Back to Journals » Drug Design, Development and Therapy » Volume 20
Untargeted Metabolomics Investigating the Intracellular Metabolic Alterations of HaCaT Cells Treated with Isotretinoin
Authors Zhu S, He Y, Wu L ![]() , Ouyang X
, Ouyang X ![]() , Wei Q, Ye F
, Wei Q, Ye F ![]() , Li C
, Li C ![]()
Received 30 November 2025
Accepted for publication 11 February 2026
Published 18 February 2026 Volume 2026:20 585422
DOI https://doi.org/10.2147/DDDT.S585422
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Anastasios Lymperopoulos
Shengcai Zhu,1 Yang He,1 Liang Wu,1 Xiaoliang Ouyang,2 Quan Wei,1 Fan Ye,1 Chunming Li1
1Department of Dermatology, The Second Affiliated Hospital, Jiangxi Medical College, Nanchang University, Nanchang, Jiangxi, People’s Republic of China; 2Department of Plastic Surgery, The Second Affiliated Hospital, Jiangxi Medical College, Nanchang University, Nanchang, Jiangxi, People’s Republic of China
Correspondence: Chunming Li, Department of Dermatology, The Second Affiliated Hospital, Jiangxi Medical College, Nanchang University, Nanchang, Jiangxi, People’s Republic of China, Email [email protected]
Background: Isotretinoin has been increasingly utilized for various dermatological diseases.However, the mechanism of action of isotretinoin on the skin, particularly the epidermis, remains unclear.
Purpose: This study aimed to investigate the intracellular metabolic alterations induced by isotretinoin in HaCaT cells.
Methods: HaCaT cells were divided into three groups: Group A (control group), Group B(treated with 10 μM isotretinoin for 48 h), and Group C (treated with 10 μM isotretinoin for 7 d). An untargeted metabolomics analysis was employed using ultra-high performance liquid chromatography coupled with high-resolution mass spectrometry (UHPLC-Q-TOF/MS). Multivariate data analyses were employed to identify distinguishing metabolites, and the Kyoto Encyclopedia of Genes and Genomes (KEGG) was utilized to assess potential metabolic pathways.
Results: Isotretinoin inhibited the proliferation of HaCaT cells in a dose-dependent manner. The OPLS-DA model displayed clear segmentation between the Group B and Group A, as well as between Group C and Group A. Compared to Group A, there were 115 significantly differentially metabolites in Group B, including glycerophosphocholine,adenosine,acetylcarnitine,creatine,Dl-lactate,hypoxanthine,uracil,LPC 18:1,creatine phosphate,5′-S-Methyl-5′-thioadenosine. In Group C, there were 140 significantly differentially expressed metabolites compared to Group A, including glycerophosphocholine, niacinamide, creatine, phytosphingosine, oleic acid, hypoxanthine, 1-Palmitoyl-sn-glycero-3-phosphocholine, Dl-lactate, 5′-S-Methyl-5′-thioadenosine and NG,NG-Dimethyl-L-arginine.A total of 30 metabolic pathways were significantly changed (p < 0.05) in Group A and Group B.The most significant metabolic pathways included purine metabolism,nucleotide metabolism,ABC transporters, and the taurine and hypotaurine metabolism signaling pathway. Additionally, 46 metabolic pathways were significantly altered (p < 0.05) in Group A and Group C, with the most significant pathways being nucleotide metabolism, ABC transporters, purine metabolism, and glycerophospholipid metabolism.
Conclusion: Important metabolites and metabolic pathways were identified in this study,which will help clarify the underlying mechanism of action of isotretinoin on keratinocytes at the metabolite level and the mucocutaneous adverse effects associated with isotretinoin in the treatment of dermatological diseases.
Keywords: Isotretinoin, metabolomics, HaCaT cells, purine metabolism
Introduction
Isotretinoin (13-cis-retinoic acid) is a first-generation non-aromatic retinoid derived from β-carotene (provitamin A). This compound was first synthesized in 1955; however, it was not until 1973 that it was utilized for the treatment of psoriasis in Europe. Subsequently, in 1982, isotretinoin received approval for the treatment of acne. Beyond its well-established role in acne management, isotretinoin has increasingly been used off-label for various dermatological diseases, including rosacea,1 seborrheic dermatitis,2 periorificial dermatitis,3 granuloma annulare,4 and sarcoidosis.5 Despite its clinical efficacy, isotretinoin is associated with a range of adverse reactions including skin symptoms (dry skin, erythema, and cheilosis), teratogenic effects,6 mood alterations,7 hepatotoxicity,8 and dyslipidemia.9
The impact of isotretinoin on the skin is responsible for various mucocutaneous adverse effects. However, the mechanisms by which isotretinoin acts on the skin, particularly the epidermis, remain poorly understood. A previous study found that the dryness of the skin observed in patients with acne treated with isotretinoin is related to an increase in transepidermal water loss (TEWL) and a decrease in the hydration of the epidermal stratum corneum.10 Furthermore, a subsequent study indicated that the skin dryness caused by isotretinoin might be associated with its effects on aquaporin 3 (AQP3) expression, a water and glycerol channel protein.11 Moreover, Tadini et al revealed that the topical application of isotretinoin on hairless mice epidermis leads to an enhancement in epidermis thickness and the number of epidermal cell layers.12 However, in vitro studies show that isotretinoin inhibits the proliferation of HaCaT keratinocyte cells and decreases their survival.13,14 Additionally, isotretinoin enhances the expression of Forkhead box protein O1 (FoxO1) in human primary keratinocytes (HPKs), promoting differentiation and apoptosis in these cells.15
Metabolomics is the qualitative and quantitative analysis of all endogenous low-molecular-weight metabolites (< 1.5 kDa) in the biological organism at a given time.Presently, several research techniques are commonly employed in metabolomics, such as ultra-high performance liquid chromatography coupled with time-of-flight mass spectrometry (UHPLC-Q-TOF/MS), liquid chromatography-mass spectrometry (LC-MS), gas chromatography-mass spectrometry (GC-MS), and nuclear magnetic resonance (NMR), and UHPLC-Q-TOF/MS has become a powerful tool for metabolomics research.16 In addition, non-targeted analysis has a wide application in drug research to reveal the potential efficacy and mechanisms of action of various drugs.17–19
To date, no study has investigated the intracellular metabolic alterations in keratinocyte cells treated with isotretinoin. Although HaCaT cells are spontaneously immortalized through p53 mutation, this cell line closely resembles normal keratinocytes, making it a valuable experimental model. In this study, we employed a non-targeted metabolomics strategy utilizing UHPLC-Q-TOF/MS to investigate the intracellular metabolic alterations in HaCaT cells treated with isotretinoin, thereby providing a comprehensive perspective on the effects of isotretinoin on keratinocytes at the metabolomic level.
Materials and Methods
Cell Culture
A spontaneously immortalized skin keratinocyte cell line HaCaT (10 to 30 passage numbers), was obtained from iCell Bioscience Inc (Shanghai, China) and cultured in high glucose DMEM supplemented with 10% fetal bovine serum (FBS) and penicillin–streptomycin (Boster, China). The cells were maintained under standard cell culture conditions at 37°C, 5% CO2 in a humid environment.
CCK8 Assay
Cell proliferation was evaluated using cell counting kit-8 (CCK8) (Solarbio, China) in accordance with the manufacturer’s instructions. Briefly, HaCaT cells were seeded at a density of 1×104 per well (100 μL) in 96-well plates. After 24h incubation, the cells were treated with isotretinoin (MedChem Express,USA) at various concentrations (0, 1, 5, 10μM) for 48 h. Subsequently, 10 μL of CCK-8 was added to each well, and cells were incubated for 2 h at 37 °C. The absorbance value at 450 nm was read by the enzyme-labeled instrument. The experiments were performed in triplicate.
Sample Preparation for untargeted Metabolic Analysis
After 48 hours of culture, the cells reached an 80% adherent density in the T-75 culture flask. The cells were divided into three groups: Group A (control group), Group B(treated with 10 μM isotretinoin for 48 h), and Group C (treated with 10 μM isotretinoin for 7 d). In Group C, the medium was changed every two days with 10 μM isotretinoin.Consistency in cell culture conditions was meticulously monitored to ensure that the cells did not exceed 90% confluence.The culture medium harvested from the HaCaT cells was aspirated using a pipette. Subsequently, the cells were rinsed with PBS at 37°C.
Eight biological replicates were used for each group for metabolomics analysis.The samples were thawed at 4°C and mixed with 1 mL of cold methanol/acetonitrile/H2O (2:2:1, v/v/v). The homogenate was sonicated at low temperature for 30 min, repeated twice. The mixture was then centrifuged for 20 min at 14,000 g and 4°C. The supernatant was dried using a vacuum centrifuge. For LC-MS analysis, the samples were re-dissolved in 100 μL acetonitrile/water (1:1, v/v) solvent.To ensure the reliability and stability of the instrument analysis, quality control (QC) samples were created by pooling 10 μL from each sample, which were then analyzed alongside the other samples. The QC samples were inserted regularly and analyzed every five samples.
UHPLC-Q-TOF MS
Analysis was performed using an UHPLC (1290 Infinity LC, Agilent Technologies) coupled to a quadrupole time-of-flight (AB SciexTriple TOF 6600) in Shanghai Applied Protein Technology Co., Ltd.
For hydrophilic interaction liquid chromatography (HILIC) separation, samples were analyzed using a 2.1 mm × 100 mm ACQUIY UPLC BEH Amide 1.7 µm column (waters, Ireland). In both ESI positive and negative modes, the mobile phase contained A=25 mM ammonium acetate and 25 mM ammonium hydroxide in water and B= acetonitrile. The gradient was 95% B for 0.5 min and was linearly reduced to 65% in 6.5 min, and then was reduced to 40% in 1 min and kept for 1 min, and then increased to 95% in 0.1 min, with a 3 min re-equilibration period employed.
The ESI source conditions were set as follows: Ion Source Gas1 (Gas1) as 60, Ion Source Gas2 (Gas2) as 60, curtain gas (CUR) as 30, source temperature: 600°C, IonSpray Voltage Floating (ISVF) ± 5500 V. In MS only acquisition, the instrument was set to acquire over the m/z range 60–1000 Da, and the accumulation time for TOF MS scan was set at 0.20 s/spectra. In auto MS/MS acquisition, the instrument was set to acquire over the m/z range 25–1000 Da, and the accumulation time for product ion scan was set at 0.05 s/spectra. The product ion scan is acquired using information dependent acquisition (IDA) with high sensitivity mode selected. The parameters were set as follows: the collision energy (CE) was fixed at 35 V with ± 15 eV; declustering potential (DP), 60 V (+) and −60 V (−); exclude isotopes within 4 Da, candidate ions to monitor per cycle: 10.
Data Processing
The raw MS data were converted to MzXML files using ProteoWizard MSConvert before importing into freely available XCMS software. For peak picking, the following parameters were used: centWave m/z = 10 ppm, peakwidth = c (10, 60), prefilter = c (10, 100). For peak grouping, bw = 5, mzwid = 0.025, minfrac = 0.5 were used. CAMERA (Collection of Algorithms of MEtabolite pRofile Annotation) was sued for annotation of isotopes and adducts. In the extracted ion features, only the variables having more than 50% of the nonzero measurement values in at least one group were kept. Compound identification of metabolites was performed by comparing of accuracy m/z value (<10 ppm), and MS/MS spectra with an in-house database established with available authentic standards.
Metabolites were compared with free online databases KEGG (http://www.genome.jp/kegg/) and HMDB (http://www.hmdb.ca/), and the corresponding KEGG pathways were extracted. KEGG enrichment analysis was performed using MetaboAnalyst (www.metaboanalyst.ca).
Statistical Analysis
For non-targeted metabolomics,after sum-normalization, the processed data were analyzed by R package (ropls), where it was subjected to multivariate data analysis, including Pareto-scaled principal component analysis (PCA) and orthogonal partial least-squares discriminant analysis (OPLS-DA). The 7-fold cross-validation and response permutation testing were used to evaluate the robustness of the model. The variable importance in the projection (VIP) value of each variable in the OPLS-DA model was calculated to indicate its contribution to the classification. Student’s t test was applied to determine the significance of differences between two groups of independent samples. VIP > 1 and P value < 0.05 were used to screen significant changed metabolites.One-way analysis of variance (ANOVA) method (GraphPad Prism 10.1.2) was applied for statistical analysis of the CCK-8 assay. P < 0.05 indicates a statistically significant difference.
Results
Effects of Isotretinoin on Proliferation of HaCaT Cells
The effect of isotretinoin on proliferation of HaCaT cells was determined by CCK8 assay. The results indicated that isotretinoin inhibited the proliferation of HaCaT cells by 21.3 ± 6.32% at a concentration of 1 μM (P = 0.652), 29.57 ± 5.15% at 5 μM (P = 0.002), and 40.14 ± 2.88% at 10 μM (P < 0.001).These results indicate that isotretinoin may block the proliferation of HaCaT cells in a dose-dependent manner (Figure 1).
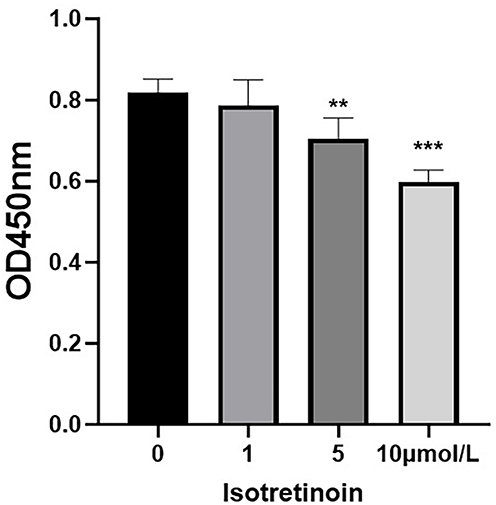 |
Figure 1 The effect of isotretinoin on proliferation of HaCaT cells. Cells were exposed to various concentrations of isotretinoin (0–10 μmol/L) and CCK8 assay was used to analyze cell proliferation.Data are expressed as means ± SD from three independent experiments, **P < 0.01, ***P < 0.001 compared to control by one-way ANOVA. |
Multivariate Analysis
Unsupervised principal component analysis (PCA) and orthogonal partial least squares discriminant analysis (OPLS-DA) were utilized to discriminate the overall distribution among samples. As shown in the PCA score plot, PC1 and PC2 accounted for 26.3 and 16.2% of the total variability, respectively. A clear cluster of the pooled QC samples was exhibited, indicating that the sample analysis sequence had satisfactory stability and repeatability (Figure 2). Furthermore, PCA revealed distinct differences among these groups, suggesting significant metabolic differences between them. Similarly, the OPLS-DA model also displayed clear segmentation between Group B and Group A (Figure 3A), as well as between Group C and Group A (Figure 3B). The model was evaluated by monitoring the model R2 (goodness of fit) and Q2 (predictive ability) values and permutation tests (n = 200) were performed on R2 and Q2. The OPLS-DA parameters of R2Y and Q2 for each comparison were as follows:Group B and Group A (R2Y = 0.972, Q2 = 0.88), Group C and Group A (R2Y = 0.994, Q2 =0.949), suggesting a good fit and predictability of these models.Moreover,the permutations plots showed that the permuted R2 (in green) and Q2 (in blue) values were lower than the original ones, and Q2 regression line intersected the vertical axis below zero, indicating all the OPLS-DA models were valid (Figure 3C,3D).
 |
Figure 2 The score plots of principal component analysis. |
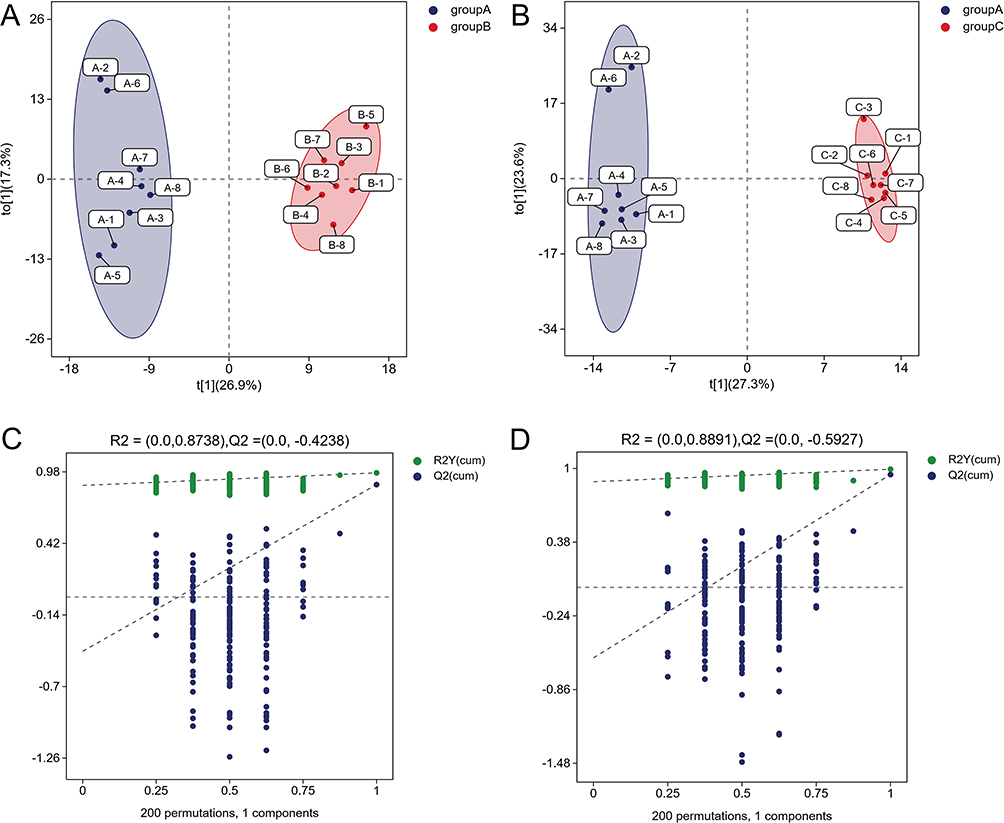 |
Figure 3 OPLS scores and 200 permutation tests for OPLS-DA models. The score plots of OPLS-DA: (A) Group A and Group B; (B): Group A and Group C. The permutation tests for OPLS-DA: (C) Group A and Group B; (D): Group A and Group C. |
Identification of Differentially Expressed Metabolites
VIP reflected the significance of those variables in the OPLS-DA model and was used to select the significant variables.To further identify the differentially metabolites in the samples, we screened the metabolites using variable importance projection (VIP)>1 and p-value<0.05 in the OPLS-DA model. Compared to Group A, we identified 115 significantly differentially metabolites in Group B,including glycerophosphocholine, adenosine, acetylcarnitine, creatine,Dl-lactate, hypoxanthine,uracil,LPC 18:1,creatine phosphate,5′-S-Methyl-5′-thioadenosine(43 upregulated, 72 downregulated)(Table 1). The chemical classification of these metabolites primarily consists of 35.96% lipids and lipid-like molecules, 21.93% organic acids and derivatives, 13.16% nucleosides nucleotides and analogues, 11.40% Organoheterocyclic compounds, and other metabolites (Figure 4A). In total, 140 significantly differential metabolites (81 upregulated and 59 downregulated) were identified in Group C compared to Group A. These metabolites include glycerophosphocholine, niacinamide, creatine, phytosphingosine, oleic acid, hypoxanthine, 1-Palmitoyl-sn-glycero-3-phosphocholine, Dl-lactate, 5’-S-Methyl-5’-thioadenosine and NG,NG-Dimethyl-L-arginine (Table 2). The highest proportion of differential metabolites was observed in lipids and lipid-like molecules (32.86%), followed by organic acids and derivatives (20.00%), organoheterocyclic compounds (13.57%), and nucleosides, nucleotides, and analogues (10.00%) (Figure 4B). Furthermore, to visualize the distribution of relative levels of the differentially metabolites in each group, the Euclideandistance matrix and complete linkage method were applied for the hierarchical clustering.The heatmap illustrated the top 50 differential metabolites and indicated that the metabolites in Group A were significantly separated from those in both Group B and Group C (Figure 5).
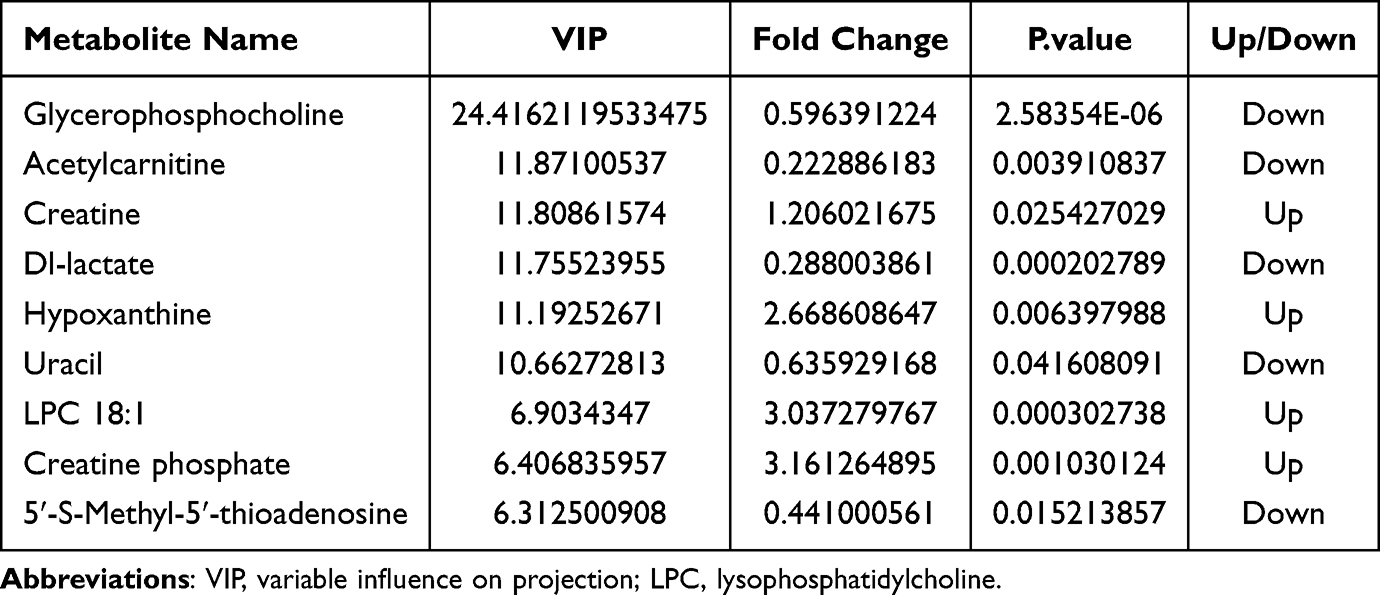 |
Table 1 The Top10 Significant Differential Metabolites Among Group A and B |
 |
Table 2 The Top10 Significant Differential Metabolites Among Group A and C |
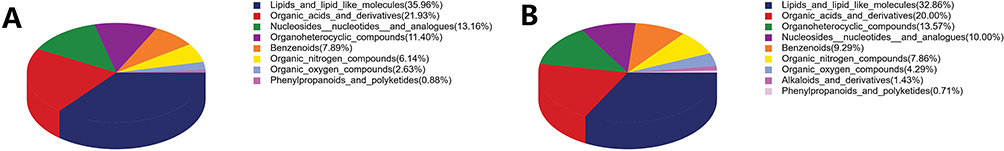 |
Figure 4 Proportion of identified metabolites in each chemical classification. (A) Differential metabolites between Group A and Group B. (B) Differential metabolites between Group A and Group C. |
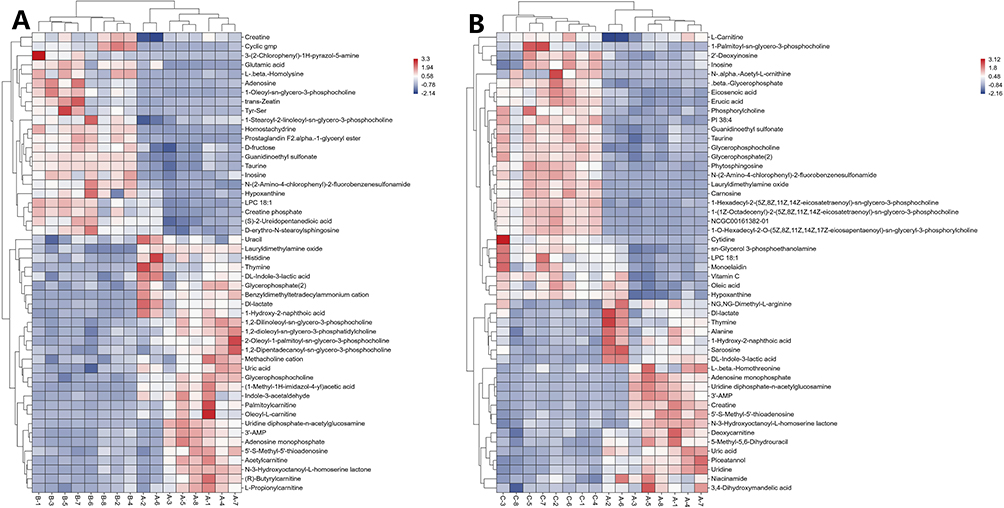 |
Figure 5 Hierarchical clustering heat map. (A) The top 50 differential metabolites between Group A and Group B. (B) The top 50 differential metabolites between Group A and Group C. Color scale on the right of the heat map represented normalized values of each identified metabolites content. Red represents significant upregulation; blue represents significant downregulation. |
Pathway Enrichment Analysis
Fisher’s exact test was employed to analyze and calculate the significance level of metabolite enrichment for each pathway.The p-value was used to identify the metabolic pathways that are significantly affected:the smaller the p-value, the more significant the difference in the metabolic pathway. Compared with the Group A,30 metabolic pathways were significantly changed (p < 0.05) in Group B. The most significant metabolic pathways included purine metabolism, nucleotide metabolism, ABC transporters, taurine and hypotaurine metabolism, and cGMP-PKG signaling pathway (Figure 6A). In total, 46 metabolic pathways were significantly changed (p < 0.05) in Group C compared with Group A. The enriched pathways primarily involved nucleotide metabolism, ABC transporters, purine metabolism, glycerophospholipid metabolism, and choline metabolism in cancer (Figure 6B).
 |
Figure 6 Statistics of KEGG enrichment. (A)The top 10 enriched pathways between Group A and Group B. (B) The top 10 enriched pathways between Group A and Group C. The x axis indicates the rich factor corresponding to each pathway, and the y axis indicates name of the KEGG metabolic pathway. The size and color of bubbles represent the number and degree of enrichment of different metabolites, respectively. |
Discussion
In this study, we investigated the therapeutic mechanism of isotretinoin on HaCaT cells at two different time points: short-term (48 h) or long-term (7 d), utilizing a metabolomics approach. Our findings revealed that, in comparison to Group A, there were 115 significantly different metabolites identified in Group B and 140 in Group C. Furthermore, when compared to Group A, 30 metabolic pathways were significantly altered in Group B, while 46 pathways were perturbed in Group C.
Nucleotide Metabolism and Purine Metabolism
Our data demonstrated that nucleotide metabolism and purine metabolism were significantly perturbed pathways at all time points (48 h and 7 d), implicating their important roles in the mechanism of action of isotretinoin on keratinocytes in both short-term and long-term contexts. Nucleotides are vital metabolites in organisms and participate in almost all biochemical processes within cells.20 Nucleotide metabolism fuels DNA replication and RNA synthesis, which are indispensable for cell proliferation. In our study, isotretinoin inhibited the proliferation of HaCaT cells in a dose-dependent manner. Previous evidence has demonstrated that isotretinoin regulates keratinocyte proliferation by binding to the nuclear receptors all-trans retinoic acid receptor (RAR) and all-trans retinoid X receptor (RXR).21 Our results suggest that the inhibition of keratinocyte proliferation by isotretinoin may be associated with its influence on the nucleotide metabolism pathway through the activation of RAR/RXR.Furthermore, seven metabolites related to the purine metabolic pathway changed significantly, including adenosine monophosphate, hypoxanthine, uric acid, inosine, 2′,3′-cyclic adenosine monophosphate, 3′-adenosine monophosphate, and 2′-deoxyinosine. Adenosine monophosphate is not only a synthesis product of purine nucleotides, but also a product of their oxidative decomposition.22 Both hypoxanthine and inosine serve as key intermediates in the degradation of purines, while uric acid is the final product of purine catabolism in humans.23 In our study, the treatment with isotretinoin disrupted the metabolism of these substances. Hence,the effect of isotretinoin on keratinocytes is associated with the regulation of purine synthesis and degradation.
ABC Transporters
ATP-binding cassette (ABC) transporters are a class of transmembrane proteins that are extensively expressed in various tissues, including keratinocytes, hepatocytes, intestinal epithelial cells, and adipocytes.24 They play multiple key functions in keratinocytes, including lipid transport, barrier formation, drug metabolism, defense responses, and responses to environmental stress such as ultraviolet radiation.25,26 In our study, ABC transporters was another significantly perturbed pathway in HaCaT cells treated with isotretinoin for 48 h and 7 d. Previous research has indicated that ABC transporters was the most altered in patients with moderate-to-severe acne.27 Similarly, in a rat acne model, ABC transporters were identified as a significantly perturbed pathway.28 Therefore, the regulation of the ABC pathway in keratinocytes by isotretinoin may contribute to its therapeutic efficacy in treating acne.
Glycerophospholipid Metabolism
Glycerophospholipids, commonly referred to as phospholipids, are essential components of the lipid bilayer of cellular membranes and play a pivotal role in cell signaling and metabolism.29 Our study revealed that after isotretinoin treatment, the glycerophospholipid metabolism pathways in HaCaT cells were significantly altered at 7 d, rather than at 48 h.This indicates the differences in the mechanism of action of isotretinoin on keratinocytes in short-term and long-term contexts. Furthermore, seven metabolites associated with the glycerophospholipid metabolic pathway exhibited significant changes, including glycerol 3-phosphate, O-phosphoethanolamine, cytidine 5′-diphosphate ethanolamine, phosphorylcholine, glycerophosphocholine, sn-glycerol 3-phosphoethanolamine, and 1-palmitoyl-sn-glycero-3-phosphocholine. Notably, phosphorylcholine is the most abundant phospholipid in biological membranes, contributing to the characteristic bilayer structure of cells and regulating membrane integrity.30 Glycerol 3-phosphate serves as a key precursor for the synthesis of membrane lipids,31 while phosphoethanolamine is involved in phospholipid turnover, acting as a substrate for various phospholipids in cell membranes. Glycerophosphocholine is a product of the breakdown of phospholipid components in cell membranes.32 In our study, a 7-day isotretinoin treatment led to a decrease in the levels of these substances. This finding suggests that long-term isotretinoin treatment may influence the synthesis and function of cell membranes by inhibiting the production of these metabolites.
The limitations of this study include the following: the use of immortalized cell lines instead of primary keratinocyte cell lines; the lack of targeted metabolomics validation; the absence of functional assays; and the limited translational relevance.
Conclusion
This study employed UHPLC-qTOF-MS to investigate the intracellular metabolic alterations induced by isotretinoin in HaCaT keratinocytes. Through the analysis of metabolic profiles, we identified 115 metabolites that were significantly differentially expressed after 48 h of isotretinoin treatment and 140 metabolites after 7 d of treatment.Isotretinoin modulates multiple metabolic pathways, including nucleotide metabolism,purine metabolism, ABC transporters,and glycerophospholipid metabolism.Our findings will help clarify the underlying mechanism of action of isotretinoin on keratinocytes at the metabolite level and the mucocutaneous adverse effects associated with isotretinoin in the treatment of dermatological diseases.However, this study has certain limitations. Further validation studies, involving targeted metabolomics and functional assays, are required to confirm and validate these initial discovery results.
Data Sharing Statement
The data used to support the findings of this study are available from the corresponding author upon request.
Ethics Approval and Consent to Participate
This study did not involve human or animal subjects, and thus, no ethical approval was required.
Acknowledgments
We thank our collaborators and advisors for their valuable feedback and support during the preparation of this manuscript.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was funded by the National Natural Science Foundation of China (Project No. 82460623) and the Natural Science Foundation of Jiangxi Province (no. 20232BAB206126).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Shemer A, Gupta AK, Kassem R, Sharon N, Quinlan EM, Galili E. Low‐dose isotretinoin versus minocycline in the treatment of rosacea. Dermatol Ther. 2021;34(4):e14986. doi:10.1111/dth.14986
2. Demirbas A, Diremsizoglu E, Esen M, Demirbas GU. Efficacy, safety, and recurrence in seborrheic dermatitis: a dose-dependent analysis of oral isotretinoin (10 mg vs 20 mg). J Am Acad Dermatol. 2025;93(3):671–11. doi:10.1016/j.jaad.2025.05.1376
3. Tambe S, Jerajani H, Pund P. Granulomatous periorificial dermatitis effectively managed with oral isotretinoin. Indian Dermatol Online J. 2018;9(1):68–70. doi:10.4103/idoj.IDOJ_129_17
4. Looney M, Smith KM. Isotretinoin in the treatment of granuloma annulare. Ann Pharmacother. 2004;38(3):494–497. doi:10.1345/aph.1D312
5. Georgiou S, Monastirli A, Pasmatzi E, et al. Cutaneous sarcoidosis: complete remission after oral isotretinoin therapy. Acta Derm Venereol. 1998;78(6):457–459. doi:10.1080/000155598442782
6. Brzezinski P, Zonda GI, Hincu MA, et al. A multicenter cohort study evaluating the teratogenic effects of isotretinoin on neonates. Children. 2022;9(11):1612. doi:10.3390/children9111612
7. Botsali A, Kocyigit P, Uran P. The effects of isotretinoin on affective and cognitive functions are disparate in adolescent acne vulgaris patients. J Dermatolog Treat. 2020;31(7):734–738. doi:10.1080/09546634.2019.1606396
8. Fernández-Crehuet P, Fernández-Crehuet JL, Allam MF, Fernández-Crehuet Navajas R. Hepatotoxicity of isotretinoin in patients with acne and Gilbert’s syndrome: a comparative study. BMJ Open. 2014;4(3):e004441. doi:10.1136/bmjopen-2013-004441
9. Al Dhafiri M, Kaliyadan F, Almukhaimar S, et al. Isotretinoin-induced dyslipidemia: a single-center study in Saudi Arabia. Patient Prefer Adherence. 2024;18:197–206. doi:10.2147/PPA.S445904
10. Kmieć ML, Pajor A, Broniarczyk-Dyła G. Evaluation of biophysical skin parameters and assessment of hair growth in patients with acne treated with isotretinoin. Postepy Dermatol Allergol. 2013;30(6):343–349. doi:10.5114/pdia.2013.39432
11. Xing F, Liao W, Jiang P, Xu W, Jin X. Effect of retinoic acid on aquaporin 3 expression in keratinocytes. Genet Mol Res. 2016;15(1):15016951 doi:10.4238/gmr.15016951
12. Tadini KA, Gaspar LR, RMaia Campos PM. Epidermal effects of tretinoin and isotretinoin: influence of isomerism. Pharmazie. 2006;61(5):453–456.
13. Wang C, Li H, Ma P, et al. The third‐generation retinoid Adapalene triggered DNA damage to induce S‐phase arrest in HaCat cells. Fundam Clin Pharmacol. 2020;34(3):380–388. doi:10.1111/fcp.12527
14. Pani B, Cornatzer E, Cornatzer W, et al. Up-regulation of transient receptor potential canonical 1 (TRPC1) following Sarco(endo)plasmic reticulum Ca2+ ATPase 2 gene silencing promotes cell survival: a potential role for TRPC1 in Darier’s disease. Mol Biol Cell. 2006;17(10):4446–4458. doi:10.1091/mbc.e06-03-0251
15. Shi G, Liao PY, Cai XL, et al. FoxO1 enhances differentiation and apoptosis in human primary keratinocytes. Exp Dermatol. 2018;27(11):1254–1260. doi:10.1111/exd.13775
16. Want EJ. LC-MS Untargeted Analysis. Methods Mol Biol. 2018;1738:99–116. doi:10.1007/978-1-4939-7643-0_7
17. Xu M, Chen J, Peng C, Mo L. Non-targeted metabolomics analysis of indoleamine 2,3-dioxygenase inhibitor treatment in a mouse model of early-stage lung adenocarcinoma. Transl Cancer Res. 2024;13(2):900–915. doi:10.21037/tcr-23-1236
18. Ogiso H, Miura K, Nagai R, Osaka H, Aizawa K. Non-targeted metabolomics reveal apomorphine’s therapeutic effects and lysophospholipid alterations in steatohepatitis. Antioxidants. 2024;13(11):1293. doi:10.3390/antiox13111293
19. Pei D, Huang J, Chen S, et al. The study of the protection mechanism of Calycosin-7-O-β-d-glucoside against oxygen–glucose deprivation/reperfusion in HT22 cells based on non-targeted metabolomics and network analysis. Molecules. 2025;30(3):549. doi:10.3390/molecules30030549
20. Cao S, Wang J, You X, Zhou B, Wang Y, Zhou Z. Purine metabolism and pyrimidine metabolism alteration is a potential mechanism of BDE-47-induced apoptosis in marine rotifer Brachionus plicatilis. Int J Mol Sci. 2023;24(16):12726. doi:10.3390/ijms241612726
21. Beckenbach L, Baron JM, Merk HF, Löffler H, Amann PM. Retinoid treatment of skin diseases. Eur J Dermatol. 2015;25(5):384–391. doi:10.1684/ejd.2015.2544
22. Chen L, Qu B, Wang H, et al. The effect of curculigo orchioides (Xianmao) on kidney energy metabolism and the related mechanism in rats based on metabolomics. Food Sci Nutr. 2021;9(11):6194–6212. doi:10.1002/fsn3.2573
23. Van Der Pol KH, Koenderink J, Van Den Heuvel JJMW, et al. Effects of allopurinol and febuxostat on uric acid transport and transporter expression in human umbilical vein endothelial cells. PLoS One. 2024;19(6):e0305906. doi:10.1371/journal.pone.0305906
24. Wenzel JJ, Piehler A, Kaminski WE. ABC A-subclass proteins: gatekeepers of cellular phospho- and sphingolipid transport. Front Biosci. 2007;12(8–12):3177–3193. doi:10.2741/2305
25. Gęgotek A, Skrzydlewska E. The role of ABC transporters in skin cells exposed to UV radiation. Int J Mol Sci. 2022;24(1):115. doi:10.3390/ijms24010115
26. Takenaka S, Itoh T, Fujiwara R. Expression pattern of human ATP -binding cassette transporters in skin. Pharmacol Res Perspect. 2013;1(1):e00005. doi:10.1002/prp2.5
27. Yu S, Xiao Z, Ou Yang X, Wang X, Zhang D, Li C. Untargeted metabolomics analysis of the plasma metabolic signature of moderate-to-severe acne. Clin Chim Acta. 2022;533:79–84. doi:10.1016/j.cca.2022.06.012
28. Ou-Yang XL, Zhang D, Wang XP, et al. Nontargeted metabolomics to characterize the effects of isotretinoin on skin metabolism in rabbit with acne. Front Pharmacol. 2022;13:963472. doi:10.3389/fphar.2022.963472
29. Ivanova PT, Milne SB, Byrne MO, Xiang Y, Brown HA. Glycerophospholipid identification and quantitation by electrospray ionization mass spectrometry. Methods Enzymol. 2007;432:21–57. doi:10.1016/S0076-6879(07)32002-8
30. Canlet C, Tremblay-Franco M, Gautier R, et al. Specific metabolic fingerprint of a dietary exposure to a very low dose of endosulfan. J Toxicol. 2013;2013:545802. doi:10.1155/2013/545802
31. Wang S, Ma C, Long J, et al. Impact of CRAMP-34 on Pseudomonas aeruginosa biofilms and extracellular metabolites. Front Cell Infect Microbiol. 2023;13:1295311. doi:10.3389/fcimb.2023.1295311
32. Gonzales MM, Tarumi T, Kaur S, et al. Aerobic fitness and the brain: increased N-acetyl-aspartate and choline concentrations in endurance-trained middle-aged adults. Brain Topogr. 2013;26(1):126–134. doi:10.1007/s10548-012-0248-8
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.