Back to Journals » Journal of Blood Medicine » Volume 10
The immunomodulatory-drug, lenalidomide, sustains and enhances interferon-α production by human plasmacytoid dendritic cells
Authors Kibata K, Ito T ![]() , Inaba M, Tanaka A
, Inaba M, Tanaka A ![]() , Iwata R, Inagaki-Katashiba N, Phan V, Satake A
, Iwata R, Inagaki-Katashiba N, Phan V, Satake A ![]() , Nomura S
, Nomura S ![]()
Received 22 February 2019
Accepted for publication 31 May 2019
Published 12 July 2019 Volume 2019:10 Pages 217—226
DOI https://doi.org/10.2147/JBM.S206459
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Martin Bluth
Kayoko Kibata,1 Tomoki Ito,1 Muneo Inaba,1 Akihiro Tanaka,1 Ryoichi Iwata,2 Noriko Inagaki-Katashiba,1 Vien Phan,1 Atsushi Satake,1 Shosaku Nomura1
1Kansai Medical University, First Department of Internal Medicine, Osaka, Japan; 2Kansai Medical University, Department of Neurosurgery, Osaka, Japan
Background: Lenalidomide (LEN), an immunomodulatory drug (IMiD), is currently used for treatment of multiple myeloma (MM). LEN potentiates T cell and natural killer cell functions. However, the cellular and molecular mechanisms underlying the immunomodulatory effects of LEN remain unclear. We focused on the effects of LEN on human plasmacytoid dendritic cells (pDCs), which are the major source of interferon (IFN)-α in the blood and play a central role in innate immune responses.
Results: We found that bortezomib, a proteasome inhibitor used to treat MM, killed pDCs but that 0.1–3 μM LEN (covering clinical plasma concentration range) did not affect pDC survival or CD86 expression. Bortezomib inhibited pDC-derived IFN-α production in a dose-dependent fashion, but 0.1–3 μM LEN sustained pDC-derived IFN-α production when stimulated with an optimal concentration of CpG-ODN 2216 (3 μM). In pDCs stimulated with a low concentration of CpG-ODN (0.1 μM), LEN enhanced IFN-α production. These results indicated that LEN, when used at a clinically relevant concentration, can potentially enhance IFN-α production by pDCs.
Conclusion: Collectively, our findings unveiled a novel target of LEN and extend the repertoire of the drug’s known immunomodulatory effects. These effects may explain the low incidence of herpes zoster viral infection observed during LEN treatment compared with bortezomib treatment. LEN may function as an IMiD affecting a wide array of immune cells, including pDCs, leading to amplification of a positive immune axis able to eliminate MM cells.
Keywords: lenalidomide, IMiDs, plasmacyotid DCs, type I IFNs, multiple myeloma
Introduction
Lenalidomide (LEN) and pomalidomide (POM) are classified as an immunomodulatory drugs (IMiDs) with both direct tumoricidal and immunomodulatory effects. Both drugs are used in the treatment of multiple myeloma (MM). LEN is an integral backbone drug for MM. At present, the standard of care for patients with newly diagnosed or relapsed/refractory MM is to administer LEN combined with proteasome inhibitors or antibody-drugs. Continuous treatment with LEN until diseases progression confers a survival benefit for MM patients.1
Recent studies have elucidated the diversity of immune cells targeted by LEN. LEN promotes the proliferation of some immune effector cells in vivo. The total percentage of proliferating S-phase CD4+ T cells, CD8+ T cells, and natural killer (NK) cells progressively increased after administration of LEN in high-risk smoldering MM patients.2 In addition to this quantitative enhancement, several studies have shown that LEN induces qualitative activation of several immune cells. For instance, LEN augments NK cell cytotoxicity and cytotoxic lymphocyte (CTL) activity,3–8 and inhibits the proliferation and function of Treg cells.9,10
PD-L1 and PD-1 expression are increased in MM cells and immune effector cells (such as CTLs and NK cells), respectively, in MM patients.11,12 In addition to the enhancing effect of LEN on immune effectors, LEN can downregulate PD-L1 on primary MM cells and PD-1 on NK cells and CTL cells of MM patients, leading to enhanced immune responses induced by immune checkpoint blockade.11,12 Thus, LEN has a potent and complex immunostimulatory effect, facilitating the attack of MM cells by activated immune effectors. This function of LEN is relevant to the treatment of MM patients under immune dysfunction. However, the cellular and molecular mechanisms underlying the immunomodulatory effects of LEN remain unclear.
A series of analyses have clarified the functional plasticity of dendritic cells (DCs) in inducing T-helper (Th)1 or Th2 responses. DCs, as cellular commanders of the immune cascade, are pivotal in orchestrating both innate and acquired immune responses. Though they make up only a small fraction of peripheral blood mononuclear cells (PBMCs), plasmacytoid DCs (pDCs) represent a major source of type I interferons (IFNs) amongst PBMCs and lymphoid tissues in both humans and mice.13,14 Human pDCs produce less IL-12 but dedicate a large proportion of their transcriptional machinery to producing type I IFNs. pDCs play a central role in the innate antiviral immune response through their ability to robustly produce type I IFNs upon viral infection.15 Type I IFNs can enhance immune effector cells such as T cells, B cells, monocytes, and NK cells16–18 pDCs respond to viral infection through their selective expression of Toll-like receptor (TLR)7 and TLR9, which sense viral RNA and DNA, respectively, within early endosomes.19 Although there is evidence demonstrating the immunomodulatory effects of LEN on mouse conventional DCs,8,20 only a few reports have examined its effects on human DC subsets. We studied the effect of LEN on the type I IFN-producing capacity of human pDCs to elucidate the immunopotentiating activity of LEN in regards to the total immune system. We show that LEN has a novel function in sustaining and enhancing the IFN-α production by human pDCs without affecting their survival. These data suggest that LEN functions as a positive immunomodulator, modulating the initial phases of the immune cascade.
Materials and methods
Media and reagents
RPMI-1640 supplemented with 2 mM L-glutamine, 100 U/mL penicillin, 100 ng/mL streptomycin and 10% heat-inactivated fetal bovine serum (Biosource International) was used for all cell culture experiments. For human cell stimulation, we used 0.1–3 μM CpG-ODN 2216 (Invivogen). LEN (SelleckChem), POM (Sigma), and bortezomib (LC Laboratories) were dissolved in 100% dimethyl sulfoxide (DMSO). Diluted DMSO was used as a vehicle control.
Cell isolation and culture
Human blood DCs were isolated from healthy adult volunteers as previously described.21,22 Briefly, PBMCs from normal healthy donors were isolated by Ficoll Hypaque density gradient centrifugation, and the DC-enriched population (CD4+/CD3−/CD14− cells) was isolated from PBMCs by negative and subsequent positive immunoselections. The CD11c−/lineage−/BDCA4+/CD4+ fraction as pDCs was sorted with a FACS Aria® (BD Biosciences) using phycoerythrin (PE)-labeled anti-CD304 (AD5-17F6: BDCA-4) (Miltenyi Biotec), allophycocyanin (APC)-labeled anti-CD11c (B-ly6: BD Biosciences), a mixture of fluorescein isothiocyanate (FITC)-labeled monoclonal antibodies (mAbs) against lineage markers (CD3 [M2AB: Exalpha], CD14 [M5E2: BD Biosciences], CD15 [M5E2: BD Biosciences], CD16 [J5511: Exalpha], CD19 [HIB19: BD Biosciences], and CD56 [NCAM16.2: BD Biosciences]), and PE-Cy5.5-labeled anti-CD4 (RPA-T4: BD Biosciences). Sorted cells reached >99% purity, which was confirmed by anti-BDCA2 staining following sorting.
CD4+ T cells were isolated using a human CD4+ T cell isolation kit (Miltenyi Biotec) according to the manufacturer’s instructions. The purified cells reached >94% purity, which was confirmed using PE-labeled anti-CD8 (17D8: Exalpha), FITC-labeled CD3, and PE-Cy5.5-labeled anti-CD4 Ab staining.
Purified pDCs were seeded at a density of 4×104 cells per 200 μL of medium in flat-bottomed 96-well plates and PBMCs were seeded at a density of or 5×105 cells per 500 µL medium in flat-bottomed 48-well plates for 24 h in the presence of CpG-ODN 2216 and 0.01–30 µM of LEN, 0.3 µM of POM, or 3–1000 nM of bortezomib.
We obtained peripheral blood (PB) from six cases with newly diagnosed MM (not plasma cell leukemia) before and after receiving one cycle of LEN with dexamethasone (LEN-DEX) (oral LEN, 5–25 mg per day on days 1–21 of each 28 day cycle with 20–40 mg oral DEX on days 1, 8, 15, and 22 of each 28 day cycle) between January 2011 and December 2015. Frequencies of pDCs were calculated amongst total PBMCs (at least 2×106 cells) in PB samples using flow cytometry (a minimum of 100,000 events were gated). pDCs were detected as the PE-labeled anti-BDCA-2 (CD303) mAb (AC144: Mitenyi Biotec)-positive population in the lineage (CD3, CD14, CD15, CD16, CD19, and CD56) negative, CD11c-negative, and CD4-positive fraction.
DC–T cell coculture
2×104 freshly isolated CD4+ T cells were stimulated with allogenic pDCs pre-cultured under different conditions for 24 h (DC:T cell ratio, 1:2) in 96-well round-bottom culture plates. After 7 days of culture, cells were collected and resuspended in ethylenediaminetetraacetic (EDTA)-containing medium to dissociate the clusters. Viable cells were counted by trypan blue exclusion.
Analysis of cells
Human pDCs were stained with FITC-labeled CD86 (2331: BD Biosciences) and then analyzed using a FACScalibur® (BD Biosciences). Dead cells were excluded based on side- and forward-scatter characteristics. Cells were washed with phosphate-buffered saline containing 2 mM EDTA, and viable cells were counted in triplicate using trypan blue exclusion. Viable cells were also evaluated as annexin V-negative fractions using an annexin V-FITC Apoptosis Detection kit (Sigma-Aldrich). Production of IFN-α in culture supernatants after 24 h was assessed by ELISA (PBL Biomedical Laboratories).
Statistical analysis
Data were analyzed using the student’s t-test. Values of p<0.05 were considered statistically significant. Data analysis was carried out using GraphPad Prism (GraphPad Software).
Results
Clinical concentrations of LEN do not affect human pDC survival and maturation
Since LEN, as well as bortezomib, have direct tumoricidal effects against MM cells, we first analyzed the viability of pDCs upon activation in the presence of different doses of LEN. Clinical pharmacokinetics show that clinical peak plasma LEN concentrations of MM patients administered 10 or 25 mg of oral LEN are around 1.2 μM (311 ng/mL) and 2.7 μM (714 ng/mL), respectively.23 Analysis of annexin V-staining (Figure 1) showed that 0.1–3 μM of LEN (covering the clinical in vivo plasma concentration range of oral LEN administration) did not affect pDC survival in the presence of CpG-ODN 2216, although pDCs were susceptible to the cytotoxic effects of bortezomib. We confirmed this finding using trypan-blue exclusion of the dead cells (data not shown). We next investigated the effects of LEN on pDC maturation. pDCs treated with 0.1–3 μM of LEN showed no change in CD86 expression in response to CpG-ODN 2216 (Figure 2A). pDCs exposed to 3–300 nM of bortezomib (covering the clinical in vivo plasma concentration range following intravenous bortezomib administration24) showed greatly reduced viability and CD86 expression after exposure to CpG-ODN 2216 (Figures 1 and 2A). Consistent with CD86 expression, CpG-prestimulated pDCs and LEN-conditioned CpG-prestimulated pDCs induced expansion of CD4+ T cells comparably. By contrast, bortezomib-conditioned CpG-prestimulated pDCs showed an impaired ability to expand CD4+ T cells (Figure 2B).
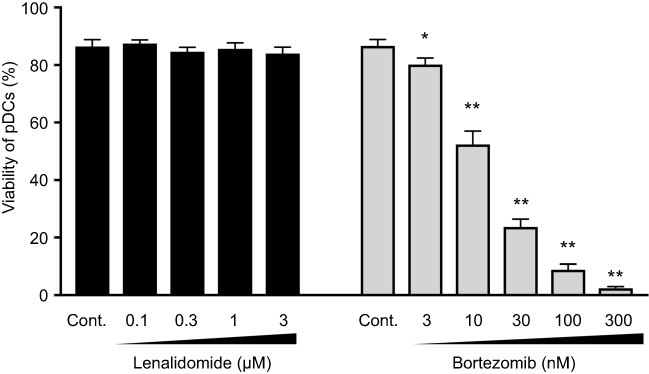 |
Figure 1 Effects of lenalidomide on plasmacytoid dendritic cell survival. Human plasmacytoid dendritic cells (pDCs) were incubated for 24 h with the indicated concentrations of lenalidomide (LEN), bortezomib, or vehicle in the presence of 3 μM CpG-ODN 2216. After 24 h, viable cells were measured by annexin V staining and flow cytometry. Percentages of annexin V-positive cells are indicated. Data are shown as means ± SEMs of six independent donors. Statistical significance was determined using paired Student’s t-tests (*=p<0.05, **=p<0.01; p-values were calculated for each concentration of LEN or bortezomib versus vehicle control). |
 |
Figure 2 Effects of lenalidomide on plasmacytoid dendritic cell maturation. Human plasmacytoid dendritic cells (pDCs) were incubated for 24 h with the indicated concentrations of lenalidomide (LEN), bortezomib (BOR), or vehicle in the presence of 3 μM CpG-ODN 2216. (A) After 24 h, CD86 expression on pDCs was analyzed using flow cytometry. Data were quantified as the mean fluorescence intensity (MFI), which was calculated by subtraction of the MFI for the isotype control from the MFI for cells stained with CD86 mAb. Data are shown as means ± SEMs of six independent donors. (B) CD4+ T cells were stimulated for 7 days with allogeneic pDCs pretreated with CpG, CpG + 1 μM LEN, or CpG + 30 nM bortezomib for 24 h. After culture, cells were quantitated by trypan blue dye-exclusion. Data are shown from four independent sets of experiments. Statistical significance was determined using paired Student’s t-tests (*=p<0.05, **=p<0.01; p-values were calculated for each concentration of LEN or bortezomib versus vehicle control). Abbreviation: N.E., not evaluated. |
Frequency of pDCs was not decreased in MM patients treated with LEN
To confirm that LEN was not cytotoxic against pDCs in the MM patients after treatment, we next investigated the frequency of pDCs in the PB of MM patients before and after LEN administration. pDCs were detected using flow cytometry as BDCA-2-positive population in the lineage (CD3, CD14, CD15, CD16, CD19, and CD56)-negative, CD11c-negative, and CD4-positive fraction (Figure 3A). We examined PB samples from 10 normal donors and six newly-diagnosed MM patients. No significant difference in the percentage of pDCs was observed between PBMCs from healthy donors (mean: 0.36%, range: 0.18%–0.73%) and PBMCs from MM patients prior to LEN administration (mean: 0.39%, range: 0.13%–0.62%) (Figure 3B). After one cycle of LEN-DEX treatment, the percentage of pDCs was not significantly decreased (mean: 0.38%, range: 0.21%–0.54%) compared with values obtained prior to treatment. This finding suggests that, in a clinical setting, LEN does not influence the viability of blood pDCs after initial treatment.
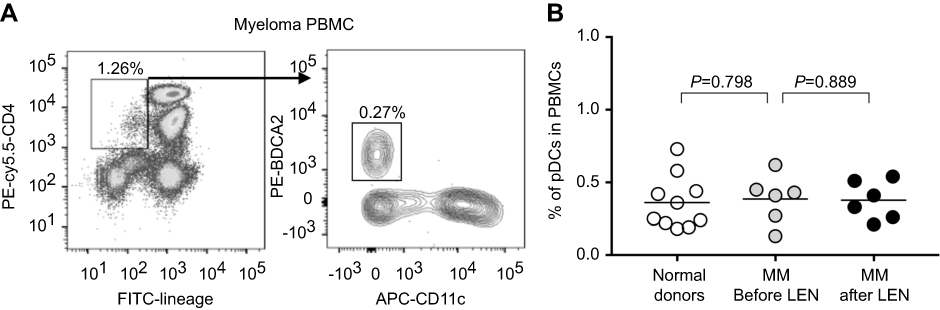 |
Figure 3 Frequency of blood plasmacytoid dendritic cells in multiple myeloma patients before and after treatment with lenalidomide. Frequency of plasmacytoid dendritic cells (pDCs) was calculated amongst total peripheral blood mononuclear cells (PBMCs) (at least 2×106 cells) from peripheral blood using flow cytometry. pDCs were detected as BDCA-2-positive population in the lineage (CD3, CD14, CD15, CD16, CD19, and CD56)-negative, CD11c-negative, and CD4-positive fraction. (A) A representative flow cytometry analysis showing pDCs in PBMCs from six newly-diagnosed multiple myeloma (MM) patients before treatment. Numbers indicate percentages of the gated fraction amongst total PBMCs. (B) Each dot shown is derived from 10 healthy donors and six newly-diagnosed MM patients before and after one cycle of lenalidomide (LEN)-dexamethasone administration. Statistical significance was determined using unpaired Student’s t-tests of healthy donors versus MM patients before LEN treatment, or paired Student’s t-tests of MM patients before LEN treatment versus MM patients after LEN treatment. |
Exposure to clinical concentrations of LEN sustained and potentially enhanced IFN-α production by human pDCs
We next examined the immunomodulatory properties of LEN toward human PBMCs and purified pDCs. We measured IFN-α production by human PBMCs and purified pDCs, which are the major source of type I IFNs,14 in the presence or absence of clinical concentrations of LEN. We found that IFN-α production by PBMCs in response to 3 μM CpG-ODN 2216 was slightly but not significantly enhanced after exposure to a clinical concentration range of LEN (0.01–3 μM) (Figure 4). This slight enhancement was observed even at a lower concentration of LEN (0.01–0.3 μM). We next investigated whether LEN and bortezomib act directly on pDCs. As shown in Figure 5A, bortezomib dose-dependently inhibited pDC-derived IFN-α production, in line with a previous report.25 However, exposure to 0.01–3 μM of LEN resulted in sustained IFN-α production in pDC cultures when stimulated with an optimal concentration of CpG-ODN 2216 (3 μM) (Figure 5B). To address the possibility that LEN enhanced IFN-α production by pDCs that could not be observed because of exhaustion following maximal CpG-ODN 2216 stimulation, we used a suboptimal concentration of CpG-ODN 2216 to stimulate pDCs. Upon stimulation with a suboptimal concentration of CpG-ODN 2216 (0.1 µM), 1 µM of LEN enhanced pDC-derived IFN-α production (185% of vehicle control culture; p=0.023) (Figure 5C). These results indicated that exposure to a clinical concentration of LEN sustained and potentially enhanced, IFN-α production by pDCs in response to weak CpG stimulation. POM is another IMiD26 and the clinical peak plasma concentration following oral administration of 4 mg POM is 0.27 μM (75 ng/mL).27 We found that 0.3 μM POM also did not affect human pDC survival and sustained IFN-α production by pDCs in response to CpG-ODN 2216 (3 μM) (Figure S1). This result suggested that POM may have a similar immunomodulatory effect on pDC function.
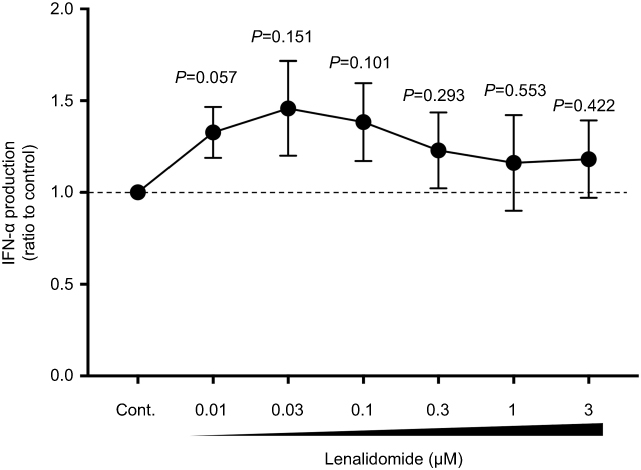 |
Figure 4 Clinical concentrations of lenalidomide (LEN) sustained IFN-α production by peripheral blood mononuclear cells (PBMCs). Human PBMCs were incubated for 24 h with the indicated concentrations of LEN or vehicle in the presence of 3 μM CpG-ODN 2216. After 24 h, the concentrations of IFN-α in the culture supernatants were measured by ELISA. Data are shown as means ± SEMs of seven independent donors. The data were normalized to the value obtained for the vehicle control. The mean (range) of absolute concentrations of CpG-ODN 2216 + vehicle control was 6396 pg/mL (1006–12,938 pg/mL). Statistical significance was determined using paired Student’s t-tests; p-values were calculated for each concentration of LEN versus vehicle control. |
 |
Figure 5 Clinical concentrations of lenalidomide (LEN) sustained and potentially enhanced IFN-α production by human plasmacytoid dendritic cells. (A,B) Purified plasmacytoid dendritic cells (pDCs) were incubated for 24 h with the indicated concentrations of bortezomib, LEN, or vehicle in the presence of 3 µM CpG-ODN 2216. Data are shown as means ± SEMs of five independent donors for bortezomib experiments (A) and 12 independent donors for LEN experiments (B). The data were normalized to the value obtained from cells treated with vehicle control. The mean (range) of absolute concentrations of CpG-ODN 2216 + vehicle control was 29,786 pg/mL (12,921–56,217 pg/mL). (C) Purified pDCs were incubated for 24 h with 1 μM LEN or vehicle in the presence of the indicated concentrations of CpG-ODN 2216. Data are derived from at least six independent donors at each CpG-ODN 2216 concentration and are shown as means ± SEMs. The concentrations of IFN-α in culture supernatants were measured by ELISA. Statistical significance was determined using paired Student’s t-tests (*=p<0.05, **=p<0.01; p-values were calculated for each concentration of LEN or bortezomib versus vehicle control). |
Discussion
MM involves an element of immune dysfunction,12,28,29 including an increased number of Tregs, which is associated with poor clinical outcomes.30,31 Therefore, immunotherapies or drugs that can improve immune status are considered effective therapeutic strategies, making IMiDs backbone drugs for MM. Although LEN was the first anti-MM drug with an immunomodulatory effect, its mechanism of action had been elusive. The present study showed that LEN potentially amplified the ability of pDCs to produce endogenous IFN-α and elucidated an additional cellular target of LEN. Our results provide new insight into a possible mechanism through which LEN operates as a pleiotropic immunomodulator.
In our study, LEN significantly enhanced IFN-α production by pDCs stimulated with the low concentrations of CpG-ODN but not with an optimal high concentration. Although pDCs rapidly produce vast amounts of type I IFNs following stimulation by viruses or CpG-ODN,13 pDCs are incapable of mounting a secondary type I IFN response to further stimulation.15 In this context, pDCs do not retain sufficient capacity to further produce type I IFNs by the maximal response to optimal stimulation. However, LEN was able to promote the residual capacity of pDCs to produce IFN-α when stimulated with a low concentration of CpG-ODN in our study.
Type I IFNs directly inhibit the proliferation of tumors and virus-infected cells, and are known to play a central role in antitumor and antiviral immune responses.32 Furthermore, type I IFNs directly activate diverse immune cell types, such as NK cells, B cells, monocytes, antigen-presenting DCs, and T cells.16–18 Type I IFNs are able to enhance cellular activities, such as the ability of NK cells to kill target cells, monocyte function, and the differentiation of monocytes into DCs.33–35 Moreover, type I IFNs suppress generation of Treg cells.36 These functions of type I IFNs might work synergistically to mediate the effect of LEN in enhancing cellular immunity.3–10 Furthermore, type I IFNs are directly involved in the maturation and sustainment of conventional DCs,37 and also affect the ability of myeloid DCs to secrete IL-12p70.15,38 B cell functions are enhanced by type I IFNs during antiviral humoral responses,39 and enhancement of antibody responses occurs through direct stimulation of B and T cells by type I IFNs.40 Thus, type I IFNs contribute to elimination of microbes and tumors through activation of cytotoxic effector cells involved in innate and adaptive immunity. Because of the pleiotropic effects of type I IFNs on various immune cells, pDCs can enhance the functions of NK cells,41 monocytes,42 blood myeloid DCs,43 B cells,44 and CD8 T cells.45 The potential cross-talk between pDCs and other immune cells occurs mainly through type I IFNs. Thus, pDCs trigger activation of the immune system as a part of their antiviral and antitumor responses through type I IFN production. In this sense, pDCs may participate in various immune aspects of IMiD treatment for MM. Our results suggest that LEN may function as an enhancer of endogenous IFN and a positive immunomodulator which activates surrounding immune cells. In supports of this hypothesis, there is relatively low incidence of herpes zoster viral infection during LEN treatment compared with bortezomib treatment.46–48
No decrease in blood pDCs was observed in MM patients following administration of LEN in our study. This finding was further supported by the result that LEN did not adversely affect pDC survival and maturation in vitro. No conclusion could be reached regarding the blood DC numbers in MM patients. One report showed a decreased number of blood pDCs in MM patients compared with healthy donors,49 whereas two other studies found that blood pDC numbers in MM patients were nearly identical to those of normal donors.28,50 We have previously reported that blood pDC numbers were unaltered in patients with tuberculosis51 and Sjögren syndrome,52 but decreased in patients with sarcoidosis53 and acute graft-versus-host disease.54 Although only a small number of patients were included in the current study, we also found no significant difference in blood pDC numbers between healthy donors and MM patients. It is possible that pDCs in MM patients, within the normal numeric range, can function better when LEN is administered.
Continuous treatment with low dose LEN as a maintenance therapy contributes to better survival in MM patients.55,56 The clinical pharmacokinetics of LEN show a gentle curve from the peak plasma concentration: 10 mg of LEN (administered orally) to MM patients results in an approximate clinical peak plasma concentration of 1.2 µM.23 Our results showed that, even at a low concentration (0.1–1.0 μM, equivalent to the clinical plasma concentration range resulting from oral administration of 10 mg LEN), LEN sustained IFN-α production by PBMCs and pDCs. This finding suggested that low dose LEN (ie, 10 mg oral administration) could function as an immunostimulator during maintenance therapy.
Conclusions
The results of our study showed that LEN sustained and potentially enhanced the ability of pDCs to produce type I IFNs without affecting their survival and maturation. LEN orchestrates the activities of a wide variety of both innate and adaptive immune cells, leading to amplification of a positive immune axis for eliminating MM cells. Our findings unveiled one part of the mechanism responsible for the immunomodulatory functions of LEN in the treatment of MM. In addition, our data provide a plausible explanation for the observed benefit of LEN in boosting immune defense against herpes zoster virus.
Ethics approval and informed consent
All studies involving human samples were performed under institutional review board-approved protocols at Kansai Medical University. All subjects provided written informed consent in accordance with the recommendations of the International Conference on Harmonization Guidelines for Good Clinical Practice and the Declaration of Helsinki.
Acknowledgments
We thank Sarah Bubeck, Ph.D., from Edanz Group, for editing a draft of this manuscript. This study was supported in part by a grant (17K09963 to TI) from the Ministry of Education, Science and Culture of Japan.
Disclosure
Tomoki Ito received honoraria from Celgene, Bristol-Myers Squibb, and Takeda. Tomoki Ito also reports personal fees from Celgene, Bristol-Myers Squibb, and Takeda, during the conduct of the study. The authors report no other conflicts of interest in this work.
References
1. Benboubker L, Dimopoulos MA, Dispenzieri A, et al. Lenalidomide and dexamethasone in transplant-ineligible patients with myeloma. N Engl J Med. 2014;371(10):906–917. doi:10.1056/NEJMoa1402551
2. Paiva B, Mateos MV, Sanchez-Abarca LI, et al. Immune status of high-risk smoldering multiple myeloma patients and its therapeutic modulation under LenDex: a longitudinal analysis. Blood. 2016;127(9):1151–1162. doi:10.1182/blood-2015-10-662320
3. Gandhi AK, Kang J, Capone L, et al. Dexamethasone synergizes with lenalidomide to inhibit multiple myeloma tumor growth, but reduces lenalidomide-induced immunomodulation of T and NK cell function. Curr Cancer Drug Targets. 2010;10(2):155–167.
4. Davies FE, Raje N, Hideshima T, et al. Thalidomide and immunomodulatory derivatives augment natural killer cell cytotoxicity in multiple myeloma. Blood. 2001;98(1):210–216. doi:10.1182/blood.v98.1.210
5. Wu L, Adams M, Carter T, et al. lenalidomide enhances natural killer cell and monocyte-mediated antibody-dependent cellular cytotoxicity of rituximab-treated CD20+ tumor cells. Clin Cancer Res. 2008;14(14):4650–4657. doi:10.1158/1078-0432.CCR-07-4405
6. Mitsiades N, Mitsiades CS, Poulaki V, et al. Apoptotic signaling induced by immunomodulatory thalidomide analogs in human multiple myeloma cells: therapeutic implications. Blood. 2002;99(12):4525–4530. doi:10.1182/blood.v99.12.4525
7. Schafer PH, Gandhi AK, Loveland MA, et al. Enhancement of cytokine production and AP-1 transcriptional activity in T cells by thalidomide-related immunomodulatory drugs. J Pharmacol Exp Ther. 2003;305(3):1222–1232. doi:10.1124/jpet.102.048496
8. Reddy N, Hernandez-Ilizaliturri FJ, Deeb G, et al. Immunomodulatory drugs stimulate natural killer-cell function, alter cytokine production by dendritic cells, and inhibit angiogenesis enhancing the anti-tumour activity of rituximab in vivo. Br J Haematol. 2008;140(1):36–45. doi:10.1111/j.1365-2141.2007.06841.x
9. Galustian C, Meyer B, Labarthe MC, et al. The anti-cancer agents lenalidomide and pomalidomide inhibit the proliferation and function of T regulatory cells. Cancer Immunol Immunother. 2009;58(7):1033–1045. doi:10.1007/s00262-008-0620-4
10. Luptakova K, Rosenblatt J, Glotzbecker B, et al. Lenalidomide enhances anti-myeloma cellular immunity. Cancer Immunol Immunother. 2013;62(1):39–49. doi:10.1007/s00262-012-1308-3
11. Benson DM, Bakan CE, Mishra A, et al. The PD-1/PD-L1 axis modulates the natural killer cell versus multiple myeloma effect: a therapeutic target for CT-011, a novel monoclonal anti-PD-1 antibody. Blood. 2010;116(13):2286–2294. doi:10.1182/blood-2010-02-271874
12. Görgün G, Samur MK, Cowens KB, et al. Lenalidomide enhances immune checkpoint blockade-induced immune response in multiple myeloma. Clin Cancer Res. 2015;21(20):4607–4618. doi:10.1158/1078-0432.CCR-15-0200
13. Siegal FP, Kadowaki N, Shodell M, et al. The nature of the principal type 1 interferon-producing cells in human blood. Science. 1999;284(5421):1835–1837.
14. Liu YJ. IPC: professional type 1 interferon-producing cells and plasmacytoid dendritic cell precursors. Annu Rev Immunol. 2005;23:275–306. doi:10.1146/annurev.immunol.23.021704.115633
15. Ito T, Kanzler H, Duramad O, Cao W, Liu Y-J. Specialization, kinetics, and repertoire of type 1 interferon responses by human plasmacytoid predendritic cells. Blood. 2006;107(6):2423–2431. doi:10.1182/blood-2005-07-2709
16. Trinchieri G, Santoli D. Anti-viral activity induced by culturing lymphocytes with tumor-derived or virus-transformed cells. Enhancement of human natural killer cell activity by interferon and antagonistic inhibition of susceptibility of target cells to lysis. J Exp Med. 1978;147(5):1314–1333. doi:10.1084/jem.147.5.1314
17. Le Bon A, Schiavoni G, D’Agostino G, Gresser I, Belardelli F, Tough DF. Type i interferons potently enhance humoral immunity and can promote isotype switching by stimulating dendritic cells in vivo. Immunity. 2001;14(4):461–470.
18. Le Bon A, Etchart N, Rossmann C, et al. Cross-priming of CD8+ T cells stimulated by virus-induced type I interferon. Nat Immunol. 2003;4(10):1009–1015. doi:10.1038/ni978
19. Ito T, Wang YH, Liu YJ. Plasmacytoid dendritic cell precursors/type I interferon-producing cells sense viral infection by Toll-like receptor (TLR) 7 and TLR9. Springer Semin Immunopathol. 2005;26(3):221–229. doi:10.1007/s00281-004-0180-4
20. Henry JY, Labarthe MC, Meyer B, Dasgupta P, Dalgleish AG, Galustian C. Enhanced cross-priming of naive CD8+ T cells by dendritic cells treated by the IMiDs® immunomodulatory compounds lenalidomide and pomalidomide. Immunology. 2013;139(3):377–385. doi:10.1111/imm.12087
21. Torii Y, Ito T, Amakawa R, et al. Imidazoquinoline acts as immune adjuvant for functional alteration of thymic stromal lymphopoietin-mediated allergic T cell response. J Immunol. 2008;181(8):5340–5349. doi:10.4049/jimmunol.181.8.5340
22. Ito T, Inaba M, Inaba K, et al. A CD1a+/CD11c+ subset of human blood dendritic cells is a direct precursor of Langerhans cells. J Immunol. 1999;163(3):1409–1419.
23. Iida S, Chou T, Okamoto S, et al. Lenalidomide plus dexamethasone treatment in Japanese patients with relapsed/refractory multiple myeloma. Int J Hematol. 2010;92(1):118–126. doi:10.1007/s12185-010-0624-7
24. Moreau P, Karamanesht II, Domnikova N, et al. Pharmacokinetic, pharmacodynamic and covariate analysis of subcutaneous versus intravenous administration of bortezomib in patients with relapsed multiple myeloma. Clin Pharmacokinet. 2012;51(12):823–829. doi:10.1007/s40262-012-0010-0
25. Hirai M, Kadowaki N, Kitawaki T, et al. Bortezomib suppresses function and survival of plasmacytoid dendritic cells by targeting intracellular trafficking of Toll-like receptors and endoplasmic reticulum homeostasis. Blood. 2011;117(2):500–509. doi:10.1182/blood-2010-05-284737
26. Chanan-Khan AA, Swaika A, Paulus A, et al. Pomalidomide: the new immunomodulatory agent for the treatment of multiple myeloma. Blood Cancer J. 2013;3:e143. doi:10.1038/bcj.2013.38
27. POMALYST® [PRESCRIBING INFORMATION]: Celgene Corporation. 2018.
28. Brown RD, Pope B, Murray A, et al. Dendritic cells from patients with myeloma are numerically normal but functionally defective as they fail to up-regulate CD80 (B7-1) expression after huCD40LT stimulation because of inhibition by transforming growth factor-beta1 and interleukin-10. Blood. 2001;98(10):2992–2998. doi:10.1182/blood.v98.10.2992
29. Ratta M, Fagnoni F, Curti A, et al. Dendritic cells are functionally defective in multiple myeloma: the role of interleukin-6. Blood. 2002;100(1):230–237. doi:10.1182/blood.v100.1.230
30. Giannopoulos K, Kaminska W, Hus I, Dmoszynska A. The frequency of T regulatory cells modulates the survival of multiple myeloma patients: detailed characterisation of immune status in multiple myeloma. Br J Cancer. 2012;106(3):546–552. doi:10.1038/bjc.2011.575
31. Muthu Raja KR, Rihova L, Zahradova L, et al. Increased T regulatory cells are associated with adverse clinical features and predict progression in multiple myeloma. PLoS One. 2012;7(10):e47077. doi:10.1371/journal.pone.0047077
32. Müller U, Steinhoff U, Reis LF, et al. Functional role of type I and type II interferons in antiviral defense. Science. 1994;264(5167):1918–1921.
33. Lee CK, Rao DT, Gertner R, Gimeno R, Frey AB, Levy DE. Distinct requirements for IFNs and STAT1 in NK cell function. J Immunol. 2000;165(7):3571–3577. doi:10.4049/jimmunol.165.7.3571
34. Bogdan C, Mattner J, Schleicher U. The role of type I interferons in non-viral infections. Immunol Rev. 2004;202:33–48. doi:10.1111/j.0105-2896.2004.00207.x
35. Trinchieri G, Santoli D, Granato D, Perussia B. Antagonistic effects of interferons on the cytotoxicity mediated by natural killer cells. Fed Proc. 1981;40(12):2705–2710.
36. Golding A, Rosen A, Petri M, Akhter E, Andrade F. Interferon-alpha regulates the dynamic balance between human activated regulatory and effector T cells: implications for antiviral and autoimmune responses. Immunology. 2010;131(1):107–117. doi:10.1111/j.1365-2567.2010.03280.x
37. Ito T, Amakawa R, Inaba M, Ikehara S, Inaba K, Fukuhara S. Differential regulation of human blood dendritic cell subsets by IFNs. J Immunol. 2001;166(5):2961–2969. doi:10.4049/jimmunol.166.5.2961
38. Gautier G, Humbert M, Deauvieau F, et al. A type I interferon autocrine-paracrine loop is involved in Toll-like receptor-induced interleukin-12p70 secretion by dendritic cells. J Exp Med. 2005;201(9):1435–1446. doi:10.1084/jem.20041964
39. Fink K, Lang KS, Manjarrez-Orduno N, et al. Early type I interferon-mediated signals on B cells specifically enhance antiviral humoral responses. Eur J Immunol. 2006;36(8):2094–2105. doi:10.1002/eji.200635993
40. Le Bon A, Thompson C, Kamphuis E, et al. Cutting edge: enhancement of antibody responses through direct stimulation of B and T cells by type I IFN. J Immunol. 2006;176(4):2074–2078. doi:10.4049/jimmunol.176.4.2074
41. Hanabuchi S, Watanabe N, Wang YH, et al. Human plasmacytoid predendritic cells activate NK cells through glucocorticoid-induced tumor necrosis factor receptor-ligand (GITRL). Blood. 2006;107(9):3617–3623. doi:10.1182/blood-2005-08-3419
42. Blanco P, Palucka AK, Gill M, Pascual V, Banchereau J. Induction of dendritic cell differentiation by IFN-alpha in systemic lupus erythematosus. Science. 2001;294(5546):1540–1543. doi:10.1126/science.1064890
43. Fonteneau JF, Larsson M, Beignon AS, et al. Human immunodeficiency virus type 1 activates plasmacytoid dendritic cells and concomitantly induces the bystander maturation of myeloid dendritic cells. J Virol. 2004;78(10):5223–5232. doi:10.1128/JVI.78.10.5223-5232.2004
44. Jego G, Palucka AK, Blanck JP, Chalouni C, Pascual V, Banchereau J. Plasmacytoid dendritic cells induce plasma cell differentiation through type I interferon and interleukin 6. Immunity. 2003;19(2):225–234.
45. Rothenfusser S, Hornung V, Ayyoub M, et al. CpG-A and CpG-B oligonucleotides differentially enhance human peptide-specific primary and memory CD8+ T-cell responses in vitro. Blood. 2004;103(6):2162–2169. doi:10.1182/blood-2003-04-1091
46. San Miguel JF, Schlag R, Khuageva NK, et al. Bortezomib plus melphalan and prednisone for initial treatment of multiple myeloma. N Engl J Med. 2008;359(9):906–917. doi:10.1056/NEJMoa0801479
47. VELCADE® [product information]: Janssen. 2016.
48. REVLIMID® [Package Insert]. Summit, NJ: Celgene Corporation; 2017.
49. Brimnes MK, Svane IM, Johnsen HE. Impaired functionality and phenotypic profile of dendritic cells from patients with multiple myeloma. Clin Exp Immunol. 2006;144(1):76–84. doi:10.1111/j.1365-2249.2006.03037.x
50. Chauhan D, Singh AV, Brahmandam M, et al. Functional interaction of plasmacytoid dendritic cells with multiple myeloma cells: a therapeutic target. Cancer Cell. 2009;16(4):309–323. doi:10.1016/j.ccr.2009.08.019
51. Uehira K, Amakawa R, Ito T, et al. Dendritic cells are decreased in blood and accumulated in granuloma in tuberculosis. Clin Immunol. 2002;105(3):296–303.
52. Ozaki Y, Amakawa R, Ito T, et al. Alteration of peripheral blood dendritic cells in patients with primary Sjögren’s syndrome. Arthritis Rheum. 2001;44(2):419–431. doi:10.1002/1529-0131(200102)44:2<419::AID-ANR61>3.0.CO;2-U
53. Ota M, Amakawa R, Uehira K, et al. Involvement of dendritic cells in sarcoidosis. Thorax. 2004;59(5):408–413. doi:10.1136/thx.2003.006049
54. Takebayashi M, Amakawa R, Tajima K, et al. Blood dendritic cells are decreased in acute graft-versus-host disease. Bone Marrow Transplant. 2004;33(10):989–996. doi:10.1038/sj.bmt.1704406
55. Attal M, Lauwers-Cances V, Marit G, et al. Lenalidomide maintenance after stem-cell transplantation for multiple myeloma. N Engl J Med. 2012;366(19):1782–1791. doi:10.1056/NEJMoa1114138
56. McCarthy PL, Owzar K, Hofmeister CC, et al. Lenalidomide after stem-cell transplantation for multiple myeloma. N Engl J Med. 2012;366(19):1770–1781. doi:10.1056/NEJMoa1114083
Supplementary material
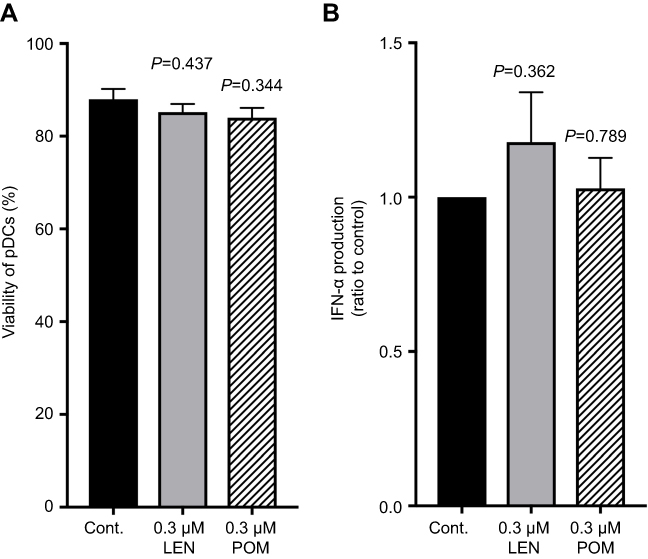 |
Figure S1 Clinical concentrations of pomalidomide sustained IFN-α production by human plasmacytoid dendritic cells. Purified plasmacytoid dendritic cells (pDCs) were incubated for 24 h with 0.3 µM lenalidomide (LEN), 0.3 µM pomalidomide (POM), or vehicle in the presence of 3 µM CpG-ODN 2216. (A) Viable cells were quantitated by annexin V staining using flow cytometry. Percentages of annexin V-positive cells are indicated. (B) The concentrations of IFN-α in culture supernatants were measured by ELISA. The data were normalized to the value obtained from cells treated with vehicle control. The mean (range) of absolute concentrations of CpG-ODN 2216 + vehicle control was 26,745 pg/mL (19,887–46,814 pg/mL). Data are shown as means ± SEMs of five independent donors (A and B). Statistical significance was determined using paired Student’s t-tests; p-values were calculated for each concentration of LEN or POM versus vehicle control. |
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.