Back to Journals » Journal of Experimental Pharmacology » Volume 13
The Effects of Furosemide on Behavioral and Hormonal Parameters in Male and Female Mice Subjected to Immobilization and Cold-Water Stress
Authors Al Za'abi M ![]() , Ali BH, Al Suleimani Y, Al-Zakwani I
, Ali BH, Al Suleimani Y, Al-Zakwani I ![]() , Al-Fulaiti B, Manoj P, Nemmar A
, Al-Fulaiti B, Manoj P, Nemmar A ![]()
Received 7 February 2021
Accepted for publication 31 May 2021
Published 5 July 2021 Volume 2021:13 Pages 637—643
DOI https://doi.org/10.2147/JEP.S305770
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Edythe London
Mohammed Al Za’abi,1 Badreldin H Ali,1 Yousuf Al Suleimani,1 Ibrahim Al-Zakwani,1 Balqees Al-Fulaiti,1 Priyadarsini Manoj,1 Abderrahim Nemmar2
1Department of Pharmacology and Clinical Pharmacy, College of Medicine and Health Sciences, Sultan Qaboos University, Muscat, Oman; 2Department of Physiology, College of Medicine and Health Sciences, UAE University, Al Ain, United Arab Emirates
Correspondence: Mohammed Al Za’abi
Department of Pharmacology, College of Medicine and Health Sciences, Sultan Qaboos University, P. O. Box 35 Al Khoud, Muscat, 123, Oman
Email [email protected]
Introduction: The diuretic agent furosemide (FUR, 25 and 50 mg/kg) has been shown in a single report to act as an anti-stressor agent in two models of acute stress in mice, viz. electric foot–shock stress and immobilization (IMS). The present work aimed to investigate the possible anti-stressor action of FUR on two models of acute stress in mice, cold-water stress (CWS) and IMS, and tried to determine whether gender has any impact on the effect of FUR.
Methods: FUR (40 mg/kg) was injected intraperitoneally, and after 30 minutes, mice were subjected to CWS (4°C for three minutes) or IMS (fixing movement for two and a half hrs using adhesive tape). Motor and exploratory activities, neuromuscular coordination, and thermal nociception were then tested. Blood was collected from the mice and used to measure the concentrations of three stress hormones (corticosterone, epinephrine and prolactin).
Results: Mice subjected to CWS and IMS had significantly reduced motor and exploratory activities, neuromuscular coordination, and increased nociception. CWS and IMS also significantly increased the plasma concentrations of the three hormones. FUR pretreatment significantly mitigated these stress-induced hormonal changes. There was no significant sex difference when CWS or IMS was applied.
Discussion: IMS and CWS stimuli in male and female mice caused significant elevations in the plasma concentrations of corticosterone, epinephrine, and prolactin, accompanied by a significant reduction of motor and exploratory activities, neuromuscular coordination, and thermal nociception. There were no sex differences when IMS was applied. In stressed mice, prior administration of FUR (40 mg/kg) significantly decreased the concentrations of stress hormones, and this effect significantly mitigated the stress-induced behavioural and motor changes.
Keywords: furosemide, immobilization stress, cold-water stress, corticosterone, prolactin, epinephrine
Introduction
Stress is a nonspecific state that can cause behavioural and physiological changes that disturbs the homeostasis of the individual organism.1,2 Coping with stress is a vital determinant of health, and failure to do so may result in the development of various pathophysiological alterations and diseases.3
Stress can be acute or chronic, depending on the frequency and duration of exposure to the stressor.4,5 Acute stress denotes a short period (24 hours or less) of exposure to a particular stressor, while a long period of exposure leads to chronic stress. The former involves activation of the hypothalamic-pituitary-adrenal (HPA) axis and the sympathetic adrenomedullary system. The latter leads to dysregulation of the HPA axis, possibly due to HPA sensitization.6 The physiological response to stress in rodents depends on the activation of both the HPA axis (leading to the release of corticosterone) and the sympathetic adrenomedullary system (causing the release of catecholamines from sympathetic nerve terminals and the adrenal medulla). Corticosterone is probably the most important hormone in the stress response in rodents, and together with epinephrine and prolactin, is used as a biological marker for assessing the stress response.7 In this work, we focused on two models of acute stress only.
Furosemide (FUR) is a high-ceiling loop diuretic that acts as a sodium-potassium chloride cotransporter inhibitor.8 The potassium chloride cotransporter (KCC) family is present in various tissues, including the loop of Henle in the nephron of the kidney and hypothalamus, and belongs to the cation-dependent chloride cotransporter family. They mainly mediate the transport of K+ and Cl− ions into the cell. The neuronal action of KCCs reverses the inhibitory action of gamma-aminobutyric acid neurotransmitter (GABA) in the brain. KCC activation is the main mechanism by which mature neurons maintain low intracellular Cl− concentrations.9
In a single report, single intraperitoneal (i.p.) doses of FUR at 25 and 50 mg/kg have been shown to act as anti-stressor agents in two models of acute stress in mice, viz. electric footshock stress and immobilization.10 In that report, only corticosterone was used as a biomarker of stress. In the present work, we used FUR at a different dose (40 mg/kg, i.p.) to assess its possible action against another stress model, viz. cold-water immersion (CWS), which has not been tested before, and to verify its action against immobilization stress (IMS) using four behavioural paradigms and three stress hormones.
Data on gender differences in preclinical neuroscience work are scarce and are mostly contradictory.11–13 Therefore, we thought it of interest to determine whether there were sex differences in response to the two acute stressful stimuli and the effect of FUR thereon.
Materials and Methods
Animals
The animals used were male and female CD1 mice (9–10 weeks old and weighing between 28–30 g) obtained from the small animal house of Sultan Qaboos University (SQU), housed under controlled environmental conditions (24 ± 2°C and 12 h light-dark cycle), and allowed free access to a standard chow diet (Oman Flour Mill, Muscat, Oman) and tap water. Ethical approval for conducting the work was approved by the Animal Ethical Committee of SQU (approval code: SQU/AEC/2017-18/9). Animal care and involved procedures were carried out following international laws and policies (EEC Council directives 2010/63/EU, 22 September 2010, and NIH guide for the Care and Use of Laboratory Animals, NIH publication no. 85–23, 1985).
Experimental Design
Male mice (n = 72) and female mice (n = 72) were each randomly divided into six equal groups (n=12 each). The experimental design was as follows:
G1 (vehicle, control): mice were injected once with normal saline intraperitoneally; i.p. (40 mg/kg) 30 min before conducting the activity tests.
G2 (FUR): mice were administered a single dose of FUR (40 mg/kg) i.p. 30 min before conducting the activity tests.
G3 (CWS-saline): mice were administered a single dose of normal saline (40 mg/kg) i.p. 30 min before being subjected to CWS for 3 min, after which the tests were performed.
G4 (CWS- FUR): mice were administered a single dose of FUR (40 mg/kg) i.p. 30 min before being subjected to CWS for 3 min, after which the tests were performed.
G5 (IMS-saline): treated similarly to G3 except that the CWS was replaced with IMS for two and a half hours.
G6 (IMS-FUR): treated similarly to G4 except that the CWS was replaced with IMS for two and a half hours.
All the tests were performed at a similar time of the 12 h light cycle of the day (8–11 am).
CWS
This acute stress model was applied to mice as described previously.14,15 Each mouse was gently dropped into a two-litre glass beaker (18 cm in height, 13 cm in diameter) filled with water at 4°C up to 11 cm for three minutes. The water level was deep enough to cover the back of the mouse, while it swam in cold water or kept its head above the water level. At the end of each stress period (3 minutes), each mouse was immediately removed and dried with towel papers to avoid hypothermia.
IMS
This stress model was applied as described previously.16 IMS was induced by fixing the entire body of each mouse using adhesive tape for two and a half hours. Each mouse was kept in the prone position with stretching all its limbs to restrict their movement in a quiet room.6
Motor Activity Test
Locomotor activity is a marker of awareness. The open-field test was employed to evaluate the spontaneous activity, general exploration, and ambulation of the rodents. Each mouse was placed in the centre of the open field in the activity metre (Ugo Basile, Comerio, Italy). Then, the number of line crossings was measured to study the effect of stress on the general locomotor activity level of stressed mice. The test was conducted for 15 min, but the values of the first five minutes were excluded from the calculation as described previously.17–19
Exploratory Activity Test (Hole Board)
Each mouse was placed in the centre of the hole board to test its exploratory activity. Then, the total number of nose dips was measured to evaluate the change related to behavioural stress of each mouse. The test was carried out within 15 min, but the values of the first five minutes were excluded from the calculation as described previously.18
Coordination Activity Test (Rota-Rod Treadmill)
Mice were gently placed on a rotarod treadmill (Ugo Basile, Comerio VA, Italy) to study neuromuscular activity. The apparatus used was subdivided into five segments by discs with a diameter of 24 cm, and the rod was 30 cm long and 3 cm in diameter. The rod was rotated at a fixed speed of 25 revolutions/min, and the time taken by the animal to fall from the rotating rod was automatically recorded.20
Thermal Nociception Activity Test (Hot Plate)
In this test, the mice were placed in an analgesia metre (Ugo Basile, Comerio VA, Italy) having a glass cylinder with a hot metal plate set to a temperature of 55 ± 0.2°C for measuring analgesia.21 The time taken by the mouse to jump off the hot plate or lick its paw was recorded and considered the responding time for its reflex. The cut-off time was 15 sec to prevent possible tissue damage.
Blood Collection and Biochemical Analysis
Animals were anaesthetized using ketamine (75 mg kg−1) and xylazine (5 mg kg−1) given i.p., and blood was immediately collected in heparinized tubes from the abdominal aorta of each animal and centrifuged at 900 g for 15 min at 5°C to separate plasma. The plasma collected was used for the measurement of the stress hormones corticosterone, epinephrine and prolactin using ELISA techniques.
Drugs and Chemicals
FUR (Sanofi Winthrop, Paris, France) was purchased from the Pharmacy of Sultan Qaboos University Hospital. A mouse epinephrine ELISA kit was purchased from Cusabio, Houston, TX, USA. Mouse prolactin and corticosterone ELISA kits were obtained from Thermo Scientific (Waltham, CA, USA).
Statistical Analysis
Descriptive statistics were used to describe the data. For categorical variables, frequencies and percentages were reported. For continuous variables, the mean and standard deviation were used to summarize the data. In Tables 1 (male mice) and 2 (female mice), the effect of drug (furosemide/saline) on motor and exploratory activities (motor/horizontal, motor/vertical, total dips/hole board, rotarod, hot plate) in male and female mice subjected to stress (CWS/IMS) was performed using three-way analysis of variance (ANOVA) with multiple pairwise comparisons conducted utilizing the Bonferroni test. In Table 3, the effect of drug (furosemide/saline) on hormone (corticosterone, epinephrine, and prolactin) concentrations of mice subjected to stress (CWS/IMS) and stratified by sex (male/female) was performed using four-way analysis of variance (ANOVA) with multiple pairwise comparisons also conducted utilizing the Bonferroni test. An a priori two-tailed level of significance was set at 0.05. Statistical analyses were performed using STATA version 16.1 (STATA Corporation, College Station, TX, USA).
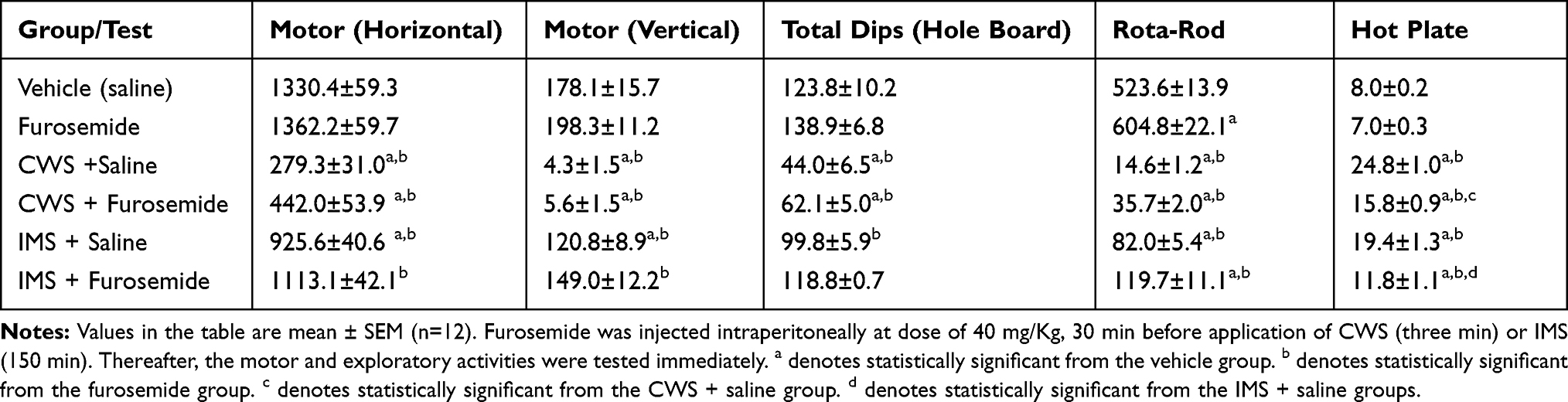 |
Table 1 The Effect of Furosemide Treatment on Motor and Exploratory Activities in Male Mice Subjected to Cold Water Stress (CWS) and Immobilization Stress (IMS) |
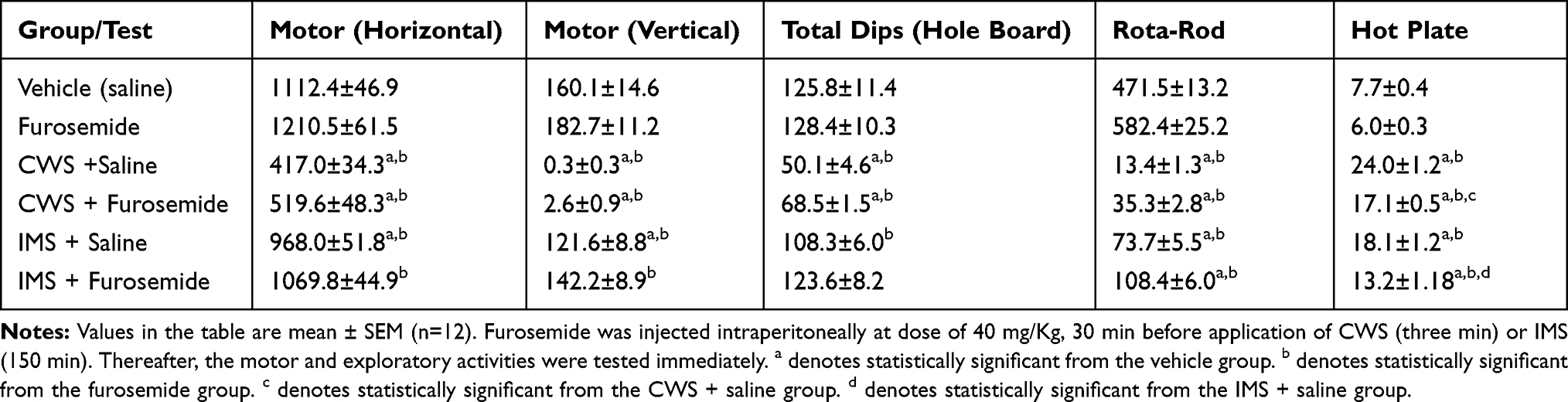 |
Table 2 The Effect of Furosemide Treatment on Motor and Exploratory Activities in Female Mice Subjected to Cold Water Stress (CWS) and Immobilization Stress (IMS) |
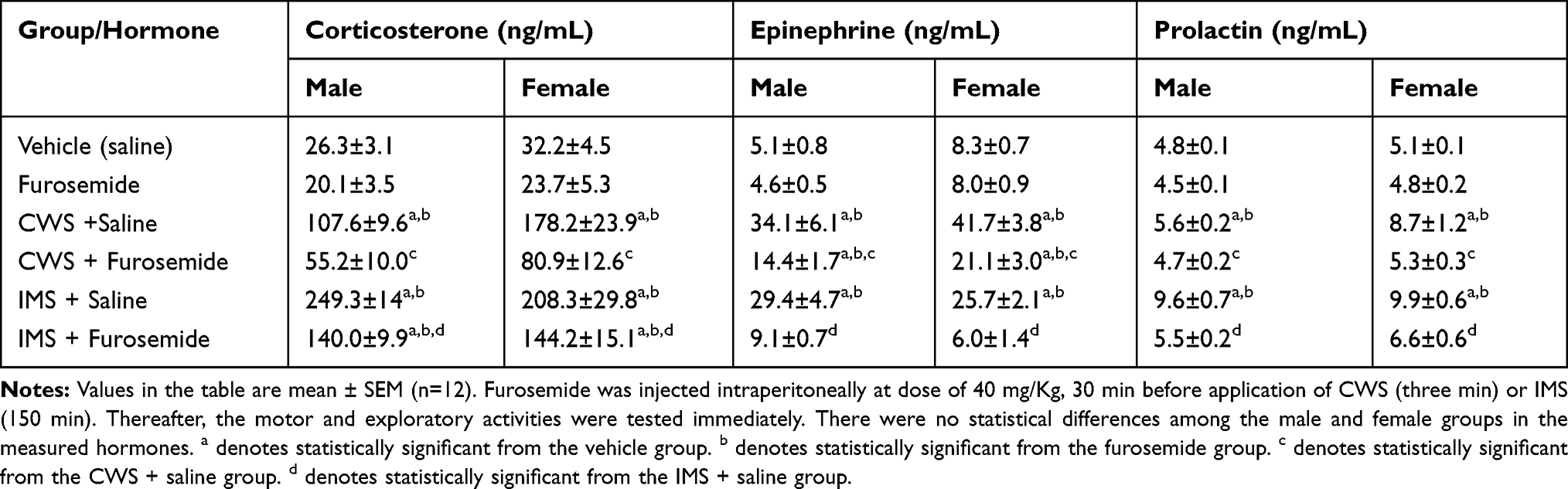 |
Table 3 The Effect of Furosemide Treatment on Corticosterone, Epinephrine and Prolactin Concentrations in Male and Female Mice Subjected to Motor and Exploratory Activities Subsequent to Cold Water Stress (CWS) and Immobilization Stress (IMS) |
Results
Tables 1 and 2 show the results obtained in the various tests in male and female mice, respectively.
Motor Activity Test
There were no differences between the FUR and vehicle groups in either horizontal or vertical motor activity. The results of the CWS-saline, CWS-FUR and IMS-saline groups were significantly different from those of the vehicle and FUR groups, while the IMS-FUR group was significantly different from the FUR group but not the vehicle group. FUR did not significantly ameliorate IMS or CWS effects on motor activities. There were no significant sex differences among the groups in response to motor activity.
Exploratory Activity Test (Hole Board)
In the hole board test, there were no significant differences in the results of the FUR and vehicle groups. The CWS-saline and CWS-FUR groups were significantly different from the vehicle and FUR groups. The IMS-saline group result was lower and significantly different from the FUR but not the vehicle group. FUR significantly mitigated the IMS but not the CWS effects. There were no significant sex differences among the groups in response to exploratory activity.
Coordination Activity Test (Rota-Rod Treadmill)
The rotarod test results for all groups were significantly different from the vehicle group. All groups’ results were also significantly different from those of the FUR group. However, FUR did not have a significant impact on the CWS and IMS effects in performing this test. There were no significant sex differences among the groups in response to rotarod activity.
Thermal Nociception Activity Test (Hot Plate)
The FUR group results were not significantly different from the vehicle group in thermal antinociception results, but all other groups were significantly different from both the vehicle and FUR groups. FUR significantly reduced the CWS and IMS effects on the hot-plate test. There were no significant sex differences among the groups in response to the hot plate test.
Effects of CWS and IMS on Mice Hormones
Table 3 shows the results of the obtained values of the measured hormones in various groups. The obtained corticosterone, epinephrine, and prolactin levels in the FUR groups were not significantly different from those in the vehicle group. Corticosterone values in the rest of the groups were significantly different from the vehicle group except for the CWS-FUR group. In contrast, epinephrine values in the IMS-FUR group were not significantly different from those in the vehicle group. Prolactin values in the other groups except for the CWS-FUR and IMS-FUR groups were significantly different than those in the vehicle group. FUR significantly reduced the impact of CWS and IMS on the levels of all three hormones. There were no significant sex differences among the groups in the measured hormones.
Discussion
This work presents experimental evidence that the two acute stressful stimuli (CWS and IMS) significantly reduced motor, exploratory activities, and neuromuscular coordination, and increased nociception in mice. It also showed that these two stressful stimuli significantly elevated the levels of three hormonal biomarkers of stress. Pretreatment with FUR significantly reduced these effects.
IMS is one of the intensive stress models employed extensively for studying stress-related biochemical, biological, and physiological responses in animals.14 It is a relatively easy and convenient method to induce both physical (eg, muscular struggle, extension, and exertion) and psychologically stressful stimuli (eg, restricting the animal and limiting its activity).22 In our experiment, IMS induced significant alterations in motor and exploratory activities, neuromuscular coordination, and thermal nociception in both sexes to an equal extent. Some of these effects have been reported earlier only in one prior publication on this subject.10
The adverse actions caused by IMS can cause alterations in the actions of peripheral muscles and changes in the flow rate of blood that supplies these muscles.23 This stressful action activates the HPA system, resulting in at least some behavioural, biochemical, and hormonal changes .
Similar to IMS, CWS is commonly used to induce acute stress in animals. In this acute stress model, there are both physical and psychological stressful stimuli.13 In this work, CWS significantly caused behavioural actions similar to those induced by IMS. CWS subjects the animal to a sudden decrease in core body temperature, resulting in alterations in the metabolic state and deleterious effects on the behaviour of the mice.24 CWS has also been reported to induce the release of catecholamines (mainly epinephrine) and other stress hormones.14
The behavioural changes induced by stress are strongly associated with the release of stress hormones.25 Certain areas of the brain are responsible for vehicle animals’ behaviour during a stressful stimulus (viz. cingulate gyrus, fornix, hippocampus, amygdala, hypothalamus, and anterior pituitary). These areas are highly susceptible to stress due to high expression of corticosterone receptors.26 Furthermore, the release of corticosterone during stress affects these brain regions, leading to alterations in behaviour.27
IMS significantly increased the plasma concentrations of stress hormones in both sexes. Elevated concentrations of plasma stress hormones were also observed in mice subjected to CWS. These changes could be explained by the slower activation effect of stress in the HPA axis that leads to activation of the adrenal cortex. Hyperactivation of the HPA axis increases the release of corticosterone from the adrenal cortex. In contrast, the rapid activation effect of stress affects the sympatho-adrenomedullary system, leading to the release of epinephrine by activating the adrenal medulla.14
Stressful stimuli can also induce the release of prolactin, which acts on the adrenal gland to increase the secretion of corticosterone hormone and increases the sensitivity of the adrenal gland to corticosterone.28 Both types of stress stimuli decreased the motor and exploratory activities of both male and female mice in this study. There were no significant sex differences in the responses to CWS and IMS in the release of the three hormones, although female values trended slightly higher than male values. It has been previously reported that male and female rodents acquire place- and response learning to acute stress at the same rate.29
FUR is clinically used as a loop diuretic in patients with mild to severe congestive heart failure and portal hypertension. Molecularly, it acts as an inhibitor of the KCC that is present in various tissues.30 Administration of FUR (40 mg/kg) significantly mitigated the stress-induced hormonal changes associated with IMS and CWS, although not all the values returned to the levels of the vehicle group. However, the hormonal effects associated with the use of FUR were not significantly translated into enough responses to restore the motor and behavioural changes that were induced by IMS and CWS, with the exception of the hot plate test. This latency in the translation of the response is not clear but can be attributed to two possible reasons. First, a possible explanation might lie in the mechanism of action of the studied hormones, ie, corticosterone and prolactin, as it involves gene expression. Such actions require several minutes and hours to be observed, and a return to near-normal levels requires a longer time than what was implemented in our study. Second, KCCs are distributed in the hypothalamus, pituitary gland, hippocampus, amygdala, and fornix. However, there is a lack of evidence for the functional relationship to the behavioural and biochemical effects of KCCs, and therefore a lack of direct impact of FUR on the motor and behavioural effects.10
It should be mentioned that different drugs may induce different hormonal and behavioural responses in animals and humans subjected to stressful stimuli. For example, the antidepressant drug amitriptyline has been reported to cause differential hormonal and behavioural actions in rats subjected to chronic stress.31
Further studies are warranted to study the neurochemical basis of these effects in specific regions of the brain and to compare the effects of FUR reported here with other sodium potassium chloride cotransporter inhibitors.
The possible clinical implications of the anti-stressor action of FUR warrant further studies. It is of interest to note that FUR is illegally used by competing human athletes and in racing animals.32,33 Whether this is related to the purported anti-stressor action is unknown.
Acknowledgments
The authors are grateful to Professor Gerald Blunden for reading the manuscript. We thank the staff of the Animal House for looking after the animals.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors declare that they have no conflicts of interest in this work.
References
1. Agnati LF, Tiengo M, Ferraguti F, et al. Pain, analgesia, and stress: an integrated view. Clin J Pain. 1991;7(Suppl 1):S23–37. doi:10.1097/00002508-199108000-00005
2. Beery AK, Kaufer D. Stress, social behavior, and resilience: insights from rodents. Neurobiol Stress. 2015;1:116–127. doi:10.1016/j.ynstr.2014.10.004
3. Peters A, McEwen BS, Friston K. Uncertainty and stress: why it causes diseases and how it is mastered by the brain. Prog Neurobiol. 2017;156:164–188. doi:10.1016/j.pneurobio.2017.05.004
4. Biagini G, Pich EM, Carani C, et al. Pyruvic acid, a tryptophan ketoanalogue, antagonizes the endocrine but not the behavioural effects of repeated stress in a model of depression. Biol Psychiatry. 1993;33(10):712–719. doi:10.1016/0006-3223(93)90121-s
5. Harris RB. Chronic and acute effects of stress on energy balance: are there appropriate animal models? Am J Physiol Regul Integr Comp Physiol. 2015;308(4):R250–65. doi:10.1152/ajpregu.00361
6. Bali A, Jaggi AS. Preclinical experimental stress studies: protocols, assessment and comparison. Euro J Pharmacol. 2015;746:282–292. doi:10.1016/j.ejphar.2014.10.017
7. Oláh S, Cservenák M, Keller D, et al. Prolactin-induced and neuronal activation in the brain of mother mice. Brain Struct Funct. 2018;223(7):3229–3250. doi:10.1007/s00429-018-1686-1
8. Ellison DH. Clinical Pharmacology in Diuretic Use. Clin J Am Soc Nephrol. 2019;14(8):1248–1257. doi:10.2215/CJN.09630818
9. Tanis JE, Bellemer A, Moresco JJ, Forbush B, Koelle MR. The Potassium Chloride Cotransporter KCC-2 Coordinates Development of Inhibitory Neurotransmission and Synapse Structure in Caenorhabditis elegans. J Neurosci. 2009;29(32):9943–9954. doi:10.1523/JNEUROSCI.1989-09.2009
10. Kaur A, Bali A, Singh N, Jaggi AS. Investigating the stress attenuating potential of furosemide in immobilization and electric foot-shock stress models in mice. Naunyn Schmiedeberg’s Arch Pharmacol. 2015;388(5):497–507. doi:10.1007/s00210-015-1084-7
11. Beery AK, Zucker I. Sex bias in neuroscience and biomedical research. Neurosci Biobehav Rev. 2011;35(3):565–572. doi:10.1016/j.neubiorev.2010.07.002
12. Zakiniaeiz Y, Cosgrove KP, Potenza MN, Mazure CM. Balance of the sexes: addressing sex Differences in preclinical research. Yale J Biol Med. 2016;89(2):255–259.
13. Krupina NA, Shirenova SD, Khlebnikova NN. Prolonged social isolation, started early in life, impairs cognitive abilities in rats depending on sex. Brain Sci. 2020;10(11):799. doi:10.3390/brainsci10110799
14. Jaggi AS, Bhatia N, Kumar N, Singh N, Anand P, Dhawan R. A review on animal models for screening potential anti-stress agents. Neuro Sci. 2011;32(6):993–1005. doi:10.1007/s10072-011-0770-6
15. Belay T, Woart A, Graffeo V. Effect of cold water-induced stress on immune response, pathology and fertility in mice during Chlamydia muridarum genital infection. Patho Dis. 2017;75(5):ftx045. doi:10.1093/femspd/ftx045
16. Buynitsky T, Mostofsky DI. Restraint stress in biobehavioural research: recent developments. Neurosci Biobehav Rev. 2009;33(7):1089–1098. doi:10.1016/j.neubiorev.2009.05.004
17. Kumar N, Singh N, Jaggi AS. Anti-stress effects of cilnidipine and nimodipine in immobilization subjected mice. Physiol Behav. 2012;105(5):1148–1155. doi:10.1016/j.physbeh.2011.12.011
18. Ali BH, Ziada A, Al Husseni I, Beegam S, Nemmar A. Motor and behavioural changes in rats with adenine-induced chronic renal failure: influence of acacia gum treatment. Exp Biol Med (Maywood). 2011;236(1):107–112. doi:10.1258/ebm.2010.010163
19. Manchanda RK, Jaggi AS, Singh N. Ameliorative potential of sodium cromoglycate and diethyldithiocarbamic acid in restraint stress-induced behavioural alterations in rats. Pharmacol Rep. 2011;63(1):54–63. doi:10.1016/s1734-1140(11)70398-x
20. Ali BH, Bashir AK, Tanira MO. Some effects of Cassia italica on the central nervous system in mice. J Pharm Pharmacol. 1997;49(5):500–504. doi:10.1111/j.2042-7158.1997.tb06831.x
21. Ali BH, Ramkumar A, Madanagopal TT, et al. Motor and behavioural changes in mice with cisplatin-induced acute renal failure. Physiol Res. 2014;63(1):35–45. doi:10.33549/physiolres.932585
22. Fontella FU, Siqueira IR, Vasconcellos AP, Tabajara AS, Netto CA, Dalmaz C. Repeated restraint stress induces oxidative damage in rat hippocampus. Neurochem Res. 2005;30(1):105–111. doi:10.1007/s11064-004-9691-6
23. Wan JJ, Qin Z, Wang PY, Sun Y, Liu X. Muscle fatigue: general understanding and treatment. Exp Mol Med. 2017;49(10):e384. doi:10.1038/emm.2017.194
24. Ranabir S, Reetu K. Stress and hormones. Indian J Endocrinol Metab. 2011;15(1):18–22. doi:10.4103/2230-8210.77573
25. Tsigos C, Kyrou I, Kassi E, et al. Stress: endocrine Physiology and Pathophysiology. Feingold KR, Anawalt B, Boyce A, et al. editors. Endotext [Internet]. South Dartmouth (MA): MDText.com, Inc; 2000.
26. Bali A, Jaggi AS. Angiotensin as stress mediator: role of its receptor and interrelationships among other stress mediators and receptors. Pharmacol Res. 2013;76:49–57. doi:10.1016/j.phrs.2013.07.004
27. Socała K, Nieoczym D, Pieróg M, Szuster-Ciesielska A, Wyska E, Wlaź P. Antidepressant-like activity of sildenafil following acute and subchronic treatment in the forced swim test in mice: effects of restraint stress and monoamine depletion. Metab Brain Dis. 2016;31(5):1095–1104. doi:10.1007/s11011-016-9852-8
28. Lennartsson AK, Jonsdottir IH. Prolactin in response to acute psychosocial stress in healthy men and women. Psychoneuroendocrinology. 2011;36(10):1530–1539. doi:10.1016/j.psyneuen.2011.04.007
29. Lipatova O, Campolattaro MM, Dixon DC, Durak A. Sex differences and the role of acute stress in the open-field tower maze. Physiol Behav. 2018;189:16–25. doi:10.1016/j.physbeh.2018.02.046
30. Delpire E, Weaver CD. Challenges of Finding Novel Drugs Targeting the K-Cl Cotransporter. ACS Chem Neurosci. 2016;7(12):1624–1627. doi:10.1021/acschemneuro.6b00366
31. Cotella EM, Durando PE, Suárez MM. A double-hit model of stress dysregulation in rats: implications for limbic corticosteroid receptors and anxious behaviour under amitriptyline treatment. Stress. 2014;17(3):235–246. doi:10.3109/10253890.2014.910649
32. Cadwallader AB, de la Torre X, Tieri A, Botrè F. The abuse of diuretics as performance-enhancing drugs and masking agents in sport doping: pharmacology, toxicology and analysis. Br J Pharmacol. 2010;161(1):1–16. doi:10.1111/j.1476-5381.2010.00789.x
33. Ali BH, Wong YC, Alhadrami GA, Charles BG, Bashir AK. Plasma pharmacokinetics of intravenous and intramuscular furosemide in the camel (Camelus dromedarius). Res Vet Sci. 1998;64(1):69–72. doi:10.1016/s0034-5288(98)90118-7
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.