Back to Journals » Clinical, Cosmetic and Investigational Dermatology » Volume 12
The regenerative potential of skin and the immune system
Authors Tsepkolenko A, Tsepkolenko V, Dash S ![]() , Mishra A, Bader A, Melerzanov A
, Mishra A, Bader A, Melerzanov A ![]() , Giri S
, Giri S ![]()
Received 29 November 2018
Accepted for publication 22 May 2019
Published 15 July 2019 Volume 2019:12 Pages 519—532
DOI https://doi.org/10.2147/CCID.S196364
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Jeffrey Weinberg
Anna Tsepkolenko,1 Vladimir Tsepkolenko,1 Sabyasachi Dash,2 Apoorva Mishra,3 Augustinus Bader,4 Alexander Melerzanov,3 Shibashish Giri4,5
1Virtus Cliniс, Odessa, Ukraine; 2Department of Pathology and Laboratory Medicine, Weill Cornell Medicine, New York, NY 10044, USA; 3Moscow Institute of Physics and Technology, Dolgoprudny, Moscow Region 141700, Russia; 4Applied Stem Cell Biology and Cell Technology, Biomedical and Biotechnological Center (BBZ), Medical Faculty, University of Leipzig, Leipzig, D-04103, Germany; 5Department of Plastic and Hand Surgery, University Hospital Rechts der Isar, Munich Technical University, Munich, Germany
Abstract: Skin has the natural ability to heal and replace dead cells regulated by a network of complex immune processes. This ability is conferred by the population of resident immune cells that act in coordination with other players to provide a homeostatic environment under constant challenge. Other than providing structure and integrity, the epidermis and dermis also house distinct immune properties. The dermal part is represented by fibroblasts and endothelial cells followed by an array of immune cells which includes dendritic cells (DCs), macrophages, mast cells, NK-cells, neutrophils, basophils, eosinophils, αβ T lymphocytes, B-cells and platelets. On the other hand, the functionally active immune cells in the epidermis comprise keratinocytes, DCs, NKT-cells, γδ T cells and αβ T cells (CD4+ and CD8+). Keratinocytes create a unique microenvironment for the cells of the immune system by promoting immune recognition and cellular differentiation. T lymphocytes exhibit tissue-specific tropism toward the epidermis and the lymphatic drainage system important for their function in immune regulation. This diversity in immune regulators makes the skin a unique organ to overcome pathogenic or foreign invasion. In addition, the highly coordinated molecular events make the skin an attractive model to understand and explore its regenerative potential.
Keywords: skin regeneration, immune system of skin, dendritic cells, macrophages, mast cells, NK cells, neutrophils, basophils, eosinophils, αβ T lymphocytes, B cells
Introduction
Skin is the largest complex organ that provides protection against constantly evolving changes in the environment and pathogenic intrusion.1 The breach of this protective barrier induces fine coordination between cells, molecular factors and matrix remodelers to reestablish and maintain structural and functional integrity.1 The skin’s immune system can be categorized into two parts: the epidermal region as isolated and described by J. Streilin2 and the dermal region with subcutaneous adipose tissue. These two compartments are in close connection with each other further synchronized with the organism’s immune system as a whole. Both possess immune cells from the innate and adaptive immune systems with well-established connections between them. In immunological context, the skin is considered to be a highly diverse yet immunocompetent landscape. It provides the immune system with ideal conditions and an environment to carry out a rapid immune response. Specifically, the integumentary epithelium creates an efficient immunoregulatory microenvironment which forms the basis of the skin’s immune system. In addition to this immune environment, other key players include resident stem cells, with the ability of self-renewal to regulate homeostasis indicative of tissue repair and regeneration.1 For instance, the epithelial stem cells ensure a constant renewal of the epidermis via their regenerative potential. The epidermis then forms an external barrier resembling a filter more than an impenetrable wall. This barrier also houses a rich and diverse microbiota comprising of pathogenic and opportunistic microorganisms, which could be beneficial for the skin microenvironment, according to recent evidence.1 The skin naturally acts a protective barrier against such microorganisms that strive to seize new territories on the host organism. The immune system acts as a deterrent and in the case of microbial aggression intensifies its response.3,4
Infection and stress-related factors activate keratinocytes and immune cells via cascades of cytokine activation.3,4 This activates the proliferation and migration of various immune cell types to the sites of injury or shock. A few examples of such immune responses include the activation of a proinflammatory response (major histompatibilty complex [MHC] and cytokine receptor expression), phagocytosis, and involvement of various immunocompetent cells.3,4 It should be noted that these responses are strong enough to initiate both an innate and adaptive immune response based on the severity of pathogenic invasion or challenge. Immunity defines the ability of multicellular organisms to maintain its macromolecular composition by removing foreign molecules, which in turn provides resistance to infectious agents and neoplastic processes.4 The presented ideas are reflected in one of the latest definitions of immunology which could not have been developed without understanding the role of barrier tissues. Thus, to build an immunological defense system for the body, the existence of a suitable barrier efficient enough to restrict penetration and recognition of foreign invasion is of utmost importance. Importantly, the skin is an ideal candidate that meets these evolutionary requirements. From a pathogen's perspective it is a naturally difficult obstacle to cross due to its dense and durable layer of keratinized epithelium. The skin has many immune cascades in effect at all times, as a result of which it is regarded as one of the emerging model systems to study the immune system.3–5
The dermis
The circulatory system is responsible for uniform distribution of immune cells after their origin from the bone marrow. The population of immune cells comprises of class of T and B lymphocytes including neutrophils, a small number of eosinophils, fibroblasts, dendritic cells (DCs), macrophages, granulocytes, mast cells, basophils, natural killer (NK) cells and natural killer T (NKT) cells. All the aforementioned cells excluding the T- and B-lymphocytes detect and act upon stress signals originating from both foreign and internal stimuli. Pathogenic penetrating microflora are detected by pathogen-associated molecular patterns (PAMPs), and nonpathogenic stress signals are detected via danger-associated molecular patterns (DAMPs). Cells of the innate immune system contain Pathogen Recognizing Receptors (PRR) both on their membranes – Toll-like receptors (TLRs) – and in their cytosols – NOD-like receptors (NLRs). These receptors remain unchanged since the time of birth and hence are associated with innate immunity. All these cells express MHC-I in their dormant state while constitutively expressing MHC-II upon activation. However, in dormancy, induced expression of adhesion molecules on the dermal endothelial cells can trigger proinflammatory response, leading to MHC-II expression in dermal keratinocytes and Langerhans cells (LCs) to recruit leukocytes.5 They also express receptors toward chemokines and cytokines, complement immune system, integrins and other molecules.5–7 The quantity and ability to synthesize certain biologically active molecules differ with each cell and the composition of their receptors. Keeping in mind the diverse properties of various skin immunocytes, in this article,we will discuss the cell populations essential for skin’s immune and regenerative features. The different types of immune cells of epidermis and dermis are presented in Figure 1.
 |
Figure 1 Different types of immune cells such as Langerhans cells, dendritic epidermal γδT cells (DETC), memory αβT cells (TRM), dendritic cell macrophages, neutrophils, mast cells, T-regulatory cells (T-reg) and CD8+ TEM Cells (Effector-Memory cells) are present in epidermis and dermis. |
Fibroblasts
Fibroblasts are immature cells that are highly abundant in the connective tissue. These cells are important in providing a structural basis to the skin. Contemporary history of these cells begins with their discovery by A. Ya. Fridenstein, who observed the ability of cells with a unique morphology to form colonies indicative of their multipotent properties.8 Some of their classical functions include contractility, locomotion, collagen and elastin fiber production and the regulation and degradation of the extracellular matrix.9 Emerging evidence indicates that fibroblasts can express a wide range of functionally active TLRs (from TLR-1 to TLR-9); synthesize antimicrobial peptides (AMPs) (LL-37) and defensins (hBD-1, and hBD-2) and secrete various proinflammatory cytokines (TNFα, INFγ, IL-6, IL-12p70 and IL-10), chemokines (CCL1, CCL2, CCL5, CXCL1, CXCL8, CXCL10 and CX3CL1),and growth factors (granulocyte/macrophage colony-stimulating factor (GM-CSF) and granulocyte colony-stimulating factor (G-CSF), essential for innate immune response against microorganisms.9 Such properties enable these cells to sense pathogens resulting in a coordinated immune response by the recruitment of inflammatory cells.9 Some of the critical immune factors of fibroblasts are presented in Figure 2.
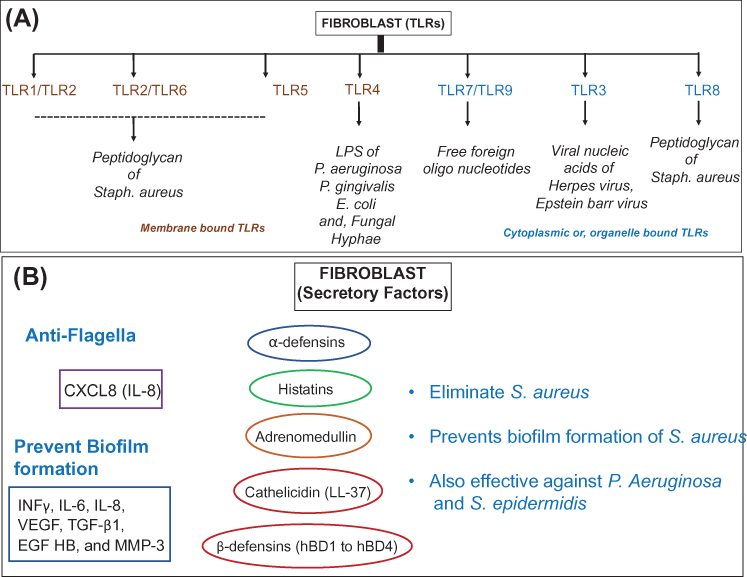 |
Figure 2 The microbial defense armor of fibroblasts. (A) Schematic representation of antimicrobial specificity for various Toll-like receptors (TLRs) expressed in the membrane and cytoplasmic organelles of fibroblasts. (B) Secretory factors responsible for elimination and prevention of microbial growth specifically in dermal and epidermal fibroblasts. |
AMPs serve as important constituents of the host innate immune response. They are known effectors against a wide range of pathogens including bacteria, fungi and viruses. In vitro studies establish the antibacterial properties of cathelicidin (LL-37) against S. aureus and mycobacterial species.9 Other studies also indicate the role β-defensin 3 (hBD-3) in the suppression of S. aureus biofilm formation.10 Additional factors also include cytokines that are secreted by the cells of the innate or adaptive immune system. Studies have shown that skin fibroblasts can synthesize proinflammatory cytokines such as INFγ, IL-6 and IL-8 when exposed to biofilm cultures of S. aureus.10 Another class of secretory factors includes chemokines that serve as recruiting factors for cytokines and facilitate the migration of immune cells at the site of immune response. In a study, it was shown that human gingival fibroblasts are capable of producing secretory IL8 in response to S. typhimurium flagellin-induced TLR5 activation.11
Similarly, fibroblasts are also capable of synthesizing growth factors including vascular endothelial growth factor (VEGF-A). VEGF-A is a factor known to promote inflammatory response-induced neovascularization, allowing recruitment of monocytes/macrophages.12 These studies indicate fibroblasts as one of the important factors responsible towards skin's immune defense system. In addition, fibroblasts also secrete bioactive factors into the intracellular matrix of the connective tissue essential for the maintenance of the cellular environment (Table 1).13 Along this line of thought, the classical role of the fibroblasts has always been perceived for collagen synthesis and maintenance of extracellular matrix including the formation of scar tissue. However, the possibility towards the differentiation ability of fibroblasts into tissue is an emerging and attractive concept.
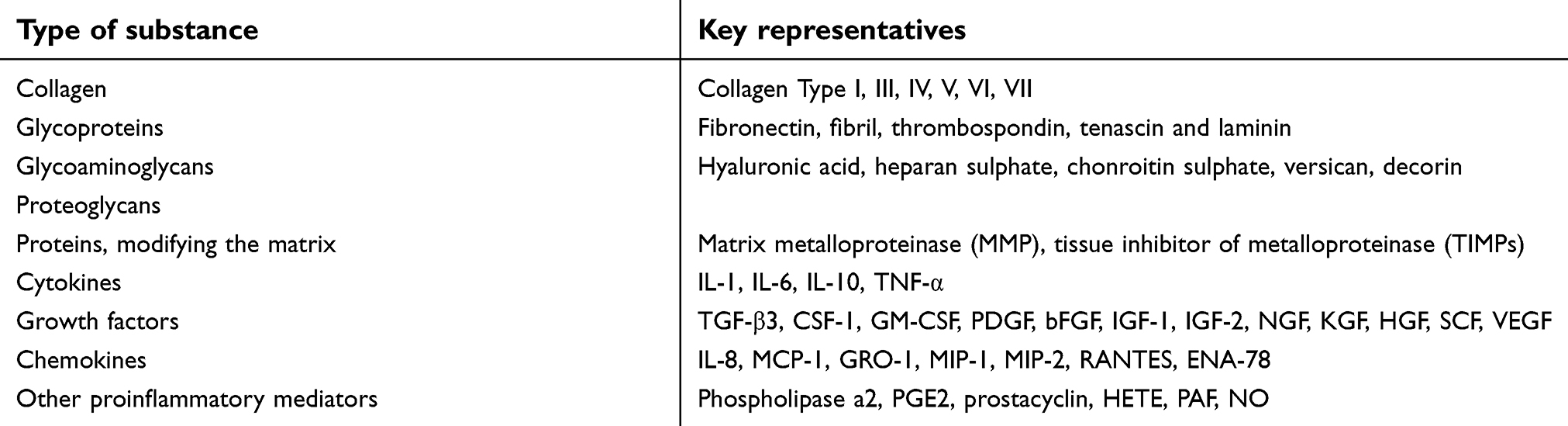 |
Table 1 Components of the intracellular matrix synthesized by dermal fibroblasts in humans |
Considering this idea, research has been conducted to study the criterias for hematopoietic multipotent stromal cells (MSCs) differentiation were determined in fibroblasts by studying features including cellular morphology, adhesiveness and expression of cell surface markers (CD44, CD73, CD90, CD105 and CD271). Additionally, these cells possess the ability to undergo osteogenic, chondrogenic and adipogenic differentiation when grown in specialized media in vitro.14 Depending on the availability of interacting cells and membrane affinity, lymphoid cells can also facilitate this process. The properties mentioned are also inherent in dermal MSCs.15 It was found that MSCs have the ability to interact with lymphocytes and form clusters – fibroblast-lymphocytic rosettes (FLR) – in vitro.14 They constitute majority of the hematopoietic stem cells (HSCs) capable of secreting various cytokines into their microenvironment which helps in establishing immunity as a whole.14,16 MSCs found in the skin resemble those in the bone marrow with the exception of a few unique properties.17 Peripheral MSCs primarily have antiproliferative, immunomodulatory and proinflammatory effects.16 The functionality of MSCs is largely in part due to the presence of TLRs.18 MSCs facilitate phagocytosis in macrophages,19 but inhibit the differentiation and activation of classical monocytes.20 They reduce inflammation, accelerate the elimination of bacteria, promote the conversion of pro-inflammatory macrophages M1 to anti-inflammatory M221 and increase survival during sepsis.22 MSCs have been also observed to induce the differentiation of T-regulatory cells,23 maturation and differentiation of DCs24 and on the other hand can inhibit their migration.25
Recent years have witnessed emerging interest in mechanistic understanding of fibroblast function for the development of novel therapeutic interventions.11 For example, Wnt signaling pathway has been shown to be critical for skin cell differentiation. As a result, Wnt-3a and fibroblast growth factor FGF-9 agonist for Wnt signaling in dermal fibroblasts are sought as therapeutic targets.26 Despite the advances made in the context of fibroblasts in skin healing and regeneration, the field demands further work to fully elucidate the contributions of different dermal fibroblast lineages. Detailed study on the heterogeneity of dermal fibroblasts may yield mechanistic insights into existing therapies and provide cues for developing novel intervention strategies. Thus, we anticipate that the field of regenerative skin biology will witness tremendous advancements, leading to novel therapeutics in the near future.
Dendritic cells
The basic defining properties of DCs are their peculiar branch-like appearance; high MHC-I and MHC-II expression; enhanced active production of cytokines and the ability to capture, process and present antigens. DCs act as critical effectors for T-cell-mediated immune response in the skin. DCs have been shown to produce TNF-alpha- and iNOS-mediated proinflammatory signals in response to bacterial infections. Importantly in psoriasis, dermal DCs expressing CD11c+ surface marker play a major role in the formation of lesions.27
Dermal (myeloid) DCs
Myeloid dermal DCs can exist in the immature state and express various PRRs. After maturing, they migrate toward the lymph node drainage system in the skin where they present the antigen to T cells28 In the dermis, myeloid DCs are located deeper and primarily in the perivasculature which upon myeloid differentiation give rise to two distinct subsets of DCs: (CD141+, CD103+) and (CD1C+, CD11b+).27 Studies conducted in atopic dermatitis (AD) model have shown the activity of myeloid DCs in immune response by initiating recruitment of TH2 immune cells via secretion of chemokine such as CCL22 and CCL17.27 Due to the divergent immunogenic properties, they are often more efficient than the resident macrophages.27 They exist in an intermediate stage within the blood and constitute up to 0.5% of the total number of immunocompetent cells in it. DCs display their membrane affinity toward lymphocytes in situ by forming rosettes around them.29 As part of the innate immune system, they begin to secrete cytokines and chemokines30 and accelerate regeneration.31
Plasmacytoid DCs
Plasmacytoid dendritic cells (pDCs) are specialized type I interferon (IFN-α/β)-producing cells. They express intracellular TLRs TLR7 and TLR9 that mediate recognition of viral nucleic acids.32 In the blood, they resemble plasmacytes and are also categorized under lymphoid cells. These cells comprise a large portion of immature DCs in circulation, with the exception of a small amount that also exists in the dermis. It is here that DCs differ in their functional abilities: they produce x10,000 the amount of IFN-1 during a viral infection as compared to other cells.33 pDCs have been observed to infiltrate both murine and human skin wounds to produce type I IFNs via TLR7- and TLR9-dependent recognition of nucleic acids.32 These events have been accounted critical for the induction of early inflammation and reepithelization of injured skin. pDCs significantly secrete IL-6, an inflammatory cytokine indirectly involved in the re-epithelization of skin wounds.34
Monocytes and macrophages
The mononuclear phagocytic system contains macrophages and monocytes, where the former is found in tissues and the latter in circulation.7,35 Due to the wide range of TLRs, mononuclear cells can recognize practically all the major groups of PAMPs. There are two types of resident macrophage populations in the skin such as the Langerhans cells (LC's) found in the epidermis and dermal macrophages found in abundance in the dermis. LCs originate from progenitors derived from the yolk sac during phases of early embryonic development and have the ability to self-renew.35 LCs are defined by their expression of the lectin receptor langerin (CD 207). In addition, LCs express CD11c and CD11b and are positive for F4/80 and MHC-II surface markers. In contrast, dermal macrophages express F4/80 and CD11b cell surface markers without any expression for CD11c or Langerin followed by low levels of MHC-II.36 The contribution of macrophages during wound healing has been well studied; however, the specific repair function of LCs is yet to be determined.36 Studies have shown that LCs undergo activation during events of wound closure that have been implicated in the healing of foot ulcer in diabetic patients.36
Resident macrophages also occur as a result of constitutive migration of monocytes from the blood. Inflammatory macrophages (M1) are formed that migrate as an emergency response toward the site of inflammation.37 These cells are marked by high phagocytic and bactericidal activity along with high levels of cytokine secretion. These properties enable the M1 macrophages to function as effector cells in the skin’s innate immune response. The classical activation of M1 produces pro-inflammatory cytokines thereby activating antigen presentation, phagocytosis, and the production of growth factors essential for wound repair. Alternatively, M2 activation produces antiinflammatory cytokines, primarily IL-10, and induces type 2 helper T cells (Th2 cells).38 In a model of skin repair, macrophages were found to induce IL-4-dependent collagen fibril assembly following injury.39 Absence of IL-4Ra or the IL-4-associated gene Retnla (resistin-like alpha) reduces the levels of lysyl hydroxylase 2 (LH2), an enzyme responsible for persistent profibrotic collagen cross-links, in injured skin.39 Thus, macrophages harness the potential to produce several important wound-healing and profibrotic mediators including regulation of skin collagen fibril synthesis to maintain the three-dimensional structure.
Similarly, the role of monocytes has been well studied in various stages of wound healing utilizing skin injury models in rodent models. Studies have shown that monocyte infiltration occurs in two distinct phases 24 hrs post wounding in skin. Two unique subsets of monocytes have been identified to infiltrate the site of wound formation. The first subset is marked by the expression of CCR-2 and the second subset is characterized by the expression of chemokine receptor CX3CR-1.36 It was shown that CCR-2-expressing monocytes are predominantly recruited to skin wound 48 hrs post wounding using CCR-2/GFP reporter mice. In addition, there is also a reduction in the number of macrophages at the site of wounding. Other than the early influx of CCR-2-expressing monocytes, an influx of CX3CR-1-expressing monocytes is also observed during subsequent stages of wound healing in the skin. Studies performed using excisional punch biopsy of the skin indicate macrophages as the dominant immune cells expressed during tissue formation phase.40 Additionally, macrophage-derived growth factors also induce extracellular matrix(ECM) deposition.36 For instance, macrophages induce VEGF-mediated neoangiogenesis-mediating formation of neoepithelium at the site of wound bed formation as observed in reporter mice.41 Taken together, these findings demonstrate the critical role of macrophages and their interplay with monocytes during early and late stages of wound healing by regulating the synthesis and organization of de novo collagen depositions in the wound.
Neutrophils
Neutrophils are part of the innate immune system, and these cells carry out a variety of functions during the normal wound repair process 42,43. They are crucial since they migrate from the blood to the site of inflammation significantly faster than monocytes and are the first to fulfill a protective function. Their effectiveness is also owed to them being able to produce the metabolites needed for a “respiratory burst” within seconds. It is important to note that the chemokines are released by activated keratinocytes35 followed by platelets that attract neutrophils to the site of inflammation. Neutrophils recognize pathogenicity via TLR-1, TLR-2 and TLR-10.42 Secretory factors including G-CSF, IL-17 and IL-23 determine the development, migration and activation of neutrophils, and IL-8 (CXCR1 and CXCR2) acts as the main factors of chemotaxis.5,42 Important receptors for the immune system of the skin which determine the link between neutrophils and the skin are CXCL12 (SDF1) and CXCR4.5,42 Granular neutrophils contain several enzymes that enable them to penetrate tissues and destroy particles within phagosomes.42 Classically, neutrophils have been considered as one of the important mediators for wound repair. According to clinical data, patients with defects in neutrophil trafficking or function exhibit higher risk for developing wound infections and impaired wound healing.2 Studies in animal models have also shown that neutrophils are important for cutaneous wound repair.42 Neutrophils also serve as storehouse for matrix metalloproteases (MMPs) in their granules and secretory vesicles. These MMPs include MMP-2, MMP-8 and MMP-9 whose functions have been widely investigated in the context of wound repair. Studies highlight induced expression of MMP-8 (also known as collagenase-2) mRNA levels and activity in chronic wounds.42 In a study performed using MMP-8-knockout mice, researchers observed a delay in wound closure marked by reduced neutrophil infiltration, suggesting that MMP-8 may also aid in neutrophil trafficking.43
Eosinophils
Eosinophils also reside within the dermis and are a good indicator of allergies7 TLR-1, TLR-4, TLR-7 and TLR-10 can be found on eosinophils.7,44 The main role of eosinophils is to combat multicellular parasites by undergoing extracellular cytolysis which releases special granules containing a host of proteins: major basic protein, eosinophil cationic protein (ECP), eosinophilic peroxidase and eosinophil-derived neurotoxin (EDN). ECP and EDN have an antiviral effect which activates mast cells and basophils.7,44–46 However, some of these proteins damage normal tissue as well.45,46 Importantly, eosinophils have been shown to stimulate fibroblasts to thereby induce α-smooth muscle actin and collagen secretion in rodent models of fibrosis.47 Presence of eosinophils facilitates the secretion of growth factors that promote keratinocyte migration in vitro, suggesting a possible role in the formation of dermal scars.47
Basophils and mast cells
Another defense against multicellular parasites is pro-inflammatory basophils and mast cells that express TLR-2, TLR-3 and TLR-4.46,48 Basophils belong to the group of granulocytes which constitute rather short-lived effector cells of the innate immune system. These cells can live from a few months to years retaining their proliferative ability. It has been reported that basophils can regulate the entry of eosinophils into the skin layers via induction of VCAM-1 expression on endothelial cells.49 Furthermore, basophils exhibit anti-inflammatory properties in models of UV-B-induced skin.49 Similarly, increased population of basophils have also been found in skin biopsies from patients with various skin disorders, suggesting their role as an inflammation regulator in human skin.49
Mast cells belong to the population of resident immune cells that undergo activation in response to tissue injury, thereby releasing granules into the extracellular space.50,51 Mast cells react immediately at the site of inflammation by secreting histamines, serotonin, heparin and a host of enzymes. They are the main effector cells that mediate hypersensitivity. Mastocytes secrete a number of growth factors (vascular/endothelial, fibroblast and nerve).50 They display a pronounced affinity toward lymphocytes in vitro as seen by rosette formations (MLR).51 Upon activation, mast cells acquire the ability of antigen presentation.50,51 Mast cells have also been found to secrete TGF-β1, including other proteases that promote fibroblast-mediated effects and collagen fibril formation.47 Mast cells also drive the formation of gap junctions with fibroblasts that are essential for the stimulation of cellular differentiation, proliferation and ECM contraction.47
NK lymphocytes
The morphology of NK cells is that of large granular lymphocytes that lack T- and B-lymphocyte markers. NK cells are directed to carry out cytolysis and produce cytokines (primarily IFN) upon contact with tumor cells and cells with viral infections. These cells express many different TLRs. They also possess various receptors that identity and react with molecules sent out by dead or stressed cells (NKG2D-MYC) which enable them to directly affect the process of regeneration. Upon inflammation, NK cells migrate to the skin expressing chemokine receptors that correspond to ligands in keratinocytes.52,53
NKT lymphocytes
NKT lymphocytes are cells of the innate immune system though they develop in the thymus.5,54,55 Unlike the T cells which carry CD4+ or CD8+ receptors, these cells only express an invariant Valpha-14 rearrangement receptor.5,7 Typical molecules associated with NK cells (NK 1.1, NKR-P1, CD56, CD16, KIR and NKG2D) are only found in the periphery. It is the Valpha-14 rearrangement receptor that allows the NKT lymphocytes to recognize glycolipid antigens with respect to antigen-presenting molecule CD1d (MHC – I).54 Among lymphocytes in the blood, NKT constitutes only 1% of the population, and at an even lesser quantity in the dermis. These cells are extremely valuable because other than their cytotoxic function, they are practically the only source of cytokines (primarily IFNγ) in the initial stages of a defense against pathogens.55 NKT cells have been observed to produce cytokines such as IL-4 and IFN-γ that facilitate resident antigen-specific CD8+T-cell and dendritic cell function.5
Platelets
Currently, the functionality of these cells has come from an unexpected and novel direction. Apparently, platelets participate in inducing inflammation, regeneration and the formation of an immune reaction (TLR-2, TLR-4, TLR-7 and TLR-9).56 They produce a majority of the growth factors and other biologically active substances. Data show that granular platelets contain 827 proteins,57 whose secretion creates a platform for interactions between platelets and majority of other immune cells and stromal cells. Platelets activate neutrophils and monocytes via P selectin on their membranes, and their complementary ligand P selectin glycoprotein ligand 1 (PSG-L-1).58 Neutrophils, monocytes, macrophages and endothelial cells are greatly influenced by the following chemokines: CXCL4, CXCL7, β-thromboglobulin, neutrophil-activating peptide (NAP), CCL3 (macrophage inhibitory protein-1α, MIP 1a), CCL5 (RANTES), CXCL1 (GROa) and CXCL5.59 Thrombocidin 1 and 2 realize any microbial threat and growth factors (PDGF, TGF-β, EGF, and VEGF) cause an angiogenic and reparative action as part of the response.60 Effective recruitment of platelets is essential for wound healing.61 The use of platelet-rich autoplasma (PRP) has clinical application in wound healing and tissue regeneration in muscoskeletal injuries and their subsequent complications.62 The introduction of PRP has been successfully used in dermatology, cosmetology, obstetrics, gynecology, cervical diseases, leukoplakia, endocervicitis, cervical erosion and kraurosis of the vulva.62
Epithelial and endothelial cells
Proinflammatory cytokines released by immunocytes of the innate immunity upon exposure to tissue pathogens can induce a reaction from nontraditional cells of the immune system (epithelial and endothelial cells) as they contain PAMP and proinflammatory cytokine receptors. These cells begin to imitate traits of innate immunocytes: adhesion molecules, complimentary Fcγ-receptors, MHC-II molecules, costimulators (CD40, CD80, and CD86) and secretion of proinflammatory cytokines (IL-1, IL-6, TNFα). Although these nontraditional cells have significantly inferior signals in comparison, they are enough to act as APCs to memory cells, αβ T lymphocytes and B-lymphocytes in the skin.63
The epidermis
The epidermis is marked by characteristic features and occupies a distinct area.1 The passage of cells in this compartment is monitored by membrane-bound adhesion molecules that interact with complementary structures found on keratinocytes. An equally important role is played by chemokine receptors secreted by keratinocytes. The specificity of the latter represents the skin as an organ of the immune system. The epidermis is composed of keratinocytes, LCs, β T-cells and αβ T-lymphocytes; among which are present a significant amount of CD4+ and CD8+ lymphocytes with a predominance of memory cells.1 Interesting enough, T lymphocytes comprise 90% of all lymphocytes in the epidermis.
Keratinocytes
Keratinocytes constitute the structural element of the outer layer of the skin. Other than their role as a structural component, emerging evidence highlight their role in both innate and adaptive immunity.5 Keratinocytes can be rightly called a part of the innate immune system since they occupy a strategic place as the primary physical barrier against pathogens. Keratinocytes together with neutrophils and epithelial cells serve as one of the reservoirs of AMPs that serve as the first line of defense.5 They also express certain pathogen pattern receptors including TLRs. These cells are also capable of secreting immune factors and chemoattractants as depicted in Figure 3. Prolonged activation of TLRs in the keratinocytes causes the synthesis of IFN1 and the polarization of a Th1 reaction.64
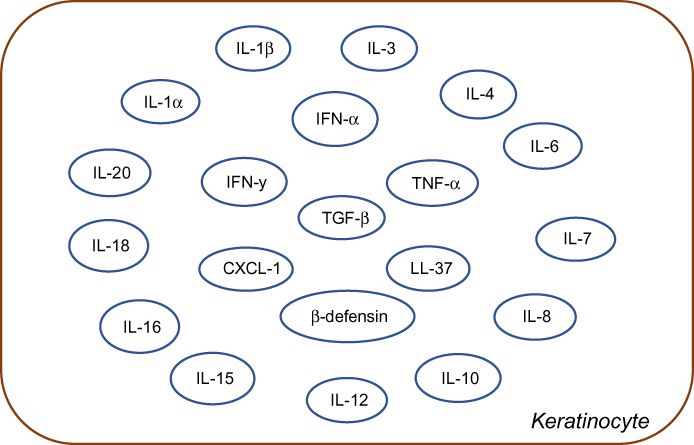 |
Figure 3 Schematic representation of various cytokines and chemokines secreted by keratinocytes in response to immune activation in skin injury. IFN represents Interferon and IL's represent Interleukin family of cytokines, LL-37 is an important anti-microbial cathelicidin molecule whereas, CXCl, TGF-beta, TNF-alpha and beta-defensins are indicative of predominant chemoattractants. |
Keratinocytes can recruit, stimulate and activate multiple cell types in healing process to regain the epidermal structure. Upon epidermal damage, keratinocytes release IL-1, which acts in both autocrine and paracrine cascades to further that activate and facilitate migration and proliferation of keratinocytes to improve injury-induced healing.65 Keratinocytes also secrete VEGF and platelet-derived growth factor (PDGF),65 which are critical for endothelial cell migration and angiogenesis in the wound bed.65 Studies have also shown that keratinocytes can also secrete PDGF to promote fibroblast proliferation and production of extracellular matrix.8,64,65 One can imagine that the synchronous activity of fibroblasts and keratinocytes is important for maintaining skin-tissue homeostasis. It is this cross talk that leads to the recruitment of necessary cellular secretory factors for efficient wound closure.65 Our understanding of the function of keratinocytes has also led to the development of bioengineered products to promote skin-tissue regeneration. Upon topical application, the combination of keratinocyte and fibroblasts aids in secretion of growth factors, cytokines and matrix proteins to promote healing and regeneration. Commercially available therapies include products such as EpiCel, Dermagraft and Apligraf.66
Defective function of some antimicrobial peptides such as cathelicidin and β-defensins has been implicated in increased predisposition to skin infections and psoriatic lesions5 One of these cathelicidins, LL-37, is also produced by keratinocytes and has an essential role in promoting angiogenesis and wound healing.67 Keratinocytes also express TLRs on their cell surface and on the surface of their endosomal vesicles. Keratinocytes are able to generate proinflammatory cytokines such as IL-1β and IL-18 via inflammasome signaling pathway.68 IL-1 produced by keratinocytes can upregulate expression of intercellular adhesion molecule (ICAM)-1. Upregulation of MHC class II on keratinocytes along with LCs facilitates leukocyte trafficking into the skin, hallmark of skin injury response. In disease conditions, keratinocytes express several chemokine ligands such as CXCL9, CXCL10, CXCL11 and CCL20.69 For instance, ligands such as CXCL1 and CXCL8 signal neutrophils for epidermal infiltration.69 Similarly, studies also show that keratinocytes possess the ability to induce T-cell activation or antigen-specific tolerization.
Langerhans cells
Another type of cells that are in close proximity to the epidermis are the LCs that are related to myeloid cells (histologically they are classified as pigmented epidermal cells). Initially, they were classified by a distinct organelle in their cytoplasm – Birbeck granules.70 LCs are scattered throughout the epidermis which enables them to monitor the penetration of the epidermis by foreign materials. LCs are identified by their expression of langerin (CD207) and CD1a.71 The receptor langerin recognized mannosylated ligands on the surface of a wide range of PAMPs. Upon contact, the processing of an antigen begins with its displacement via exocytosis by forming a complex with CD1a and langerin in Birbeck granules. Presenting antigen is the primary function of DCs, whose effectiveness is far superior to other cells, including macrophages. Epidermal LCs have a few unique properties. They express specific proteinases including MMPs which enable them to penetrate through the basal membrane and reach the lymphatic pathway. Expression of MMP8 in LC inhibits TNFα and IL-1β, and its migration can prevent the inhibition of MMP and antibodies to MMP9 and MMP2.53,72
γδ T cells
The epidermis contains a specific subpopulation of lymphocytes - γδ T cells that contain TCR containing γ and δ chains.73 Usually, these cells are generated in the thymus during embryonic development, but a small number are developed elsewhere and survive in the periphery. After their synthesis, they comprise only about 1% of all cells in the thymus and 5% of the periphery. γδ T cells selectively reside in barrier tissues, particularly the epidermis. In mice, the migration of these cells from the thymus begins on the 14th day of embryogenesis. The tropism of these cells does not vary with species, but certain unique features in the cells do exist. These cells in the epidermis of mice acquire a dendritic morphology (dendritic γδ T cells). The epidermal localization of γδ T cells is mainly due to their chemokine receptor expression, synthesized from keratocyte chemokines. More than 80% of cells express CCR5 which causes their migration to the site of inflammation.73 γδ T cell receptors have a narrow antigen-recognizing repertoire. They have the ability to recognize and respond to antigens without the main histocompatibility complex and costimulating molecules74 due to the presence of TLR-1 and TLR-3 that recognize PAMPs. Inflammation causes an increased expression of NLR-4. γδ lymphocytes can strengthen anti-infective resistance by being one of the main types of cells that produce IL-17 by migrating to the dermal region postbacterial infection or injury.74,75 They also reduce the activity of delayed-type hypersensitivity effector cells and their production of IFNγ and inhibit the activity of αβ lymphocytes in allergic reactions.76 γδ T cells activate DC and lead to their maturation and cytokine production.77 Due to the presence of NKG2D and NKG2 CD94, γδ lymphocytes are able to recognize stress molecules after which they locally synthesize chemokines, cytokines, growth factors and cytokine effectors including IFN-γ.73–75 These can regulate inflammation, cellular cytotoxicity and wound healing74 γδ lymphocytes interact with keratinocytes and support their survival and functionality which contribute to the regeneration of the epidermis when damaged.78 They are among the first cells to react to skin trauma79 and change their morphology within minutes. They express activation markers (CD69) and produce chemokines attracting other inflammation cells to the matrix. In many ways, the reaction is maintained by mitogens secreted by keratinocytes and the subsequent response of the epithelial cells by ligand expression for γδ lymphocytes.80 Important factors stimulating epithelial regeneration are keratinocyte growth factors: KGF1, KGF2, IRT1 and insulin -ike growth factor (IGF).81 The importance for skin regeneration by γδ lymphocytes indicates that the isolation of prolonged wounds of these cells is anergic and unable to produce IGF1.
αβ T lymphocytes
αβ T lymphocytes appear at the epidermis from circulating blood by the attraction of synthesized keratinocyte chemokines (CTACK) and the intercellular interaction of general adhesives – selectin and integrins – with the endothelial receptors (CD34, GlyCAM-1, ICAM1, ICAM2, VCAM1 and MadCAM1).82 Tropism of classical T lymphocytes to the epidermis in significant quantities is determined by the expression of so-called cutaneous lymphocyte antigen.83 Practically all αβ T lymphocytes belong to memory cells.84
Differences in immune cell types during skin repair
All cells of the immune system take part during the reparative process in the regeneration of skin. Their individual roles change depending on the phase. Damage to the skin causes an inflammatory response. The first reaction is the synthesis of proinflammatory cytokines (IL-1, IL-6, TNFα, IFNγ, etc.), chemokines, receptors and adherent molecules that activate keratinocytes. This is followed by the activation of platelets, endothelial cells, mast cells and NKT cells.85 Within the first few days, chemokines attract a large quantity of neutrophils42 to the wound, followed by its replacement with phagocytic proinflammatory M1 macrophages.86 The act of inflammation is linked to the effect of proinflammatory cytokines, by the transition of M2 macrophages (anti-inflammatory) to M1 phenotype (proinflammatory),87 mast cells, Th2 and T-regulatory cells (TREG). γδ T cells work in a similar manner by secreting factors that stimulate and maintain keratinocytes (IGF-1 and KGF1 and KGF2 growth factors). αβ and γδ T cells are important for the maintenance and homeostasis of the skin via recognition and clearance of stressed or damaged cells.88 Current studies lack the distinct roles of αβ and γδ cells in skin immunity. However, it is certain that their role in immune mechanisms is conserved.88 γδ T cells can recognize a wide variety of peptide and nonpeptide antigens released as a result of constant changes in the skin, while αβ T cells recognize pathogen-specific peptides or tumors presented by MHC molecules.88
Fibroblasts play one of the main roles in the vivification of damaged skin. In a wounded area, the surface of these cells is attracted to the growth factors released by platelets (PDGFs). However, in such conditions, the fibroblasts proliferate and synthesize matrix molecules that are not very effective. It has been shown that in the presence of immune cells the performance of fibroblasts is greatly enhanced89 At the same time, HSC, MSC and endothelial precursors mobilize from the bone marrow.90 Under the influence of the chemokines, stem cells migrate to the wound site and differentiate and proliferate along with resident elements.91 This is followed by angiogenesis, granulation and epithelialization. Effective remodeling occurs in the event of sufficient intervention of MSC, endothelial precursors and embryonic stem cells. Of great importance in this context are cells synthesizing MMPs and extracellular matrix molecules.
It has been shown that keratinocytes and immune cells are capable of secreting steroidal hormones de novo.83 It is possible that this may be limited only to the inflammatory phase: catecholamine92 and acetylcholine. Anti-inflammatory effects are apparently similar to those of the parasympathetic nervous system and influence acetylcholine receptors (nAchR) which are located on the membranes of immune cells.93,94 It is through these receptors that the synthesis of an extracellular matrix by keratinocytes is facilitated along with factors that influence differentiation. The activation of β-adrenergic receptors also has an influence on the migration and differentiation of keratinocytes.
Conclusion and future remarks
Although the immune system of the skin is a nonstructured diffused lymphoid tissue, it has a distinct spatial and functional organization that is unique to it. Its cellular composition is distinguished by a huge variety of cellular components belonging to both the innate and adaptive immunity. Our current knowledge on the cellular immune repertoire of the skin highlights its importance. In the era of emerging awareness toward cancer diagnosis, autoimmune disorders, aging and vaccine development, the current and emerging knowledge on skin’s immune competence holds greater significance. Immune cells of the skin are broadly categorized based on their prevalence in the two distinct layers. It is not as specific in terms of cellular composition pertaining to the dermis and subcutaneous adipose tissue. Various cells from within circulation accumulate and become activated within this space. Classical T- and B lymphocytes and their subpopulations in their absolute and relative state quantitatively correspond to that in the periphery.5,7 This depends on NK and NKT lymphocytes and circulating myeloid cells: neutrophils, basophils, eosinophils and monocytes.53–55
Progenitors of mast cells, myeloid cells and lymphoid DC travel through the bloodstream and migrate to the dermis where they mature as a result of resident elements. The predominant tissues in the bone marrow are MSCs and fibroblasts. MSCs from hair follicles arising from the neural crest are the closest to this sort judging by their properties. It Is worth a mention to state that the quantity of platelets increases during an immune response due to their special properties. The dermis is the structural and functional backbone of the epidermis. It provides access to cells within circulation which are strictly regulated. These selective cells can practically only be found within this area. This is primarily keratinocytes, LC and γδ T lymphocytes. The last two function as part of the innate as well as the adaptive immunity, which links both the skin compartments with the entire body. This is required to present antigens traveling through the lymphatic path in the dermis followed by the lymphatic follicle. γδ T lymphocytes support keratinocytes and promote their regeneration.
Of the classical lymphocytes, only those with the appropriate tropism are found in the epidermis. This is due to the expression of adhesion molecules in the epidermis, which is attracted to chemokines released by keratinocytes from γδ T lymphocytes. B lymphocytes, on the other hand, are absent. The overwhelming majority of the population are memory cells.
Cells of the innate immune system are activated by PRR that recognizes pathogenic and stress signals (PAMP and DAMP). “Intermediate” cells viz. NKT and γδ T lymphocytes express PRRs as do T-cell receptors, though the latter has a different structure and limited capabilities. Upon activation, many cells express high levels of MHC-I and monocyte chemoattractant protein 1 (MCP1), cytokines and their receptors, chemokines and growth factors. This cytokine network when in contact with the many available cell types triggers the required immune response. Both sections of the immune system participate in the regeneration of the skin and its cellular elements. They are also involved in the process of specialization and its various stages. Keratinocytes, platelets, γδ T lymphocytes and mast cells can induce inflammation by attracting phagocytes against any infection and activate mechanisms to trigger adaptive immunity. Attenuation of inflammation converts the proinflammatory M1-Mø into the anti-inflammatory M2-Mø. It also marks the beginning of the proliferative phase of fibroblasts and stem cells in various regions, both local and in the bone marrow, thereby regulating their immune-cooperative interaction and the process of remodeling multichannel tissue design features.
In-depth understanding on the contribution of skin-resident and skin-infiltrating immune populations bolstered by their cross-talk with other cell types can open avenues toward the development of novel therapeutics. Many questions remain unanswered in this emerging area of skin immunology and skin’s regeneration potential. For instance, the establishment of experimental in vitro human and induced pluoripotent stem cell-based systems will allow assessment of the functional capacity of skin’s immune cell repertoire in the context of immune response and their alterations in skin diseases. Further, use of lineage-tracing concepts in such regenerative experimental model systems will enhance our understanding of molecular events and trigger factors that are responsible for immune cell trafficking to sites of regeneration, post skin injury. Taken together, we conclude that near future discoveries using such innovative strategies will not only help us achieve better therapeutic products for skin-related immune disorders but will also foster ideas toward novel cosmetic formulations and topical applications for improving skin’s regenerative potential.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Gonzales KAU, Fuchs E. Skin and its regenerative powers: an alliance between stem cells and their niche. Dev Cell. 2017;43(4):387–401. doi:10.1016/j.devcel.2017.10.001
2. Streilein JW. Lymphocyte traffic, T-cell malignancies and the skin. J Invest Dermatol. 1978;71(3):167–171. doi:10.1111/1523-1747.ep12547071
3. Cogen AL, Nizet V, Gallo RL. Skin microbiota: a source of disease or defence? Br J Dermatol. 2008;158(3):442–455. doi:10.1111/j.1365-2133.2008.08437.x
4. Grice EA, Segre JA. The skin microbiome. Nat Rev Microbiol. 2011;9(4):244. doi:10.1038/nrmicro2537
5. Matejuk MA. Skin Immunity. Arch Immunol Ther Exp (Warsz). 2017;66(1):45–54. doi:10.1007/s00005-017-0477-3
6. de Koning HD, Simon A, Zeeuwen PLJM, Schalkwijk J. Pattern recognition receptors in infectious skin diseases. Microbes Infect. 2012;14(11):881–893. doi:10.1016/j.micinf.2012.03.004
7. Janeway CA
8. Friedenstein AJ, Gorskaja JF, Kulagina NN. Fibroblast precursors in normal and irradiated mouse hematopoietic organs. Exp Hematol. 1976;4(5):267–274.
9. Bautista-Hernández LA, Gómez-Olivares JL, Buentello-Volante B, Bautista-de Lucio VM. Fibroblasts: the unknown sentinels eliciting immune responses against microorganisms. Eur J Microbiol Immunol (Bp). 2017:7(3):151–157. Published 2017 Aug 19. doi:10.1556/1886.2017.00009.
10. Huang Q, Fei J, Yu HJ, Gou YB, Huang XK. Effects of human β-defensin-3 on biofilm formation-regulating genes dltB and icaA in Staphylococcus aureus. Mol Med Resp. 2014;10:825–831.
11. Mahanonda R, SardAIam N, Montreekachon P, et al. IL-8 and IDO expression by human gingival fibroblasts via TLRs. J Immunol. 2007;178(2):1157. doi:10.4049/jimmunol.178.2.1151
12. Kirker KR, James GA, Fleckman P, Orelund JE, Stewart PS. Differential effects of planktonic and biofilm MRSA on human fibroblasts. Wound Repair Regen. 2012;20:253–261. doi:10.1111/j.1524-475X.2012.00769.x
13. Stappers MHT, Thys Y, Oosting M, et al. TLR1, TLR2, and TLR6 gene polymorphisms are associated with increased susceptibility to complicated skin and skin structure infections. J Infect Dis. 2014;210(2):311–318. doi:10.1093/infdis/jiu080
14. Dominici MLBK, Le Blanc K, Mueller I, et al. Minimal criteria for defining multipotent mesenchymal stromal cells. The International society for cellular therapy position statement. Cytotherapy. 2006;8(4):315–317. doi:10.1080/14653240600855905
15. Janson D, Rietveld M, Mahé C, Saintigny G, El Ghalbzouri A. Differential effect of extracellular matrix derived from papillary and reticular fibroblasts on epidermal development in vitro. Eur J Dermatol. 2017;27(3):237–246. doi:10.1684/ejd.2017.2984
16. Ma T, Wang X, Jiang D. Immune tolerance of mesenchymal stem cells and induction of skin allograft tolerance. Curr Stem Cell Res Ther. 2017;12(5):409–415. doi:10.2174/1574888x12666170301122744
17. Kühbacher A, Henkel H, Stevens P, et al. Dermal fibroblasts play a central role in skin model protection against C. albicans invasion. J Infect Dis. 2017;215. doi:10.1093/infdis/jix153.
18. Wang X, Cheng Q, Li L, et al. Toll-like receptors 2 and 4 mediate the capacity of mesenchymal stromal cells to support the proliferation and differentiation of CD34+ cells. Exp Cell Res. 2012;318(3):196–206. doi:10.1016/j.yexcr.2011.11.001
19. Jackson MV, Morrison TJ, Doherty DF, et al. Mitochondrial transfer via tunneling nanotubes is an important mechanism by which mesenchymal stem cells enhance macrophage phagocytosis in the in vitro and in vivo models of ARDS. Stem Cells. 2016;34(8):2210–2223. doi:10.1002/stem.2372
20. Rocher BD, Mencalha AL, Gomes BE, Abdelhay E. Mesenchymal stromal cells impair the differentiation of CD14++ CD16− CD64+ classical monocytes into CD14++ CD16+ CD64++ activate monocytes. Cytotherapy. 2012;14(1):12–25. doi:10.3109/14653249.2011.594792
21. Abumaree MH, Al Jumah MA, Kalionis B, et al. Human placental mesenchymal stem cells (pMSCs) play a role as immune suppressive cells by shifting macrophage differentiation from inflammatory M1 to anti-inflammatory M2 macrophages. Stem Cell Rev Rep. 2013;9(5):620–641. doi:10.1007/s12015-013-9455-2
22. Mei SHJ, Haitsma JJ, Dos Santos CC, et al. Mesenchymal stem cells reduce inflammation while enhancing bacterial clearance and improving survival in sepsis. Am J Respir Crit Care Med. 2010;182(8):1047–1057. PMID: 20558630. doi:10.1164/rccm.201001-0010OC.
23. Burr SP, Dazzi F, Garden OA. Mesenchymal stromal cells and regulatory T cells: the Yin and Yang of peripheral tolerance? Immunol Cell Biol. 2013;91(1):12–18. doi:10.1038/icb.2012.60
24. Asadi M, Farokhi F, Delirezh N, Ganji Bakhsh M, Nejati V, Golami K. Fibroblast and T cells conditioned media induce maturation dendritic cell and promote T helper immune response. Veterinary research forum : an international quarterly journal. 2012;3(2):111–118.
25. English K, Barry FP, Mahon BP. Murine mesenchymal stem cells suppress dendritic cell migration, maturation and antigen presentation. Immunol Lett. 2008;115(1):50–58. doi:10.1016/j.imlet.2007.10.002
26. Gay D, Kwon O, Zhang Z, et al. Fgf9 from dermal γδ T cells induces hair follicle neogenesis after wounding. Nat Med. 2013;19(7):916–923. doi:10.1038/nm.3181
27. Kashem SW, Haniffa M, Kaplan DH. Antigen-presenting cells in the skin. Annu Rev Immunol. 2017;35:469–499. doi:10.1146/annurev-immunol-051116-052215
28. Dos Santos VG, Orfali RL, de Oliveira Titz T, Da Silva Duarte AJ, Sato MN, Aoki V. Evidence of regulatory myeloid dendritic cells and circulating inflammatory epidermal dendritic cells‐like modulated by Toll‐like receptors 2 and 7/8 in adults with atopic dermatitis. Int J Dermatol. 2017;56(6):630–635. doi:10.1111/ijd.13537
29. Kyewski BA, Momburg F, Schirrmacher V. Phenotype of stromal cell‐associated thymocytes in situ is compatible with selection of the T cell repertoire at an “immature” stage of thymic T cell differentiation. Eur J Immunol. 1987;17(7):961–967. doi:10.1002/eji.1830170711
30. Khasawneh A, Baráth S, Medgyesi B, et al. Myeloid but not plasmacytoid blood DCs possess Th1 polarizing and Th1/Th17 recruiting capacity in psoriasis. Immunol Lett. 2017;189:109–113. doi:10.1016/j.imlet.2017.04.005
31. Han Z, Chen Y, Zhang Y, et al. MiR‐21/PTEN axis promotes skin wound healing by dendritic cells enhancement. J Cell Biochem. 2017;118(10):3511–3519. doi:10.1002/jcb.26026
32. Gregorio J, Meller S, Conrad C, et al. Plasmacytoid dendritic cells sense skin injury and promote wound healing through type I interferons. J Exp Med. 2010;207(13):2921–2930. doi:10.1084/jem.20101102
33. Jacquemin C, Rambert J, Guillet S, et al. Heat shock protein 70 potentiates interferon alpha production by plasmacytoid dendritic cells: relevance for cutaneous lupus and vitiligo pathogenesis. Br J Dermatol. 2017;177(5):1367–1375. doi:10.1111/bjd.15550
34. Gallucci RM, Sloan DK, Heck JM, Murray AR, O’Dell SJ. Interleukin 6 indirectly induces keratinocyte migration. J Invest Dermatol. 2004;122:764–772. doi:10.1111/j.0022-202X.2004.22323.x
35. Hoeffel G, Wang Y, Greter M, et al. Adult Langerhans cells derive predominantly from embryonic fetal liver monocytes with a minor contribution of yolk sac-derived macrophages. J Exp Med. 2012;209(6):1167–1181. doi:10.1084/jem.20120340
36. Minutti CM, Knipper JA, Allen JE, Zaiss DMW. Tissue-specific contribution of macrophages to wound healing. Semin Cell Dev Biol. 2017;61:3–11. ISSN 1084-9521. doi:10.1016/j.semcdb.2016.08.006
37. Mantovani A, Sica A, Sozzani S, Allavena P, Vecchi A, Locati M. The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol. 2004;25(12):677–686. doi:10.1016/j.it.2004.09.015
38. Gerber JS, Mosser DM. Reversing lipopolysaccharide toxicity by ligating the macrophage Fcγ receptors. J Immunol. 2001;166(11):6861–6868. doi:10.4049/jimmunol.166.11.6861
39. Knipper JA, Willenborg S, Brinckmann J, et al. Interleukin-4 receptor α signaling in myeloid cells controls collagen fibril assembly in skin repair. Immunity. 2015;43(4):803–816. doi:10.1016/j.immuni.2015.09.005
40. Lucas T, Waisman A, Ranjan R, et al. Differential roles of macrophages in diverse phases of skin repair. J.Immunol. 2010;184(7):3964–3977. doi:10.4049/jimmunol.0903356
41. Willenborg S, Lucas T, vanLoo G, et al. CCR2 recruits an inflammatory macrophage subpopulation critical for angiogenesis in tissue repair. Blood. 2012;120(3):613–625. doi:10.1182/blood-2012-01-403386
42. Wilgus TA, Roy S, McDaniel JC. Neutrophils and wound repair: positive actions and negative reactions. Adv Wound Care. 2013;2(7):379–388. doi:10.1089/wound.2012.0383
43. Gutiérrez-Fernández, Inada M, Balbin M, et al. Increased inflammation delays wound healing in mice deficient in collagenase-2 (MMP-8). FASEB J. 2017;21(10):2580–2591. doi:10.1096/fj.06-7860com
44. Skrzeczynska-Moncznik J, Zabieglo K, Bossowski JP, et al. Eosinophils regulate interferon alpha production in plasmacytoid dendritic cells stimulated with components of neutrophil extracellular traps. Int J Interferon Cytokine Res. 2017;37(3):119–128. doi:10.1089/jir.2016.0036
45. Lamback EB, Resende FSS, Lenzi TCR. Eosinophilic fasciitis. An Bras Dermatol. 2016;91(5):57–59. doi:10.1590/abd1806-4841.20164683
46. Otsuka A, Kabashima K. Contribution of basophils to cutaneous immune reactions and Th2-mediated allergic responses. Front Immunol. 2015;6:393. doi:10.3389/fimmu.2015.00393
47. Larouche J, Sheoran S, Maruyama K, Martino MM. Immune regulation of skin wound healing: mechanisms and novel therapeutic targets. Adv Wound Care. 2018;7(7):209–231. doi:10.1089/wound.2017.0761
48. Schwartz C, Eberle JU, Voehringer D. Basophils in inflammation. Eur J Pharmacol. 2016;778:90–95. doi:10.1016/j.ejphar.2015.04.049
49. Voehringer D. Recent advances in understanding basophil functions in vivo. F1000Res. 2017;6:1464. Published 2017 Aug 15. doi:10.12688/f1000research.11697.1
50. Cardamone C, Parente R, Feo GD, Triggiani M. Mast cells as effector cells of innate immunity and regulators of adaptive immunity. Immunol Lett. 2016;178:10–14. doi:10.1016/j.imlet.2016.07.003
51. Igawa S, Nardo AD. Skin microbiome and mast cells. Transl Res. 2017;184:68–76. doi:10.1016/j.trsl.2017.03.003
52. Bonefeld CM, Geisler C. The role of innate lymphoid cells in healthy and inflamed skin. Immunol Lett. 2016;179:25–28. doi:10.1016/j.imlet.2016.01.005
53. Levin C, Bonduelle O, Nuttens C, et al. Critical role for skin-derived migratory DCs and Langerhans cells in TFH and GC responses after intradermal immunization. J Invest Dermatol. 2017;137(9):1905–1913. doi:10.1016/j.jid.2017.04.016
54. Godfrey DI, Pellicci DG, Smyth MJ. The elusive NKT cell antigen–is the search over? Science. 2004;306(5702):1687–1689. doi:10.1126/science.1106932
55. Linsen L, Somers V, Stinissen P. Immunoregulation of autoimmunity by natural killer T cells. Hum Immunol. 2005;66(12):1193–1202. doi:10.1016/j.humimm.2006.02.020
56. Koupenova M, Vitseva O, MacKay CR, et al. “Platelet-TLR7 mediates host survival and platelet count during viral infection in the absence of platelet-dependent thrombosis. Blood. 2014; blood-2013. doi:10.1182/blood-2013-11-536003.
57. Zufferey A, Schvartz D, Nolli S, Reny J-L, Sanchez J-C, Fontana P. Characterization of the platelet granule proteome: evidence of the presence of MHC1 in alpha-granules. J Proteomics. 2014;101:130–140. doi:10.1016/j.jprot.2014.02.008
58. Wagner DD, Frenette PS. The vessel wall and its interactions. Blood. 2008;111(11):5271–5281. doi:10.1182/blood-2008-01-078204
59. Hartwig H, Drechsler M, Lievens D, et al. Platelet-derived PF4 reduces neutrophil apoptosis following arterial occlusion. Thromb Haemost. 2014;111(3):562–564. doi:10.1160/TH13-08-0699
60. Barrientos S, Stojadinovic O, Golinko MS, Brem H, Tomic-Canic M. Growth factors and cytokines in wound healing. Wound Repair Regen. 2008;16(5):585–601. doi:10.1111/j.1524-475x.2008.00410.x
61. Bayer A, Lammel J, Lippross S, et al. Platelet-released growth factors induce psoriasin in keratinocytes: implications for the cutaneous barrier. Ann Anat Anz. 2017;213:25–32. doi:10.1016/j.aanat.2017.04.002
62. Chicharro-Alcántara D, Rubio-Zaragoza M, Damiá-Giménez E, et al. Platelet rich plasma: new insights for cutaneous wound healing management. J Funct Biomater. 2018;9(1):10. Published 2018 Jan 18. doi:10.3390/jfb9010010
63. Mann ER, Smith KM, Bernardo D, Al-Hassi HO, Knight SC, Hart AL. Review: Skin and the Immune System. J Clin Exp Dermatol Res. 2012;S2:003. doi:10.4172/2155-9554.S2-003
64. Luger TA, Kock A, Kirnbauer R, Schwarz T, Ansel JC. Keratinocyte‐Derived Interleukin 3. Ann N Y Acad Sci. 1988;548(1):253–261. doi:10.1111/j.1749-6632.1988.tb18813.x
65. Wojtowicz AM, Oliveira S, Carlson MW, Zawadzka A, Rousseau CF, Baksh D. The importance of both fibroblasts and keratinocytes in a bilayered living cellular construct used in wound healing. Wound Repair Regen. 2014;22(2):246–255. doi:10.1111/wrr.12154
66. Auger FA, Lacroix D, Germain L. Skin substitutes and wound healing. Skin Pharmacol Physiol. 2009;22:94–102. doi:10.1159/000178868
67. Zanetti M. Cathelicidins, multifunctional peptides of the innate immunity. J Leukoc Biol. 2004;75:39–48. doi:10.1189/jlb.0403147
68. Martinon F, Mayor A, Tschopp J. The inflammasomes: guardians of the body. Annu Rev Immunol. 2009;27:229–265. doi:10.1146/annurev.immunol.021908.132715
69. Albanesi C, Scarponi C, Giustizieri M, Girolomoni G. Keratinocytes in inflammatory skin diseases. Curr Drug Targets Inflamm Allergy. 2005;4(3):329–334. doi:10.2174/1568010054022033
70. Mc Dermott R, Ziylan U, Spehner D, et al. Birbeck granules are subdomains of endosomal recycling compartment in human epidermal Langerhans cells, which form where Langerin accumulates. Mol Biol Cell. 2002;13(1):317–335. doi:10.1091/mbc.01-06-0300
71. Bursch LS, Wang L, Igyarto B, et al. Identification of a novel population of Langerin+ dendritic cells. J Exp Med. 2007;204(13):3147–3156. doi:10.1084/jem.20071966
72. Johnson LA, Banerji S, Lawrance W, et al. Dendritic cells enter lymph vessels by hyaluronan-mediated docking to the endothelial receptor LYVE-1. Nat Immunol. 2017;18(7):762. doi:10.1038/ni.3750
73. Fay NS, Larson EC, Jameson JM. Chronic inflammation and γδ T cells. Front Immunol. 2016;7:210. doi:10.3389/fimmu.2016.00210
74. MacLeod AS, Havran WL. Functions of skin-resident γδ T cells. Cell Mol Life Sci. 2011;68(14):2399–2408. doi:10.1007/s00018-011-0702-x
75. Hartwig H, Drechsler M, Lievens D, et al. Platelet-derived PF4 reduces neutrophil apoptosis following arterial occlusion. Thromb Haemost. 2014;111(3):562–564. doi:10.1160/TH13-08-0699
76. McMenamin C, Pimm C, McKersey M, Holt PG. Regulation of IgE responses to inhaled antigen in mice by antigen-specific gamma delta T cells. Science. 1994;265(5180):1869–1871. doi:10.1126/science.7916481
77. Ramírez-Valle F, Gray EE, Cyster JG. Inflammation induces dermal Vγ4+ γδT17 memory-like cells that travel to distant skin and accelerate secondary IL-17–driven responses. Proc Natl Acad Sci. 2015;112(26):8046–8051. doi:10.1073/pnas.1508990112
78. Sharp LL, Jameson JM, Cauvi G, Havran WL. Dendritic epidermal T cells regulate skin homeostasis through local production of insulin-like growth factor 1. Nat Immunol. 2005;6(1):73. doi:10.1038/ni1152
79. Komori HK, Witherden DA, Kelly R, Sendaydiego K, Jameson JM, Teyton L, Havran WL. Cutting edge: dendritic epidermal γδ T cell ligands are rapidly and locally expressed by keratinocytes following cutaneous wounding. J Immunology. 2012;188(7):2972–2976. doi:10.4049/jimmunol.1100887
80. Xu P, Fu X, Xiao N, et al. Involvements of γδT lymphocytes in acute and chronic skin wound repair. Inflammation. 2017;40(4):1416–1427. doi:10.1007/s10753-017-0585-6
81. Toulon A, Breton L, Taylor KR, et al. A role for human skin–resident T cells in wound healing. J Exp Med. 2009;206(4):743–750. doi:10.1084/jem.20081787
82. Campbell JJ, O’Connell DJ, Wurbel M-A. Cutting edge: chemokine receptor CCR4 is necessary for antigen-driven cutaneous accumulation of CD4 T cells under physiological conditions. J Immunol. 2007;178(6):3358–3362. doi:10.4049/jimmunol.178.6.3358
83. Slominski AT, Manna PR, Tuckey RC. Cutaneous glucocorticosteroidogenesis: securing local homeostasis and the skin integrity. Exp Dermatol. 2014;23(6):369–374. doi:10.1111/exd.12376
84. Fuhlbrigge RC, Kieffer JD, Armerding D, Kupper TS. Cutaneous lymphocyte antigen is a specialized form of PSGL-1 expressed on skin-homing T cells. Nature. 1997;389(6654):978. doi:10.1038/40166
85. Richmond JM, Harris JE. Immunology and skin in health and disease. Cold Spring Harb Perspect Med. 2014;4(12):a15339. doi:10.1101/cshperspect.a015339
86. Barinova ME, Barinov EF, Sulaieva OM. Functional activity of monocytes and mechanisms of iNOS intracellular regulation during wound process. Fiziol Zh. 2011;57(1):36–44.
87. Gilroy D, De Maeyer R. New insights into the resolution of inflammation. Semin Immunol. 2015:27(3):161–168. Academic Press. doi:10.1016/j.smim.2015.05.003.
88. Cruz MS, Diamond A, Russell A, Jameson JM. Human αβ and γδ T cells in skin immunity and disease. Front Immunol. 2018;9:1304. Published 2018 Jun 6. doi:10.3389/fimmu.2018.01304
89. Supp DM, Wilson-Landy KAILA, Boyce ST. Human dermal microvascular endothelial cells form vascular analogs in cultured skin substitutes after grafting to athymic mice. Faseb J. 2002;16(8):797–804. doi:10.1096/fj.01-0868com
90. Nikolsky IS, Serebrovska TV. Role of hypoxia in stem cell development and functioning. Int J Physiol Pathophysiol Pharmacol. 2010;1(1). doi:10.1615/IntJPhysPathophys.v1.i1.90
91. Serebrovskaya TV, Nikolsky IS, Nikolska VV, Mallet RT, Ishchuk VA. Intermittent hypoxia mobilizes hematopoietic progenitors and augments cellular and humoral elements of innate immunity in adult men. High Alt Med Biol. 2011;12(3):243–252. doi:10.1089/ham.2010.1086
92. Grando SA, Pittelkow MR, Schallreuter KU. Adrenergic and cholinergic control in the biology of epidermis: physiological and clinical significance. J Invest Dermatol. 2006;126(9):1948–1965. doi:10.1038/sj.jid.5700151
93. Pongratz G, Straub RH. Role of peripheral nerve fibres in acute and chronic inflammation in arthritis. Nat Rev Rheumatol. 2013;9(2):117. doi:10.1038/nrrheum.2012.181
94. Vinik AI, Strotmeyer ES. Diabetic neuropathy. Pathy’s Principles Pract Geriatric Med. 2012;1:751–767. doi:10.1002/9781119952930.ch65
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.