Back to Journals » Infection and Drug Resistance » Volume 13
A Multidrug-resistant Monophasic Salmonella Typhimurium Co-harboring mcr-1, fosA3, blaCTX-M-14 in a Transferable IncHI2 Plasmid from a Healthy Catering Worker in China
Authors Wang Z, Xu H, Tang Y, Li Q ![]() , Jiao X
, Jiao X ![]()
Received 15 July 2020
Accepted for publication 29 August 2020
Published 13 October 2020 Volume 2020:13 Pages 3569—3574
DOI https://doi.org/10.2147/IDR.S272272
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Suresh Antony
Zhenyu Wang,1– 3,* Haiyan Xu4 ,* Yuanyue Tang,1– 3 Qiuchun Li,1– 3 Xinan Jiao1– 3
1Key Laboratory of Prevention and Control of Biological Hazard Factors (Animal Origin) for Agri-Food Safety and Quality, Ministry of Agriculture of China, Yangzhou University, Yangzhou, People’s Republic of China; 2Jiangsu Key Laboratory of Zoonosis/Jiangsu Co-Innovation Center for Prevention and Control of Important Animal Infectious Diseases and Zoonoses, Yangzhou University, Yangzhou, People’s Republic of China; 3Joint International Research Laboratory of Agriculture and Agri-Product Safety, Yangzhou University, Yangzhou, People’s Republic of China; 4Nantong Center for Disease Control and Prevention, Nantong, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Qiuchun Li; Xinan Jiao
Jiangsu Key Laboratory of Zoonosis, Yangzhou University, Yangzhou, Jiangsu Province, People’s Republic of China
Tel +86-514-87971136
Email [email protected]; [email protected]
Background: Polymyxins are currently regarded as a possible last-resort therapy to eradicate multidrug-resistant (MDR) gram-negative bacteria. Meanwhile, the old antimicrobial agent fosfomycin has recently been reintroduced into clinical use for the treatment of extended-spectrum β-lactamase (ESBL)-producing and carbapenem-resistant Enterobacteriaceae. This study investigated a multidrug-resistant Salmonella 4,[5],12:i:- strain from a food catering handler, which had the potential to act as a vehicle for transmitting MDR foodborne pathogens.
Methods: A Salmonella 4,[5],12:i:- YZU1189 strain was isolated from the fecal sample of a food catering worker according to the standard protocol of the Salmonella detection method from World Health Organization in 2003. Serotyping of YZU1189 was performed according to the Kauffmann–White scheme. The antimicrobial resistance phenotype of the strain was determined by the agar dilution method according to the instruction from Clinical and Laboratory Standards Institute (CLSI). Plasmid conjugation was performed between the donor strain Salmonella 4,[5],12:i:- YZU1189 and the recipient strain Escherichia coli C600. The genetic locations of mcr-1, blaCTX-M-14 and fosA3 genes were determined by the whole genome sequence analysis.
Results: Salmonella 4,[5],12:i:- YZU1189 was an ESBL-producing stain isolated from a healthy catering worker. The strain displayed resistance to aminoglycosides, beta-lactams, polymyxins, fosfomycins, phenicols, trimethoprims, sulfonamides, tetracyclines and fluoroquinolones. Whole genome sequence analysis and plasmid conjugation revealed that the strain had a transferable IncHI2 plasmid carrying the mcr-1, blaCTX-M-14 and fosA3 genes. Sequence homology analysis showed that this plasmid possessed high sequence similarity to previously reported mcr-1, blaCTX-M-14 and fosA3 positive plasmids in China.
Conclusion: This study reported a the multidrug-resistant Salmonella 4,[5],12:i:- isolate harboring mcr-1, blaCTX-M-14 and fosA3 from human for the first time in China. The occurrence of mcr-1 and fosA3 genes in the transferable IncHI2 plasmid pYZU1189 from the ESBL-producing Salmonella 4,[5],12:i:- isolate showed a potential threat to public health. Great concern should be taken for the spread of multidrug-resistant ESBL-producing Salmonella isolates from food catering workers to consumers.
Keywords: Salmonella 4[5] 12:i:-, whole genome sequencing analysis, multidrug resistant, ESBL, colistin resistance, fosA3
Introduction
Salmonella enterica serovar Typhimurium is well known to be associated with foodborne diseases and outbreaks, as well as causing infectious diseases in animals and humans.1 In recent year, the Salmonella Typhimurium monophasic variant (Salmonella 4,[5],12:i:-) was frequently reported from human salmonellosis in both Europe and the US.2,3 Salmonella 4,[5],12:i:- is described as a variant of Salmonella Typhimurium lacking the second phase flagellum, which mostly belonged to the pandemic ST34 clone with an antibiotic resistance pattern of ASSuT (ampicillin, streptomycin, sulphonamides and tetracycline).4 In addition, the mcr-1 gene responsible for the colistin resistance has frequently occurred in ST34 Salmonella Typhimurium and Salmonella 4,[5],12:i:-.4–7 In China, Salmonella 4,[5],12:i:- has increased to being one of the most predominant serovars causing human salmonellosis, while ST34 is the most common ST type of Salmonella 4,[5],12:i:-.8 ESBL-producing Salmonella isolates have raised concern due to their resistance to cephalosporins, which have been widely applied for treating severe salmonellosis.9 The ESBL-producing Salmonella strains showed a higher prevalence (8.58%) on retail chicken in China than in the US.10,11 Meanwhile, fosfomycin has recently been reintroduced into clinical treatment for ESBL-producing Enterobacteriaceae. The occurrence of multidrug resistance in Salmonella 4,[5],12:i:- isolated from animal origin food products has increased the potential infection risk while treatment options are becoming limited.
In this study, we identified a MDR ST34 Salmonella 4,[5],12:i:- isolate carrying an IncHI2 plasmid with mcr-1, fosA3, blaCTX-M-14 genes, which was from a healthy catering industry worker in Jiangsu province, China. To our best knowledge, this is the first report of the multidrug-resistant Salmonella 4,[5],12:i:- isolate harboring mcr-1, blaCTX-M-14 and fosA3 from a human in China.
Materials and Methods
Bacterial Isolate
A surveillance study for foodborne pathogens was performed toward food catering workers and diarrheal patients by Nantong CDC, Jiangsu, China.12 The initial aim of this project was to evaluate the role of food handler as a vehicle for transmitting foodborne pathogens by screening human fecal samples. In order to selectively enrich Salmonella, samples were added to the selenite cysteine broth and incubated at 35°C for 24 h. The 10 µL of each incubated sample was inoculated onto xylose lysine deoxycholate (XLD, OXOID, England) and Salmonella Shigella (SS) agar plate, and cultured for 24 h at 37°C. Two or more presumptive Salmonella colonies were subcultured in the triple sugar-iron-agar medium. The serotyping of Salmonella was based on Kauffmann–White (KW) scheme. During the process, we identified Salmonella Typhimurium and its monophasic variant by multiplex-PCR for the fliB-fliA intergenic region and the fljB gene.13
Antimicrobial Susceptibility Testing
Minimum inhibitory concentrations (MICs) were determined by the agar dilution method. Antibiotics used in this study were listed in Table 1. MICs were interpreted by CLSI breakpoints,14,15 except that florfenicol (>16 mg/L) was interpreted according to EUCAST epidemiological cutoff values (http://mic.eucast.org/Eucast2).
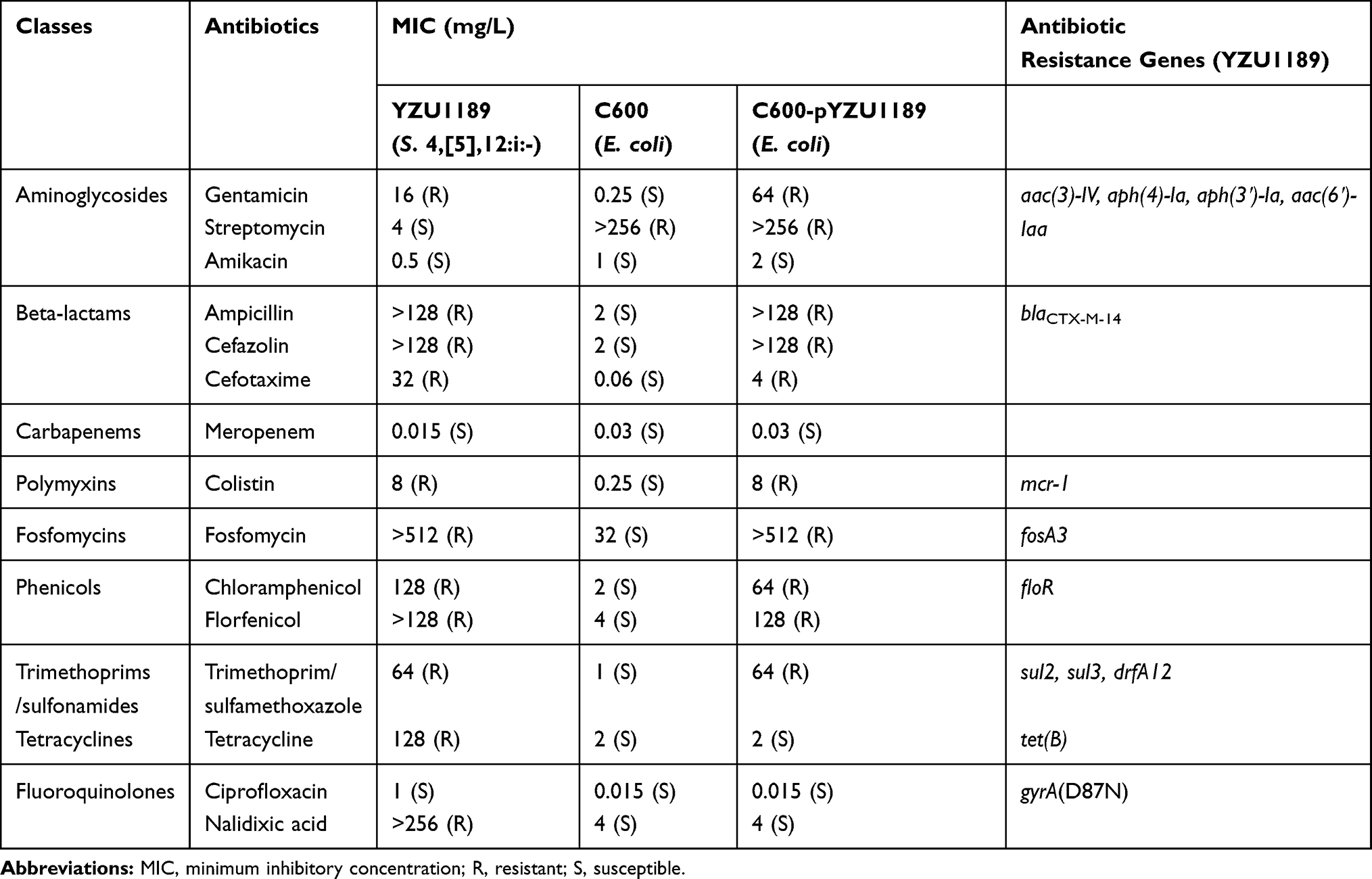 |
Table 1 The Minimum Inhibitory Concentration (MIC) Values of the Donor (Salmonella 4,[5],12:i:- YZU1189), Transconjugant (E. coli C600-pYZU1189) and Recipient (E. coli C600) |
Plasmid Conjugation Experiments
Conjugation experiments were performed between donor Salmonella 4,[5],12:i:- and recipient streptomycin-resistant E. coli C600, and transconjugants were selected on the Luria–Bertani agar plate containing streptomycin (3000 mg/L) and colistin (2 mg/L). The mcr-1 and mdh genes were used to distinguish donor, recipient and transconjugant based on the PCR analysis.16,17
Whole Genome Sequencing Analysis
To understand the genetic background of Salmonella 4,[5],12:i:- YZU1189, the strain YZU1189 was sequenced using the Hiseq 2500 platform. Raw sequencing reads were deposited in the European Nucleotide Archive database under the accession number PRJEB38934. The raw reads were trimmed and filtered by NGSQC toolkit (v2.3.3), and subjected to de novo assembly by SPAdes 3.6.18 The assembled genome was annotated by Prokka version 1.12.19 The acquired antimicrobial resistance genes and chromosomal mutations was identified by ResFinder 3.2.20 The multilocus sequence type of the strain YZU1189 was obtained by MLST 2.0.21 The virulence factors of strain YZU1189 was detected by BLASTn based on seven typical virulence genes in Salmonella Typhimurium, of which four genes located on prophages including gipA (encoding a Peyer’s patch-specific virulence factor), sspH1 (Salmonella-type III effector protein), sodC1 (putative Cu/Zn superoxide dismutase) and sopE1 (Salmonella-type III effector protein), and three genes (spvC, pefA, rck) in plasmids.22
Ethical Approval
The study protocol was performed following the ethical guidelines of the 1975 Declaration of Helsinki, and was approved by the Ethics Committee of the Chinese Centers for Disease Control and Prevention (CDC). Fresh feces of all healthy catering workers were sampled after obtaining written informed consents.
Results and Discussion
In the surveillance study, we observed a Salmonella 4,[5],12:i:- strain YZU1189 harboring mcr-1, fosA3 and blaCTX-M-14 from a healthy female catering worker. This MDR isolate was defined using the criteria established by Magiorakos et al.23 MICs of 15 antibiotics for the strain YZU1189 showed that YZU1189 was resistant to antibiotics including aminoglycosides, beta-lactams, polymyxins, fosfomycins, phenicols, trimethoprims, sulfonamides, tetracyclines and fluoroquinolones (Table 1). Compared to the typical ASSuT resistance pattern in most of Salmonella 4,[5],12:i:- isolates, this strain lacked the phenotypic streptomycin resistance, while showed resistance to colistin and fosfomycin.
According to the MLST analysis, Salmonella 4,[5],12:i:- strain YZU1189 belonged to sequence type (ST) 34. Virulence factors analysis of seven genes showed that this isolate carried virulence factors gipA and sodC1, which were prevalent in Salmonella 4,[5],12:i:- isolates from foods in China.8 Strain YZU1189 contained several antimicrobial resistance (AMR) genes including aminoglycosides (aac(3)-IV, aph(4)-Ia, aph(3ʹ)-Ia, aac(6ʹ)-Iaa), β-lactamase (blaCTX-M-14), polymyxins (mcr-1), fosfomycins (fosA3), phenicols (floR), sulphonamides (sul2 and sul3), tetracycline (tet(B)), and trimethoprims (drfA12). In addition, the mutation of codon 87 (D87N) in the GyrA protein causing the quinolone resistance was also detected in this strain. The mcr-1 gene was previously observed in ST34 Salmonella Typhimurium from pigs in China, which located mainly on IncHI2-like plasmids.5 The PCR analysis showed that the transconjugant was positive for the mcr-1 gene and negative for the Salmonella Typhimurium specific mdh gene, indicating the successful transmission of mcr-1 positive plasmid into the recipient strain E. coli C600. The plasmid conjugation experiment showed that the mcr-1 gene in YZU1189 could be co-transferred with blaCTX-M-14 and fosA3 genes (Table 1).
By whole genome sequencing analysis, we confirmed several antimicrobial resistance genes located on an IncHI2 plasmid (246Kb), named pYZU1189 (accession number ERS4951189), which harbored mcr-1, fosA3, floR, sul2, sul3, drfA12, aac(3)-IV, aph(4)-Ia, aph(3ʹ)-Ia, blaCTX-M-14 genes (Figure 1). The mcr-1 gene was located in the common genetic background of ISApl1-mcr-1-orf, which was also detected in pECHN-15–61(IncI2), pECFJ-B42-63(IncI2), and pECJS-B60-267(IncHI2) plasmids,24 while the fosA3 gene had the common genetic background of ΔISEcp1-blaCTX-M-14-IS903B-fosA3-orf located in pYZU1189. IncHI2 plasmids harboring antimicrobial genes found in this study were also reported in Enterobacteriaceae, such as E. coli and Salmonella spp.6,7,24 The sequence alignment showed that an IncHI2-type plasmid pHNSHP45-2 harboring a similar genetic background with mcr-1, blaCTX-M-14 and fosA3 in E. coli (Figure 1). Five Salmonella Typhimurium isolates carrying co-transferred fosA3, blaCTX-M-14, mcr-1, oqxAB and floR genes from food animals between 2016 and 2017 in China have been previously reported.6 More importantly, mcr-positive Salmonella 4,[5],12:i:- isolates with multiple antimicrobial resistance genes have been frequently reported from different sources in multiple countries worldwide,25–32 indicating the increasing risk potential of this pathogen transmitting worldwide. Also, mcr-positive Salmonella 4,[5],12:i:- isolates from difference sources showed a potential dissemination between food animals and humans, which remains a significant threat to human health.
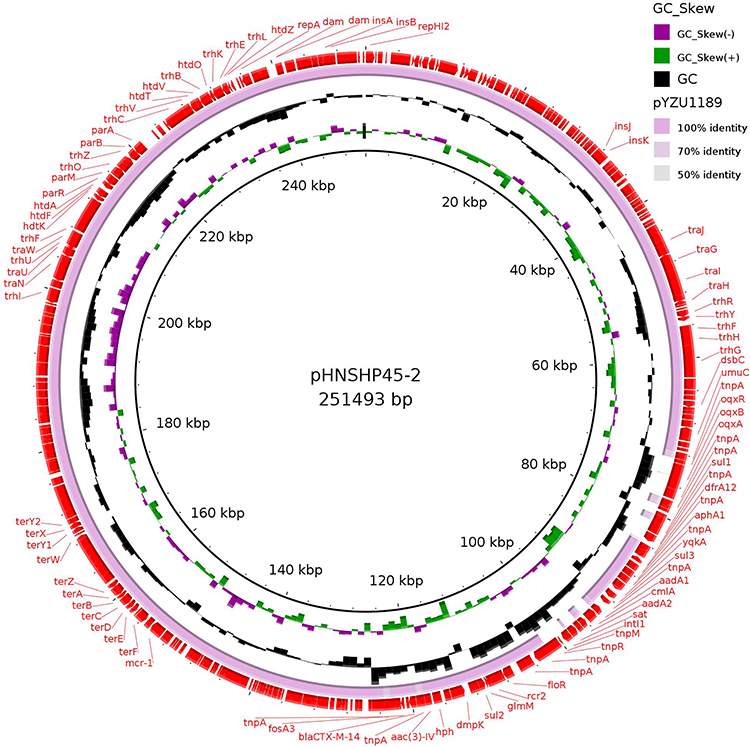 |
Figure 1 Sequence alignment of pYZU1189 and pHNSHP45-2 (GenBank no. KU341381). The pHNSHP45-2 was used as a reference to compare with mcr-1-bearing plasmid which possess the IncHI2 replicon in this study. Red arrows represent the plasmid pHNSHP45-2. |
Conclusion
In conclusion, this study reported an ESBL-producing Salmonella 4,[5],12:i- isolate of human origin carrying a colistin resistance gene mcr-1, a fosfomycin resistance gene fosA3 and a rare sulfonamide resistance gene sul3. The occurrence of mcr-1 and fosA3 genes in a transferable IncHI2 plasmid pYZU1189 from a ESBL-producing Salmonella 4,[5],12:i:- isolate brings great considerable public health threat. Great concern should be taken for the spread of multidrug-resistant ESBL-producing Salmonella isolates from food catering workers to consumers. In the “One Health” aspect, quick and thorough action should be taken to reduce the use of colistin in food-producing animals in the farming industry. Interventions of restricting antibiotic use in food-producing animals should also be applied to reduce the prevalence of MDR bacteria.
Funding
This research was funded by National Natural Science Foundation of China, grant number 3192010301, and 31730094; The Priority Academic Program Development of Jiangsu Higher Education Institution (PAPD) and Jiangsu Key Laboratory of Zoonosis (R1703).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Zhang J, Jin H, Hu J, et al. Serovars and antimicrobial resistance of non-typhoidal Salmonella from human patients in Shanghai, China, 2006–2010. Epidemiol Infect. 2014;142:826–832.
2. European Food Safety Authority and European Centre for Disease Prevention and Control (EFSA and ECDC). The European union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2017. EFSA J. 2018;16(12):e5500.
3. CDC (Centers for Disease Control and Prevention). National Salmonella surveillance annual report, 2016; 2018. Available from: https://www.cdc.gov/nationalsurveillance/salmonella-surveillance.html.
4. Biswas S, Li Y, Elbediwi M, Yue M. Emergence and dissemination of mcr-carrying clinically relevant Salmonella Typhimurium monophasic clone ST34. Microorganisms. 2019;7.
5. Yi LX, Wang J, Gao YL, et al. Mcr-1-harboring Salmonella enterica serovar Typhimurium sequence type 34 in pigs, China. Emerg Infect Dis. 2017;23:291–295.
6. Mulvey MR, Bharat A, Boyd DA, Irwin RJ, Wylie J. Characterization of a colistin-resistant Salmonella enterica 4,[5],12:i:- harbouring mcr-3.2 on a variant IncHI-2 plasmid identified in Canada. J Med Microbiol. 2018;67:1673–1675.
7. Monte DF, Nelson V, Cerdeira L, et al. Multidrug- and colistin-resistant Salmonella enterica 4,[5],12:i:- sequence type 34 carrying the mcr-3.1 gene on the IncHI2 plasmid recovered from a human. J Med Microbiol. 2019;68:986–990.
8. Yang X, Wu Q, Zhang J, et al. Prevalence and characterization of monophasic Salmonella serovar 1,4,[5],12:i:- of food origin in China. PLoS One. 2015;10:e0137967.
9. Medalla F, Gu W, Mahon BE, et al. Estimated incidence of antimicrobial drug–resistant nontyphoidal salmonella infections, United States, 2004–2012. Emerg Infect Dis. 2017;23:29–37.
10. Food and Drug Administration (FDA). National antimicrobial resistance monitoring system-enteric bacteria (narms): 2015 integrated report. Rockville,MD, USA: US Department of Health and Human Services, FDA; 2017. Available from: https://www.fda.gov/downloads/AnimalVeterinary/SafetyHealth/AntimicrobialResistance/NationalAntimicrobialResistanceMonitoringSystem/UCM581468.pdf.
11. Wu H, Xia X, Cui Y, et al. Prevalence of extended-spectrum b-lactamase producing Salmonella on retail chicken in six provinces and two national cities in the People’s Republic of China. J Food Prot. 2013;76:2040–2044.
12. Xu H, Zhang W, Guo C, et al. Prevalence, serotypes, and antimicrobial resistance profiles among Salmonella isolated from food catering workers in Nantong, China. Foodborne Pathog Dis. 2019;16(5):346–351.
13. Tennant SM, Diallo S, Levy H, et al. Identification by PCR of non-typhoidal Salmonella enterica serovars associated with invasive infections among febrile patients in Mali. PLoS Negl Trop Dis. 2010;4.
14. Clinical and Laboratory Standards Institute. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically. M07 Standard.
15. Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing. M100 Standard.
16. Liu YY, Wang Y, Walsh TR, et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: a microbiological and molecular biological study. Lancet Infect Dis. 2016;16:161–168.
17. Amavisit P, Boonyawiwat W, Bangtrakulnont A. Characterization of Salmonella enterica serovar Typhimurium and monophasic Salmonella serovar 1,4,[5],12:i:- isolates in Thailand. J Clin Microbiol. 2005;43:2736–2740.
18. Bankevich A, Nurk S, Antipov D, et al. SPAdes: a new genome assembly algorithm and its applications to single-cell sequencing. J Comput Biol. 2012;19:455–477.
19. Seemann T. Prokka: rapid prokaryotic genome annotation. Bioinformatics. 2014;30:2068–2069.
20. Zankari E, Hasman H, Cosentino S, et al. Identification of acquired antimicrobial resistance genes. J Antimicrob Chemother. 2012;67:2640–2644.
21. Larsen MV, Cosentino S, Rasmussen S, et al. Multilocus sequence typing of total-genome-sequenced bacteria. J Clin Microbiol. 2012;50:1355–1361.
22. Capuano F, Mancusi A, Capparelli R, Esposito S, Proroga YTR. Characterization of drug resistance and virulotypes of Salmonella strains isolated from food and humans. Foodborne Pathog Dis. 2013;10:963–968.
23. Magiorakos AP, Srinivasan A, Carey RB, et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance. Clin Microbiol Infect. 2012;18(3):268–281.
24. Li RC, Xie MM, Zhang JF, et al. Genetic characterization of mcr-1-bearing plasmids to depict molecular mechanisms underlying dissemination of the colistin resistance determinant. J Antimicrob Chemother. 2017;72:393–401.
25. Fang LX, Jiang Q, Deng GH, et al. Diverse and flexible transmission of fosA3 associated with heterogeneous multidrug resistance regions in Salmonella enterica serovar Typhimurium and indiana isolates. Antimicrob Agents Chemother. 2020;64.
26. Li XP, Fang LX, Song JQ, et al. Clonal spread of mcr-1 in PMQR-carrying ST34 Salmonella isolates from animals in China. Sci Rep. 2016;6:38511.
27. Doumith M, Godbole G, Ashton P, et al. Detection of the plasmid-mediated mcr-1 gene conferring colistin resistance in human and food isolates of Salmonella enterica and Escherichia coli in England and Wales. J Antimicrob Chemother. 2016;71:2300–2305.
28. Campos J, Cristino L, Peixe L, Antunes P. MCR-1 in multidrug-resistant and copper-tolerant clinically relevant Salmonella 1,4,[5],12:i:- and S. Rissen clones in Portugal, 2011 to 2015. Euro Surveill. 2016;21.
29. Litrup E, Kiil K, Hammerum AM, et al. Plasmid-borne colistin resistance gene mcr-3 in Salmonella isolates from human infections, Denmark, 2009–17. Euro Surveill. 2017;22.
30. Saavedra SY, Diaz L, Wiesner M, et al. Genomic and molecular characterization of clinical isolates of Enterobacteriaceae harboring mcr-1 in Colombia, 2002 to 2016. Antimicrob Agents Chemother. 2017;61.
31. Arnott A, Wang Q, Bachmann N, et al. Multidrug-resistant Salmonella enterica 4,[5],12:i:- sequence type 34, New South Wales, Australia, 2016–2017. Emerg Infect Dis. 2018;24:751–753.
32. Borowiak M, Hammerl JA, Deneke C, et al. Characterization of mcr-5-harboring Salmonella enterica subsp. enterica serovar Typhimurium isolates from animal and food origin in Germany. Antimicrob Agents Chemother. 2019;63.
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.