Back to Journals » Drug Design, Development and Therapy » Volume 18
The Culprit Behind HBV-Infected Hepatocytes: NTCP
Authors Li S, Hao L, Deng J, Zhang J ![]() , Yu F
, Yu F ![]() , Ye F
, Ye F ![]() , Li N
, Li N ![]() , Hu X
, Hu X
Received 27 May 2024
Accepted for publication 16 October 2024
Published 28 October 2024 Volume 2024:18 Pages 4839—4858
DOI https://doi.org/10.2147/DDDT.S480151
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Yan Zhu
Shenghao Li,1,2,* Liyuan Hao,1,2,* Jiali Deng,1,2 Junli Zhang,3 Fei Yu,1,2 Fanghang Ye,1,2 Na Li,1,2 Xiaoyu Hu2
1School of Clinical Medicine, Chengdu University of Traditional Chinese Medicine, Chengdu, Sichuan Province, People’s Republic of China; 2Department of Infectious Diseases, Hospital of Chengdu University of Traditional Chinese Medicine, Chengdu, Sichuan Province, People’s Republic of China; 3Department of Infectious Diseases, Jiangsu Province Hospital of Chinese Medicine, Nanjing, Jiangsu Province, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Xiaoyu Hu, Department of Infectious Diseases, Hospital of Chengdu University of Traditional Chinese Medicine, No. 39 Shi-er-qiao Road, Chengdu, 610072, Sichuan Province, People’s Republic of China, Email [email protected]
Abstract: Hepatitis B virus (HBV) is a globally prevalent human DNA virus responsible for over 250 million cases of chronic liver infections, leading to conditions such as liver inflammation, cirrhosis and hepatocellular carcinoma (HCC). Sodium taurocholate co-transporting polypeptide (NTCP) is a transmembrane protein highly expressed in human hepatocytes and functions as a bile acid (BA) transporter. NTCP has been identified as the receptor that HBV and its satellite virus, hepatitis delta virus (HDV), use to enter hepatocytes. HBV entry into hepatocytes is tightly regulated by various signaling pathways, and NTCP plays an important role as the initial stage of HBV infection. NTCP acts as an initiation signal, causing metabolic changes in hepatocytes and facilitating the entry of HBV into hepatocytes. Thus, a comprehensive understanding of NTCP’s role is crucial. In this review, we will examine the regulatory mechanisms governing HBV pre-S1 binding to liver membrane NTCP, the role of NTCP in HBV internalization, and the transcriptional and translational regulation of NTCP expression. Additionally, we will discuss clinical drugs targeting NTCP, including combination therapies involving NTCP inhibitors, and consider the safety of NTCP as a therapeutic target.
Keywords: NTCP, HBV, HBV entry, Bulevirtide
Introduction
Hepatitis B virus (HBV) infection is a major global public health challenge. According to data from the World Health Organization (WHO) showed that in 2019, 290 million people were living with chronic hepatitis B (CHB) infection, and 1.5 million new infections were diagnosed each year.1 As a prototype virus of the Hepadnaviridae family, HBV has partial double-linked ring DNA and encodes a variety of functional proteins.2 Chronic HBV infection is a major cause of cirrhosis and hepatocellular carcinoma (HCC).3
Currently, the primary treatments for HBV infection are nucleotide analogues (NAs) and interferons (IFNs).4 Nucleotides and NAs inhibit the reverse transcription of pre-genomic RNA (pgRNA) into HBV DNA but are associated with the risk of drug resistance and often fail to achieve hepatitis B surface antigen (HBsAg) clearance.5,6 Some cccDNA degradation can be achieved by immune-mediated degradation, such as interferon alpha, lymphosin-beta receptor agonists, tumor necrosis factor alpha, and interferon gamma, by upregating apolipoprotein B mRNA editing enzymes to catalyse cellular effectors such as apolipoprotein B mRNA editing enzyme catalytic polypeptide-like 3A/3B (also known as APOBEC3A/3B cytidine deaminase). The cccDNA is deammoniated in the nucleus. In addition, IFNs therapy can regulate innate and adaptive immunity and also affect the activity of HBV covalently closed circular DNA (cccDNA). However, it has the disadvantages of poor tolerance and serious side effects. While these treatments can help control HBV infection, long-term or even lifelong therapy is often required. Patients must also contend with high treatment costs, frequent disease monitoring, and strict adherence to their treatment regimen. Despite treatment, the risk of cirrhosis and liver cancer remains elevated. Therefore, the discovery of new anti-HBV drugs is very important and urgent.
Given the current inability to completely eradicate cccDNA from infected liver nuclei, therapies have been developed to inhibit the entry, assembly and secretion of HBV and to clear cccDNA from infected liver cells.7 In particular, in 2012, the cellular entry receptor for HBV, sodium taurocholate co-transporting polypeptide (NTCP), was discovered. NTCP is a transmembrane bile acid (BAs) transporter expressed in hepatocytes and also serves as the entry receptor for HBV’s satellite virus, HDV.7 This provides new clues to potential treatments for HBV infection.
NTCP plays a crucial role in the entry of HBV into hepatocytes. It serves as a key mediator of the liver’s innate antiviral immune response, and its involvement in various viral infections has been demonstrated through multiple mechanisms.8 During hepatocyte proliferation, the down-regulation of membrane-localized NTCP expression renders newly formed hepatocytes resistant to HBV reinfection. This process may accelerate viral clearance during immune-mediated cell death and the compensatory proliferation of surviving hepatocytes.9 Inhibition of Ntcp has been shown to prevent the spread of HBV-infected hepatocytes in mice.9 NTCP-targeted therapy has potential for regulating HBV infection in patients with CHB.10 Post-translational glycosylation of NTCP is critical for its proper localization to the hepatocyte membrane, which may help explain physiological cholestasis in newborns and also play a significant role in HBV infection after birth.11 In addition, as an important target of HBV entry, NTCP is not only related to new HBV infection, but also closely related to the addition of cccDNA library in HBV-infected hepatocytes.12,13
In this review, we review the complex regulatory mechanisms of NTCP. However, the role of these mechanisms in different conditions remains largely unknown, and further elucidation may lead to the identification of other anti-HBV drugs with novel modes of action.
NTCP and HBV
The Mechanism of HBV Enters Cells
HBV is a pathogen responsible for acute and chronic hepatitis, as well as HCC. It has a circular, partially double stranded DNA genome that selectively enters liver cells and transmits the genome, thus initiating various processes of virus replication.14 The HBV envelope contains three forms of HBsAg, namely large (L), middle (M), and small (S) proteins15 (Figure 1). Notably, the pre-S1 domain of the large envelope protein plays a crucial role in HBV attachment and entry into hepatocytes. And NTCP is the key to pre-S1 binding and HBV infection.16
 |
Figure 1 Schematic diagram of hepatitis B virus. HBV is a partially double-stranded DNA genome of approximately 3.2 kilobases(kb). HBV contains three envelope proteins: large (L), middle (M), and small (S). The C-terminal S domain is common to all three envelope proteins. The preS1, preS2, and S domains are indicated. The viral polymerase is covalently attached to the partially double-stranded DNA genome. The core protein (HBc) forms the capsid of viral particles. |
HBV entry into hepatocytes occurs in several stages. It begins with HBV’s low-affinity interactions with heparan sulfate proteoglycan (HSPG), including glypican-5 (GPC5), on the surface of hepatocytes.17,18 HSPG exists in the extracellular matrix or cell surface of almost all cells19 Studies have shown that highly sulfated inhibitors are more effective at blocking HBV binding to cells.20 Previous studies have found that HSPG is related with HBV cell attachment.20,21 This attachment is facilitated by the interactions of HSPG with the Arg122 and Lys141 residues in the S-domain antigenic ring of all HBV envelope proteins, creating electrostatic interactions with negatively charged HSPG.22 This low-affinity binding stabilizes HBV on the cell surface, allowing the virus to engage with its high-affinity receptors23 After this initial interaction, HBV binds to NTCP, which oligomerizes before the virus enters the cell, following its attachment to HSPG.24 HBV is attached by HSPG, but the expression of NTCP significantly affects HBV internalization. GPC5 is an HBV entry factor, promoting HBV infection. Studies have shown that GPC5 adheres to the surface of HBV particles and facilitates the entry of host cells.18 HBV’s preferential binding to GPC5 may partly explain its strong hepatotropism.18 Once attached to the high-affinity NTCP receptor,25 HBV undergoes endocytosis, leading to further viral infection.26 NTCP is an important receptor in HBV binding and entry (Figure 2). These studies suggest that the initial interaction between HBV and NTCP occurs before the virus enters the cell.27
 |
Figure 2 Diagram of viral entry of hepatitis B virus. HBV first enters the liver with the blood, then enters the lesion space and reaches the sinuses of hepatocytes. HBV virus first binds to HSPG with low affinity on the cell surface. Subsequently, HBV pre-S1 binds to NTCP with high specificity. HBV binding to NTCP can lead to endocytosis of the receptor, and eventually the HBV nucleocapsid enters the nucleus and establishes infection. Created with BioRender.com. |
An Overview of NTCP
NTCP is a member of the SLC10 transporter family and a sodium-dependent transporter of BAs, also known as solute carrier family 10A1 (SLC10A1). Human NTCP (hNTCP) is a protein with a molecular weight of 56kDa and consists of 349 amino acids.28,29 Of course, NTCP is also responsible for transporting BAs to enter hepatocytes. Therefore, competition between BAs and HBV for NTCP binding is an important consideration in future research on NTCP inhibitors. The effect of NTCP inhibitors on Bas is a key factor in evaluating their safety, as significant alterations in BAs levels could lead to hepatotoxicity or other adverse effects. Therefore, careful monitoring of BAs homeostasis is essential to ensure the safe use of NTCP inhibitors. NTCP’s mainly function is to uptake conjugated BAs from plasma into hepatocytes in a sodium-dependent manner, which makes a vital part in the enterohepatic circulation of BAs.30 Moreover, certain hormones, drugs, and combinations of drugs and BAs are also substrates for NTCP uptake.31–33 Some studies have shown that the HBV envelope protein pre-S1 regulates HBV invasion and cell infection by specifically binding to NTCP.34 There are five different papers showing different structures of NTCP in different conformations that give hinds for different proposed transport modes.
The protein NTCP has the following characteristics. In terms of structural characteristics, its overall structure consists of nine transmembrane helices but lacks the first transmembrane helix. The N-terminus is exposed on the extracellular surface and shows similarities and differences compared with bacterial bile acid transporters. It has a panel domain composed of TM1, TM5, and TM6 and a core domain composed of TM2-TM4 and TM7-TM9. Regarding transport mode hints, when binding with Fab YN69083, the formed complex structure provides atomic-level details for studying the interactions between NTCP, HBV, and bile acids, indicating specific conformational changes in the N-terminal region of NTCP upon antibody binding. For bile acid transport, through mutant studies, key sites related to bile acid transport such as the cross-region of TM3 and TM8 and amino acid residues like Leu27, Leu31, and Leu35 have been identified. A structure-based transport mechanism is proposed where bile acid molecules are co-transported with two sodium ions, and conformational changes of NTCP may play an important role. In terms of interaction with HBV, it interacts with the myristoylated N-terminal preS1 domain of the large surface protein of HBV. Amino acid residues at positions 9–18 of this domain are crucial for binding to NTCP. The infection mechanism is that HBV induces viral internalization and infection through the binding of preS1 to NTCP, while compounds such as Bulevirtide can inhibit this process.35
The protein NTCP exhibits the following characteristics. In terms of structural features, it adopts the SLC10 fold structure, comprising nine transmembrane α-helices and having two domains, core and panel. When binding with nanobodies, NTCP undergoes conformational changes to form a wide transmembrane pore that serves as the transport route for bile salts. The important sodium binding sites (Na1 and Na2) and substrate binding-related sites are structurally conserved, and some mutations can affect the transport function of NTCP. Regarding the transport mode hints, in the transport process, NTCP undergoes a conformational change from an inward-facing state to an open pore state, which is caused by the relative movement of the core and panel domains. The open pore structure indicates that NTCP can transport various amphipathic bulky substrates including bile salts, sulfated steroids, and statins, and the substrate interacts with amino acid residues within the transmembrane pore during transport. In terms of receptor recognition for HBV/HDV, the receptor binding domain preS1 of HBV/HDV interacts with amino acid residues within the transmembrane pore of NTCP. The open pore state of NTCP is conducive to the binding of preS1, while the inward-facing state inhibits the binding of preS1. This mechanism explains why myr-preS1 binding inhibits bile salt transport and why certain mutations of NTCP can affect HBV infection and bile acid transport.36
The overall structure of hNTCP in complex with Fab fragments, nanobodies and substrates was resolved by cryo-electron microscopy. It has a tunnel structure connecting the extracellular environment and cytoplasm. The external part of the tunnel is partially open to the outer leaflet of the membrane, while the internal part is open to the inner leaflet of the membrane and cytoplasm. Substrate molecules (such as glycochenodeoxycholic acid) are arranged in parallel in the tunnel, and their hydrophilic tails can move in the hydrophilic part of the tunnel. In terms of transport mechanism, NTCP co-transports bile acid molecules with two sodium ions. Based on conformational changes during transportation, a transport mechanism is proposed, explaining the stoichiometry of sodium ions and bile acid transportation. The broad specificity of NTCP for bile salts can be explained by the binding mode and conformational changes of substrate molecules in the tunnel. The disorder of the glycine part enables NTCP to bind different bile acid molecules. The research results provide clues for understanding the transport mechanisms of human transport proteins ASBT and SOAT, and suggest that NTCP, ASBT and SOAT may share a common transport mechanism.37
The cryo-electron microscopy structures of human, bovine and rat NTCPs in the absence of substrates and the structure of hNTCP in the presence of the myristoylated preS1 domain of hepatitis B virus envelope protein LHBs are reported. NTCP has nine transmembrane helices. The panel domain is composed of TM1, TM5 and TM6, and the core domain is composed of TM2-TM4 and TM7-TM9. There are some special regions between transmembrane helices, such as the crossover region of TM3 and TM8, which is of great significance for sodium binding and substrate transportation. Substrate transportation occurs through transmembrane tunnels. Lipid-like densities in the tunnel suggest that this region may be part of substrate transport. PreS1 and substrate compete for the extracellular opening of the NTCP tunnel. This competitive binding mode explains why there is mutual interference between HBV infection and bile acid transport. The key binding sites for HBV preS1 are determined, such as amino acid residues in regions such as the TM2-TM3 loop and TM5. The binding of preS1 to NTCP forms a lasso-like structure, in which the N-terminal region folds into the tunnel and forms extensive interactions with amino acid residues in the tunnel, and the C-terminal region binds to the extracellular surface region.38
The complex structure of hNTCP and myristoylated preS1 (2–48) peptide segment was analyzed. In this complex, NTCP and preS1 form a ternary complex with a ratio of 1:1:1. After NTCP binds to preS1, the overall conformation is similar to that in the ligand-free state, but conformational changes occur in some regions. For example, the conformational change of TM5 is closely related to the binding of preS1 and virus infection. The N-terminal region (amino acids 2–34) of preS1 fills the extracellular opening of the NTCP tunnel and forms multiple hydrogen bonds and hydrophobic interactions with amino acid residues in the tunnel. The C-terminal region (amino acids 35–48) of preS1 interacts with the extracellular surface region of NTCP (such as the TM2-TM3 loop), including hydrogen bonds and hydrophobic interactions. This study reveals the mechanism of NTCP as an HBV receptor. That is, preS1 induces the attachment and infection of the virus to the host cell through specific binding to NTCP. It also provides a structural basis for the development of anti-HBV drugs targeting the HBV entry step and identifies some potential drug targets, such as amino acid residues Q264, T268, and V272 on TM8b.39
The Regulatory Factors of HBV Entry into Hepatocytes and Its Relationship with NTCP
However, NTCP alone may not be sufficient for the effective internalization of HBV into hepatocytes. Other regulatory factors likely contribute to HBV susceptibility, suggesting a complex, multi-step entry process (Table 1).
 |
Table 1 The Regulatory Factors of HBV Entry into Hepatocytes |
The entry of HBV into liver cells begins with the low-affinity binding of the pre-S1 domain on HBV’s surface to HSPG on hepatocyte membranes.18,21,22 Low density lipoprotein receptor (LDLR) is a known apoE-binding receptor like HSPGs and plays an important role in HBV infection.51 LDLR plays a key role in lipid and cholesterol metabolism by binding to very low-density lipoprotein (VLDL), apolipoprotein E (ApoE), and high-density lipoprotein (HDL) particles on ApoB100 on LDL and chylomicrons. Studies have shown that knocking down of LDLR significantly reduces HBV infection.40 The enrichment of human ApoE on the HBV envelope is vital for the effective infection and production of HBV.52,53 In fact, strong evidence links HBV to ApoE, as specific ApoE antibodies can block HBV infection and capture HBV particles.52 Interestingly, patients infected with HBV who carry the ApoE3 allele have lower rates of HBsAg clearance.41,42 Viral envelope proteins play a key role in mediating cell entry.
The high expression of beta2-glycoprotein I (beta2-GPI) has been shown to promote the transfer of viral particles to NTCP, and its interaction with annexin II facilitates viral membrane fusion.43 Endothelial lipase (LIPG) has been reported to enhance HBV adherence to HSPG or promotes HBV adherence to NTCP.44 The introduction of SLC10A1 by mRNA transfection increases the sensitivity of hepatoma cells to HBV. Conversely, transfection of SLC10A1 mRNA into non-hepatocytes can support the uptake of BAs, but still does not make the cells susceptible to HBV, suggesting the need for additional host factors.54
Next, the pre-S1 structural region of the HBV envelope binds to NTCP.16,45,55 Knocking down of EGFR has been shown to reduce both HBV internalization and NTCP expression.45 Additionally, activation of EGFR triggers time-dependent relocalization of HBV preS1 to early and late endosomes and lysosomes in coordination with EGFR trafficking. Inhibition of EGFR ubiquitination by site-directed mutagenesis or by knockdown of two EGFR sorting molecules, the signal-transducing adaptor molecule (STAM) and lysosomal protein transmembrane 4β (LAPTM4B), indicates that EGFR trafficking to late endosomes is crucial for efficient HBV infection.46 Studies have shown that knocking out of EGFR significantly impairs NTCP transport to the cell surface.47
Interferon-induced transmembrane protein 1–3 (IFITM1-3) is a limiting factor for the entry of many viruses.48 The upregulation of IFITM3 may be an adaptive upregulation during the process of HBV infecting hepatocytes. Knocking out of IFITM3 significantly inhibits HBV infection in NTCP-expressing in HuH7 and PHH cells, which proves from the side that IFITM3 is conducive to HBV infection.48 IFITM expression is regulated by interferon stimulated response elements (ISRE).56,57 A proteomic study has shown that FITM3 is one of the most upregulated proteins during HBV infection.49 In addition, IFITM3 expression increases when hepatocytes are exposed to myr-pre-S1 lipopeptides that mimic HBV/HDV binding to NTCP.8
ATP5B, the beta subunit of the F1 component of F1F0 ATP synthase, is responsible for most ATP synthesis in many organisms.58 Establish ATP5B knockdown cells to test the importance of ATP5B for the life cycle of HBV. Lower ATP5B expression is associated with lower production of HBeAg and HBsAg. Total HBV DNA and cccDNA are significantly reduced. Studies have shown that knocking out of ATP5B reduces HBV infectivity.59
SLC10A1 Mutation
HBV only infects humans and chimpanzees, posing major challenges for modeling HBV infection and chronic viral hepatitis.50 Studies have shown that the expression of NTCP in liver organs is significantly increased, and infection with HBV in vitro differentiated organoids leads to higher levels of infection and replication. Because high expression of NTCP alone does not result in more effective HBV infection, NTCP by itself may not be sufficient to facilitate optimal viral entry.60 The natural course of hepatitis virus infection in woodchucks closely resembles that of HBV infection in humans. However, woodchuck hepatocytes expressing hNTCP are only susceptible to HDV infection, indicating that other key factors mediate HBV infection, which are subject to strict species-specific limitations. This suggests that while hNTCP is essential for HDV entry, HBV infection likely requires additional human-specific factors, suggesting that while hNTCP is crucial for HDV entry, HBV infection likely requires additional human-specific factors. These factors may include specific co-receptors, host proteins involved in viral replication, or immune evasion mechanisms that are absent or function differently in woodchucks, leading to species-specific limitations on HBV infection.61 Studies have shown that exogenous expression of hNTCP has established a robust model of HBV infection in rhesus monkeys.62 SLC10A1 gene mutations also affected the efficiency of HBV infection. Detection of the primary protein sequence of human NTCP shows that 139YIYSRGIY146 is a highly conserved tyrosine-rich motif. To study the roles of amino acids Y139, Y141, and Y146 in NTCP biology, site-directed mutagenesis was used to replace the above residues with alanine, phenylalanine, or glutamic acid (simulating phosphorylation). Only Y141E has a transport defect, most likely due to the accumulation of mutant proteins intracellularly. Most importantly, the Y146A and Y146E mutations completely eliminate the binding of the viral preS1 peptide to NTCP, while the Y146F mutant of NTCP shows some residual binding ability to preS1. Therefore, tyrosine 146 and tyrosine 141 both belong to the tyrosine-rich motif 139YIYSRGIY146 in human NTCP to a certain extent. They are newly discovered amino acid residues that play an important role in the interaction between HBV and its receptor NTCP, and thus play an important role in the process of virus entry into hepatocytes.63 The NTCP S267F variant of the SLC10A1 gene provides protection against both HBV and HDV infection, significantly reducing the risk of developing cirrhosis and HCC64,65 In patients with CHB, the p.Ser267Phe variant serves as a protective factor, lowering the risk of liver failure, cirrhosis, and HCC.66 The rs4646287 G > A genotype of the SLC10A1 gene may be linked to a reduced risk of HBV vaccine failure in children born to highly infectious mothers.67 The SLC10A1 p.Ser267Phe variant has also been associated with a faster anti-HBV response to first-line nucleoside analogues.68 Additionally, species-specific differences and mutations in the SLC10A1 gene influence the efficiency of HBV infection.
Regulation of NTCP
NTCP plays a critical role in BAs transport and HBV binding, making its regulation at both the transcriptional and translational levels crucial for BA metabolism and HBV-related diseases. NTCP expression is regulated by many factors (Figure 3).
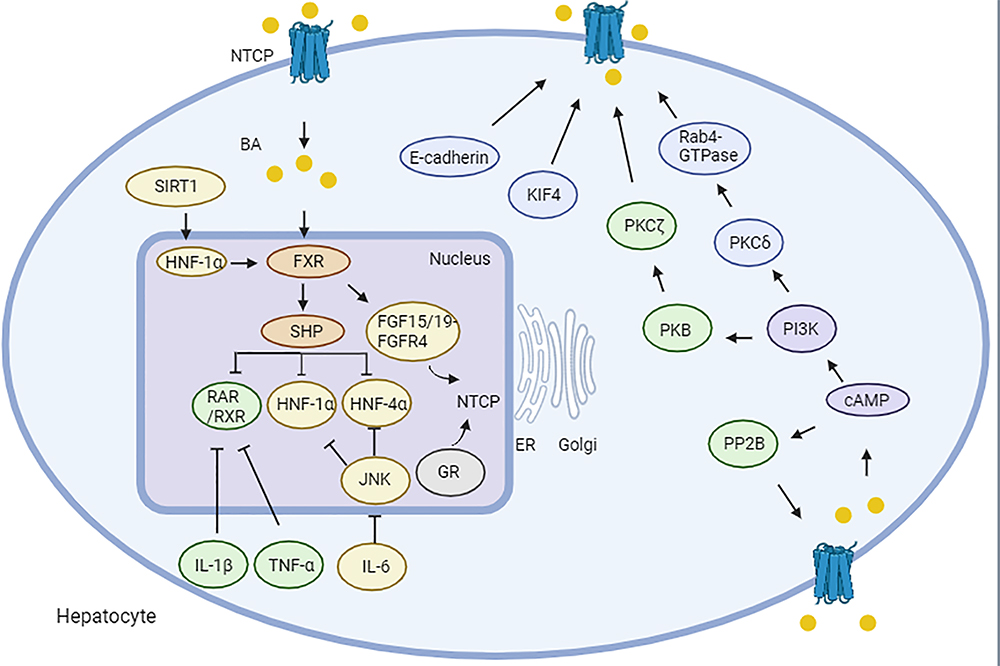 |
Figure 3 Regulation of sodium taurocholate co-transporting polypeptide expression. BA-induced, FXR-mediated induction of nuclear suppressor SHP is a key mechanism to reduce NTCP expression by interfering with RXR, RAR, HNF-1α and HNF-4α with binding sites within the NTCP promoter. SIRT1 regulates FXR expression through HNF-1α. FXR further regulates the expression of NTCP mRNA through the FGF15/19 signaling pathway inhibits the down-regulation of NTCP by HNF1α and HNF4α via JNK-dependent pathway. And IL-1β and TNF-α down-regulated the expression of NTCP through inhibition of RAR/RXR complex. cAMP stimulates Ca2+ or PP2B to dephosphorylate NTCP. After cAMP activation, the PI3K/PKB/PKC isomers jointly promote the intracellular movement of NTCP into the plasma membrane. Other factors, including KIF4, E-cadherin, that promote the transport of NTCP to the membrane. Among them, yellow represents mouse, green represents rat, blue represents human, Orange represents mouse and rat, and purple represents rat and human, showing the effect. Created with BioRender.com. Abbreviations: BA, bile acid; FXR, farnesoid X receptor; SHP, small heterodimer partner; HNF-1α, hepatocyte nuclear factor 1 alpha; SIRT1, hepatic sirtuin 1; RXR, retinoid X receptor; RAR, RXR-retinoic acid receptor; PP2B, protein phosphatase 2B; IL-6, interleukin 6; TNF-α, tumor necrosis factor-alpha. |
Transcriptional Regulation of NTCP
The transcriptional regulation of NTCP is mediated by BAs, nuclear receptors and nuclear factors. Other factors, such as hormones and cytokines, also regulate the expression of NTCP. NTCP mRNA regulation is primarily associated with BA concentration. Farnesoid X receptor (FXR), a liver-enriched nuclear receptor activated by BAs, forms a heterodimer with retinoid X receptor alpha (RXR-α) to recognize response elements.69 FXR activation by chenodeoxycholic acid (CDCA) increases the synthesis of pgRNA and DNA replication intermediates in the HBV genome.70,71 FXR binds to two response elements in the HBV core promoter region, which are activated by ligands to modulate HBV core promoter.70,71 However, in HBV transgenic models, FXR only participates to a limited extent in HBV biosynthesis.72 Although FXR activation regulates SLC10A1 expression, FXR does not directly interact with the SLC10A1 promoter. Instead, it induces the expression of small heterodimer partner (SHP), which inhibits the activation of SLC10A1 gene by retinoic acid receptor (RAR) and retinoid X receptor (RXR) response elements in rat.73 FXR also modulates Ntcp through the FGF15/19 signaling pathway in mice.74 Other key regulators of BA metabolism include HNF-1α, HNF-4α and HNF-3β. HNF-1α binds to and activates Ntcp promoters in rat, while HNF-3β regulates transcriptional inhibition of NTCP/Ntcp promoters by directly binding to response elements in all three species.75 HNF-1α regulates the expression of FXR in mice,76 and the absence of Sirt1 has been shown to reduce HNF-1α binding to the FXR promoter, disrupting BA metabolism.77 HNF-4α directly binds to Slc10a1 promoters via HNF-4α response elements in mouse, with HNF-4α-induced Slc10a1 activation further enhanced by proliferator-activated receptor-γ coactivator-1α (PGC-1α).78 Over-expression of NTCP has been shown to inhibit adenosine production and HIF1α expression.79
As a ligand-activated transcription factor, the glucocorticoid receptor (GR) binds to Slc10a1 promoter gene, downregulating its expression and reducing NTCP-mediated BAs transport in GR-deficient mice.80 Other hormones, including growth hormone, prolactin, thyroid hormone and estrogen, also play significant roles in regulating the expression of Ntcp and maintaining its homeostasis. The maximum secretion rate of taurocholate also increased by 10%, 31%, and 24% on days 2, 14, and 21 postpartum, respectively, which is associated with increased mRNA and protein expression of Ntcp and Bsep. Infusion of ovine prolactin (oPRL) to ovariectomized rats increases the mRNA and protein expression of Ntcp and Bsep. These data indicate that the coordinated expression of bile salt transporters postpartum and when affected by prolactin is increased.81 Prolactin promotes the binding of Stat5 to Slc10a1-GLEs through the long form prolactin receptor PRLRL, thereby transcriptionally regulating Slc10a1.82 In addition, studies have confirmed that prolactin (PRL) and placental prolactin can induce Slc10a1 expression through the prolactin receptor and the signal transducer and activator of transcription (Stat) 5a pathway. However, in pregnant rats, elevated levels of placental prolactin do not increase the expression of Slc10a1. Since plasma estradiol (E₂) levels also increase during pregnancy, there is an inhibitory effect of E₂ on PRL-induced Slc10a1 activation. E₂ treatment can inhibit the increase in liver Slc10a1 induced by PRL to the same level as in rats treated with E₂ alone. In HepG2 cells, in the presence of co-transfected ERα, E₂ inhibits PRL-induced SLC10A1 reporter gene expression in a dose-dependent manner.83 Studies have shown that both thyroid hormones and glucocorticoids can directly induce the level of Slc10a1 mRNA without being affected by the secretion pattern of growth hormone. However, the effect of estrogen in reducing Slc10a1 mRNA is not due to a direct effect on the liver but requires the stable secretion of pituitary hormones, especially growth hormone.84 Estradiol inhibits the expression of NTCP, thereby inhibiting viral entry, limiting infection, and spreading in the liver. In some cases, women may have a stronger immune response to HBV than men,85 which may have a potential impact on the progression of HBV from acute to chronic infection. Cytokines play a crucial role in the immune response to HBV. During acute infection, their balance is important for virus clearance. Pro - inflammatory cell factors such as IFN-γ and tumor necrosis factor-α (TNF-α) can activate immune cells to clear infected cells. However, if the balance is disrupted, the immune response may be ineffective.86 High levels of interleukin-6 (IL-6) during chronic inflammation or due to other factors may lead to immune dysregulation. Chronic elevation of cytokines can cause immune exhaustion, making acute infection likely to turn into chronic infection.87
During cholestasis, the chronic pathologic elevation of BAs concentrations triggers the release of cytokines such as IL-6, IL-1β, and TNF-α, which inhibit SLC10A1 transcription. IL-6 is an important cytokine that plays a role in inflammation and liver regeneration.88 The expression of NTCP is regulated by IL-6.89 The regulation of NTCP expression by IL-6 has been mainly studied in rodents. These studies show that the down - regulation of Slc10a1 can be mediated by activating c-Jun-N-terminal kinase (JNK), resulting in the decreased expression and binding activity of multiple nuclear factors such as HNF1α, HNF4α and nuclear receptor dimer RXR.90 The transcriptional regulation of the human and mouse NTCP/Ntcp genes was compared with that of rats. HNF1α, HNF4α as well as the retinol X receptor/retinoic acid receptor dimer (RXRα/RARα) bind and activate the rat NTCP/Ntcp promoter but do not bind and activate the human or mouse NTCP/Ntcp promoters. Conversely, the activation of CCAAT/enhancer-binding protein-β is specific to human and mouse NTCP/Ntcp. The only common motif present in these three species is HNF3β. In electrophoretic mobility shift assays, HNF3β forms specific DNA-protein complexes and inhibits NTCP/Ntcp promoter activity in co-transfection analyses.91 In rats, inactivation of TNF-α completely prevents the downregulation of Ntcp and reduces the binding activity of HNF-1. In endotoxemia, the downregulation of Mrp2 and partial downregulation of Ntcp can be prevented by blocking IL-1β but not by blocking TNF-α.92 In summary, the expression of NTCP/Ntcp is regulated by BAs, hormones and cytokines.
There are studies on whether HBV infection is partially determined by diet. Research on HBV transgenic mice fed different levels of casein diets showed that reducing casein to 6% significantly inhibited HBV expression. Serum HBsAg concentrations differed among different casein diets. Low casein diet is an inhibitor of HBV transgene and liver injury. This suggests that a low casein diet is an effective inhibitor of HBV transgene and HBV-induced liver injury, indicating that diet management could be a practical way to help control HBV infection.93 Epidemiological observations show chronic alcoholics have more HBV markers. In HBV transgenic SCID mice, ethanol diet increased HBsAg and viral DNA levels. Changes were observed in liver antigens. However, no such changes were observed in transgenic mice on an isocaloric diet.91
Translational Regulation of NTCP
NTCP localization and membrane expression are regulated by various factors, including cyclic adenosine monophosphate (cAMP), phosphoinositide-3-kinase (PI3K), KIF4 and E-cadherin.94 It has been found that cAMP promotes protein phosphatase 2B (PP2B) or Ca2+ to dephosphorylate Ntcp in rat.95 cAMP leads to dephosphorylation of Ntcp and translocation to the plasma membrane through the Ser-226 site of Ntcp cytoplasm in rat.96 This finding raises questions about the precise localization of NTCP on the basolateral membrane. Tyr-321 and Tyr-307 in the C-terminal cytoplasmic domain of NTCP have been implicated in sorting NTCP to basolateral membrane.97 Moreover, two Asn residues (N5, N11) are believed to be crucial for NTCP expression on the plasma membrane, though there is some controversy surrounding this claim in HepG2.98 While the complete molecular mechanisms governing NTCP localization remain unclear, these Tyr and Asn residues likely play a role in basolateral membrane sorting via interaction with specific host signaling pathways.
The PI3K signaling pathway, including three downstream effectors (P70 S6 kinase [p70S6K], PKC and protein kinase B [PKB]), also influences cAMP-mediated Ntcp translocation in isolated rat hepatocytes.99 Activation of PKB, for example, has been shown to promote Ntcp translocation into the plasma membrane in rat.100 However, only a subset of PKC isoforms affect this process. Specifically, PKCζ promotes cAMP-induced Ntcp plasma membrane localization,101,102 while PKCδ mediates Ntcp transport by activating Rab4 and an endosomal marker in HuH-Ntcp cells.103 The activation of PKCδ stimulates Ntcp localization to the plasma membrane.103 Upon cAMP activation, the PI3K/PKB/PKC pathways work together to facilitate the intracellular movement of Ntcp to the membrane. Additionally, elevated BA concentrations in liver cells expedite Ntcp recovery on the basolateral membrane, involving PKC, Src family kinases (Yes and Fyn), and reactive oxygen species and reactive oxygen species.104–106 Furthermore, phosphodiesterase-induced cAMP degradation restricts NTCP expression on cells, thereby limiting HBV infection.107
Kinesin family member 4 (KIF4), a highly conserved member of the kinesin family,108–110 plays a critical role in NTCP expression. KIF4 is elevated in HBV-related liver malignancies, and its loss reduces surface NTCP levels in HepG2-hNTCP.111,112 Interestingly, RXR agonists like bexarotene and alitretinoin have been found to reduce KIF4 expression.112
E-cadherin is vital for desmosomes and adhesive junctions113,114, and it has implications for pathogen infections, including HBV. Overexpression of E-cadherin has been shown to promote viral transmission.115,116 Adhesion junction proteins (nectins and cadherins) are involved in host-pathogen interactions. Nectins are entry receptors for several major human viruses such as herpes simplex virus, measles virus and poliovirus.117 Many pathogens also target cadherins to enter host cells and survive in them. A study involving siRNA-mediated E-cadherin silencing in Huh7.5.1 cells (a liver cell-derived cancer cell line) showed that the entry of HCV pseudoparticles was inhibited, indicating that E-cadherin plays an important role in mediating host-HCV interaction and regulating virus entry.117 In addition, viral protein U (a viral pore protein important for the viral transmission and pathogenesis of human immunodeficiency virus type 1 and simian immunodeficiency virus) is reported to interfere with the interaction between E-cadherin and β-catenin in addition to down-regulating E-cadherin expression.118 This leads to the destruction of the E-cadherin-β-catenin complex and thus the release of viral particles. It also facilitates HBV infection by promoting the localization of glycosylated NTCP on the cell surface.119 E-cadherin may interact with the cellular surface NTCP-HBV complex,119 and silencing of E-cadherin reduces HBV pre-S1 binding and internalization in primary human hepatocytes and HepG2-NTCP cells.119
NTCP as a Target of Drug Development
Viral entry is the first step in viral infection and is a potential target for the development of anti-viral drugs. NTCP has the potential to be a therapeutic target for HBV. Currently, there are many drugs that target NTCP, such as Bulevirtide, cyclosporin and its derivatives, Thiazolidinediones and their derivatives, BAs derivatives, antibodies and natural products (Table 2).
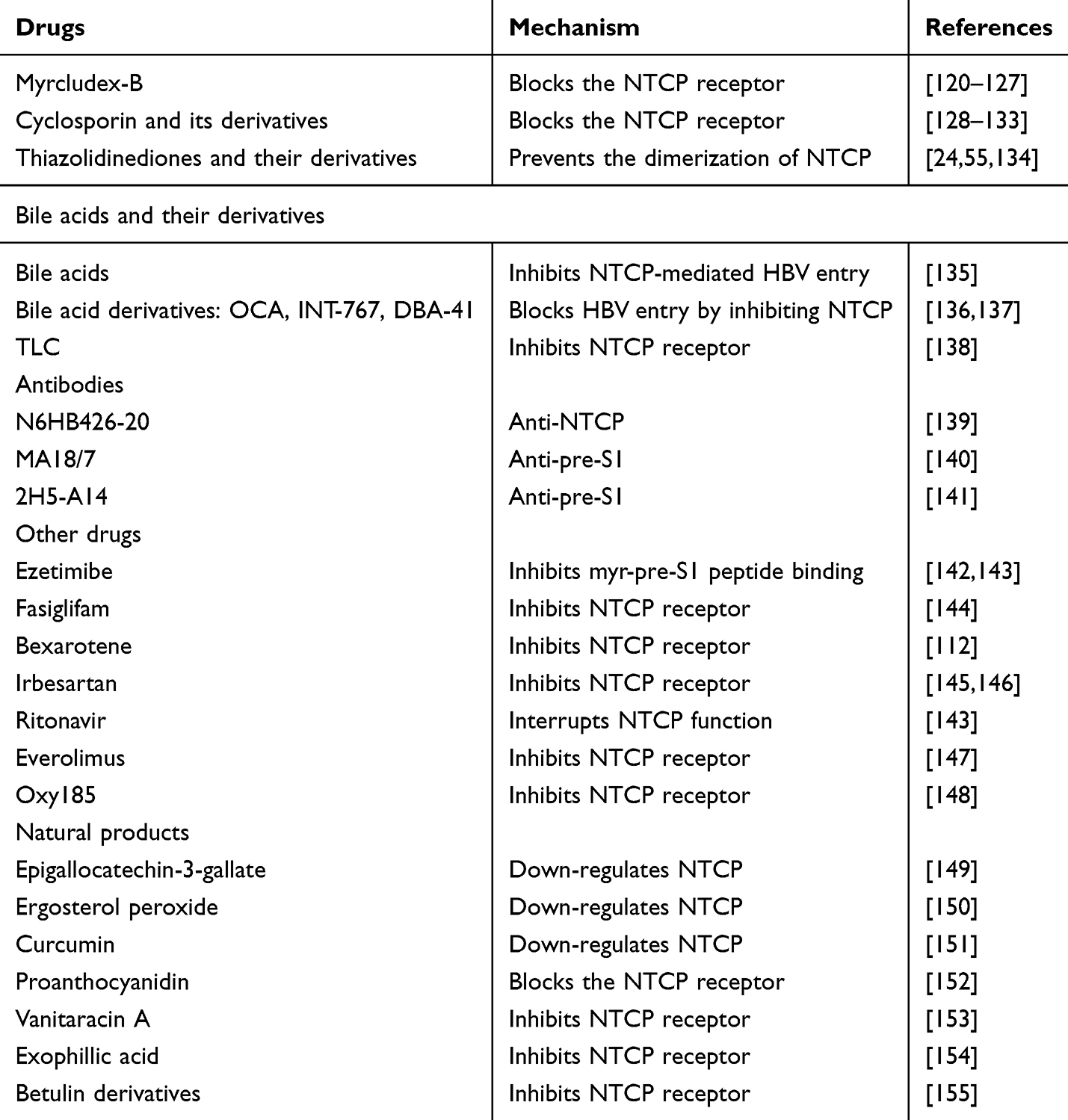 |
Table 2 Examples of Drugs and Natural Products Targeting NTCP |
Clinical study
Bulevirtide
Bulevirtide (Myrcludex B) has been marketed in Europe since 2020 as a treatment for HDV.156,157 As a drug to treat HBV, bulevirtide still in the Phase II clinical trial stage,156 is a potential drug for Phase III clinical trials.158 Bulevirtide is a nutmeg acylating peptide synthesized from 2–48 amino acids of pre-S1.159 Bulevirtide has demonstrated significant efficacy in reducing HBV infection in humanized mice.160 In Phase I/II clinical trials, it exhibited strong anti-viral activity against HDV in humans.120,121 Bulevirtide analogs (2a, 2b, and 2d) have also effectively reduced HBV infection in HepG2-NTCP cells.122 Recent advancements in structural biology have provided detailed cryo-EM structures of NTCP bound to either preS1 or Bulevirtide, offering new insights into how NTCP functions as an HBV receptor and how Bulevirtide inhibits both BAs transport and HBV particle entry.161 Bulevirtide interacts with NTCP in a highly specific manner, forming two distinct domains: a plug that occupies the bile salt transport tunnel of NTCP and a string that covers its extracellular surface. The N-terminally attached myristoyl group of Bulevirtide interacts with the lipid-exposed surface of NTCP. These structures reveal how Bulevirtide inhibits bile salt transport, rationalizes NTCP mutations that decrease the risk of HBV/HDV infection, and provides a basis for understanding the host specificity of HBV/HDV. Such insights not only elucidate the molecular mechanisms by which Bulevirtide works but also pave the way for novel HBV-targeted drug development.35 To disrupt the HBV-NTCP interaction without impairing BAs uptake, targeting the exposed hydrophobic patches between Asn87 and Glu277 could be effective. These specific residues, such as Ile88, Pro281, and Leu282, were chosen because they are located within the critical regions responsible for HBV binding, while maintaining NTCP’s bile acid transport function. Their hydrophobic nature makes them ideal candidates for selective disruption of HBV attachment without compromising BAs transport.35 Approximately 950 Ų of the NTCP surface area is buried in the formation of the Fab complex. There are two interaction clusters centered on Glu277 of the receptor and Tyr104 of the Fab heavy chain. Glu277 of NTCP is located on the loop between TM8 and TM9. It forms a hydrogen bond with the side chain of light chain Arg95, while its carbonyl oxygen obtains a hydrogen bond from the side chain of Trp90. At the same time, the indole ring of tryptophan is located on the side chain of NTCP Glu277. At about 15 Å, Tyr104 of the heavy chain is located near Asp24 and Asn209 and forms hydrophobic interactions with the side chains of Lys20, Phe283 and Phe284. By filling the pocket between the N-terminal regions of TM1 and TM9, Tyr104 of the heavy chain completely prevents the movement of the panel domain relative to the core. As mentioned above, because this closes the pocket. Therefore, this antibody seems to have strict specificity for the outward-open form of NTCP.35
Clinical studies have confirmed the efficacy of Bulevirtide both as a monotherapy and in combination with other antiviral drugs for treating HBV and/or HDV infections.123,124 As a satellite virus of HBV, HDV relies on the envelope protein of HBV for assembly. In fact, HDV and HBV share the envelope protein and use the same cell entry factors. Bulevirtide has shown a differentiated impact on BAs transport and viral entry: while a high dose (47 nM) affects bile salt transport, a much lower dose (IC50 of 80 pM) is sufficient to inhibit HBV and HDV entry.125 Blocking NTCP as a receptor for HBV and/or HDV can be achieved without blocking the function of the NTCP transporter.126 However, this is a mechanistic view based on in vitro data, which is not the case in patients receiving Bulevirtide, and more animal or human studies are needed to confirm this conclusion. The IC50 of Bulevirtide inhibiting NTCP was 4 nM. Notably, the IC50 for inhibiting HBV infection (about 100 pM) and bile salt transport (about 5 nM) varied widely, which is related to the observed binding saturation that does not require NTCP to inhibit infection.162 A long-term clinical study involving the administration of high-dose Bulevirtide (10 mg) over 48 weeks demonstrated its safety and efficacy. In this study, HBV DNA and HBV RNA levels became undetectable, and HDV RNA levels gradually decreased, with two patients achieving complete viral clearance, and a third patient showing a significant reduction in HDV RNA (from 6.8 log10 cp/mL to 500 cp/mL). Despite a significant increase in BAs, the patients remained asymptomatic, further supporting the safety profile of high-dose Bulevirtide.127 In addition, Slc10a1−/− mice are alive and show few NTCP-specific abnormalities.163 In a case study, a person with high bile salt levels was diagnosed with functional knockdown of NTCP but did not show obvious clinical symptoms.164 Moreover, Bulevirtide in combination with pegylated interferon alpha (Peg-IFN-alpha) has shown enhanced synergistic effects on HDV and HBV. This suggests that Peg-IFN-alpha and Bulevirtide have a synergistic effect. However, clinical follow-ups have revealed a common issue of viral rebound upon discontinuation of treatment.120 Given the persistent global burden of chronic HBV/HDV co-infections, these results suggest that Bulevirtide holds significant potential as a therapeutic option for HBV and HDV treatment, though strategies to manage viral rebound will be essential for sustained long-term outcomes. However, more clinical data are needed to demonstrate the safety and efficacy of Bulevirtide.
Basic study
Cyclosporin and Its Derivatives
Cyclosporin A, derived from fungal metabolism, is used as a very effective immunosuppressant. Cyclosporin compounds block the interaction between HBV preS1 and NTCP, inhibiting HBV entry. Studies have shown that Cyclosporin A blocks BAs uptake through NTCP transport,128 and blocks the binding of HBV pre-S to NTCP.129,130 In order to eliminate the negative effects of immunosuppression and BAs uptake inhibition, a series of Cyclosporin derivatives have been screened.131 Cyclosporin A derivatives SCYX827830 and SCYX11454139 not only have no immunosuppressive activity, but also have stronger anti-HBV activity in HepG2-NTCP cells.130 Another Cyclosporin A derivative 27A also inhibits HBV infection without causing immunosuppression.132 In addition, the Cyclosporin derivatives SCY450 or SCY995 inhibit HBV entry without effect on NTCP transporter activity. SCY446, on the other hand, was able to inhibit both HBV entry and NTCP transporter function.131 It has also been confirmed that Cyclosporin B is a stronger HBV entry inhibitor.133 However, the immunosuppressive nature of Cyclosporin A may limit its utility as an anti-HBV therapeutic.
Thiazolidinediones and Their Derivatives
Troglitazone is an insulin sensitizer for the treatment of insulin resistance in type 2 diabetes.165 Recently, troglitazone has been identified as a potential inhibitor of HBV entry by targeting NTCP. Troglitazone inhibits HBV internalization and NTCP oligomerization.24,55,134 Although troglitazone disrupts NTCP oligomerization, it does not impair the interaction between NTCP and EGFR. After HBV attachment, NTCP first binds to EGFR, followed by its oligomerization. Troglitazone prevents NTCP oligomerization, and phenylalanine at NTCP 274 is essential for this process. The formation of a complex involving HBV virions, EGFR, and NTCP oligomers, followed by EGFR autophosphorylation, enables HBV internalization via endocytosis. Therefore, NTCP-EGFR interaction is required for NTCP oligomer formation, and NTCP oligomerization downstream of this association is essential for HBV entry. Administration of troglitazone and pioglitazone significantly reduces the levels of HBsAg without inducing significant cytotoxic effects. Moreover, thiazolidinedione derivatives 1, 2, and 6 inhibit NTCP oligomerization and HBV infection in HepG2-NTCP cells.24 In conclusion, the discovery of thiazolidinediones and their derivatives provides a better state strategy for inhibiting HBV infection.
BAs and Their Derivatives
In addition, multiple substrates of NTCP competitively reduce NTCP-mediated HBV entry. BAs such as tauroursodeoxycholate, taurocholate, bromosulfophthalein competitively reduce NTCP-mediated HBV entry.135 Among BAs derivatives, obeticholic acid (OCA) and INT-767 inhibit HBV entry through reducing the expression of NTCP.136 It has been found that dimeric BA derivatives (DBADs) synthesized from ursodeoxycholic acid (UDCA), have also been found to effectively inhibit NTCP function. One potent compound, DBA-41, with high selectivity, affinity, and bioavailability for human NTCP, was optimized for better efficacy.137 Intraperitoneal injection of hNTCP-TG induces an increase in serum total BAs.137 In addition, secondary BAs taurolithocholic acid (TLC) significantly inhibits the transport function of NTCP, but NTCP protein still exists on the plasma membrane. Preincubation with TLC also significantly reduces HBV/HDV myr-preS1 peptide binding, thereby reducing HDV infection.138 Therefore, BAs and their derivatives have the potential to become new therapeutic agents against HBV infection.
Antibodies
Several monoclonal antibodies have been developed that target the pre-S1 region and inhibit HBV entry. The N6HB426-20 antibody, which recognizes the extracellular domain of human NTCP, effectively blocked HBV entry in vitro but had a minimal effect on BAs uptake. It also induced HBV viremia without strongly inhibiting BA absorption.139 In addition, the MA18/7 antibody blocks HBV infection by targeting the pre-S1 domain of the large HBs protein.140 Similarly, the 2H5-A14 antibody obstructs the binding of pre-S1 to NTCP, effectively neutralizing both HBV and HDV.141
Other Drugs
As the first selective cholesterol absorption inhibitor, Ezetimibe is primarily used to reduce cholesterol uptake by intestinal cells to lower blood lipids.166 In addition to its lipid-lowering effects, Ezetimibe has been shown to inhibit HBV infection in HepG2 cells transfected with NTCP,142 as well as in Huh7 cell lines expressing hNTCP.143 Fasiglifam, a partial agonist of G-protein-coupled receptor 40 (GPR40), has been used in the treatment of type 2 diabetes.167 Interestingly, pretreatment with Fasiglifam was found to reduce HBV DNA levels in NTCP-overexpressing HepG2 cells, human liver cell lines, and PXB cells, suggesting its potential as a novel HBV entry inhibitor.144 Fasiglifam has the potential to be a novel HBV entry inhibitor. Bexarotene is an RXR agonist that has an inhibitory effect on HBV infection.168,169 KIF4 is a key host factor for HBV infection, and KIF4 controls surface NTCP through anterograde NTCP transport to the cell surface. Bexarotene not only inhibits HBV/HDV entry by targeting KIF4 but also suppresses NTCP-mediated bile salt uptake in primary hepatocytes.112 Irbesartan is used to fight hypertension in patients with type 2 diabetes.170 It has also shown some activity in inhibiting liver fibrosis.171 It was found that irbesartan effectively inhibited HBV by interfering with NTCP activity, stabilized the expression of NTCP, and reduced the formation of HBeAg and cccDNA.145,146 Ritonavir, a protease inhibitor class of anti-retroviral drugs used to treat HIV infection, has an inhibitory effect on the metabolic function of NTCP.172 Studies have shown that Ritonavir can block the function of NTCP and inhibit the entry of HBV.143 Everolimus is a mammalian target of rapamycin inhibitor (mTOR)/proliferation-signal inhibitor with potent immunosuppressive and anti-proliferative effects.173 Studies have shown that everolimus significantly inhibits NTCP.147 Oxy185 is a semi-synthetic oxysterol. Studies have shown that Oxy185 interacts with NTCP to inhibit the oligomerization of NTCP, reducing the efficiency of HBV internalization.148
Natural Products
Natural products are an important source for new drug discovery.174,175 Numerous natural compounds, including terpenoids, alkaloids, and flavonoids, exhibit anti-HBV activity176 and have been shown to inhibit NTCP expression.
Epigallocatechin-3-gallate (EGCG), a kind of flavonoid in green tea extracts, has a variety of properties.177 EGCG induces clathrin-dependent endocytosis and protein degradation of NTCP from the plasma membrane, while also reducing clathrin-mediated transferrin endocytosis.149 Ergosterol peroxide (EP) is a kind of sterol that exists in medicinal mushrooms, sponges, etc. It has anti-viral, inflammatory and oxidation functions.178 EP directly interferes with the NTCP-LHBsAg interaction by acting on NTCP. EP has the potential to treat HBV infection.150 Curcumin is a polyphenol derived from herbs and dietary spices that has anti-inflammatory, anti-cancer and anti-viral properties.179,180 It inhibits HBV attachment and internalization, reducing HBV entry and NTCP density.151 Proanthocyanidin (PAC), also known as condensed tannins, is the most abundant class of natural phenolic compounds found in a variety of plants.181 PAC prevents the pre-S1 region in LHBs from attaching to NTCP, targeting HBV particles and impelling their infectiveness without affecting the NTCP-mediated BAs transport activity.152 Vanitaracin A is isolated from the culture medium of fungus Talaromyces sp,182 inhibits BAs uptake and blocks HBV and HDV entry by interacting directly with NTCP.153 Studies have shown that exophillic acid, derived from fungi, also interacts with NTCP, impairing its BAs uptake function.154 Betulin, a pentacyclic triterpenes, is obtained from natural sources and with a variety of biological activities, such as anti-inflammatory and anti-tumor activities.183 Betulin derivatives have been shown to effectively and selectively inhibit the NTCP receptor for HBV/HDV, demonstrating a clear structure-activity relationship while preserving NTCP’s BAs transport function.184
Safety of NTCP as a Drug Research Target
The inhibition of NTCP activity provides a new approach to the field of anti-HBV drug discovery. The advantage of HBV entry inhibitors that target NTCP is that they remain effective regardless of the presence or absence of viral mutations or subviral particles. Importantly, an important concern with the use of NTCP inhibitors is that these drugs may also block NTCP-mediated BAs transport, interfere with the physiological function of NTCP, and may cause adverse reactions. While studies have shown that some NTCP knockout mice exhibited reduced clearance of conjugated BAs, no significant liver damage was observed.163 Furthermore, no serious disease that associated with NTCP variants have been reported in humans,185 and no direct correlation between NTCP inhibition and drug-induced liver injury has been found.155 One patient with the NTCP R252H homozygous mutation exhibited elevated plasma bile salt levels without any signs of liver damage.186 Interestingly, 26 of 1828 healthy individuals homozygous for the p.Ser267Phe variant had no functional human NTCP, yet did not experience any related health issues.160,187 These findings suggest that humans may tolerate some degree of NTCP function loss, potentially compensated by other transporters. Nevertheless, the complete loss of human NTCP function and its long-term consequences remain unclear. Given that many drugs are transported via NTCP in the liver, more information is needed to understand drug-drug interactions when NTCP is inhibited.
Conclusion and Future Perspectives
Chronic hepatitis, which can progress to cirrhosis and liver cancer, remains a major global public health problem.188 Human NTCP plays a critical role as a receptor for HBV entry, yet the mechanisms regulating human NTCP during HBV infection remain largely unknown, and many relevant host factors have yet to be identified. Future research should focus not only on NTCP expression at the cell membrane but also on its cytoplasmic activity after binding to HBV. The relationship between NTCP, HBV replication, and HBV DNA warrants further investigation. A deeper understanding of these factors will be instrumental in the development of targeted NTCP inhibitors and the elucidation of HBV infection mechanisms. Given the important role of NTCP in HBV infection and other HBV-related diseases, it is important to find more effective drugs or natural products. Although many molecules and natural products have been tested for their inhibition of NTCP, most show high IC50 values, making it unlikely that they will work clinically or even pharmacologically. Bulevirtide may be a potential drug for the treatment of HBV and HDV, and has no obvious side effects, and has a good application prospect, however has to be daily injected.
While efficacy is paramount, the ideal treatment would involve compounds that are both effective and exhibit minimal side effects. Therefore, future drug development for hepatitis B must carefully balance the inhibition of NTCP’s physiological functions with the need to avoid serious adverse effects.
Data Sharing Statement
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Acknowledgments
The authors acknowledge using Biorender (https://app.biorender.com/user/signin) to create the schemata (Figures 2 and 3).
Funding
The present study was financially supported by the National Natural Science Foundation of China (No. 81973840 and No. 81273748); National science and Technology major projects of the 13th Five-Year Plan (2018ZX10303502); Science and Technology Program of Hebei (223777156D); Sichuan Provincial Administration of Traditional Chinese Medicine Major science and technology projects (2021XYCZ004).
Disclosure
The authors declare that they have no competing interests in this work.
References
1. Li Y, Zhou J, Li T. Regulation of the HBV Entry Receptor NTCP and its Potential in Hepatitis B Treatment. Review. Front Mol Biosci. 2022;9(879817).
2. Seeger C, Mason WS. Hepatitis B virus biology. Research Support, Non-U S Gov’t Research Support, U S Gov’t, P H S Review. Microbiol Mol Biol Rev. 2000;64(1):51–68.
3. Lee HW, Lee JS, Ahn SH. Hepatitis B Virus Cure: targets and Future Therapies. Review. Int J Mol Sci. 2020;22(1).
4. Wang J, Huang H, Liu Y, et al. HBV Genome and Life Cycle. Review Adv Exp Med Biol. 2020;9151–4_2.
5. Asselah T, Loureiro D, Boyer N, Mansouri A. Targets and future direct-acting antiviral approaches to achieve hepatitis B virus cure. The Lancet Gastroenterology & Hepatology. 2019;4(11):883–892. doi:10.1016/s2468-1253(19)30190-6
6. Murr R. Interplay between different epigenetic modifications and mechanisms. Adv Genet. 2010;70:101–141. doi:10.1016/b978-0-12-380866-0.60005-8
7. Fanning GC, Zoulim F, Hou J, Bertoletti A. Therapeutic strategies for hepatitis B virus infection: towards a cure. Nat Rev Drug Discov. 2019;18(11):827–844. doi:10.1038/s41573-019-0037-0
8. Verrier ER, Colpitts CC, Bach C, et al. Solute Carrier NTCP Regulates Innate Antiviral Immune Responses Targeting Hepatitis C Virus Infection of Hepatocytes. Cell Rep. 2016;17(5):1357–1368. doi:10.1016/j.celrep.2016.09.084
9. Yan Y, Allweiss L, Yang D, et al. Down-regulation of cell membrane localized NTCP expression in proliferating hepatocytes prevents hepatitis B virus infection. Emerging Microbes & Infections. 2019;8(1):879–894. doi:10.1080/22221751.2019.1625728
10. Nakabori T, Hikita H, Murai K, et al. Sodium taurocholate cotransporting polypeptide inhibition efficiently blocks hepatitis B virus spread in mice with a humanized liver. Sci Rep. 2016;6(1):27782. doi:10.1038/srep27782
11. Sargiacomo C, El-Kehdy H, Pourcher G, Stieger B, Najimi M, Sokal E. Age-dependent glycosylation of the sodium taurocholate cotransporter polypeptide: from fetal to adult human livers. Hepatology Communications. Jun. 2018;2(6):693–702. doi:10.1002/hep4.1174
12. Tian J, Li C, Li W. Entry of hepatitis B virus: going beyond NTCP to the nucleus. Curr Opin Virol. 2021;50:97–102. doi:10.1016/j.coviro.2021.08.001
13. Zhao K, Liu S, Chen Y, et al. Upregulation of HBV transcription by sodium taurocholate cotransporting polypeptide at the postentry step is inhibited by the entry inhibitor Myrcludex B. Emer microb infecti. 2018;7(1):186. DOI:10.1038/s41426-018-0189-8
14. Iannacone M, Guidotti LG. Immunobiology and pathogenesis of hepatitis B virus infection. Research Support, Non-U S Gov’t Review. Nat Rev Immunol. 2022;22(1):19–32.
15. Zakrzewicz D, Geyer J. Multitasking Na(+)/Taurocholate Cotransporting Polypeptide (NTCP) as a Drug Target for HBV Infection: from Protein Engineering to Drug Discovery. Review. Biomedicines. 2022;10(1).
16. Yan H, Zhong G, Xu G, et al. Sodium taurocholate cotransporting polypeptide is a functional receptor for human hepatitis B and D virus. Elife. 2012;13(1):00049.
17. Li J, Wands J. Hepatitis B and D viral receptors. Hepatology. 2016;63(1):11–13. doi:10.1002/hep.28131
18. Verrier ER, Colpitts CC, Bach C, et al. A targeted functional RNA interference screen uncovers glypican 5 as an entry factor for hepatitis B and D viruses. Hepatology. 2016;63(1):35–48.
19. Sarrazin S, Lamanna WC, Esko JD. Heparan sulfate proteoglycans. Research Support, N I H, ExtramuralResearch Support, Non-U S Gov’t Review. Cold Spring Harb Perspect Biol. 2011;3(7).
20. Leistner CM, Gruen-Bernhard S, Glebe D. Role of glycosaminoglycans for binding and infection of hepatitis B virus. Cell Microbiol. 2008;10(1):122–133.
21. Schulze A, Gripon P, Urban S. Hepatitis B virus infection initiates with a large surface protein-dependent binding to heparan sulfate proteoglycans. Hepatology. 2007;46(6):1759–1768. doi:10.1002/hep.21896
22. Sureau C, Salisse J. A conformational heparan sulfate binding site essential to infectivity overlaps with the conserved hepatitis B virus a-determinant. Hepatology. 2013;57(3):985–994.
23. Herrscher C, Roingeard P, Blanchard E. Hepatitis B Virus Entry into Cells. Cells. 2020;9(6).
24. Fukano K, Tsukuda S, Oshima M, et al. Troglitazone Impedes the Oligomerization of Sodium Taurocholate Cotransporting Polypeptide and Entry of Hepatitis B Virus Into Hepatocytes. Front Microbiol. 2018;9:3257. doi:10.3389/fmicb.2018.03257
25. Watashi K, Wakita T. Hepatitis B Virus and Hepatitis D Virus Entry, Species Specificity, and Tissue Tropism. Cold Spring Harbor perspect med. 2015;5(8):a021378. doi:10.1101/cshperspect.a021378
26. Li W, Urban S. Entry of hepatitis B and hepatitis D virus into hepatocytes: basic insights and clinical implications. J Hepatol. 2016;64(1 Suppl):S32–s40. doi:10.1016/j.jhep.2016.02.011
27. Chakraborty A, Ko C, Henning C, et al. Synchronised infection identifies early rate-limiting steps in the hepatitis B virus life cycle. Cellular Microbiology. 2020;22(12):e13250. doi:10.1111/cmi.13250
28. Hagenbuch B, Meier PJ. Molecular cloning, chromosomal localization, and functional characterization of a human liver Na+/bile acid cotransporter. J Clin Invest. 1994;93(3):1326–1331. doi:10.1172/jci117091
29. Hallén S, Björquist A, Ostlund-Lindqvist AM, Sachs G. Identification of a region of the ileal-type sodium/bile acid cotransporter interacting with a competitive bile acid transport inhibitor. Biochemistry. 2002;41(50):14916–14924. doi:10.1021/bi0205404
30. Appelman MD, Wettengel JM, Protzer U, Oude Elferink RPJ, van de Graaf SFJ. Molecular regulation of the hepatic bile acid uptake transporter and HBV entry receptor NTCP. Biochim Biophys Acta Mol Cell Biol Lipids. 2021;8(158960):29.
31. Keitel V, Burdelski M, Warskulat U, et al. Expression and localization of hepatobiliary transport proteins in progressive familial intrahepatic cholestasis. Hepatology. 2005;41(5):1160–1172.
32. Mita S, Suzuki H, Akita H, et al. Vectorial transport of unconjugated and conjugated bile salts by monolayers of LLC-PK1 cells doubly transfected with human NTCP and BSEP or with rat Ntcp and Bsep. Am J Physiol Gastrointest Liver Physiol. 2006;290(3).
33. Visser WE, Wong WS, van Mullem AA, Friesema EC, Geyer J, Visser TJ. Study of the transport of thyroid hormone by transporters of the SLC10 family. Mol Cell Endocrinol. 2010;315(1–2):138–145.
34. Watashi K, Urban S, Li W, Wakita T. NTCP and beyond: opening the door to unveil hepatitis B virus entry. Research Support, Non-U S Gov’t Review. Int J Mol Sci. 2014;15(2):2892–2905.
35. Park JH, Iwamoto M, Yun JH, et al. Structural insights into the HBV receptor and bile acid transporter NTCP. Nature. 2022;606(7916):1027–1031. doi:10.1038/s41586-022-04857-0
36. Goutam K, Ielasi FS, Pardon E, Steyaert J, Reyes N. Structural basis of sodium-dependent bile salt uptake into the liver. Nature. 2022;606(7916):1015–1020. doi:10.1038/s41586-022-04723-z
37. Liu H, Irobalieva RN, Bang-Sørensen R, et al. Structure of human NTCP reveals the basis of recognition and sodium-driven transport of bile salts into the liver. Cell Res. 2022;32(8):773–776. doi:10.1038/s41422-022-00680-4
38. Asami J, Kimura KT, Fujita-Fujiharu Y, et al. Structure of the bile acid transporter and HBV receptor NTCP. Nature. 2022;606(7916):1021–1026. doi:10.1038/s41586-022-04845-4
39. Asami J, Park JH, Nomura Y, et al. Structural basis of hepatitis B virus receptor binding. Nat Struct Mol Biol. 2024;31(3):447–454. doi:10.1038/s41594-023-01191-5
40. Li Y, Luo G. Human low-density lipoprotein receptor plays an important role in hepatitis B virus infection. PLoS Pathogens. 2021;17(7):e1009722. doi:10.1371/journal.ppat.1009722
41. Ahn SJ, Kim DK, Kim SS, et al. Association between apolipoprotein E genotype, chronic liver disease, and hepatitis B virus. Clinic molec hepatol. 2012;18(3):295–301. DOI:10.3350/cmh.2012.18.3.295
42. Toniutto P, Fattovich G, Fabris C, et al. Genetic polymorphism at the apolipoprotein E locus affects the outcome of chronic hepatitis B. J med virol. 2010;82(2):224–331. doi:10.1002/jmv.21642
43. Liu YM, Zhang WY, Wang ZF, Yan CY, Gao PJ. High expression of beta2-glycoprotein I is associated significantly with the earliest stages of hepatitis B virus infection. J Med Virol. 2014;86(8):1296–1306.
44. Shirasaki T, Murai K, Ishida A, et al. Functional involvement of endothelial lipase in hepatitis B virus infection. Hepatol Commun. 2023;7(9):1.
45. Iwamoto M, Saso W, Sugiyama R, et al. Epidermal growth factor receptor is a host-entry cofactor triggering hepatitis B virus internalization. Proc Natl Acad Sci U S A. 2019;116(17):8487–8492.
46. Iwamoto M, Saso W, Nishioka K, et al. The machinery for endocytosis of epidermal growth factor receptor coordinates the transport of incoming hepatitis B virus to the endosomal network. J Biol Chem. 2020;295(3):800–807. doi:10.1074/jbc.AC119.010366
47. Wang X, Wang P, Wang W, Murray JW, Wolkoff AW. The Na(+)-Taurocholate Cotransporting Polypeptide Traffics with the Epidermal Growth Factor Receptor. Research Support, N I H, Extramural. Traffic. 2016;17(3):230–244.
48. Palatini M, Müller SF, Kirstgen M, et al. IFITM3 Interacts with the HBV/HDV Receptor NTCP and Modulates Virus Entry and Infection. Viruses. 2022;14(4). doi:10.3390/v14040727
49. Makjaroen J, Somparn P, Hodge K, Poomipak W, Hirankarn N, Pisitkun T. Comprehensive Proteomics Identification of IFN-λ3-regulated Antiviral Proteins in HBV-transfected Cells. Molecul Cell Proteomi. 2018;17(11):2197–2215. doi:10.1074/mcp.RA118.000735
50. Liu Y, Cafiero TR, Park D, et al. Targeted viral adaptation generates a simian-tropic hepatitis B virus that infects marmoset cells. Research Support, N I H, Extramural. Nat Commun. 2023;14(1):23–39148.
51. Jeon H, Blacklow SC. Structure and physiologic function of the low-density lipoprotein receptor. Annu. Rev. Biochem. 2005;74:535–562. doi:10.1146/annurev.biochem.74.082803.133354
52. Qiao L, Luo GG. Human apolipoprotein E promotes hepatitis B virus infection and production. PLoS Pathogens. 2019;15(8):e1007874. doi:10.1371/journal.ppat.1007874
53. Esser K, Lucifora J, Wettengel J, et al. Lipase inhibitor orlistat prevents hepatitis B virus infection by targeting an early step in the virus life cycle. Antiviral Res. 2018;151:4–7. doi:10.1016/j.antiviral.2018.01.001
54. Oswald A, Chakraborty A, Ni Y, Wettengel JM, Urban S, Protzer U. Concentration of Na(+)-taurocholate-cotransporting polypeptide expressed after in vitro-transcribed mRNA transfection determines susceptibility of hepatoma cells for hepatitis B virus. Sci Rep. 2021;11(1):021–99263.
55. Fukano K, Oshima M, Tsukuda S, et al. NTCP Oligomerization Occurs Downstream of the NTCP-EGFR Interaction during Hepatitis B Virus Internalization. J Virol. 2021;95(24):e0093821. doi:10.1128/jvi.00938-21
56. Li P, Shi ML, Shen WL, et al. Coordinated regulation of IFITM1, 2 and 3 genes by an IFN-responsive enhancer through long-range chromatin interactions. Biochimica et biophys acta Gene reg mech. 2017;1860(8):885–893. DOI:10.1016/j.bbagrm.2017.05.003
57. Bedford JG, O’Keeffe M, Reading PC, Wakim LM. Rapid interferon independent expression of IFITM3 following T cell activation protects cells from influenza virus infection. PLoS One. 2019;14(1):e0210132. doi:10.1371/journal.pone.0210132
58. Neupane P, Bhuju S, Thapa N, Bhattarai HK. ATP Synthase: structure, Function and Inhibition. Biomolecular Concepts. 2019;10(1):1–10. doi:10.1515/bmc-2019-0001
59. Ueda K, Suwanmanee Y. ATP5B Is an Essential Factor for Hepatitis B Virus Entry. Int J Mol Sci. 2022;23(17). doi:10.3390/ijms23179570
60. De Crignis E, Hossain T, Romal S, et al. Application of human liver organoids as a patient-derived primary model for HBV infection and related hepatocellular carcinoma. Elife. 2021;30(10):60747.
61. Yang L, Zhou D, Martin K, et al. Aborted infection of human sodium taurocholate cotransporting polypeptide (hNTCP) expressing woodchuck hepatocytes with hepatitis B virus (HBV). Virus Genes. 2023;59(6):823–830.
62. Wettengel JM, Burwitz BJ. Innovative HBV Animal Models Based on the Entry Receptor NTCP. Review. Viruses. 2020;12(8).
63. Zakrzewicz D, Leidolf R, Kunz S, et al. Tyrosine 146 of the Human Na(+)/Taurocholate Cotransporting Polypeptide (NTCP) Is Essential for Its Hepatitis B Virus (HBV) Receptor Function and HBV Entry into Hepatocytes. Viruses. 2022;14(6).
64. Binh MT, Hoan NX, Van Tong H, et al. NTCP S267F variant associates with decreased susceptibility to HBV and HDV infection and decelerated progression of related liver diseases. Int J Infect Dis. 2019;80:147–152.
65. An P, Zeng Z, Winkler CA. The Loss-of-Function S267F Variant in HBV Receptor NTCP Reduces Human Risk for HBV Infection and Disease Progression. Research Support, NIH, Extramural Research Support, N I H, Intramural. J Infect Dis. 2018;218(9):1404–1410.
66. Yang F, Wu L, Xu W, et al. Diverse Effects of the NTCP p.Ser267Phe Variant on Disease Progression During Chronic HBV Infection and on HBV preS1 Variability. Front Cell Infect Microbiol. 2019;9(18).
67. Chen YH, Tsuei DJ, Lai MW, et al. Genetic variants of NTCP gene and hepatitis B vaccine failure in Taiwanese children of hepatitis B e antigen positive mothers. Hepatol Int. 2022;16(4):789–798.
68. Wu L, Xu W, Li X, et al. The NTCP p.Ser267Phe Variant Is Associated With a Faster Anti-HBV Effect on First-Line Nucleos(t)ide Analog Treatment. Front Pharmacol. 2021;12(616858).
69. Ananthanarayanan M, Balasubramanian N, Makishima M, Mangelsdorf DJ, Suchy FJ. Human bile salt export pump promoter is transactivated by the farnesoid X receptor/bile acid receptor. Research Support, Non-U S Gov’tResearch Support, U S Gov’t, P H S. J Biol Chem. 2001;276(31):28857–28865.
70. Kim HY, Cho HK, Choi YH, Lee KS, Cheong J. Bile acids increase hepatitis B virus gene expression and inhibit interferon-alpha activity. FEBS J. 2010;277(13):2791–2802. doi:10.1111/j.1742-4658.2010.07695.x
71. Ramière C, Scholtès C, Diaz O, et al. Transactivation of the hepatitis B virus core promoter by the nuclear receptor FXRalpha. J Virol. 2008;82(21):10832–10840. doi:10.1128/jvi.00883-08
72. Reese VC, Moore DD, McLachlan A. Limited effects of bile acids and small heterodimer partner on hepatitis B virus biosynthesis in vivo. J Virol. 2012;86(5):2760–2768. doi:10.1128/jvi.06742-11
73. Denson LA, Sturm E, Echevarria W, et al. The orphan nuclear receptor, shp, mediates bile acid-induced inhibition of the rat bile acid transporter, ntcp. Gastroenterology. 2001;121(1):140–147. doi:10.1053/gast.2001.25503
74. Slijepcevic D, Roscam Abbing RLP, Katafuchi T, et al. Hepatic uptake of conjugated bile acids is mediated by both sodium taurocholate cotransporting polypeptide and organic anion transporting polypeptides and modulated by intestinal sensing of plasma bile acid levels in mice. Hepatology. 2017;66(5):1631–1643. doi:10.1002/hep.29251
75. Jung D, Hagenbuch B, Fried M, Meier PJ, Kullak-Ublick GA. Role of liver-enriched transcription factors and nuclear receptors in regulating the human, mouse, and rat NTCP gene. Am J Physiol Gastrointest Liver Physiol. 2004;286(5):30.
76. Shih DQ, Bussen M, Sehayek E, et al. Hepatocyte nuclear factor-1 alpha is an essential regulator of bile acid and plasma cholesterol metabolism. Research Support, Non-U S Gov’tResearch Support, U S Gov’t, P H S. Nat Genet. 2001;27(4):375–382.
77. Purushotham A, Xu Q, Lu J, et al. Hepatic deletion of SIRT1 decreases hepatocyte nuclear factor 1α/farnesoid X receptor signaling and induces formation of cholesterol gallstones in mice. Research Support, N I H, Extramural Research Support, N I H, Intramural. Mol Cell Biol. 2012;32(7):1226–1236.
78. Geier A, Martin IV, Dietrich CG, et al. Hepatocyte nuclear factor-4 alpha is a central transactivator of the mouse Ntcp gene. Research Support, N I H, Extramural. Am J Physiol Gastrointest Liver Physiol. 2008;295(2):15.
79. Wang J, Tian R, Shan Y, et al. Metabolomics study of the metabolic changes in hepatoblastoma cells in response to NTCP/SLC10A1 overexpression. Int J Biochem Cell Biol. 2020;125(105773):22.
80. Rose AJ, Berriel Díaz M, Reimann A, et al. Molecular control of systemic bile acid homeostasis by the liver glucocorticoid receptor. Cell Metabolism. 2011;14(1):123–130. doi:10.1016/j.cmet.2011.04.010
81. Cao J, Huang L, Liu Y, et al. Differential regulation of hepatic bile salt and organic anion transporters in pregnant and postpartum rats and the role of prolactin. Hepatology. 2001;33(1):140–147. doi:10.1053/jhep.2001.20895
82. Ganguly TC, O’Brien ML, Karpen SJ, Hyde JF, Suchy FJ, Vore M. Regulation of the rat liver sodium-dependent bile acid cotransporter gene by prolactin. Mediation of transcriptional activation by Stat5. J Clin Invest. 1997;99(12):2906–2914. doi:10.1172/jci119485
83. Cao J, Wood M, Liu Y, et al. Estradiol represses prolactin-induced expression of Na+/taurocholate cotransporting polypeptide in liver cells through estrogen receptor-alpha and signal transducers and activators of transcription 5a. Endocrinology. 2004;145(4):1739–1749. doi:10.1210/en.2003-0752
84. Simon FR, Fortune J, Iwahashi M, Qadri I, Sutherland E. Multihormonal regulation of hepatic sinusoidal Ntcp gene expression. Am J Physiol Gastrointest Liver Physiol. 2004;287(4):G782–94. doi:10.1152/ajpgi.00379.2003
85. Buettner N, Thimme R. Sexual dimorphism in hepatitis B and C and hepatocellular carcinoma. Semin Immunopathol. 2019;41(2):203–211. doi:10.1007/s00281-018-0727-4
86. Ribeiro CRA, Beghini DG, Lemos AS, et al. Cytokines profile in patients with acute and chronic hepatitis B infection. Microbiol Immunol. 2022;66(1):31–39. doi:10.1111/1348-0421.12947
87. Ribeiro CRA, Martinelli KG, de Mello VDM, et al. Cytokine, Genotype, and Viral Load Profile in the Acute and Chronic Hepatitis B. Viral Immunology. 2020;33(10):620–627. doi:10.1089/vim.2020.0176
88. Naseem S, Hussain T, Manzoor S. Interleukin-6: a promising cytokine to support liver regeneration and adaptive immunity in liver pathologies. Cytokine & Growth Factor Reviews. 2018;39:36–45. doi:10.1016/j.cytogfr.2018.01.002
89. Bouezzedine F, Fardel O, Gripon P. Interleukin 6 inhibits HBV entry through NTCP down regulation. Virology. 2015;481:34–42.
90. Jung D, Hagenbuch B, Fried M, Meier PJ, Kullak-Ublick GA. Role of liver-enriched transcription factors and nuclear receptors in regulating the human, mouse, and rat NTCP gene. Am J Physiol Gastrointest Liver Physiol. 2004;286(5):G752–61. doi:10.1152/ajpgi.00456.2003
91. Larkin J, Clayton MM, Liu J, Feitelson MA. Chronic ethanol consumption stimulates hepatitis B virus gene expression and replication in transgenic mice. Hepatology. 2001;34(4 Pt 1):792–797. doi:10.1053/jhep.2001.27565
92. Geier A, Dietrich CG, Voigt S, et al. Effects of proinflammatory cytokines on rat organic anion transporters during toxic liver injury and cholestasis. Hepatology. 2003;38(2):345–354. doi:10.1053/jhep.2003.50317
93. Hu JF, Cheng Z, Chisari FV, Vu TH, Hoffman AR, Campbell TC. Repression of hepatitis B virus (HBV) transgene and HBV-induced liver injury by low protein diet. Oncogene. 1997;15(23):2795–2801. doi:10.1038/sj.onc.1201444
94. Anwer MS, Stieger B. Sodium-dependent bile salt transporters of the SLC10A transporter family: more than solute transporters. Research Support, N I H, Extramural Review. Cold Spring Harb Perspect Biol. 2011;3(7):77–89.
95. Webster CR, Blanch C, Anwer MS. Role of PP2B in cAMP-induced dephosphorylation and translocation of NTCP. Am J Physiol Gastrointest Liver Physiol. 2002;283(1):G44–50. doi:10.1152/ajpgi.00530.2001
96. Anwer MS, Gillin H, Mukhopadhyay S, Balasubramaniyan N, Suchy FJ, Ananthanarayanan M. Dephosphorylation of Ser-226 facilitates plasma membrane retention of Ntcp. J Biol Chem. 2005;280(39):33687–33692. doi:10.1074/jbc.M502151200
97. Sun AQ, Arrese MA, Zeng L, Swaby I, Zhou MM, Suchy FJ. The rat liver Na(+)/bile acid cotransporter. Importance of the cytoplasmic tail to function and plasma membrane targeting. J Biol Chem. 2001;276(9):6825–6833. doi:10.1074/jbc.M008797200
98. Appelman MD, Chakraborty A, Protzer U, McKeating JA, van de Graaf SF. N-Glycosylation of the Na+-Taurocholate Cotransporting Polypeptide (NTCP) Determines Its Trafficking and Stability and Is Required for Hepatitis B Virus Infection. PLoS One. 2017;12(1):e0170419. doi:10.1371/journal.pone.0170419
99. Webster CRL, Blanch CJ, Phillips J, Anwer MS. Cell Swelling-induced Translocation of Rat Liver Na+/Taurocholate Cotransport Polypeptide Is Mediated via the Phosphoinositide 3-Kinase Signaling Pathway. J Biol Chem. 2000;275(38):29754–29760. doi:10.1074/jbc.M002831200
100. Webster CRL, Srinivasulu U, Ananthanarayanan M, Suchy FJ, Anwer MS. Protein kinase B/Akt mediates cAMP- and cell swelling-stimulated Na+/taurocholate cotransport and Ntcp translocation. J Biol Chem. 2002;277(32):28578–28583. doi:10.1074/jbc.M201937200
101. Sarkar S, Bananis E, Nath S, Anwer MS, Wolkoff AW, Murray JW. PKCζ Is Required for Microtubule-Based Motility of Vesicles Containing the ntcp Transporter. Traffic. 2006;7(8):1078–1091. doi:10.1111/j.1600-0854.2006.00447.x
102. McConkey M, Gillin H, Webster CRL, Anwer MS. Cross-talk between Protein Kinases Cζ and B in Cyclic AMP-mediated Sodium Taurocholate Co-transporting Polypeptide Translocation in Hepatocytes. J Biol Chem. 2004;279(20):20882–20888. doi:10.1074/jbc.M309988200
103. Park SW, Schonhoff CM, Webster CRL, Anwer MS. Protein kinase Cδ differentially regulates cAMP-dependent translocation of NTCP and MRP2 to the plasma membrane. American Journal of physiology-Gastrointestinal and Liver Physiology. 2012;303(5):G657–65. doi:10.1152/ajpgi.00529.2011
104. Grüne S, Engelking LR, Anwer MS. Role of intracellular calcium and protein kinases in the activation of hepatic Na+/taurocholate cotransport by cyclic AMP. J Biol Chem. 1993;268(24):17734–17741. doi:10.1016/S0021-9258(17)46766-4
105. Stross C, Helmer A, Weissenberger K, et al. Protein kinase C induces endocytosis of the sodium taurocholate cotransporting polypeptide. American Journal of physiology-Gastrointestinal and Liver Physiology. 2010;299(2):G320–8. doi:10.1152/ajpgi.00180.2010
106. Mayer PGK, Qvartskhava N, Sommerfeld A, Görg B, Häussinger D. Regulation of Plasma Membrane Localization of the Na⁺-Taurocholate Co-Transporting Polypeptide by Glycochenodeoxycholate and Tauroursodeoxycholate. Cellul Physio Biochem. 2019;52(6):1427–1445. doi:10.33594/000000100
107. Evripioti AA, Ortega-Prieto AM, Skelton JK, Bazot Q, Dorner M. Phosphodiesterase-induced cAMP degradation restricts hepatitis B virus infection. Philos Trans R Soc Lond B Biol Sci. 2019;374:1773.
108. Powers J, Rose DJ, Saunders A, Dunkelbarger S, Strome S, Saxton WM. Loss of KLP-19 polar ejection force causes misorientation and missegregation of holocentric chromosomes. J Cell Biol. 2004;166(7):991–1001. doi:10.1083/jcb.200403036
109. Williams BC, Riedy MF, Williams EV, Gatti M, Goldberg ML. The Drosophila kinesin-like protein KLP3A is a midbody component required for central spindle assembly and initiation of cytokinesis. J Cell Biol. 1995;129(3):709–723. doi:10.1083/jcb.129.3.709
110. Vernos I, Raats J, Hirano T, Heasman J, Karsenti E, Wylie C. Xklp1, a chromosomal Xenopus kinesin-like protein essential for spindle organization and chromosome positioning. Cell. 1995;81(1):117–127. doi:10.1016/0092-8674(95)90376-3
111. Zhu CL, Cheng DZ, Liu F, et al. Hepatitis B virus upregulates the expression of kinesin family member 4A. Molecular Medicine Reports. 2015;12(3):3503–3507. doi:10.3892/mmr.2015.3792
112. Gad SA, Sugiyama M, Tsuge M, et al. The kinesin KIF4 mediates HBV/HDV entry through the regulation of surface NTCP localization and can be targeted by RXR agonists in vitro. PLoS Pathogens. 2022;18(3):e1009983. doi:10.1371/journal.ppat.1009983
113. Fonseca I, da Luz FAC, Uehara IA, Silva MJB. Cell-adhesion molecules and their soluble forms: promising predictors of ”tumor progression” and relapse in leukemia. Tumour Biology: the Journal of the International Society for Oncodevelopmental Biology and Medicine. 2018;40(11):1010428318811525. doi:10.1177/1010428318811525
114. Shneider BL, Fox VL, Schwarz KB, et al. Hepatic basolateral sodium-dependent-bile acid transporter expression in two unusual cases of hypercholanemia and in extrahepatic biliary atresia. Hepatology. 1997;25(5):1176–1183. doi:10.1002/hep.510250521
115. Bonazzi M, Cossart P. Impenetrable barriers or entry portals? The role of cell-cell adhesion during infection. J Cell Biol. 2011;195(3):349–358. doi:10.1083/jcb.201106011
116. Connolly SA, Landsburg DJ, Carfi A, et al. Potential nectin-1 binding site on herpes simplex virus glycoprotein d. J Virol. 2005;79(2):1282–1295. doi:10.1128/jvi.79.2.1282-1295.2005
117. Mateo M, Generous A, Sinn PL, Cattaneo R. Connections matter--how viruses use cell–cell adhesion components. J Cell Sci. 2015;128(3):431–439. doi:10.1242/jcs.159400
118. Salim A, Ratner L. Modulation of beta-catenin and E-cadherin interaction by Vpu increases human immunodeficiency virus type 1 particle release. J Virol. 2008;82(8):3932–3938. doi:10.1128/jvi.00430-07
119. Hu Q, Zhang F, Duan L, et al. E-cadherin Plays a Role in Hepatitis B Virus Entry Through Affecting Glycosylated Sodium-Taurocholate Cotransporting Polypeptide Distribution. Front Cell Infect Microbiol. 2020;10:74. doi:10.3389/fcimb.2020.00074
120. Bogomolov P, Alexandrov A, Voronkova N, et al. Treatment of chronic hepatitis D with the entry inhibitor myrcludex B: first results of a phase Ib/IIa study. J Hepatol. 2016;65(3):490–498. doi:10.1016/j.jhep.2016.04.016
121. Blank A, Markert C, Hohmann N, et al. First-in-human application of the novel hepatitis B and hepatitis D virus entry inhibitor myrcludex B. J Hepatol. 2016;65(3):483–489.
122. Shepperson OA, Cameron AJ, Wang CJ, Harris PWR, Taylor JA, Brimble MA. Thiol-ene enabled preparation of S-lipidated anti-HBV peptides. Org Biomol Chem. 2021;19(1):220–232.
123. Wedemeyer H, Schöneweis K, Bogomolov P, et al. Safety and efficacy of bulevirtide in combination with tenofovir disoproxil fumarate in patients with hepatitis B virus and hepatitis D virus coinfection (MYR202): a multicentre, randomised, parallel-group, open-label, Phase 2 trial. The Lancet Infectious Diseases. 2023;23(1):117–129. doi:10.1016/s1473-3099(22)00318-8
124. Herta T, Hahn M, Maier M, et al. Efficacy and Safety of Bulevirtide plus Tenofovir Disoproxil Fumarate in Real-World Patients with Chronic Hepatitis B and D Co-Infection. Pathogens. 2022;11(5).
125. Urban S, Bartenschlager R, Kubitz R, Zoulim F. Strategies to inhibit entry of HBV and HDV into hepatocytes. Gastroenterology. 2014;147(1):48–64. doi:10.1053/j.gastro.2014.04.030
126. Lempp FA, Ni Y, Urban S. Hepatitis delta virus: insights into a peculiar pathogen and novel treatment options. Nature reviews Gastroenterology & hepatology. 2016;13(10):580–589. doi:10.1038/nrgastro.2016.126
127. Loglio A, Ferenci P, Uceda Renteria SC, et al. Excellent safety and effectiveness of high-dose myrcludex-B monotherapy administered for 48 weeks in HDV-related compensated cirrhosis: a case report of 3 patients. J Hepatol. 2019;71(4):834–839. doi:10.1016/j.jhep.2019.07.003
128. Mita S, Suzuki H, Akita H, et al. Inhibition of bile acid transport across Na+/taurocholate cotransporting polypeptide (SLC10A1) and bile salt export pump (ABCB 11)-coexpressing LLC-PK1 cells by cholestasis-inducing drugs. Drug Metabo Disposit. 2006;34(9):1575–1581. doi:10.1124/dmd.105.008748
129. Nkongolo S, Ni Y, Lempp FA, et al. Cyclosporin A inhibits hepatitis B and hepatitis D virus entry by cyclophilin-independent interference with the NTCP receptor. J Hepatol. 2014;60(4):723–731.
130. Watashi K, Sluder A, Daito T, et al. Cyclosporin A and its analogs inhibit hepatitis B virus entry into cultured hepatocytes through targeting a membrane transporter, sodium taurocholate cotransporting polypeptide (NTCP). Hepatology. 2014;59(5):1726–1737.
131. Shimura S, Watashi K, Fukano K, et al. Cyclosporin derivatives inhibit hepatitis B virus entry without interfering with NTCP transporter activity. J Hepatol. 2017;66(4):685–692.
132. Liu Y, Ruan H, Li Y, et al. Potent and Specific Inhibition of NTCP-Mediated HBV/HDV Infection and Substrate Transporting by a Novel, Oral-Available Cyclosporine A Analogue. J Med Chem. 2021;64(1):543–565.
133. Iwamoto M, Watashi K, Tsukuda S, et al. Evaluation and identification of hepatitis B virus entry inhibitors using HepG2 cells overexpressing a membrane transporter NTCP. Biochem Biophys Res Commun. 2014;443(3):808–813.
134. Tanaka T, Okuyama-Dobashi K, Motohashi R, et al. Inhibitory effect of a novel thiazolidinedione derivative on hepatitis B virus entry. Antiviral Res. 2021;194(105165):19.
135. Yan H, Peng B, Liu Y, et al. Viral entry of hepatitis B and D viruses and bile salts transportation share common molecular determinants on sodium taurocholate cotransporting polypeptide. J Virol. 2014;88(6):3273–3284.
136. Ito K, Okumura A, Takeuchi JS, et al. Dual Agonist of Farnesoid X Receptor and Takeda G Protein-Coupled Receptor 5 Inhibits Hepatitis B Virus Infection In Vitro and In Vivo. Hepatology. 2021;74(1):83–98.
137. Liu Y, Zhang L, Yan H, et al. Design of Dimeric Bile Acid Derivatives as Potent and Selective Human NTCP Inhibitors. J Med Chem. 2021;64(9):5973–6007.
138. Lowjaga K, Kirstgen M, Müller SF, et al. Long-term trans-inhibition of the hepatitis B and D virus receptor NTCP by taurolithocholic acid. Am J Physiol Gast Liver Physiol. 2021;320(1):G66–G80.
139. Takemori T, Sugimoto-Ishige A, Nishitsuji H, et al. Establishment of a Monoclonal Antibody against Human NTCP That Blocks Hepatitis B Virus Infection. J Virol. 2022;96(5):e0168621. doi:10.1128/jvi.01686-21
140. Glebe D, Aliakbari M, Krass P, Knoop EV, Valerius KP, Gerlich WH. Pre-s1 antigen-dependent infection of Tupaia hepatocyte cultures with human hepatitis B virus. J Virol. 2003;77(17):9511–9521.
141. Li D, He W, Liu X, et al. A potent human neutralizing antibody Fc-dependently reduces established HBV infections. Elife. 2017;26(6):26738.
142. König A, Döring B, Mohr C, Geipel A, Geyer J, Glebe D. Kinetics of the bile acid transporter and hepatitis B virus receptor Na+/taurocholate cotransporting polypeptide (NTCP) in hepatocytes. J Hepatol. 2014;61(4):867–875.
143. Blanchet M, Sureau C, Labonté P. Use of FDA approved therapeutics with hNTCP metabolic inhibitory properties to impair the HDV lifecycle. Antiviral Res. 2014;106:111–115.
144. Nio Y, Akahori Y, Okamura H, Watashi K, Wakita T, Hijikata M. Inhibitory effect of fasiglifam on hepatitis B virus infections through suppression of the sodium taurocholate cotransporting polypeptide. Biochem Biophys Res Commun. 2018;501(3):820–825.
145. Ko C, Park WJ, Park S, Kim S, Windisch MP, Ryu WS. The FDA-approved drug irbesartan inhibits HBV-infection in HepG2 cells stably expressing sodium taurocholate co-transporting polypeptide. Antivir Ther. 2015;20(8):835–842.
146. Wang XJ, Hu W, Zhang TY, Mao YY, Liu NN, Wang SQ. Irbesartan, an FDA approved drug for hypertension and diabetic nephropathy, is a potent inhibitor for hepatitis B virus entry by disturbing Na(+)-dependent taurocholate cotransporting polypeptide activity. Antiviral Res. 2015;120:140–146.
147. Saran C, Ho H, Honkakoski P, Brouwer KLR. Effect of mTOR inhibitors on sodium taurocholate cotransporting polypeptide (NTCP) function in vitro. Front Pharmacol. 2023;14(1147495).
148. Oshima M, Stappenbeck F, Ohashi H, et al. Selective inhibition of hepatitis B virus internalization by oxysterol derivatives. Biochem Biophys Res Commun. 2023;675:139–145.
149. Huang HC, Tao MH, Hung TM, Chen JC, Lin ZJ, Huang C. (-)-Epigallocatechin-3-gallate inhibits entry of hepatitis B virus into hepatocytes. Antiviral Res. 2014;111:100–111.
150. Huang H, Huang HC, Chiou WC, et al. Ergosterol peroxide inhibits HBV infection by inhibiting the binding of the pre-S1 domain of LHBsAg to NTCP. Antiviral Res. 2021;195:105184. doi:10.1016/j.antiviral.2021.105184
151. Thongsri P, Pewkliang Y, Borwornpinyo S, Wongkajornsilp A, Hongeng S, Sa-Ngiamsuntorn K. Curcumin inhibited hepatitis B viral entry through NTCP binding. Sci Rep. 2021;11(1):19125. doi:10.1038/s41598-021-98243-x
152. Tsukuda S, Watashi K, Hojima T, et al. A new class of hepatitis B and D virus entry inhibitors, proanthocyanidin and its analogs, that directly act on the viral large surface proteins. Hepatology. 2017;65(4):1104–1116.
153. Kaneko M, Watashi K, Kamisuki S, et al. A Novel Tricyclic Polyketide, Vanitaracin A, Specifically Inhibits the Entry of Hepatitis B and D Viruses by Targeting Sodium Taurocholate Cotransporting Polypeptide. J Virol. 2015;89(23):11945–11953.
154. Kobayashi C, Watanabe Y, Oshima M, et al. Fungal Secondary Metabolite Exophillic Acid Selectively Inhibits the Entry of Hepatitis B and D Viruses. Viruses. 2022;14(4).
155. Dong Z, Ekins S, Polli JE. Quantitative NTCP pharmacophore and lack of association between Dili and NTCP Inhibition. European j Pharmaceutical Scien. 2015;66:1–9. doi:10.1016/j.ejps.2014.09.005
156. Kang C, Syed YY. Bulevirtide: first Approval. Drugs. 2020;80(15):1601–1605. doi:10.1007/s40265-020-01400-1
157. Yardeni D, Koh C. Bulevirtide for HBV and HDV infections. Drugs Today. 2021;57(7):433–448.
158. Cheng D, Han B, Zhang W, Wu W. Clinical effects of NTCP-inhibitor myrcludex B. Research Support, Non-U S Gov’t Review. J Viral Hepat. 2021;28(6):852–858.
159. Meier A, Mehrle S, Weiss TS, Mier W, Urban S. Myristoylated PreS1-domain of the hepatitis B virus L-protein mediates specific binding to differentiated hepatocytes. Hepatology. 2013;58(1):31–42. doi:10.1002/hep.26181
160. Volz T, Allweiss L, Ben MM, et al. The entry inhibitor Myrcludex-B efficiently blocks intrahepatic virus spreading in humanized mice previously infected with hepatitis B virus. J Hepatol. 2013;58(5):861–867. doi:10.1016/j.jhep.2012.12.008
161. Liu H, Zakrzewicz D, Nosol K, et al. Structure of antiviral drug bulevirtide bound to hepatitis B and D virus receptor protein NTCP. Nat Commun. 2024;15(1):2476. doi:10.1038/s41467-024-46706-w
162. Ni Y, Lempp FA, Mehrle S, et al. Hepatitis B and D viruses exploit sodium taurocholate co-transporting polypeptide for species-specific entry into hepatocytes. Gastroenterology. 2014;146(4):1070–1083. doi:10.1053/j.gastro.2013.12.024
163. Slijepcevic D, Kaufman C, Wichers CG, et al. Impaired uptake of conjugated bile acids and hepatitis b virus pres1-binding in na(+) -taurocholate cotransporting polypeptide knockout mice. Hepatology. 2015;62(1):207–219. doi:10.1002/hep.27694
164. Vaz FM, Paulusma CC, Huidekoper H, et al. Sodium taurocholate cotransporting polypeptide (SLC10A1) deficiency: conjugated hypercholanemia without a clear clinical phenotype. Hepatology (Baltimore. 2015;61(1):260–267. doi:10.1002/hep.27240
165. Scheen AJ, Lefèbvre PJ. Troglitazone: antihyperglycemic activity and potential role in the treatment of type 2 diabetes. Diabetes Care. 1999;22(9):1568–1577. doi:10.2337/diacare.22.9.1568
166. Nutescu EA, Shapiro NL. Ezetimibe: a selective cholesterol absorption inhibitor. Pharmacotherapy. 2003;23(11):1463–1474. doi:10.1592/phco.23.14.1463.31942
167. Negoro N, Sasaki S, Mikami S, et al. Discovery of TAK-875: a Potent, Selective, and Orally Bioavailable GPR40 Agonist. ACS Med Chem Lett. 2010;1(6):290–294.
168. Song M, Sun Y, Tian J, et al. Silencing Retinoid X Receptor Alpha Expression Enhances Early-Stage Hepatitis B Virus Infection In Cell Cultures. J Virol. 2018;92(8). doi:10.1128/jvi.01771-17
169. Gaunt CM, Rainbow DB, Mackenzie RJ, et al. The MS Remyelinating Drug Bexarotene (an RXR Agonist) Promotes Induction of Human Tregs and Suppresses Th17 Differentiation In Vitro. Front Immunol. 2021;12:712241. doi:10.3389/fimmu.2021.712241
170. Lewis EJ, Hunsicker LG, Clarke WR, et al. Renoprotective effect of the angiotensin-receptor antagonist irbesartan in patients with nephropathy due to type 2 diabetes. Clinical Trial Comparative Study Multicenter Study Randomized Controlled Trial. N Engl J Med. 2001;345(12):851–860.
171. Kim MY, Baik SK, Park DH, et al. Angiotensin receptor blockers are superior to angiotensin-converting enzyme inhibitors in the suppression of hepatic fibrosis in a bile duct-ligated rat model. J Gastroenterol. 2008;43(11):889–896.
172. Abbas Z, Abbas M. Management of hepatitis delta: need for novel therapeutic options. World J Gastroenterol. 2015;21(32):9461–9465. doi:10.3748/wjg.v21.i32.9461
173. Tedesco-Silva H, Saliba F, Barten MJ, et al. An overview of the efficacy and safety of everolimus in adult solid organ transplant recipients. Transplant Rev. 2022;36(1):24.
174. Demain AL, Sanchez S. Microbial drug discovery: 80 years of progress. The Journal of Antibiotics. 2009;62(1):5–16. doi:10.1038/ja.2008.16
175. Harvey AL, Edrada-Ebel R, Quinn RJ. The re-emergence of natural products for drug discovery in the genomics era. Nature revi Drug discov. 2015;14(2):111–129. doi:10.1038/nrd4510
176. Wu YH. Naturally derived anti-hepatitis B virus agents and their mechanism of action. World Journal of Gastroenterology. 2016;22(1):188–204. doi:10.3748/wjg.v22.i1.188
177. Calland N, Albecka A, Belouzard S, et al. (-)-Epigallocatechin-3-gallate is a new inhibitor of hepatitis C virus entry. Hepatology. 2012;55(3):720–729.
178. Liu Y, Wang X, Wang J, Zhang J, Duan C. Ergosterol Peroxide Inhibits Porcine Epidemic Diarrhea Virus Infection in Vero Cells by Suppressing ROS Generation and p53 Activation. Viruses. 2022;14(2).
179. Sharma RA, Gescher AJ, Steward WP. Curcumin: the story so far. Eur J Cancer. 2005;41(13):1955–1968. doi:10.1016/j.ejca.2005.05.009
180. Jennings MR, Parks RJ. Curcumin as an Antiviral Agent. Viruses. 2020;12(11). doi:10.3390/v12111242
181. Li Z, Liu J, You J, Li X, Liang Z, Du J. Proanthocyanidin Structure-Activity Relationship Analysis by Path Analysis Model. Int J Mol Sci. 2023;24(7).
182. Kamisuki S, Shibasaki H, Ashikawa K, et al. Determining the absolute configuration of vanitaracin A, an anti-hepatitis B virus agent. J Antibiot. 2022;75(2):92–97.
183. Gonçalves SMC, Silva GN, Pitta IDR, Rêgo M, Gnoato SCB, Pitta M. Novel betulin derivatives inhibit IFN-γ and modulates COX-2 expression. Nat Prod Res. 2020;34(12):1702–1711.
184. Kirstgen M, Lowjaga K, Müller SF, et al. Selective hepatitis B and D virus entry inhibitors from the group of pentacyclic lupane-type betulin-derived triterpenoids. Sci Rep. 2020;10(1):020–78618.
185. Pan W, Song IS, Shin HJ, et al. Genetic polymorphisms in Na+-taurocholate co-transporting polypeptide (NTCP) and ileal apical sodium-dependent bile acid transporter (ASBT) and ethnic comparisons of functional variants of NTCP among Asian populations. Xenobiotica fate foreig compou biologic sys. 2011;41(6):501–510. doi:10.3109/00498254.2011.555567
186. Vaz FM, Huidekoper HH, Paulusma CC. Extended Abstract: deficiency of Sodium Taurocholate Cotransporting Polypeptide (SLC10A1): a New Inborn Error of Metabolism with an Attenuated Phenotype. Digestive Diseas. 2017;35(3):259–260. doi:10.1159/000450984
187. Peng L, Zhao Q, Li Q, et al. The p.Ser267Phe variant in SLC10A1 is associated with resistance to chronic hepatitis B. Hepatology. 2015;61(4):1251–1260. doi:10.1002/hep.27608
188. Liang TJ. Hepatitis B: the virus and disease. Consensus Development Conference, NIHResearch Support, N I H, Intramural. Hepatology. 2009;49(5):22881.
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.