studies have demonstrated that BMSC-sEV can upregulate the expression of autophagy-related proteins (Beclin-1, ATG13, etc.) and downregulate the expression of apoptosis-related proteins (Apaf1, Bax, etc.) in rat cardiomyocytes under MI/RI conditions, thereby enhancing cardiac function in MI/RI-afflicted rats. Oridonin-sEV, derived from BMSCs pretreated with oridonin, effectively encapsulates oridonin and exhibits more pronounced myocardial protective capabilities compared to BMSC-sEV alone.118 Irisin, a glycosyloxyisoflavone extract from iris rhizomes, not only ameliorates myocardial injury induced by ischemia but also augments the protective effects of ADMSCs in the context of ischemic heart injury by promoting myocardial homing of ADMSCs.119–121 Simultaneously, sEV can serve as carriers for transporting Irisin to injured myocardial tissue. Research indicates that irisin-sEV, obtained through irisin pretreatment of BMSCs, can inhibit pyroptosis of cardiomyocytes under conditions of H/R and protect cardiomyocyte vitality by suppressing the NLRP3 inflammasome pathway. This protective effect surpasses that achieved by BMSC-sEV intervention alone.122 Myocardial resident macrophages encompass various subtypes, with C-C chemokine receptor 2-positive macrophages (MφCCR2+) demonstrating a significant increase under MI/RI conditions. Myocardial-resident MφCCR2+ serve as inducers for monocyte homing from the circulatory system to the myocardium, thereby exacerbating myocardial inflammatory damage. Conversely, myocardial-resident MφCCR2- exhibit an inhibitory effect on inflammation.123 The expression level of miR-223-5p in tanshinone IIA (TSC)-pretreated UCMSC-sEV (TSC-sEV) was significantly elevated, and miR-223-5 targeted and suppressed the expression of CCR2. TSC-sEV mitigated the accumulation of MφCCR2+ within the myocardium through the miR-223-5/CCR2 pathway, thereby ameliorating the inflammatory milieu in the myocardial microenvironment. Moreover, TSC-sEV exhibited a stronger pro-angiogenesis effect compared to umbilical cord mesenchymal stem cell-derived sEV alone.124 Thus, the encapsulation of natural compound extracts within sEV not only facilitates drug delivery but also amplifies the efficacy of sEV, meriting further investigation.
Conclusion and Prospect
Drawing from the aforementioned investigations, it becomes evident that MSC-sEV wield a plethora of merits in the realm of mitigating MI/RI. These advantages encompass the following: (1) Given that MSC-sEV are excreted by MSCs, which are plentiful in origin and amenable to substantial in vitro cultivation, the scalable production of MSC-sEV is an attainable endeavor. (2) MSC-sEV therapy hinges upon the conveyance of signaling moieties such as proteins, RNA, and assorted molecules. Consequently, MSC-sEV therapy boasts the distinct advantage of eliciting minimal immunogenicity. (3) MSC-sEV encapsulate a diverse array of signaling molecules, engendering an expansive signal network that orchestrates MI/RI modulation, thereby imparting multi-target effects. (4) An additional facet of note pertains to the exploration of sEV freeze-drying methodologies, wherein the incorporation of a cryoprotective excipient bestows the convenience of prolonged storage and streamlined transportation.125 Concomitantly, the augmentation of MSC-sEV via the integration of membrane proteins from monocytes and platelets or the inclusion of immune evasion factors onto their membrane surface serves to significantly enhance tissue tropism within damaged areas. This augmentation extends their residence duration and safeguards them from monocyte phagocytosis. Furthermore, the prospect of coupling MSC-sEV with biomaterials for the surgical restoration of injured myocardium harbors considerable potential.
It is imperative to acknowledge that MSC-sEV therapy confronts a host of challenges: (1) Presently, researchers have proposed a plethora of divergent methodologies for MSC-sEV production and purification, a majority of which lack standardized specifications and quality control. (2) MSC-sEV assimilate disparate signal factors contingent upon their source, necessitating the meticulous classification and functional discernment of MSC-sEV as prerequisites for the standardized production and clinical deployment of these vesicles. (3) MSC-sEV offer a range of administration routes, including intravenous, intranasal, and local injection, among others. While evidence underscores the efficacy of these routes, instances dictate that intramyocardial injection is indispensable for certain MSC-sEV to ameliorate MI/RI. However, this requirement poses practical challenges within the clinical domain.
Of course, not only MSC-sEV but sEV from other sources have also been applied in research related to MI/RI. For instance, studies have reported that sEV isolated from serum of myocardial infarction patients can reduce myocardial damage area in MI/RI rat models and improve cardiac function.126 However, using sEV isolated from patient serum for MI/RI treatment is challenging due to difficulties in mass production and ethical concerns. Obtaining MSC-sEV through the cultivation of MSCs helps overcome these issues, even though research on MSC-sEV is still at an early stage. In summation, MSC-sEV therapy is a strategy teeming with advantages for MI/RI treatment, yet it contends with an array of challenges. Therefore, we aspire to see more comparative studies, standardization of the MSC-sEV preparation process, and clarification of the administration routes for MSC-sEV. Concurrently, conducting clinical research will facilitate the early application of MSC-sEV in the clinical treatment of MI/RI.
Abbreviations
MI/RI, Myocardial ischemia-reperfusion injury; sEV, small Extracellular vesicles; MSCs, Mesenchymal stem cells; CABG, Coronary artery bypass grafting; PMVs, Platelet membrane vesicles; Mons, Mononuclear cell membrane; SJP, Suxiao Jiuxin pills; ACS, Acute coronary syndromes; PCI, Percutaneous coronary intervention; TCM, Traditional Chinese Medicine.
Acknowledgments
Thanks to FigDraw (www.figdraw.com) for giving the drawing support.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the Innovation Team and Talents Cultivation Program of National Administration of Traditional Chinese Medicine (No. ZYYCXTD-C-202203) and the National Natural Science Foundation of China (No.82174326).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Collet JP, Thiele H, Barbato E, et al. 2020 ESC Guidelines for the management of acute coronary syndromes in patients presenting without persistent ST-segment elevation. Eur Heart J. 2021;42(14):1289–1367. doi:10.1093/eurheartj/ehaa575
2. Ibanez B, James S, Agewall S, et al. 2017 ESC Guidelines for the management of acute myocardial infarction in patients presenting with ST-segment elevation: the Task Force for the management of acute myocardial infarction in patients presenting with ST-segment elevation of the European Society of Cardiology (ESC). Eur Heart J. 2018;39(2):119–177. doi:10.1093/eurheartj/ehx393
3. Vaduganathan M, Mensah GA, Turco JV, et al. The global burden of cardiovascular diseases and risk: a compass for future health. J Am Coll Cardiol. 2022;80(25):2361–2371. doi:10.1016/j.jacc.2022.11.005
4. Heusch G, Gersh BJ. The pathophysiology of acute myocardial infarction and strategies of protection beyond reperfusion: a continual challenge. Eur Heart J. 2017;38(11):774–784. doi:10.1093/eurheartj/ehw224
5. Schäfer A, König T, Bauersachs J, et al. Novel therapeutic strategies to reduce reperfusion injury after acute myocardial infarction. Curr Probl Cardiol. 2022;47(12):101398. doi:10.1016/j.cpcardiol.2022.101398
6. Wang J, Liu Y, Liu Y, et al. Recent advances in nanomedicines for imaging and therapy of myocardial ischemia-reperfusion injury. J Control Release. 2023;353:563–590. doi:10.1016/j.jconrel.2022.11.057
7. Heusch G. Myocardial stunning and hibernation revisited. Nat Rev Cardiol. 2021;18(7):522–536. doi:10.1038/s41569-021-00506-7
8. Jiang Y, Sun S-J, Zhen Z, et al. Myocardial repair of bioengineered cardiac patches with decellularized placental scaffold and human-induced pluripotent stem cells in a rat model of myocardial infarction. Stem Cell Res Ther. 2021;12(1):13. doi:10.1186/s13287-020-02066-y
9. Thavapalachandran S, Le TYL, Romanazzo S, et al. Pluripotent stem cell-derived mesenchymal stromal cells improve cardiac function and vascularity after myocardial infarction. Cytotherapy. 2021;23(12):1074–1084. doi:10.1016/j.jcyt.2021.07.016
10. Lutter G, Puehler T, Cyganek L, et al. Biodegradable Poly-ε-caprolactone scaffolds with ECFCs and iMSCs for tissue-engineered heart valves. Int J Mol Sci. 2022;23(1):527. doi:10.3390/ijms23010527
11. Naji A, Eitoku M, Favier B, et al. Biological functions of mesenchymal stem cells and clinical implications. Cell Mol Life Sci. 2019;76(17):3323–3348. doi:10.1007/s00018-019-03125-1
12. Bolli R, Mitrani RD, Hare JM, et al. A Phase II study of autologous mesenchymal stromal cells and c-kit positive cardiac cells, alone or in combination, in patients with ischaemic heart failure: the CCTRN CONCERT-HF trial. Eur J Heart Fail. 2021;23(4):661–674. doi:10.1002/ejhf.2178
13. Bartolucci J, Verdugo FJ, González PL, et al. Safety and efficacy of the intravenous infusion of umbilical cord mesenchymal stem cells in patients with heart failure: a Phase 1/2 randomized controlled trial (RIMECARD Trial [Randomized Clinical Trial of Intravenous Infusion Umbilical Cord Mesenchymal Stem Cells on Cardiopathy]). Circ Res. 2017;121(10):1192–1204. doi:10.1161/CIRCRESAHA.117.310712
14. Can A, Ulus AT, Cinar O, et al. Human umbilical cord mesenchymal stromal cell transplantation in myocardial ischemia (HUC-HEART Trial). A study protocol of a Phase 1/2, controlled and randomized trial in combination with coronary artery bypass grafting. Stem Cell Rev Rep. 2015;11(5):752–760. doi:10.1007/s12015-015-9601-0
15. Ulus AT, Mungan C, Kurtoglu M, et al. Intramyocardial transplantation of umbilical cord mesenchymal stromal cells in chronic ischemic cardiomyopathy: a controlled, randomized clinical trial (HUC-HEART Trial). Int J Stem Cells. 2020;13(3):364–376. doi:10.15283/ijsc20075
16. Li M, Jiang Y, Hou Q, et al. Potential pre-activation strategies for improving therapeutic efficacy of mesenchymal stem cells: current status and future prospects. Stem Cell Res Ther. 2022;13(1):146. doi:10.1186/s13287-022-02822-2
17. Phermthai T, Thongbopit S, Pokathikorn P, et al. Carcinogenicity, efficiency and biosafety analysis in xeno-free human amniotic stem cells for regenerative medical therapies. Cytotherapy. 2017;19(8):990–1001. doi:10.1016/j.jcyt.2017.04.004
18. Lee JY, Kang MH, Jang JE, et al. Comparative analysis of mesenchymal stem cells cultivated in serum free media. Sci Rep. 2022;12:8620. doi:10.1038/s41598-022-12467-z
19. Wei F, Wang T-Z, Zhang J, et al. Mesenchymal stem cells neither fully acquire the electrophysiological properties of mature cardiomyocytes nor promote ventricular arrhythmias in infarcted rats. Basic Res Cardiol. 2012;107(4):274. doi:10.1007/s00395-012-0274-4
20. Yáñez-Mó M, Siljander PR, Andreu Z, et al. Biological properties of extracellular vesicles and their physiological functions. J Extracell Vesicles. 2015;4:27066. doi:10.3402/jev.v4.27066
21. Xiao C, Wang K, Xu Y, et al. Transplanted mesenchymal stem cells reduce autophagic flux in infarcted hearts via the exosomal transfer of miR-125b. Circ Res. 2018;123(5):564–578. doi:10.1161/CIRCRESAHA.118.312758
22. Nasser MI, Masood M, Adlat S, et al. Mesenchymal stem cell-derived exosome microRNA as therapy for cardiac ischemic injury. Biomed Pharmacother. 2021;143:112118. doi:10.1016/j.biopha.2021.112118
23. Sun Y, Liu G, Zhang K, et al. Mesenchymal stem cells-derived exosomes for drug delivery. Stem Cell Res Ther. 2021;12(1):561. doi:10.1186/s13287-021-02629-7
24. Liu Y, Li L, Wang Z, et al. Myocardial ischemia-reperfusion injury; Molecular mechanisms and prevention. Microvasc Res. 2023;149:104565. doi:10.1016/j.mvr.2023.104565
25. Ma Q, Ma Y, Wang X, et al. Intracoronary compared with intravenous bolus tirofiban on the microvascular obstruction in patients with STEMI undergoing PCI: a cardiac MR study. Int J Cardiovasc Imaging. 2020;36(6):1121–1132. doi:10.1007/s10554-020-01800-0
26. Schulz S, Richardt G, Laugwitz KL, et al. Prasugrel plus bivalirudin vs. clopidogrel plus heparin in patients with ST-segment elevation myocardial infarction. Eur Heart J. 2014;35(34):2285–2294. doi:10.1093/eurheartj/ehu182
27. Qian G, Zhang Y, Dong W, et al. Effects of nicorandil administration on infarct size in patients with ST-segment-elevation myocardial infarction undergoing primary percutaneous coronary intervention: the CHANGE trial. J Am Heart Assoc. 2022;11(18):e026232. doi:10.1161/JAHA.122.026232
28. Li X, Zhang S, Wang Z, et al. Platelet function and risk of bleeding in patients with acute coronary syndrome following tirofiban infusion. Front Pharmacol. 2019;10:1158. doi:10.3389/fphar.2019.01158
29. Wang Y, Shen Y. Exosomal miR-455-3p from BMMSCs prevents cardiac ischemia-reperfusion injury. Hum Exp Toxicol. 2022;41:9603271221102508. doi:10.1177/09603271221102508
30. Zhang L, Wei Q, Liu X, et al. Exosomal microRNA-98-5p from hypoxic bone marrow mesenchymal stem cells inhibits myocardial ischemia-reperfusion injury by reducing TLR4 and activating the PI3K/Akt signaling pathway. Int Immunopharmacol. 2021;101:107592. doi:10.1016/j.intimp.2021.107592
31. Nieuwland R, Falcón-Pérez JM, Théry C, et al. Rigor and standardization of extracellular vesicle research: paving the road towards robustness. J Extracell Vesicles. 2020;10(2):e12037. doi:10.1002/jev2.12037
32. Witwer KW, Goberdhan DC, O’Driscoll L, et al. Updating MISEV: evolving the minimal requirements for studies of extracellular vesicles. J Extracell Vesicles. 2021;10(14):e12182. doi:10.1002/jev2.12182
33. Chu DT, Phuong TNT, Tien NLB, et al. An update on the progress of isolation, culture, storage, and clinical application of human bone marrow mesenchymal stem/stromal cells. Int J Mol Sci. 2020;21(3):708. doi:10.3390/ijms21030708
34. Al-Ghadban S, Bunnell BA. Adipose tissue-derived stem cells: immunomodulatory effects and therapeutic potential. Physiology. 2020;35(2):125–133. doi:10.1152/physiol.00021.2019
35. Ma H, Jiang S, Du L, et al. Conditioned medium from primary cytotrophoblasts, primary placenta-derived mesenchymal stem cells, or sub-cultured placental tissue promoted HUVEC angiogenesis in vitro. Stem Cell Res Ther. 2021;12(1):141. doi:10.1186/s13287-021-02192-1
36. Semenova E, Grudniak MP, Machaj EK, et al. Mesenchymal stromal cells from different parts of umbilical cord: approach to comparison & characteristics. Stem Cell Rev Rep. 2021;17(5):1780–1795. doi:10.1007/s12015-021-10157-3
37. Suzuki S, Mizuno M, Sakamaki Y, et al. Morphological changes in synovial mesenchymal stem cells during their adhesion to the meniscus. Lab Invest. 2020;100(7):916–927. doi:10.1038/s41374-020-0421-8
38. Masuda K, Han X, Kato H, et al. Dental pulp-derived mesenchymal stem cells for modeling genetic disorders. Int J Mol Sci. 2021;22(5):2269. doi:10.3390/ijms22052269
39. Margiana R, Markov A, Zekiy AO, et al. Clinical application of mesenchymal stem cell in regenerative medicine: a narrative review. Stem Cell Res Ther. 2022;13(1):366. doi:10.1186/s13287-022-03054-0
40. Xu J, Liao W, Gu D, et al. Neural ganglioside GD2 identifies a subpopulation of mesenchymal stem cells in umbilical cord. Cell Physiol Biochem. 2009;23(4–6):415–424. doi:10.1159/000218188
41. Pittenger MF, Mackay AM, Beck SC, et al. Multilineage potential of adult human mesenchymal stem cells. Science. 1999;284(5411):143–147. doi:10.1126/science.284.5411.143
42. De Ugarte DA, Alfonso Z, Zuk PA, et al. Differential expression of stem cell mobilization-associated molecules on multi-lineage cells from adipose tissue and bone marrow. Immunol Lett. 2003;89(2–3):267–270. doi:10.1016/S0165-2478(03)00108-1
43. Zuk PA, Zhu M, Ashjian P, et al. Human adipose tissue is a source of multipotent stem cells. Mol Biol Cell. 2002;13(12):4279–4295. doi:10.1091/mbc.e02-02-0105
44. Schäffler A, Büchler C. Concise review: adipose tissue-derived stromal cells--basic and clinical implications for novel cell-based therapies. Stem Cells. 2007;25(4):818–827. doi:10.1634/stemcells.2006-0589
45. Zhao L, Wang J, Wang P, et al. Oct4 cooperates with c-Myc to improve mesenchymal-to-endothelial transition and myocardial repair of cardiac-resident mesenchymal stem cells. Stem Cell Res Ther. 2022;13(1):445. doi:10.1186/s13287-022-03120-7
46. Álvarez-Viejo M, Menéndez-Menéndez Y, Otero-Hernández J. CD271 as a marker to identify mesenchymal stem cells from diverse sources before culture. World J Stem Cells. 2015;7(2):470–476. doi:10.4252/wjsc.v7.i2.470
47. Kacham S, Bhure TS, Eswaramoorthy SD, et al. Human umbilical cord-derived mesenchymal stem cells promote corneal epithelial repair in vitro. Cells. 2021;10(5):1254. doi:10.3390/cells10051254
48. Du WJ, Chi Y, Yang ZX, et al. Heterogeneity of proangiogenic features in mesenchymal stem cells derived from bone marrow, adipose tissue, umbilical cord, and placenta. Stem Cell Res Ther. 2016;7(1):163. doi:10.1186/s13287-016-0418-9
49. Xu L, Liu Y, Sun Y, et al. Tissue source determines the differentiation potentials of mesenchymal stem cells: a comparative study of human mesenchymal stem cells from bone marrow and adipose tissue. Stem Cell Res Ther. 2017;8(1):275. doi:10.1186/s13287-017-0716-x
50. Choudhery MS, Badowski M, Muise A, et al. Comparison of human mesenchymal stem cells derived from adipose and cord tissue. Cytotherapy. 2013;15(3):330–343. doi:10.1016/j.jcyt.2012.11.010
51. Li CY, Wu XY, Tong JB, et al. Comparative analysis of human mesenchymal stem cells from bone marrow and adipose tissue under xeno-free conditions for cell therapy. Stem Cell Res Ther. 2015;6:55.
52. Chen Y, Shen H, Ding Y, et al. The application of umbilical cord-derived MSCs in cardiovascular diseases. J Cell Mol Med. 2021;25(17):8103–8114. doi:10.1111/jcmm.16830
53. Beane OS, Fonseca VC, Cooper LL, et al. Impact of aging on the regenerative properties of bone marrow-, muscle-, and adipose-derived mesenchymal stem/stromal cells. PLoS One. 2014;9(12):e115963. doi:10.1371/journal.pone.0115963
54. Sattayaprasert P, Vasireddi SK, Bektik E, et al. Human cardiac mesenchymal stem cells remodel in disease and can regulate arrhythmia substrates. Circ Arrhythm Electrophysiol. 2020;13(10):e008740. doi:10.1161/CIRCEP.120.008740
55. Bei Y, Lu D, Bär C, et al. miR-486 attenuates cardiac ischemia/reperfusion injury and mediates the beneficial effect of exercise for myocardial protection. Mol Ther. 2022;30(4):1675–1691. doi:10.1016/j.ymthe.2022.01.031
56. Wang Z, Hao W, Hu J, et al. Maltol improves APAP-induced hepatotoxicity by inhibiting oxidative stress and inflammation response via NF-κB and PI3K/Akt signal pathways. Antioxidants. 2019;8(9):395. doi:10.3390/antiox8090395
57. Tu Y, Wu W, Guo Y, et al. Up-regulation of hsa-miR-221-3p induced by UVB affects proliferation and apoptosis of keratinocytes via Bcl-xL/Bax pathway. Photodermatol Photoimmunol Photomed. 2021;37(4):269–277. doi:10.1111/phpp.12647
58. Lai TC, Lee TL, Chang YC, et al. MicroRNA-221/222 mediates ADSC-exosome-induced cardioprotection against ischemia/reperfusion by targeting PUMA and ETS-1. Front Cell Dev Biol. 2020;8:569150. doi:10.3389/fcell.2020.569150
59. Nakamura M, Sadoshima J. Mechanisms of physiological and pathological cardiac hypertrophy. Nat Rev Cardiol. 2018;15(7):387–407. doi:10.1038/s41569-018-0007-y
60. Ni W, Zhan Y, He H, et al. Ets-1 Is a Critical Transcriptional Regulator of Reactive Oxygen Species and p47(phox) gene expression in response to angiotensin II. Circ Res. 2007;101(10):985–994. doi:10.1161/CIRCRESAHA.107.152439
61. Sun XH, Wang X, Zhang Y, et al. Exosomes of bone-marrow stromal cells inhibit cardiomyocyte apoptosis under ischemic and hypoxic conditions via miR-486-5p targeting the PTEN/PI3K/AKT signaling pathway. Thromb Res. 2019;177:23–32. doi:10.1016/j.thromres.2019.02.002
62. Huang Y, Xu W, Zhou R. NLRP3 inflammasome activation and cell death. Cell Mol Immunol. 2021;18(9):2114–2127. doi:10.1038/s41423-021-00740-6
63. Tang J, Jin L, Liu Y, et al. Exosomes derived from mesenchymal stem cells protect the myocardium against ischemia/reperfusion injury through inhibiting pyroptosis. Drug Des Devel Ther. 2020;14:3765–3775. doi:10.2147/DDDT.S239546
64. Shi H, Gao Y, Dong Z, et al. GSDMD-mediated cardiomyocyte pyroptosis promotes myocardial I/R injury. Circ Res. 2021;129(3):383–396. doi:10.1161/CIRCRESAHA.120.318629
65. Yue R, Lu S, Luo Y, et al. Mesenchymal stem cell-derived exosomal microRNA-182-5p alleviates myocardial ischemia/reperfusion injury by targeting GSDMD in mice. Cell Death Discov. 2022;8(1):202. doi:10.1038/s41420-022-00909-6
66. Jiang X, Stockwell BR, Conrad M. Ferroptosis: mechanisms, biology and role in disease. Nat Rev Mol Cell Biol. 2021;22(4):266–282. doi:10.1038/s41580-020-00324-8
67. Ursini F, Maiorino M. Lipid peroxidation and ferroptosis: the role of GSH and GPx4. Free Radic Biol Med. 2020;152:175–185. doi:10.1016/j.freeradbiomed.2020.02.027
68. Zhao WK, Zhou Y, Xu TT, et al. Ferroptosis: opportunities and challenges in myocardial ischemia-reperfusion injury. Oxid Med Cell Longev. 2021;2021:9929687. doi:10.1155/2021/9929687
69. Zhang JK, Zhang Z, Guo ZA, et al. The BMSC-derived exosomal lncRNA Mir9-3hg suppresses cardiomyocyte ferroptosis in ischemia-reperfusion mice via the Pum2/PRDX6 axis. Nutr, Metab Cardiovasc Dis. 2022;32(2):515–527. doi:10.1016/j.numecd.2021.10.017
70. Lu B, Chen XB, Hong YC, et al. Identification of PRDX6 as a regulator of ferroptosis. Acta Pharmacol Sin. 2019;40(10):1334–1342. doi:10.1038/s41401-019-0233-9
71. Song Y, Wang B, Zhu X, et al. Human umbilical cord blood-derived MSCs exosome attenuate myocardial injury by inhibiting ferroptosis in acute myocardial infarction mice. Cell Biol Toxicol. 2021;37(1):51–64. doi:10.1007/s10565-020-09530-8
72. Klionsky DJ, Petroni G, Amaravadi RK, et al. Autophagy in major human diseases. EMBO J. 2021;40(19):e108863. doi:10.15252/embj.2021108863
73. Yan Y, Finkel T. Autophagy as a regulator of cardiovascular redox homeostasis. Free Radic Biol Med. 2017;109:108–113. doi:10.1016/j.freeradbiomed.2016.12.003
74. Azad MB, Chen Y, Henson ES, et al. Hypoxia induces autophagic cell death in apoptosis-competent cells through a mechanism involving BNIP3. Autophagy. 2008;4(2):195–204. doi:10.4161/auto.5278
75. Galluzzi L, Vitale I, Aaronson SA, et al. Molecular mechanisms of cell death: recommendations of the nomenclature committee on cell death 2018. Cell Death Differ. 2018;25(3):486–541. doi:10.1038/s41418-017-0012-4
76. Chen G, Wang M, Ruan Z, et al. Mesenchymal stem cell-derived exosomal miR-143-3p suppresses myocardial ischemia-reperfusion injury by regulating autophagy. Life Sci. 2021;280:119742. doi:10.1016/j.lfs.2021.119742
77. Guo QQ, Wang SS, Zhang SS, et al. ATM-CHK2-Beclin 1 axis promotes autophagy to maintain ROS homeostasis under oxidative stress. EMBO J. 2020;39(10):e103111. doi:10.15252/embj.2019103111
78. Ma L, Zou R, Shi W, et al. SGLT2 inhibitor dapagliflozin reduces endothelial dysfunction and microvascular damage during cardiac ischemia/reperfusion injury through normalizing the XO-SERCA2-CaMKII-coffilin pathways. Theranostics. 2022;12(11):5034–5050. doi:10.7150/thno.75121
79. Diao L, Zhang Q. Transfer of lncRNA UCA1 by hUCMSCs-derived exosomes protects against hypoxia/reoxygenation injury through impairing miR-143-targeted degradation of Bcl-2. Aging. 2021;13(4):5967–5985. doi:10.18632/aging.202520
80. Frangogiannis NG, Rosenzweig A. Regulation of the inflammatory response in cardiac repair. Circ Res. 2012;110(1):159–173. doi:10.1161/CIRCRESAHA.111.243162
81. Lafuse WP, Wozniak DJ, Rajaram MVS. Role of cardiac macrophages on cardiac inflammation, fibrosis and tissue repair. Cells. 2020;10(1):51. doi:10.3390/cells10010051
82. Ong SB, Hernández-Reséndiz S, Crespo-Avilan GE, et al. Inflammation following acute myocardial infarction: multiple players, dynamic roles, and novel therapeutic opportunities. Pharmacol Ther. 2018;186:73–87. doi:10.1016/j.pharmthera.2018.01.001
83. Shen D, He Z. Mesenchymal stem cell-derived exosomes regulate the polarization and inflammatory response of macrophages via miR-21-5p to promote repair after myocardial reperfusion injury. Ann Transl Med. 2021;9(16):1323. doi:10.21037/atm-21-3557
84. Zhao J, Li X, Hu J, et al. Mesenchymal stromal cell-derived exosomes attenuate myocardial ischaemia-reperfusion injury through miR-182-regulated macrophage polarization. Cardiovasc Res. 2019;115(7):1205–1216. doi:10.1093/cvr/cvz040
85. Morishita M, Takahashi Y, Nishikawa M, et al. Quantitative analysis of tissue distribution of the B16BL6-derived exosomes using a streptavidin-lactadherin fusion protein and iodine-125-labeled biotin derivative after intravenous injection in mice. J Pharm Sci. 2015;104(2):705–713. doi:10.1002/jps.24251
86. Imai T, Takahashi Y, Nishikawa M, et al. Macrophage-dependent clearance of systemically administered B16BL6-derived exosomes from the blood circulation in mice. J Extracell Vesicles. 2015;4(1):26238. doi:10.3402/jev.v4.26238
87. Matsumoto A, Takahashi Y, Chang H-Y, et al. Blood concentrations of small extracellular vesicles are determined by a balance between abundant secretion and rapid clearance. J Extracell Vesicles. 2020;9(1):1696517. doi:10.1080/20013078.2019.1696517
88. Gallet R, Dawkins J, Valle J, et al. Exosomes secreted by cardiosphere-derived cells reduce scarring, attenuate adverse remodelling, and improve function in acute and chronic porcine myocardial infarction. Eur Heart J. 2017;38(3):201–211. doi:10.1093/eurheartj/ehw240
89. Dehaini D, Wei X, Fang RH, et al. Erythrocyte-platelet hybrid membrane coating for enhanced nanoparticle functionalization. Adv Mater. 2017;29(16). doi:10.1002/adma.201606209
90. Springer TA. von Willebrand factor, Jedi knight of the bloodstream. Blood. 2014;124(9):1412–1425. doi:10.1182/blood-2014-05-378638
91. Lu Y, Hu Q, Jiang C, et al. Platelet for drug delivery. Curr Opin Biotechnol. 2019;58:81–91. doi:10.1016/j.copbio.2018.11.010
92. Wrigley BJ, Lip GY, Shantsila E. The role of monocytes and inflammation in the pathophysiology of heart failure. Eur J Heart Fail. 2011;13(11):1161–1171. doi:10.1093/eurjhf/hfr122
93. Imhof BA, Aurrand-Lions M. Adhesion mechanisms regulating the migration of monocytes. Nat Rev Immunol. 2004;4(6):432–444. doi:10.1038/nri1375
94. Nahrendorf M, Swirski FK, Aikawa E, et al. The healing myocardium sequentially mobilizes two monocyte subsets with divergent and complementary functions. J Exp Med. 2007;204(12):3037–3047. doi:10.1084/jem.20070885
95. Zhang J, Hao W, Zhang J, et al. CXCL16 promotes Ly6Chigh monocyte infiltration and impairs heart function after acute myocardial infarction. J Immunol. 2023;210(6):820–831. doi:10.4049/jimmunol.2200249
96. Ziegler M, Wang X, Peter K. Platelets in cardiac ischaemia/reperfusion injury: a promising therapeutic target. Cardiovasc Res. 2019;115(7):1178–1188. doi:10.1093/cvr/cvz070
97. Li Q, Song Y, Wang Q, et al. Engineering extracellular vesicles with platelet membranes fusion enhanced targeted therapeutic angiogenesis in a mouse model of myocardial ischemia reperfusion. Theranostics. 2021;11(8):3916–3931. doi:10.7150/thno.52496
98. Li Q, Huang Z, Wang Q, et al. Targeted immunomodulation therapy for cardiac repair by platelet membrane engineering extracellular vesicles via hitching peripheral monocytes. Biomaterials. 2022;284:121529. doi:10.1016/j.biomaterials.2022.121529
99. Zhang N, Song Y, Huang Z, et al. Monocyte mimics improve mesenchymal stem cell-derived extracellular vesicle homing in a mouse MI/RI model. Biomaterials. 2020;255:120168. doi:10.1016/j.biomaterials.2020.120168
100. van der Meel R, Fens MH, Vader P, et al. Extracellular vesicles as drug delivery systems: lessons from the liposome field. J Control Release. 2014;195:72–85. doi:10.1016/j.jconrel.2014.07.049
101. Chao MP, Weissman IL, Majeti R. The CD47-SIRPα pathway in cancer immune evasion and potential therapeutic implications. Curr Opin Immunol. 2012;24(2):225–232. doi:10.1016/j.coi.2012.01.010
102. Wei Z, Chen Z, Zhao Y, et al. Mononuclear phagocyte system blockade using extracellular vesicles modified with CD47 on membrane surface for myocardial infarction reperfusion injury treatment. Biomaterials. 2021;275:121000. doi:10.1016/j.biomaterials.2021.121000
103. Radhakrishnan J, Krishnan UM, Sethuraman S. Hydrogel based injectable scaffolds for cardiac tissue regeneration. Biotechnol Adv. 2014;32(2):449–461. doi:10.1016/j.biotechadv.2013.12.010
104. Hao T, Qian M, Zhang Y, et al. An injectable dual-function hydrogel protects against myocardial ischemia/reperfusion injury by modulating ROS/NO disequilibrium. Adv Sci. 2022;9(15):e2105408. doi:10.1002/advs.202105408
105. Rao SV, Zeymer U, Douglas PS, et al. Bioabsorbable intracoronary matrix for prevention of ventricular remodeling after myocardial infarction. J Am Coll Cardiol. 2016;68(7):715–723. doi:10.1016/j.jacc.2016.05.053
106. Zou Y, Li L, Li Y, et al. Restoring cardiac functions after myocardial infarction-ischemia/reperfusion via an exosome anchoring conductive hydrogel. ACS Appl Mater Interfaces. 2021;13(48):56892–56908. doi:10.1021/acsami.1c16481
107. Li X, Zhang J, Huang J, et al. A multicenter, randomized, double-blind, parallel-group, placebo-controlled study of the effects of qili qiangxin capsules in patients with chronic heart failure. J Am Coll Cardiol. 2013;62(12):1065–1072. doi:10.1016/j.jacc.2013.05.035
108. Bai R, Yang Q, Xi R, et al. The effectiveness and safety of Chinese Patent Medicines based on syndrome differentiation in patients following percutaneous coronary intervention due to acute coronary syndrome (CPM trial): a nationwide Cohort Study. Phytomedicine. 2023;109:154554. doi:10.1016/j.phymed.2022.154554
109. Mao S, Taylor S, Chen Q, et al. Sodium tanshinone IIA sulfonate prevents the adverse left ventricular remodelling: focus on polymorphonuclear neutrophil-derived granule components. J Cell Mol Med. 2019;23(7):4592–4600. doi:10.1111/jcmm.14306
110. Li C, Jia WW, Yang JL, et al. Multi-compound and drug-combination pharmacokinetic research on Chinese herbal medicines. Acta Pharmacol Sin. 2022;43(12):3080–3095. doi:10.1038/s41401-022-00983-7
111. Wei D, Yang H, Zhang Y, et al. Nano-traditional Chinese medicine: a promising strategy and its recent advances. J Mater Chem B. 2022;10(16):2973–2994. doi:10.1039/D2TB00225F
112. Shen Z, Chen T, Deng B, et al. Effects on suxiao jiuxin pills in the treatment of patients with acute coronary syndrome undergoing early percutaneous coronary intervention: a multicenter randomized double-blind placebo-controlled trial. J Altern Complement Med. 2020;26(11):1055–1063. doi:10.1089/acm.2020.0014
113. Ruan XF, Ju CW, Shen Y, et al. Suxiao Jiuxin pill promotes exosome secretion from mouse cardiac mesenchymal stem cells in vitro. Acta Pharmacol Sin. 2018;39(4):569–578. doi:10.1038/aps.2018.19
114. Ruan XF, Li YJ, Ju CW, et al. Exosomes from Suxiao Jiuxin pill-treated cardiac mesenchymal stem cells decrease H3K27 demethylase UTX expression in mouse cardiomyocytes in vitro. Acta Pharmacol Sin. 2018;39(4):579–586. doi:10.1038/aps.2018.18
115. Ai S, Yu X, Li Y, et al. Divergent requirements for EZH1 in heart development versus regeneration. Circ Res. 2017;121(2):106–112. doi:10.1161/CIRCRESAHA.117.311212
116. Li D, Han T, Liao J, et al. Oridonin, a promising ent-kaurane diterpenoid lead compound. Int J Mol Sci. 2016;17(9):1395. doi:10.3390/ijms17091395
117. Cheng W, Huang C, Ma W, et al. Recent development of oridonin derivatives with diverse pharmacological activities. Mini Rev Med Chem. 2019;19(2):114–124. doi:10.2174/1389557517666170417170609
118. Fu M, Xie D, Sun Y, et al. Exosomes derived from MSC pre-treated with oridonin alleviates myocardial IR injury by suppressing apoptosis via regulating autophagy activation. J Cell Mol Med. 2021;25(12):5486–5496. doi:10.1111/jcmm.16558
119. Ibrahim SR, Mohamed GA, Al-Musayeib NM. New constituents from the rhizomes of Egyptian Iris germanica L. Molecules. 2012;17(3):2587–2598. doi:10.3390/molecules17032587
120. Zhao YT, Wang J, Yano N, et al. Irisin promotes cardiac progenitor cell-induced myocardial repair and functional improvement in infarcted heart. J Cell Physiol. 2019;234(2):1671–1681. doi:10.1002/jcp.27037
121. Yan W, Chen Y, Guo Y, et al. Irisin promotes cardiac homing of intravenously delivered MSCs and protects against ischemic heart injury. Adv Sci. 2022;9(7):e2103697. doi:10.1002/advs.202103697
122. Deng J, Zhang T, Li M, et al. Irisin-pretreated BMMSCs secrete exosomes to alleviate cardiomyocytes pyroptosis and oxidative stress to hypoxia/reoxygenation injury. Curr Stem Cell Res Ther. 2023;18(6):843–852. doi:10.2174/1574888X18666221117111829
123. Bajpai G, Bredemeyer A, Li W, et al. Tissue resident CCR2- and CCR2+ cardiac macrophages differentially orchestrate monocyte recruitment and fate specification following myocardial injury. Circ Res. 2019;124(2):263–278. doi:10.1161/CIRCRESAHA.118.314028
124. Li S, Yang K, Cao W, et al. Tanshinone IIA enhances the therapeutic efficacy of mesenchymal stem cells derived exosomes in myocardial ischemia/reperfusion injury via up-regulating miR-223-5p. J Control Release. 2023;358:13–26. doi:10.1016/j.jconrel.2023.04.014
125. Susa F, Limongi T, Borgione F, et al. Comparative studies of different preservation methods and relative freeze-drying formulations for extracellular vesicle pharmaceutical applications. ACS Biomater Sci Eng. 2023;9(10):5871–5885. doi:10.1021/acsbiomaterials.3c00678
126. D’Ascenzo F, Femminò S, Ravera F, et al. Extracellular vesicles from patients with Acute Coronary Syndrome impact on ischemia-reperfusion injury. Pharmacol Res. 2021;170:105715. doi:10.1016/j.phrs.2021.105715
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.