Back to Journals » Drug Design, Development and Therapy » Volume 17
0.5–5% Supramolecular Salicylic Acid Hydrogel is Safe for Long-Term Topical Application and Improves the Expression of Genes Related to Skin Barrier Homeostasis in Mice Models
Authors Zhou C, Hua C ![]() , Liang Q, Al Rudaisat M
, Liang Q, Al Rudaisat M ![]() , Chen S
, Chen S ![]() , Song Y, Zhu J, Cheng H
, Song Y, Zhu J, Cheng H ![]()
Received 24 November 2022
Accepted for publication 12 May 2023
Published 26 May 2023 Volume 2023:17 Pages 1593—1609
DOI https://doi.org/10.2147/DDDT.S397541
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Georgios Panos
Can Zhou,* Chunting Hua,* Qichang Liang,* Mus’ab Al Rudaisat, Siji Chen, Yinjing Song, Jiang Zhu, Hao Cheng
Department of Dermatology, Sir Run Run Shaw Hospital, School of Medicine, Zhejiang University, Hangzhou, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Hao Cheng; Jiang Zhu, Email [email protected]; [email protected]
Background: As a keratolytic, salicylic acid (SA) can be topically applied in various formulations and doses in dermatology. Supramolecular SA hydrogel, a new SA formulation with higher bioavailability, is developed and commercially available nowadays. However, there still remain concerns that the long-term and continual application of SA at low concentrations may jeopardize the cutaneous barrier properties.
Aim of the Study: To reveal the long-term effects of 0.5– 5% supramolecular SA hydrogel on the skin barrier in normal mice models.
Materials and Methods: The 0.5%, 1%, 2%, and 5% supramolecular SA hydrogel or hydrogel vehicle without SA was applied to mice’s shaved dorsal skin once per day respectively. Tissue samples of the dorsal skin were harvested on day 14 and 28 of the serial application of SA for histopathological observation and transcriptomic analysis.
Results: Following topical supramolecular SA hydrogel therapy with various concentrations of SA (0.5%, 1%, 2%, and 5%) for 14 days and 28 days, there were no obvious macroscopic signs of impaired cutaneous health and no inflammatory or degenerative abnormalities were observed in histological results. Additionally, the transcriptomic analysis revealed that on day 14, SA dramatically altered the expression of genes related to the extracellular matrix structural constituent. And on day 28, SA regulated gene expression profiles of keratinization, cornified envelope, and lipid metabolism remarkably. Furthermore, the expression of skin barrier related genes was significantly elevated after the application of SA based on RNA-seq results, and this is likely to be associated with the PPAR signaling pathway according to the enrichment analysis.
Conclusion: Our findings demonstrated that the sustained topical administration of the 0.5– 5% supramolecular SA hydrogel for up to 28 days did no harm to normal murine skin and upregulated the expression of genes related to the epidermal barrier.
Keywords: supramolecular salicylic acid hydrogel, skin barrier, PPAR signaling pathway, transcriptomic analysis
Introduction
The skin barrier, which is essential for preserving homeostasis, enables the skin to resist hazardous substances from the external environment (i.e., the outside-in barrier) and to prevent excessive loss of water and electrolytes from the body (i.e., the inside-out barrier).1 Maintaining the integrity and wellness of the body requires a well-balanced cutaneous barrier. A vicious cycle of inflammation and the emergence of skin illnesses like atopic dermatitis may arise if the barrier system is disrupted and not compensated for.2,3
In terms of function levels, the cutaneous barrier can be divided into the physical barrier, the chemical barrier, and the immune barrier.4 The physical barrier, also known as the permeability barrier, mainly consists of the stratum corneum (SC) and the system of cell-cell junctions.5 The SC architecture is usually compared to the “bricks and mortar model”.6 Corneocytes are the ‘bricks’, while the “mortar” refers to the inter-corneocyte lipid lamellae.6 Especially, the cornified envelope (CE), a highly insoluble layer of densely cross-linked proteins, replaces the plasma membrane of corneocytes.7 And cell-cell junctions, including desmosomes, adherens junctions, tight junctions, and gap junctions, are crucial for enhancing the skin’s mechanical strength.1 Furthermore, the ‘natural moisturizing factors’ (NMFs) and substances that contribute to the acidic surface pH make up the chemical barrier.5 Additionally, antimicrobial peptides (AMPs), which are produced in the epidermis to defend against invading pathogens, form an important part of the immune barrier.4
Salicylic acid (SA), a naturally occurring beta-hydroxy acid, has both cosmetic and medicinal benefits.8 With various formulations and concentrations, SA is applied topically for its keratolytic, anti-inflammatory, bacteriostatic, fungicidal, and photoprotective characteristics.8,9 However, there are some limitations in SA applications, such as poor water solubility and easy recrystallization in skin-irritating organic solvents. And it is not uncommon that certain acute side effects occur when applying SA topically, such as prolonged erythema, stinging sensation, intense exfoliation, crusting, dryness, pigmentary dyschromia, and contact sensitization.10–13 Furthermore, salicylic acid toxicity (salicylism), which is typically associated with high concentrations, takes place due to systemic absorption of SA when applied to a large area of skin.14 To enhance the bioavailability and lessen complications of SA, a novel formulation of SA, namely supramolecular SA hydrogel, is developed and commercially available nowadays. It is a kind of reversely thermo-reversible hydrogel, having an adjustable solution-to-gel transition temperature in the range of 4–45° C.15 The term “reversely thermo-reversible” means that the process of gelation takes place upon an increase in temperature rather than a decrease in temperature.15 The hydrogel form contributes to preventing the possible re-crystallization or settling of active ingredients during storage, easy and uniform spreading during application, prolonged residence time, and controlled or sustained release of active ingredients.15 Furthermore, with the aid of supramolecular technology, SA can be administered without alcohol or any other irritant solvents by selectively self-assembled into water-soluble organized entities in the form of intermolecular complexes. Under the action of skin temperature, the non-covalent bonds between molecules break so that SA can be slowly and continuously released to exert its pharmacological effects.15
Additionally, as a keratolytic, the mechanism of SA is not fully elucidated, which might be relevant to the disruption of the inter-corneocyte cohesion and the facilitation of desquamation.10,16 For superficial chemical peeling of the face, which is often performed 4–6 times at intervals of 2–4 weeks, high SA concentrations of up to 20–30% are recommended.17 Meanwhile, because of its anti-inflammatory and comedolytic effects, the long-term and continual application of 0.5–5%SA is suggested for the entire process of treating acne, including maintenance therapy to prevent relapses and the formation of scars.8,18 However, there remain concerns that the long-term use of SA may jeopardize the cutaneous barrier properties due to its capacity to weaken intercellular cohesion and remove the SC.19,20 Therefore, in our study, we assessed the safety of supramolecular SA hydrogel on normal murine skin at a concentration of up to 5% for 28 days, as well as whether and how it influenced epidermal permeability barrier homeostasis and the possible underlying mechanisms.
Materials and Methods
Animals and SA Topical Treatment
Female C57BL/6 mice (7 weeks old) were acquired from Shanghai SLAC Laboratory Animal Co., Ltd and kept in a specific pathogen-free environment at the Zhejiang University Laboratory Animal Center. All animal experiments were authorized by Zhejiang University’s Animal Care and Use Committee and carried out in accordance with their rules. The Zhejiang University School of Medicine’s Sir Run Run Shaw Hospital’s Ethics Committee gave its permission to this work (approval no 20220103–01).
Supramolecular SA hydrogel with different concentrations was obtained from Broda Company (Broda, Shanghai Rui Zhi Medicine Technology, Shanghai, China). It is composed of de-ionized water, poloxamer 407, polyethylene glycol, and salicylic acid. The hydrogel vehicle consists of de-ionized water, poloxamer 407, and polyethylene glycol.
The mice were divided into 5 groups, a hydrogel vehicle group without SA as control, and four SA-applied groups with concentrations of 0.5%, 1%, 2%, and 5%. The supramolecular SA hydrogel or vehicle was applied to mice’s shaved dorsal skin at 20 mg/cm2 once per day. Each group had six mice, three of which were executed on day 14 of the serial application of SA and the other three on day 28. Tissue samples of the dorsal skin were harvested for histopathological observation and transcriptomic analysis.
Histopathology and Immunohistochemistry Staining
The dorsal skin tissue samples from mice were fixed in formalin and dehydrated before being embedded in paraffin. Hematoxylin-eosin (H&E) or an anti-Ki67 antibody were utilized to stain paraffin sections taken from paraffin-embedded tissues after they had been deparaffinized by xylene and rehydrated through a graduated series of ethanol. Olympus Image Viewer was used to quantify the epithelial thickness after images were taken with an Olympus microscope (IX73).
RNA Extraction, Library Construction, and Sequencing
Total RNA from mice’s dorsal skin tissues was extracted utilizing TRIzol (Takara). Following the vendor-recommended protocol, LC-Bio Technology Company (Hangzhou, China) produced the library and conducted the transcriptome sequencing using NovaseqTM 6000. DESeq2 software was used to analyze gene expression differences between SA-treated and control groups. The parameters of false discovery rate (FDR) below 0.05 and absolute fold change ≥ 2 were used to identify the genes that were differentially expressed.
Short Time-Series Expression Miner (STEM) Analysis
Genes with comparable expression patterns for various groups (at least 3 in a specific time point, space, or treatment dose size order) were clustered via gene expression pattern analysis. Each sample’s expression data were standardized and then clustered by STEM program (STEM, v1.3.11) to examine the expression patterns in the order of concentration size. Each profile has a unique number that STEM assigned to it. Significant clustered profiles were those with a p-value less than 0.05.
Enrichment Analysis
Molecular function, cellular component, and biological process are the three components of Gene Ontology (GO), a worldwide standardized classification system for genes. GO terms with p < 0.05 were defined as significantly enriched GO terms in DEGs. When compared to the backdrop of the total genome, DEGs have significantly enriched metabolic or signal transduction pathways, according to the Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis. A value of p < 0.05 was considered statistically significant. Software GSEA (v4.1.0) and MSigDB were used to conduct Gene Set Enrichment Analysis (GSEA) to determine whether a set of genes in certain GO keywords or KEGG pathways exhibits appreciable differences between the two groups. GO terms and KEGG pathways that met the requirement that |NES|>1, NOM p-value<0.05, and FDR q-value<0.25 were thought to differ between the two groups.
Results
Effects of Topical SA with Different Concentrations on the Mice’s Skin Morphology
To investigate the SA’s impacts on murine skin, SA with various concentrations (0.5%, 1%, 2%, and 5%) or hydrogel vehicle without SA was administered to the back skin of mice (Figure 1A). The dorsal skin did not show erythema, scaling, or other macroscopic lesions after smearing different concentrations of SA for 14 days and 28 days (not shown). Besides, the results of hematoxylin and eosin (H&E) staining revealed that there were no discernible degenerative or inflammatory abnormalities after the application of SA as well (Figure 1B). However, 5%SA induced transient morphological alterations on day 14 that the epidermal thickness of mice dorsal skin increased compared with the vehicle group under the optic microscope. While after 28 days of exposure to 5% SA, the epidermis was restored to normal thickness (Figure 1B and C). Correspondingly, immunohistochemical analysis showed an increased level of proliferation marker Ki67 in mice treated with 5%SA for 14 days but a normal level on day 28 (Figure 1D). And the quantification of epidermal thickness preliminarily demonstrated that other concentration groups represented no increase over the normal range (Figure 1C).
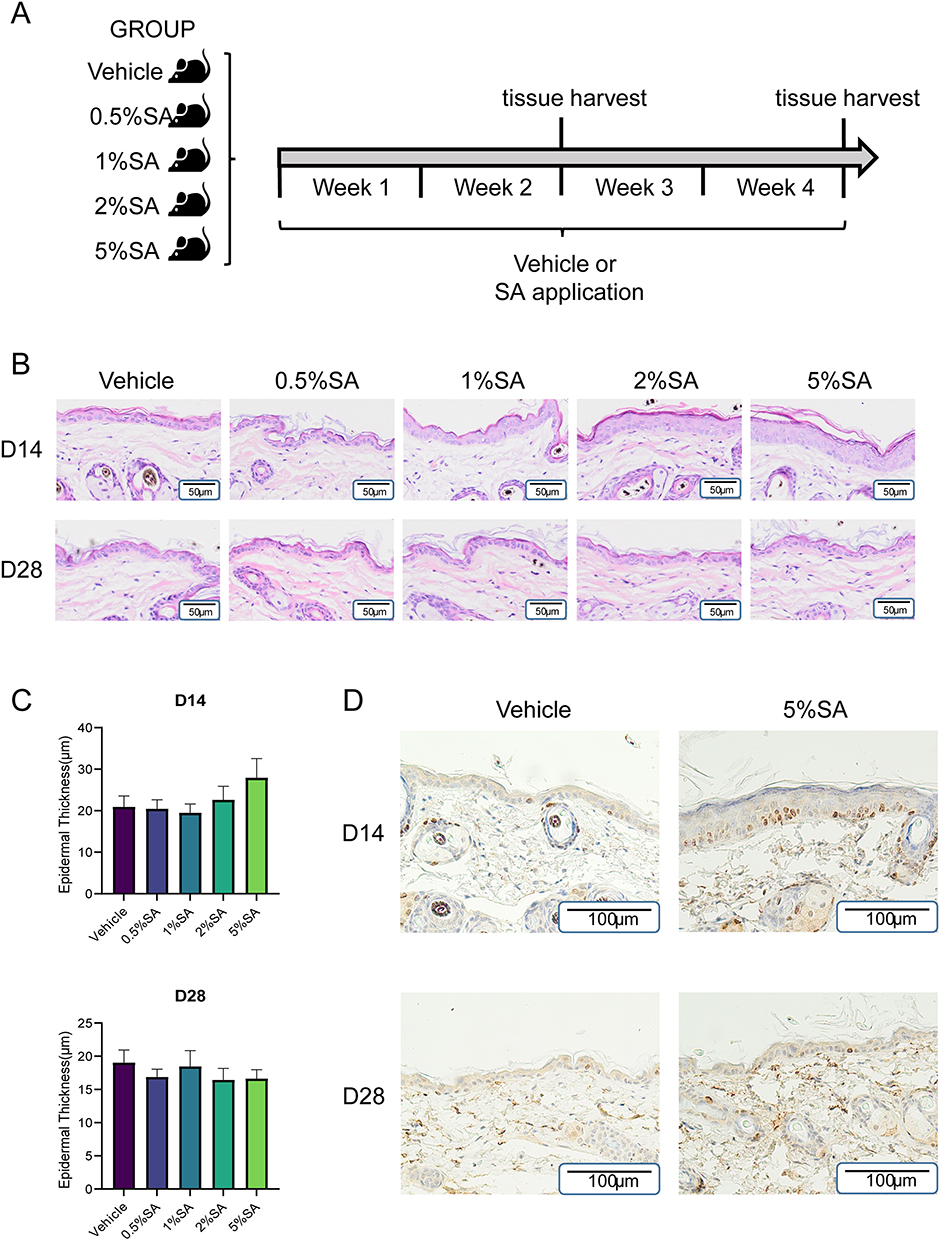 |
Figure 1 Effects of SA with different concentrations on the skin morphology of mice. (A) Protocol for evaluating the effect of SA on the dorsal skin of normal murine. The murine skin was treated with vehicle control or 0.5%, 1%, 2%, and 5% SA for 28 consecutive days. At the end of day 14 and 28, the samples were harvested and subsequently analyzed. (B) H&E staining of the back skin on day 14 and day 28. Each picture is representative of data from three different individuals. Scale Bars= 50µm. (C) Quantitative evaluation of epidermal thickness in mice skin specimens treated with SA for 14 days and 28 days. Values are expressed as the mean ± SD (n = 3). (D) Immunohistochemistry for Ki67 in dorsal skin sections obtained from the control group and 5%SA group on day 14 and 28. Abbreviations: SA, salicylic acid; H&E, hematoxylin and eosin. |
Expression Patterns Clustering of Genes Regulated by Topical SA on Day 14 and 28
To comprehend the pattern of gene expression alterations as SA concentration rises, we conducted STEM analysis on the basis of entire gene sets. Each profile had a profile number that STEM allocated to it which was represented on the upper-left corner of the profile box, and a black line inside each box symbolized the trend in gene expression variance by dosage size. The three most significant co-expression modules on day 14 together with the corresponding enriched GO terms were displayed in Figure 2A. Module 37 was the most abundant profile, in which gene expression rose after SA application compared to the control group while changing inappreciably as SA concentration fluctuated. Genes in this module were enriched for extracellular matrix (GO:0031012), cell adhesion (GO:0007155), and extracellular matrix structural constituent (GO:0005201). Module 40, which presented continuous upregulation patterns, was the second significant pattern. Genes in this module were enriched for cornified envelope (GO:0001533), keratinization (GO:0031424), and lipid metabolic process (GO:0006629) (Figure 2A and C).
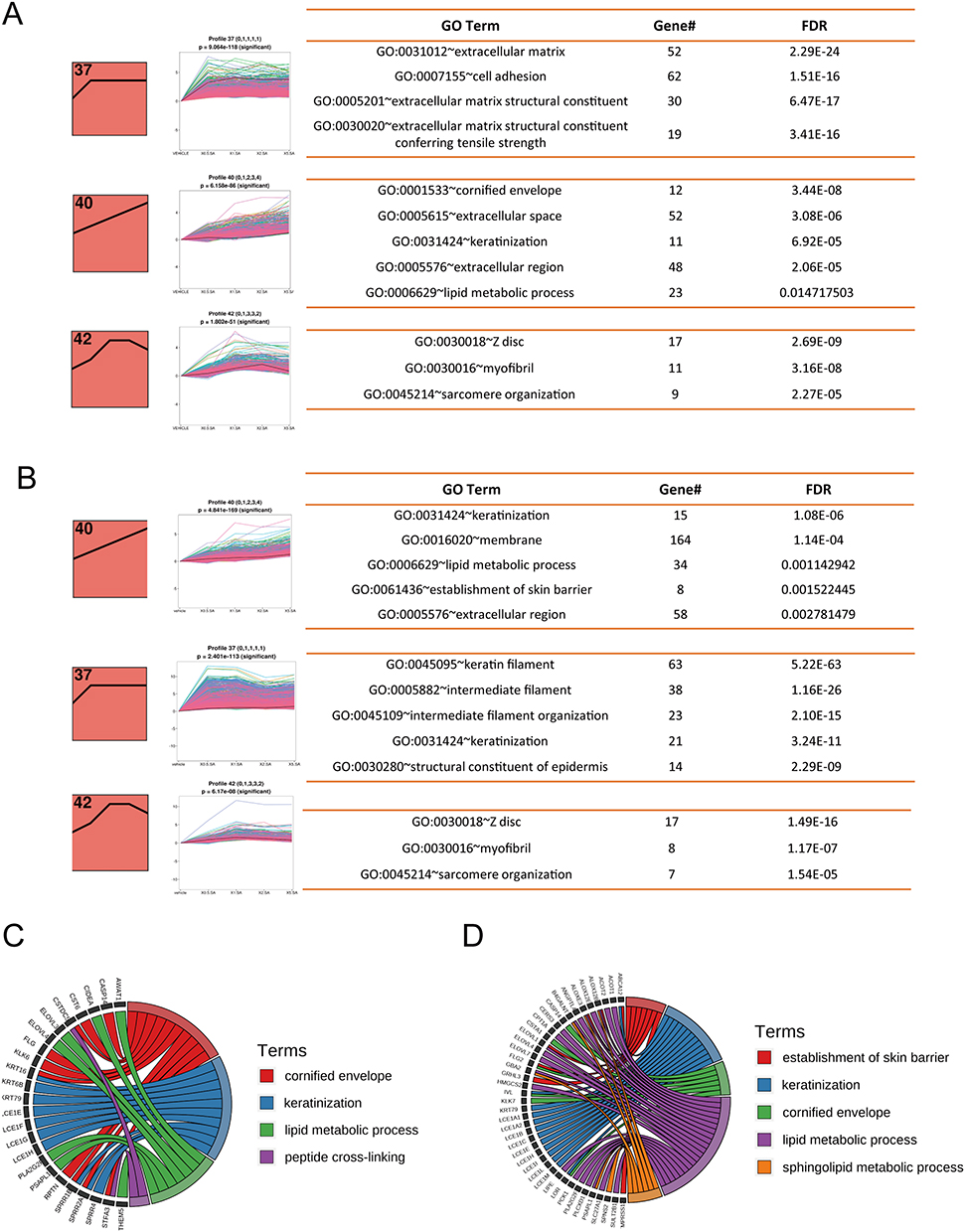 |
Figure 2 Co-expression patterns of genes regulated by SA on day 14 and 28. (A and B) The three most significant co-expression modules on day 14 (A) and day 28 (B) and their respective enriched GO terms. The profile number on the top left corner of each red box was assigned by STEM. The black lines in the red profile boxes depict the gene expression patterns in dose size order. The p-value of each profile is above the white profile box. Each line in the white box represents the expression pattern of a gene. Enriched GO terms for each module, the number of genes for the term, and the corresponding FDR (False discovery rate) are shown at the right. (C and D) Chord diagram of enriched GO terms and their genes in profile 40 on (C) day 14 and (D) day 28. Abbreviations: SA, salicylic acid; STEM, short time-series expression miner. |
The three most significant co-expression modules on day 28 were the same as the modules on day 14, and their respective enriched GO terms were displayed in Figure 2B. The most significant profile on day 28 was module 40, which was closely related to keratinization (GO:0031424), lipid metabolic process (GO:0006629), and establishment of skin barrier (GO:0061436) (Figure 2B and D). And genes in module 37 were associated with keratin filament (GO:0045095), intermediate filament (GO:0005882), and intermediate filament organization (GO:0045109) (Figure 2B). Taken together, our results revealed that the expression patterns of genes regulated by topical SA on day 14 and 28 were similar and dominated by upward trends.
Topical SA Regulated the Gene Expression Profiles of Extracellular Matrix on Day 14
Next, we looked into the gene expression profiles influenced by SA for 14 days in the mice’s dorsal skin by GO analysis. Considering the morphology of murine skin and the findings of the STEM analysis, we decided on the 5%SA group as the primary research area where alterations in the majority of gene expression levels would manifest. As shown in the heat map, 534 significant DEGs were found after 14 days of 5%SA treatment, including 461 upregulated and 73 downregulated genes (Figure 3A). A bubble plot showed 20 GO terms with the greatest significant p-value (Figure 3B). It appeared that SA had impacts on the constituent of extracellular region since most DEGs were related to extracellular region (GO:0005576), collagen-containing extracellular matrix (GO:0062023), extracellular matrix structural constituent (GO:0005201), and extracellular matrix structural constituent conferring tensile strength (GO:0030020). Additionally, lipid metabolic process (GO:0006629), keratinization (GO:0031424), cornified envelope (GO:0001533), negative regulation of endopeptidase activity (GO:0010951), and response to bacterium (GO:0009617), which all have connections with the epidermal barrier function, were enriched noticeably as well (Figure 3B). The 0.5%, 1%, and 2%SA groups showed comparable enrichment results on day 14, which altered the expression of the genes related to extracellular space as well (Supplementary Figure 1).
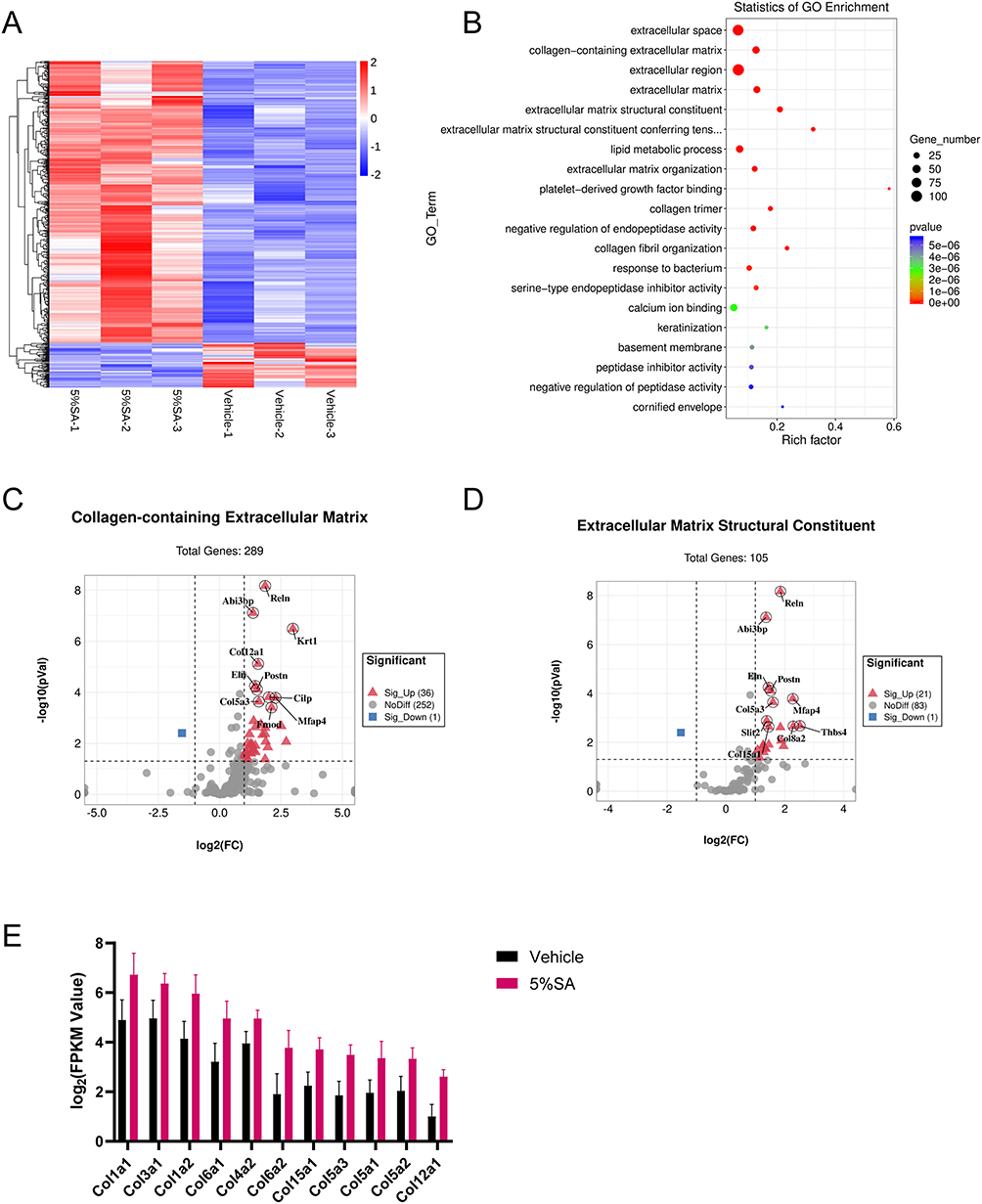 |
Figure 3 5% SA mainly regulates the gene expression profiles of extracellular matrix on day 14. (A) Heatmap of DEGs in 5%SA group (left) and control group (right) on day 14. (B) The most significant top 20 GO terms of DEGs. The dot size indicates the number of DEGs enriched in the corresponding GO term. Different p-values are indicated by dot colors. The rich factor is the ratio of the DEG number to the total number of genes in the GO term. (C and D) Volcano plots of genes included in GO terms for (C) collagen-containing extracellular matrix and (D) extracellular matrix structural constituent. (E) Representation of DEGs related to extracellular matrix structural constituent conferring tensile strength. Abbreviations: SA, salicylic acid; DEGs, differentially expressed genes; Col, collagen. |
There were 37 DEGs between the 5%SA group and control group that clustered to the collagen-containing extracellular matrix (GO:0062023) of which expression was increased for 36 genes and decreased for 1 gene (Figure 3C). Moreover, for extracellular matrix structural constituent (GO:0005201), SA significantly suppressed the expression of 1 gene and increased the expression of 21 genes (Figure 3D). Noticeably, SA upregulated genes associated with collagen fibril organization which enhances skin tensile strength. The expression of related genes was shown in Figure 3E. As a result, our findings demonstrated that SA controlled the genes which were linked to the extracellular matrix that contains collagen.
Topical SA Regulated the Gene Expression Profiles of Keratin Filament and Keratinization on Day 28
Afterwards, we examined the gene expression profiles in the mice’s dorsal skin which were applied with SA for 28 days. In 2558 genes, the 5%SA-treated mice’s gene expression profile was significantly different from the vehicle group mice’s, with almost equal amounts of up- and down-regulation. The overall dramatic difference in gene expression between the 5%SA group and the control group was visible on the heat map (Figure 4A). The top 20 GO terms were listed in the bubble plot, showing that a vast majority of DEGs were associated with extracellular region (GO:0005576), intermediate filament (GO:0005882), keratin filament (GO:0045095), keratinization (GO:0031424), and lipid metabolic process (GO:0006629) (Figure 4B). Similar enrichment results were found in the SA groups of 0.5%, 1%, and 2% on day 28, which also significantly changed the expression of genes related to keratin filament and keratinization (Supplementary Figure 2).
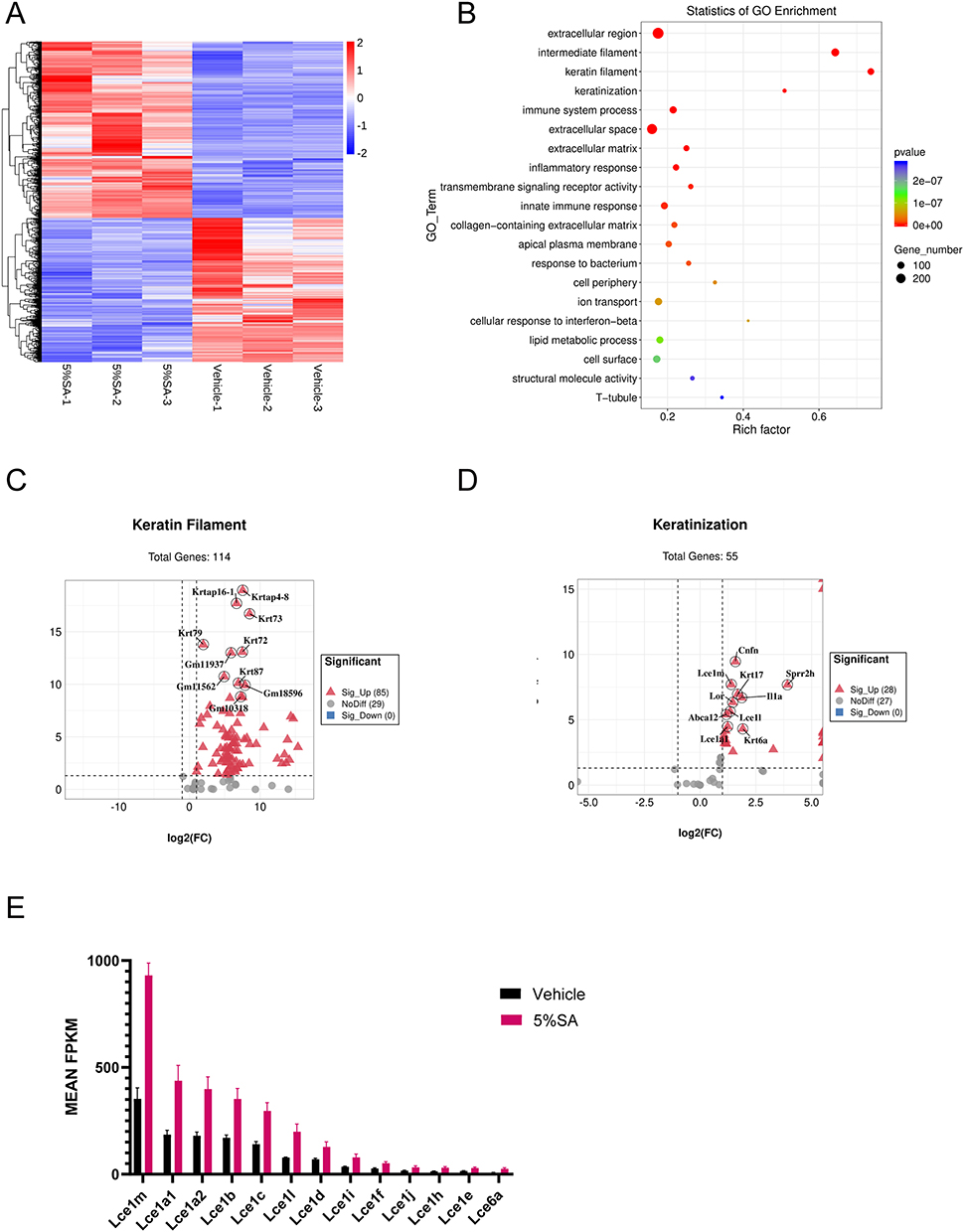 |
Figure 4 5% SA upregulates genes associated with keratin filament and keratinization on day 28. (A) Heatmap of DEGs in 5%SA group (left) and control group (right) on day 28. (B) The most significant top 20 GO terms of DEGs. The dot size indicates the number of DEGs enriched in the corresponding GO term. Different p-values are indicated by dot colors. The rich factor is the ratio of the DEG number to the total number of genes in the GO term. (C and D) Volcano plots of genes included in GO terms for (C) keratin filament and (D) keratinization. (E) Representation of DEGs related to the late cornified envelope. Abbreviations: SA, salicylic acid; DEGs, differentially expressed genes; LCE, late cornified envelope. |
All 85 DEGs which were associated with keratin filament (GO:0045095) were dramatically increased after applying SA (Figure 4C). What’s more, SA treatment also upregulated 28 DEGs which were clustered into keratinization (GO:0031424) (Figure 4D). And the top 10 genes with the highest p-value were labeled in the respective volcano plots. Notably, SA increased the expression of genes associated with the late cornified envelope which is the composition of the cornified envelope and involved in keratinocyte differentiation.21 The expression of related genes was shown in Figure 4E. Hence, our results revealed that SA influenced the expression of genes linked to keratin filament and keratinization.
Topical SA Upregulated Genes Associated with Skin Barrier
To gain further insight into the SA’s potential impacts on the skin, we selected 15 GO terms associated with the skin barrier function (e.g., keratinization, skin development, and sphingolipid biosynthetic process) to compare their proportion of associated DEGs on day 14 and 28. As illustrated in the bidirectional bar graph (Figure 5A), there were more DEGs enriched in these barrier-related GO terms on day 28, suggesting that SA may have greater effects on the epidermal barrier after 28 days of application than after 14 days. For example, as to the establishment of skin barrier (GO:0061436), there were 10 DEGs in this term on day 28 and only 3 DEGs on day 14.
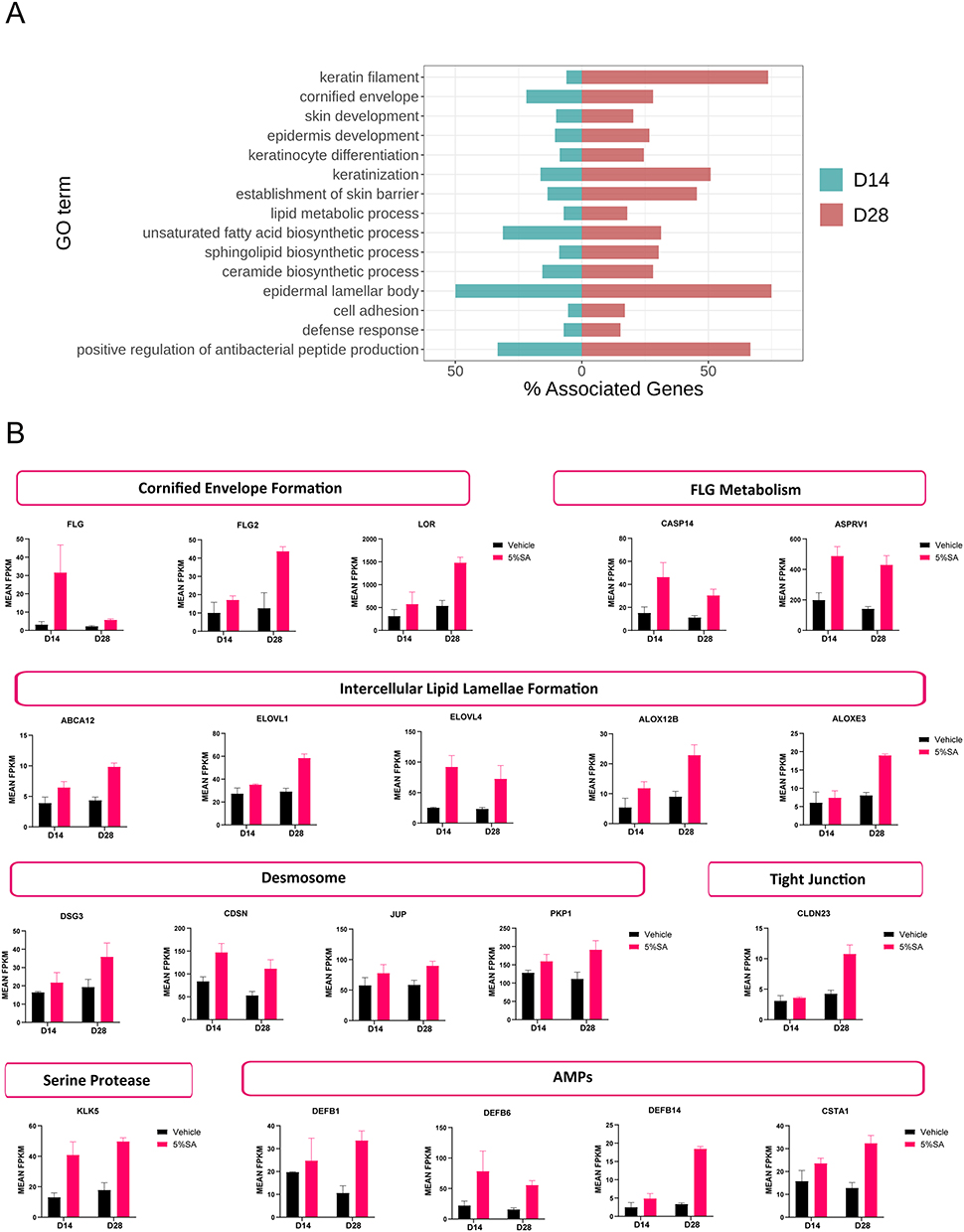 |
Figure 5 SA upregulates genes associated with the skin barrier. (A) Barrier-related significantly enriched GO terms between the 5%SA group and the control group. Green bars represent day 14, and red bars represent day 28. (B) Barrier biomarkers show greater differential expression across 5% SA vs the control group. Abbreviation: SA, salicylic acid. |
Skin barrier function is multifactorial and is dependent on a tightly controlled keratinocyte differentiation process that results in keratinization and desquamation.4 We looked at the expression of genes pertaining to the main categories of skin barrier molecules. These main categories included:
- Cornified envelope formation: filaggrin (Flg), filaggrin family member 2 (Flg2), and loricrin (Lor);
- Filaggrin metabolism: caspase 14 (Casp14) and aspartic peptidase retroviral like 1 (Asprv1);
- Intercellular lipid lamellae formation: ATP-binding cassette sub-family A member 12 (Abca12), fatty acid elongase 1 (Elovl1), fatty acid elongase 4 (Elovl4), arachidonate 12-lipoxygenase 12r type (Alox12b), and arachidonate lipoxygenase 3 (Aloxe3);
- Desmosome organization: desmoglein 3 (Dsg3), corneodesmosin (Cdsn), junction plakoglobin (Jup), and plakophilin 1 (Pkp1);
- Tight junction formation: claudin 23 (Cldn23);
- Serine protease: kallikrein 5 (Klk5);
- Antimicrobial peptides: defensin beta 1 (Defb1), defensin beta 6 (Defb6), defensin beta 14 (Defb14), and cystatin A (Csta1).
The expression levels for these genes increased in the 5%SA group versus the control group on day 14 or day 28 or both (Figure 5B). These results indicated that SA plays a significant role in affecting the epidermal barrier from diverse aspects.
Topical SA Treatment Altered the Expression of PPAR Signaling Pathway Associated Genes
To investigate the mechanisms of SA on the skin barrier in mice, we performed the GSEA analysis of KEGG pathways. On day 14, we discovered that 5%SA had the greatest impacts on the gene expression profiles of inflammatory mediator regulation of TRP channels (ko04750), aldosterone synthesis and secretion (ko04925), cGMP-PKG signaling pathway (ko04022), calcium signaling pathway (ko04020), ECM-receptor interaction (ko04512), vascular smooth muscle contraction (ko04270), biosynthesis of unsaturated fatty acids (ko01040), cortisol synthesis and secretion (ko04927), PPAR signaling pathway (ko03320), and linoleic acid metabolism (ko00591) (Figure 6A). The enriched KEGG pathways were comparable on day 14 after applications of 0.5%, 1%, 2%, and 5%SA (Supplementary Figure 3).
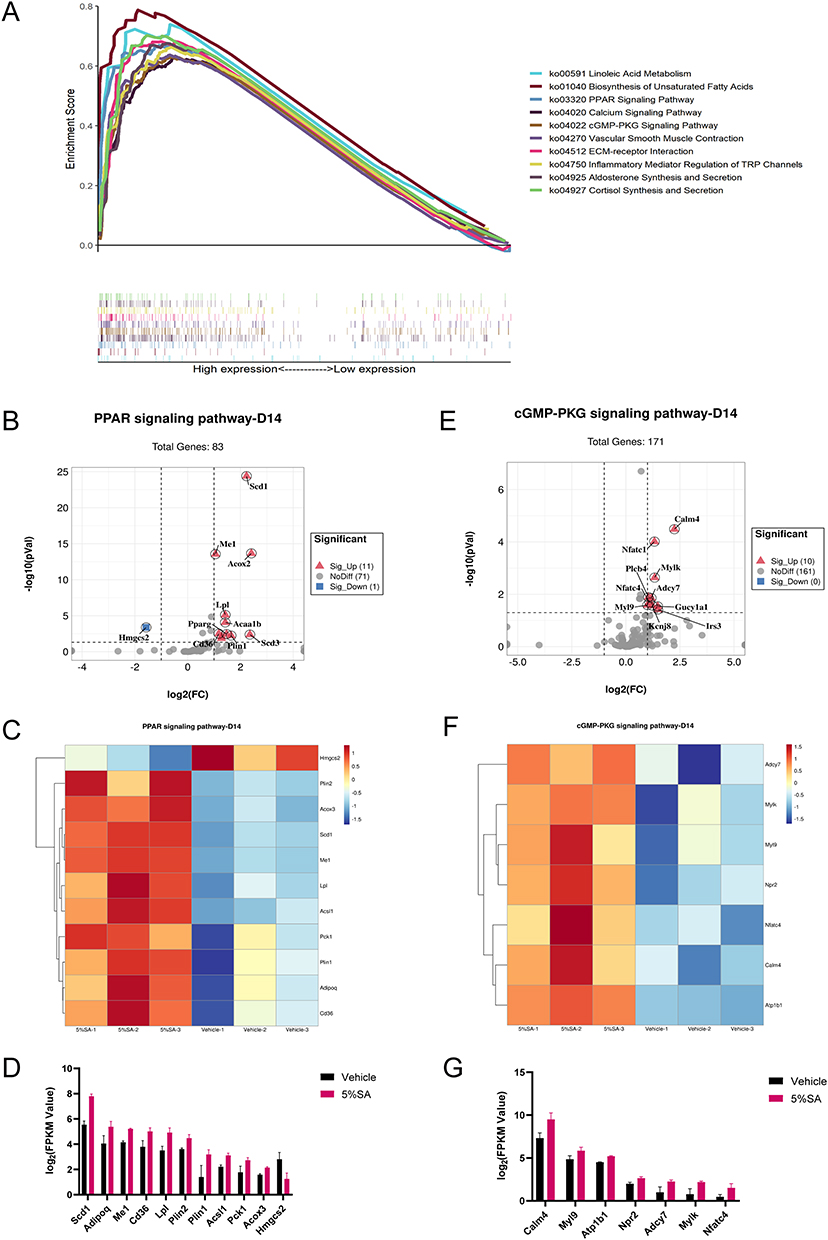 |
Figure 6 KEGG pathway analysis on day 14. (A) Multiple GSEA analysis of the KEGG pathway which achieved the top 10 highest enrichment scores. (B) The volcano plot shows the expression of PPAR signaling pathway associated genes. (C) Heatmap of PPAR signaling pathway associated genes between 5%SA and vehicle on day 14. (D) The FPKM values of genes associated with the PPAR signaling pathway. (E) The volcano plot shows the expression of cGMP-PKG signaling pathway associated genes. (F) Heatmap of cGMP-PKG signaling pathway associated genes between 5%SA and vehicle on day 14. (G) The FPKM values of genes associated with the cGMP-PKG signaling pathway. Abbreviations: SA, salicylic acid; FPKM, fragments per kilobase million. |
On day 28, the KEGG pathways with the top 10 highest enrichment scores were as follows: PPAR signaling pathway (ko03320), spliceosome (ko03040), steroid hormone biosynthesis (ko00140), axon guidance (ko04360), Notch signaling pathway (ko04330), retinol metabolism (ko00830), estrogen signaling pathway (ko04915), ribosome biogenesis in eukaryotes (ko03008), endocrine resistance (ko01522) and cell cycle (ko04110) (Figure 7A). Applications of 0.5%, 1%, and 2%SA resulted in similar enrichment results compared with the vehicle group on day 28 (Supplementary Figure 3).
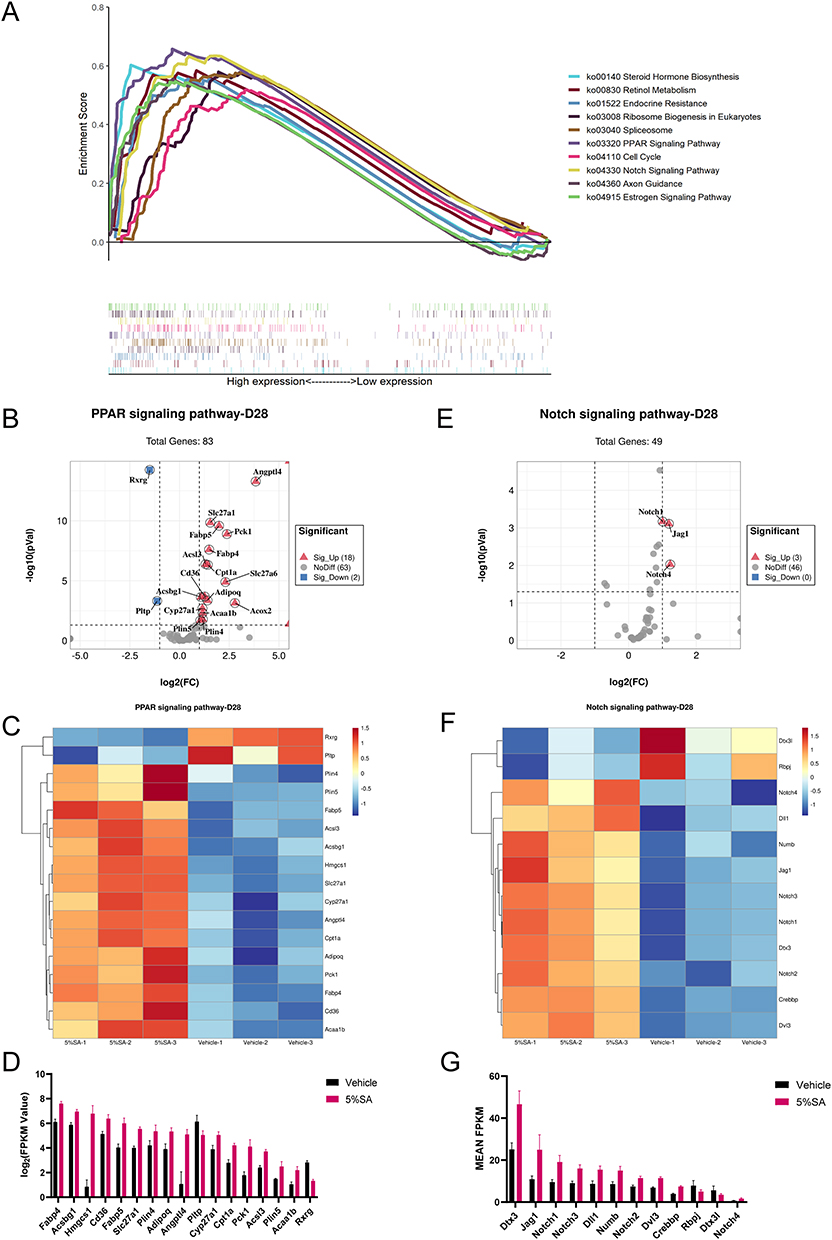 |
Figure 7 KEGG pathway analysis on day 28. (A) Multiple GSEA analysis of the KEGG pathway which achieved the top 10 highest enrichment scores. (B) The volcano plot shows the expression of PPAR signaling pathway associated genes. (C) Heatmap of PPAR signaling pathway associated genes between 5%SA and vehicle on day 28. (D) The FPKM values of genes associated with the PPAR signaling pathway. (E) The volcano plot shows the expression of NOTCH signaling pathway associated genes. (F) Heatmap of NOTCH signaling pathway associated genes between 5%SA and vehicle on day 28. (G) The FPKM values of genes associated with the NOTCH signaling pathway. Abbreviations: SA, salicylic acid; FPKM, fragments per kilobase million. |
Notably, the PPAR signaling pathway was significantly enriched following different concentrations of SA hydrogel application for 14 days and 28 days (Figure 7A and B). Peroxisome proliferator-activated receptors (PPARs) are ligand-activated nuclear receptors, whose activation is associated with the improvement of the epidermal barrier.22 There were 12 DEGs of the 5%SA group versus the control group associated with the PPAR signaling pathway on day 14, consisting of 11 upregulated genes and 1 downregulated gene (Figure 6B). According to the heatmap and the bar graph, the expression of genes related to the PPAR signaling pathway was dramatically changed by 5%SA on day 14 (Figure 6C and D). As for 28 days, there were 20 DEGs associated with the PPAR signaling pathway, including 18 upregulated genes and 2 downregulated genes (Figure 7B). And the expression of these related genes which varied between the 5%SA group and the vehicle group was shown in Figure 7C and D.
We also found that 5%SA affected the gene expression profile of the cGMP-PKG signaling pathway on day 14, including 10 upregulated DEGs (Figure 6E). The expression level of these related genes was displayed in Figure 6F and G. In addition, the Notch signaling pathway was enriched markedly on day 28, including 3 upregulated DEGs (Figure 7E). In the skin, the Notch signaling pathway is a well-established regulator of keratinocyte differentiation that promotes and maintains the differentiated state.23 The heatmap and bar graph revealed that the gene expression related to the Notch signaling pathway was significantly modulated by 5%SA on day 28 (Figure 7F and G). Taken together, our results suggested that SA may enhance the epidermal barrier homeostasis through various pathways.
Discussion
SA is a kind of lipid-soluble organic acid that is widely used in dermatology. Numerous studies have revealed that SA is effective for acne vulgaris, rosacea, seborrheic dermatitis, psoriasis,24–27 and solar-induced skin aging.28,29 SA has previously been demonstrated to disrupt intercellular cement that is covalently attached to the CE without breaking or lysing intracellular keratin filaments.16,20 This property of SA results in the desquamation of the SC, which has the potential to impair the epidermal barrier.20 Clinical investigations have already revealed that transepidermal water loss (TEWL), a measure of skin barrier impairment, either decreased or showed no statistical difference after 30% supramolecular SA chemical peeling 4 times with 2-week intervals.12,24 However, there is little research on the safety evaluation of long-term consecutive SA application with relatively low concentration and its impacts on the epidermal barrier. In our study, we discovered that following topical SA treatment with doses ranging from 0.5% to 5% for 14 days and 28 days, there were no prominent alterations in the gross appearance of murine dorsal skin. However, the epidermal thickness of the dorsal skin, as determined in the H&E sections, increased after smearing 5%SA for 14 days and returned to normal on day 28. Meanwhile, the expression of skin barrier related genes was considerably elevated after the application of SA based on RNA-seq, and this is likely to be associated with the activation of the PPAR signaling pathway according to the enrichment analysis.
In the study of Reeder et al,19 they evaluated the epidermis’s acute proliferation response to SA therapy at various doses using the hairless SKH-1 mouse. For 3.5 weeks, 4, 8, and 10% SA-treated groups showed an increase in epidermal thickness, which was accompanied by a dose-dependent rise in epidermal proliferation as determined by BrdU staining. These findings are in agreement with other studies.30,31 Other researchers, however, have not seen a rise in cell division after SA therapy.20,32 And an anti-hyperplastic effect of SA was noticed in animal models with pathological proliferation as well.33,34 The reason for this discrepancy is still not clear. In our study, the temporal epidermal thickness might be ascribed to keratinocytes proliferation irritated by 5%SA, which was verified by Ki67 staining. This indicated that there may exist transitory perturbation of the skin barrier because early research connected the exfoliation process to moderate inflammation and injury.20,35 Nonetheless, the anti-inflammatory property of SA is well-known,36 therefore, the effects of SA on epidermal proliferation still need to be further studied.
The previous study reported that incomplete expression of Flg and Ivl in UVB-irradiated mice skin was improved after the application of SA and immature CEs were replaced with mature CEs after 4 weeks of SA treatment in photo-exposed human facial skin.37 According to our findings, following the SA application, Flg, Lor, and Lce, which are all components of the CE,7 showed significantly higher expression levels compared with vehicle groups. What’s more, we discovered that there was a dose-dependent elevation in the expression of Asprv1 and Casp14, which are linked to the process of filaggrin metabolism and NMFs production.3 It is reported that mutations in Asprv1 contribute to dominantly inherited ichthyosis38 and patients with psoriasis39 and AD40 have lower expression level of Casp14. Furthermore, Geertrui et al41 found that the epidermis of caspase-14-deficient mice exhibited reduced skin-hydration levels, increased water loss, and vulnerability to UVB photodamage. Therefore, it is conceivable that SA enhances epidermal permeability homeostasis by encouraging CE formation and NMFs production. However, further analysis is still required to determine whether the induction of filaggrin-degrading enzymes occurred as a consequence of the up-modulation of filaggrin expression.
The cohesion between adjacent corneocytes, which is facilitated by intercellular junction apparatus and lipids, plays an imperative role in the epidermal defensive function.42,43 The expression of epidermal junction proteins, which support intercellular bonds to establish intact morphogenesis,44 was modulated in murine models in the presence of SA. A comparative ultrastructural study has shown that hydroxy acids targeted the central desmosomes in the SC, resulting in reduced coherence between corneocytes and excessive desquamation.45 In the current study, mRNA levels for desmosome apparatus components (Dsg3, Cdsn, Jup, Pkp1) and tight junction components (Cldn23) were prominently elevated after topical treatment of SA in comparison to controls. What’s more, mRNA levels for Klk5, which catalyzes the degradation of junction proteins and is believed to be the cascade initiator of desquamation,46 increased remarkably as well. Therefore, it is possible that SA accelerates the decomposition of epidermal junction proteins by raising Klk expression, which then likely causes an increase in the synthesis of junction components as a result of negative feedback. Besides, in the transition from the stratum granulosum to the SC, a series of lipids (ceramides, cholesterol, and free fatty acids) are generated, stored in the lamellar bodies (LB), and released into the extracellular space.42 Some of these intercellular lipids are covalently bound to the CE components and form the corneocyte lipid envelope (CLE), which contributes to the generation of an intact barrier.47 In the current study, Elovl1, Elovl4, Alox12b, Aloxe3, and Abca12, which are essential to lipid synthesis and transport,48 increased significantly following SA treatment based on RNA-seq data. Similarly, T-H. Kim et al49 found that a 2-week application of 5% α-hydroxy acid enhanced the number and secretion of the LB in the epidermis without increasing TEWL. Nevertheless, due to its potent lipophilic properties, SA can easily penetrate into the intercellular spaces of the epidermis and destroy the CLE.20,50 Therefore, it still needs to be affirmed whether the synthesis of intercellular lipids that SA induced can be formed in a proper structure.
AMPs are crucial and multifunctional components for the skin’s immunological defense against infection and damage. They take part in many different activities, including direct microbial killing, chemotaxis, altering inflammatory responses, angiogenesis, and wound healing.51 The process of AMPs induction is significantly reduced in AD patients’ lesional skin, which results in the diminished antimicrobial barrier correlated with increased susceptibility to microbial superinfections.52 Topical urea, which is known to enhance the SC hydration, has been shown to elevate AMPs expression levels together with increased barrier function determined by reduced TEWL.53 And the present study revealed that topical SA caused a marked elevation in murine epidermal β-defensins mRNA expression, suggesting that it may be conducible to enhancing the antibacterial capacity of the skin.
PPAR signaling pathway, which is involved in a variety of biological activities throughout the body, has been well demonstrated to have associations with the improvement of the skin barrier, and it is mainly mediated by promoting keratinocyte differentiation,54 epidermal lipids synthesis, and lipid trafficking processes.55,56 It is reported that topical administration of PPAR activators to normal murine skin did not affect baseline TEWL.57 While after acute disruption of the permeability barrier, recovery of permeability barrier function was hastened in animals treated with PPAR agonists topically.57,58 What’s more, 5-aminosalicylic acid (5-ASA), a derivative of SA, is a medication frequently used in the management of inflammatory bowel disease. It is confirmed that 5-ASA could exert anti-inflammatory,59 and anti-proliferation effects60 and restrict dysbiotic Escherichia coli expansion by activating PPAR signaling in the intestinal epithelium.61 As shown in our KEGG enrichment analysis results, SA with various concentrations modulated the gene expression profiles of the PPAR signaling pathway, suggesting that SA treatment may improve the epidermal barrier and serve an anti-inflammatory role by triggering the PPAR pathway.
There are still some limitations in our study, for example, protein levels and enzyme activities were not assessed. And SA was solely applied to normal animal skin, without examining the impacts on the epidermal barrier in animal models with the disturbed barrier. Additionally, the anti-inflammatory mechanisms of SA in the inflammation model as well as in clinical patients are worth future investigation.
Conclusion
In summary, we discovered that the topical application of 0.5–5%SA for 14 or 28 days did not induce damage in normal murine skin based on gross appearance and histological examination. Furthermore, the transcriptomic results revealed that the expression of skin barrier related genes was considerably increased following the application of 5%SA which may be attributed to the activation of the PPAR signaling pathway. Thus, the supramolecular SA hydrogel with a concentration of up to 5% is safe for prolonged topical administration without impairing skin barrier homeostasis in murine models and has the potential to become a preventive or therapeutic approach for the treatment of dermatoses accompanied by barrier abnormalities.
Abbreviations
SA, salicylic acid; CE, cornified envelope; SC stratum corneum; AMPs, antimicrobial peptides; NMFs, natural moisturizing factors; STEM, short time-series expression miner; GO, Gene Ontology; KEGG, Kyoto Encyclopedia of Genes and Genomes; GSEA, Gene Set Enrichment Analysis; TEWL, transepidermal water loss; PPAR, peroxisome proliferator-activated receptor.
Acknowledgments
The authors thank Broda company (Shanghai Rui Zhi Medicine Technology, Shanghai, China) for providing supramolecular salicylic acid hydrogel for this study.
Funding
This work was supported by grants from the National Natural Science Foundation of China (81573057, 81703135, and 82103743). All authors have read and approved the final manuscript.
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Natsuga K. Epidermal barriers. Cold Spring Harb Perspect Med. 2014;4(4):a018218. doi:10.1101/cshperspect.a018218
2. Lee AY. Molecular mechanism of epidermal barrier dysfunction as primary abnormalities. Int J Mol Sci. 2020;21(4):1.
3. Egawa G, Kabashima K. Barrier dysfunction in the skin allergy. Allergol Int. 2018;67(1):3–11. doi:10.1016/j.alit.2017.10.002
4. Lefevre-Utile A, Braun C, Haftek M, Aubin F. Five functional aspects of the epidermal barrier. Int J Mol Sci. 2021;22(21):11676. doi:10.3390/ijms222111676
5. Eyerich S, Eyerich K, Traidl-Hoffmann C, Biedermann T. Cutaneous barriers and skin immunity: differentiating a connected network. Trends Immunol. 2018;39(4):315–327. doi:10.1016/j.it.2018.02.004
6. Nemes Z, Steinert PM. Bricks and mortar of the epidermal barrier. Exp Mol Med. 1999;31(1):5–19. doi:10.1038/emm.1999.2
7. Candi E, Schmidt R, Melino G. The cornified envelope: a model of cell death in the skin. Nat Rev Mol Cell Biol. 2005;6(4):328–340. doi:10.1038/nrm1619
8. Arif T. Salicylic acid as a peeling agent: a comprehensive review. Clin Cosmet Investig Dermatol. 2015;8:455–461. doi:10.2147/CCID.S84765
9. Kornhauser A, Coelho SG, Hearing VJ. Applications of hydroxy acids: classification, mechanisms, and photoactivity. Clin Cosmet Investig Dermatol. 2010;3:135–142. doi:10.2147/CCID.S9042
10. Bashir SJ, Dreher F, Chew AL, et al. Cutaneous bioassay of salicylic acid as a keratolytic. Int J Pharm. 2005;292(1–2):187–194. doi:10.1016/j.ijpharm.2004.11.032
11. Sultana R, Alam M, Khondker L, Ahamed R. Safety in use of cryotherapy and topical salicylic acid with lactic acid combination in treating verruca vulgaris. Mymensingh Med J. 2012;21(4):715–722.
12. Ye D, Xue H, Huang S, et al. A prospective, randomized, split-face study of concomitant administration of low-dose oral isotretinoin with 30% salicylic acid chemical peeling for the treatment of acne vulgaris in Asian population. Int J Dermatol. 2022;61(6):698–706. doi:10.1111/ijd.16127
13. Costa IMC, Damasceno PS, Costa MC, Gomes KGP. Review in peeling complications. J Cosmet Dermatol. 2017;16(3):319–326. doi:10.1111/jocd.12329
14. Madan RK, Levitt J. A review of toxicity from topical salicylic acid preparations. J Am Acad Dermatol. 2014;70(4):788–792. doi:10.1016/j.jaad.2013.12.005
15. Lv SX, Lu J, Liu L. Reversely thermo-reversible hydrogel compositions. US Pat. 8865143. 2014 October 21.
16. Huber C, Christophers E. ”Keratolytic” effect of salicylic acid. Arch Dermatol Res. 1977;257(3):293–297. doi:10.1007/BF00741846
17. O’Connor AA, Lowe PM, Shumack S, Lim AC. Chemical peels: a review of current practice. Australas J Dermatol. 2018;59(3):171–181. doi:10.1111/ajd.12715
18. Araviiskaia E, Dréno B. The role of topical dermocosmetics in acne vulgaris. J Eur Acad Dermatol Venereol. 2016;30(6):926–935. doi:10.1111/jdv.13579
19. Sams RL, Couch LH, Miller BJ, et al. Basal cell proliferation in female SKH-1 mice treated with α- and β-hydroxy acids. Toxicol Appl Pharmacol. 2001;175(1):76–82. doi:10.1006/taap.2001.9232
20. Davies M, Marks R. Studies on the effect of salicylic acid on normal skin. Br J Dermatol. 1976;95:187–192. doi:10.1111/j.1365-2133.1976.tb00825.x
21. Marshall D, Hardman M, Nield K, Byrne C. Differentially expressed late constituents of the epidermal cornified envelope. Proc Natl Acad Sci. 2001;98(23):13031–13036. doi:10.1073/pnas.231489198
22. Man MQ, Choi EH, Schmuth M, et al. Basis for improved permeability barrier homeostasis induced by PPAR and LXR activators: liposensors stimulate lipid synthesis, lamellar body secretion, and post-secretory lipid processing. J Invest Dermatol. 2006;126(2):386–392. doi:10.1038/sj.jid.5700046
23. Nowell C, Radtke F. Cutaneous Notch signaling in health and disease. Cold Spring Harb Perspect Med. 2013;3(12):a017772. doi:10.1101/cshperspect.a017772
24. Zhang L, Shao X, Chen Y, et al. 30% supramolecular salicylic acid peels effectively treats acne vulgaris and reduces facial sebum. J Cosmet Dermatol. 2022;21:3398–3405. doi:10.1111/jocd.14799
25. Wang L, Li X, Wen X, et al. Retrospective analysis of 19 papulopustular rosacea cases treated with oral minocycline and supramolecular salicylic acid 30% chemical peels. Exp Ther Med. 2020;20(2):1048–1052. doi:10.3892/etm.2020.8740
26. Draelos ZJP. The efficacy and tolerability of turmeric and salicylic acid in psoriasis treatment. Psoriasis. 2022;12:63–71. doi:10.2147/PTT.S360448
27. Gu R, Wang SJC. Clinical study on treatment of facial seborrheic dermatitis with intense pulsed light combined with 30% supramolecular salicylic acid. Clinics. 2020;75:e1875. doi:10.6061/clinics/2020/e1875
28. Kligman D, Kligman AM. Salicylic acid peels for the treatment of photoaging. Dermatol Surg. 1998;24(3):325–328. doi:10.1111/j.1524-4725.1998.tb04162.x
29. Oresajo C, Yatskayer M, Hansenne I. Clinical tolerance and efficacy of capryloyl salicylic acid peel compared to a glycolic acid peel in subjects with fine lines/wrinkles and hyperpigmented skin. J Cosmet Dermatol. 2008;7(4):259–262. doi:10.1111/j.1473-2165.2008.00403.x
30. Abdel-Motaleb AA, Abu-Dief EE, Hussein MR. Dermal morphological changes following salicylic acid peeling and microdermabrasion. J Cosmet Dermatol. 2017;16(4):e9–e14. doi:10.1111/jocd.12315
31. Shuhei I, Setsuko U, Midori I. Histologic changes in the skin of hairless mice following peeling with salicylic acid. Arch Dermatol. 2000;136(11):1390–1395. doi:10.1001/archderm.136.11.1390
32. Roberts D, Marshall R, Marks R. Detection of the action of salicylic acid on the normal stratum corneum. Br J Dermatol. 1980;103:191–196. doi:10.1111/j.1365-2133.1980.tb06590.x
33. Weirich EG, Longauer JK, Kirkwood AH. Dermatopharmacology of salicylic acid. II. Epidermal antihyperplastic effect of salicylic acid in animals. Dermatologica. 1975;151(6):321–332. doi:10.1159/000251356
34. Isoda M, Ueda S, Imayama S, Tsukahara K. New formulation of chemical peeling agent: histological evaluation in sun-damaged skin model in hairless mice. J Dermatol Sci. 2001;27(Suppl 1):S60–S67. doi:10.1016/S0923-1811(01)00111-6
35. Park KS, Kim HJ, Kim EJ, et al. Effect of glycolic acid on UVB-induced skin damage and inflammation in Guinea pigs. Skin Pharmacol Appl Skin Physiol. 2002;15(4):236–245. doi:10.1159/000065970
36. Weirich EG, Longauer JK, Kirkwood AH. Dermatopharmacology of salicylic acid. III. Topical contra-inflammatory effect of salicylic acid and other drugs in animal experiments. Dermatologica. 1976;152(2):87–99. doi:10.1159/000251167
37. Dainichi T, Amano S, Matsunaga Y, et al. Chemical peeling by SA-PEG remodels photo-damaged skin: suppressing p53 expression and normalizing keratinocyte differentiation. J Invest Dermatol. 2006;126(2):416–421. doi:10.1038/sj.jid.5700066
38. Boyden LM, Zhou J, Hu R, et al. Mutations in ASPRV1 cause dominantly inherited ichthyosis. Am J Hum Genet. 2020;107(1):158–163. doi:10.1016/j.ajhg.2020.05.013
39. Hoste E, Denecker G, Gilbert B, et al. Caspase-14-deficient mice are more prone to the development of parakeratosis. J Invest Dermatol. 2013;133(3):742–750. doi:10.1038/jid.2012.350
40. Jung M, Choi J, Lee SA, Kim H, Hwang J, Choi EH. Pyrrolidone carboxylic acid levels or caspase-14 expression in the corneocytes of lesional skin correlates with clinical severity, skin barrier function and lesional inflammation in atopic dermatitis. J Dermatol Sci. 2014;76(3):231–239. doi:10.1016/j.jdermsci.2014.09.004
41. Denecker G, Hoste E, Gilbert B, et al. Caspase-14 protects against epidermal UVB photodamage and water loss. Nat Cell Biol. 2007;9(6):666–674. doi:10.1038/ncb1597
42. Elias PM. Stratum corneum defensive functions: an integrated view. J Invest Dermatol. 2005;125(2):183–200. doi:10.1111/j.0022-202X.2005.23668.x
43. Elias PM. Epidermal lipids, barrier function, and desquamation. J Invest Dermatol. 1983;80(1 Suppl):44s–49s. doi:10.1038/jid.1983.12
44. Sumigray KD, Lechler T. Cell adhesion in epidermal development and barrier formation. Curr Top Dev Biol. 2015;112:383–414.
45. Corcuff P, Fiat F, Minondo A, Lévèque J, Rougier A. A comparative ultrastructural study of hydroxyacids induced desquamation. Eur J Dermatol. 2002;12(4):XXXIX–XLIII.
46. Rawlings AV, Voegeli R. Stratum corneum proteases and dry skin conditions. Cell Tissue Res. 2013;351(2):217–235. doi:10.1007/s00441-012-1501-x
47. Jia Y, Gan Y, He C, Chen Z, Zhou C. The mechanism of skin lipids influencing skin status. J Dermatol Sci. 2018;89(2):112–119. doi:10.1016/j.jdermsci.2017.11.006
48. Kruse V, Neess D, Faergeman NJ. The significance of epidermal lipid metabolism in whole-body physiology. Trends Endocrinol Metab. 2017;28(9):669–683. doi:10.1016/j.tem.2017.06.001
49. Kim TH, Choi EH, Kang YC, Lee SH, Ahn SK. The effects of topical α-hydroxyacids on the normal skin barrier of hairless mice. Br J Dermatol. 2001;144:267–273. doi:10.1046/j.1365-2133.2001.04011.x
50. Lazo ND, Meine JG, Downing DT. Lipids are covalently attached to rigid corneocyte protein envelopes existing predominantly as beta-sheets: a solid-state nuclear magnetic resonance study. J Invest Dermatol. 1995;105(2):296–300. doi:10.1111/1523-1747.ep12318985
51. Clausen ML, Agner T. Antimicrobial peptides, infections and the skin barrier. Curr Probl Dermatol. 2016;49:38–46.
52. Ong PY, Ohtake T, Brandt C, et al. Endogenous antimicrobial peptides and skin infections in atopic dermatitis. N Engl J Med. 2002;347(15):1151–1160. doi:10.1056/NEJMoa021481
53. Grether-Beck S, Felsner I, Brenden H, et al. Urea uptake enhances barrier function and antimicrobial defense in humans by regulating epidermal gene expression. J Invest Dermatol. 2012;132(6):1561–1572. doi:10.1038/jid.2012.42
54. Schmuth M, Haqq CM, Cairns WJ, et al. Peroxisome proliferator-activated receptor (PPAR)-beta/delta stimulates differentiation and lipid accumulation in keratinocytes. J Invest Dermatol. 2004;122(4):971–983. doi:10.1111/j.0022-202X.2004.22412.x
55. Rivier M, Castiel I, Safonova I, Ailhaud G, Michel S. Peroxisome proliferator-activated receptor-alpha enhances lipid metabolism in a skin equivalent model. J Invest Dermatol. 2000;114(4):681–687. doi:10.1046/j.1523-1747.2000.00939.x
56. Wallmeyer L, Lehnen D, Eger N, et al. Stimulation of PPARalpha normalizes the skin lipid ratio and improves the skin barrier of normal and filaggrin deficient reconstructed skin. J Dermatol Sci. 2015;80(2):102–110. doi:10.1016/j.jdermsci.2015.09.012
57. Kömüves LG, Hanley K, Lefebvre AM, et al. Stimulation of PPARalpha promotes epidermal keratinocyte differentiation in vivo. J Invest Dermatol. 2000;115(3):353–360. doi:10.1046/j.1523-1747.2000.00073.x
58. Mao-Qiang M, Fowler AJ, Schmuth M, et al. Peroxisome-proliferator-activated receptor (PPAR)-gamma activation stimulates keratinocyte differentiation. J Invest Dermatol. 2004;123(2):305–312. doi:10.1111/j.0022-202X.2004.23235.x
59. Rousseaux C, Lefebvre B, Dubuquoy L, et al. Intestinal antiinflammatory effect of 5-aminosalicylic acid is dependent on peroxisome proliferator-activated receptor-gamma. J Exp Med. 2005;201(8):1205–1215. doi:10.1084/jem.20041948
60. Rousseaux C, El-Jamal N, Fumery M, et al. The 5-aminosalicylic acid antineoplastic effect in the intestine is mediated by PPARgamma. Carcinogenesis. 2013;34(11):2580–2586. doi:10.1093/carcin/bgt245
61. Cevallos SA, Lee JY, Velazquez EM, et al. 5-aminosalicylic acid ameliorates colitis and checks dysbiotic Escherichia coli expansion by activating PPAR-gamma signaling in the intestinal epithelium. mBio. 2021;12(1). doi:10.1128/mBio.03227-20
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.