Back to Journals » International Journal of Nanomedicine » Volume 20
The Potential Application of Exosomes as Therapeutic Agents, Carriers, and Biomarkers in Skin Diseases
Authors Li W, Li H, Fan L, Zhao R, Dong F, Jia X, Yin L, Yang S, Zhao D ![]() , Wang J
, Wang J
Received 3 July 2025
Accepted for publication 28 September 2025
Published 15 October 2025 Volume 2025:20 Pages 12627—12658
DOI https://doi.org/10.2147/IJN.S547871
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Krishna Nune
Wenjing Li,1 Haiyan Li,2 Liangliang Fan,1 Ronghua Zhao,1 Fusong Dong,1 Xuxia Jia,1 Lijia Yin,1 Song Yang,1 Daqing Zhao,1 Jiawen Wang1
1Northeast Asia Institute of Traditional Chinese Medicine, Changchun University of Chinese Medicine, Changchun, Jilin, People’s Republic of China; 2Experimental Centre, The Affiliated Hospital of Changchun University of Chinese Medicine, Changchun, Jilin, People’s Republic of China
Correspondence: Jiawen Wang, Northeast Asia Institute of Traditional Chinese Medicine, Changchun University of Chinese Medicine, Boshuo Road, Nanguan District, Changchun, Jilin, 130117, People’s Republic of China, Email [email protected]
Abstract: Exosomes, owing to their potent capabilities for intercellular communication, have great potential in modern medicine. The pathogenesis of skin diseases is predominantly characterized by immune imbalance and impaired regenerative processes. Exosomes are rich in microRNAs, lipids, and proteins, which play a crucial role in immune regulation and regenerative medicine. Recent studies have highlighted the involvement of exosomes in the therapeutic management of dermatological disorders. This review provides a comprehensive overview of the potential therapeutic applications of exosomes in autoimmune skin conditions, such as systemic lupus erythematosus, systemic sclerosis, psoriasis, and vitiligo, as well as their roles in skin wound healing, skin aging, scar formation, skin flap regeneration, hair regeneration, and other skin regenerative disorders. The paper examines various exosome delivery methods and evaluates current clinical trials, identifying challenges related to exosome isolation, preservation, active components, mechanistic understanding, and clinical translation. Furthermore, it delineates future research directions and proposes innovative strategies for advancing exosome-based therapies in dermatology.
Keywords: Exosomes, Autoimmune skin diseases, Skin regeneration, Engineered exosomes
Introduction
The skin, recognized as the largest organ in vertebrates, consists of diverse layers, including the epidermis, dermis, and subcutaneous tissue, along with their associated appendages, all supported by a complex neurovascular network1,2 (Figure 1). The stratum corneum (SC), the outermost layer of the skin, measures approximately 10–20 μm in thickness. Within the intercellular spaces of the SC, a highly organized lipid layer is present. These lipids are essential for the permeation of compounds through the SC, thereby facilitating its barrier function on the skin surface.1,3 Therefore, examining the lipid composition of the SC is essential for elucidating both normal skin function and conditions involving a compromised skin barrier. Located beneath the epidermis, the dermis serves as the primary structural component of the skin, composed of collagen, elastin, and glycosaminoglycans (GAGs). Fibroblasts, which are the most abundant cell type in the dermis, can synthesize remodeling enzymes like proteases and collagenases. These enzymes play a vital role in the wound repair process. The deepest layer of the skin is the subcutaneous tissue, primarily made up of highly vascularized adipose tissue and a layer of loose connective tissue. This composition plays a vital role in regulating skin temperature and mechanical properties. Collectively, these three layers are essential for protecting the body against mechanical injury.2 Skin appendages, such as hair follicles, sweat glands, and sebaceous glands, are fundamental components of the integumentary system. During embryogenesis, these structures originate from epidermal buds that extend into the dermis, forming the respective appendages. Sweat glands are characterized by a coiled secretory portion and an elongated duct that traverses the dermis to reach the epidermis. These glands are crucial for sweat secretion, maintaining electrolyte balance, regulating body temperature, and enhancing skin defense by inhibiting bacterial growth through lactate production.4,5 Sebaceous glands are situated within the middle layer of the dermis, with an outer layer of proliferating cells and an inner layer comprising sebocytes. Sebaceous glands secrete sebum into the ducts at the junction of the hair follicle. This secretion plays a crucial role in minimizing water loss from the epithelium located between hair follicles and contributes to the regulation of body temperature.5 Hair follicles are embedded within the dermis and subcutaneous tissue and consist of the hair papilla and dermal fibers. These follicles are dynamic structures characterized by their cyclical regenerative capabilities.6 They are integral to temperature regulation, the distribution of sebum and sweat, and the provision of physical protection.5 Collectively, these tissues function to protect the body from harmful microorganisms and other external environmental threats.
 |
Figure 1 Composition of skin. |
Cells possess the capacity to release extracellular vesicles enveloped by a phospholipid bilayer. Among these vesicles, those with diameters ranging from 30 to 150 nm are classified as exosomes. Intracellular lipids, RNA, and proteins are selectively incorporated into the intraluminal vesicles (ILVs) within multivesicular bodies (MVBs). Cells release exosomes via exocytosis, following the fusion of the plasma membrane with MVBs7,8 (Figure 2).
 |
Figure 2 Source, Secretion, Composition and Function of Exosomes. |
Exosomes originate from a wide range of sources, including human body fluids, cells, and tumors; animal-derived substances such as bovine milk and cells; plant materials like ginger, ginseng, and grapefruit; and microorganisms, including bacteria and fungi.9 The methods employed for the extraction and isolation of exosomes differ based on their source, as well as the desired extraction efficiency and purity. Techniques utilized for exosome extraction encompass ultracentrifugation, ultrafiltration, size-exclusion chromatography, field-flow fractionation, immunoaffinity approaches, ion exchange chromatography, and microfluidic-based technologies.10 Of these, differential ultracentrifugation is the most commonly employed method. Researchers frequently utilize differential ultracentrifugation to isolate exosomes from cells and plant sources, subsequently employing sucrose gradient centrifugation to enhance the purity of the exosomes obtained11 (Figure 2).
Exosomes fulfill diverse roles, encompassing diagnostics, therapy, and drug delivery, and are integral to human medicine, particularly in the treatment of cancer,12 cardiovascular diseases,9 immune disorders,13 degenerative diseases,14 diabetes,15 cardiac protection,16 liver injury,17 peripheral nerve regeneration,18 inflammation,19,20 and aging21 (Figure 2).
Dermatological conditions, also known as dermatoses, encompass all diseases affecting the skin, hair, nails, and their associated structures. As a global public health concern, skin diseases exert complex impacts across physiological, psychological, and social dimensions. Conventional therapies are often protracted, susceptible to recurrence, and may induce systemic side effects due to improper administration methods, leading to suboptimal treatment outcomes.22 Exosomes, as nanoscale therapeutic agents, exhibit enhanced targeting capabilities, functioning as carriers to deliver therapeutic molecules to affected sites. Due to their unique regenerative properties, exosomes hold significant therapeutic potential in the treatment of skin disorders.
This review provides a comprehensive summary of exosome delivery methods within the field of dermatology. It elucidates their therapeutic potential as therapeutic agents, carriers, and biomarkers in the context of autoimmune skin diseases, skin regenerative disorders, and cutaneous malignancies. Additionally, it addresses the limitations associated with exosomes development, particularly in terms of extraction, characterization, preservation, compositional and mechanistic research, and clinical trials. By building upon existing research foundations and acknowledging current constraints, the review delineates future developmental directions and explores the therapeutic potential of exosomes in dermatological conditions.
Administration Methods of Exosomes in the Treatment of Skin Diseases
In disease management, exosomes can be administered through diverse routes including systemic delivery, topical application, and targeted delivery. For dermatological conditions, topical application and subcutaneous/intradermal injection are the primary approaches. Herein, we summarise and compare the administration methods of exosomes for treating skin diseases, alongside their respective advantages and disadvantages (Table 1).
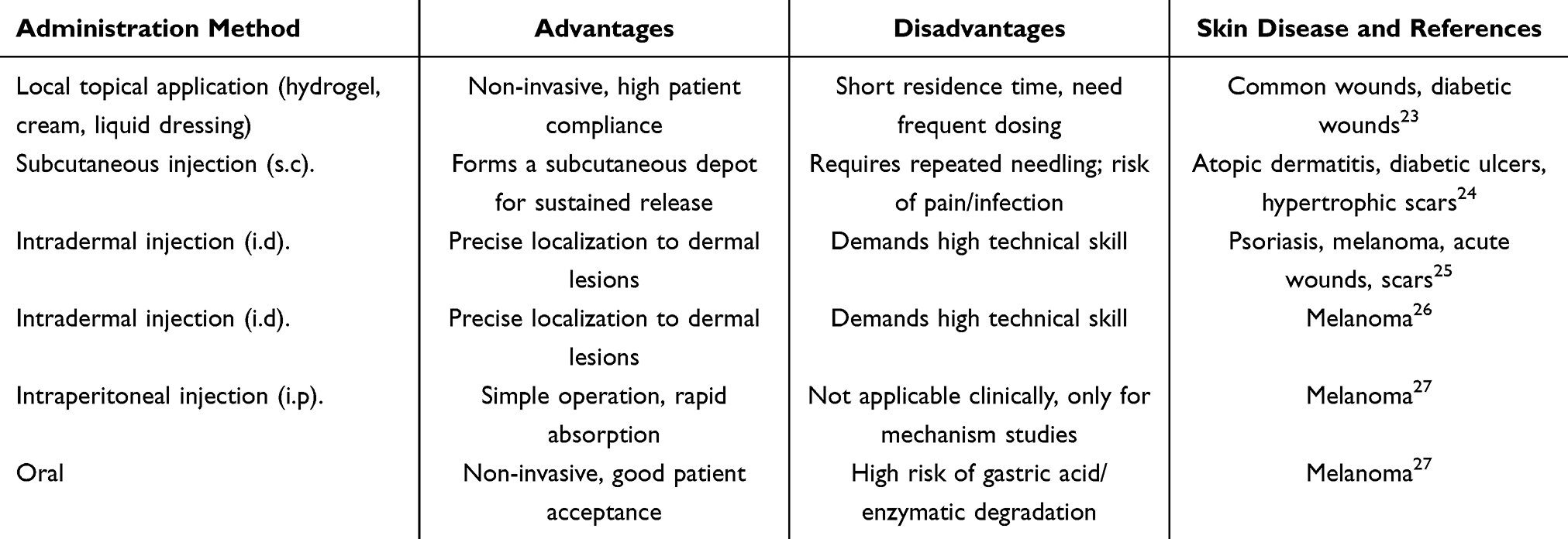 |
Table 1 Administration Methods of Exosomes in the Treatment of Skin Diseases |
The Therapeutic Role of Exosomes in Autoimmune Skin Diseases
Autoimmune skin diseases are characterized by heterogeneity and complex pathogenesis, where genetic, infectious, environmental, and psychological factors interact to trigger innate and adaptive immune responses.28 Cutaneous manifestations of autoimmune diseases encompass conditions such as systemic lupus erythematosus (SLE), systemic sclerosis (SSc), psoriasis, and vitiligo, among others. Exosomes contribute to the pathogenesis of autoimmune diseases through intercellular communication and modulation of immune cell responses.29 (Figure 3).
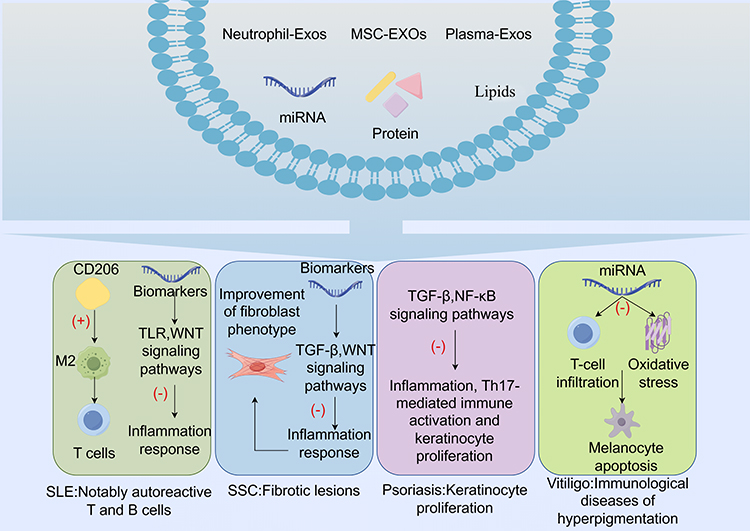 |
Figure 3 The therapeutic role of exosomes in autoimmune skin diseases (“+”:activate, “-”: inhibit). |
SLE
SLE is characterized as a multisystem autoimmune disease. This disruption in immune tolerance results in the aberrant activation of various immune cells, notably autoreactive T and B cells, leading to clinical manifestations such as butterfly or discoid rash, mucosal ulcers, and alopecia.30 The predominant causes of mortality in patients with SLE are renal disease, cardiovascular complications, and infections.31 Existing pharmacological treatments for SLE are associated with numerous side effects and exhibit limited efficacy in certain patient subgroups. Recent studies have demonstrated that exosomes have the capacity to modulate immune responses, either stimulating or inhibiting them based on the environmental context, thereby highlighting their therapeutic potential in the management of SLE.32
Mesenchymal stem cells (MSCs), derived from diverse tissues and organs throughout the human body, are multipotent stromal cells capable of differentiating into various cell types, such as osteoblasts, adipocytes, and chondrocytes.33 MSCs demonstrate advantageous immunomodulatory properties that can be leveraged for the treatment of SLE.34 Exosomes derived from MSCs (MSC-Exos) also exhibit extensive immunomodulatory and regenerative functions.13 Research suggests that exosomes from bone marrow MSCs (BMMSC-Exos) ameliorate SLE by promoting the upregulation of cytokines such as CD206 and B7H4, inducing macrophage polarization, and recruiting IL-17+ regulatory T cells.35 Similarly, exosomes derived from human umbilical cord MSCs (hUCMSC-Exos) can enhance the polarization of CD206-induced M2 macrophages and T regulatory (Treg) cells, as well as facilitate the expansion of regulatory T cells, thereby improving SLE.36 Stem cells from human exfoliated deciduous teeth (SHED) exhibit MSC-like characteristics.37 SHED-Exos enhance the functionality of recipient BMMSCs in SLE by restoring Tert mRNA-associated telomerase activity, facilitating hematopoietic niche formation, and modulating immune responses, thereby playing a pivotal role in the disease process.38
The expression of microRNAs (miRNAs) is stringently regulated during the development, differentiation, and effector functions of immune cells, including those involved in the immunological disorders associated with SLE.39 miRNAs derived from exosomes in the plasma of SLE patients specifically target TLR7 and TLR8 signaling pathways, thereby inhibiting autoimmune-mediated inflammation in SLE.40 Exosomes are found in all biological fluids and their unique contents are regarded as indicators of autoimmune diseases. In patients with SLE, the miRNA profile within exosomes markedly deviates from that of healthy individuals, underscoring their potential as key biomarkers for renal damage.41 For example, miR-21, miR-29a, miR-29b, miR-146a, miR-26a, miR-150, and miR-146a-5p are typically found to be abnormally elevated in patients with SLE, thereby facilitating early diagnosis and treatment of the disease.40–45
Beyond the role of miRNAs as biomarkers for SLE, additional substances have been identified that may aid in diagnosing the condition. Yan et al found the metabolic fingerprint of urinary exosomes that could serve as a potential biomarker for SLE.46 Renal damage represents one of the most severe complications associated with SLE. In plasma exosomes, four long non-coding RNAs and two miRNAs (miR-16-5p and miR-101-3p) linked to renal damage target the TGF-β, WNT/β-catenin, and fibroblast growth factors (FGFs), which may emerge as potential diagnostic and therapeutic targets for SLE.47 Bactericidal/Permeability-Increasing Protein (BPI), originating from SLE T cells and T cell-derived exosomes, functions as a negative regulator of regulatory T cell differentiation. The overexpression of BPI in T cell-derived exosomes has the potential to serve as both a biomarker and a pathogenic factor for human SLE.48 Identifying these biomarkers could enhance the early diagnosis and treatment of SLE in clinical settings.
SSc
SSc, commonly referred to as scleroderma, is an immune-mediated rheumatic disease characterized by fibrosis and vascular alterations in the skin and internal organs. Complications associated with SSc include inflammatory skin conditions, digital swelling, musculoskeletal inflammation, pulmonary fibrosis, and pulmonary arterial hypertension.49 The pathogenesis of SSC involves vascular lesions, immune system dysfunction, and ultimately cutaneous fibrosis.50 Due to the characteristics of exosomes, potentially establishing a communication network among endothelial cells (ECs), immune cells, and specific target organs, thereby acting as mediators of vascular damage and parenchymal fibrosis.51 Exosomes are implicated in various pathological and physiological aspects of SSc, playing a crucial role in its diagnosis and treatment.
MSCs exhibit significant immunomodulatory properties; however, MSC-Exos may offer distinct advantages over the use of MSCs alone in the treatment of autoimmune diseases.52 Human myofibroblasts induced by TGFβ1 contribute to the pathogenesis of SSc. Research has shown that exosomes derived from adipose MSCs (AMSC-Exos) more effectively ameliorate the myofibroblast phenotype than their parental cells, suggesting that extracellular vesicles exert more potent anti-fibrotic effects in vitro models of SSc compared to adipose mesenchymal stromal cells.52 Furthermore, as a cell-free therapeutic approach, MSC-Exos present a reduced risk of immune rejection and tumorigenesis compared to MSCs. This study further underscores the importance of investigating MSC-Exos in SSc. MSC-Exos, which are abundant in various bioactive factors, have the potential to ameliorate SSc through multiple mechanisms, potentially including the promotion of angiogenesis and the modulation of inflammation and fibrosis.53
The miRNAs and proteins encapsulated within exosomes are crucial components of signal transduction and play a significant role in the diagnosis and treatment of SSc (Table 2). A distinctive feature of SSc is the extension of fibrotic changes to previously unaffected tissues. This phenomenon may be mediated by exosomes derived from SSc-affected cells, which can induce the activation of profibrotic phenotypes in normal or unaffected cells.54 Skin ulcers are among the common complications experienced by SSc patients.55 The vascular abnormalities observed in SSc may be attributed to a disruption in the transfer of exosomes from skin tissue to the bloodstream, resulting in decreased serum exosome levels. This disruption may lead to delayed wound healing due to the downregulation of collagen, thereby increasing susceptibility to atrophic scars and/or ulcers.56 These findings suggest that modulating exosome activity in SSc patients could potentially arrest the progression of the disease.
 |
Table 2 Components and Mechanisms of Effect of Exosomes in the Treatment of SSC |
Psoriasis
Psoriasis is a prevalent chronic papulosquamous dermatosis characterized by clinical manifestations such as chronic plaques, erythroderma, and pustular psoriasis. The etiology of psoriasis is primarily attributed to genetic predispositions, environmental influences, and immune-related factors.64 Key elements in its pathogenesis include tumor necrosis factor alpha (TNF-α), dendritic cells, and T cells.65 In psoriasis, there is excessive proliferation of keratinocytes (KCs), accompanied by parakeratosis, which ultimately results in acanthosis. Exosomes serve as long-distance molecular signals in immune responses, positioning them as crucial contributors to the pathogenesis of psoriasis.66
The immunomodulatory properties of MSCs have shown significant therapeutic efficacy in psoriasis, advancing into clinical research.67 Numerous studies have confirmed that MSC-Exos can mitigate the onset and progression of psoriasis by modulating inflammatory factors and immune responses.68–70 Moreover, MSC-Exos have been shown to mitigate the proliferation of psoriasis by inducing TGF-β1/2.71,72 Similarly, UCMSC-Exos have demonstrated significant efficacy in repairing tissue damage in psoriasis mouse models by modulating the local immune microenvironment.73 Researchers have innovatively combined exosomes derived from grapefruit with those from gingiva-derived mesenchymal stem cells (GMSCs) to create multifunctional fusion vesicles for the treatment of autoimmune skin diseases. These vesicles effectively reduce the secretion of inflammatory factors, inhibit the activation of Th17 cells, and promote the infiltration of Treg cells. Furthermore, they reestablish the disrupted immune microenvironment and have shown remarkable therapeutic effects in psoriasis disease models.74 These multifunctional fusion vesicles offer advantages such as enhanced targeting and reduced side effects in the treatment of immune-related diseases.
In the pathogenesis of psoriasis, the interaction among inflammation, Th17-mediated immune activation, and keratinocyte hyperproliferation plays a pivotal role.75 Engineered keratinocyte-derived extracellular vesicles have demonstrated therapeutic potential in psoriasis by inhibiting LAT1-mTOR-mediated keratinocyte overproliferation and Th17 cell expansion, as well as suppressing IL-1/NF-κB-mediated inflammatory responses.76 Similarly, small extracellular vesicles derived from umbilical cord blood mononuclear cells (UCB-EVs) and neutrophil-derived exosomes have shown efficacy in treating psoriasis through the modulation of inflammation, immune responses, and keratinocyte proliferation.77,78
Beyond immune dysregulation, dysbiosis of the skin microbiome also contributes to the etiology of psoriasis.79 In patients with psoriasis, there is an increased expression of Bacillales, Staphylococcus, and Sphingomonas, along with a higher abundance of Propionibacterium acnes in plasma exosomes. Conversely, there is a decreased abundance of Lactobacillales, Brucellaceae, Streptococcus, Kingella, and Aquabacterium.80 Exosomes derived from Cutibacterium acnes have been shown to ameliorate psoriasis symptoms by modulating the differentiation of innate lymphoid cell (ILC) subsets and re-establishing the long-term homeostasis of the skin microbiome.81 Similarly, exosomes from Staphylococcus epidermidis have demonstrated efficacy in mitigating imiquimod-induced psoriasis by reducing acanthosis, cellular infiltration, and the expression of vascular endothelial growth factor (VEGF).82
Psoriasis is linked to abnormalities in epidermal Langerhans cells (LC),83 with CD1a, a molecule that is highly and constitutively expressed by LC, playing a crucial role in the disease’s pathogenesis.84 Exosomes released from mast cells, upon induction by IFN-α, transfer cytosolic phospholipase A2 (PLA2) activity to adjacent CD1a-expressing cells, resulting in the formation of novel lipid antigens. These antigens are subsequently recognized by lipid-specific CD1a-reactive T cells, which in turn produce IL-22 and IL-17A, thereby exacerbating the inflammatory response characteristic of psoriasis.85 In a similar vein, the interaction between keratinocytes and macrophages through extracellular vesicles enriched with leucine-rich alpha-2-glycoprotein 1 (LRG1) has been shown to exacerbate psoriasis-like dermatitis.86 This finding suggests that inhibiting specific exosome-mediated communications could potentially serve as a therapeutic strategy for psoriasis.
miRNAs, which act as essential mediators of cellular communication within exosomes, play a significant role in the diagnosis of psoriasis. Studies have demonstrated that various miRNAs exhibit abnormal expression levels, either increased or decreased, in the plasma and skin keratinocyte exosomes of patients with psoriasis, with mechanisms primarily involving anti-inflammatory and immunomodulatory effects (Table 3). Psoriatic arthritis (PsA), a prevalent complication of psoriasis, affects approximately 30% of individuals with the condition.87 Exosome miRNAs not only serve as biomarkers for the onset and progression of psoriasis but also facilitate the differentiation between PsA and Psoriasis vulgaris (PsV). For example, in patients with PsA, serum extracellular vesicles exhibit a marked decrease in hsa-miR-671-3p levels, while hsa-miR-33-5p levels tend to increase.88 Furthermore, lipids, which are critical constituents of exosomes, exhibit significant alterations in the plasma exosomes of patients with psoriasis. Notably, there is an observed increase in the levels of phosphatidylcholine (PC), phosphatidylethanolamine (PE), phosphatidylglycerol, and PC.89 These findings suggest that exosomal miRNAs and lipids have potential as biomarkers for the early detection and monitoring of the onset and progression of psoriasis.
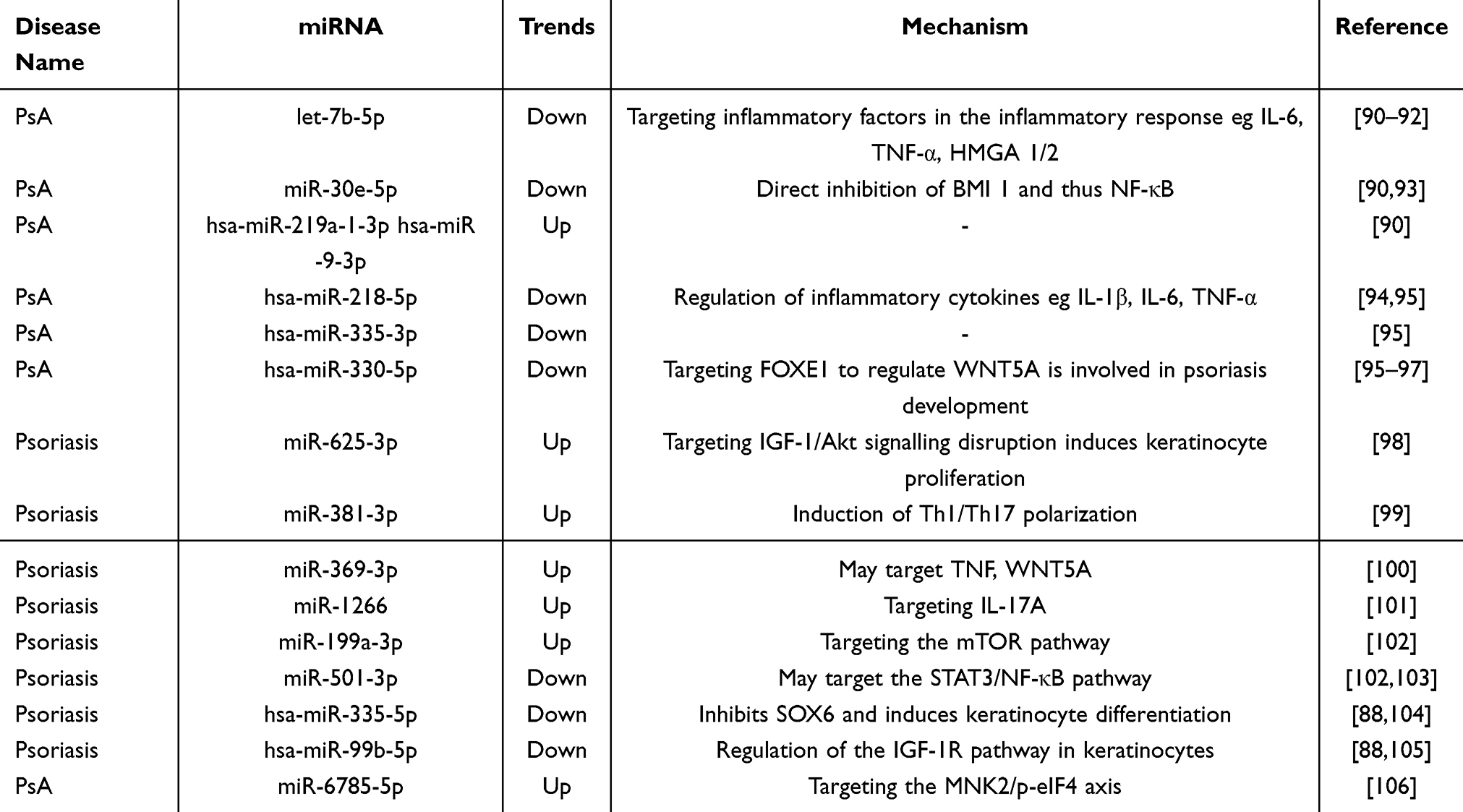 |
Table 3 Biomarkers in Psoriasis |
Vitiligo
Vitiligo is an idiopathic acquired pigmentation disorder characterized by the formation of white patches resulting from the loss of functional melanocytes in the skin, hair, or both. The pathogenesis of vitiligo is linked to autoimmune mechanisms, inflammation, and oxidative stress.107 Exosomes exhibit extensive immunomodulatory properties, participating in antigen presentation, synapse formation, and acting as mediators of intercellular communication to exert immunomodulatory effects.77,108,109
In patients with vitiligo, autoreactive cytotoxic CD8+ T cells interact with melanocytes through the local production of IFN-γ, thereby promoting disease progression.110 hUMSCs-Exos effectively ameliorate vitiligo by delivering miR-132-3p and miR-125b-5p, which concurrently reduce CD8+ T cell infiltration, enhance Treg cell-mediated immune suppression, and inhibit oxidative stress-induced melanocyte damage.111 Circulating exosomal miRNAs have emerged as promising indicators for a range of diseases. In vitiligo patients, circulating exosomal miR-493-3p is significantly elevated, which is associated with its capacity to induce reactive oxygen species in keratinocytes and promote melanocyte apoptosis.112 miR-200c activates β-catenin and upregulates genes associated with melanogenesis through the suppression of SOX1. Consequently, it is significantly downregulated in exosomes derived from keratinocytes in vitiligo lesions.113 These exosomes are internalized by NK cells, thereby enhancing their proliferative activity and capacity for IFN-γ secretion, which contributes to the elevated levels of miR-1469 in the circulating exosomes of non-segmental vitiligo (NSV) patients. CD122, a potential target of miR-1469, can partially mitigate the effects of miR-1469 on NK cells.114 In conclusion, considering the pathogenesis of vitiligo and the role of exosomes in immune modulation, exosomes may offer significant potential for the diagnosis and treatment of vitiligo.
The Therapeutic Role of Exosomes in Skin Rejuvenation
Skin injury and regeneration are tightly coupled through intricate interactions among skin cells. Exosomes, acting as endogenous mediators of tissue repair, exert potent regenerative effects.115 In the context of skin regeneration, exosomes commonly utilized are derived from sources such as stem cells, neutrophils, body fluids, macrophages, plasma, and plants. These exosomes have demonstrated beneficial diagnostic and therapeutic effects on a range of skin regenerative disorders, including skin wound,116 skin aging,117 hypertrophic scarring,118 flap regeneration,119 and hair regeneration120 (Figure 4).
 |
Figure 4 The therapeutic role of exosomes in skin rejuvenation. |
Wound Healing of the Skin
The skin is frequently exposed to chronic and acute wounds resulting from conditions like diabetic foot ulcers or extensive burns. The wound healing process is intricate, comprising four distinct phases: hemostasis, inflammation, proliferation, and tissue remodeling or regression.121 Initially, a clot forms at the site of tissue injury to provide a temporary seal over the wound gap. Following the initial response, innate immune cells, including neutrophils and macrophages, are recruited to the wound site. These cells orchestrate the development of granulation tissue, which functions to replace the lost connective tissue and drive processes such as angiogenesis, fibroblast migration, and collagen matrix deposition. As granulation tissue accumulates, epidermal cells migrate and proliferate, aiding in the restoration of the skin surface and barrier function. Wound healing is ultimately achieved when the wound edges approximate.122,123 Exosomes are involved in a range of biological processes, such as cell differentiation and proliferation, angiogenesis, stress response, and immune signaling.124 As a cell-free therapeutic approach, Exosomes have shown notable efficacy in promoting wound healing.
General Wounds
Stem cells, defined by their undifferentiated status and capacity for self-renewal, self-replication, and multidirectional differentiation, are pivotal in tissue repair. They help minimize scar formation, reduce wound contraction, and enhance collagen synthesis.116 Exosomes derived from stem cells (SC-Exos) have been the subject of extensive research due to their significant therapeutic potential in wound healing, attributed to their exceptional biocompatibility and targeted delivery capabilities. Exosomes derived from multiple types of stem cells, such as adipose-derived mesenchymal stem cells, human umbilical cord mesenchymal stem cells, human amniotic stem cells, and mesenchymal stem cells from induced pluripotent stem cells, have demonstrated the ability to promote fibroblast and vascular endothelial cell proliferation, as well as collagen synthesis at the wound site. These actions collectively accelerate the wound healing process (Table 4).
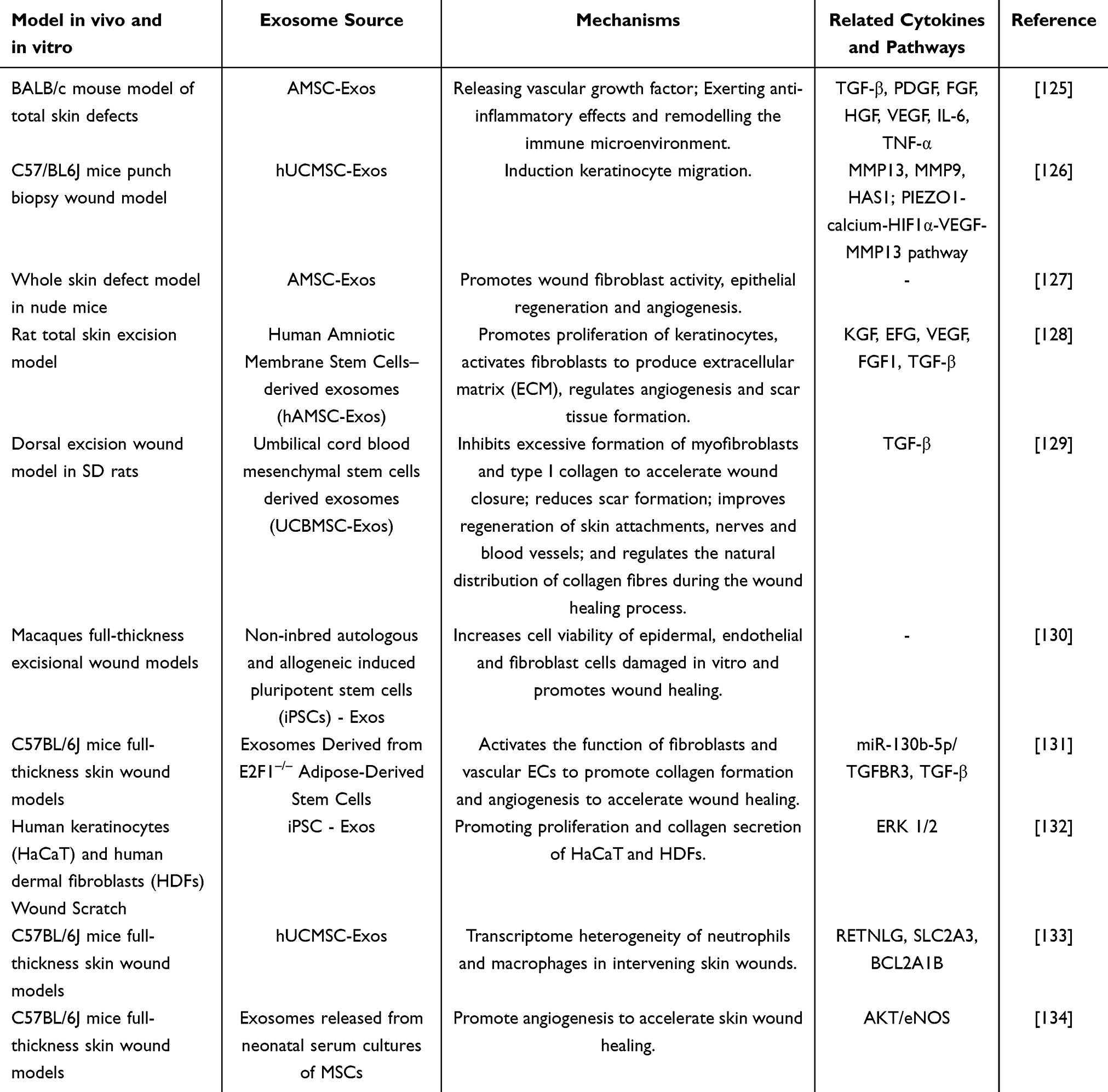 |
Table 4 Effects and Mechanisms of Stem Cell Exosomes in Promoting Wound Healing |
Effective wound healing following skin injury relies on dynamic cellular processes that are orchestrated through bidirectional communication between cells. In particular, autophagy is essential for coordinating the interactions between keratinocytes and fibroblasts during the skin repair process.135 To prevent wound infection, neutrophils are recruited to recognize inflammatory signals, while macrophages phagocytose apoptotic neutrophils to mitigate wound inflammation.123 SHED-Exos transport miR-1246 to regulate the AKT/ERK1/2/STAT3 signaling pathway, which in turn enhances autophagy and facilitates wound healing.136 Engineered keratinocyte-derived exosomes can be selectively phagocytosed by wound macrophages within granulation tissue, leading to elevated levels of pro-inflammatory cytokines and epithelial junction proteins, and preserving barrier function in murine models. This highlights the interplay between keratinocytes and macrophages during the wound healing process.137 Bovine milk exosomes (BM-Exos) encompass a variety of anti-inflammatory agents that facilitate the shift from the inflammatory phase to the proliferative phase.138 Additionally, they contain factors essential for tissue remodeling and angiogenesis, thereby promoting epithelial regeneration, activating angiogenesis, and enhancing extracellular matrix (ECM) maturation to support wound healing.139 Exosomes derived from activated neutrophils, which are enriched with antimicrobial proteins, contribute to the healing of infected wounds by reducing inflammatory cytokines and enhancing collagen deposition.140
As significant paracrine mediators, exosomes can facilitate regenerative processes through the delivery of bioactive molecules. Exosomes sourced from human umbilical cord plasma (hUCP-Exos) expedite wound re-epithelialization, diminish scar width, and promote angiogenesis by delivering miR-21-3p. This miRNA inhibits key regulators of fibroblast and endothelial cell functions, specifically phosphatase and tensin homolog (PTEN) and sprouty homolog 1 (SPRY1).141 Exosomes derived from saliva are enriched with ubiquitin-conjugating enzyme E2O (UBE2O), which diminishes the levels of SMAD family member 6 (SMAD6), subsequently activating bone morphogenetic protein 2 (BMP2) and inducing angiogenesis.142 Additionally, exosomes mediate the transfer of miR-125b from young to senescent fibroblasts, where it inhibits sirtuin 7, thereby facilitating fibroblast migration and transition, ultimately promoting wound healing.143 Exosomes originating from primary skin fibroblasts that encapsulate Gstm2 mRNA mitigate oxidative stress damage in senescent dermal fibroblasts by modulating mitochondrial oxidative phosphorylation. They also promote the regulation of epidermal cell function by dermal fibroblasts through the paracrine secretion of the nascent peptide-associated complex alpha subunit (NACA), thereby facilitating the repair of aged wounds.144
The skin microbiota is vital to the skin diseases, and skin injuries can disrupt this balance.145 Furthermore, bacteria have been reported to secrete exosomes, which contribute to wound healing. Exosomes secreted by Synechococcus elongatus PCC7942 have been shown to exert anti-inflammatory effects by inhibiting the expression of IL-6, thereby promoting angiogenesis and facilitating wound healing.146 Similarly, extracellular vesicles derived from Lactobacillus rhamnosus GG contribute to wound healing by mediating re-epithelialization and angiogenesis through miR-21-5p.147
In recent years, exosomes derived from naturally green plants have demonstrated significant therapeutic potential in regenerative medicine, attributed to their excellent tissue compatibility and low toxicity.148 Exosomes isolated from ginseng have been shown to enhance the secretion of MMP-1, fibronectin-1, elastin-1, and COL1A1, and to regulate the ERK/AKT/mTOR signaling pathway, thereby enhancing cell proliferation, facilitating skin wound healing, and reducing inflammation.149 Furthermore, exosomes derived from Aloe vera exhibit characteristics of inflammatory modulators, reducing the differentiation and contraction of myofibroblasts, and are essential in the initial phases of burn wound healing.150 Wheat-derived exosomes have been shown to enhance the proliferation and migration of endothelial, epithelial, and dermal fibroblast cells, as well as to facilitate the formation of tubular structures in endothelial cells, indicating their potential therapeutic application in wound healing.151 Similarly, exosomes derived from Dendrobium have demonstrated efficacy in suppressing IL-1β expression and expediting the repair of skin wound tissues.152 Collectively, these findings suggest that plant-derived exosomes could represent an effective and environmentally friendly therapeutic strategy for wound healing.
Currently, the primary routes of exosome administration include oral gavage, direct injection, and topical application to wounds. However, free exosomes may rapidly disperse within the body or at wound sites, potentially diminishing their therapeutic efficacy. To preserve the biological activity of exosomes and regulate their effective concentration during treatment, researchers have developed modified exosomes and engineered delivery systems.153 Such delivery systems have been extensively utilized in exosome-facilitated wound healing (Table 5). Hydrogels, which consist of a three-dimensional polymer network that mimics the extracellular matrix, offer high water content, excellent biocompatibility, and plasticity. By modifying the network structure and cross-linking density of hydrogels, as well as their degradability, the release rate and duration of exosomes can be precisely controlled. This modulation facilitates improved adaptation to complex wound environments, thereby enhancing the biological activity and therapeutic efficacy of exosomes.154 Delayed wound healing is primarily attributed to bacterial infection, immune response, and inflammatory response. Exosomes have the capacity to transport immunomodulatory factors, anti-inflammatory agents, and antimicrobial compounds, enabling them to modulate systemic immune responses or specifically target damaged tissues, thereby augmenting their tissue regeneration capabilities.155,156
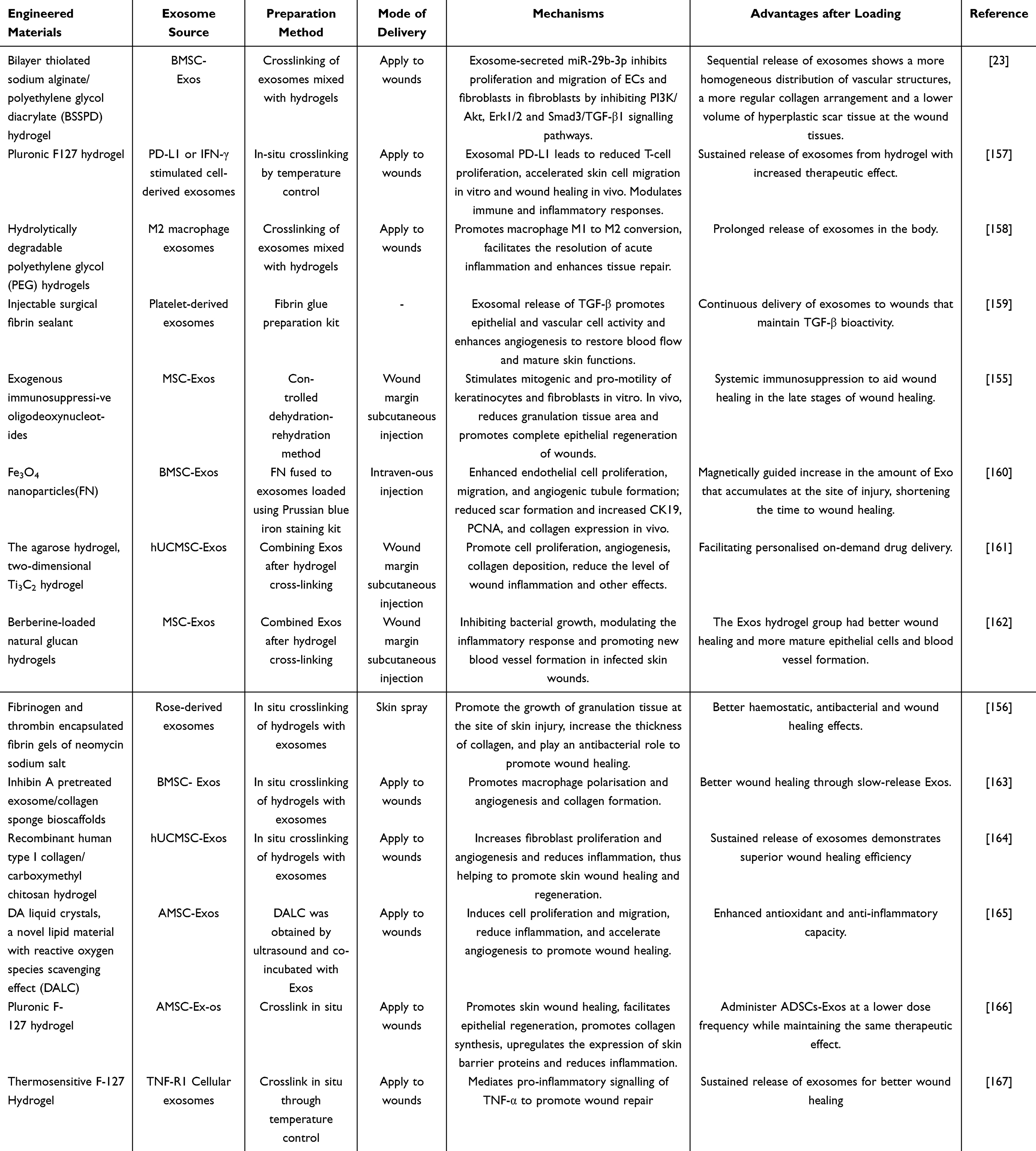 |
Table 5 The Role of Engineered Exosomes in Wound Healing |
Diabetic Wounds
Diabetic wounds represent a category of chronic wound diseases. Chronic wounds often become arrested in the inflammatory phase, leading to compromised angiogenesis and postponed re-epithelialization.123 The pathogenesis of diabetic wounds is associated with chronic bacterial infection, oxidative stress, inflammation, and angiogenic impairment.168–170 Consequently, diabetic wounds necessitate more effective and diversified therapeutic strategies.
Analogous to the treatment of common wounds, stem cell-derived exosomes(SC-Exos) play a crucial role in addressing diabetic wounds. A significant challenge in treating diabetic wounds is angiogenic impairment. BMSC-Exos promote the proliferation, migration, and tubule formation of vascular ECs and simultaneously upregulate the expression of VEGF.171,172 The PI3K/AKT signaling pathway plays a vital role in modulating cell proliferation and migration. In diabetic wounds, BMSC-Exos activate this pathway, which in turn enhances angiogenesis and facilitates wound healing.172–174 Human urine stem cell-derived exosomes (USC-Exos) promote endothelial cell angiogenesis by highly expressing deleted in malignant brain tumors 1.175 Vascular ECs are highly heterogeneous, with CD31hiEMCNhi ECs promoting angiogenesis176 MSC-Exos induce the formation of CD31hiEMCNhi vessels in skin regeneration and promote diabetic wound healing.177 In patients with diabetic ulcers, cellular Ca2+ homeostasis is disrupted, and TRPC6 can mediate Ca2+ influx. MSC-Exos facilitate the restoration of TRPC6 and mitochondrial function by delivering the transcription factor SP2 and the deubiquitinase USP9, thereby contributing to the healing of diabetic wounds.178 Autophagy plays a critical role in the removal of damaged mitochondria through various pathways, mitigating skin cell dysfunction induced by high-glucose conditions.179 ADSC-Exos improve skin wound healing in diabetic mice through the promotion of epidermal autophagy and acceleration of re-epithelialization.180 Furthermore, iPSC-Exos demonstrate reduced donor variability and exhibit significant therapeutic potential in diabetic wounds by modulating immune cell infiltration and exerting anti-inflammatory and pro-angiogenic effects.181
In addition to SC-Exos, exosomes from various other cellular sources or body fluids have also demonstrated significant therapeutic potential in diabetic wound healing. Sphingosine-1-phosphate, a key regulator of vascular homeostasis and angiogenesis, is enriched in platelet-rich plasma-derived exosomes (PRP-Exos). These exosomes enhance angiogenesis in diabetic wounds by modulating the S1PR1/AKT/FN1 signaling pathway.182 Hyperglycemia causes vascular dysfunction in diabetic wounds by increasing inflammatory factors and reactive oxygen species.183 Macrophages help reduce inflammation through chemotaxis and phagocytosis.184 Macrophage-derived exosomes loaded with curcumin mitigates oxidative stress and inflammation induced by high glucose, thereby facilitating the proliferation, migration, and angiogenesis of HUVECs.185 Exosomes from adipose tissue macrophages adjust miR-222-3p levels, influencing macrophage polarization and speeding up diabetic wound healing.186 Interestingly, plant-derived exosomes promote macrophage polarization and enhance diabetic ulcer wound healing, showing promise for glycolytic reprogramming in these ulcers.187,188
Analogous to the use of exosomes in the treatment of common wounds, engineered materials combined with exosomes have been extensively employed in addressing diabetic wounds, including multifunctional hydrogels, smart microneedles, protein patches, and multifunctional bio-hybrid nanorobotic platforms (Table 6). These engineered exosomes exhibit enhanced therapeutic efficacy at diabetic wound sites, offering more advantageous strategies for the application of exosomes in diabetic wound treatment.
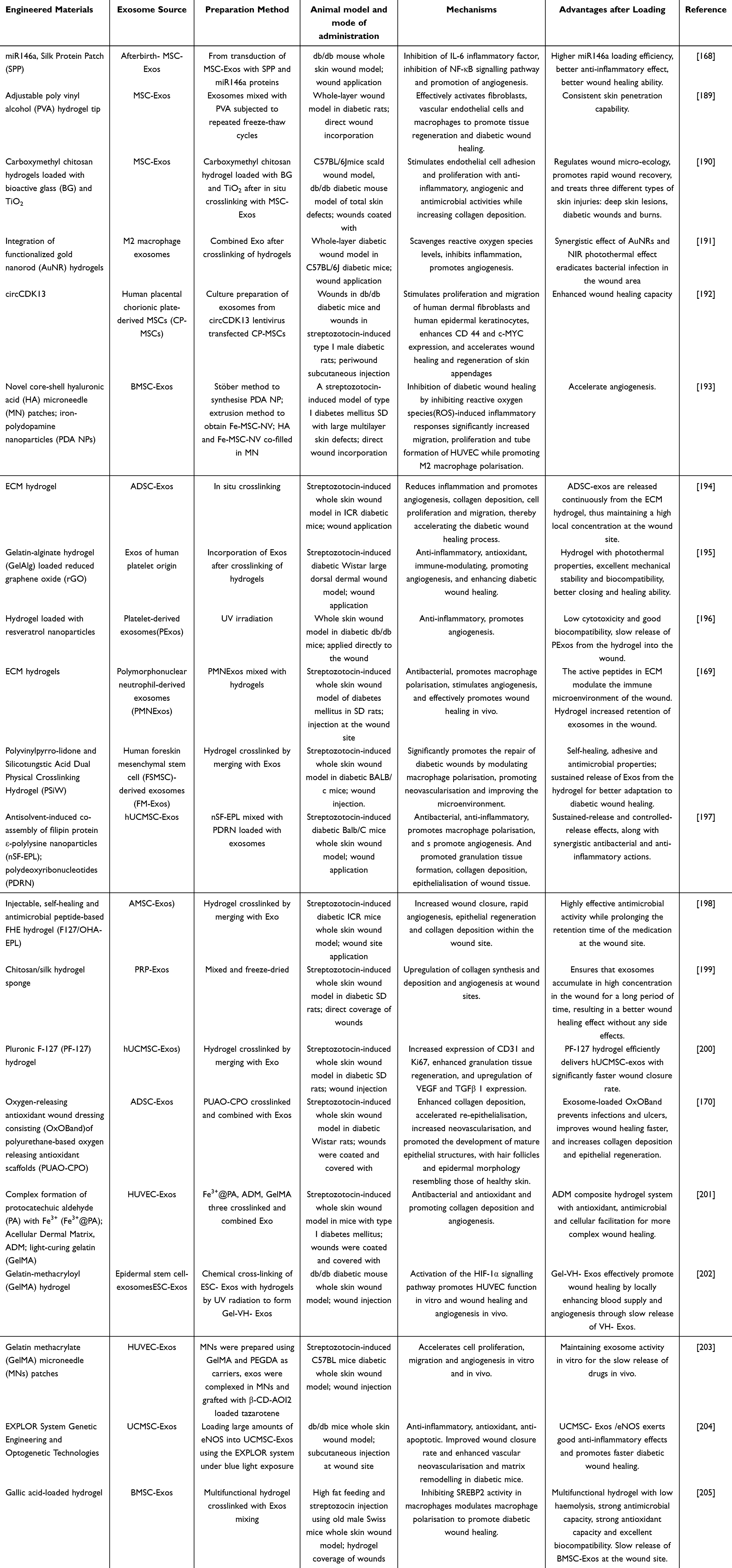 |
Table 6 The Role of Engineered Exosomes in Diabetic Wounds |
Scars
Following skin injury, the wound healing process often culminates in scar formation, which can be classified into four types: mature scars, immature scars, hypertrophic scars, and keloids.206 Scar formation is linked to metabolic dysfunction of connective tissue during the wound healing process, as well as the overproduction of activated fibroblasts and various components of the ECM.207 Exosomes are instrumental in scar management through mediating cell-to-cell communication.
Hypertrophic scars represent a category of fibroproliferative disorders that arise from surgical wounds or burns. In the course of wound healing, the prolonged persistence of myofibroblasts leads to excessive collagen accumulation, which ultimately results in the development of hypertrophic scars.208 ADSC-Exos ameliorate hypertrophic scarring by inhibiting collagen deposition and the transdifferentiation of fibroblasts into myofibroblasts, specifically delivering miR-125b-5p and miR-192-5p to suppress Smad signaling.208,209 MSC-Exos mitigate hypertrophic scarring by suppressing fibroblast proliferation through the TNFSF-13/HSPG2 signaling pathway.210 Additionally, exosomes derived from Lactobacillus delbrueckii can inhibit the expression of collagen I/III and α-SMA in fibroblasts, thereby suppressing the formation of hypertrophic scars in scleroderma mouse models.211 Additionally, engineered exosomes have been employed in the treatment of hypertrophic scars. Exosomes functionalized with miR-141-3p and incorporated into soluble microneedle arrays can modulate Smad expression, thereby enhancing the distribution of fibroblasts and the alignment of collagen fibers within hypertrophic scar tissue.118 These exosomes, particularly MSC-Exos, offer a promising therapeutic strategy for hypertrophic scars.
Skin Aging
Skin aging is affected by a combination of intrinsic, time-related factors and extrinsic, environmental factors. As skin ages, it undergoes structural, cellular, and molecular changes, including the buildup of senescent cells.212 Intrinsic aging leads to skin thinning due to reduced cell proliferation and changes in dermal components, with a decrease in ECM components like collagen and elastin causing fine lines. Additionally, the skin’s ability to repair itself diminishes, making it more susceptible to oxidative stress.212–214 Extrinsic aging, primarily caused by UV radiation, smoking, and pollution, results in impaired keratinocyte differentiation, reduced collagen, loss of melanocytes.215 Exosomes, especially those from stem cells, have shown anti-aging benefits by restoring skin functions and repairing damage through mechanisms like reducing oxidative stress, lowering MMPs, boosting collagen and elastin production, and enhancing cellular communication.117,216
Photoaging represents a major contributor to extrinsic skin aging, and several studies have reported the efficacy of exosomes in addressing skin photoaging. For instance, needle-free administration of exosomes sourced from human dermal fibroblast spheroids has been demonstrated to ameliorate UVB-induced photoaging by augmenting type I procollagen expression and dermal collagen deposition while concurrently reducing MMP-1 levels.217 Furthermore, exosome-like nanovesicles derived from Phellinus linteus, containing miR-CM1, have been shown to inhibit the expression of Mical2 in HaCaT cells through cross-species regulation, thereby reducing ROS levels and attenuating UV-induced skin aging.218 Nanovesicles from dermal papilla MSCs combat photoaging by reducing cell cycle arrest, cellular senescence, and macrophage infiltration, while enhancing cell proliferation, ECM production, and antioxidant enzyme activity.219 Cow’s milk exosomes (MK-Exos) aid fibroblast migration and restore collagen expression after UV exposure.220 Plant-derived exosomes offer antioxidant and anti-inflammatory benefits against skin photoaging. Exosome-like nanovesicles from olive leaf in a hydrogel deliver miR168a-5p to inhibit the NF-κB pathway, reducing inflammation and promoting skin repair after UV damage.221 Exosome-like nanoparticles from ginseng roots protect skin from UV and oxidative stress by inhibiting AP-1 signaling and reducing senescence and inflammation markers.222 Apple-derived exosomes boost type I collagen and lower MMPs by downregulating the NF-κB pathway, aiding in skin aging.223
Similarly, Engineered nanovesicles from human iPSCs improve aging signs in dermal fibroblasts by reducing SA-β-Gal activity and inhibiting p53 and p21.224 In addition, Combining exosomes with bioactive substances enhances their anti-aging effects. Co-administering hydrolyzed collagen oligopeptides and SC-Exos boosts cell migration and proliferation, reduces ROS production, and inhibits senescence-related genes, enhancing skin’s anti-aging abilities.225
Flap Regeneration
A flap, consisting of skin and subcutaneous fat with its own blood supply, is used in wound repair, organ reconstruction, or cosmetic procedures. Flap regeneration, the process of restoring blood flow and function post-transplantation, can be hindered by insufficient blood supply causing necrosis.226 SC-Exos improve flap regeneration due to their regenerative properties, with studies showing BMSC-Exos enhance flap survival after ischemia by reducing oxidative stress, inflammation, and apoptosis.227,228 Similarly, ADSC-Exos enhance neovascularization and reduce inflammation and apoptosis in skin grafts after ischemia-reperfusion injury.229 Moreover, Curcumin-loaded ADSC-Exos boost antioxidant and anti-inflammatory effects in flap regeneration.230 Exosomes from human dental pulp stem cells (hDPSC-Exos) enhance HUVEC proliferation, migration, and lumen formation via the PI3K/AKT pathway, improving flap survival.231 miRNAs in exosomes are crucial for flap regeneration, with hypoxia-treated BMSC-Exos carrying miR-421-3p targeting the mTOR/ULK1/FUNDC1 axis to activate autophagy and support flap survival after ischemia-reperfusion injury.119 ADSC-Exos treated with FGF1 deliver miR-183-5p, targeting GPR137 via the PI3K/Akt/mTOR pathway to reduce oxidative stress and endothelial cell apoptosis, enhancing ischemic flap survival.232 Beyond stem cells, M2 macrophages exhibit angiogenic potential during tissue repair. M2-derived exosomes facilitate angiogenesis and improve flap survival via the HIF-1AN/HIF-1α/VEGFA axis.233 Consequently, exosomes represent a significant therapeutic approach for flap regeneration.
Hair Regeneration
Alopecia, both scarring and non-scarring, affects the hair growth cycle, often shortening the anagen phase and extending the telogen phase due to various factors.234,235 Exosomes from different cell types have shown potential in promoting hair growth. These cell types include stem cells,236 keratinocytes,237 dermal papilla cells,238 and macrophages.239
Dermal papilla cells (DPCs) function as the signaling hub within hair follicles, orchestrating hair formation and cycling through paracrine mechanisms.240 Exosomes derived from DPCs (DPC-Exos) enhance intercellular communication and serve as a crucial factor in the treatment of alopecia. DPC-Exos can activate LEF1, thereby regulating the proliferation of hair follicle stem cells.241 Moreover, miR-218-5p and miR-181a-5p, which are conveyed by DPC-Exos, target the Wnt/β-catenin signaling pathway,120,242 while miR-140-5p targets the BMP/TGF-β signaling pathway, thus promoting hair follicle development.243 Engineered DPC-Exo hydrogels have been explored as a therapeutic strategy for alopecia, aiming to achieve more sustained and comprehensive therapeutic outcome.240 Additionally, reNcell-derived nanovesicles (ReN-NV) and MK-Exos, which exhibit stem cell-like properties, can activate the Wnt/β-catenin pathway to enhance DPC proliferation and expedite the transition of the hair follicle cycle.244,245 Exosomes derived from fibroblasts increase the expression of PCNA, pAKT, pERK, and VEGF receptor-2 (VEGFR2) in DPCs, thereby promoting their proliferation.246
Androgenetic alopecia (AGA) is a non-cicatricial hair loss disorder attributed to elevated levels of androgens. Recent advancements have seen the application of various SC-Exos in the therapeutic management of AGA. iPSC-Exos have been shown to ameliorate AGA by modulating growth factors and cytokines, as well as activating the androgen receptor (AR)-associated Wnt/β-catenin signaling pathway.247 ADSC-Exos, which carry miR-122-5p, target the TGF-β/SMAD3 axis to facilitate hair follicle regeneration in AGA.248 Additionally, hUCMSC-Exos enhance the stemness of hair follicle stem cells via the RAS/ERK pathway, thereby promoting hair proliferation in AGA mouse models.249 Moreover, the combination of SC-Exos with various hair loss treatment agents can yield a multifaceted synergistic effect in the management of AGA.250,251 In contrast, alopecia areata, an autoimmune-mediated condition, results from immune attacks on hair follicles.252 In this context, hMSC-Exos have demonstrated efficacy in promoting hair regeneration and hair shaft elongation in hair follicles of imiquimod-induced alopecia areata mice.253 Collectively, these exosomes offer a comprehensive therapeutic strategy for addressing hair loss.
Therapeutic Role of Exosomes in Skin Cancer
Malignant melanoma (MM) is a skin cancer. Upon exposure to external radiation, such as UV light, the skin initiates a DNA damage response, leading to the secretion of melanocyte-stimulating hormone (MSH) by keratinocytes. MSH subsequently binds to the melanocortin 1 receptor (MC1R) on melanocytes, stimulating melanin production. The metastasis of melanoma is primarily driven by genetic mutations and modifications within the tumor microenvironment, which are characterized by increased immunogenicity and the infiltration of immune cells into the tumor.254,255 Exosomes, which mediate intercellular communication, have emerged as important players in the diagnosis and treatment of skin cancer. Specifically, miRNAs present in exosomes derived from MM cells, contribute to tumor immune evasion and represent potential therapeutic targets.256 Furthermore, exosomes isolated from the lymphatic drainage of melanoma patients post-lymph node dissection contain proteins that are enriched and analogous to those associated with melanoma progression. Evaluating surrogate markers indicative of tumor progression within circulating extracellular vesicles could provide a robust non-invasive approach.257 Conversely, exosomes originating from MM cells have been implicated in the modulation of MM treatment. Research indicates that WNT5A stimulates the release of MM cell-derived exosomes, which contain immunomodulatory and pro-angiogenic proteins.258 Furthermore, exosomes carrying PD-L1, secreted by MM cells, facilitate the polarization of M2 macrophages, thereby enhancing resistance to therapies targeting programmed cell death 1 (PD-1)/programmed death ligand 1 (PD-L1).259
In addition to cancer cell-derived exosomes, those derived from normal cells and plants also contribute to MM treatment. Exosomes derived from NK cells have demonstrated the ability to suppress the proliferation of MM cells.260 Similarly, adipose stem cell-derived exosomes inhibit the progression of skin cancer through the miR-199a-5p/SOX4 pathway.261 Moreover, grapefruit-derived microvesicles and nanovesicles demonstrate a distinct metabolomic profile and exhibit anti-MM activity in the A375 human melanoma cell line.262
In addition to their diagnostic and therapeutic roles, exosomes have the potential to function as carriers for cancer therapeutic agents, specifically targeting MM cells. This facilitates the development of targeted drug delivery systems that offer synergistic enhancement and reduced toxicity. Nanovesicles derived from aloe vera, when loaded with indocyanine green (ICG), demonstrate efficient uptake by MM cells over prolonged periods, resulting in more effective inhibition of melanoma growth compared to free ICG.263 Furthermore, hUCMSC-Exos complexed with the cyclic peptide arginine-glycine-aspartic acid (cRGD) and encapsulating triptolide (TP) exhibit tumor-targeting capabilities. This formulation not only extends the half-life of TP but also significantly inhibits MM growth and prolongs the survival of mice bearing MM.264
Clinical Trials of Exosomes in Dermatological Conditions
Numerous case studies on exosome therapy for dermatological conditions have progressed to the stage of clinical research. These studies primarily focus on MSC-EXOs, plasma-derived exosomes, and include one clinical investigation involving plant-derived exosomes. Among these, MSC-EXOs are the most extensively utilized, aligning with foundational research findings. They have been employed in the treatment of conditions such as hair loss, psoriasis, skin aging, diabetic foot ulcers, melanoma, melasma, and dry eye syndrome, as outlined in the accompanying (Table 7). These clinical trials not only explore the application of exosomes as therapeutic agents for skin conditions but also investigate their potential as biomarkers. This dual functionality—as therapeutic agents and biomarkers—has been validated at the clinical trial level. Unfortunately, as of now, no exosome-based treatments for skin diseases have received approval for market release as medicinal products.
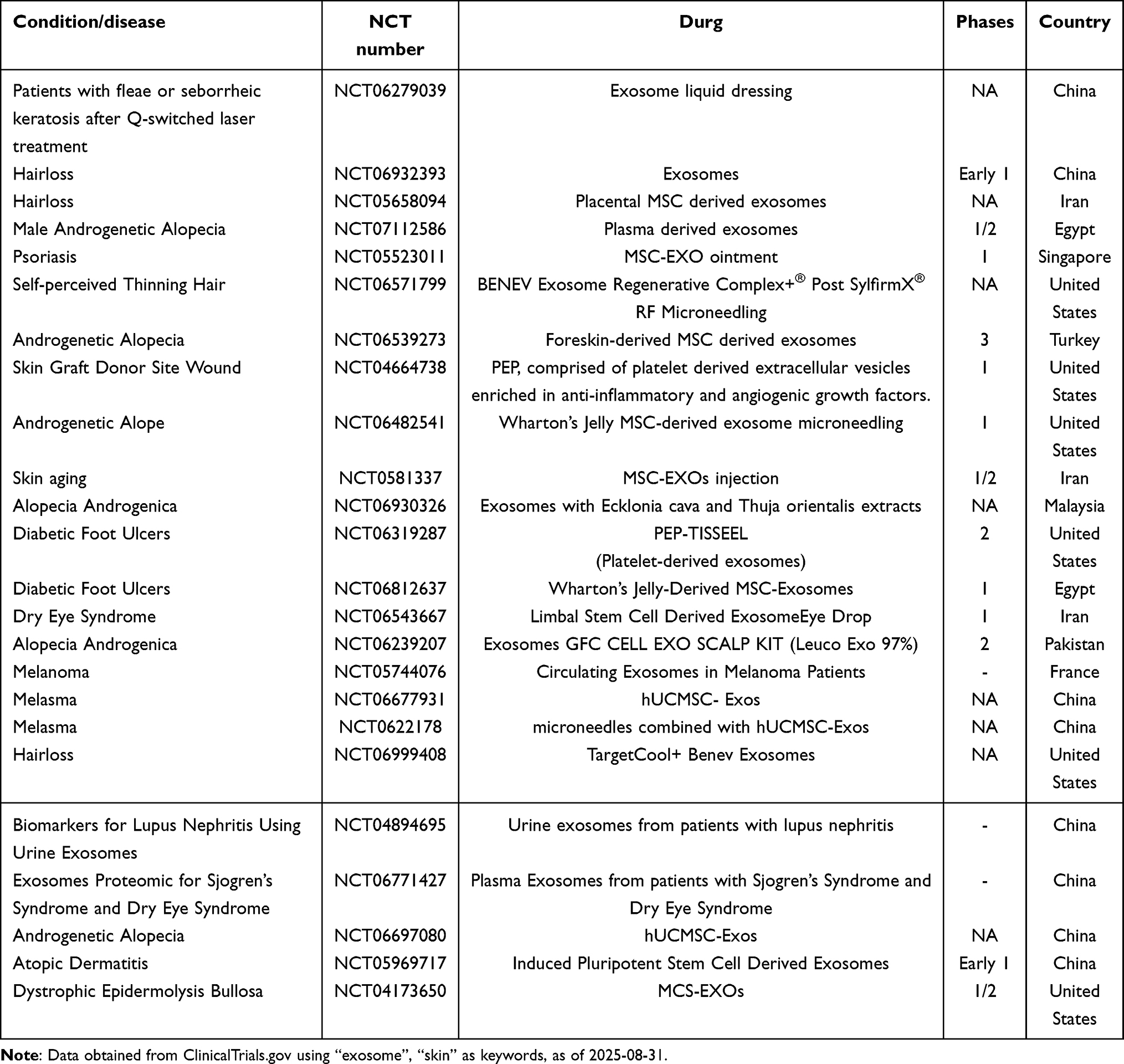 |
Table 7 Clinical Trials of Exosomes in Skin Diseases |
Limitations and Future Prospects of Exosomes in Skin Therapy
Despite significant advancements in fundamental research on exosomes within the field of dermatology in recent years, numerous challenges remain concerning their clinical application as therapeutic agents. Firstly, exosomes originate from a wide array of sources, resulting in compositional variations between those derived from different cell types. Even among exosomes from the same cell type, variations in composition are observed at different stages of cellular growth.116 The nucleic acids, proteins, and lipids within exosomes constitute a highly complex mixture. Although exosomes exhibit promising therapeutic potential across various diseases, the specific therapeutic components have yet to be clearly identified. Research has predominantly focused on microRNAs, with limited studies exploring the detailed composition and mechanisms of action of proteins and lipids. Therefore, future research should prioritize elucidating the therapeutic constituents of exosomes and their specific mechanisms of action. 2) Researchers have devised a range of methodologies for exosome isolation, grounded in principles such as density, size, and immunoreactivity. Nevertheless, each method is accompanied by distinct limitations. For example, differential ultracentrifugation, the most frequently utilized technique, requires expensive equipment and skilled personnel. Size-exclusion chromatography is prone to equipment blockage, whereas immunocapture methods are restricted to isolating specific subsets of exosomes. Furthermore, the exosome isolation and purification process is complex and time-consuming, with operator variability potentially resulting in inconsistencies in the composition of isolated exosomes.10,265 These inconsistencies hinder the precise identification of exosomes and subsequent experimental analyses. Therefore, the development of cost-effective, scalable, and reliable methods for exosome isolation is crucial for advancing their application in the medical field. 3) The harvested exosomes necessitate long-term storage at −80°C, as their biological activity is only preserved for approximately one week when stored at 4°C. Nevertheless, storage at −80°C is not without its drawbacks, as it adversely affects the stability of exosomes, with degradation observed even after extended storage periods of up to two years. This presents considerable challenges for the storage and transportation of exosomes. Although exosome lyophilization techniques have been developed to facilitate stable preservation at room temperature, there is a lack of sufficient experimental evidence to ascertain whether lyophilization affects their biological activity.266 Consequently, there is an urgent need for more convenient, durable, and stable preservation methods in exosome research. 4) Due to the ambiguous composition of exosomes, particularly the immunogenicity associated with cell-derived exosomes, may result in significant heterogeneity between different donors and recipients. While studies suggest that plant-derived exosomes exhibit lower immunogenicity and enhanced safety profiles, their application in the treatment of skin diseases remains relatively unexplored. Additionally, the identification of plant exosomes continues to pose a significant challenge. The therapeutic application of exosomes in disease management faces several challenges.267 Establishing standardized protocols for the identification of plant-derived exosomes, in conjunction with fundamental research and clinical applications in dermatology, is of paramount importance for future investigations. 5) Currently, there are over a hundred instances of exosomes being evaluated in clinical trials across various disciplines, including orthopaedics (NCT05060107), neurosurgery (NCT03384433), plastic surgery (NCT02565264), and ophthalmology (NCT05413148), with the majority still in the preclinical research phase.268 Despite extensive foundational research on exosome therapies for dermatological conditions, only a limited number of studies have advanced to clinical trials. Future research efforts should focus on translating basic scientific findings into clinical applications and developing safe and effective exosome formulations specifically designed for dermatological therapies. Although there are inherent challenges in the medical application of exosomes, particularly in the treatment of skin diseases, we propose that interdisciplinary advancements in medicine, bioengineering, nanotechnology, and related fields will facilitate significant progress in dermatological therapeutics. This advancement is expected to result in the development of safer, more effective, and varied therapeutic modalities for a wide spectrum of human diseases.
Conclusion
In recent years, exosomes have been extensively utilized across various domains of medical treatment, including cardiovascular diseases, immune disorders, degenerative diseases, inflammation, aging, and regenerative medicine. This is attributed to their diverse origins, favorable biocompatibility, and capacity to transport nucleic acids, proteins, lipids, and other components involved in intercellular communication. Notably, the phospholipid bilayer structure of exosomes confers exceptional biocompatibility with the skin, enhancing their absorption. This review consolidates recent studies on the utilization of exosomes as biomarkers, therapeutic agents, and drug delivery systems in dermatological disorders. It offers a comprehensive analysis of the therapeutic potential of exosomes in autoimmune skin disorders, such as SLE, SC, psoriasis, and vitiligo, as well as in skin regenerative conditions, including wound healing, skin aging, hypertrophic scarring, flap regeneration, and hair regeneration. Furthermore, exosomes demonstrate promising therapeutic roles in the treatment of skin cancers. These results offer compelling therapeutic approaches for managing skin diseases and establish a strong basis for the advancement of exosome-based therapies in dermatology.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was supported by Jilin Province Changbai Yingcai-Young Postdoctoral Program.
Disclosure
The authors declare that they have no known competing financial interests or personal relationships.
References
1. Raszewska-Famielec M, Flieger J. Nanoparticles for topical application in the treatment of skin dysfunctions—an overview of dermo-cosmetic and dermatological products. Int J Mol Sci. 2022;23(24). doi:10.3390/ijms232415980
2. Metcalfe AD, Ferguson MWJ. Bioengineering skin using mechanisms of regeneration and repair. Biomaterials. 2007;28(34):5100–5113. doi:10.1016/j.biomaterials.2007.07.031
3. Bouwstra JA, Nădăban A, Bras W, et al. The skin barrier: an extraordinary interface with an exceptional lipid organization. Prog lipid res. 2023:92. doi:10.1016/j.plipres.2023.101252
4. Chen R, Zhu Z, Ji S, et al. Sweat gland regeneration: current strategies and future opportunities. Biomaterials. 2020;255. doi:10.1016/j.biomaterials.2020.120201
5. Hosseini M, Koehler KR, Shafiee A. Biofabrication of human skin with its appendages. Adv Healthcare Mater. 2022;11(22). doi:10.1002/adhm.202201626
6. Schneider MR, Schmidt-Ullrich R, Paus R. The hair follicle as a dynamic miniorgan. Curr Biol. 2009;19(3):R132–R142. doi:10.1016/j.cub.2008.12.005
7. Pegtel DM, Gould SJ. Exosomes. Annu Rev Biochem. 2019;88(1):487–514. doi:10.1146/annurev-biochem-013118-111902
8. Jeppesen DK, Fenix AM, Franklin JL, et al. Reassessment of exosome composition. Cell. 2019;177(2):428–445.e18. doi:10.1016/j.cell.2019.02.029
9. Ibrahim A, Marbán E. Exosomes: fundamental biology and roles in cardiovascular physiology. Annual Rev Physiol. 2016;78(1):67–83. doi:10.1146/annurev-physiol-021115-104929
10. Kimiz-Gebologlu I, Oncel SS. Exosomes: large-scale production, isolation, drug loading efficiency, and biodistribution and uptake. J Control Release. 2022;347:533–543. doi:10.1016/j.jconrel.2022.05.027
11. Yang D, Zhang W, Zhang H, et al. Progress, opportunity, and perspective on exosome isolation - efforts for efficient exosome-based theranostics. Theranostics. 2020;10(8):3684–3707. doi:10.7150/thno.41580
12. Zhang F, Jiang J, Qian H, et al. Exosomal circRNA: emerging insights into cancer progression and clinical application potential. J hematol oncol. 2023;16(1). doi:10.1186/s13045-023-01452-2
13. Ha DH, Kim HK, Lee J, et al. Mesenchymal stem/stromal cell-derived exosomes for immunomodulatory therapeutics and skin regeneration. Cells. 2020;9(5). doi:10.3390/cells9051157
14. Wu RJ, Li HT, Sun CW, et al. Exosome-based strategy for degenerative disease in orthopedics: recent progress and perspectives. J Orthopaedic Transl. 2022;36:8–17. doi:10.1016/j.jot.2022.05.009
15. Wang Y, Li MW, Chen L, et al. Natural killer cell-derived exosomal miR-1249-3p attenuates insulin resistance and inflammation in mouse models of type 2 diabetes. Signal Transd Targeted Ther. 2021;6(1):1.
16. Li JL, Sun SH, Zhu DS, et al. Inhalable stem cell exosomes promote heart repair after myocardial infarction. Circulation. 2024;150(9):710–723. doi:10.1161/Circulationaha.123.065005
17. Li W, Yang S, Zhao Y, et al. Ginseng-derived nanoparticles alleviate alcohol-induced liver injury by activating the Nrf2/HO-1 signaling pathway and inhibiting the NF-κB signaling pathway in vitro and in vivo. Phytomedicine. 2024:127. doi:10.1016/j.phymed.2024.155428
18. Zhang YY, Yi D, Hong Q, et al. Platelet-rich plasma-derived exosomes boost mesenchymal stem cells to promote peripheral nerve regeneration. J Control Release. 2024;367:265–282. doi:10.1016/j.jconrel.2024.01.043
19. Pan Y, Hui XY, Hoo RLC, et al. Adipocyte-secreted exosomal microRNA-34a inhibits M2 macrophage polarization to promote obesity-induced adipose inflammation. J Clin Investig. 2019;129(2):834–849. doi:10.1172/Jci123069
20. Yang S, Li WJ, Bai XY, et al. Ginseng-derived nanoparticles alleviate inflammatory bowel disease via the TLR4/MAPK and p62/Nrf2/Keap1 pathways. J Nanobiotechnol. 2024;22(1). doi:10.1186/s12951-024-02313-x
21. Zou WC, Lai MQ, Jiang YJ, et al. Exosome release delays senescence by disposing of obsolete biomolecules. Adv Sci. 2023;10(8). doi:10.1002/advs.202204826
22. Qu F, Geng R, Liu Y, et al. Advanced nanocarrier- and microneedle-based transdermal drug delivery strategies for skin diseases treatment. Theranostics. 2022;12(7):3372–3406. doi:10.7150/thno.69999
23. Shen Y, Xu G, Huang H, et al. Sequential release of small extracellular vesicles from bilayered thiolated alginate/polyethylene glycol diacrylate hydrogels for scarless wound healing. ACS Nano. 2021;15(4):6352–6368. doi:10.1021/acsnano.0c07714
24. Keshtkar S, Asvar Z, Najafi H, et al. Exosomes as natural vectors for therapeutic delivery of bioactive compounds in skin diseases. Front Pharmacol. 2025. doi:10.3389/fphar.2025.1485769
25. Dal’Forno-Dini T, Birck MS, Rocha M, et al. Exploring the reality of exosomes in dermatology. Anais Brasileiros de Dermatologia. 2025;100(1):121–130. doi:10.1016/j.abd.2024.09.002
26. Chen H, Jiang Y, Li X. Adriamycin‐loaded exosome with anti‐CD20 aptamers selectively suppresses human CD20+ melanoma stem cells. Skin Res Technol. 2022;29(1). doi:10.1111/srt.13259
27. Kumar DN, Chaudhuri A, Dehari D, et al. Enhanced therapeutic efficacy against melanoma through exosomal delivery of hesperidin. Mol Pharmaceut. 2024;21(6):3061–3076. doi:10.1021/acs.molpharmaceut.4c00490
28. Yao Y, Wang Z, Li J, et al. Pyroptosis and its role in autoimmune skin disease. Exp Dermatol. 2024;33(7). doi:10.1111/exd.15135
29. Fang Y, Ni J, Wang Y-S, et al. Exosomes as biomarkers and therapeutic delivery for autoimmune diseases: opportunities and challenges. Autoimmunity Rev. 2023;22(3). doi:10.1016/j.autrev.2022.103260
30. Bieber K, Hundt JE, Yu X, et al. Autoimmune pre-disease. Autoimmunity Rev. 2023;22(2). doi:10.1016/j.autrev.2022.103236
31. Kiriakidou M, Ching CL. Systemic lupus erythematosus. Ann Internal Med. 2020;172(11):ITC81–ITC96. doi:10.7326/aitc202006020
32. Perez-Hernandez J, Redon J, Cortes R. Extracellular vesicles as therapeutic agents in systemic lupus erythematosus. Int J Mol Sci. 2017;18(4). doi:10.3390/ijms18040717
33. Lin Z, Wu Y, Xu Y, et al. Mesenchymal stem cell-derived exosomes in cancer therapy resistance: recent advances and therapeutic potential. Mol Cancer. 2022;21(1). doi:10.1186/s12943-022-01650-5
34. Ma W, Che J, Chen W, et al. Dexamethasone-integrated mesenchymal stem cells for systemic lupus erythematosus treatment via multiple immunomodulatory mechanisms. ACS Nano. 2024;18(20):13249–13265. doi:10.1021/acsnano.4c02420
35. Zhang M, Johnson-Stephenson TK, Wang W, et al. Mesenchymal stem cell-derived exosome-educated macrophages alleviate systemic lupus erythematosus by promoting efferocytosis and recruitment of IL-17+ regulatory T cell. Stem Cell Res Ther. 2022;13(1). doi:10.1186/s13287-022-03174-7
36. Sun W, Yan S, Yang C, et al. Mesenchymal stem cells-derived exosomes ameliorate lupus by inducing M2 macrophage polarization and regulatory T cell expansion in MRL/lpr mice. Immunol invest. 2022;51(6):1785–1803. doi:10.1080/08820139.2022.2055478
37. T YY, Kentaro A, Chen C, et al. Immunomodulatory properties of stem cells from human exfoliated deciduous teeth. Stem Cell Res Ther. 2010;1(1):5. doi:10.1186/scrt5
38. Sonoda S, Murata S, Kato H, et al. Targeting of deciduous tooth pulp stem cell–derived extracellular vesicles on telomerase-mediated stem cell niche and immune regulation in systemic lupus erythematosus. J Immunol. 2021;206(12):3053–3063. doi:10.4049/jimmunol.2001312
39. Luo B, Zhou K, Liufu Y, et al. Novel insight into miRNA biology and its role in the pathogenesis of systemic lupus erythematosus. Front Immunol. 2022:13. doi:10.3389/fimmu.2022.1059887
40. Young NA, Schwarz E, Zeno BM, et al. Inhibition of miRNA associated with a disease-specific signature and secreted via extracellular vesicles of systemic lupus erythematosus patients suppresses target organ inflammation in a humanized mouse model. Front Immunol. 2024:14. doi:10.3389/fimmu.2023.1090177
41. Wang W, Yue C, Gao S, et al. Promising roles of exosomal microRNAs in systemic lupus erythematosus. Front Immunol. 2021. doi:10.3389/fimmu.2021.757096
42. Perez-Hernandez J, Martinez-Arroyo O, Ortega A, et al. Urinary exosomal miR-146a as a marker of albuminuria, activity changes and disease flares in lupus nephritis. J Nephrol. 2020;34(4):1157–1167. doi:10.1007/s40620-020-00832-y
43. Solé C, Moliné T, Vidal M, et al. An exosomal urinary miRNA signature for early diagnosis of renal fibrosis in lupus nephritis. Cells. 2019;8(8). doi:10.3390/cells8080773
44. Rastaldi MP, Ichii O, Otsuka-Kanazawa S, et al. Decreased miR-26a expression correlates with the progression of podocyte injury in autoimmune glomerulonephritis. PLoS One. 2014;9(10). doi:10.1371/journal.pone.0110383
45. Tan L, Zhao M, Wu H, et al. Downregulated serum exosomal miR-451a expression correlates with renal damage and its intercellular communication role in systemic lupus erythematosus. Front Immunol. 2021. doi:10.3389/fimmu.2021.630112
46. Yan S, Huang Z, Chen X, et al. Metabolic profiling of urinary exosomes for systemic lupus erythematosus discrimination based on HPL-SEC/MALDI-TOF MS. Anal Bioanal Chem. 2023;415(26):6411–6420. doi:10.1007/s00216-023-04916-z
47. Flores-Chova A, Martinez-Arroyo O, Riffo-Campos AL, et al. Plasma exosomal non-coding RNA profile associated with renal damage reveals potential therapeutic targets in lupus nephritis. Int J Mol Sci. 2023;24(8). doi:10.3390/ijms24087088
48. Chuang H-C, Chen M-H, Chen Y-M, et al. BPI overexpression suppresses treg differentiation and induces exosome-mediated inflammation in systemic lupus erythematosus. Theranostics. 2021;11(20):9953–9966. doi:10.7150/thno.63743
49. Denton CP, Khanna D. Systemic sclerosis. Lancet. 2017;390(10103):1685–1699. doi:10.1016/s0140-6736(17)30933-9
50. Jerjen R, Nikpour M, Krieg T, et al. Systemic sclerosis in adults. Part I: clinical features and pathogenesis. J Am Acad Dermatol. 2022;87(5):937–954. doi:10.1016/j.jaad.2021.10.065
51. De Lorenzis E, Rindone A, Di Donato S, et al. Circulating extracellular vesicles in the context of interstitial lung disease related to systemic sclerosis: a scoping literature review. Autoimmunity Rev. 2023;22(9). doi:10.1016/j.autrev.2023.103401
52. Rozier P, Maumus M, Bony C, et al. Extracellular vesicles are more potent than adipose mesenchymal stromal cells to exert an anti-fibrotic effect in an in vitro model of systemic sclerosis. Int J Mol Sci. 2021;22(13). doi:10.3390/ijms22136837
53. Zhao K, Kong C, Shi N, et al. Potential angiogenic, immunomodulatory, and antifibrotic effects of mesenchymal stem cell-derived extracellular vesicles in systemic sclerosis. Front Immunol. 2023. doi:10.3389/fimmu.2023.1125257
54. Imenez SA, Piera-Velazquez S. Probable role of exosomes in the extension of fibrotic alterations from affected to normal cells in systemic sclerosis. Rheumatology. 2023;62(3):999–1008. doi:10.1093/rheumatology/keac451/6659115
55. Kanemaru H, Kajihara I, Yamanaka K, et al. Platelet-rich plasma therapy is effective for the treatment of refractory skin ulcers in patients with systemic sclerosis. Mod Rheumatol. 2014;25(4):660–661. doi:10.3109/14397595.2014.924204
56. Nakamura K, Jinnin M, Harada M, et al. Altered expression of CD63 and exosomes in scleroderma dermal fibroblasts. J Dermatological Sci. 2016;84(1):30–39. doi:10.1016/j.jdermsci.2016.06.013
57. Li L, Zuo X, Liu D, et al. The profiles of miRNAs and lncRNAs in peripheral blood neutrophils exosomes of diffuse cutaneous systemic sclerosis. J Dermatological Sci. 2020;98(2):88–97. doi:10.1016/j.jdermsci.2020.02.009
58. Jin J, Ou Q, Wang Z, et al. BMSC-derived extracellular vesicles intervened the pathogenic changes of scleroderma in mice through miRNAs. Stem Cell Res Ther. 2021;12(1). doi:10.1186/s13287-021-02400-y
59. Xie L, Long X, Mo M, et al. Bone marrow mesenchymal stem cell-derived exosomes alleviate skin fibrosis in systemic sclerosis by inhibiting the IL-33/ST2 axis via the delivery of microRNA-214. Mol Immunol. 2023;157:146–157. doi:10.1016/j.molimm.2023.03.017
60. Rozier P, Maumus M, Maria ATJ, et al. Mesenchymal stromal cells-derived extracellular vesicles alleviate systemic sclerosis via miR-29a-3p. J Autoimmun. 2021;121. doi:10.1016/j.jaut.2021.102660
61. Baral H, Uchiyama A, Yokoyama Y, et al. Antifibrotic effects and mechanisms of mesenchymal stem cell-derived exosomes in a systemic sclerosis mouse model: possible contribution of miR-196b-5p. J Dermatological Sci. 2021;104(1):39–47. doi:10.1016/j.jdermsci.2021.08.006
62. Zhu K, Liu C, Guo X, et al. Exosomal miR‐126‐3p: potential protection against vascular damage by regulating the SLC7A5/mTOR signaling pathway in human umbilical vein endothelial cells. Scand J Immunol. 2024;99(4). doi:10.1111/sji.13354
63. Li L, Zuo X, Xiao Y, et al. Neutrophil-derived exosome from systemic sclerosis inhibits the proliferation and migration of endothelial cells. Biochem Biophys Res Commun. 2020;526(2):334–340. doi:10.1016/j.bbrc.2020.03.088
64. Griffiths CEM, Armstrong AW, Gudjonsson JE, et al. Psoriasis. Lancet. 2021;397(10281):1301–1315. doi:10.1016/s0140-6736(20)32549-6
65. Griffiths CEM, Barker JNWN. Pathogenesis and clinical features of psoriasis. Lancet. 2007;370(9583):263–271. doi:10.1016/s0140-6736(07)61128-3
66. Iuliano M, Grimaldi L, Rosa P, et al. Extracellular vescicles in psoriasis: from pathogenesis to possible roles in therapy. Front Immunol. 2024:15. doi:10.3389/fimmu.2024.1360618
67. Cheng L, Wang S, Peng C, et al. Human umbilical cord mesenchymal stem cells for psoriasis: a Phase 1/2a, single-arm study. Signal Transd Targeted Ther. 2022;7(1). doi:10.1038/s41392-022-01059-y
68. Meybodi MAM, Nilforoushzadeh MA, KhandanDezfully N, et al. The safety and efficacy of adipose tissue-derived exosomes in treating mild to moderate plaque psoriasis: a clinical study. Life Sci. 2024:353. doi:10.1016/j.lfs.2024.122915
69. Zhang W, Lin J, Shi P, et al. Small extracellular vesicles derived from MSCs have immunomodulatory effects to enhance delivery of ASO-210 for Psoriasis Treatment. Front Cell Develop Biol. 2022. doi:10.3389/fcell.2022.842813
70. Ye Z, Liang Y, Lin B, et al. Gingiva-derived mesenchymal stem cells attenuate imiquimod- (IMQ-) induced murine psoriasis-like skin inflammation. Stem Cells Int. 2022;2022:1–16. doi:10.1155/2022/6544514
71. Abed ZI, Arianejad M, Azizi Z. Mesenchymal stem cell-derived exosomes decrease hyperplasia in psoriasis by inducing transforming growth factor β2 (TGF-β2). Mol Biol Rep. 2024;51(1). doi:10.1007/s11033-024-09337-4
72. Song M, Lim KM, Song K, et al. Efficient treatment of psoriasis using conditioned media from mesenchymal stem cell spheroids cultured to produce transforming growth factor-β1-enriched small-sized extracellular vesicles. Int J Stem Cells. 2024. doi:10.15283/ijsc24089
73. Xu F, Fei Z, Dai H, et al. Mesenchymal stem cell‐derived extracellular vesicles with high PD‐L1 expression for autoimmune diseases treatment. Adv Mater. 2021;34(1). doi:10.1002/adma.202106265
74. Huang R, Jia B, Su D, et al. Plant exosomes fused with engineered mesenchymal stem cell‐derived nanovesicles for synergistic therapy of autoimmune skin disorders. J Extracell Vesicles. 2023;12(10). doi:10.1002/jev2.12361
75. Subudhi I, Konieczny P, Prystupa A, et al. Metabolic coordination between skin epithelium and type 17 immunity sustains chronic skin inflammation. Immunity. 2024;57(7):1665–1680.e7. doi:10.1016/j.immuni.2024.04.022
76. Jiang X, Jiang Z, Huang S, et al. Ultraviolet B radiation-induced JPH203-loaded keratinocyte extracellular vesicles exert etiological interventions for psoriasis therapy. J Control Release. 2023;362:468–478. doi:10.1016/j.jconrel.2023.08.059
77. Rodrigues SC, Cardoso RMS, Freire PC, et al. Immunomodulatory properties of umbilical cord blood-derived small extracellular vesicles and their therapeutic potential for inflammatory skin disorders. Int J Mol Sci. 2021;22(18). doi:10.3390/ijms22189797
78. Shao S, Fang H, Zhang J, et al. Neutrophil exosomes enhance the skin autoinflammation in generalized pustular psoriasis via activating keratinocytes. FASEB J. 2019;33(6):6813–6828. doi:10.1096/fj.201802090RR
79. Boix-Amorós A, Badri MH, Manasson J, et al. Alterations in the cutaneous microbiome of patients with psoriasis and psoriatic arthritis reveal similarities between non-lesional and lesional skin. Ann Rheumatic Dis. 2023;82(4):507–514. doi:10.1136/ard-2022-223389
80. Chang C-J, Zhang J, Tsai Y-L, et al. Compositional features of distinct microbiota base on serum extracellular vesicle metagenomics analysis in moderate to severe psoriasis patients. Cells. 2021;10(9). doi:10.3390/cells10092349
81. Xu Y, Gan Y, Qi F, et al. Innate lymphoid cell-based immunomodulatory hydrogel microspheres containing Cutibacterium acnes extracellular vesicles for the treatment of psoriasis. Acta Biomater. 2024;184:296–312. doi:10.1016/j.actbio.2024.06.006
82. Gómez-Chávez F, Cedillo-Peláez C, Zapi-Colín LA, et al. The extracellular vesicles from the commensal staphylococcus epidermidis ATCC12228 strain regulate skin inflammation in the imiquimod-induced psoriasis murine model. Int J Mol Sci. 2021;22(23). doi:10.3390/ijms222313029
83. Eaton LH, Chularojanamontri L, Ali FR, et al. Guttate psoriasis is associated with an intermediate phenotype of impaired langerhans cell migration. Br J Dermatol. 2014;171(2):409–411. doi:10.1111/bjd.12960
84. Cully M. Anti-CD1a antibody reduces inflammation in psoriasis. Nat Rev Drug Discov. 2016;15(10):678. doi:10.1038/nrd.2016.192
85. Cheung KL, Jarrett R, Subramaniam S, et al. Psoriatic T cells recognize neolipid antigens generated by mast cell phospholipase delivered by exosomes and presented by CD1a. J Exp Med. 2016;213(11):2399–2412. doi:10.1084/jem.20160258
86. Jiang W, Zhang T, Qiu Y, et al. Keratinocyte-to-macrophage communication exacerbate psoriasiform dermatitis via LRG1-enriched extracellular vesicles. Theranostics. 2024;14(3):1049–1064. doi:10.7150/thno.89180
87. Ritchlin CT, Longo DL, Colbert RA, et al. Psoriatic Arthritis. N Engl J Med. 2017;376(10):957–970. doi:10.1056/NEJMra1505557
88. Lättekivi F, Guljavina I, Midekessa G, et al. Profiling blood serum extracellular vesicles in plaque psoriasis and psoriatic arthritis patients reveals potential disease biomarkers. Int J Mol Sci. 2022;23(7). doi:10.3390/ijms23074005
89. Paolino G, Buratta S, Mercuri SR, et al. Lipidic profile changes in exosomes and microvesicles derived from plasma of monoclonal antibody-treated psoriatic patients. Front Cell Develop Biol. 2022. doi:10.3389/fcell.2022.923769
90. Pasquali L, Svedbom A, Srivastava A, et al. Circulating microRNAs in extracellular vesicles as potential biomarkers for psoriatic arthritis in patients with psoriasis. J Eur Acad Dermatol Venereol. 2020;34(6):1248–1256. doi:10.1111/jdv.16203
91. Nematian SE, Mamillapalli R, Kadakia TS, et al. Systemic inflammation induced by microRNAs: endometriosis-derived alterations in circulating microRNA 125b-5p and Let-7b-5p regulate macrophage cytokine production. J Clin Endocrinol Metab. 2018;103(1):64–74. doi:10.1210/jc.2017-01199
92. Li X, Sun M, Long Y. Cyanidin-3-O-glucoside attenuates lipopolysaccharide-induced inflammation in human corneal epithelial cells by inducing let-7b-5p-mediated HMGA2/PI3K/Akt pathway. Inflammation. 2020;43(3):1088–1096. doi:10.1007/s10753-020-01194-0
93. Li J, Sugihara H, Ishimoto T, et al. Identification of miR-30e* regulation of bmi1 expression mediated by tumor-associated macrophages in gastrointestinal cancer. PLoS One. 2013;8(11). doi:10.1371/journal.pone.0081839
94. Iwamoto N, Fukui S, Takatani A, et al. Osteogenic differentiation of fibroblast-like synovial cells in rheumatoid arthritis is induced by microRNA-218 through a ROBO/Slit pathway. Arthritis Res Ther. 2018;20(1). doi:10.1186/s13075-018-1703-z
95. Yan K, Zhu J, Zhang M, et al. Differential expression of plasma extracellular vesicle miRNAs as biomarkers for distinguishing psoriatic arthritis from psoriasis. Chin Med J. 2024. doi:10.1097/cm9.0000000000003288
96. Wang Y, Liu Z, Ren X, et al. Hsa-miR-330-5p aggravates thyroid carcinoma via targeting FOXE1. J Oncol. 2021;2021:1–9. doi:10.1155/2021/1070365
97. Liu M, Zhang G, Wang Z, et al. FOXE1 contributes to the development of psoriasis by regulating WNT5A. J Invest Dermatol. 2023;143(12):2366–2377.e7. doi:10.1016/j.jid.2023.04.035
98. Park YJ, Kim DC, Lee S-J, et al. Keratinocyte-derived circulating microRNAs in extracellular vesicles: a novel biomarker of psoriasis severity and potential therapeutic target. J Transl Med. 2024;22(1). doi:10.1186/s12967-024-05030-z
99. Jiang M, Fang H, Dang E, et al. Small extracellular vesicles containing miR-381-3p from keratinocytes promote T helper type 1 and T helper type 17 polarization in psoriasis. J Invest Dermatol. 2021;141(3):563–574. doi:10.1016/j.jid.2020.07.009
100. Guo S, Zhang W, Wei C, et al. Serum and skin levels of miR-369-3p in patients with psoriasis and their correlation with disease severity. Eur J Dermatol. 2013;23(5):608–613. doi:10.1684/ejd.2013.2148
101. Seifeldin NS, Sayed SBE, Asaad MK. Increased MicroRNA‐1266 levels as a biomarker for disease activity in psoriasis vulgaris. Int J Dermatol. 2016;55(11):1242–1247. doi:10.1111/ijd.13102
102. Wang Z-Y, Yan B-X, Zhou Y, et al. miRNA profiling of extracellular vesicles reveals biomarkers for psoriasis. J Invest Dermatol. 2021;141(1):185–189.e4. doi:10.1016/j.jid.2020.04.021
103. Nie X, Deng W, Zhou H, et al. Long noncoding RNA MCM3AP-AS1 attenuates sepsis-induced cardiomyopathy by improving inflammation, oxidative stress, and mitochondrial function through mediating the miR-501-3p/CADM1/STAT3 axis. Int Immunopharmacol. 2024:128. doi:10.1016/j.intimp.2024.111500
104. Liew WC, Sundaram GM, Quah S, et al. Belinostat resolves skin barrier defects in atopic dermatitis by targeting the dysregulated miR-335:SOX6 axis. J Allergy Clin Immunol. 2020;146(3):606–620.e12. doi:10.1016/j.jaci.2020.02.007
105. Capogrossi MC, Lerman G, Avivi C, et al. MiRNA expression in psoriatic skin: reciprocal regulation of hsa-miR-99a and IGF-1R. PLoS One. 2011;6(6). doi:10.1371/journal.pone.0020916
106. Wang R, Huang Y, Shao K, et al. High expression of miR-6785-5p in the serum exosomes of psoriasis patients alleviates psoriasis-like skin damage by interfering with the MNK2/p-eIF4E Axis in keratinocytes. Inflammation. 2024;47(5):1585–1599. doi:10.1007/s10753-024-01995-7
107. Ezzedine K, Eleftheriadou V, Whitton M, et al. Vitiligo. Lancet. 2015;386(9988):74–84. doi:10.1016/s0140-6736(14)60763-7
108. Colletti M, Galardi A, De Santis M, et al. Exosomes in systemic sclerosis: messengers between immune, vascular and fibrotic components? Int J Mol Sci. 2019;20(18). doi:10.3390/ijms20184337
109. Wong PM, Yang L, Yang L, et al. New insight into the role of exosomes in vitiligo. Autoimmunity Rev. 2020;19(11). doi:10.1016/j.autrev.2020.102664
110. Frisoli ML, Essien K, Harris JE. Vitiligo: mechanisms of pathogenesis and treatment. Ann Rev Immunol. 2020;38(1):621–648. doi:10.1146/annurev-immunol-100919-023531
111. Wang Q, Guo W, Niu L, et al. 3D‐hUMSCs exosomes ameliorate vitiligo by simultaneously potentiating treg cells‐mediated immunosuppression and suppressing oxidative stress‐induced melanocyte damage. Adv Sci. 2024;11(31). doi:10.1002/advs.202404064
112. Li D, Zhou T, She Q, et al. Circulating exosomal miR-493-3p affects melanocyte survival and function by regulating epidermal dopamine concentration in segmental vitiligo. J Invest Dermatol. 2022;142(12):3262–3273.e11. doi:10.1016/j.jid.2022.05.1086
113. Zhao C, Wang D, Wang X, et al. Down‐regulation of exosomal miR‐200c derived from keratinocytes in vitiligo lesions suppresses melanogenesis. J Cell Mol Med. 2020;24(20):12164–12175. doi:10.1111/jcmm.15864
114. Wei Y, Zhou T, Pan R, et al. Exosomes containing miR-1469 regulate natural killer cells by targeting CD122 in non-segmental vitiligo. J Dermatological Sci. 2024;113(2):42–50. doi:10.1016/j.jdermsci.2023.12.006
115. Al-Masawa ME, Alshawsh MA, Ng CY, et al. Efficacy and safety of small extracellular vesicle interventions in wound healing and skin regeneration: a systematic review and meta-analysis of animal studies. Theranostics. 2022;12(15):6455–6508. doi:10.7150/thno.73436
116. Ding J-Y, Chen M-J, Wu L-F, et al. Mesenchymal stem cell-derived extracellular vesicles in skin wound healing: roles, opportunities and challenges. Military Med Res. 2023;10(1). doi:10.1186/s40779-023-00472-w
117. Najafabadi AH, Soheilifar MH, Masoudi-Khoram N. Exosomes in skin photoaging: biological functions and therapeutic opportunity. Cell Commun Signaling. 2024;22(1). doi:10.1186/s12964-023-01451-3
118. Meng S, Wei Q, Chen S, et al. MiR‐141‐3p‐functionalized exosomes loaded in dissolvable microneedle arrays for hypertrophic scar treatment. Small. 2023;20(8). doi:10.1002/smll.202305374
119. Deng C, Dong K, Liu Y, et al. Hypoxic mesenchymal stem cell-derived exosomes promote the survival of skin flaps after ischaemia–reperfusion injury via mTOR/ULK1/FUNDC1 pathways. J Nanobiotechnol. 2023;21(1). doi:10.1186/s12951-023-02098-5
120. Hu S, Li Z, Lutz H, et al. Dermal exosomes containing miR-218-5p promote hair regeneration by regulating β-catenin signaling. Sci Adv. 2020;6(30):eaba1685. doi:10.1126/sciadv.aba1685
121. Rani S, Ritter T. The exosome ‐ A naturally secreted nanoparticle and its application to wound healing. Adv Mater. 2015;28(27):5542–5552. doi:10.1002/adma.201504009
122. Martin P, Nunan R. Cellular and molecular mechanisms of repair in acute and chronic wound healing. Br J Dermatol. 2015;173(2):370–378. doi:10.1111/bjd.13954
123. Peña OA, Martin P. Cellular and molecular mechanisms of skin wound healing. Nat Rev Mol Cell Biol. 2024;25(8):599–616. doi:10.1038/s41580-024-00715-1
124. Hade MD, Suire CN, Mossell J, et al. Extracellular vesicles: emerging frontiers in wound healing. Med Res Rev. 2022;42(6):2102–2125. doi:10.1002/med.21918
125. Wang Y, Zhang Y, Li T, et al. Adipose mesenchymal stem cell derived exosomes promote keratinocytes and fibroblasts embedded in collagen/platelet‐rich plasma scaffold and accelerate wound healing. Adv Mater. 2023;35(40). doi:10.1002/adma.202303642
126. Liu Y, Zhang M, Wang C, et al. Human umbilical cord mesenchymal stromal cell-derived extracellular vesicles induce fetal wound healing features revealed by single-cell RNA sequencing. ACS Nano. 2024;18(21):13696–13713. doi:10.1021/acsnano.4c01401
127. Liu K, Chen C, Zhang H, et al. Adipose stem cell‐derived exosomes in combination with hyaluronic acid accelerate wound healing through enhancing re‐epithelialization and vascularization. Br J Dermatol. 2019;181(4):854–856. doi:10.1111/bjd.17984
128. Noh CH, Park S, Seong H-R, et al. An exosome-rich conditioned medium from human amniotic membrane stem cells facilitates wound healing via increased reepithelization, collagen synthesis, and angiogenesis. Cells. 2023;12(23). doi:10.3390/cells12232698
129. Zhang Y, Pan Y, Liu Y, et al. Exosomes derived from human umbilical cord blood mesenchymal stem cells stimulate regenerative wound healing via transforming growth factor-β receptor inhibition. Stem Cell Res Ther. 2021;12(1). doi:10.1186/s13287-021-02517-0
130. Lu M, Peng L, Ming X, et al. Enhanced wound healing promotion by immune response-free monkey autologous iPSCs and exosomes vs. their allogeneic counterparts. EBioMedicine. 2019;42:443–457. doi:10.1016/j.ebiom.2019.03.011
131. Yu H, Wu Y, Zhang B, et al. Exosomes derived from E2F1–/– adipose-derived stem cells promote skin wound healing via miR-130b-5p/TGFBR3 axis. Int J Nanomed. 2023;18:6275–6292. doi:10.2147/ijn.S431725
132. Kim S, Lee S, Kim H, et al. Exosomes secreted from induced pluripotent stem cell-derived mesenchymal stem cells accelerate skin cell proliferation. Int J Mol Sci. 2018;19(10). doi:10.3390/ijms19103119
133. Liu Y, Zhang M, Liao Y, et al. Human umbilical cord mesenchymal stem cell-derived exosomes promote murine skin wound healing by neutrophil and macrophage modulations revealed by single-cell RNA sequencing. Front Immunol. 2023:14. doi:10.3389/fimmu.2023.1142088
134. Qiu X, Liu J, Zheng C, et al. Exosomes released from educated mesenchymal stem cells accelerate cutaneous wound healing via promoting angiogenesis. Cell Proliferation. 2020;53(8). doi:10.1111/cpr.12830
135. Qiang L, Yang S, Cui Y-H, et al. Keratinocyte autophagy enables the activation of keratinocytes and fibroblastsand facilitates wound healing. Autophagy. 2020;17(9):2128–2143. doi:10.1080/15548627.2020.1816342
136. Xie Y, Yu L, Cheng Z, et al. SHED-derived exosomes promote LPS-induced wound healing with less itching by stimulating macrophage autophagy. J Nanobiotechnol. 2022;20(1). doi:10.1186/s12951-022-01446-1
137. Zhou X, Brown BA, Siegel AP, et al. Exosome-mediated crosstalk between keratinocytes and macrophages in cutaneous wound healing. ACS Nano. 2020;14(10):12732–12748. doi:10.1021/acsnano.0c03064
138. Jabłońska M, Sawicki T, Żulewska J, et al. The role of bovine milk-derived exosomes in human health and disease. Molecules. 2024;29(24). doi:10.3390/molecules29245835
139. Kim H, Kim DE, Han G, et al. Harnessing the natural healing power of colostrum: bovine milk‐derived extracellular vesicles from colostrum facilitating the transition from inflammation to tissue regeneration for accelerating cutaneous wound healing. Adv Healthcare Mater. 2021;11(6). doi:10.1002/adhm.202102027
140. Jin H, Wen X, Sun R, et al. Engineered nanovesicles from activated neutrophils with enriched bactericidal proteins have molecular debridement ability and promote infectious wound healing. Burns Trauma. 2024:12. doi:10.1093/burnst/tkae018
141. Hu Y, Rao -S-S, Wang Z-X, et al. Exosomes from human umbilical cord blood accelerate cutaneous wound healing through miR-21-3p-mediated promotion of angiogenesis and fibroblast function. Theranostics. 2018;8(1):169–184. doi:10.7150/thno.21234
142. Mi B, Chen L, Xiong Y, et al. Saliva exosomes-derived UBE2O mRNA promotes angiogenesis in cutaneous wounds by targeting SMAD6. J Nanobiotechnol. 2020;18(1). doi:10.1186/s12951-020-00624-3
143. Xia W, Li M, Jiang X, et al. Young fibroblast-derived exosomal microRNA-125b transfers beneficial effects on aged cutaneous wound healing. J Nanobiotechnol. 2022;20(1). doi:10.1186/s12951-022-01348-2
144. Wu H, Yao Z, Li H, et al. Improving dermal fibroblast-to-epidermis communications and aging wound repair through extracellular vesicle-mediated delivery of Gstm2 mRNA. J Nanobiotechnol. 2024;22(1). doi:10.1186/s12951-024-02541-1
145. Bay L, Barnes CJ, Fritz BG, et al. Universal dermal microbiome in human skin. mBio. 2020;11(1). doi:10.1128/mBio.02945-19
146. Yin H, Chen C-Y, Liu Y-W, et al. Synechococcus elongatus PCC7942 secretes extracellular vesicles to accelerate cutaneous wound healing by promoting angiogenesis. Theranostics. 2019;9(9):2678–2693. doi:10.7150/thno.31884
147. Wang J, Li X, Zhao X, et al. Lactobacillus rhamnosus GG-derived extracellular vesicles promote wound healing via miR-21-5p-mediated re-epithelization and angiogenesis. J Nanobiotechnol. 2024;22(1). doi:10.1186/s12951-024-02893-8
148. Feng H, Yue Y, Zhang Y, et al. Plant-derived exosome-like nanoparticles: emerging nanosystems for enhanced tissue engineering. Int J Nanomed. 2024;19:1189–1204. doi:10.2147/ijn.S448905
149. Yang S, Lu S, Ren L, et al. Ginseng-derived nanoparticles induce skin cell proliferation and promote wound healing. J Ginseng Res. 2023;47(1):133–143. doi:10.1016/j.jgr.2022.07.005
150. Ramírez O, Pomareda F, Olivares B, et al. Aloe vera peel-derived nanovesicles display anti-inflammatory properties and prevent myofibroblast differentiation. Phytomedicine. 2024:122. doi:10.1016/j.phymed.2023.155108
151. Şahin F, Koçak P, Güneş MY, et al. In vitro wound healing activity of wheat-derived nanovesicles. Appl Biochem Biotechnol. 2018;188(2):381–394. doi:10.1007/s12010-018-2913-1
152. Tu J, Jiang F, Fang J, et al. Anticipation and verification of dendrobium-derived nanovesicles for skin wound healing targets, predicated upon immune infiltration and senescence. Int J Nanomed. 2024;19:1629–1644. doi:10.2147/ijn.S438398
153. Yang C, Xue Y, Duan Y, et al. Extracellular vesicles and their engineering strategies, delivery systems, and biomedical applications. J Control Release. 2024;365:1089–1123. doi:10.1016/j.jconrel.2023.11.057
154. Li Z, Liu J, Song J, et al. Multifunctional hydrogel-based engineered extracellular vesicles delivery for complicated wound healing. Theranostics. 2024;14(11):4198–4217. doi:10.7150/thno.97317
155. Camões SP, Bulut O, Yazar V, et al. 3D-MSCs A151 ODN-loaded exosomes are immunomodulatory and reveal a proteomic cargo that sustains wound resolution. J Adv Res. 2022;41:113–128. doi:10.1016/j.jare.2022.01.013
156. Su L-Y, Yao M, Xu W, et al. Cascade encapsulation of antimicrobial peptides, exosomes and antibiotics in fibrin-gel for first-aid hemostasis and infected wound healing. Int J Biol Macromol. 2024:269. doi:10.1016/j.ijbiomac.2024.132140
157. Su D, Tsai HI, Xu Z, et al. Exosomal PD‐L1 functions as an immunosuppressant to promote wound healing. J Extracell Vesicles. 2019;9(1). doi:10.1080/20013078.2019.1709262
158. Kwak G, Cheng J, Kim H, et al. Sustained exosome‐guided macrophage polarization using hydrolytically degradable PEG hydrogels for cutaneous wound healing: identification of key proteins and MiRNAs, and sustained release formulation. Small. 2022;18(15). doi:10.1002/smll.202200060
159. Shi A, Li J, Qiu X, et al. TGF-β loaded exosome enhances ischemic wound healing in vitro and in vivo. Theranostics. 2021;11(13):6616–6631. doi:10.7150/thno.57701
160. Li X, Wang Y, Shi L, et al. Magnetic targeting enhances the cutaneous wound healing effects of human mesenchymal stem cell-derived iron oxide exosomes. J Nanobiotechnol. 2020;18(1). doi:10.1186/s12951-020-00670-x
161. Teng X, Liu T, Zhao G, et al. A novel exosome-based multifunctional nanocomposite platform driven by photothermal-controlled release system for repair of skin injury. J Control Release. 2024;371:258–272. doi:10.1016/j.jconrel.2024.05.049
162. Yang P, Ju Y, Liu X, et al. Natural self-healing injectable hydrogels loaded with exosomes and berberine for infected wound healing. Mater Today Bio. 2023. doi:10.1016/j.mtbio.2023.100875
163. Wang T, Xue Y, Zhang W, et al. Collagen sponge scaffolds loaded with trichostatin A pretreated BMSCs-derived exosomes regulate macrophage polarization to promote skin wound healing. Int J Biol Macromol. 2024:269. doi:10.1016/j.ijbiomac.2024.131948
164. Wu Q, Guo Y, Li H, et al. Recombinant human collagen I/carboxymethyl chitosan hydrogel loaded with long-term released hUCMSCs derived exosomes promotes skin wound repair. Int J Biol Macromol. 2024;265. doi:10.1016/j.ijbiomac.2024.130843
165. Li L, Wang Y, Xu Y, et al. ROS-scavenging lipid-based liquid crystalline as a favorable stem cell extracellular vesicles delivery vector to promote wound healing. J Control Release. 2024;371:298–312. doi:10.1016/j.jconrel.2024.05.048
166. Zhou Y, Zhang X-L, Lu S-T, et al. Human adipose-derived mesenchymal stem cells-derived exosomes encapsulated in pluronic F127 hydrogel promote wound healing and regeneration. Stem Cell Res Ther. 2022;13(1). doi:10.1186/s13287-022-02980-3
167. Xi L, Wang L, Zhang M, et al. TNF-R1 cellular nanovesicles loaded on the thermosensitive F-127 hydrogel enhance the repair of scalded skin. ACS Biomater Sci Eng. 2023;9(10):5843–5854. doi:10.1021/acsbiomaterials.2c01257
168. Li Q, Hu W, Huang Q, et al. MiR146a-loaded engineered exosomes released from silk fibroin patch promote diabetic wound healing by targeting IRAK1. Signal Transd Targeted Ther. 2023;8(1). doi:10.1038/s41392-022-01263-w
169. Yu Y, Jin H, Li L, et al. An injectable, activated neutrophil-derived exosome mimetics/extracellular matrix hybrid hydrogel with antibacterial activity and wound healing promotion effect for diabetic wound therapy. J Nanobiotechnol. 2023;21(1). doi:10.1186/s12951-023-02073-0
170. Shiekh PA, Singh A, Kumar A. Exosome laden oxygen releasing antioxidant and antibacterial cryogel wound dressing OxOBand alleviate diabetic and infectious wound healing. Biomaterials. 2020;249. doi:10.1016/j.biomaterials.2020.120020
171. Pomatto M, Gai C, Negro F, et al. Differential therapeutic effect of extracellular vesicles derived by bone marrow and adipose mesenchymal stem cells on wound healing of diabetic ulcers and correlation to their cargoes. Int J Mol Sci. 2021;22(8). doi:10.3390/ijms22083851
172. Yu M, Liu W, Li J, et al. Exosomes derived from atorvastatin-pretreated MSC accelerate diabetic wound repair by enhancing angiogenesis via AKT/eNOS pathway. Stem Cell Res Ther. 2020;11(1). doi:10.1186/s13287-020-01824-2
173. Wang J, Wu H, Peng Y, et al. Hypoxia adipose stem cell-derived exosomes promote high-quality healing of diabetic wound involves activation of PI3K/Akt pathways. J Nanobiotechnol. 2021;19(1). doi:10.1186/s12951-021-00942-0
174. Hu Y, Tao R, Chen L, et al. Exosomes derived from pioglitazone-pretreated MSCs accelerate diabetic wound healing through enhancing angiogenesis. J Nanobiotechnol. 2021;19(1). doi:10.1186/s12951-021-00894-5
175. Chen C-Y, Rao -S-S, Ren L, et al. Exosomal DMBT1 from human urine-derived stem cells facilitates diabetic wound repair by promoting angiogenesis. Theranostics. 2018;8(6):1607–1623. doi:10.7150/thno.22958
176. Kusumbe AP, Ramasamy SK, Adams RH. Coupling of angiogenesis and osteogenesis by a specific vessel subtype in bone. Nature. 2014;507(7492):323–328. doi:10.1038/nature13145
177. Liu L, Zheng CX, Zhao N, et al. Mesenchymal stem cell aggregation‐released extracellular vesicles induce CD31+EMCN+ vessels in skin regeneration and improve diabetic wound healing. Adv Healthcare Mater. 2023;12(20). doi:10.1002/adhm.202300019
178. Wang M, Yang D, Li L, et al. A dual role of mesenchymal stem cell derived small extracellular vesicles on TRPC6 protein and mitochondria to promote diabetic wound healing. ACS Nano. 2024;18(6):4871–4885. doi:10.1021/acsnano.3c09814
179. Condello M, Pellegrini E, Caraglia M, et al. Targeting autophagy to overcome human diseases. Int J Mol Sci. 2019;20(3). doi:10.3390/ijms20030725
180. Ren H, Su P, Zhao F, et al. Adipose mesenchymal stem cell-derived exosomes promote skin wound healing in diabetic mice by regulating epidermal autophagy. Burns Trauma. 2024:12. doi:10.1093/burnst/tkae001
181. Levy D, Abadchi SN, Shababi N, et al. Induced pluripotent stem cell‐derived extracellular vesicles promote wound repair in a diabetic mouse model via an anti‐inflammatory immunomodulatory mechanism. Adv Healthcare Mater. 2023;12(26). doi:10.1002/adhm.202300879
182. Chen T, Song P, He M, et al. Sphingosine-1-phosphate derived from PRP-exos promotes angiogenesis in diabetic wound healing via the S1PR1/AKT/FN1 signaling pathway. Burns Trauma. 2023:11. doi:10.1093/burnst/tkad003
183. Qu KS, Ru Y, Yang D, et al. Fu-Huang ointment ameliorates impaired wound healing associated with diabetes through PI3K-AKT signaling pathway activation. Comput Biol Med. 2023;155. doi:10.1016/j.compbiomed.2023.106660
184. Zhu L, Zang J, Liu B, et al. Oxidative stress‐induced RAC autophagy can improve the HUVEC functions by releasing exosomes. J Cell Physiol. 2020;235(10):7392–7409. doi:10.1002/jcp.29641
185. Li D, Zhang C, Gao Z, et al. Curcumin-loaded macrophage-derived exosomes effectively improve wound healing. Mol Pharmaceut. 2023;20(9):4453–4467. doi:10.1021/acs.molpharmaceut.3c00062
186. Xia W, Liu Y, Jiang X, et al. Lean adipose tissue macrophage derived exosome confers immunoregulation to improve wound healing in diabetes. J Nanobiotechnol. 2023;21(1). doi:10.1186/s12951-023-01869-4
187. Tan M, Liu Y, Xu Y, et al. Plant-derived exosomes as novel nanotherapeutics contrive glycolysis reprogramming-mediated angiogenesis for diabetic ulcer healing. Biomater Res. 2024:28. doi:10.34133/bmr.0035
188. Fan L, Jia X, Dong F, et al. Ginseng-derived nanoparticles accelerate diabetic wound healing by modulating macrophage polarization and restoring endothelial cell function. Mater Today Bio. 2025. doi:10.1016/j.mtbio.2025.102143
189. Zhang X, Gan J, Fan L, et al. Bioinspired adaptable indwelling microneedles for treatment of diabetic ulcers. Adv Mater. 2023;35(23). doi:10.1002/adma.202210903
190. Shang S, Zhuang K, Chen J, et al. A bioactive composite hydrogel dressing that promotes healing of both acute and chronic diabetic skin wounds. Bioact Mater. 2024;34:298–310. doi:10.1016/j.bioactmat.2023.12.026
191. Li W, Wu S, Ren L, et al. Development of an antiswelling hydrogel system incorporating M2-exosomes and photothermal effect for diabetic wound healing. ACS Nano. 2023;17(21):22106–22120. doi:10.1021/acsnano.3c09220
192. Huang Q, Chu Z, Wang Z, et al. circCDK13-loaded small extracellular vesicles accelerate healing in preclinical diabetic wound models. Nat Commun. 2024;15(1). doi:10.1038/s41467-024-48284-3
193. Ma W, Zhang X, Liu Y, et al. Polydopamine decorated microneedles with Fe‐MSC‐derived nanovesicles encapsulation for wound healing. Adv Sci. 2022;9(13). doi:10.1002/advs.202103317
194. Song Y, You Y, Xu X, et al. Adipose‐derived mesenchymal stem cell‐derived exosomes biopotentiated extracellular matrix hydrogels accelerate diabetic wound healing and skin regeneration. Adv Sci. 2023;10(30). doi:10.1002/advs.202304023
195. Hao P-C, Burnouf T, Chiang C-W, et al. Enhanced diabetic wound healing using platelet-derived extracellular vesicles and reduced graphene oxide in polymer-coordinated hydrogels. J Nanobiotechnol. 2023;21(1). doi:10.1186/s12951-023-02068-x
196. Zhu W, Dong Y, Xu P, et al. A composite hydrogel containing resveratrol-laden nanoparticles and platelet-derived extracellular vesicles promotes wound healing in diabetic mice. Acta Biomater. 2022;154:212–230. doi:10.1016/j.actbio.2022.10.038
197. Li X, Qu S, Ouyang Q, et al. A multifunctional composite nanoparticle with antibacterial activities, anti-inflammatory, and angiogenesis for diabetic wound healing. Int J Biol Macromol. 2024;260. doi:10.1016/j.ijbiomac.2024.129531
198. Wang C, Wang M, Xu T, et al. Engineering bioactive self-healing antibacterial exosomes hydrogel for promoting chronic diabetic wound healing and complete skin regeneration. Theranostics. 2019;9(1):65–76. doi:10.7150/thno.29766
199. Xu N, Wang L, Guan J, et al. Wound healing effects of a curcuma zedoaria polysaccharide with platelet-rich plasma exosomes assembled on chitosan/silk hydrogel sponge in a diabetic rat model. Int J Biol Macromol. 2018;117:102–107. doi:10.1016/j.ijbiomac.2018.05.066
200. Yang J, Chen Z, Pan D, et al. Umbilical cord-derived mesenchymal stem cell-derived exosomes combined pluronic F127 hydrogel promote chronic diabetic wound healing and complete skin regeneration. Int J Nanomed. 2020;15:5911–5926. doi:10.2147/ijn.S249129
201. Xiang K, Chen J, Guo J, et al. Multifunctional ADM hydrogel containing endothelial cell-exosomes for diabetic wound healing. Mater Today Bio. 2023. doi:10.1016/j.mtbio.2023.100863
202. Wang Y, Cao Z, Wei Q, et al. VH298-loaded extracellular vesicles released from gelatin methacryloyl hydrogel facilitate diabetic wound healing by HIF-1α-mediated enhancement of angiogenesis. Acta Biomater. 2022;147:342–355. doi:10.1016/j.actbio.2022.05.018
203. Yuan M, Liu K, Jiang T, et al. GelMA/PEGDA microneedles patch loaded with HUVECs-derived exosomes and Tazarotene promote diabetic wound healing. J Nanobiotechnol. 2022;20(1). doi:10.1186/s12951-022-01354-4
204. Zhao X, Fu L, Zou H, et al. Optogenetic engineered umbilical cord MSC-derived exosomes for remodeling of the immune microenvironment in diabetic wounds and the promotion of tissue repair. J Nanobiotechnol. 2023;21(1). doi:10.1186/s12951-023-01886-3
205. Shi Y, Wang S, Wang K, et al. Relieving macrophage dysfunction by inhibiting SREBP2 activity: a hypoxic mesenchymal stem cells‐derived exosomes loaded multifunctional hydrogel for accelerated diabetic wound healing. Small. 2024;20(25). doi:10.1002/smll.202309276
206. Lin X, Lai Y. Scarring skin: mechanisms and therapies. Int J Mol Sci. 2024;25(3). doi:10.3390/ijms25031458
207. Coentro JQ, Pugliese E, Hanley G, et al. Current and upcoming therapies to modulate skin scarring and fibrosis. Adv Drug Delivery Rev. 2019;146:37–59. doi:10.1016/j.addr.2018.08.009
208. Xu C, Zhang H, Yang C, et al. miR-125b-5p delivered by adipose-derived stem cell exosomes alleviates hypertrophic scarring by suppressing Smad2. Burns Trauma. 2024:12. doi:10.1093/burnst/tkad064
209. Li Y, Zhang J, Shi J, et al. Exosomes derived from human adipose mesenchymal stem cells attenuate hypertrophic scar fibrosis by miR-192-5p/IL-17RA/Smad axis. Stem Cell Res Ther. 2021;12(1). doi:10.1186/s13287-021-02290-0
210. Zhang H, Zang C, Zhao W, et al. Exosome derived from mesenchymal stem cells alleviates hypertrophic scar by inhibiting the fibroblasts via TNFSF-13/HSPG2 signaling pathway. Int J Nanomed. 2023;18:7047–7063. doi:10.2147/ijn.S433510
211. Han F, Wang K, Shen K, et al. Extracellular vesicles from lactobacillus druckerii inhibit hypertrophic scar fibrosis. J Nanobiotechnol. 2023;21(1). doi:10.1186/s12951-023-01861-y
212. Franco AC, Aveleira C, Cavadas C. Skin senescence: mechanisms and impact on whole-body aging. Trends Mol Med. 2022;28(2):97–109. doi:10.1016/j.molmed.2021.12.003
213. Farage MA, Miller KW, Elsner P, Maibach HI. Intrinsic and extrinsic factors in skin ageing: a review. Int J Cosmet Sci. 2008;30(2):87–95. 10.1111/j.1468-2494.2007.00415.x.
214. Shin J-W, Kwon S-H, Choi J-Y, et al. Molecular mechanisms of dermal aging and antiaging approaches. Int J Mol Sci. 2019;20(9). doi:10.3390/ijms20092126
215. Low E, Alimohammadiha G, Smith LA, et al. How good is the evidence that cellular senescence causes skin ageing? Ageing Res Rev. 2021. doi:10.1016/j.arr.2021.101456
216. Xiao X, Xu M, Yu H, et al. Mesenchymal stem cell-derived small extracellular vesicles mitigate oxidative stress-induced senescence in endothelial cells via regulation of miR-146a/Src. Signal Transd Targeted Ther. 2021;6(1). doi:10.1038/s41392-021-00765-3
217. Hu S, Li Z, Cores J, et al. Needle-free injection of exosomes derived from human dermal fibroblast spheroids ameliorates skin photoaging. ACS Nano. 2019;13(10):11273–11282. doi:10.1021/acsnano.9b04384
218. Han J, Wu T, Jin J, et al. Exosome-like nanovesicles derived from Phellinus linteus inhibit Mical2 expression through cross-kingdom regulation and inhibit ultraviolet-induced skin aging. J Nanobiotechnol. 2022;20(1). doi:10.1186/s12951-022-01657-6
219. Jiang Z, Cheng H, Qian X, et al. The role and mechanism of engineered nanovesicles derived from hair follicle mesenchymal stem cells in the treatment of UVB‐induced skin photoaging. J Cosmet Dermatol. 2024;23(9):3005–3020. doi:10.1111/jocd.16336
220. Lu L, Bai W, Wang M, et al. Novel roles of bovine milk‐derived exosomes in skin antiaging. J Cosmet Dermatol. 2023;23(4):1374–1385. doi:10.1111/jocd.16112
221. Wang Z, Yuan J, Xu Y, et al. Olea europaea leaf exosome-like nanovesicles encapsulated in a hyaluronic acid / tannic acid hydrogel dressing with dual “defense-repair” effects for treating skin photoaging. Mater Today Bio. 2024. doi:10.1016/j.mtbio.2024.101103
222. Choi W, Cho JH, Park SH, et al. Ginseng root-derived exosome-like nanoparticles protect skin from UV irradiation and oxidative stress by suppressing activator protein-1 signaling and limiting the generation of reactive oxygen species. J Ginseng Res. 2024;48(2):211–219. doi:10.1016/j.jgr.2024.01.001
223. Trentini M, Zanolla I, Zanotti F, et al. Apple derived exosomes improve collagen Type I production and decrease MMPs during aging of the skin through downregulation of the NF-κB pathway as mode of action. Cells. 2022;11(24). doi:10.3390/cells11243950
224. Lee H, Cha H, Park JH. Derivation of cell-engineered nanovesicles from human induced pluripotent stem cells and their protective effect on the senescence of dermal fibroblasts. Int J Mol Sci. 2020;21(1). doi:10.3390/ijms21010343
225. Zhu H, Guo X, Zhang Y, et al. The combined anti-aging effect of hydrolyzed collagen oligopeptides and exosomes derived from human umbilical cord mesenchymal stem cells on human skin fibroblasts. Molecules. 2024;29(7). doi:10.3390/molecules29071468
226. Hsueh -Y-Y, Wang D-H, Huang T-C, et al. Novel skin chamber for rat ischemic flap studies in regenerative wound repair. Stem Cell Res Ther. 2016;7(1). doi:10.1186/s13287-016-0333-0
227. Niu Q, Yang Y, Li D, et al. Exosomes derived from bone marrow mesenchymal stem cells alleviate ischemia-reperfusion injury and promote survival of skin flaps in rats. Life. 2022;12(10). doi:10.3390/life12101567
228. Ding J-P, Sun Y, Chen B, et al. Sequential transplantation of exosomes and BMSCs pretreated with hypoxia efficiently facilitates perforator skin flap survival area in rats. Br J Oral Maxillofac Surg. 2024;62(4):361–366. doi:10.1016/j.bjoms.2023.12.012
229. Bai Y, Han Y-D, Yan X-L, et al. Adipose mesenchymal stem cell-derived exosomes stimulated by hydrogen peroxide enhanced skin flap recovery in ischemia-reperfusion injury. Biochem Biophys Res Commun. 2018;500(2):310–317. doi:10.1016/j.bbrc.2018.04.065
230. Liu X, Chen H, Lei L, et al. Exosomes-carried curcumin based on polysaccharide hydrogel promote flap survival. Int J Biol Macromol. 2024;270. doi:10.1016/j.ijbiomac.2024.132367
231. Shi X, Yang G, Liu MY, et al. Exosomes derived from human dental pulp stem cells increase flap survival with ischemia-reperfusion injuries. Regener Med. 2023;18(4):313–327. doi:10.2217/rme-2022-0206
232. Zhang X, Jiang X, Deng H, et al. Engineering exosomes from fibroblast growth factor 1 pre-conditioned adipose-derived stem cells promote ischemic skin flaps survival by activating autophagy. Mater Today Bio. 2024;29. doi:10.1016/j.mtbio.2024.101314
233. Luo G, Zhou Z, Cao Z, et al. M2 macrophage-derived exosomes induce angiogenesis and increase skin flap survival through HIF1AN/HIF-1α/VEGFA control. Arch Biochem Biophys. 2024:751. doi:10.1016/j.abb.2023.109822
234. Mubki T, Rudnicka L, Olszewska M, et al. Evaluation and diagnosis of the hair loss patient. J Am Acad Dermatol. 2014;71(3):415.e1–415.e15. doi:10.1016/j.jaad.2014.04.070
235. Devjani S, Ezemma O, Kelley KJ, et al. Androgenetic alopecia: therapy update. Drugs. 2023;83(8):701–715. doi:10.1007/s40265-023-01880-x
236. Tang X, Cao C, Liang Y, et al. Adipose-derived stem cell exosomes antagonize the inhibitory effect of dihydrotestosterone on hair follicle growth by activating Wnt/β-catenin pathway. Stem Cells Int. 2023;2023:1–20. doi:10.1155/2023/5548112
237. Ogawa M, Udono M, Teruya K, et al. Exosomes derived from fisetin-treated keratinocytes mediate hair growth promotion. Nutrients. 2021;13(6). doi:10.3390/nu13062087
238. Kwack MH, Seo CH, Gangadaran P, et al. Exosomes derived from human dermal papilla cells promote hair growth in cultured human hair follicles and augment the hair‐inductive capacity of cultured dermal papilla spheres. Exp Dermatol. 2019;28(7):854–857. doi:10.1111/exd.13927
239. Rajendran RL, Gangadaran P, Kwack MH, et al. Engineered extracellular vesicle mimetics from macrophage promotes hair growth in mice and promotes human hair follicle growth. Exp Cell Res. 2021;409(1). doi:10.1016/j.yexcr.2021.112887
240. Chen Y, Huang J, Chen R, et al. Sustained release of dermal papilla-derived extracellular vesicles from injectable microgel promotes hair growth. Theranostics. 2020;10(3):1454–1478. doi:10.7150/thno.39566
241. Li J, Zhao B, Yao S, et al. Dermal PapillaCell-derived exosomes regulate hair follicle stem cell proliferation via LEF1. Int J Mol Sci. 2023;24(4). doi:10.3390/ijms24043961
242. Shang Y, Li M, Zhang L, et al. Exosomes derived from mouse vibrissa dermal papilla cells promote hair follicle regeneration during wound healing by activating Wnt/β-catenin signaling pathway. J Nanobiotechnol. 2024;22(1). doi:10.1186/s12951-024-02689-w
243. Chen Y, Huang J, Liu Z, et al. miR-140-5p in small extracellular vesicles from human papilla cells stimulates hair growth by promoting proliferation of outer root sheath and hair matrix cells. Front Cell Develop Biol. 2020:8. doi:10.3389/fcell.2020.593638
244. Cao L, Tian T, Huang Y, et al. Neural progenitor cell-derived nanovesicles promote hair follicle growth via miR-100. J Nanobiotechnol. 2021;19(1). doi:10.1186/s12951-020-00757-5
245. Kim H, Jang Y, Kim EH, et al. Potential of colostrum-derived exosomes for promoting hair regeneration through the transition from telogen to anagen phase. Front Cell Develop Biol. 2022. doi:10.3389/fcell.2022.815205
246. Rajendran RL, Gangadaran P, Kwack MH, et al. Engineered nanovesicles from fibroblasts modulate dermal papillae cells in vitro and promote human hair follicle growth ex vivo. Cells. 2022;11(24). doi:10.3390/cells11244066
247. Oh HG, Jung M, Jeong S-Y, et al. Improvement of androgenic alopecia by extracellular vesicles secreted from hyaluronic acid-stimulated induced mesenchymal stem cells. Stem Cell Res Ther. 2024;15(1). doi:10.1186/s13287-024-03906-x
248. Liang Y, Tang X, Zhang X, et al. Adipose mesenchymal stromal cell-derived exosomes carrying MiR-122-5p antagonize the inhibitory effect of dihydrotestosterone on hair follicles by targeting the TGF-β1/SMAD3 signaling pathway. Int J Mol Sci. 2023;24(6). doi:10.3390/ijms24065703
249. Mao Y, Liu P, Wei J, et al. Exosomes derived from umbilical cord mesenchymal stem cell promote hair regrowth in C57BL6 mice through upregulation of the RAS/ERK signaling pathway. J Transl Internal Med. 2024;12(5):478–494. doi:10.1515/jtim-2024-0012
250. Xiong J, Liu Z, Jia L, et al. Bioinspired engineering ADSC nanovesicles thermosensitive hydrogel enhance autophagy of dermal papilla cells for androgenetic alopecia treatment. Bioact Mater. 2024;36:112–125. doi:10.1016/j.bioactmat.2024.02.023
251. Wu X, Huang X, Zhu Q, et al. Hybrid hair follicle stem cell extracellular vesicles co-delivering finasteride and gold nanoparticles for androgenetic alopecia treatment. J Control Release. 2024;373:652–666. doi:10.1016/j.jconrel.2024.07.066
252. Skogberg G, Jackson S, Åstrand A. Mechanisms of tolerance and potential therapeutic interventions in Alopecia Areata. Pharmacol Ther. 2017;179:102–110. doi:10.1016/j.pharmthera.2017.05.008
253. Hu S, Zhang J, Ji Q, et al. Exosomes derived from uMSCs promote hair regrowth in alopecia areata through accelerating human hair follicular keratinocyte proliferation and migration. Cell Biol Int. 2023;48(2):154–161. doi:10.1002/cbin.12099
254. Kalaora S, Nagler A, Wargo JA, et al. Mechanisms of immune activation and regulation: lessons from melanoma. Nat Rev Cancer. 2022;22(4):195–207. doi:10.1038/s41568-022-00442-9
255. Leonardi G, Falzone L, Salemi R, et al. Cutaneous melanoma: from pathogenesis to therapy (Review). Int J Oncol. 2018. doi:10.3892/ijo.2018.4287
256. Vignard V, Labbé M, Marec N, et al. MicroRNAs in tumor exosomes drive immune escape in melanoma. Cancer Immunol Res. 2020;8(2):255–267. doi:10.1158/2326-6066.Cir-19-0522
257. García-Silva S, Benito-Martín A, Sánchez-Redondo S, et al. Use of extracellular vesicles from lymphatic drainage as surrogate markers of melanoma progression and BRAF V600E mutation. J Exp Med. 2019;216(5):1061–1070. doi:10.1084/jem.20181522
258. Ekström EJ, Bergenfelz C, Von Bülow V, et al. WNT5A induces release of exosomes containing pro-angiogenic and immunosuppressive factors from malignant melanoma cells. Mol Cancer. 2014;13:88. doi:10.1186/1476-4598-13-88
259. Liu N, Zhang J, Yin M, et al. Inhibition of xCT suppresses the efficacy of anti-PD-1/L1 melanoma treatment through exosomal PD-L1-induced macrophage M2 polarization. Mol Ther. 2021;29(7):2321–2334. doi:10.1016/j.ymthe.2021.03.013
260. Zhu L, Kalimuthu S, Gangadaran P, et al. Exosomes derived from natural killer cells exert therapeutic effect in melanoma. Theranostics. 2017;7(10):2732–2745. doi:10.7150/thno.18752
261. Liu M, Wang H, Liu Z, et al. Exosomes from adipose-derived stem cells inhibits skin cancer progression via miR-199a-5p/SOX4. Biotechnol Genet Eng Rev. 2023:1–13. doi:10.1080/02648725.2023.2204702
262. Stanly C, Alfieri M, Ambrosone A, et al. Grapefruit-derived micro and nanovesicles show distinct metabolome profiles and anticancer activities in the A375 human melanoma cell line. Cells. 2020;9(12). doi:10.3390/cells9122722
263. Zeng L, Wang H, Shi W, et al. Aloe derived nanovesicle as a functional carrier for indocyanine green encapsulation and phototherapy. J Nanobiotechnol. 2021;19(1). doi:10.1186/s12951-021-01195-7
264. Gu Y, Du Y, Jiang L, et al. αvβ3 integrin-specific exosomes engineered with cyclopeptide for targeted delivery of triptolide against malignant melanoma. J Nanobiotechnol. 2022;20(1). doi:10.1186/s12951-022-01597-1
265. Kostyusheva A, Romano E, Yan N, et al. Breaking barriers in targeted therapy: advancing exosome isolation, engineering, and imaging. Adv Drug Delivery Rev. 2025;218. doi:10.1016/j.addr.2025.115522
266. Charoenviriyakul C, Takahashi Y, Nishikawa M, et al. Preservation of exosomes at room temperature using lyophilization. Int J Pharm. 2018;553(1–2):1–7. doi:10.1016/j.ijpharm.2018.10.032
267. Suharta S, Barlian A, Hidajah AC, et al. Plant‐derived exosome‐like nanoparticles: a concise review on its extraction methods, content, bioactivities, and potential as functional food ingredient. Journal of Food Science. 2021;86(7):2838–2850. doi:10.1111/1750-3841.15787
268. Tan F, Li X, Wang Z, et al. Clinical applications of stem cell-derived exosomes. Signal Transd Targeted Ther. 2024;9(1). doi:10.1038/s41392-023-01704-0
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.