Back to Journals » OncoTargets and Therapy » Volume 12
14-Deoxycoleon U-induced endoplasmic reticulum stress-mediated apoptosis, autophagy, and cell cycle arrest in lung adenocarcinoma
Authors Peng X, Tu Y, Fu S ![]() , Xia Y, Ma C, Yang Y
, Xia Y, Ma C, Yang Y ![]() , Wu H, Liu Y, You P
, Wu H, Liu Y, You P ![]()
Received 11 April 2019
Accepted for publication 13 June 2019
Published 30 July 2019 Volume 2019:12 Pages 5955—5965
DOI https://doi.org/10.2147/OTT.S211933
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Federico Perche
Xiaozhi Peng,*,1 Yijun Tu,*,1 San Fu,2 Yu Xia,1 Chaozhi Ma,1 Yanfang Yang,1 Hezhen Wu,3 Yanwen Liu,1 Pengtao You1
1Hubei Key Laboratory of Resources and Chemistry of Chinese Medicine, Department of Pharmacy, Hubei University of Chinese Medicine, Wuhan, Hubei, 430065, People’s Republic of China; 2State Key Laboratory of Natural Medicines, Research Department of Pharmacognosy, China Pharmaceutical University, Nanjing 211198, People’s Republic of China; 3Department of Pharmacy, College of Pharmacy, Hubei University of Chinese Medicine, Wuhan, Hubei, 430065, People’s Republic of China
*These authors contributed equally to this work
Objective: 14-Deoxycoleon U is a natural abietane-type diterpene and exerts an inhibitory effect on tumor cells proliferation, which suggests that 14-Deoxycoleon U may be a potent anti-cancerous lead compound for lung cancer treatment. This study was to evaluate potential of 14-Deoxycoleon U to treat lung adenocarcinoma in vitro and in vivo.
Methods: In the present study, the cell viability and apoptosis morphology of 14-Deoxycoleon U-treated A549 and LLC cells were explored using cell counting kit-8 assay and Hoechst 33258 staining. Then, the protein expressions about apoptosis, endoplasmic reticulum (ER) stress, autophagy and cell cycle were measured using Western blot. The autophagosome formation of 14-Deoxycoleon U-treated A549 cells was visualized using a confocal microscopy. LLC lung adenocarcinoma model was established.
Results: The results indicated that 14-Deoxycoleon U significantly inhibited A549 and LLC cell proliferation in a dose-dependent manner via caspase-dependent apoptosis. Furthermore, apoptosis of both cells was mediated by 14-Deoxycoleon U-induced ER stress. In addition, 14-Deoxycoleon U-induced A549 and LLC cell autophagy, thus promoting their death. Moreover, 14-Deoxycoleon U-induced cell cycle arrest in both cells via inhibition of cyclin D3, cyclin-dependent kinase 6, CDC2 and up-regulation of p21. In vivo results showed that administration of 14-Deoxycoleon U significantly suppressed LLC growth and adverse effects of 14-Deoxycoleon U on organs might be lower than of adriamycin.
Conclusion: Overall, our results demonstrated that 14-Deoxycoleon U represses in vitro and in vivo growth of lung adenocarcinoma through ER stress-mediated apoptosis accompanied by autophagy and cell cycle arrest.
Keywords: 14-Deoxycoleon U, apoptosis, endoplasmic reticulum, autophagy, cell cycle
Introduction
As one of the top 10 malignant tumors worldwide, lung cancer which includes small cell lung cancer (SCLC) and non-small cell lung cancer (NSCLC), is associated with a highest morbidity and mortality and became a serious threat to human health.1 About 80% of lung cancer patients are diagnosed as NSCLC, and the survival rate of NSCLC patients remains approximately 15% in the last 5 years despite various progressive therapeutic anti-tumor strategies.2–4 Up to now, surgical resection is the first choice for primary tumor treatment at an initial stage, but a mere handful of patients are diagnosed in such stage.5 In NSCLC advanced stage, combination of chemotherapy and radiotherapy is still the preferred option against the malignant tumor. However, the development of pleiotropic drug resistance becomes an obstacle for a conventional drug therapy against NSCLC.6 The present therapy status indicated the inefficiency of most current treatment strategies, meaning that the discovery of novel effective compounds against NSCLC is urgently needed.
14-Deoxycoleon U is a natural abietane-type diterpene extracted from Coleus forskohlii, which has been used for relieving asthma and acute pneumonia as a folk medicine in clinical practice for many years.7,8 Aromatic abietanes as a natural diterpenoids family are characterized by the aromatic ring C with different degrees of oxygenation at several locations, Aromatic abietanes exhibit a wide variety of interesting biological activities including anti-inflammatory, hepato-protective and anti-tumor effects, becoming dominant in their role as anti-cancer drugs.9–12 In addition, a previous study reported that 14-Deoxycoleon U possesses a significant cytotoxicity effect on a human ovarian cancer cell line.13 All these evidence suggest that 14-Deoxycoleon U may be a potent anti-cancerous lead compound for lung cancer treatment. Therefore, in our current study, we evaluated the effect of this compound on lung cancer cells and our results showed that 14-Deoxycoleon U had inhibitory effects on both the lung cancer cell lines we used and it is also effective on inducing apoptosis. Thus, 14-Deoxycoleon U might be considered a novel anti-cancer natural compound.
Materials and methods
Cell culture and treatment
A549 and LLC cell lines were obtained from the China Center for Type Culture Collection (CCTCC). RMPI 1640 medium (Thermo Fisher Scientific, Waltham, MA, USA) complemented with 10% FBS (Thermo Fisher Scientific), 100 U/mL penicillin and 100 μg/mL streptomycin were used for cells culture. And, cells were incubated at 37°C under 5% CO2 and 95% humidified atmosphere. A549 and LLC cells were treated with 14-Deoxycoleon U at different concentrations (0–40 μg/mL), while untreated cells were considered as control cells. 14-Deoxycoleon U was dissolved in DMSO and stored at −20 °C as a stock solution at the concentration of 4 mg/mL, then diluted with culture medium during each treatment.
Cell viability assay
A549 and LLC cells (6×103) were seeded in 96-well plates, in 100 µL culture medium per well, for a 12 hrs incubation, then cells were treated with 0–40 μg/mL 14-Deoxycoleon U for 48 hrs. After treatment, 100 µL culture medium containing 10% cell counting kit-8 (CCK8) were replaced into each well, and cells with CCK8 were incubated at 37°C for a 4 hrs reaction. Then, OD values were measured at 450 nm by Spark 10M microplate reader (Tecan, Männedorf, Switzerland).
Hoechst 33258 staining
A549 cells (2×105 cells in 2 mL per well) were seeded in a 6-well plate and incubated overnight to allow adherence. Then, cells were treated with 14-Deoxycoleon U (0–20 μg/mL) for 12, 24 and 36 hrs. Subsequently, cells were washed 3 times with PBS (Thermo Fisher Scientific) and fixed in Carnoy’s solution for 20 mins at 4°C. Next, after 3 times washing, cells were stained with Hoechst 33258 (10 μg/mL) (Beyotime, Shanghai, China) and incubated for 15 mins at room temperature away from light. After PBS washing, nuclear morphology changes were observed under fluorescence microscope (Olympus, Tokyo, Japan).
Western blot
A549 and LLC cells were seeded in 6-well plates at the same concentration described in the previous paragraph and treated with 14-Deoxycoleon U (0–20 μg/mL) for 12, 24, and 36 hrs. After 14-Deoxycoleon U treatment, cell lysis was performed using RIPA lysis kit (Beyotime) and mixed with phenylmethanesulfonyl fluoride. Total protein quantification of the whole cell lysate was performed using BCA protein assay kit (Takara, Kyoto, Japan). The same amount of total proteins was separated on 10–12% SDS-PAGE gel electrophoresis and subsequently electrophoretically transferred onto a nitrocellulose membrane by electrophoresis (Millipore, Billerica, MA, USA). Then, the membranes were blocked in tris-buffered saline containing 0.1% Tween-20 (TBS-T) (pH 7.6) with 5% skim milk for 1 hrs, followed by an overnight incubation with primary antibodies of GAPDH, XBP1s (Proteintech Group, Wuhan, China), Bax, Bcl-2, PARP, caspase-3, cleaved-caspase-3, caspase-4, caspase-12, CHOP, Bip, IRE1-α, cdc2, cdk2, cdk6, cyclinB1, cyclind3, p21, Atg5, beclin-1, LC3 I/II [(Cell Signaling Technology, Danvers, MA, USA) (1:1,000 dilution in TBS-T)]. GAPDH was used as an internal control. Next, after 3 times washing with TBS-T, membranes were probed with horseradish peroxidase-conjugated secondary antibody (1:2,000) (Cell Signaling Technology) for 1.5 hrs at room temperature. After three times washing, blots were detected using enhanced chemiluminescence detection reagent (Bio-Rad, Hercules, CA, USA) under FluorChem FC3 system (Protein Simple, San Francisco, CA, USA), and the OD was measured using a FluorChem FC3 system (Protein Simple).
Confocal microscopy
A549 cells (5×105 cells/mL in 1 mL per well) were seeded into 6-well plates and treated with 5, 10, 20 μg/mL 14-Deoxycoleon U for 12 hrs. Then, cells were treated with 4% paraformaldehyde for a 20-min fixation, then washed with PBS and subsequently permeabilized with 0.1% Triton X-100 for 30 mins. Then, slides were washed with PBS followed by 1 hrs blocking using 5% BSA at room temperature and followed by an overnight incubation with Alexa Fluor® 488 Conjugated LC3 I/II XP® Rabbit mAb (1:400) (Cell Signaling Technology) at 4°C in the dark. Next, a 5-min incubation of DAPI (1 μg/mL) (Biosharp, Hefei, China) was used for counterstain of nuclei, and the signal was visualized using a laser scanning confocal microscopy system (Leica TCS SP2, Wetzlar, Germany).
Murine model
6-8 week C57BL/6 male mice (25±2 g) were purchased from Hubei Research Center of Laboratory Animals. All animals were fed in SPF lab of Experimental Animal Center of Hubei University of Chinese Medicine. All animal experiments were in accordance with the National Institute of Health Guide for the Care and Use of Laboratory Animals, and were approved by the Committee on the Ethics of Animal Experiments of Hubei University of Chinese Medicine. Tumors were implanted by injection of 1.5×106 cells/150 μL LLC cell suspension into the subcutaneous tissue of the right back of the mice. Mice were then randomly divided into three groups (n=10). When the tumors volume reached about 40 mm3, 5 mg/kg 14-Deoxycoleon U, 5 mg/kg adriamycin, and PBS were peritumorally injected into LLC bearing mice every day for 14 times and mice weight was recorded every day. Mice were sacrificed 24 hrs after the last drug injection. The solid tumor sizes were measured every day, and the tumor volumes and tumor growth inhibition rate were calculated by the following formula:
Tumor volume (mm3) = (length × width2)/2, where the length and width are given in mm.
Statistical analysis
Data statistics were analyzed by IBM SPSS Statistics 24.0 (IBM, Armonk, NY, USA) for statistical analysis. Results were presented as mean ± SD of 3 replicates for 3 independent experiments. A Student's t-test was performed for evaluation of statistical significance between two groups. A P<0.05 was considered statistically significant.
Results
14-Deoxycoleon U inhibited the proliferation of A549 and LLC cells
As shown in Figure 1A and B, there was a dose-dependent decrease of proliferation in 14-Deoxycoleon U treated A549 and LLC cells, viability inhibition of A549 and LLC cells was statistically evident compared to the control group (P<0.05 and P<0.01) from 2.5 μg/mL onward. The IC50 values of 14-Deoxycoleon U on A549 and LLC cells were 18.99 and 10.04 μg/mL, respectively, at 48 hrs. Indeed, cell viability of A549 treated with 20 μg/mL 14-Deoxycoleon U and LLC treated with 10 μg/mL 14-Deoxycoleon U for 48 hrs were 49.98% and 46.68%, respectively. Thus, 5, 10 and 20 μg/mL 14-Deoxycoleon U were chosen for further experiments on A549 cells and 2.5, 5 and 10 μg/mL were used on LLC cells.
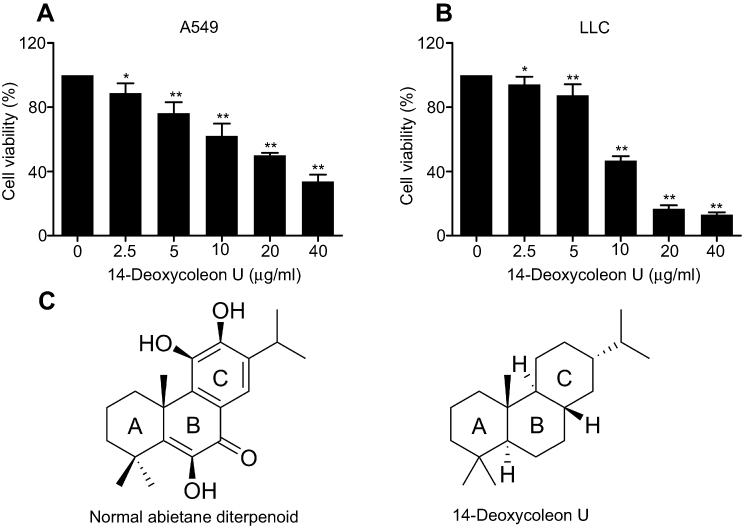 |
Figure 1 14-Deoxycoleon U inhibited the proliferation of A549 and LLC cells. A549 (A) and LLC (B) cells were treated with 14-Deoxycoleon U (0–40 μg/mL) for 48 hrs and cell viability was measured by cell counting kit-8 (CCK8) assay. (C) Structures for normal abietane diterpenoid and 14-Deoxycoleon U. *P<0.05, **P<0.01 vs control. |
14-Deoxycoleon U-induced A549 cell apoptosis
In order to further investigate whether the inhibitory effect of 14-Deoxycoleon U on proliferation was associated with apoptosis, Hoechst 33258 staining and Western blot were performed. Hoechst 33258 staining revealed that the control cells displayed the uniform rounded or elliptical nuclei with uniformly distributed chromatin, while 14-Deoxycoleon U-treated cells exhibited the typical apoptotic morphological changes, such as shrunken cell body, hyperchromatic nuclear chromatin, karyorrhexis and apoptotic bodies’ formation (Figure 2A). Apoptosis is not clearly manifested at 12 hrs, starting to appear at 36 hrs, revealing an increased apoptosis over time.
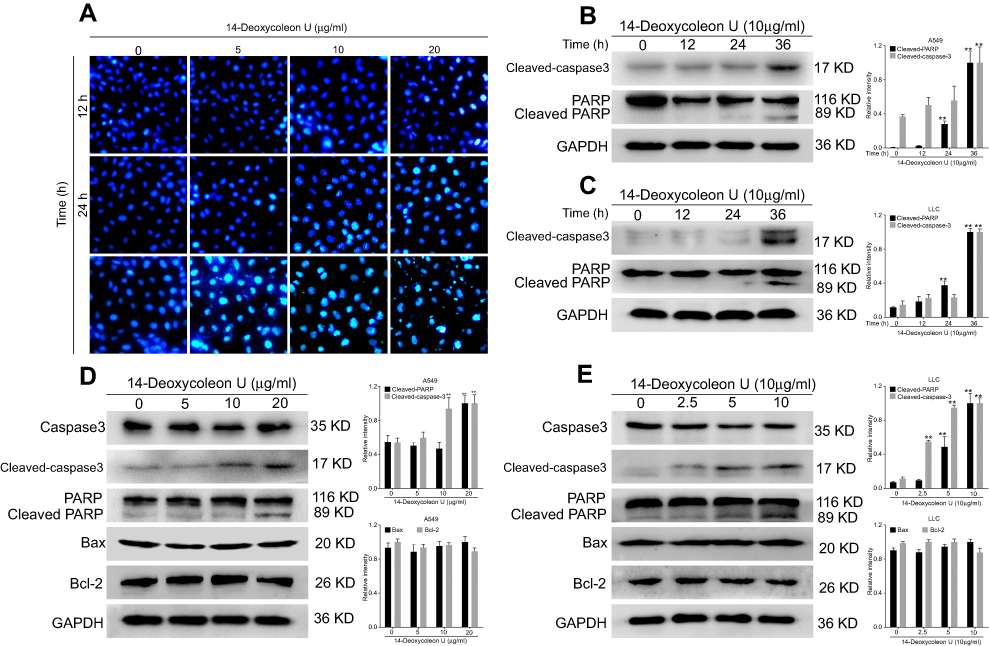 |
Figure 2 14-Deoxycoleon U-induced cell apoptosis. (A) A549 cells were treated with 0, 5, 10 and 20 μg/mL 14-Deoxycoleon U for 12, 24, 36 hrs and the nuclear morphology was detected by Hoechst 33258 staining (200× magnification). A549 and LCC cells were treated with 10 μg/mL 14-Deoxycoleon U for 12, 24, and 36 hrs (B and C, respectively) or with 0, 5, 10 and 20 μg/mL 14-Deoxycoleon U for 36 hrs (D and E, respectively) and apoptotic-related protein expression was evaluated by Western blot. **P<0.01 vs control. |
Next, apoptosis-related protein expression was evaluated by Western blot. As shown in Figure 2B–E, cleaved-caspase-3 and cleaved-PARP as the apoptotic markers were significantly increased at 36 hrs in the 14-Deoxycoleon U treated group compared to the control group (P<0.01), and this increase was dose- and time-dependent. Further investigation on Bcl-2 family proteins (Bcl-2 and Bax) which are associated with mitochondrial-mediated apoptosis was performed, revealing no difference in their expression between the control group and 14-Deoxycoleon U-treated group. These results indicated that 14-Deoxycoleon U-induced apoptosis of A549 and LLC cells probably via a non-mitochondrial pathway.
14-Deoxycoleon U-induced A549 cell apoptosis via endoplasmic reticulum (ER) stress
To further investigate apoptosis induced by 14-Deoxycoleon U, the expression ER stress related-proteins was evaluated and 12 hrs incubation time was selected according to our previous experiments (data not shown). Our results indicated that the expression of Bip, IRE1α, XBP1s and CHOP was significantly up-regulated after 12 hrs treatment with 14-Deoxycoleon U in A549 and LLC cells compared to the control group (P<0.05 and P<0.01, respectively) (Figure 3A–D). The key proteins of ER stress-mediated apoptosis, such as caspase-4 (human) and caspase-12 (mouse), were significantly decreased (P<0.01) and cleaved caspase-12 was significantly increased (P<0.01) after the treatment with 14-Deoxycoleon U in A549 and LLC cells compared to the control group (P<0.05 and P<0.01). Overall, these results indicated that 14-Deoxycoleon U-induced ER stress-mediated apoptosis in A549 and LLC cells.
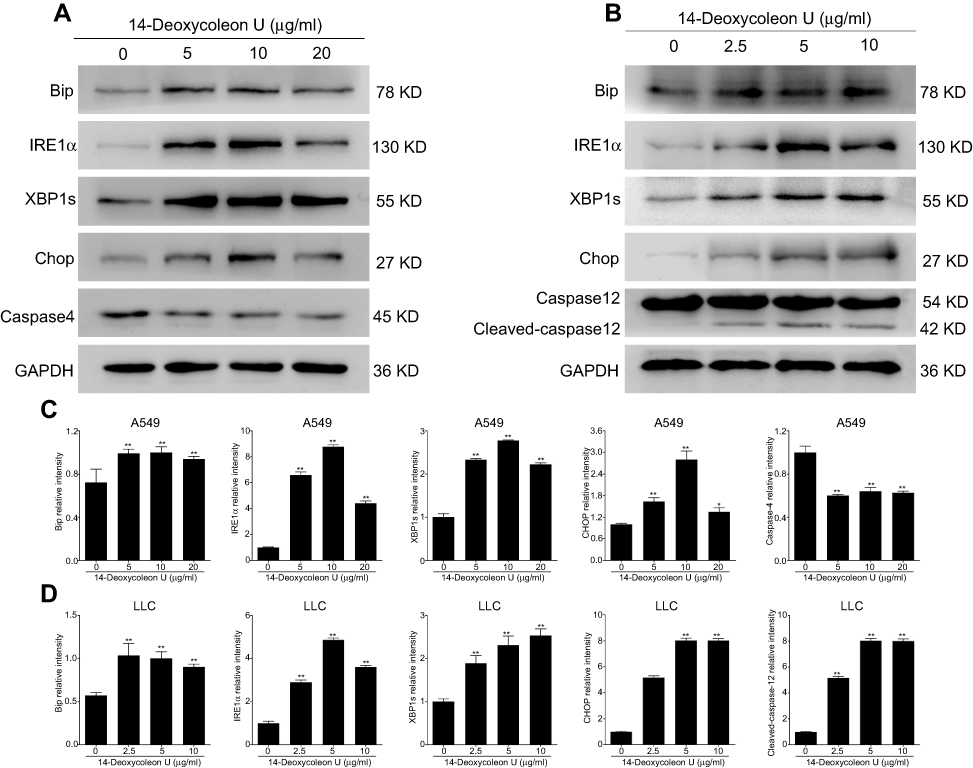 |
Figure 3 14-Deoxycoleon U-induced A549 and LCC cell apoptosis via ER stress. A549 (A) and LLC (B) cells were treated with 0, 5, 10 and 20 μg/mL 14-Deoxycoleon U for 12 hrs and ER stress-related protein expression was determined by Western blot. Quantification of these proteins is shown in (C) and (D). **P<0.01 vs control. |
14-Deoxycoleon U-induced A549 cell autophagy
Since it is reported that severe ER stress could also induce autophagy in tumor cells, the expressions of LC3, Atg5 and Beclin-1 were evaluated in both A549 and LLC cells. Our results indicated that 14-Deoxycoleon U effectively up-regulated Atg5 and LC3 II expression (both P<0.01) without modifying Beclin-1 (Figure 4A and B). Consistent with the Western blot results, the immunofluorescent staining visualized by confocal microscopy also showed the increase of LC3 II induced by 14-Deoxycoleon U (Figure 4C), suggesting that 14-Deoxycoleon U could induce the autophagy of A549 and LLC cells.14-Deoxycoleon U-induced A549 cell cycle arrest.
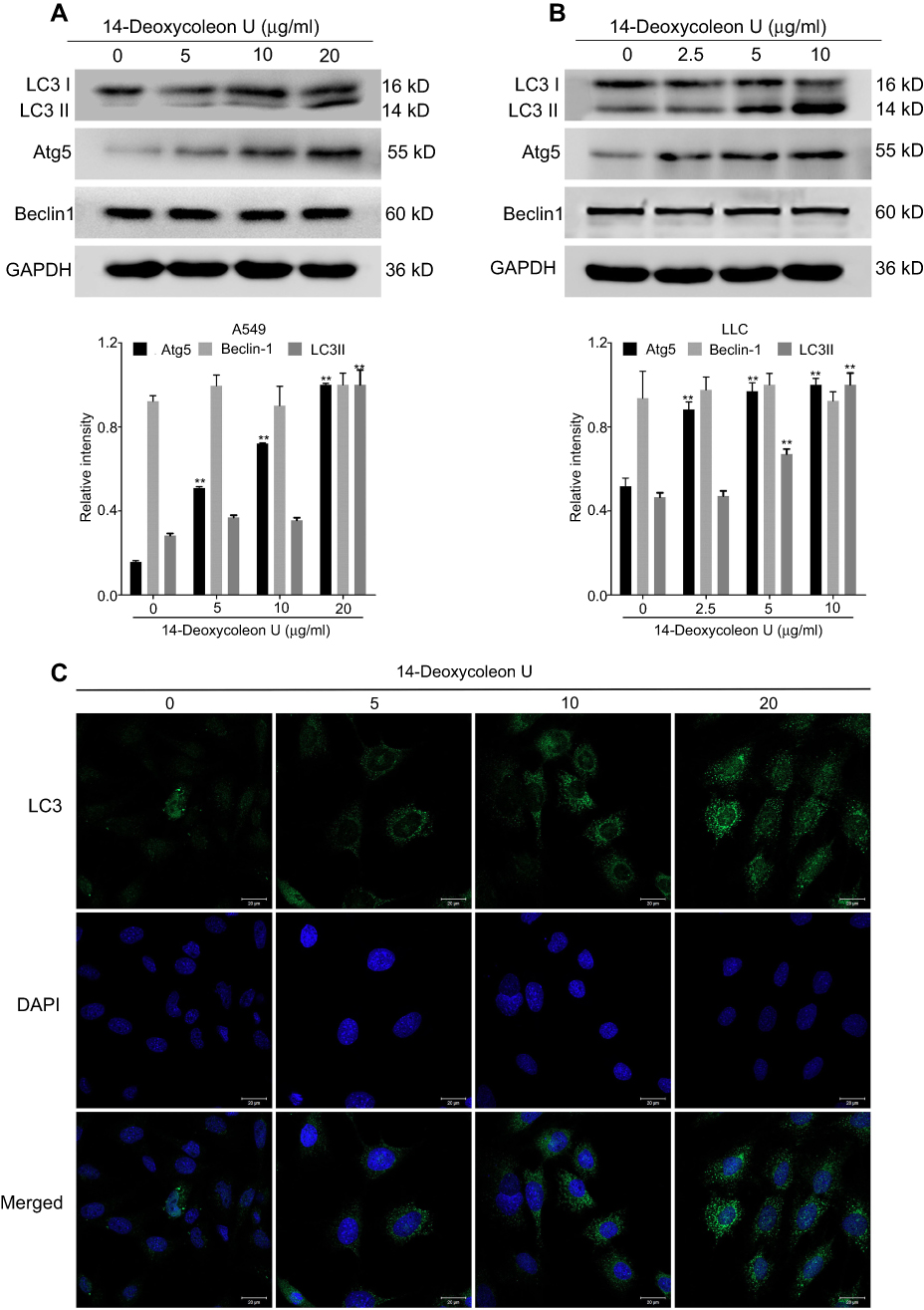 |
Figure 4 14-Deoxycoleon U-induced A549 and LCC cell autophagy. A549 (A) and LLC (B) cells were treated with 0, 5, 10 and 20 μg/mL and 0, 2.5, 5 and 10 μg/mL 14-Deoxycoleon U, respectively, for 12 hrs and autophagy-related protein expression was determined by Western blot. (C) LC3 expression and autophagosome formation were analyzed by confocal microscopy (200× magnification) in A549 cell. *P<0.05, **P<0.01 vs control. |
Since cell cycle arrest plays an important role in apoptosis, the expression of proteins involved in the cell cycle was evaluated. Our results (Figure 5A–D) showed that the expression of CDC2, cyclin-dependent kinase 6 (CDK6), and cyclin D3 was significantly decreased (P<0.01), p21 was increased (P<0.05 and P<0.01), and CDK2 and cyclin B1 expression was not significantly changed after treatment with 14-Deoxycoleon U for 12 hrs compared to the control group. Our results indicated that 14-Deoxycoleon U could induce cell cycle arrest via regulation of cell cycle-related proteins.
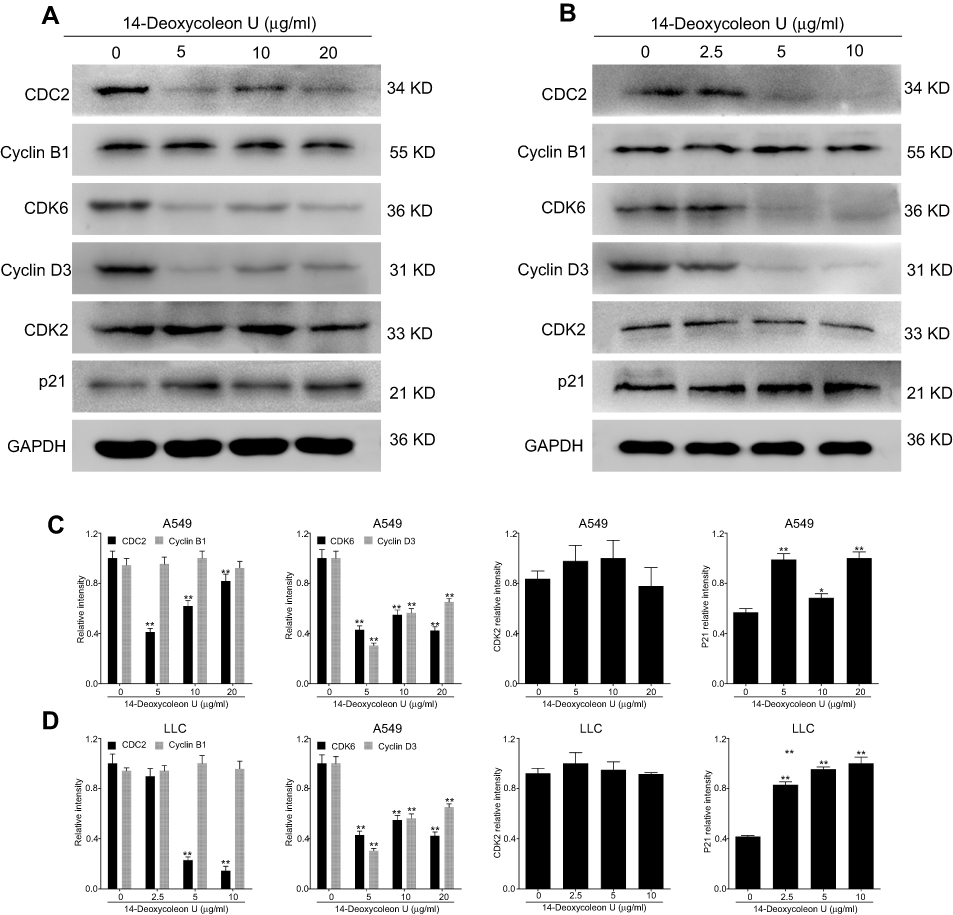 |
Figure 5 14-Deoxycoleon U-induced A549 and LCC cell cycle arrest. A549 (A) and LLC (B) cells were treated with 0, 5, 10 and 20 μg/mL and 0, 2.5, 5 and 10 μg/mL 14-Deoxycoleon U, respectively, for 12 hrs and cell cycle-related protein expression was determined by Western blot. Quantification of these proteins is shown in (C) and (D). *P<0.05, **P<0.01 vs control. |
14-Deoxycoleon U inhibited tumor growth in vivo
Tumor weights of mice treated with 14-Deoxycoleon U and adriamycin for 14 days were significantly lower compared to the control group (P<0.05 and P<0.01, respectively), while the weight of those treated with adriamycin was significantly lower than that of the 14-Deoxycoleon U group (P<0.01) (Figure 6A). The body weight of adriamycin groups was lower than the control group (P<0.01), while the weight of those treated with adriamycin was significantly lower than that of the 14-Deoxycoleon U group (P<0.01) (Figure 6B). The kidney and lung to body weight ratio of both treatment groups were similar to the control group (P>0.05) (Figure 6C and D). The liver and spleen to body weight ratio of adriamycin groups were lower than the control group (P<0.01), while the weight of those treated with adriamycin was significantly lower than that of the 14-Deoxycoleon U group (P<0.01) (Figure 6E and F).
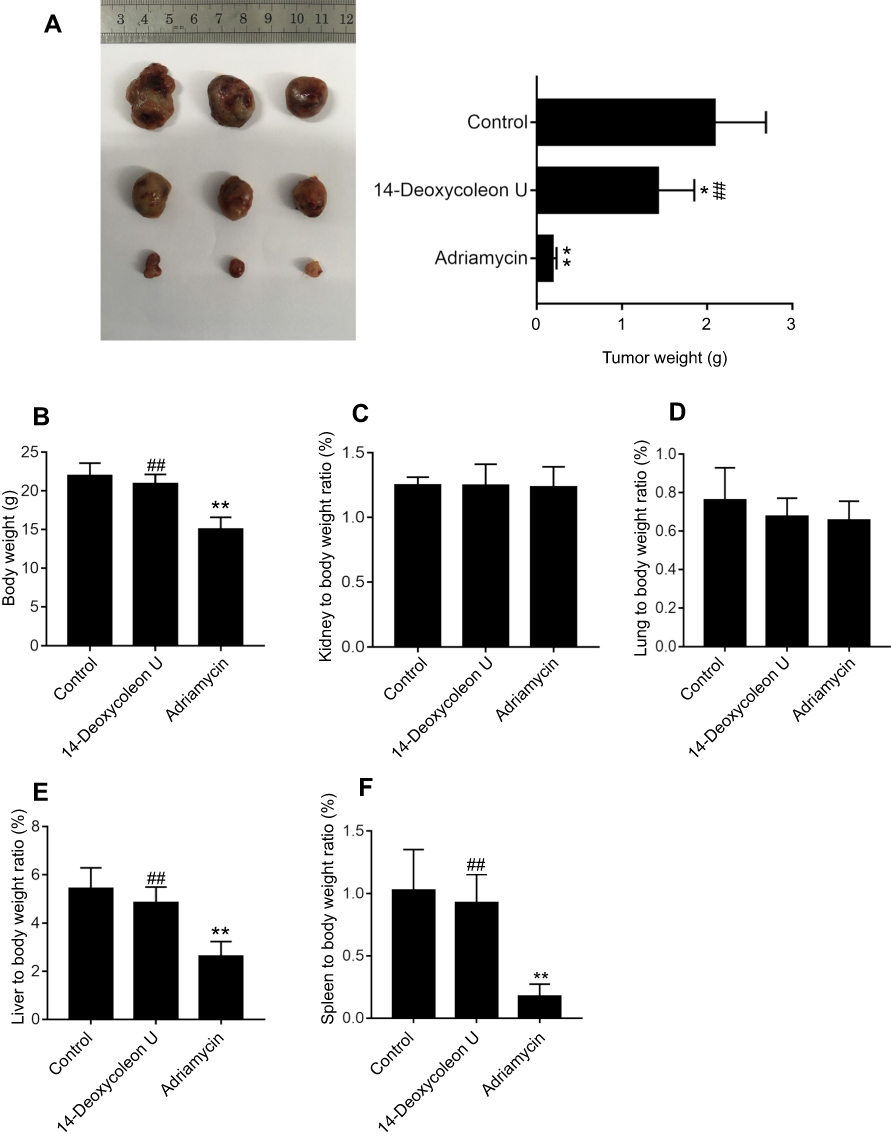 |
Figure 6 Tumor growth inhibited by 14-Deoxycoleon U. Tumor-bearing mice was sacrificed after treatment with 5 mg/kg 14-Deoxycoleon U and adriamycin for 14 days. (A) Mice tumor weight. (B) Mice body weight. (C) Kidney to body weight ratio of the mice. (D) Lung to body weight ratio of the mice. (E) Liver to body weight ratio of the mice. (F) Spleen to body weight ratio of the mice. *P<0 0.05 and **P<0 0.01 vs control, ##P<0.01 vs adriamycin. |
Discussion
Since the last few decades, natural products have always been the focus in the discovery of anti-cancer agents and show significant therapeutic efficacy against many malignancies.14–16 Abietane diterpenes display a wide variety of interesting biological activities, especially anti-proliferative activities against tumor cells.12,17,18 14-Deoxycoleon U is a natural abietane-type diterpene characterized by a cyclic structure possessing the abietane carbon framework and an aromatic ring C (I, C20) (Figure 1C) and possesses an anti-tumor activity confirmed in our previous research. Our current research indicated that 14-Deoxycoleon U exerted a significant anti-proliferative effect on two NSCLC cell lines via apoptosis.
Apoptosis mechanism is well known since it is the key mechanism of physiological cell death in vertebrates, and it is also considered the main indicator of anticancer ability. Caspase-3 is a key death executor in apoptotic pathway, which induces the cleavage of PARP, resulting in apoptosis.19,20 The increase of cleaved-caspase-3 and PARP triggered by 14-Deoxycoleon U suggested that 14-Deoxycoleon U-induced A549 and LLC cell apoptosis through the activation of caspase. However, the expression of Bcl-2 family proteins which are involved in the regulation of the mitochondrial pathway of caspase activation, such as the pro-apoptotic protein Bax and anti-apoptotic protein Bcl-2,21,22 which were not significantly changed by 14-Deoxycoleon U treatment. These results suggested that 14-Deoxycoleon U might induce A549 and LLC cell apoptosis without involving the mitochondrial pathway.
In addition to the mitochondria pathway, ER stress that induces perturbations in ER function also plays a key role in apoptosis.23 ER stress triggers the unfolded protein response, which can inhibit the protein translation, degrade misfolded proteins or generate molecular chaperones aiming to maintain ER homeostasis.23,24 The critical player Bip (also called GRP78) which acts as an ER resident chaperone for the neutralization of chronic stress was remarkably increased after 14-Deoxycoleon U treatment in A549 and LLC cells. Under the conditions of severe ER stress, the intolerably high level of Bip is unable to establish ER homeostasis, and, as a consequence of that, the oligomerization and auto-phosphorylation of IRE1α, a key transmembrane protein which acts as a variable resistance capable of regulating cell fate, occur.25,26 Because of the endoribonuclease activity, activated IRE1α splices XBP-1 mRNA to produce spliced XBP-1 (XBP1s), which binds to promoters of several genes related to ER membrane biogenesis and cell homeostasis.27 The up-regulation of XPB1s indicated that the cellular response switched from an adaptive to apoptotic. In addition, CHOP that is the downstream of IRE1, which causes changes in expression of gene involving for example DR5, which favor apoptosis, is one of the critical mediators in ER stress-induced apoptosis.28,29 Our results showed that 14-Deoxycoleon U increased IRE1α, XBP1s and CHOP expression, suggesting that ER stress was induced. On the other side, ER stress response also triggers the activation of an ER resident caspase-12 (caspase-4 possesses the same function in human cells), which translocated from ER to cytosol and activates a subsequent effector caspase mediating apoptosis.30,31 According to this mechanism, our results showed that 14-Deoxycoleon U induced the activation of ER resident caspase-12 or caspase-4, which consisted the activation of caspase-3 and resulted in cell apoptosis.
However, ER stress not only triggers apoptosis but also induces autophagy as a secondary response in the mitigation of ER stress and the degradation of accumulated proteins that were previously misfolded or unfolded.32 Moderate autophagy is considered as a survival mechanism during starvation and some other detrimental conditions, whereas prolonged autophagy disturbs cellular processes and triggers cell death. Thus, autophagy also plays a critical role as a tumor cell death mechanism in cancer chemotherapy and radiotherapy.33 Our research indicated that 14-Deoxycoleon U increased the ratio of LC3 II/LC3 I and the expression of Atg5, but without significant changes in Beclin-1 expression in both NSCLC cell lines. Newly synthesized LC3 is firstly processed to a soluble form LC3 I, but after its combination with phosphatidylethanolamine, it modifies to the lipidated and autophagosomes-associated form LC3 II because of the E3 ligase-like activity of Atg5-Atg12 complex, which is a marker for autophagy.34 Studies about cell apoptosis-autophagy crosstalk indicated that Beclin-1 is cleaved by caspases and interacts with Bcl-2 for autophagy suppression, while calpain-mediated Atg5 cleavage is associated with apoptosis, and Atg5-FADD interaction may serve to regulate apoptosis rather than autophagy.35,36 In our research, 14-Deoxycoleon U induced both cell autophagy and apoptosis, although both Bcl-2 and Beclin-1 were not significantly changed, suggesting that 14-Deoxycoleon U induce A549 and LLC cell autophagy associated with Atg5. However, the crosstalk between autophagy and apoptosis induced by 14-Deoxycoleon U requires further studies, and the mechanism of the 14-Deoxycoleon U-induced autophagy will be the focus of our future study.
The apoptosis of the two NSCLC cell lines was not visible and detectable in the early stage after 14-Deoxycoleon U treatment, but cell proliferation was significantly inhibited. Thus, cell cycle arrest in 14-Deoxycoleon U treated A549 and LLC cells was further investigated. Cell cycle and apoptosis synergistically function to maintain cell homeostasis using or controlling a shared set of factors, and failure of DNA repair during cell cycle arrest causes apoptosis.37–39 CDKs and cyclins are two group of proteins that regulate cell cycle progression by their interaction.40 14-Deoxycoleon U induced a significant degradation of cyclin D3, an ubiquitously expressed member of the cyclin D family, and its combination with CDK6 forming a CDK/cyclin complex promotes cell progression from G0 into G1 phase.41 In addition, one of the CDK inhibitory subunits such as p21 was also increased in 14-Deoxycoleon U-treated NSCLC cells, and p21 predominantly inhibits the CDKs involved in the transition of G1- to-S-phase, working as an important tumor suppressor protein.38 Thus, the down-regulation of cyclin D3 and CDK6 accompanied by the up-regulation of p21 suggested that 14-Deoxycoleon U may result in the suppression of cell progression from the G1 to the S phase of cell growth. In addition to cyclin D family, cyclin B and CDC2 play a similar role in controlling the G2 to M phase transition. In our results, CDC2 was down-regulated, but cyclin B1 expression was not modified, suggesting that 14-Deoxycoleon U could also influence cell progression from G2 to M phase by the inhibition of CDC2. However, CDK2, which is activated by cyclin E and lead to G1/S transition, was not inhibited by 14-Deoxycoleon U. Thus, 14-Deoxycoleon U might induce cell cycle arrest through the down-regulation of CDC2, CDK6, cyclin D3 and the up-regulation of p21, which probably results in apoptosis. However, further investigations are required to clarify the relationship between 14-Deoxycoleon U-induced apoptosis and cell cycle arrest.
In conclusion, the current research demonstrated that 14-Deoxycoleon U effectively inhibited lung adenocarcinoma growth and induced A549 and LLC cell apoptosis via the ER stress pathway, induced cell autophagy and cell cycle arrest, consequently inhibiting cell proliferation, which supported the development of 14-Deoxycoleon U into a potential anti-tumor compound against lung adenocarcinoma.
Acknowledgments
This study was supported by Hubei Provincial Natural Science Foundation of China (No. 2018CFB657) and China Postdoctoral Science Foundation (No. 2016M592320).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2018. CA Cancer J Clin. 2018;68(1):7–30. doi:10.3322/caac.21442
2. Soda M, Choi YL, Enomoto M, et al. Identification of the transforming EML4-ALK fusion gene in non-small-cell lung cancer. Nature. 2007;448(7153):561–566. doi:10.1038/nature05945
3. Jemal A, Bray F, Center MM, Ferlay J, Ward E, Forman D. Global cancer statistics. CA Cancer J Clin. 2011;61(2):69–90. doi:10.3322/caac.20107
4. Siegel R, Naishadham D, Jemal A. Cancer statistics, 2013. CA Cancer J Clin. 2013;63(1):11–30. doi:10.3322/caac.21166
5. Osmani L, Askin F, Gabrielson E, Li QK. Current WHO guidelines and the critical role of immunohistochemical markers in the subclassification of non-small cell lung carcinoma (NSCLC): moving from targeted therapy to immunotherapy. Semin Cancer Biol. 2018;52(Pt 1):103–109. doi:10.1016/j.semcancer.2017.11.019
6. Hensing T, Chawla A, Batra R, Salgia R. A personalized treatment for lung cancer: molecular pathways, targeted therapies, and genomic characterization. Adv Exp Med Biol. 2014;799:85–117. doi:10.1007/978-1-4614-8778-4_5
7. Nilani P, Duraisamy B, Dhamodaran P, Ravichandran S, Elango K. Effect of selected antiasthmatic plant constituents against micro organism causing upper respiratory tract infection. Anc Sci Life. 2010;29(3):30–32.
8. Ma C, Zou L, Xia Y, et al. Extracts of Coleus forskohlii relieves cough and asthma symptoms via modulating inflammation and the extracellular matrix. J CellBiochem. 2019;120(6):9648–9655. doi:10.1002/jcb.28243
9. Gonzalez MA. Aromatic abietane diterpenoids: their biological activity and synthesis. Nat Prod Rep. 2015;32(5):684–704. doi:10.1039/c4np00110a
10. Ramnath MG, Thirugnanasampandan R, Mathusalini S, Mohan PS. Hepatoprotective and cytotoxic activities of abietic acid from Isodon wightii (Bentham) H. Hara. Pharmacognosy Res. 2016;8(3):206–208. doi:10.4103/0974-8490.182920
11. Son KH, Oh HM, Choi SK, Han DC, Kwon BM. Anti-tumor abietane diterpenes from the cones of Sequoia sempervirens. Bioorg Med Chem Lett. 2005;15(8):2019–2021. doi:10.1016/j.bmcl.2005.02.057
12. Fronza M, Lamy E, Gunther S, Heinzmann B, Laufer S, Merfort I. Abietane diterpenes induce cytotoxic effects in human pancreatic cancer cell line MIA PaCa-2 through different modes of action. Phytochemistry. 2012;78:107–119. doi:10.1016/j.phytochem.2012.02.015
13. Topcu G, Turkmen Z, Schilling JK, Kingston DGI, Pezzuto JM, Ulubelen A. Cytotoxic activity of some Anatolian salvia. Extracts and isolated abietane diterpenoids. Pharm Biol. 2008;46(3):180–184. doi:10.1080/13880200701735411
14. Hsieh YS, Yang SF, Sethi G, Hu DN. Natural bioactives in cancer treatment and prevention. Biomed Res Int. 2015;2015:182835. doi:10.1155/2015/182835
15. Bishayee A, Sethi G. Bioactive natural products in cancer prevention and therapy: progress and promise. Semin Cancer Biol. 2016;40-41:1–3. doi:10.1016/j.semcancer.2016.08.006
16. Kim JY, Cho JH, Choi JR, et al. A novel anti-cancer role of beta-apopicropodophyllin against non-small cell lung cancer cells. Toxicol Appl Pharmacol. 2018;357:39–49. doi:10.1016/j.taap.2018.08.022
17. Burmistrova O, Simoes MF, Rijo P, Quintana J, Bermejo J, Estevez F. Antiproliferative activity of abietane diterpenoids against human tumor cells. J Nat Prod. 2013;76(8):1413–1423. doi:10.1021/np400172k
18. Wang X, Morris-Natschke SL, Lee KH. New developments in the chemistry and biology of the bioactive constituents of Tanshen. Med Res Rev. 2007;27(1):133–148. doi:10.1002/med.20077
19. Fatokun AA, Dawson VL, Dawson TM. Parthanatos: mitochondrial-linked mechanisms and therapeutic opportunities. Br J Pharmacol. 2014;171(8):2000–2016. doi:10.1111/bph.12416
20. Liu SY, Song JY, Fan B, et al. Resveratrol protects photoreceptors by blocking caspase- and PARP-dependent cell death pathways. Free Radic Biol Med. 2018;129:569–581. doi:10.1016/j.freeradbiomed.2018.10.431
21. Cleary ML, Smith SD, Sklar J. Cloning and structural analysis of cDNAs for bcl-2 and a hybrid bcl-2/immunoglobulin transcript resulting from the t(14;18) translocation. Cell. 1986;47(1):19–28.
22. Tsujimoto Y, Finger LR, Yunis J, Nowell PC, Croce CM. Cloning of the chromosome breakpoint of neoplastic B cells with the t(14;18) chromosome translocation. Science. 1984;226(4678):1097–1099.
23. Sano R, Reed JC. ER stress-induced cell death mechanisms. Biochim Biophys Acta. 2013;1833(12):3460–3470. doi:10.1016/j.bbamcr.2013.06.028
24. Zheng Y, Wang K, Wu Y, et al. Pinocembrin induces ER stress mediated apoptosis and suppresses autophagy in melanoma cells. Cancer Lett. 2018;431:31–42. doi:10.1016/j.canlet.2018.05.026
25. Shamu CE, Walter P. Oligomerization and phosphorylation of the Ire1p kinase during intracellular signaling from the endoplasmic reticulum to the nucleus. EMBO J. 1996;15(12):3028–3039.
26. Gardner BM, Walter P. Unfolded proteins are Ire1-activating ligands that directly induce the unfolded protein response. Science. 2011;333(6051):1891–1894. doi:10.1126/science.1209126
27. Hassler J, Cao SS, Kaufman RJ. IRE1, a double-edged sword in pre-miRNA slicing and cell death. Dev Cell. 2012;23(5):921–923. doi:10.1016/j.devcel.2012.10.025
28. Yamaguchi H, Wang HG. CHOP is involved in endoplasmic reticulum stress-induced apoptosis by enhancing DR5 expression in human carcinoma cells. J Biol Chem. 2004;279(44):45495–45502. doi:10.1074/jbc.M406933200
29. Oyadomari S, Mori M. Roles of CHOP/GADD153 in endoplasmic reticulum stress. Cell Death Differ. 2004;11(4):381–389. doi:10.1038/sj.cdd.4401373
30. Szegezdi E, Fitzgerald U, Samali A. Caspase-12 and ER-stress-mediated apoptosis: the story so far. Ann N Y Acad Sci. 2003;1010(1):186–194. doi:10.1196/annals.1299.032
31. Hetz C, Russelakis-Carneiro M, Maundrell K, Castilla J, Soto C. Caspase-12 and endoplasmic reticulum stress mediate neurotoxicity of pathological prion protein. Embo J. 2003;22(20):5435–5445. doi:10.1093/emboj/cdg537
32. Bernales S, McDonald KL, Walter P. Autophagy counterbalances endoplasmic reticulum expansion during the unfolded protein response. PLoS Biol. 2006;4(12):e423. doi:10.1371/journal.pbio.0040423
33. Schonthal AH. Endoplasmic reticulum stress and autophagy as targets for cancer therapy. Cancer Lett. 2009;275(2):163–169. doi:10.1016/j.canlet.2008.07.005
34. Tanida I, Ueno T, Kominami E. LC3 conjugation system in mammalian autophagy. Int J Biochem Cell Biol. 2004;36(12):2503–2518. doi:10.1016/j.biocel.2004.05.009
35. Pattingre S, Tassa A, Qu X, et al. Bcl-2 antiapoptotic proteins inhibit Beclin 1-dependent autophagy. Cell. 2005;122(6):927–939. doi:10.1016/j.cell.2005.07.002
36. Djavaheri-Mergny M, Maiuri MC, Kroemer G. Cross talk between apoptosis and autophagy by caspase-mediated cleavage of Beclin 1. Oncogene. 2010;29(12):1717–1719. doi:10.1038/onc.2009.519
37. Azzopardi M, Farrugia G, Balzan R. Cell-cycle involvement in autophagy and apoptosis in yeast. Mech Ageing Dev. 2017;161(Pt B):211–224. doi:10.1016/j.mad.2016.07.006
38. Karimian A, Ahmadi Y, Yousefi B. Multiple functions of p21 in cell cycle, apoptosis and transcriptional regulation after DNA damage. DNA Repair (Amst). 2016;42:63–71. doi:10.1016/j.dnarep.2016.04.008
39. Chen J. The cell-cycle arrest and apoptotic functions of p53 in tumor initiation and progression. Cold Spring Harb Perspect Med. 2016;6(3):a026104. doi:10.1101/cshperspect.a026104
40. Pucci B, Kasten M, Giordano A. Cell cycle and apoptosis. Neoplasia. 2000;2(4):291–299. doi:10.1038/sj.neo.7900101
41. Musgrove EA, Lee CS, Buckley MF, Sutherland RL. Cyclin D1 induction in breast cancer cells shortens G1 and is sufficient for cells arrested in G1 to complete the cell cycle. Proc Natl Acad Sci U S A. 1994;91(17):8022–8026. doi:10.1073/pnas.91.17.8022
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.