Back to Journals » Journal of Pain Research » Volume 9
TRPA1 in the spinal dorsal horn is involved in post-inflammatory visceral hypersensitivity: in vivo study using TNBS-treated rat model
Authors Li Q, Guo C, Chowdhury MA, Dai T, Han W
Received 1 August 2016
Accepted for publication 3 November 2016
Published 2 December 2016 Volume 2016:9 Pages 1153—1160
DOI https://doi.org/10.2147/JPR.S118581
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Michael Schatman
Qian Li,1,* Cheng-Hao Guo,2,* Mohammed Ali Chowdhury,1 Tao-Li Dai,1 Wei Han,1,3
1Department of Gastroenterology, Qilu Hospital of Shandong University, 2Department of Pathology, Medical School of Shandong University, 3Laboratory of Translational Gastroenterology, Shandong University, Qilu Hospital, Jinan, Shandong Province, People’s Republic of China
*These authors contributed equally to this work
Introduction: The transient receptor potential ankyrin-1 (TRPA1) channel, a pain transducer and amplifier, is drawing increasing attention in the field of visceral hypersensitivity, commonly seen in irritable bowel syndrome and inflammatory bowel disease. However, the role of TRPA1 in visceral nociception during post-inflammatory states is not well defined. Here, we explore the correlation between TRPA1 expression in the spinal dorsal horn (SDH) and persistent post-inflammatory visceral hypersensitivity.
Methods: We injected rats intracolonically with 2,4,6-trinitrobenzene sulfonic acid (TNBS) or vehicle (n=12 per group). Post-inflammatory visceral hypersensitivity was assessed by recording the electromyographic activity of the external oblique muscle in response to colorectal distension. TRPA1 expression and distribution in the spinal cord and colon were examined by Western blotting and immunohistochemistry.
Results: Animals exposed to TNBS had more abdominal contractions than vehicle-injected controls (P<0.05), which corresponded to a lower nociceptive threshold. Expression of TRPA1 in the SDH (especially in the substantia gelatinosa) and the colon was significantly greater in the TNBS-treated group than in controls (P<0.05). In the SDH, the number of TRPA1-immunopositive neurons was 25.75±5.12 in the control group and 34.25±7.89 in the TNBS-treated group (P=0.023), and integrated optical density values of TRPA1 in the control and TNBS-treated groups were 14,544.63±6,525.54 and 22,532.75±7,608.11, respectively (P=0.041).
Conclusion: Our results indicate that upregulation of TRPA1 expression in the SDH is associated with persistent post-inflammatory visceral hypersensitivity in the rat and provides insight into potential therapeutic targets for the control of persistent visceral hypersensitivity.
Keywords: TRPA1, post-inflammatory visceral hypersensitivity, spinal dorsal horn, substantia gelatinosa
Introduction
Chronic abdominal pain is one of the common and major complaints of the irritable bowel syndrome (IBS) and inflammatory bowel disease (IBD) remission. In the last 2 decades, studies have found that altered intestinal permeability, gut microbiota, and dysregulated intestinal immune function along with changes in function or structure of the enteric nervous system are important contributing factors of chronic abdominal pain,1–3 but the detailed mechanisms remain unclear due to the undetectable structural abnormalities in the peripheral organs. Visceral hypersensitivity is considered to play an important role in the development of chronic visceral pain, and it refers to the exaggerated sensory reflex of visceral organs to the noxious stimuli.4–6 However, there is a lack of knowledge in the mechanisms underlying the visceral hypersensitivity.
Recently, a growing number of studies have tried to explore the molecular mechanism of the visceral hypersensitivity.7,8 The transient receptor potential (TRP) family of ion channels are emerging interested targets, as they are known to serve many important roles, such as in mechanosensation, pain, regulation of gastrointestinal motility, absorptive and secretory processes, and mucosal homeostasis.9–11 TRP vanilloid-1 (TRPV1) was the first cloned prototypical vanilloid TRP channel in mammalian sensory neurons and the most widely studied member in the family.12 TRPV1 was seen to have been closely correlated with the post-inflammatory visceral pain in rodents, given that the upregulation of TRPV1 expression persists long after the initial inflammatory has subsided.13 It is reported that 97% of TRP ankyrin-1 (TRPA1)-expressing sensory neurons express TRPV1,14,15 which suggests that they have similar functions and features. TRPA1, the sole member of the TRPA subfamily, is a widely expressed channel present in the sensory neurons (dorsal root ganglia, trigeminal ganglia, and nodose ganglia)16,17 and many non-neuronal tissues, including lung, pancreas, small intestine, and colon.18 Moreover, TRPA1 channel is highly conserved across the animal kingdom. It contains 14–16 ankyrin repeats within the N-terminal domain, which is an unusual structural feature that might be relevant to its proposed mechanosensory role. TRPA1 channel can be activated by noxious cold (<17°C) and many chemical stimulus, including menthol, tetrahydrocannabinol, allyl isothiocyanate (AITC), and cinnamaldehyde.14,19,20 The activation of TRPA1 could facilitate Ca2+ influx in sensory afferent nerves and potentiate signal transmission between sensory and central nervous system neurons.20,21 Activated TRPA1 could also induce neurogenic inflammation and somatic pain by stimulating the release of some nociceptive peptides containing substance P (SP) and calcitonin gene-related peptide (CGRP).22,23 Our previous study, likewise, showed that TRPA1 expression was upregulated in the colonic afferent dorsal root ganglia of the IBS visceral hyperalgesia rat model.24 All these previous studies indicate that TRPA1 plays multiple key roles in the pain and/or visceral hypersensitivity. It is worth noting that previous studies have demonstrated that transient colitis could cause long-lasting alterations of visceral sensory function,25,26 that is, post-inflammatory visceral hypersensitivity, but the exact mechanisms were unknown.
To determine whether post-inflammatory visceral hypersensitivity is associated with changes in TRPA1, we examined the distribution and expression of TRPA1 in the spinal cord and colon of a well-known rat model of this condition using Western blotting and immunohistochemistry.
Materials and methods
Animals
All animals used in the study were female Wistar rats weighing between 180 and 200 g (obtained from the experimental animal center of Shandong University). Animals had free access to tap water and standard laboratory food. They were individually housed in polypropylene cages in a room with controlled temperature (21°C±1°C) and light–dark cycle (12–12 hours). This study adhered to the ethical guidelines of the International Association for the Study of Pain. All experimental protocols described in this study were approved by the Animal Care and Use Committee of the Medical school of Shandong University (Permit Number: 2011066). All efforts were made to minimize animal suffering and the number of animals necessary for reliable scientific data.
Experimental animal models and grouping
A total of 24 Wistar rats were randomly allocated equally into two groups: the 2,4,6-trinitrobenzene sulfonic acid (TNBS)-treated group (n=12) and the control group (n=12). After an overnight fast, anesthesia was induced via intraperitoneal injection of acepromazine (0.6 mg/kg; Sigma-Aldrich, St Louis, MO, USA) and ketamine (120 mg/kg; Sigma-Aldrich), respectively. For the TNBS-treated group, colonic inflammation was induced by the intracolonic injection of 14–16 mg/rat TNBS (Sigma-Aldrich) dissolved in 25% ethanol at 6 cm proximal to the anus through a 24-gauge catheter. The control group received the same volume of vehicle (normal saline) injected in the same manner. Then, the rats were kept in a position with heads low and tails high for several minutes to avoid leakage of the instilled intracolonic solutions. After the model was established, the rats were put back into the humidity chamber. They were observed daily; feces characteristics, eating situation, and mental state were recorded along with the weight. Six weeks after TNBS administration, the visceral hypersensitivity of rats was measured by the electromyographic (EMG) recording.
The visceral hypersensitivity to colorectal distension (CRD) testing
The visceral sensitivity of rats was assessed by recording the EMG activity of the external oblique muscle in response to CRD. EMG recordings were performed as follows. After anesthetization with an intraperitoneal administration of acepromazine (0.6 mg/kg) and ketamine (120 mg/kg), the electrodes were implanted in the striated muscles of the abdomen, and electrodes were exteriorized on the back of the neck, which was protected by a glass tube attached to the skin. Rats were allowed to recover from the surgery for a week. They were fasted overnight (with free access to water) before the experiment. On the day of the experiment, rats were lightly anesthetized by an intraperitoneal administration of acepromazine and ketamine at doses of 0.6 and 120 mg/kg, respectively, and the balloon (4 cm) made of a surgical glove finger attached to a Tygon tubing with thread was carefully inserted into the colon at 2 cm distal from the end of the balloon and secured to the base of the tail with tape. Rats were placed in plastic tunnels (8 cm diameter, 20 cm length) and allowed to recover and acclimate for 30 minutes before testing. After the animals were fully awake and acclimatized, CRD was performed by rapidly inflating the balloon to constant pressure. The first distension was performed at a pressure of 15 mmHg and an increment of 15 mmHg was added at each successive following step, until a maximal pressure of 45 mmHg, each distension step lasted 5 minutes.
Sample collection and processing
Following decapitation, the lumbosacral segment (L4–S2) of the spinal cord was dissected out, and the distal colon was excised and rinsed with saline. Part of each tissue was used for Western blotting, and the remaining part was immediately placed in 4% formalin for immunohistochemistry.
Western blotting procedure
Tissue samples (~1 g) of the distal colon and the spinal cord were extracted in three volumes of extract buffer, centrifuged at 13,000× g for 10 minutes at 4°C; the supernatant was saved at −70°C until usage. Western blotting protocol was developed for the Bio-Rad protein gel and transfer apparatus. The membranes were blocked and incubated with the primary antibodies at 4°C, overnight, and subsequently with the secondary antibodies for 2 hours. The gel was transferred to a polyvinylidene difluoriizde membrane and blotted with primary (overnight) and then with secondary (1 hour) antibodies. The primary antibodies included a rabbit polyclonal anti-TRPA1 antibody (1:200; Novus Biologicals) and a mouse monoclonal anti-glyceraldehyde-3-phosphate dehydrogenase (GAPDH) antibody (1:500; Sigma). The secondary antibodies included anti-mouse Ig for GAPDH (1:1,000) and anti-rabbit Ig for TRPA1 (1:3,000). The integrated density of Western blotting bands was quantitatively analyzed by the Gel Image analysis software. TRPA1 protein expression was normalized to GAPDH.
Immunohistochemical localization of TRPA1 procedure
The distal colon tissue samples and the lumbosacral segment (L4–S2) of the spinal cord were fixed for 12 hours in 4% formalin, dehydrated through graded ethanol, and embedded in paraffin. Sections (5 mm) were rehydrated and submerged in antigen retrieval solution (citrate buffer, 10 mM, pH 6, 95°C, 3 minutes). After inhibition of endogenous peroxidases with 0.6% H2O2 in 100% methanol for 30 minutes and incubation in blocking solution (phosphate-buffered saline [PBS] containing 1% bovine serum albumin and 2% normal serum), they were washed with room temperature PBS for 5 minutes (2×). Later tissues were incubated with a rabbit polyclonal anti-TRPA1 antibody (1:100; Novus Biologicals) overnight at 4°C. They were then washed with room temperature PBS for 5 minutes (2×). The anti-rabbit IgG antibody (1:500; Sigma) on to the tissue sections were incubated at room temperature for 30 minutes. Slides were then washed with room temperature PBS for 2 minutes (2×). Tissue slides were incubated with ABC reagent (1:200; Thermo Fisher Scientific) for 30 minutes, and slides were washed with room temperature PBS for 2 minutes (2×). Tissues were then strained with DAB solution for ~1–2 minutes (or until brown stain developed, maximum 15 minutes). Tissue slides were rinsed in distilled H2O2 for 1 minute (1×) and then for 3 minutes (2×). Tissues were then strained with hematoxylin solution for 1–2 minutes, and the tissue slides were then rinsed in distilled H2O2 for 1 minute (1×) and then for 3 minutes (2×). Then TRPA1-immunopositive neurons count and the integrated optical density (IOD) values were performed using IPP6.0 software (Media Cybernetics, Bethesda, MD, USA).
Statistical analysis
Data are presented as mean ± standard deviation, and all statistical analysis were performed using SPSS 18.0 statistical software (SPSS Inc, Chicago, IL, USA). EMG data were analyzed by two-way repeated-measures analysis of variance (ANOVA). Other data were analyzed by the Student’s t-test or one-way ANOVA where appropriate. A comparison was considered statistically significant when the P-value was <0.05.
Results
The visceral hypersensitivity
The visceral hypersensitivity to CRD was assessed after 6 weeks of TNBS infusion with no microscopic evidence of inflammation, which was observed in rat colonic tissues collected by that time. The number of abdominal contractions as an index of visceral pain was measured by EMG recording. For the TNBS-treated rats, the number of abdominal contractions was significantly increased in response to CRD at 15 mmHg (P=0.012), 30 mmHg (P=0.012), and 45 mmHg (P=0.011; Figure 1). These data indicate that the TNBS-treated rats were more sensitive to CRD than controls, suggesting that rats of TNBS treatment produce a persistent post-inflammatory visceral hypersensitivity.
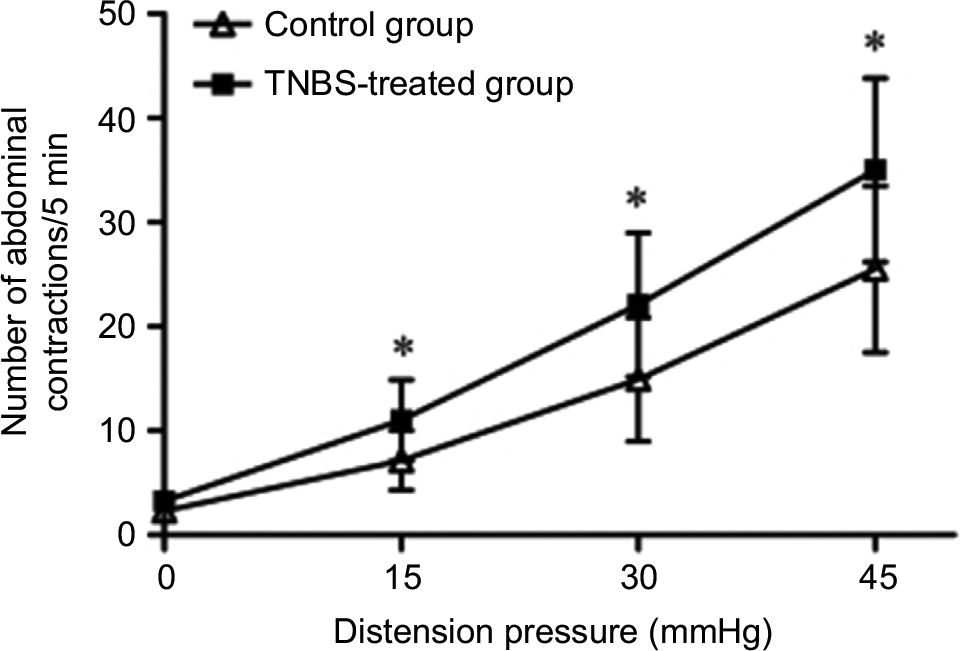 | Figure 1 Abdominal contractions in response to graded CRD between control and TNBS-treated groups in Wistar rats (n=12 per group). Note: Values shown are the mean ± SD. *P<0.05 significantly different from the control group. Abbreviations: CRD, colorectal distension; TNBS, 2,4,6-trinitrobenzene sulfonic acid. |
Expression of TRPA1 in the colon
This step determined whether colonic TRPA1 expression was different between TNBS-treated rats and controls. After extracting rat colon protein, the expression of TRPA1 was tested by Western blotting. Densitometric analysis of the bands revealed expression of TRPA1 was 26.27±12.05 in the control group and 37.53±14.19 in the TNBS group, respectively. These data indicate that the expression of TRPA1 was significantly greater in the TNBS-treated group than in controls (P=0.045; Figure 2). Moreover, immunohistochemical staining was used to further detect colonic TRPA1 expression, but revealed negative result.
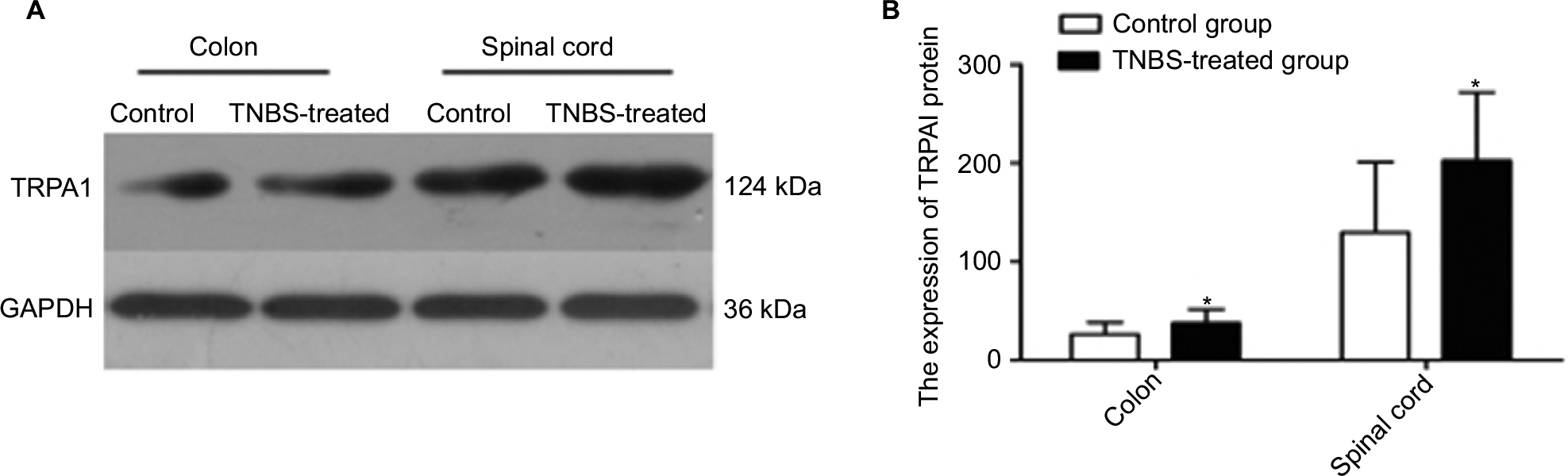 | Figure 2 (A) Western blotting image of TRPA1 and GAPDH expression in the colon and spinal cord of the rats from the control and TNBS-treated groups (n=12 per group). The top panel was the target band, TRPA1 protein (124 kDa) and the bottom one was for the loading control GAPDH (36 kDa). For the TRPA1, the TNBS-treated group showed much stronger and wider signal compared with the control group accordingly. (B) Western blotting quantitative analysis of the TRPA1 expression in the rat’s colon and spinal cord of the control and TNBS-treated groups. Expression of TRPA1 was significantly greater in the TNBS-treated group than in controls (P<0.05). Notes: Values shown are the mean ± SD. *P<0.05 significantly different from the control group. Abbreviations: TNBS, 2,4,6-trinitrobenzene sulfonic acid; TRPA1, transient receptor potential ankyrin-1. |
Expression and localization of TRPA1 in the spinal cord
Western blotting detection
Western blotting was used to determine whether TRPA1 was expressed in the spinal cord. Results of this analysis showed that the expression of TRPA1 was 129.63±71.28 in the control group and 202.74±69.44 in the TNBS-treated group. This suggests that TRPA1 expressed in the spinal cord and the expression of TRPA1 from the TNBS-treated rats were significantly greater than that from the control rats (P=0.018; Figure 2).
Immunohistochemical detection
To further investigate the expression and localization of TRPA1 in the spinal dorsal horn (SDH), we examined the distribution features of TRPA1 using immunohistochemical staining. The expression of TRPA1 was observed in the SDH from the typical photomicrographs of TRPA1 staining, and TRPA1 expression in TNBS-treated group was significantly stronger and thicker compared to the control group. Interestingly, for the TRPA1 expression localization in the SDH, we found that the more superficial layer, the stronger expression was. This means that TRPA1-immunopositive neurons were mainly localized in the superficial layers of the SDH, especially in the substantia gelatinosa (SG; lamina II of Rexed). Figure 3 shows four representative immunohistochemical staining images of TRPA1 distribution characterization in the SDH of the control and TNBS-treated groups. We counted the number of TRPA1-immunopositive neurons and measured IOD values of TRPA1 in the SDH and SG by using IPP6.0 software. In the SDH, the number of TRPA1-immunopositive neurons was 25.75±5.12 in the control group and 34.25±7.89 in the TNBS-treated group (P=0.023; Figure 4), and IOD values of TRPA1 in the control and TNBS-treated groups were 14,544.63±6,525.54 and 22,532.75±7,608.11, respectively (P=0.041; Figure 5). These results indicate that expression of TRPA1 in the SDH from the TNBS-treated group was significantly greater than that from the controls. In parallel with the SDH, expression of TRPA1 in the SG was also significantly greater in the TNBS-treated group than in controls. The number of TRPA1-immunopositive neurons was 8.13±2.03 and 12.25±3.88 (P=0.019; Figure 4), and IOD values of TRPA1 were 3,504.25±1,237.86 and 5,353.5±1,427.04 (P=0.015; Figure 5) in the control and TNBS-treated groups, respectively.
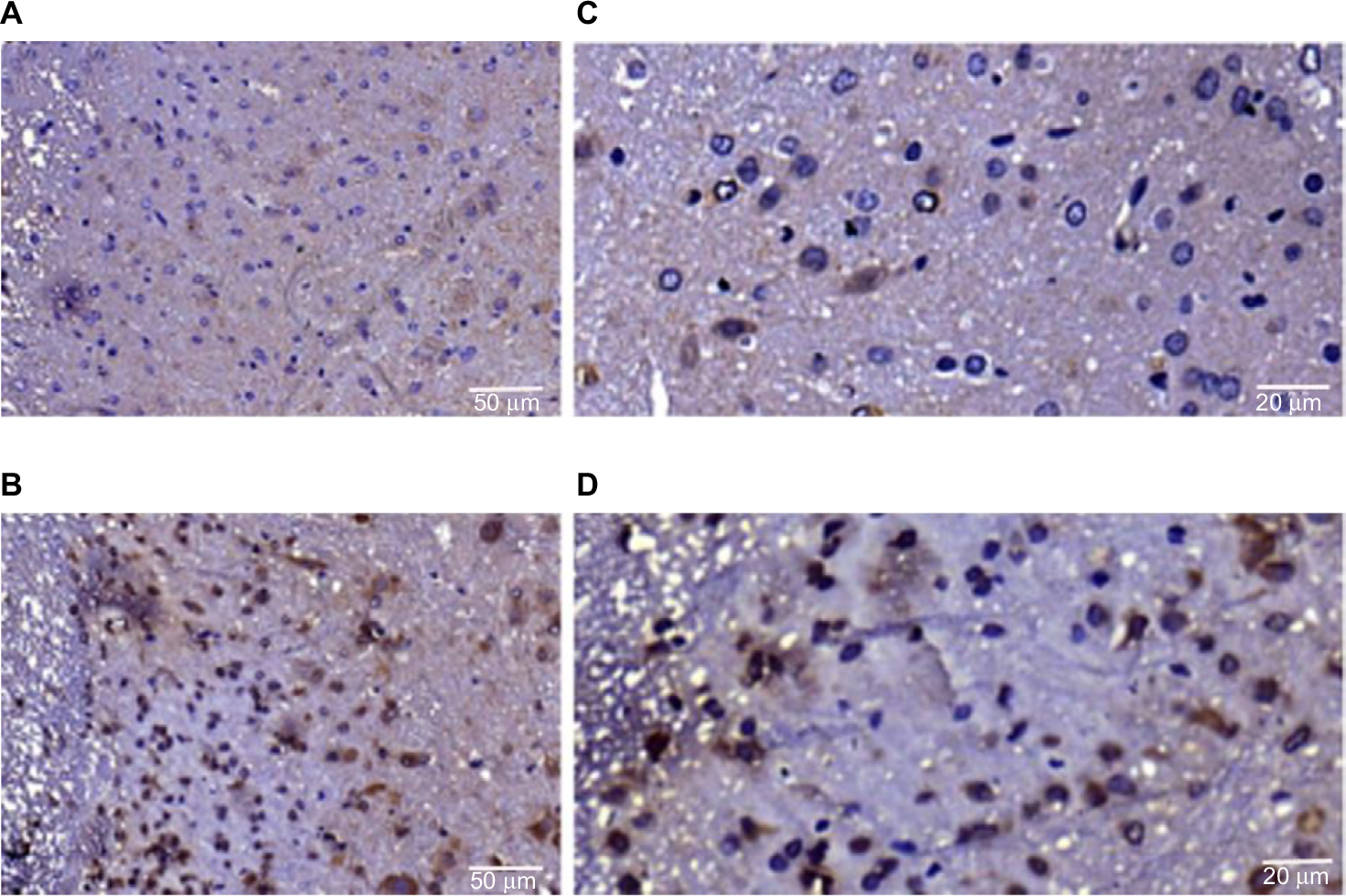 | Figure 3 Representative immunostaining for the TRPA1 in paraffin sections of the spinal dorsal horn from the control group (A) and TNBS-treated group (B). Scale bar = 50 μm. Representative immunostaining for the TRPA1 in paraffin sections of substantia gelatinosa from the control group (C) and TNBS-treated group (D). Scale bar = 20 μm. Abbreviations: TNBS, 2,4,6-trinitrobenzene sulfonic acid; TRPA1, transient receptor potential ankyrin-1. |
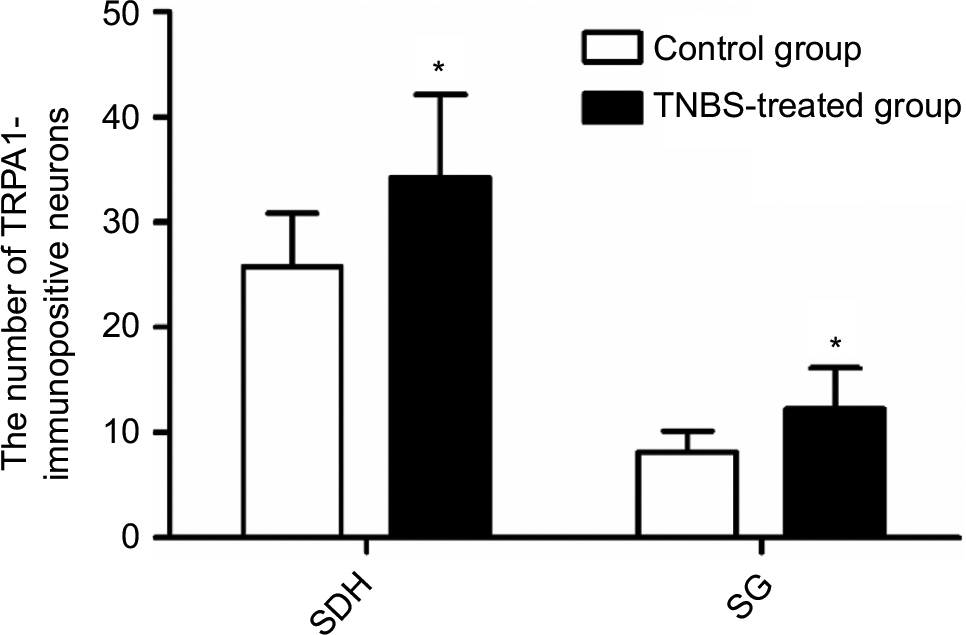 | Figure 4 A comparison of the number of TRPA1-immunopositive neurons in the SDH and the SG between the two groups (n=8 per group). Notes: TNBS-treated rats had a greater number of TRPA1-immunopositive neurons than control rats, P<0.05. Values shown are the mean ± SD. *P<0.05 significantly different from the control group. Abbreviations: SDH, spinal dorsal horn; SG, substantia gelatinosa; TNBS, 2,4,6-trinitrobenzene sulfonic acid; TRPA1, transient receptor potential ankyrin-1. |
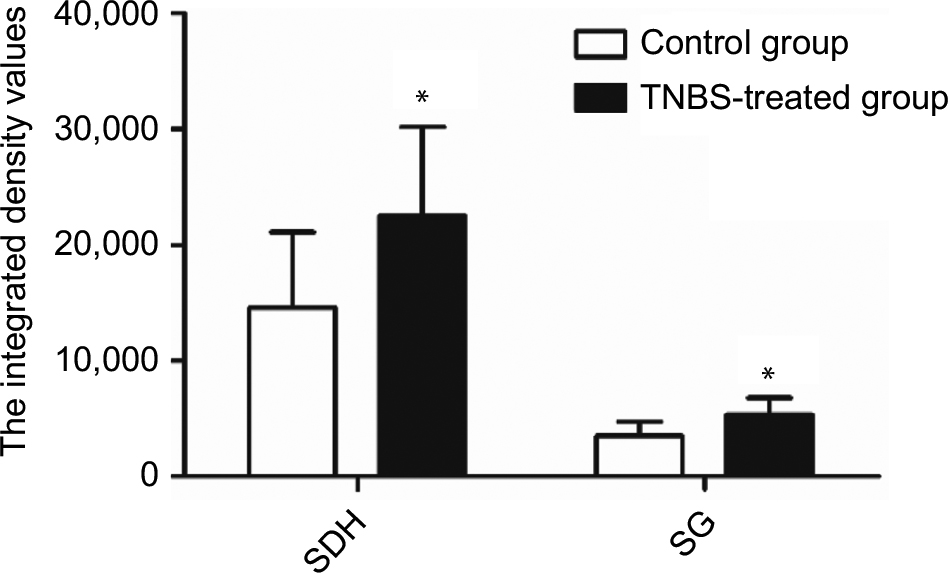 | Figure 5 A IOD values of TRPA1 in the SDH and the SG between the two groups (n=8 per group). Notes: TNBS-treated rats had higher IOD values of TRPA1 than control rats, P<0.05. Values shown are the mean ± SD. *P<0.05 significantly different from the control group. Abbreviations: IOD, integrated optical density; SDH, spinal dorsal horn; SG, substantia gelatinosa; TNBS, 2,4,6-trinitrobenzene sulfonic acid; TRPA1, transient receptor potential ankyrin-1. |
Given these findings, we speculate that upregulation of TRPA1 expression in the colon and SDH after cessation of inflammation is related to persistent post-inflammatory visceral hypersensitivity.
Discussion
Visceral hypersensitivity was assumed as one of the major mechanisms in explaining the chronic abdominal pain in IBS and IBD remission. Increasing amount of evidence suggests that TRPs family members are widely involved in visceral hyperalgesia.27–29 However, there are only limited number of the studies regarding the role and mechanism of TRPA1 with the visceral hypersensitivity,30 and none is extensively involved into both the peripheral and central nervous systems of the pathophysiological aspects.
Here, we established a widely used TNBS-induced rat model of post-inflammatory visceral hypersensitivity to study the features and mechanisms of this disorder. We found that visceral sensitivity was significantly elevated in the TNBS-treated group compared with the vehicle-injected control group, manifesting as a greater number of abdominal contractions, which corresponded to a lower nociceptive threshold. Next, we used Western blotting to quantify the expression of TRPA1 protein in the colon. Colonic TRPA1 expression in the TNBS-treated group was significantly greater than in the control group, suggesting that post-inflammatory visceral hypersensitivity involves changes in the expression of TRPA1. Previously, we also demonstrated that TNBS could induce a significant increase in visceral sensitivity to colonic distension and chemical irritation accompanied by the upregulation of TRPA1 expression in colonic afferent dorsal root ganglia.31 In addition, the contribution of TRPA1 to visceral pain has been defined using TRPA1 knockout mice. Cattaruzza et al32 demonstrated that TRPA1 mediated mechanosensory transduction and participated in visceral hypersensitivity in colonic afferents; intracolonic administration of mustard oil, a TRPA1 agonist, led to visceral mechanical hyperalgesia in wild-type mice but not in TRPA1 knockout mice. However, these studies only investigated peripheral mechanisms of visceral hypersensitivity. Persistent visceral pain is chronic in nature. In the present study, visceral hypersensitivity was still observed in the rats 6 weeks after induction by TNBS, despite there being no microscopic evidence of inflammation in rat colonic tissues by that time. This indicates that central mechanisms contribute to visceral hypersensitivity. Therefore, we also explored TRPA1 protein expression in the spinal cord using Western blotting, which revealed that spinal TRPA1 expression was significantly higher in the TNBS group than in the control group. This suggests that changes in TRPA1 expression in the spinal cord participate in the mediation and maintenance of post-inflammatory visceral hypersensitivity.
The SDH is a relay station for pain information and contains numerous receptors, ion channels, and signal transduction molecules related to pain. Recent study has demonstrated that the expression of P2X3 receptor,33 one of pain receptors, is upregulated in the SDH after an inflammatory injury, and broadly contributes to somatosensory/nociceptive transmission in models of chronic inflammatory pain. Kim et al34 observed dense TRPA1 immunoreactivity in the superficial dorsal horn of the intact side following unilateral dorsal rhizotomy in rats and revealed that TRPA1-mediated nociceptive information is processed in the SDH. With this in mind, we further explored the relationship between TRPA1 expression changes in the SDH and the persistent visceral hypersensitivity. We performed immunohistochemical staining of the SDH in our TNBS-treated and control groups, counting the number of TRPA1-immunopositive neurons and measuring the IOD values of TRPA1. TRPA1 expression was observed in the SDH. Importantly, we show for the first time that TNBS-treated rats had a greater number of TRPA1-immunopositive neurons and higher IOD values of TRPA1 than their vehicle-injected counterparts. Interestingly, this upregulation was mainly located in the superficial layers of the SDH (I and II), especially in the SG. The SG is known to be the first area in the central nervous system to process nociceptive information and plays a pivotal role in the initial modulation and integration of such information. Upregulation of TRPA1 in the SDH (particularly in the SG) could directly result in a greater nociceptive response to peripheral inputs, manifesting as visceral hypersensitivity. In addition, the TRPA1 channel was previously shown to contribute to the transduction of noxious mechanical stimuli in nociceptive C-fiber terminals.35 The SG is the principal projection territory of nociceptive C-fiber primary afferents,36 which suggests that the localization and expression levels of TRPA1 in the SG are closely related to the mediation of nociceptive information. Kosugi et al21 indicated that AITC, a TRPA1 agonist, markedly enhanced excitatory synaptic transmission in SG neurons via the release of more glutamate, a fast excitatory neurotransmitter, to convey sensory signals from the periphery. Cattaruzza et al32 showed that intracolonic administration of TRPA1 agonists, such as mustard oil and 4-hydroxynonenal, significantly increased spinal nociceptive activation, as indicated by a corresponding increase in c-Fos expression in the superficial laminae (I and II) of the SDH in TRPA1+/+, but not in TRPA1–/– mice. These previous studies indicate that TRPA1 also indirectly participates in the mediation of visceral hypersensitivity. Further studies should explore the mechanism of post-inflammatory visceral hypersensitivity by studying the relationship between the upregulation of TRPA1 expressed in the SDH and the release of l-glutamate and spinal c-Fos expression.
However, the cause of TRPA1 upregulation is still not clear. Previous studies have shown that inflammation and nerve damage could be responsible for this upregulation by increasing the release of nociceptive peptides (eg, SP and CGRP) and neurotrophic factors (eg, nerve growth factor).37–39 It therefore seems reasonable to propose the following hypothesis: early inflammatory stimuli activate afferent nerve endings in the gut and increase pain signal transmission, which induces the upregulation of TRPA1 in the SDH. This process directly or indirectly leads to an increase in excitability and responsiveness, which can persist even after the cessation of the noxious stimulus, resulting in persistent post-inflammatory visceral hypersensitivity. Recently, a novel TRPA1 receptor agonist, ASP7663, was reported to be effective in rat models of drug-induced constipation and visceral pain.40 Therefore, revealing the upregulation of TRPA1 in the rat SDH alone is not sufficient to understand the whole maintenance mechanism of visceral hypersensitivity. The causal relationships between TRPA1 expression in the SDH and post-inflammatory visceral pain or hypersensitivity, and spinal c-Fos expression and release of l-glutamate following TRPA1 agonists or antagonists will be explored in future studies.
Conclusion
In summary, our study shows that TRPA1 expression in the SDH (especially in the SG) and the colon was significantly elevated in rats with TNBS-induced visceral hypersensitivity compared with vehicle-injected control rats. This finding supports an important role for TRPA1 in the initiation and maintenance of persistent post-inflammatory visceral hypersensitivity.
Acknowledgments
The authors would like to thank Zhe Wang and Cunhui Jia for technical assistance. The study was granted and supported by National Natural Science Foundation of China (No 30700358).
Author contributions
All authors contributed toward data analysis, drafting, and revising the paper and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
Camilleri M, Madsen K, Spiller R, Greenwood-Van Meerveld B, Verne GN. Intestinal barrier function in health and gastrointestinal disease. J Neurogastroenterol Motil. 2012;24(6):503–512. | ||
Ringel Y, Maharshak N. Intestinal microbiota and immune function in the pathogenesis of irritable bowel syndrome. Am J Physiol Gastrointest Liver Physiol. 2013;305(8):G529–G541. | ||
Jeffery IB, O’Toole PW, Öhman L, Claesson MJ, Deane J, Quigley EM, Simrén M. An irritable bowel syndrome subtype defined by species-specific alterations in faecal microbiota. Gut. 2012;61(7):997–1006. | ||
Farrokhyar F, Marshall JK, Easterbrook B, Irvine EJ. Functional gastrointestinal disorders and mood disorders in patients with inactive inflammatory bowel disease: prevalence and impact on health. Inflamm Bowel Dis. 2006;12(1):38–46. | ||
Kanazawa M, Hongo M and Fukudo S. Visceral hypersensitivity in irritable bowel syndrome. J Gastroenterol Hepatol. 2011;26(Suppl 3):119–121. | ||
Hasler WL and Owyang C. Challenges of managing pain in constipation-predominant IBS: clinical perspectives on antinociceptive actions of linaclotide. Gastroenterology. 2013;145(6):1196–1199. | ||
Han W, Wang Z, Lu X, Guo C. Protease activated receptor 4 status of mast cells in post infectious irritable bowel syndrome. Neurogastroenterol Motil. 2012;24(2):113–119. | ||
Julius D and Basbaum AI. Molecular mechanisms of nociception. Nature. 2001;413(6852):203–210. | ||
Nozawa K, Kawabata-Shoda E, Doihara H, et al. TRPA1 regulates gastrointestinal motility through serotonin release from enterochromaffin cells. Proc Natl Acad Sci U S A. 2009;106(9):3408–3413. | ||
Boesmans W, Owsianik G, Tack J, Voets T, Vanden Berghe P. TRP channels in neurogastroenterology: opportunities for therapeutic intervention. Br J Pharmacol. 2011;162(1):18–37. | ||
Kono T, Kaneko A, Omiya Y, Ohbuchi K, Ohno N, Yamamoto M. Epithelial transient receptor potential ankyrin 1 (TRPA1)-dependent adrenomedullin upregulates blood flow in rat small intestine. Am J Physiol Gastrointest Liver Physiol. 2013;304(4):G428–G436. | ||
Tominaga M, Caterina MJ, Malmberg AB, et al. The cloned capsaicin receptor integrates multiple pain producing stimuli. Neuron. 1998;21(3):531–543. | ||
Lapointe TK, Basso L, Iftinca MC, et al. TRPV1 sensitization mediates postinflammatory visceral pain following acute colitis. Am J Physiol Gastrointest Liver Physiol. 2015;309(2):G87–G99. | ||
Story GM, Peier AM, Reeve AJ, et al. ANKTM1, a TRP-like channel expressed in nociceptive neurons, is activated by cold temperatures. Cell. 2003;112(6):819–829. | ||
Bautista DM, Jordt SE, Nikai T, et al. TRPA1 mediates the inflammatory actions of environmental irritants and proalgesic agents. Cell. 2006;124(6):1269–1282. | ||
Kobayashi K, Fukuoka T, Obata K, et al. Distinct expression of TRPM8, TRPA1, and TRPV1 mRNAs in rat primary afferent neurons with adelta/c-fibers and colocalization with trk receptors. J Comp Neurol. 2005;493(4):596–606. | ||
Nagata K, Duggan A, Kumar G, García-Añoveros J. Nociceptor and hair cell transducer properties of TRPA1, a channel for pain and hearing. J Neurosci. 2005;25(16):4052–4061. | ||
Poole DP, Pelayo JC, Cattaruzza F, et al. Transient receptor potential ankyrin 1 is expressed by inhibitory motoneurons of the mouse intestine. Gastroenterology. 2011;141(2):565–575. | ||
Karashima Y, Talavera K, Everaerts W, et al. TRPA1 acts as a cold sensor in vitro and in vivo. Proc Natl Acad Sci U S A. 2009;106(4):1273–1278. | ||
Jordt SE, Bautista DM, Chuang HH, et al. Mustard oils and cannabinoids excite sensory nerve fibres through the TRP channel ANKTM1. Nature. 2004;427(6971):260–265. | ||
Kosugi M, Nakatsuka T, Fujita T, Kuroda Y, Kumamoto E. Activation of TRPA1 channel facilitates excitatory synaptic transmission in substantia gelatinosa neurons of the adult rat spinal cord. J Neurosci. 2007;27(16):4443–4451. | ||
Dai Y, Wang S, Tominaga M, et al. Sensitization of TRPA1 by PAR2 contributes to the sensation of inflammatory pain. J Clin Invest. 2007;117(7):1979–1987. | ||
Trevisani M, Siemens J, Materazzi S, et al. 4-Hydroxynonenal, an endogenous aldehyde, causes pain and neurogenic inflammation through activation of the irritant receptor TRPA1. Proc Natl Acad Sci U S A. 2007;104(33):13519–13524. | ||
Yu YB, Yang J, Zuo XL, Gao LJ, Wang P, Li YQ. Transient receptor potential vanilloid-1 (TRPV1) and ankyrin-1 (TRPA1) participate in visceral hyperalgesia in chronic water avoidance stress rat model. Neurochem Res. 2010;35(5):797–803. | ||
Jones RC 3rd, Otsuka E, Wagstrom E, Jensen CS, Price MP, Gebhart GF. Short-term sensitization of colon mechanoreceptors is associated with long-term hypersensitivity to colon distention in the mouse. Gastroenterology. 2007;133(1):184–194. | ||
Winston J, Shenoy M, Medley D, Naniwadekar A, Pasricha PJ. The vanilloid receptor initiates and maintains colonic hypersensitivity induced by neonatal colon irritation in rats. Gastroenterology. 2007;132(2):615–627. | ||
Akbar A, Yiangou Y, Facer P, Walters JR, Anand P, Ghosh S. Increased capsaicin receptor TRPV1-expressing sensory fibres in irritable bowel syndrome and their correlation with abdominal pain. Gut. 2008;57(7):923–929. | ||
Zielińska M, Jarmuż A, Wasilewski A, Sałaga M, Fichna J. Role of transient receptor potential channels in intestinal inflammation and visceral pain: novel targets in inflammatory bowel diseases. Inflamm Bowel Dis. 2015;21(2):419–427. | ||
Akbar A, Yiangou Y, Facer P, Brydon WG, Walters JR, Anand P, Ghosh S. Expression of the TRPV1 receptor differs in quiescent inflammatory bowel disease with or without abdominal pain. Gut. 2010;59(6):767–774. | ||
Christianson JA, Bielefeldt K, Malin SA, Davis BM. Neonatal colon insult alters growth factor expression and TRPA1 responses in adult mice. Pain. 2010;151(2):540–549. | ||
Yang J, Li Y, Zuo X, Zhen Y, Yu Y, Gao L. Transient receptor potential ankyrin-1 participates in visceral hyperalgesia following experimental colitis. Neurosci Lett. 2008;440(3):237–241. | ||
Cattaruzza F, Spreadbury I, Miranda-Morales M, Grady EF, Vanner S, Bunnett NW. Transient receptor potential ankyrin-1 has a major role in mediating visceral pain in mice. Am J Physiol Gastrointest Liver Physiol. 2010;298(1):G81–G91. | ||
Xu J, Chu KL, Brederson JD, Jarvis MF, McGaraughty S. Spontaneous firing and evoked responses of spinal nociceptive neurons are attenuated by blockade of P2X3 and P2X2/3 receptors in inflamed rats. J Neurosci Res. 2012;90(8):1597–1606. | ||
Kim YS, Son JY, Kim TH, et al. Expression of transient receptor potential ankyrin1(TRPA1) in the rat trigeminal sensory afferents and spinal dorsal horn. J Comp Neurol. 2010;518(5):687–698. | ||
Kerstein PC, del Camino D, Moran MM, Stucky CL. Pharmacological blockade of TRPA1 inhibits mechanical firing in nociceptors. Mol Pain. 2009;5:19. | ||
Hiura A. Neuroanatomical effects of capsaicin on the primary afferent neurons. Arch Histol Cytol. 2000;63(3):199–215. | ||
Bautista DM, Pellegrino M, Tsunozaki M. TRPA1: a gatekeeper for inflammation. Annu Rev Physiol. 2013;75:181–200. | ||
Obata K, Katsura H, Mizushima T, et al. TRPA1 induced in sensory neurons contributes to cold hyperalgesia after inflammation and nerve injury. J Clin Invest. 2005;115(9):2393–2401. | ||
Chen J, Winston JH and Sarna SK. Neurological and cellular regulation of visceral hypersensitivity induced by chronic stress and colonic inflammation in rats. Neuroscience. 2013;248:469–478. | ||
Kojima R, Nozawa K, Doihara H, Keto Y, Kaku H, Yokoyama T, Itou H. Effects of novel TRPA1 receptor agonist ASP7663 in models of drug-induced constipation and visceral pain. Eur J Pharmacol. 2014;723:288–293. |
 © 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.