Back to Journals » Infection and Drug Resistance » Volume 11
A molecular epidemiological study of methicillin-resistant and methicillin-susceptible Staphylococcus aureus contamination in the airport environment
Authors Chen Z, Han C, Huang X, Liu Y, Guo D, Ye X ![]()
Received 28 June 2018
Accepted for publication 16 September 2018
Published 20 November 2018 Volume 2018:11 Pages 2363—2375
DOI https://doi.org/10.2147/IDR.S178584
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Professor Suresh Antony
Zhiyao Chen, Changlin Han, Xiaobin Huang, Yangqun Liu, Dan Guo, Xiaohua Ye
Laboratory of Molecular Epidemiology, School of Public Health, Guangdong Pharmaceutical University, Guangzhou 510310, China
Background: Methicillin-resistant Staphylococcus aureus (MRSA) causes a wide variety of serious infections worldwide. There are few studies on the prevalence, antimicrobial susceptibility, and molecular characteristics of MRSA contamination in the environment of airports.
Materials and methods: A cross-sectional survey was conducted in Guangzhou Baiyun Airport. Environmental surface sampling was conducted in frequently touched locations for S. aureus analysis. All isolates were characterized by multilocus sequence typing (MLST) and tested for antimicrobial susceptibility, resistance genes, and virulence genes. Data were analyzed by chi-squared test and correspondence analysis.
Results: Of the 1,054 surface samples, the contamination rate was 7.2% (76/1,054) for S. aureus and 2.2% (23/1,054) for MRSA. There were 62.9% (56/89) S. aureus isolates classified as multidrug resistant (MDR), with six linezolid-resistant isolates and two cfr-carrying isolates. The most prevalent S. aureus genotypes were CC6 (ST6), CC59 (ST59), and CC188 (ST188), with ST59-MRSA-IV (pvl–) as the predominant MRSA. There were significant differences between methicillin-resistant and methicillin-sensitive isolates in rates of resistance to tetracycline (P<0.001) and sek carriage (P=0.029). The correspondence analyses revealed significant corresponding relationships between genotypes and phenotype–genotype characteristics of S. aureus isolates.
Conclusion: Our findings revealed a potential risk of cross-transmission of MRSA between human beings and environments, suggesting more stringent contamination control measures. In addition, this study revealed significant corresponding relationships between genotypes and phenotype–genotype characteristics of S. aureus isolates, which may provide new ideas for monitoring the latest epidemiological trends.
Keywords: Staphylococcus aureus, methicillin-resistant S. aureus, environment, molecular characterization, antimicrobial susceptibility
Introduction
Staphylococcus aureus is a major human pathogen, and methicillin-resistant S. aureus (MRSA) causes a wide variety of infections worldwide, including hospital-associated MRSA (HA-MRSA) and community-associated MRSA (CA-MRSA) infections.1,2 Previously, many studies have focused on assessing CA-MRSA infections, which were associated with health care or hospital contact.2,3 To date, more and more epidemiological studies have reported the CA-MRSA infection in the population without established health care risk factors.4,5 The emergence of cross-transmission of CA-MRSA and HA-MRSA in hospital settings poses a potential public health risk.6,7 Therefore, future studies should pay more attention to explore the potential transmission route for MRSA isolates, especially for closely monitoring environmental MRSA contamination.
To date, most studies mainly focused on MRSA contamination in the environment of public transports, indicating that environment surfaces in public transports may be a hazardous reservoir for the transmission of S. aureus (including MRSA) to passengers.8,9 A study of airport door handles revealed that the contamination rate was 5.5% for S. aureus and 0.3% for MRSA.10 In addition, the latest report described the introduction of various multidrug-resistant (MDR) bacteria through a patient repatriated after hospitalization abroad and family members receiving this repatriated patient who were also colonized with MDR bacteria, which emphasize the importance of adherence to isolation precautions for returning patients.11 International travelers with skin and soft tissue infections also promote the emergence and spread of highly virulent or antibiotic-resistant S. aureus in their home countries, proposing a network for the surveillance of imported S. aureus.12 Airports are important travel hubs for the global spread of antimicrobial-resistant bacteria, but the characteristics of MRSA transmission in the environment of airports remain uncertain. In addition, few studies have attempted to assess the potential relationships between phenotype–genotype characteristics and molecular typing of S. aureus isolates. Therefore, this study aimed to examine the contamination, antimicrobial susceptibility, virulence genes, and molecular typing of S. aureus isolates at the Guangzhou Baiyun International Airport. In addition, this study adds to existing literature by using correspondence analysis to explore the potential relationships between phenotype–genotype characteristics and molecular typing of S. aureus isolates.
Materials and methods
Environmental sample collection
This cross-sectional study of surface sampling was conducted between October and November 2017 in Guangzhou Baiyun International Airport, which is the only airport in Guangzhou, China. In 2017, this airport was China’s third busiest and world’s 13th busiest airport by passenger traffic, with 65,806,977 passengers handled. Environmental surface sampling was conducted in six types of locations, including luggage trolleys, self-service machines, escalators, seat handles, luggage detectors, and water dispensers. These locations were chosen because they are frequently touched and are more likely to be contaminated with bacteria. Swabs moistened with sterile normal saline were used to sample surfaces, and the sample area of each swab was approximately 10 cm × 10 cm. In total, there were 1,054 non-duplicate environmental samples collected, including 251 from luggage trolleys, 244 from self-service machines, 233 from escalators, 219 from seat handles, 68 from luggage detectors, and 39 from water dispensers.
Bacterial isolation and identification
Swabs were soaked into 5 mL enrichment broth (0.25% yeast extract, 1% mannitol, 1% tryptone, and 7.5% NaCl) at 4°C during transportation for about 20 minutes and incubated at 35±1°C for 24 hours. Then, a loopful of the broth was plated onto mannitol salt agar and incubated at 37°C for 24–48 hours. Suspected colonies were selected and subcultured to 5% sheep blood agar plates and incubated at 35±1°C overnight. S. aureus isolates were identified by a combination of Gram staining, morphology, hemolysis pattern, catalase test, tube coagulase test, and DNase test. PCR assays were also used to test the staphylococci 16SrRNA and nuc and mecA genes.13 All S. aureus isolates carried the 16S rRNA and nuc genes. The mecA-positive S. aureus was identified as genotypic MRSA.
Antimicrobial susceptibility testing
The Kirby-Bauer disk diffusion method was used to test susceptibility to antibiotics, and diameter interpretations were based on the recommendations of the Clinical and Laboratory Standards Institute (CLSI, 2013). All S. aureus isolates were assessed for susceptibility to a panel of 11 antibiotics: cefoxitin (30 µg), erythromycin (15 µg), clindamycin (2 µg), tetracycline (30 µg), chloromycetin (30 µg), trimethoprim–sulfamethoxazole (23.75 µg), ciprofloxacin (5 µg), gentamicin (10 µg), rifampicin (5 µg), quinupristin–dalfopristin (15 µg), and linezolid (30 µg). The reference S. aureus ATCC 25923 was used for quality control, and S. aureus ATCC 29213 was used for positive control. Cefoxitin-resistant isolates were identified as phenotypic MRSA. The detection of inducible clindamycin resistance was done using the D-test method. Erythromycin resistance phenotypes were classified as the inducible macrolide–lincosamide–streptogramin B (MLSB; resistance to erythromycin and inducible resistance to clindamycin), constitutive MLSB (resistance to both erythromycin and clindamycin), or M resistance (resistance to erythromycin but susceptibility to clindamycin). S. aureus isolates were classified as MDR if they were non-susceptible (including both intermediate and resistant isolates) to ≥3 classes of antibiotics,14 including mild MDR (non-susceptible to 3–4 classes of antibiotics) and extended MDR (non-susceptible to ≥5 classes of antibiotics). Specifically, all MRSA isolates were classified as MDR, including mild MDR (non-susceptible to ≤4 classes of antibiotics besides beta-lactams) and extended MDR (non-susceptible to ≥5 classes of antibiotics besides beta-lactams).
Molecular characterization
Multilocus sequence typing (MLST) of the seven housekeeping genes (arcC, aroE, glpF, gmk, pta, tpi, and yqiL) was conducted for all S. aureus isolates.15 The sequence types (STs) for each isolate were determined by comparing the sequence obtained to known alleles at each locus in the MLST database (http://saureus.mlst.net), and the clonal complexes (CCs) were determined using the eBURST algorithm (http://eburst.mlst.net). In addition, specific PCR tests were used to detect the staphylococcal cassette chromosome mec element (SCCmec) type (I–V), the Panton–Valentine leukocidin toxin gene (pvl), the immune evasion cluster (IEC) genes (scn, chp, sak, sea, and sep), the hemolysin genes (hla, hlb, and hld), the exfoliative toxin genes (eta, etb, and etd), the enterotoxin genes (sea, seb, sec, sed, see, seg, seh, sei, sej, sek, sel, sem, sen, seo, sep, seq, ser, and seu), the toxic shock syndrome toxin-1 gene (tsst1), and the linezolid-resistant gene (cfr).16–21
Statistical analyses
The differences in categorical variables between groups were compared by logistic regression or exact logistic regression. Correspondence analysis shows or summarizes a set of data in two-dimensional graphical form, which will provide a useful graphic and statistical method for exploring the internal relationship between categorical variables.22 It is conceptually similar to principal component analysis, but applies to categorical rather than continuous data. So, we used the correspondence analysis to explore the potential relationships between CC typing and phenotype–genotype characteristics (including resistance pattern, IEC genes, enterotoxin genes, and hemolysin genes) of S. aureus isolates. The statistical analysis was conducted using STATA version 14.0 (StataCorp LP, College Station, TX, USA). A two-sided P-value of <0.05 was considered to be statistically significant.
Results
Prevalence of S. aureus and MRSA
A total of 1,054 surface samples were collected in this study (Table 1). Among 76 S. aureus-contaminated samples, we identified 89 S. aureus isolates (including one strain isolated from each of the 63 samples and two strains isolated from each of the other 13 samples) and 28 MRSA isolates (including 18 isolates with both phenotypic and genotypic resistance, seven isolates with only genotypic resistance and three isolates with only phenotypic resistance). The overall prevalence of S. aureus contamination was 7.2% (76/1,054) and was significantly higher in self-service machines (OR =4.55, 95% CI 2.06–10.06) and luggage detectors (OR =3.75, 95% CI 1.35–10.40) than in escalators. The overall prevalence of MRSA contamination was 2.2% (23/1,054) and was significantly higher in self-service machines (OR =12.47, 95% CI 2.77–56.15), luggage detectors (OR =11.12, 95% CI 1.82–67.82), and seat handles (OR =5.63, 95% CI 1.08–29.25) than in escalators and luggage trolleys.
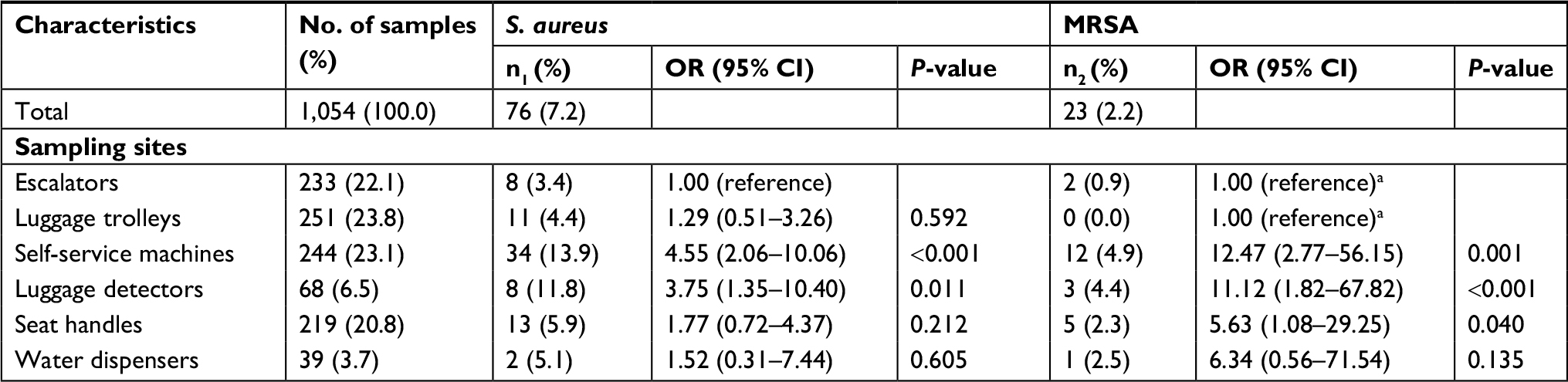 | Table 1 Prevalence of S. aureus and MRSA contamination in 1,054 samples collected from different sites Notes: aBoth escalators and luggage trolleys were used as the reference group. n1, frequency of S. aureus-positive samples; n2, frequency of MRSA-positive samples. Abbreviations: MRSA, methicillin-resistant S. aureus; S. aureus, Staphylococcus aureus. |
Antimicrobial susceptibility and resistance genes
As to resistance phenotypes for 89 S. aureus isolates, most of the isolates showed low resistance percentage to gentamicin (3.4%) and linezolid (6.7%), but one-third of the isolates were resistant to erythromycin (36.0%) and clindamycin (33.7%; Table 2). Among the 32 erythromycin-resistant isolates, 59.0% (19 isolates) expressed the constitutive MLSB and 41.0% (13 isolates) expressed the M resistance, but no isolates expressed the inducible MLSB. Notably, 56 (62.9%) isolates were classified as MDR, and the most common MDR pattern was co-non-susceptibility to erythromycin, clindamycin, and quinupristin–dalfopristin (19 isolates, including six linezolid-resistant isolates; Figure S1). Notably, there were two isolates (belonged to MRSA) carrying the cfr gene, which were resistant to linezolid, cefoxitin, erythromycin, clindamycin, quinupristin–dalfopristin, trimethoprim–sulfamethoxazole, ciprofloxacin, tetracycline, and chloromycetin.
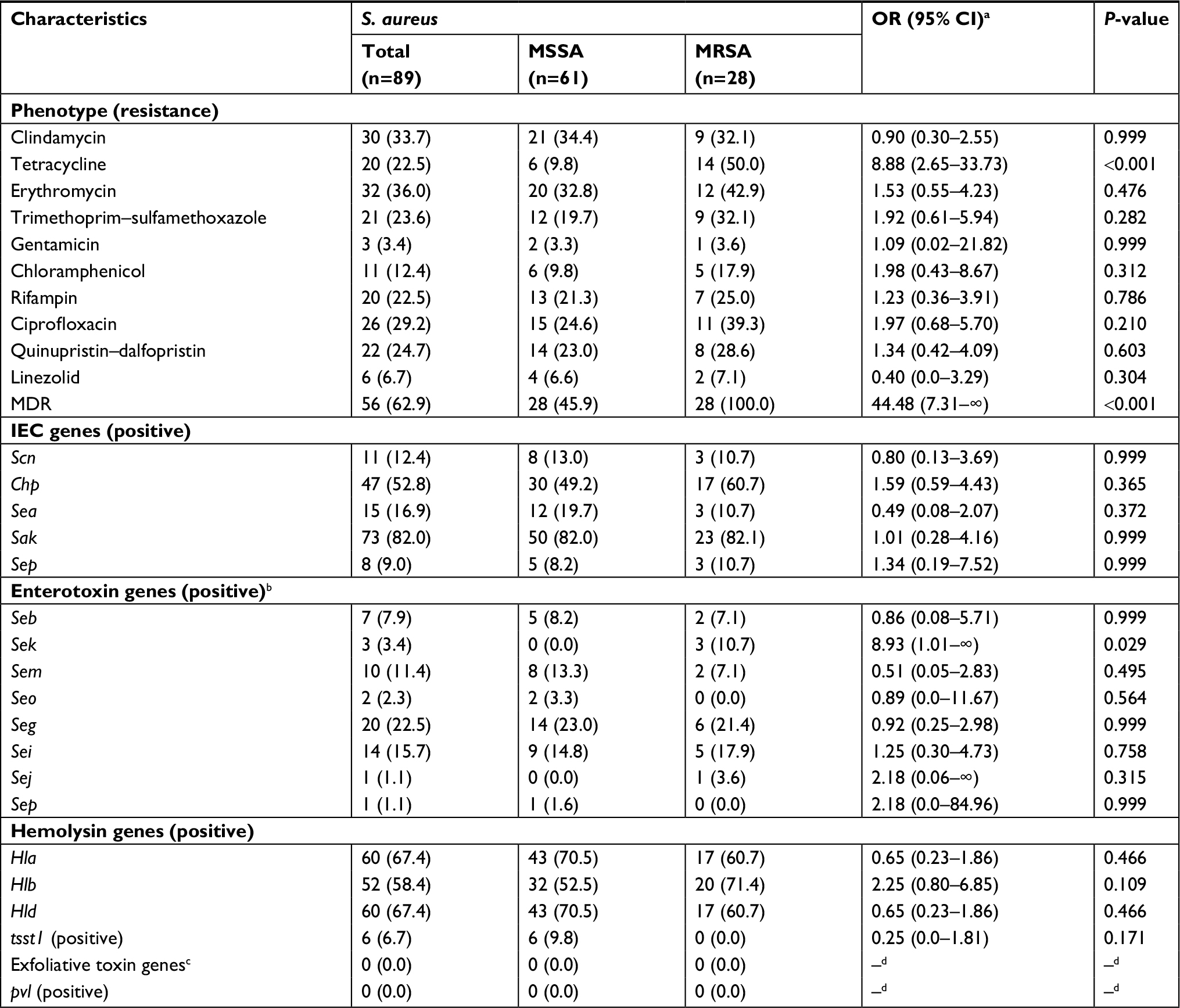 | Table 2 Phenotypic and molecular characteristics of MSSA and MRSA isolates Notes: Values are expressed as the number of isolates with characteristics (the proportion of isolates with characteristics), except where specified otherwise. aORs were estimated by the exact logistic regression. bAll isolates were negative for some enterotoxin genes, including sea, sec, sed, see, seh, sel, sen, seq, ser, and seu. cAll isolates were negative for all exfoliative toxin genes, including eta, etb, and etd. dNo estimate was provided due to the lack of occurrence of the outcome of interest in the two groups. Abbreviations: IEC, immune evasion cluster; MRSA, methicillin-resistant Staphylococcus aureus; MSSA, methicillin-sensitive Staphylococcus aureus. |
Molecular typing and virulence genes
Among 89 S. aureus isolates, we observed 33 unique STs belonging to 10 CCs, except for four untypeable isolates. The most prevalent S. aureus STs were ST6 (nine isolates), ST 188 (nine isolates), ST30 (six isolates), and ST59 (six isolates), with the predominant MRSA being ST59-MRSA-IV. Notably, two livestock-associated MRSA isolates (one for ST398 and the other for ST5) were also found in this study. The most common S. aureus CCs were CC6 (nine isolates), CC59 (nine isolates), CC188 (nine isolates), CC30 (seven isolates), CC5 (six isolates), CC20 (six isolates), CC398 (six isolates), and CC7 (five isolates), with the predominant MRSA being CC59-MRSA-IV.
As to the IEC genes (Table 2), sak (82.0%) predominated, followed by chp (52.8%), sea (16.9%), scn (12.4%), and sep (9.0%). In terms of the enterotoxin genes, the predominant gene was seg (22.5%), followed by sei (15.7%), sem (11.4%), seb (7.9%), sek (3.4%), and seo (2.3%). However, several enterotoxin genes (including sea, sec, sed, see, seh, sel, sen, seq, ser, and seu) were absent from all S. aureus isolates. With regard to the hemolysin genes, hla was detected in 67.4%, hlb in 58.4%, and hld in 67.4% of the S. aureus isolates. In addition, the tsst1 gene was only identified in six isolates, and pvl gene was absent from all S. aureus tested.
Phenotypic and molecular characteristics of MRSA and methicillin-sensitive S. aureus (MSSA)
In terms of the resistance phenotypes (Table 2), the MRSA isolates had a significantly higher rate of resistance to tetracycline (50.0% vs 9.8%; OR =8.88, 95% CI 2.65–33.73; P<0.001) than MSSA isolates. However, no significant differences were found for the rates of resistance to other antibiotics. In addition, the rate of multidrug resistance was significantly higher in MRSA isolates than in MSSA isolates (100.0% vs 45.9%; OR =44.48, 95% CI 7.31–∞; P<0.001). In terms of virulence genes, the rate of carrying sek gene was significantly higher in MRSA isolates than in MSSA isolates (10.7% vs 0.0%; OR =8.93, 95% CI 1.01–∞; P=0.029), but no significant differences were observed for the carriage of other virulence genes.
Relationship between CC typing and phenotype–genotype characteristics
The first correspondence analysis revealed a significant corresponding relationship between CC typing and resistance pattern (χ2=34.46, P=0.044; Figure 1A), suggesting that CC5/CC20/CC59/ST288 isolates were associated with being extended MDR, CC30 and CC672 isolates were associated with mild MDR, and CC6/CC7/CC8/CC45/CC188/CC398 isolates were associated with non-MDR. The second correspondence analysis revealed a significant corresponding relationship between CC typing and the number of IEC genes (χ2=41.14, P=0.008; Figure 1B). For example, CC6/CC7/CC20/CC30/CC45/CC59/CC188/CC398 isolates were associated with carrying two to four IEC genes, CC5 and CC672 isolates were associated with carrying one IEC gene, and ST288 isolates were associated with the absence of these IEC genes. As summarized in Table 3, CC6 and CC30 isolates were associated with carrying sak and sea, CC20/CC45/CC59/CC188/CC398 isolates were associated with carrying sak and chp, CC7 isolates were associated with carrying sak and sep, and CC5 and CC672 isolates were associated with carrying sak.
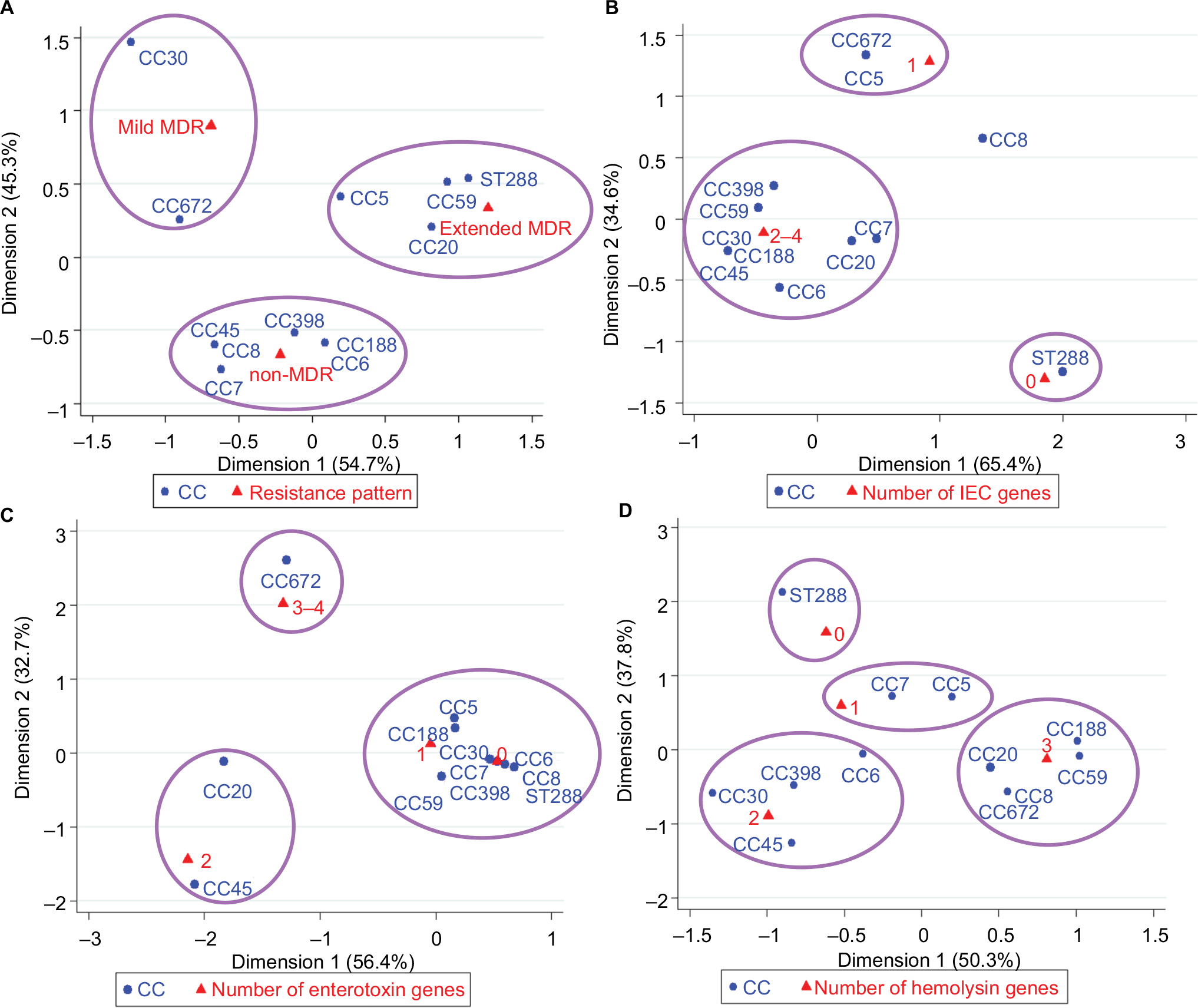 | Figure 1 Correspondence analysis for the relationship between CC and resistance pattern (A), the number of IEC genes (B), the number of enterotoxin genes (C), or the number of hemolysin genes (D) of S. aureus isolates. Abbreviations: CC, clonal complex; IEC, immune evasion cluster; MDR, multidrug resistant; S. aureus, Staphylococcus aureus. |
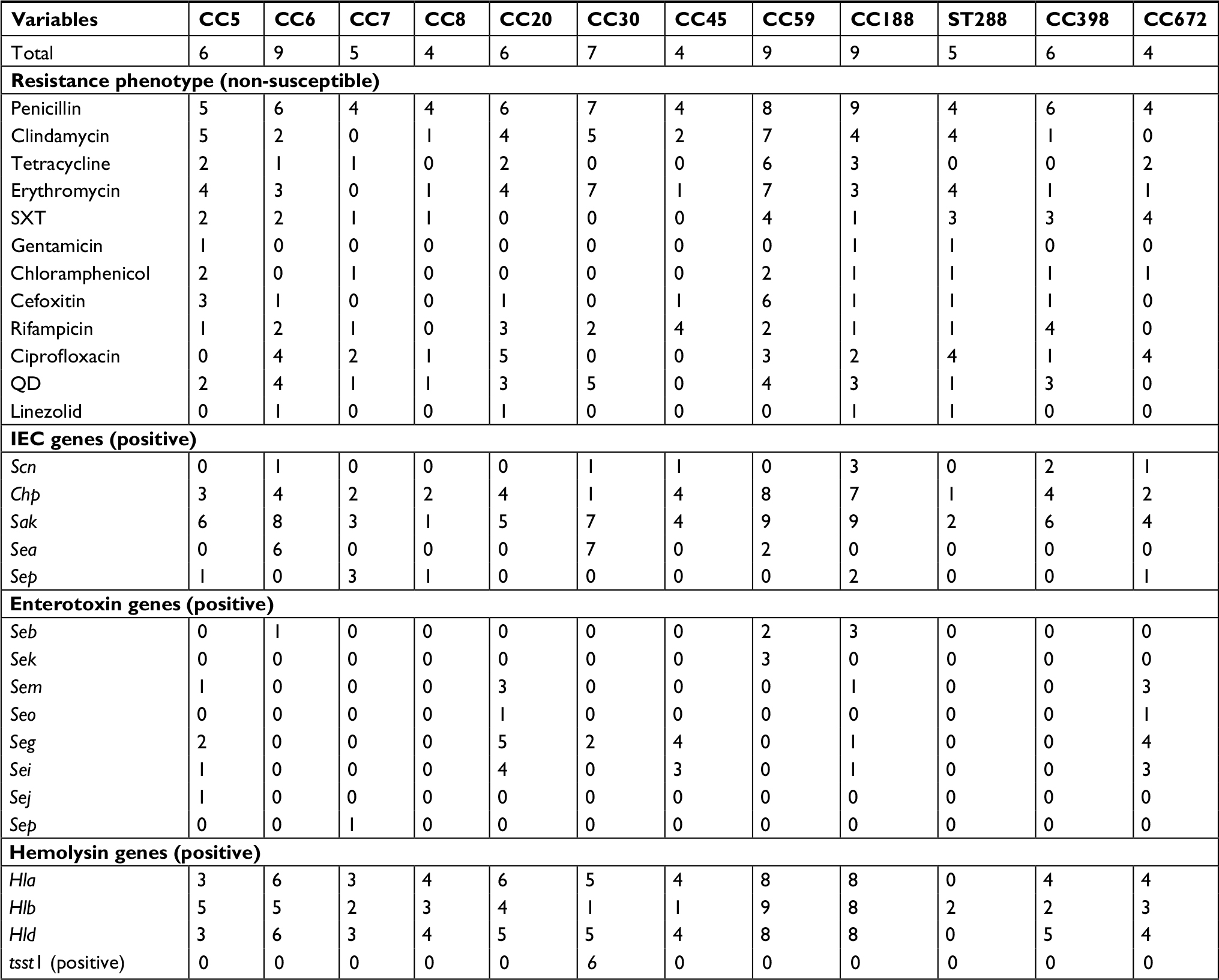 | Table 3 Relationship between predominant CCs and phenotype–genotype characteristics of S. aureus isolates Notes: Values are expressed as the number of isolates. All isolates were negative for pvl and some enterotoxin genes (sea, sec, sed, see, seh, sel, sen, seq, ser, and seu). Abbreviations: CC, clonal complex; IEC, immune evasion cluster; QD, quinupristin–dalfopristin; S. aureus, Staphylococcus aureus; SXT, trimethoprim–sulfamethoxazole. |
In addition, the third correspondence analysis indicated a significant corresponding relationship between CC typing and the number of enterotoxin genes (χ2=79.47, P<0.001; Figure 1C). For example, CC672 isolates were associated with carrying three or four enterotoxin genes (sem, seg, and sei), CC20 and CC45 isolates were associated with carrying two enterotoxin genes (seg and sei), and other isolates were associated with carrying only one enterotoxin gene or the absence of these genes (Table 3). The fourth correspondence analysis indicated a significant corresponding relationship between CC typing and the number of hemolysin genes of S. aureus isolates (χ2=61.30, P=0.002; Figure 1D). For example, CC8/CC20/CC59/CC188/CC672 isolates were associated with carrying three hemolysin genes (hla, hlb, and hld), CC6/CC30/CC45/CC398 isolates were associated with carrying two hemolysin genes (hla and hld), CC5 and CC7 isolates were associated with carrying only one hemolysin gene (hlb for CC5; hla or hld for CC7), and ST288 isolates were associated with the absence of these hemolysin genes (Table 3).
Notably, all virulence genes (including IEC genes, enterotoxin genes, and hemolysin genes) were absent from the ST288 isolates. In addition, this study showed that all tsst-1-positive isolates belonged to CC30, including five isolates for ST30 and one for ST435. The abovementioned analysis revealed that the most prevalent clones of S. aureus were CC6 (sak–sea–hla–hld), CC59 (chp–sak–hla–hlb–hld), CC188 (chp–sak–hla–hlb–hld), CC30 (sak–sea–hla–hld–tsst1), CC5 (sak–hlb), CC20 (chp–sak–seg–sei–hla–hlb–hld), and CC398 (chp–sak–hla–hld; Table 3 and Figures 2 and 3). The predominant clones of MRSA were CC59-IVd (chp–sak–hla–hlb–hld), CC6 (sak–hla–hld), CC188-II (chp–sak–hla–hlb–hld) and CC5 (sak–hlb; Figure 2).
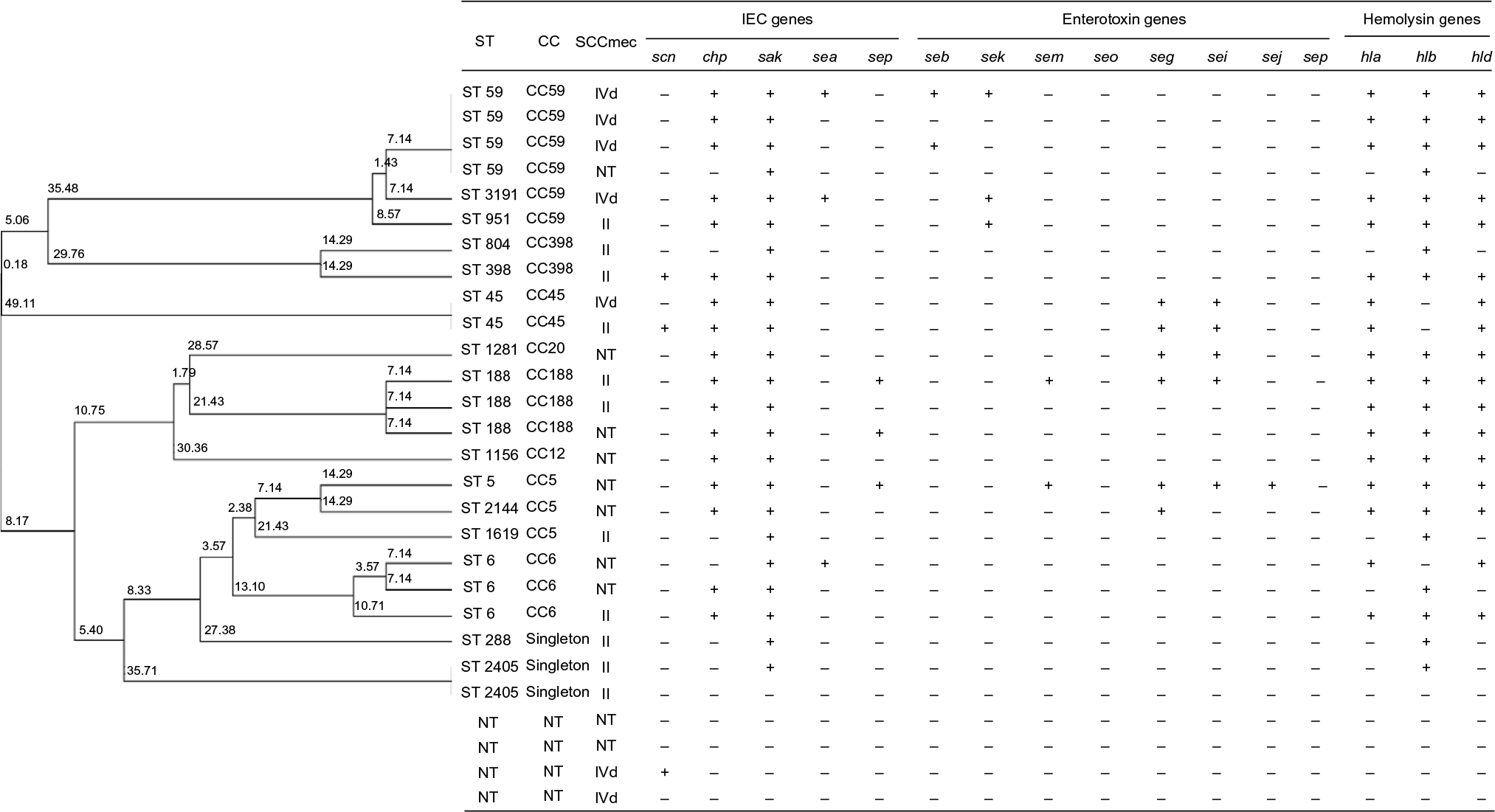 | Figure 2 Clonal dendrogram and detailed molecular characteristics of 28 MRSA isolates. Abbreviations: CC, clonal complex; IEC, immune evasion cluster; MRSA, methicillin-resistant Staphylococcus aureus; ST, sequence type; NT, non-typeable. |
 | Figure 3 Clonal dendrogram and detailed molecular characteristics of 61 MSSA isolates. Abbreviations: CC, clonal complex; IEC, immune evasion cluster; MSSA, methicillin-sensitive Staphylococcus aureus; ST, sequence type. |
Discussion
In the current study, the contamination rate was 7.2% for S. aureus and 2.2% for MRSA, which is similar to the results observed in metro systems in China (11.3% for S. aureus and 2.5% for MRSA),8,9 railway–coach stations in China (7.6% for S. aureus and 1.6% for MRSA),8,9 and public transports in Japan (2.3% for MRSA).23 However, low MRSA contamination rate was found in toilet door handles from 136 airports in 59 countries (0.25%),10 and no MRSA isolates were recovered from public transports in London and Serbia.24,25 In addition, this study revealed significantly high S. aureus and MRSA contamination in self-service machines and luggage detectors. Thus, relevant departments need to take more stringent disinfection and surveillance measures in Guangzhou Baiyun Airport, especially for self-service machines and luggage detectors.
Linezolid is one of the few clinically effective agents against MRSA infections. Data from global surveillance studies reported that resistance to linezolid was observed very rarely among S. aureus (0.03%),26 but linezolid resistance in staphylococci has been increasing in recent years.27 It should be noted that six linezolid-resistant isolates were found in this study, which is consistent with Chinese studies in metro systems and railway–coach stations.8,9 The rate of multidrug resistance among S. aureus was high in 62.9%, which is similar to the results from metro systems (55.6%), railway–coach stations (62.1%), and busses (65.0%).8,9,28 Notably, two MRSA isolates expressed the multiresistance cfr gene, and the latest studies in China and European countries have also reported the occurrence of cfr-mediated multiresistance in staphylococci from animals and livestock-related human beings.29–31 In addition, the predominant MDR pattern in this study was co-non-susceptibility to clindamycin, erythromycin, and quinupristin–dalfopristin, which is consistent with the findings from human-related and livestock-related MRSA in China.32-36 These findings suggest that cross-transmission of MRSA between animals, human beings, and environments is possible. Therefore, both linezolid-resistant and MDR isolates must be monitored in prospective surveillance programs to assess the prevalence, resistance patterns, and resistance mechanisms.
The results of molecular features may broaden our insights into S. aureus and MRSA in the nonhospital environment and their associations with human-related isolates. In this study, the most common S. aureus CCs were CC6, CC59, CC188, CC30, CC5, CC20, CC398, and CC7, with ST59-MRSA-IV(pvl–) as the predominant MRSA. For surface samples from Chinese metro systems, the predominant STs were ST5 and ST188 for S. aureus and ST398-MRSA-II for MRSA,8 while for a study in railway and coach stations in China, the most prevalent STs were ST5 and ST72 for S. aureus and ST15-MRSA-IVa for MRSA.9 In public transports in Portugal, ST22-MRSA-IV was the most prevalent genotype for MRSA.37,38 These findings exhibit marked clonal diversity and regional difference in genotype distribution. Notably, ST59 is the most common CA-MRSA in China and other Asia-Pacific regions, and many CA-MRSA ST59 isolates have spread into hospital settings, suggesting that the burden of MRSA ST59 infection in the Asia-Pacific regions has been increasing.39-41 The ST59-MRSA-IV(pvl–) was the predominant clone found in China, Taiwan, and other Asia-Pacific countries, while both ST59-MRSA-IV(pvl–) and ST59-MRSA-V(pvl+) were the major clones observed in the clinical MRSA infections in China.42 However, both ST59-MRSA-IV(pvl–) and ST59-MRSA-V(pvl+) are characterized by potential pathogenicity based on their cytotoxicity and ability to adhere. In addition, a number of clinical studies have found that MRSA ST188/ST6/ST7 clones were associated with colonization and community-associated infection and were also associated with hospital-associated infection.39–41,43 So, the environment MRSA pollution may pose an unpredictable future risk to human beings. Future research is needed to better assess the relationship between environment MRSA pollution and the risk of human MRSA infection or colonization. Surprisingly, livestock-associated MRSA ST398 and ST5 were observed in the present study, which were rarely reported in previous studies.39–41 Therefore, we cannot rule out the possibility that livestock-associated MRSA was transmitted by human beings contacting with contaminated meats, animals, and waste releases from livestock farms.
Previous studies have used some statistical methods (such as principal component analysis and cluster analysis) to explore potential relationships among multidimensional phenotype and genotype characteristics of S. aureus. For example, the study of public transport in Japan indicated that the MRSA ST5 isolates were resistant to multiple drugs,23 and MDR ST59 and ST45 isolates were found in US beaches,44 suggesting potential relationships between CC typing and resistance phenotypes by statistic tables. A novel aspect of this study was to reveal a significant corresponding relationship between CC typing and resistant pattern of S. aureus isolates using correspondence analysis, suggesting that CC5/CC20/CC59/ST288 isolates were associated with being extended MDR and CC30 and CC672 isolates were associated with mild MDR. These findings support growing concern about MDR S. aureus in the airport characterized by high population density. Livestock-associated CC398 isolates were prevalent in livestock and related workers in European–American countries and were associated with extensively drug resistance.45,46 However, CC398 isolates in this study were associated with non-MDR, which may be due to human origin of these isolates, instead of animal origin. Notably, the tsst-1 gene encoding the toxic shock syndrome toxin TSST-1 was mainly found in CC30 isolates, which is consistent with the clinical analysis of bloodstream isolates.47 In addition, the study of food S. aureus revealed that specific genotypes (eg, CC5, CC9, CC20, CC50, and CC72) carried more toxin genes than other CCs.48 Another report also found a relationship of CC5, CC20, and CC72 with a high content of toxin genes.49 This study first used the correspondence analysis to reveal good consistency between CC typing and multiple molecular characteristics, indicating that CC6/CC7/CC30/CC59/CC188/CC398 isolates were associated with carrying IEC–hemolysin genes, and CC20 and CC45 isolates were associated with carrying IEC–enterotoxin–hemolysin genes. This study also found that the dominant clones were CC6 (sak–sea–hla–hld), CC59 (chp–sak–hla–hlb–hld), and CC188 (chp–sak–hla–hlb–hld). These findings adequately reveal the necessity of differentiating S. aureus clones based on multiple phenotype–genotype characteristics, which may provide new ideas for monitoring the latest epidemiological trends.
Although increasing epidemiological studies have focused on studying MRSA and MSSA based on antimicrobial resistance, few studies have attempted to differentiate MRSA and MSSA based on multiple molecular characteristics. Previous studies of environment-related and human-related MRSA have revealed that the MRSA isolates have a higher risk of being resistance to specific antimicrobial and being multidrug resistance than MSSA isolates,8,9,32–34 which is consistent with our results. Notably, we observed that linezolid resistance was found in MSSA isolates and similar results were also found in human research in Iran and China.32–34 These results support growing concern about the emergence of linezolid-resistant MSSA, which may have potential negative effects on human beings. The previous Iranian report of clinical isolates demonstrated that MSSA and MRSA have similar distribution patterns of enterotoxin genes (eg, sea, seb, sec, sed, and sed), tsst-1gene, and eta gene, but etb was more frequent in MRSA isolates than in MSSA isolates.32–34 In addition, there were no significant differences in enterotoxin genes (eg, sea-sep, ser, and seu), tsst-1 gene, and etd gene between clinical MSSA and MRSA isolates.50 In accordance with previous data, there is similar distribution between our MSSA and MRSA in the carriage of virulence genes, except for sek. These findings support that MSSA should be taken as seriously as MRSA due to harboring broad virulence gene spectrum.
To the best of our knowledge, this study is the first attempt to explore the multidimensional corresponding relationships between CC typing and phenotype–genotype characteristics of S. aureus isolates using the correspondence analysis. However, this study also has some limitations. First, this cross-sectional study was conducted at only one time point, so we could not determine whether the contamination rate of MRSA was persistent or transient. Second, no samples were obtained from airport workers, so it could not determine whether there exists the risk of MRSA transmission to human beings through environment contact. Third, samples were cultured on mannitol salt agar to isolate S. aureus, which might be less sensitive than selective, chromogenic agar plates. Fourth, consistent with previous studies,51,52 the present study observed some MRSA isolates with only genotypic or phenotypic resistance, suggesting that the MRSA definition based only on genotypic or phenotypic resistance may underestimate the true prevalence. Future studies should direct more attention to identify any mecA variants (such as mecC). Finally, we observed four untypeable isolates. Since the novel Staphylococcus argenteus is phenotypically similar to S. aureus, these untypeable isolates may be recognized as this novel species.53,54
Conclusion
This study revealed that 62.9% S. aureus isolates were classified as MDR, with six linezolid-resistant isolates and two cfr-carrying isolates, supporting concern about linezolid-resistant and MDR S. aureus in environmental surfaces. Notably, both phenotype and genotype characteristics reveal a potential risk of cross-transmission of MRSA between human beings and environment, suggesting more stringent contamination control measures conducted in Baiyun Airport. In addition, this study first used the correspondence analysis to reveal corresponding relationships between CC typing and phenotype–genotype characteristics of S. aureus isolates, which may provide new ideas for monitoring the latest epidemiological trends.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (No. 81602901) and the Science and Technology Planning Project of Guangdong province (No. 2014A020212306). The funders had no role in study design, data collection and analysis, and interpretation of the data.
Author contributions
All authors contributed toward data analysis, drafting and critically revising the paper, gave approval of the final version to be published and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
Laupland KB, Lyytikäinen O, Søgaard M, et al; International Bacteremia Surveillance Collaborative. The changing epidemiology of Staphylococcus aureus bloodstream infection: a multinational population-based surveillance study. Clin Microbiol Infect. 2013;19(5):465–471. | ||
Woodford N, Livermore DM. Infections caused by Gram-positive bacteria: a review of the global challenge. J Infect. 2009;59(Suppl 1):S4–S16. | ||
Chambers HF, Deleo FR. Waves of resistance: Staphylococcus aureus in the antibiotic era. Nat Rev Microbiol. 2009;7(9):629–641. | ||
DeLeo FR, Otto M, Kreiswirth BN, Chambers HF. Community-associated meticillin-resistant Staphylococcus aureus. Lancet. 2010;375(9725):1557–1568. | ||
Chuang YY, Huang YC. Molecular epidemiology of community-associated meticillin-resistant Staphylococcus aureus in Asia. Lancet Infect Dis. 2013;13(8):698–708. | ||
Popovich KJ, Weinstein RA, Hota B. Are community-associated methicillin-resistant Staphylococcus aureus (MRSA) strains replacing traditional nosocomial MRSA strains? Clin Infect Dis. 2008;46(6):787–794. | ||
Tenover FC, Tickler IA, Goering RV, et al; MRSA Consortium. Characterization of nasal and blood culture isolates of methicillin-resistant Staphylococcus aureus from patients in United States Hospitals. Antimicrob Agents Chemother. 2012;56(3):1324–1330. | ||
Peng Y, Ou Q, Lin D, et al. Metro system in Guangzhou as a hazardous reservoir of methicillin-resistant Staphylococci: findings from a point-prevalence molecular epidemiologic study. Sci Rep. 2015;5:16087. | ||
Lin JL, Peng Y, Ou QT, et al. A molecular epidemiological study of methicillin-resistant Staphylococci environmental contamination in railway stations and coach stations in Guangzhou of China. Lett Appl Microbiol. 2017;64(2):131–137. | ||
Schaumburg F, Köck R, Leendertz FH, Becker K. Airport door handles and the global spread of antimicrobial-resistant bacteria: a cross sectional study. Clin Microbiol Infect. 2016;22(12):1010–1011. | ||
Bathoorn E, Friedrich AW, Zhou K, et al. Latent introduction to the Netherlands of multiple antibiotic resistance including NDM-1 after hospitalisation in Egypt, August 2013. Euro Surveill. 2013;18(42):20610–20614. | ||
Zanger P, Nurjadi D, Schleucher R, et al. Import and spread of Panton-Valentine Leukocidin-positive Staphylococcus aureus through nasal carriage and skin infections in travelers returning from the tropics and subtropics. Clin Infect Dis. 2012;54(4):483–492. | ||
Zhang K, Sparling J, Chow BL, et al. New quadriplex PCR assay for detection of methicillin and mupirocin resistance and simultaneous discrimination of Staphylococcus aureus from coagulase-negative staphylococci. J Clin Microbiol. 2004;42(11):4947–4955. | ||
Magiorakos AP, Srinivasan A, Carey RB, et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance. Clin Microbiol Infect. 2012;18(3):268–281. | ||
Enright MC, Day NP, Davies CE, Peacock SJ, Spratt BG. Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. J Clin Microbiol. 2000;38(3):1008–1015. | ||
Zhang K, McClure JA, Elsayed S, Louie T, Conly JM. Novel multiplex PCR assay for characterization and concomitant subtyping of staphylococcal cassette chromosome mec types I to V in methicillin-resistant Staphylococcus aureus. J Clin Microbiol. 2005;43(10):5026–5033. | ||
Zhang K, McClure JA, Elsayed S, Louie T, Conly JM. Novel multiplex PCR assay for simultaneous identification of community-associated methicillin-resistant Staphylococcus aureus strains USA300 and USA400 and detection of mecA and Panton-Valentine leukocidin genes, with discrimination of Staphylococcus aureus from coagulase-negative staphylococci. J Clin Microbiol. 2008;46(3):1118–1122. | ||
van Wamel WJ, Rooijakkers SH, Ruyken M, van Kessel KP, van Strijp JA. The innate immune modulators staphylococcal complement inhibitor and chemotaxis inhibitory protein of Staphylococcus aureus are located on beta-hemolysin-converting bacteriophages. J Bacteriol. 2006;188(4):1310–1315. | ||
Jarraud S, Mougel C, Thioulouse J, et al. Relationships between Staphylococcus aureus genetic background, virulence factors, agr groups (alleles), and human disease. Infect Immun. 2002;70(2):631–641. | ||
Park JY, Fox LK, Seo KS, et al. Detection of classical and newly described staphylococcal superantigen genes in coagulase-negative staphylococci isolated from bovine intramammary infections. Vet Microbiol. 2011;147(1–2):149–154. | ||
Kehrenberg C, Schwarz S. Distribution of florfenicol resistance genes fexA and cfr among chloramphenicol-resistant Staphylococcus isolates. Antimicrob Agents Chemother. 2006;50(4):1156–1163. | ||
Ayele D, Zewotir T, Mwambi H. Multiple correspondence analysis as a tool for analysis of large health surveys in African settings. Afr Health Sci. 2014;14(4):1036–1045. | ||
Iwao Y, Yabe S, Takano T, et al. Isolation and molecular characterization of methicillin-resistant Staphylococcus aureus from public transport. Microbiol Immunol. 2012;56(1):76–82. | ||
Otter JA, French GL. Bacterial contamination on touch surfaces in the public transport system and in public areas of a hospital in London. Lett Appl Microbiol. 2009;49(6):803–805. | ||
Stepanovic´ S, Cirkovic´ I, Djukic´ S, Vukovic´ D, Svabic´-Vlahovic´ M. Public transport as a reservoir of methicillin-resistant staphylococci. Lett Appl Microbiol. 2008;47(4):339–341. | ||
Jones RN, Kohno S, Ono Y, Ross JE, Yanagihara K. ZAAPS International Surveillance Program (2007) for linezolid resistance: results from 5591 Gram-positive clinical isolates in 23 countries. Diagn Microbiol Infect Dis. 2009;64(2):191–201. | ||
Gu B, Kelesidis T, Tsiodras S, Hindler J, Humphries RM. The emerging problem of linezolid-resistant Staphylococcus. J Antimicrob Chemother. 2013;68(1):4–11. | ||
Lutz JK, van Balen J, Crawford JM, et al. Methicillin-resistant Staphylococcus aureus in public transportation vehicles (buses): another piece to the epidemiologic puzzle. Am J Infect Control. 2014;42(12):1285–1290. | ||
Cuny C, Arnold P, Hermes J, et al. Occurrence of cfr-mediated multiresistance in staphylococci from veal calves and pigs, from humans at the corresponding farms, and from veterinarians and their family members. Vet Microbiol. 2017;200:88–94. | ||
Wang Y, He T, Schwarz S, et al. Multidrug resistance gene cfr in methicillin-resistant coagulase-negative staphylococci from chickens, ducks, and pigs in China. Int J Med Microbiol. 2013;303(2):84–87. | ||
Argudín MA, Vanderhaeghen W, Vandendriessche S, et al. Antimicrobial resistance and population structure of Staphylococcus epidermidis recovered from animals and humans. Vet Microbiol. 2015;178(1-2):105–113. | ||
Imani Fooladi AA, Ashrafi E, Tazandareh SG, et al. The distribution of pathogenic and toxigenic genes among MRSA and MSSA clinical isolates. Microb Pathog. 2015;81:60–66. | ||
Rahimi F, Bouzari M, Katouli M, Pourshafie MR. Antibiotic Resistance Pattern of Methicillin Resistant and Methicillin Sensitive Staphylococcus aureus Isolates in Tehran, Iran. Jundishapur J Microbiol. 2013;6(2):144–149. | ||
Fan Y, Wang X, Li L, et al. Potential Relationship between Phenotypic and Molecular Characteristics in Revealing Livestock-Associated Staphylococcus aureus in Chinese Humans without Occupational Livestock Contact. Front Microbiol. 2016;7:1517. | ||
Ye X, Fan Y, Wang X, et al. Livestock-associated methicillin and multidrug resistant S. aureus in humans is associated with occupational pig contact, not pet contact. Sci Rep. 2016;6:19184. | ||
Wang L, Liu Y, Yang Y, et al. Multidrug-resistant clones of community-associated meticillin-resistant Staphylococcus aureus isolated from Chinese children and the resistance genes to clindamycin and mupirocin. J Med Microbiol. 2012;61(Pt 9):1240–1247. | ||
Mendes Â, Martins da Costa P, Rego D, et al. Contamination of public transports by Staphylococcus aureus and its carriage by biomedical students: point-prevalence, related risk factors and molecular characterization of methicillin-resistant strains. Public Health. 2015;129(8):1125–1131. | ||
Conceição T, Diamantino F, Coelho C, de Lencastre H, Aires-de-Sousa M. Contamination of public buses with MRSA in Lisbon, Portugal: a possible transmission route of major MRSA clones within the community. PLoS One. 2013;8(11):e77812. | ||
Huh K, Chung DR. Changing epidemiology of community-associated methicillin-resistant Staphylococcus aureus in the Asia-Pacific region. Expert Rev Anti Infect Ther. 2016;14(11):1007–1022. | ||
Song Q, Wu J, Ruan P. Predominance of community-associated sequence type 59 methicillin-resistant Staphylococcus aureus in a paediatric intensive care unit. J Med Microbiol. 2018;67(3):408–414. | ||
Wang X, Li X, Liu W, Huang W, Fu Q, Li M. Molecular Characteristic and Virulence Gene Profiles of Community-Associated Methicillin-Resistant Staphylococcus aureus Isolates from Pediatric Patients in Shanghai, China. Front Microbiol. 2016;7:1818. | ||
Li J, Wang L, Ip M, et al. Molecular and clinical characteristics of clonal complex 59 methicillin-resistant Staphylococcus aureus infections in Mainland China. PLoS One. 2013;8(8):e70602. | ||
Lee JY, Chong YP, Kim T, et al. Bone and joint infection as a predictor of community-acquired methicillin-resistant Staphylococcus aureus bacteraemia: a comparative cohort study. J Antimicrob Chemother. 2014;69(7):1966–1971. | ||
Soge OO, Meschke JS, No DB, Roberts MC. Characterization of methicillin-resistant Staphylococcus aureus and methicillin-resistant coagulase-negative Staphylococcus spp. isolated from US West Coast public marine beaches. J Antimicrob Chemother. 2009;64(6):1148–1155. | ||
Goerge T, Lorenz MB, van Alen S, et al. MRSA colonization and infection among persons with occupational livestock exposure in Europe: Prevalence, preventive options and evidence. Vet Microbiol. 2017;200:6–12. | ||
Guo D, Liu Y, Han C, Chen Z, Ye X. Phenotypic and molecular characteristics of methicillin-resistant and methicillin-susceptible Staphylococcus aureus isolated from pigs: implication for livestock-association markers and vaccine strategies. Infect Drug Resist. 2018;11:1299–1307. | ||
Liu Y, Du FL, Liu PP, et al. Molecular Epidemiology and Virulence Features of Staphylococcus aureus Bloodstream Isolates in a Regional Burn Center in China, 2012-2016. Microb Drug Resist. 2018;92(2):1–7. | ||
Song M, Bai Y, Xu J, et al. Genetic diversity and virulence potential of Staphylococcus aureus isolates from raw and processed food commodities in Shanghai. Int J Food Microbiol. 2015;195:1–8. | ||
Varshney AK, Mediavilla JR, Robiou N, et al. Diverse enterotoxin gene profiles among clonal complexes of Staphylococcus aureus isolates from the Bronx, New York. Appl Environ Microbiol. 2009;75(21):6839–6849. | ||
Arabestani MR, Rastiyani S, Alikhani MY, Mousavi SF. The Relationship Between Prevalence of Antibiotics Resistance and Virulence Factors Genes of MRSA and MSSA Strains Isolated from Clinical Samples, West Iran. Oman Med J. 2018;33(2):134–140. | ||
Petersen A, Stegger M, Heltberg O, et al. Epidemiology of methicillin-resistant Staphylococcus aureus carrying the novel mecC gene in Denmark corroborates a zoonotic reservoir with transmission to humans. Clin Microbiol Infect. 2013;19(1):E16–E22. | ||
Neyra RC, Frisancho JA, Rinsky JL, et al. Multidrug-resistant and methicillin-resistant Staphylococcus aureus (MRSA) in hog slaughter and processing plant workers and their community in North Carolina (USA). Environ Health Perspect. 2014;122(5):471–477. | ||
Tong SY, Schaumburg F, Ellington MJ, et al. Novel staphylococcal species that form part of a Staphylococcus aureus-related complex: the non-pigmented Staphylococcus argenteus sp. nov. and the non-human primate-associated Staphylococcus schweitzeri sp. nov. Int J Syst Evol Microbiol. 2015;65(Pt 1):15–22. | ||
Zhang DF, Xu X, Song Q, et al. Identification of Staphylococcus argenteus in Eastern China based on a nonribosomal peptide synthetase (NRPS) gene. Future Microbiol. 2016;11:1113–1121. |
Supplementary material
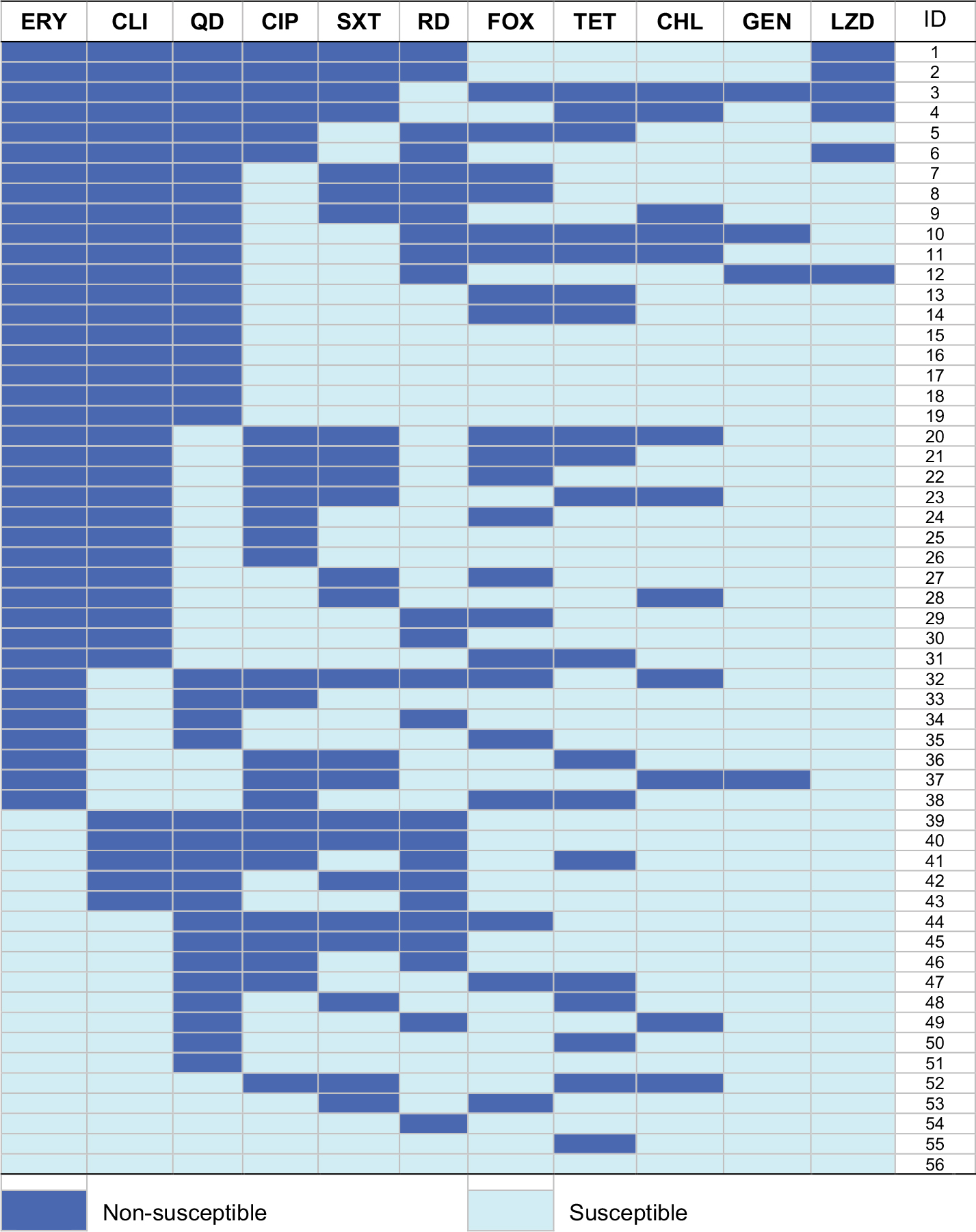 | Figure S1 Heat map showing antibiotic resistance profiles of 56 MDR S. aureus isolates. Each row represents one isolate tested for the susceptibility from an MDR S. aureus-positive participant. Abbreviations: CHL, chloramphenicol; CIP, ciprofloxacin; CLI, clindamycin; ERY, erythromycin; FOX, cefoxitin; GEN, gentamicin; LZD, linezolid; MDR, multidrug-resistant; QD, quinupristin–dalfopristin; RD, rifampicin; S. aureus, Staphylococcus aureus; SXT, trimethoprim–sulfamethoxazole; TET, tetracycline. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.