Back to Journals » Drug Design, Development and Therapy » Volume 19
Targeting THR-β for MASLD: Mechanisms and Drug Development
Authors Hua Y ![]() , Liu Y, Xing L, Yu X, Zheng P, Yang L, Song H
, Liu Y, Xing L, Yu X, Zheng P, Yang L, Song H ![]()
Received 4 August 2025
Accepted for publication 15 November 2025
Published 24 November 2025 Volume 2025:19 Pages 10473—10483
DOI https://doi.org/10.2147/DDDT.S558221
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Anastasios Lymperopoulos
Yunlong Hua,1,* Yang Liu,2,* Lianjun Xing,3 Xiao Yu,3 Peiyong Zheng,1 Lili Yang,1 Haiyan Song1
1Institute of Digestive Diseases, Longhua Hospital, Shanghai University of Traditional Chinese Medicine, Shanghai, 200032, People’s Republic of China; 2School of Integrative Medicine, Shanghai University of Traditional Chinese Medicine, Shanghai, 201203, People’s Republic of China; 3Department II of Digestive Diseases, Longhua Hospital, Shanghai University of Traditional Chinese Medicine, Shanghai, 200032, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Haiyan Song, Institute of Digestive Diseases, Longhua Hospital, Shanghai University of Traditional Chinese Medicine, 725 South Wanping Road, Shanghai, People’s Republic of China, Email [email protected] Lili Yang, Institute of Digestive Diseases, Longhua Hospital, Shanghai University of Traditional Chinese Medicine, 725 South Wanping Road, Shanghai, People’s Republic of China, Email [email protected]
Abstract: Metabolic dysfunction-associated steatotic liver disease (MASLD) has emerged as the most prevalent chronic liver disease worldwide, but there has long been a lack of effective therapeutic drugs. Thyroid hormone (TH) and its receptor THR-β play pivotal roles in hepatic metabolism, positioning THR-β as a promising therapeutic target for MASLD. Notably, Resmetirom, a selective THR-β agonist, gained FDA approval in 2024 for MASLD treatment, and several other THR-β agonists are currently undergoing preclinical or clinical studies. While these agents demonstrate effective in alleviating hepatic steatosis, inflammation, and fibrosis in MASLD, the disease heterogeneity and drugs’ adverse reactions remain key challenges. Therefore, further research is necessary to comprehensively assess their clinical efficacy and safety. This review summarizes the mechanisms by which TH/THR-β influences MASLD and recent advances in THR-β-targeted pharmacotherapy, aiming to enhance understanding of its therapeutic potential and promote drug development and clinical applications.
Keywords: thyroid hormone, thyroid hormone receptor-β, agonist, metabolic dysfunction-associated steatotic liver disease, metabolic dysfunction-associated steatohepatitis
Introduction
Metabolic dysfunction-associated steatotic liver disease (MASLD), previously named non-alcoholic fatty liver disease (NAFLD), is referred to liver manifestation of systemic metabolic symptom and has become the most prevalent chronic liver disease worldwide. A 2022 systematic meta-analysis reported a global prevalence of 32.4% (incidence: 46.9/1000 person-years).1 China is predicted to have 314 million MASLD cases by 2030.2 The disease spectrum ranges from simple steatosis to advanced metabolic dysfunction-associated steatohepatitis (MASH).3 The latter accounts for 10–30% of MASLD incidences and may progress to liver fibrosis, cirrhosis, and hepatocellular carcinoma.4 Notably, 15–25% of MASH patients develop fibrosis or cirrhosis within 10–15 years.5
The pathophysiology of MASLD involves complex network interactions among diverse cell types and signaling pathways. Initially explained by the “Two-hit” hypothesis,6 current understanding has evolved to the “Multiple parallel hits” model. Multiple factors including insulin resistance, imbalance of energy metabolism, lipotoxicity, dietary influences, gut microbiota, and genetic and epigenetic factors, inflammatory response, et al, jointly contribute to MASLD development.7
The complexity of the pathological mechanism challenges MASLD treatment. In the last decades, pharmaceutical development for MASLD/MASH has become a major research focus. As of 2024, about 700 investigational agents are in global pipelines, primarily consisting of small molecule compounds. Key therapeutic targets include Farnesoid X Receptor (FXR), glucagon-like peptide-1 (GLP-1), fibroblast growth factor (FGF), peroxisome proliferator-activated receptors (PPARs),8 and some emerging targets like iodothyronine deiodinase (DIO) 3, and thyroid hormone receptor-β (THR-β).9 However, up to now, only two drugs have been approved by the US Food and medicine Administration (FDA) for MASH treatment: the GLP-1 agonist semaglutide 3 and the THR-β agonist resmetirom, representing landmark breakthroughs in MASLD management.
This review summarizes the mechanisms and recent advancements of THR-β-targeted pharmacotherapy for MASLD, evaluating its therapeutic potential alongside existing challenges, with the aim of accelerating drug development and clinical applications.
Association Between Thyroid Hormone (TH), THR and MASLD
Action Pathway of TH/THR
TH is essential for organ development and metabolic homeostasis. Thyroid gland produces three kinds of hormones derived from tyrosine, 90% of them are tetraiodothyronine (T4), 9% are triiodothyronine (T3), and 1% are reverse triiodothyronine (rT3). Despite T4 being the predominant secretion, T3 exhibits fivefold more activity than T4 with a higher affinity for binding to THRs than T4. The minimal amount of rT3 is inactive.10 TH production is regulated by the hypothalamus–pituitary–thyroid (HPT) axis. The hypothalamus secretes thyrotropin-releasing hormone (TRH), which prompts the pituitary gland to release thyroid-stimulating hormone (TSH). Subsequently, TSH stimulates the thyroid gland to synthesize and release T4 and T3 into blood circulation. Concurrently, the release of TRH and TSH can be inhibited when TH reaches a higher concentration, establishing a negative feedback.10,11
TH is a lipophilic hormone, enabling it to traverse cell and nuclear membranes. Thus, T3 can directly access the nucleus, T4 can penetrate cytoplasm membrane and be transformed into T3 prior to nuclear entry.12 The TH transformation is governed by iodothyronine deiodinase (DIO), comprising DIO1, DIO2, and DIO3, which vary in their catalytic characteristics and tissue localization.13 DIO1 is expressed in thyroid, liver, and kidney, and facilitates the deiodination of both the phenol and tyrosine rings of T4. DIO2 exclusively catalyzes the deiodination of the phenol ring of T4, thereby contributing to T3 production in central nervous system, thyroid, skeletal muscle, and brown adipose tissues.14 DIO3 facilitates the deiodination of the tyrosine ring and is predominantly expressed in brain, placenta, and pancreas. It can inactivate T3, inhibit T4 activation, and convert T4 into rT3, serving as a physiological termination factor for TH activity.15 Importantly, in the context of MASLD, hepatic DIO3 expression and activity are significantly upregulated, a process triggered by oxidative stress and inflammation, which in turn leads to diminished local T3 levels and contributes to mitochondrial dysfunction and disease progression. Furthermore, therapeutic interventions such as ornithine aspartate have been shown to ameliorate MASLD in part by normalizing the aberrantly high expression and activity of DIO3, thereby restoring T3-mediated signaling and improving hepatic metabolism.9,16
By promoting the expression and nuclear translocation of intermediate factors like nuclear respiratory factor 1 and 2 (NRF1 and NRF2) to facilitate mitochondrial DNA (mtDNA) maintenance/transcription, mediating expression of the transcriptional coactivator peroxisome proliferator-activated receptor-gamma coactivator-1 α (PGC-1α), and stimulating mitochondrial protein synthesis via its mitochondrial matrix receptor p43, T3 orchestrates key transcriptional programs that drive mitochondrial function, including substrate oxidation, carbohydrate/lipid catabolism, increased the tricarboxylic acid cycle flux, and stimulation of mitochondrial respiration and oxidative phosphorylation.17
The regulation of target gene expression by T3 requires THR mediation.18 THR belongs to the superfamily of nuclear receptors, which are critical for development and homeostasis and major drug targets due to their involvement in some diseases like diabetes and obesity.19 These superfamily members generally contain a highly conserved DNA-binding domain to recognize highly similar DNA motifs, and a ligand-binding domain to bind natural ligands or xenobiotics.20 THR typically categorized into two subtypes: THR-α, predominantly expressed in cardiac and skeletal muscles, and THR-β, primarily expressed in liver, brain, and kidneys.21 The local availability and effect of TH varies among organs and cell types, potentially attributable to gating mechanisms including distinct TH-specific transporters like monocarboxylate transporter 8 (MCT8) for absorption, intracellular DIOs for TH conversion, and expression of THR subtype.22 In liver, DIO1 and THR-β are primarily involved in the regulation mechanism, while the role of various TH transporters remains ambiguous according to current studies.23 THR-β is the predominant isoform in hepatocytes. A study analyzing gene expression data related to TH uptake, conversion, and action in liver biopsy specimens indicated that THR-β expression correlated with serum triglycerides (TG) and glycosylated hemoglobin (HbA1c).24 Furthermore, THR-β was correlated with the severity of MASLD patients.
There are two principal pathways by which TH exerts regulatory effects through THR mediation (Figure 1). The first one is the classical pathway. Upon entry into nucleus, T3 associates with THR to create a hormone-receptor complex. THR subsequently undergoes polymerization to form either a homodimer or a heterodimer with the retinoic acid X receptor (RXR) in the nucleus. In the absence of TH, THR attaches to thyroid hormone response elements (TREs) in T3 target genes to repress their expression.25 Once the dimer formation, the co-repressor factors that were previously associated with THR and RXR separate, resulting in the activation of silent genes, which subsequently regulates the expression of target genes, and ultimately results in a number of biological effects.10,26 This regulatory mechanism, mediated through binding to TREs and directly orchestrating transcriptional programs, is classified as type 1 signaling pathway. The second one is the non-classical pathway, also called type 3 signaling pathway, that includes DNA/TRE-independent TR action, such as the phosphatidylinositol 3-kinase (PI3K) pathway signaling pathway, which is activated of intracellular signaling cascades. T3/THR exert physiological effect through activating the second messenger signaling pathway within cytoplasm. Negin P Martin et al discovered that in most Chinese hamster ovary cells in T3-free serum, over 50% of THR resided in the cytoplasm and functioned directly via PI3K pathway without binding DNA.27 G. Sebastian Hönes et al conducted experiment with TR knockout mice and demonstrating that the non-classical pathway can activate the PI3K signaling pathway via THR-β to modulate energy balance, blood glucose and triglyceride levels in mice.28
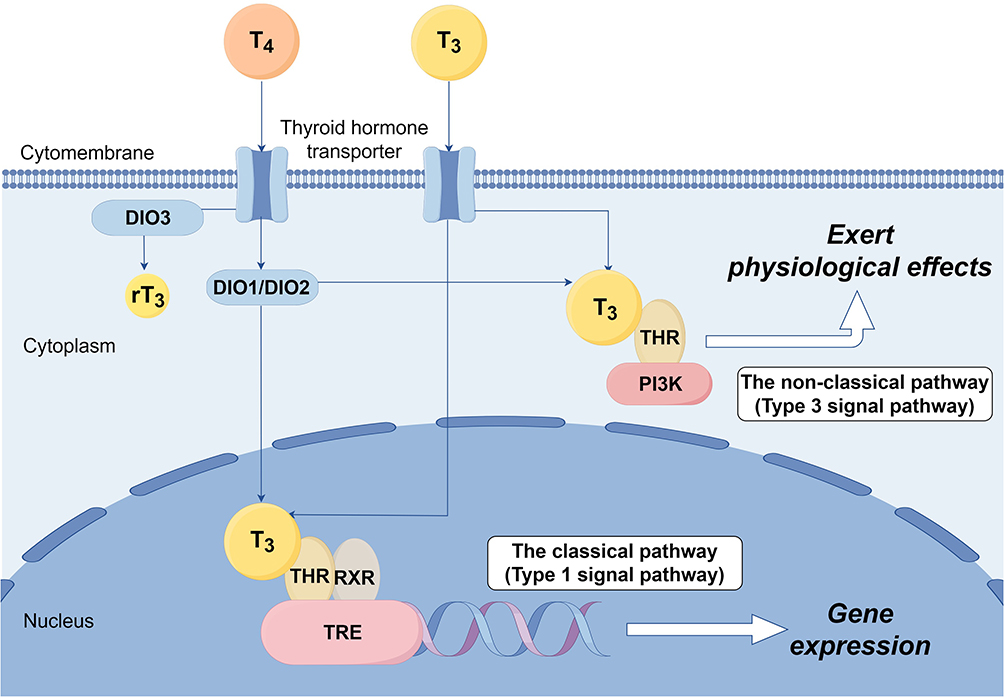 |
Figure 1 Action pathway of TH/THR. |
Correlation Between TH/THR Abnormalities and MASLD
The impact of TH on lipid metabolism was recorded over 130 years ago. In the 1930s, through measuring the lipid profiles, researchers discovered that serum total cholesterol (TC) and low-density lipoprotein cholesterol (LDL-c) were diminished in hyperthyroid patients, whereas they were elevated in those with subclinical and clinical hypothyroidism.29 Concurrently, serum triglyceride (TG) concentrations in hypothyroid patients were notably elevated, which could remain within normal range when TH levels were restored. Subsequent investigations have validated that aberrant TH levels might facilitate the onset and progression of metabolic disorders, including MASLD and hyperlipidemia.30 The reduction in T3 and T4 levels can diminish the enzyme activities associated with liver lipid metabolism, leading to impaired hepatic lipid clearance.31 A meta-analysis of cross-sectional and longitudinal studies indicated that, irrespective of different diagnostic criteria, there is a correlation between hypothyroidism and an elevated risk of MASLD, independent of prevalent metabolic risk factors.32 Researches also proved TH insufficiency could lead to liver diseases, ranging from liver steatosis to hepatocellular carcinoma (HCC).33 This was confirmed by epidemiological studies, which found that chronic hypothyroidism was positively associated with the occurrence rate of NAFLD and HCC, independent of other risk factors.34
A recent study investigated the association between TH sensitivity and fibrosis in MASLD. This study comprised 129 patients exhibiting normal thyroid function and biopsy-verified MASLD. Indicators of TH sensitivity were calculated. By using logistic regression, it was determined that, even after adjustment of gender, age, and metabolic parameters, T3 levels remained independently correlated with the advancement of liver fibrosis. Nevertheless, the causal relationship remained ambiguous.35 The study by Chung Ge et al indicated that individuals with loss-of-function mutations in THR-β gene have an elevated risk of TH resistance.36 Concurrently, Chaves et al discovered that these individuals also had a higher risk of hepatosteatosis.37 To elucidate the function of THR in lipid homeostasis, Araki et al created gene mutant mice with specific suppression of THR-α and THR-β. The findings indicated that mice with various THR mutations presented distinct metabolic consequences.38 In comparison to THR-α-inhibited mice, THR-β-inhibited mice exhibited elevated blood level of free fatty acids and TG, alongside excessive hepatic lipid deposition.
Regulatory Function of TH/THR-β Related to MASLD
The processes such as liver lipid and glucose metabolism, inflammatory response and fibrosis are closely contributed to MASLD development, and TH/THR- β has a significant regulatory effect on these processes. Researches indicate that TH and THR-β participate in the regulation of glycolysis, gluconeogenesis, glycogen synthesis, glycogenolysis, fatty acid uptake, lipid synthesis, lipid β-oxidation, mitochondrial autophagy and cholesterol synthesis.18,31
Regulation of Lipid Metabolism
Sterol regulatory element-binding protein-1c (SREBP-1c) is a major regulatory factor in fatty acid metabolism, playing a crucial role in transcriptional regulation of lipid synthesis genes. TH mediates its effect through TREs, which are repeats of a half-site consensus motif ((A/G)GG(T/A)CA),39 and functioned as a positive TRE (pTRE).40 Koshi Hashimoto et al demonstrated that the THR-β/RXR heterodimer binds to the novel negative TRE (nTRE) Site2 (GCCTGACAGGTGAAATCGGC), triggering transcriptional repression of SREBP-1c mRNA. This process enables T3 to negatively modulate the transcription of the SREBP-1c gene, consequently suppressing hepatic lipogenesis.41 Carnitine palmitoyltransferase-1 (CPT-1) influences the rate of fatty acid oxidation. Experiments have shown that THR-β interacts with TRE in the CPT-1α promoter, resulting in the upregulation of CPT-1 levels and the enhancement of fatty acid oxidation.42 THR-β can also interact with peroxisome proliferator-activated receptor α (PPARα), activate secondary transcription factors including FOXO1, promote autophagy and enhance β-oxidation of fatty acids.43 Sinha et al examined the impact of TH on cell death induced by palmitic acid (PA), and found that T3 can revert PA-induced lipotoxic injury via THR-β-mediated lipophagy and β-oxidation of fatty acids.44
A study revealed that LDL-c exhibited a significant correlation with proprotein convertase subtilisin/kexin type 9 (PCSK9).45 TH can diminish the PCSK9 level in circulation through its interaction with THR-β, consequently lowering LDL-c and alleviating lipid accumulation in liver. Moreover, researchers have discovered that a key hepatic metabolic regulator, fibroblast growth factor 21 (FGF21), is intricately associated with THR-β, RXR, and PPARα. Targeting THRβ with specific agonists represents a promising therapeutic strategy, as it induces FGF21 to stimulate insulin sensitivity, lipolysis and hepatic fatty acid oxidation.46
Regulation of Glycometabolism
TH is a crucial regulator of hepatic gluconeogenesis. It can stimulate the transcription of the essential gluconeogenic gene phosphoenolpyruvate carboxykinase (PCK1), which facilitates the conversion of pyruvate to phosphoenolpyruvate, a critical and tightly controlled phase in gluconeogenesis. TH also stimulates the transcriptional activation of glucose-6-phosphatase (G6PC), which facilitates the final phase of gluconeogenesis by converting glucose-6-phosphate into glucose.47 Sirtuin 1 (SIRT1) is a nuclear deacetylase that modulates lipid and glucose metabolism while augmenting mitochondrial function, hence conferring resistance to metabolic disorders.48 It has been revealed that TH promotes the transcription of PCK1 and G6PC genes via SIRT1-dependent deacetylation of FOXO1, which subsequently binds to the insulin response element in these gene promoters, activating transcription and enhancing expression.49
Regulation of Inflammatory Response and Fibrosis
In an investigation examining the impacts of T3 and THR-β agonist on steatohepatitis in rodents, Andrea Perra et al discovered that targeting THR-β can markedly impede the signal transduction of stress-activated protein kinase c-Jun N-terminal kinase (JNK), inhibiting the expression of cyclooxygenase-2, thereby ameliorating lipid accumulation and inflammatory response in murine liver tissues.50 Alonso-Merino et al found that liver fibrosis of mice induced by carbon tetrachloride was attenuated by TH administration, which inhibited the pro-fibrotic TGF-β/SMAD pathway.51
Research Advancements in THR-β-Related MASLD Pharmacotherapy
As mentioned above, TH/THR significantly influences numerous physiological processes, including glucose and lipid metabolism, which can benefit in ameliorating lipotoxic liver injury. This pushes THR agonists to be one of the prospective pharmacological targets for MASLD. However, it should be noted that, systemic TH replacement therapy is inappropriate for this disease, as it primarily affects THR-α signaling, potentially resulting in numerous adverse thyroid toxic reactions such as arrhythmias, osteopenia, sleep disturbances and other hyperthyroidism-like symptoms. Conversely, the incidence of adverse reactions associated with specific THR-β targeting drugs is comparatively low, rendering them favored in recent researches.52 Currently, there are over a dozen THR-β agonist drugs under development worldwide. But the research progression of them, such as CS27109, CS271011 and KB-141, has not been reported in public. Here, we described the research progress of several selective THR-β agonists that have been extensively reported in recent years (Table 1).
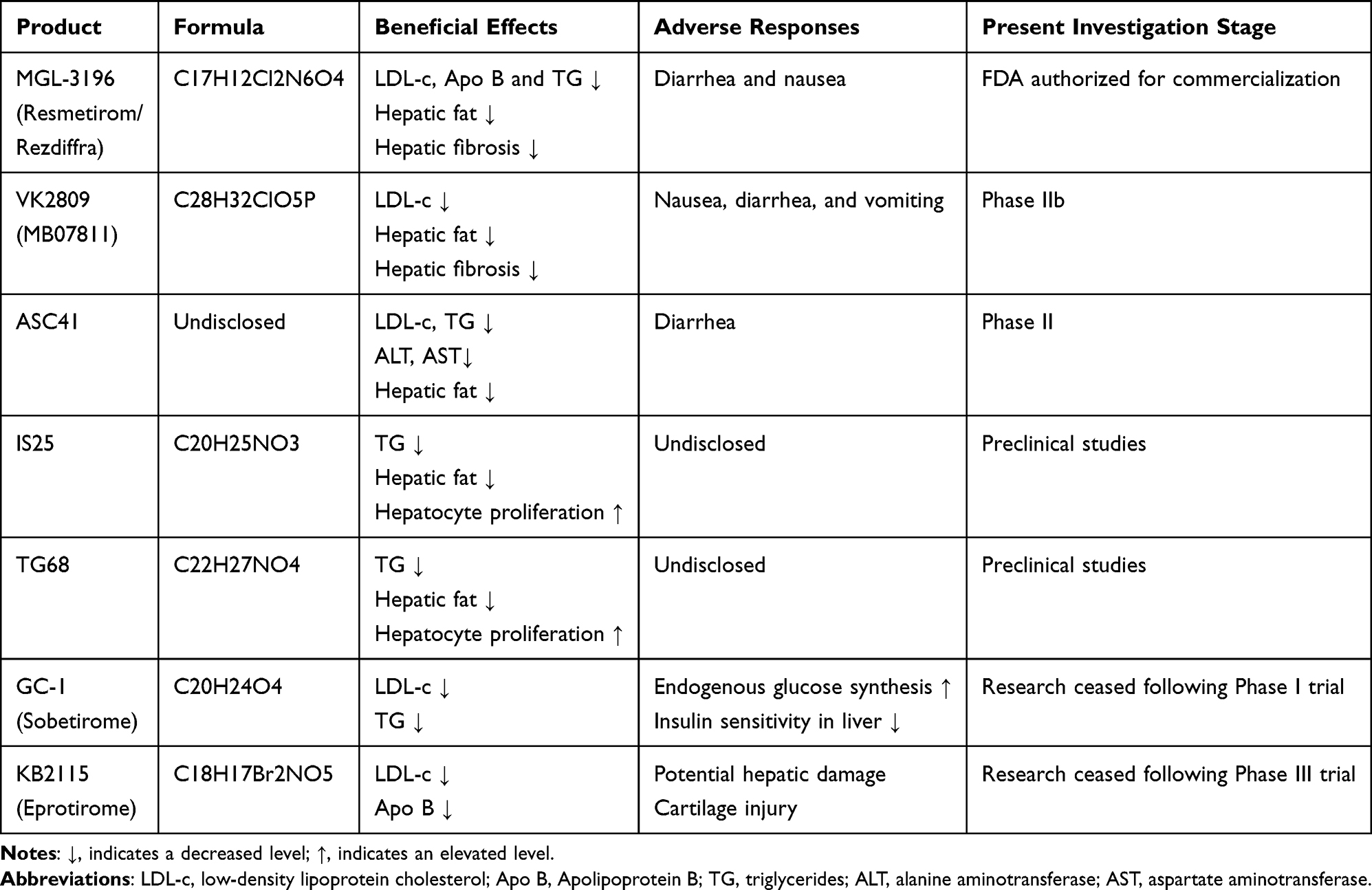 |
Table 1 THR-β-Related MASLD Pharmaceuticals |
THR-β Agonist Precursor
VK2809, originally named MB07811, is a prodrug with oral activity that acts as a phosphate-type THR agonist, produced by Viking Therapeutics. It is metabolized into a pharmacologically active form (3,5-dimethyl-4-(4′-hydroxy-3′-isopropylbenzyl) phenoxymethylphosphonic acid (named MB07344) by cytochrome P450 isozyme CYP3A4. CYP3A4 is primarily expressed in liver, thus resulting in liver-targeting characteristics of the drug. Its active form subsequently undergoes fast elimination through biliary excretion and exhibits minimal distribution level in other tissues.53 A preclinical investigation found that, VK2809 decreased serum cholesterol, serum and liver TG in diet-induced obese mice, without exerting a significant impact on body weight, blood glucose or TH level.54 The discovery endorses the clinical studies for this medication.
In a Phase I clinical trial, patients with mild hypercholesterolemia exhibited reduced levels of blood LDL-c and TG following VK2809 therapy, without any impact on heart rate or blood pressure. The findings indicated favorable safety and tolerability of the drug.55 In September 2016, a phase IIa clinical trial commenced to investigate the efficacy of VK2809 in patients with MASLD.56 The study concluded in April 2021, involving a total of 45 participants. The primary endpoint was the alteration in Magnetic Resonance Imaging-Proton Density Fat Fraction (MRI-PDFF) and plasma LDL-c level relative to baseline and placebo group. The patients administered VK2809 exhibited markedly reduced hepatic fat content compared to the placebo group. After 12 weeks, the median of relative decrease in liver lipid content was 53.8% for the 5 mg daily group, 59.7% for the 10 mg daily group, and 56.5% for the 10 mg every other day group, and the placebo group had a decrease of only 9.4%. Moreover, 70% patients in the therapy group exhibited a reduction in MRI-PDFF values of ≥50%. No significant adverse events were reported across all cohorts. A transient elevation of serum alanine aminotransferase (ALT) level has been observed by Esler et al in the early stage of drug administrations, but it restored to normal after 12 weeks.57
The encouraging outcomes endorse the Phase IIb VOYAGE study (NCT04173065), which assesses the efficacy of VK2809 (1.0 mg once daily, 2.5 mg once daily, 5 mg every other day, or 10 mg every other day) in treating biopsy-confirmed MASH and liver fibrosis in a broader patient cohort. It was a double-blind, placebo-controlled study, with a total treatment duration of 52 weeks. The trial employed MRI-PDFF-assessed liver fat reduction at 12 weeks as the primary endpoint. Secondary endpoints included histological changes in liver biopsy at week 52 and the proportion of patients with no worsening of liver fibrosis. After 12 weeks, an obvious decrease in liver fat was observed at all doses of VK2809 except 1.0 mg once daily. Patients experienced up to a 57% median reduction in MRI-PDFF from baseline. Importantly, these liver fat reductions were sustained or improved through week 52. Up to 88% of patients achieved a 30% or more decrease in liver fat after 52-week VK2809 treatment. Moreover, 75% of patients were found MASH improvement without worsening of fibrosis. At the highest 2 doses of VK2809, a significant proportion of patients achieved a 1-stage or higher improvement in fibrosis. In terms of safety, the severity of treatment-related adverse events reported in the VK2809 group was mostly mild or moderate (94%). The incidence of nausea, diarrhea, stool frequency, and vomiting in VK2809 group was similar to that of the placebo group.58 The high rates of liver fat and fibrosis improvement in MASH patients provide support for the Phase III trials of VK2809.
Selective THR-β Agonist
Sobetirome (GC-1) and Eprotirome (KB2115)
Sobetirome (GC-1) was a novel TH mimic synthesized by G. Chiellini et al in 1998. It is the first batch of liver selective THR-β agonist. The synthesis pathway is more efficient than conventional mimetics. It possesses potential characteristics of receptor binding and activation.59 However, in a Phase I clinical trial, researchers noted that GC-1 resulted in increased endogenous glucose production and diminished hepatic insulin sensitivity, prompting the cessation of future trials.60
Eprotirome (KB2115), another TH mimic, has shown beneficial effects on obesity and type 2 diabetes.61 It demonstrated advantageous effects on multiple lipid metabolic indices including levels of LDL-c, TG, and Apolipoprotein B (Apo B).62 However, in a Phase III clinical research, Eprotirome was found to induce variable degrees of hepatic damage. Simultaneously, toxicology investigations revealed that it may induce cartilage injury in canines (unpublished). The research was ultimately halted due to its side effects and adverse reactions.63
Resmetirom (MGL-3196)
Resmetirom (MGL-3196), produced by Madrigal Pharmaceuticals, is an oral, liver-targeted TH analog that preferentially binds to THR-β, promoting hepatic fat metabolism and diminishing lipotoxicity. It selectively binds to THR-β, thereby avoiding the side effects associated with THR – α. A Phase I clinical investigation assessed the safety and tolerability of Resmetirom. The trial examined the pharmacokinetics of Resmetirom at dosage ranging from 5 mg to 200 mg per day in healthy volunteers. The findings indicated its favorable tolerability and safety. Approximately the maximum effect was observed at a dose of 80 mg, LDL-c and Apo B decreased by about 30%, and TG decreased by about 60%.64
A further Phase II clinical trial recruited 125 patients with biopsy-confirmed MASH across 18 regions in the United States, and evaluated the impact of daily oral dosages of 80 ± 20 mg of Resmetirom compared to placebo after 12 and 36 weeks.65 Primary outcome was liver fat reduction evaluated by MRI-PDFF. After 12 weeks, patients with 80 mg Resmetirom administration exhibited a relative liver fat change of −32.9%, compared to −10.4% in the placebo group. After 36 weeks, the relative liver fat reduction rate for patients in Resmetirom group was −37.3%, while the placebo group showed −8.5%. The concentration of human type III collagen propeptide (PRO-C3) escalates with the progression of liver fibrosis and is frequently utilized to differentiate between MASLD and MASH.66 In this study, PRO-C3 levels in Resmetirom group were significantly diminished at both 12 and 36 weeks in comparison to the placebo group. Furthermore, Younossi et al assessed the alterations in health-related quality of life (HRQL) among individuals administered 80 mg of the medication daily in the phase II trial.67 It was found that patients in the therapy group with lower hepatic fat content also exhibited enhanced HRQL scores.
The positive preliminary outcomes triggered the initiation of Phase III clinical trials. Madrigal Pharmaceuticals has conducted 4 Phase III trials of Resmetirom: MAESTRO-NASH (to assess the efficacy and safety of Resmetirom in a substantial cohort of patients with pathologically confirmed MASH and fibrosis) (NCT03900429),68 MAESTRO-NAFLD-1 (to investigate the incidence of adverse effects and biomarkers during treatment) (NCT04197479),69 MAESTRO-NAFLD-OLE (a multicenter open-label extension study to evaluate the efficacy in approximately 1000 MAFLD patients) (NCT04951219),70 and the MAESTRO-NASH-OUTCOME (NCT05500222) study. The outcomes of the Phase III trials indicated a significant improvement in the NASH score and fibrosis progression. After 52 weeks, the percentage of MASH remission in patients administered Resmetirom 100 mg or 80 mg daily was 26% and 30%, respectively, significantly exceeding the 10% observed in the placebo group; the percentage of fibrosis improvement was 24% and 26%, respectively, also significantly surpassing the 14% in the placebo group.68 Concurrently, Younossi et al performed an additional HRQL evaluation.71 The findings indicated that, in contrast to placebo group, the patients treated with Resmetirom who attained a major histological response including MASH remission, fibrosis improvement, or either, exhibited substantial HRQL improvements. The findings further indicated that Resmetirom may offer extensive advantages to MASH patients with liver fibrosis.
Resmetirom demonstrates significant metabolic benefits, including reduction of hepatic lipid accumulation, lowering plasma lipids and lipoproteins that contribute to atherosclerosis, and improvement in quality-of-life metrics. On March 14, 2024, Resmetirom (trade name: Rezdiffra) obtained FDA accelerated approval for the treatment of adult non-cirrhotic non-alcoholic steatohepatitis (NASH) with advanced liver fibrosis (F2-F3 stage), making it the first innovative drug approved by FDA for MASH treatment. However, Resmetirom has shown limitations in clinical application, as it is only suitable for adult MASLD patients with moderate to severe liver fibrosis. Meanwhile, Adverse reactions related to liver toxicity and gallbladder have been observed during clinical trials. Therefore, its efficacy and safety still need further observation.
IS25 and TG68
Two novel THR-β selective agonists, IS25 and its prodrug TG68, have been recognized as promising candidates for the treatment of MASLD. A study conducted in vivo assessed the safety and efficacy of these two drugs in male F344 rats. Despite not being a rat model specifically for MASLD, the researchers noted a substantial reduction in TG levels.72 These two compounds may function by stimulating the adenosine 5′-monophosphate (AMP)-activated protein kinase (AMPK)/acetyl CoA carboxylase (ACC) signaling pathway, which is pivotal in cellular energy management and metabolism.73 Studies have found IS25 and TG68 could induce a significant elevation of p-AMPK/AMPK and p-ACC/ACC protein expression ratios at the higher concentration such as 10 μM.72
A study evaluated the therapeutic effects of TG68 and Resmetirom in obese mice induced by a high-fat diet. The results demonstrated that, both drugs could activate the THR-β target genes, yielding comparable beneficial outcomes including reducing liver weight, hepatic steatosis, and circulating TG without inducing significant extrahepatic damage.74 A study by Andrea Caddo et al further revealed that, whereas Resmetirom diminished hepatic steatosis, TG68 at an equivalent dosage was capable of regressing diethylnitrosamine-induced precancerous lesions and exhibited a notable anti-tumor effect, which Resmetirom did not demonstrate.75 Another study shows that both TG68 and IS25 are able to increase hepatocyte proliferation, in a way quite similar to the thyroid hormone T3, but without overt toxicity.76 These preliminary research data for IS25 and TG68 suggest significant potential for treating MASLD. Currently, they are in urgent need for further basic and clinical researches.
ASC41
ASC41, produced by Ascletis Pharmaceuticals, is the THR-β selective agonist medicine with the most rapid development in China. Ascletis conducted a Phase I randomized, double-blind, placebo-controlled clinical investigation in 2021, involving both single-dose and multiple-dose escalation.77,78 The experiment comprised 65 participants with MASLD, with dosages varying from 1 mg to 20 mg. In the highest dosage group of 20 mg, ASC41 demonstrated good tolerance and safety. Concurrently, in comparison to the placebo group, the drug-treated patients exhibited a significant reduction in LDL-c and TG levels, with both single-dose and multiple-dose escalations yielding equivalent benefits.
In 2022, Ascletis’ Ganlai Company conducted a double-blind, randomized, multicenter, placebo-controlled Phase II clinical research to further assess the safety, tolerability, and efficacy of ASC41. The experiment comprised 180 patients with liver biopsy-confirmed MASH and met the requirement of liver fat content ≥7.5% and hepatic fibrosis stage F2 and F3 evaluated by MRI-PDFF. Participants were randomly assigned to two treatment groups (2 mg and 4 mg) of ASC41 administered orally once daily, along with a placebo control group. Liver biopsies were conducted at baseline and 52 weeks, whereas MRI-PDFF evaluations were carried out at baseline, 12 weeks, and 52 weeks thereafter. The principal endpoint was a decrease of ≥2 points in the MASLD activity score (indicating improvement in inflammation or ballooning) without any deterioration of fibrosis. In 2024, Ganlai exhibited that ASC41 can markedly diminish liver fat, alanine aminotransferase, and aspartate aminotransferase, while exhibiting overall favorable tolerability and safety.79 This indicates that ASC41 possesses adequate therapeutic potential. Nonetheless, additional clinical trials are necessary for further validation.
Conclusion
Acculmulating preclinical and clinical researches have indicated that THR-β selective agonists can alleviate hepatic steatosis, inflammation and fibrosis in MASLD, without the adverse reactions to cardiovascular and central thyroid axis control. Especially, Resmetirom has made the first breakthrough in MASH clinical treatment since decades, encouraging progress of THR-β-targeted drugs. However, the disease heterogeneity and bias in clinical trial evaluation criteria may substantially affect clinical trial outcomes. Moreover, drugs’ side effects, including but not only common gastrointestinal adverse reactions and potential liver toxicity, challenge their clinical applications. On the other hand, combination therapy is currently considered as the future potential for MASH treatment by many experts. As lipid accumulation in liver crucially affects MASH development and fibrosis, effective combination therapy must incorporate at least one lipid-lowering agent.80 Thus, even some THR-β agonists previously discontinued or failed in clinical trials still have the opportunity. In summary, the THR-β targeted drugs have already demonstrated promising therapeutic potential for MASLD. But there is an urgent need for further preclinical and clinical studies to elucidate their efficacies and adverse reactions, thereby facilitating the precision medicine with improved clinical outcomes for MASLD patients.
Acknowledgments
Figure 1 was drawn through Figdraw (www.figdraw.com).
Funding
This work was supported by National Natural Science Foundation of China (82374417), Natural Science Foundation of Shanghai (24ZR1466900), Shanghai Medical Innovation Research Special Project (24Y12801100) and Scientific Research Project of Shanghai Municipal Science and Technology Commission (22Y11921500). The authors declare that financial support was received for the research or publication of this article.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Riazi K, Azhari H, Charette JH, et al. The prevalence and incidence of NAFLD worldwide: a systematic review and meta-analysis. Lancet Gastroenterol Hepatol. 2022;7(9):851–861. doi:10.1016/S2468-1253(22)00165-0
2. Zhou J, Zhou F, Wang W, et al. Epidemiological features of NAFLD from 1999 to 2018 in China. Hepatology. 2020;71(5):1851–1864. doi:10.1002/hep.31150
3. Dolgin E. NASH therapies head toward landmark approval. Nat Biotechnol. 2023;41(5):587–590. doi:10.1038/s41587-023-01787-8
4. Younossi ZM, Koenig AB, Abdelatif D, Fazel Y, Henry L, Wymer M. Global epidemiology of nonalcoholic fatty liver disease—Meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology. 2016;64(1):73–84. doi:10.1002/hep.28431
5. National Workshop on Fatty Liver and Alcoholic Liver Disease CSOH. Guideline of prevention and treatment for nonalcoholic fatty liver disease: a 2018 update. J Pract Hepatol. 2018;21:177–186.
6. Ying LI, Hong ZF. Hypothesis of “Two Hits” in NAFLD. Med Recapitulate. 2013;19(4):594–596.
7. Tilg H, Adolph TE, Moschen AR. Multiple parallel hits hypothesis in nonalcoholic fatty liver disease: revisited after a decade. Hepatology. 2021;73(2):833–842. doi:10.1002/hep.31518
8. Hanhui X, Jinsheng J, Pei C, et al. Research progress in the current treatment and drugs to popular targets for meta-bolic dysfunction-associated steatotic liver disease. Pract Pharm Clin Remedies. 2024;27(5):374–379.
9. Longo L, Marschner RA, de Freitas LBR, et al. Redefining the role of ornithine aspartate and vitamin E in metabolic dysfunction-associated steatotic liver disease through its biochemical properties. Int J Mol Sci. 2024;25(13):6839. doi:10.3390/ijms25136839
10. Wong T. Physiology.
11. Sinha RA, Bruinstroop E, Yen PM. Actions of thyroid hormones and thyromimetics on the liver. Nat Rev Gastroenterol Hepatol. 2024;22(1):9–22. doi:10.1038/s41575-024-00991-4
12. Beiyu C, Yiyang H, Qi F. New development for targeted therapy of non-alcoholic fatty liver disease with thyroid hormone receptor beta. Chin J Hepatol. 2021;29(8):807–811. doi:10.3760/cma.j.cn501113-20190520-00178
13. Luongo C, Dentice M, Salvatore D. Deiodinases and their intricate role in thyroid hormone homeostasis. Nat Rev Endocrinol. 2019;15(8):479–488. doi:10.1038/s41574-019-0218-2
14. Bianco AC, Salvatore D, Gereben B, Berry MJ, Larsen PR. Biochemistry, cellular and molecular biology, and physiological roles of the iodothyronine selenodeiodinases. Endocr Rev. 2002;23(1):38–89. doi:10.1210/edrv.23.1.0455
15. Piantanida E, Ippolito S, Gallo D, et al. The interplay between thyroid and liver: implications for clinical practice. J Endocrinol Invest. 2020;43(7):885–899. doi:10.1007/s40618-020-01208-6
16. Marschner RA, Roginski AC, Ribeiro RT, Longo L, Álvares-da-Silva MR, Wajner SM. Uncovering actions of type 3 deiodinase in the Metabolic Dysfunction-Associated Fatty Liver Disease (MAFLD). Cells. 2023;12(7):1022. doi:10.3390/cells12071022
17. Tawfik I, Gottschalk B, Jarc A, et al. T3-induced enhancement of mitochondrial Ca2+ uptake as a boost for mitochondrial metabolism. Free Radic Biol Med. 2022;181:197–208. doi:10.1016/j.freeradbiomed.2022.01.024
18. Hatziagelaki E, Paschou SA, Schön M, Psaltopoulou T, Roden M. NAFLD and thyroid function: pathophysiological and therapeutic considerations. Trends Endocrinol Metab. 2022;33(11):755–768. doi:10.1016/j.tem.2022.08.001
19. Gronemeyer H, Gustafsson JA, Laudet V. Principles for modulation of the nuclear receptor superfamily. Nat Rev Drug Discov. 2004;3(11):950–964. doi:10.1038/nrd1551
20. Dubois V, Lefebvre P, Staels B, Eeckhoute J. Nuclear receptors: pathophysiological mechanisms and drug targets in liver disease. Gut. 2024;73(9):1562–1569. doi:10.1136/gutjnl-2023-331741
21. Delitala AP, Delitala G, Sioni P, Fanciulli G. Thyroid hormone analogs for the treatment of dyslipidemia: past, present, and future. Curr Med Res Opin. 2017;33(11):1985–1993. doi:10.1080/03007995.2017.1330259
22. Friesema EC, Jansen J, Milici C, Visser TJ. Thyroid hormone transporters. Vitam Horm. 2005;70:137–167.
23. Yen PM. Physiological and molecular basis of thyroid hormone action. Physiol Rev. 2001;81(3):1097–1142. doi:10.1152/physrev.2001.81.3.1097
24. Krause C, Grohs M, El Gammal AT, et al. Reduced expression of thyroid hormone receptor β in human nonalcoholic steatohepatitis. Endocr Connect. 2018;7(12):1448–1456. doi:10.1530/EC-18-0499
25. Cheng SY. Multiple mechanisms for regulation of the transcriptional activity of thyroid hormone receptors. Rev Endocr Metab Disord. 2000;1(1–2):9–18. doi:10.1023/A:1010052101214
26. Xu R. Advances in research on nuclear receptor families. Acad J Second Mil Med Univ. 2001;22(11):2.
27. Martin NP, Marron FDVE, Mizuno F, et al. A rapid cytoplasmic mechanism for PI3 kinase regulation by the nuclear thyroid hormone receptor, TRβ, and genetic evidence for its role in the maturation of mouse hippocampal synapses in vivo. Endocrinology. 2014;155(9):3713–3724. doi:10.1210/en.2013-2058
28. Hönes GS, Geist D, Moeller LC. Noncanonical Action of Thyroid Hormone Receptors α and β. Exp Clin Endocrinol Diabetes. 2020;128(6–7):383–387. doi:10.1055/a-1088-1187
29. Duntas LH, Brenta G. A renewed focus on the association between thyroid hormones and lipid metabolism. Front Endocrinol. 2018;9:511.
30. Al Doghaither HA, Alshaikh EM, Omar UM, et al. Insulin resistance and its correlation with chemerin and visfatin in Saudi patients with hyperthyroidism. Int J Health Sci. 2019;13(5):18–21.
31. Sinha RA, Singh BK, Yen PM. Thyroid hormone regulation of hepatic lipid and carbohydrate metabolism. Trends Endocrinol Metab. 2014;25(10):538–545. doi:10.1016/j.tem.2014.07.001
32. Mantovani A, Nascimbeni F, Lonardo A, et al. Association between primary hypothyroidism and nonalcoholic fatty liver disease: a systematic review and meta-analysis. Thyroid. 2018;28(10):1270–1284. doi:10.1089/thy.2018.0257
33. Chi HC, Tsai CY, Tsai MM, Yeh CT, Lin KH. Molecular functions and clinical impact of thyroid hormone-triggered autophagy in liver-related diseases. J Biomed Sci. 2019;26(1):24. doi:10.1186/s12929-019-0517-x
34. Lee J, Ha J, Jo K, et al. Male-specific association between subclinical hypothyroidism and the risk of non-alcoholic fatty liver disease estimated by hepatic steatosis index: Korea National Health and Nutrition Examination Survey 2013 to 2015. Sci Rep. 2018;8(1):15145. doi:10.1038/s41598-018-32245-0
35. Li R, Zhou L, Chen C, et al. Sensitivity to thyroid hormones is associated with advanced fibrosis in euthyroid patients with non-alcoholic fatty liver disease: a cross-sectional study. Dig Liver Dis. 2023;55(2):254–261. doi:10.1016/j.dld.2022.06.021
36. Chung GE, Kim D, Kim W, et al. Non-alcoholic fatty liver disease across the spectrum of hypothyroidism. J Hepatol. 2012;57(1):150–156. doi:10.1016/j.jhep.2012.02.027
37. Chaves C, Bruinstroop E, Refetoff S, Yen PM, Anselmo J. Increased hepatic fat content in patients with resistance to thyroid hormone beta. Thyroid. 2021;31(7):1127–1134. doi:10.1089/thy.2020.0651
38. Araki O, Ying H, Zhu XG, Willingham MC, Cheng SY. Distinct dysregulation of lipid metabolism by unliganded thyroid hormone receptor isoforms. Mol Endocrinol. 2009;23(3):308–315. doi:10.1210/me.2008-0311
39. Cohen O, Flynn TR, Wondisford FE. Ligand-dependent antagonism by Retinoid X receptors of inhibitory thyroid hormone response elements. J Biol Chem. 1995;270(23):13899–13905. doi:10.1074/jbc.270.23.13899
40. Umesono K, Murakami KK, Thompson CC, Evans RM. Direct repeats as selective response elements for the thyroid hormone, retinoic acid, and vitamin D3 receptors. Cell. 1991;65(7):1255–1266.
41. Hashimoto K, Yamada M, Matsumoto S, Monden T, Satoh T, Mori M. Mouse sterol response element binding protein-1c gene expression is negatively regulated by thyroid hormone. Endocrinology. 2006;147(9):4292–4302. doi:10.1210/en.2006-0116
42. Jackson-Hayes L, Song S, Lavrentyev EN, et al. A thyroid hormone response unit formed between the promoter and first intron of the carnitine palmitoyltransferase-Ialpha gene mediates the liver-specific induction by thyroid hormone. J Biol Chem. 2003;278(10):7964–7972. doi:10.1074/jbc.M211062200
43. Musso G, Gambino R, Cassader M. Cholesterol metabolism and the pathogenesis of non-alcoholic steatohepatitis. Prog Lipid Res. 2013;52(1):175–191. doi:10.1016/j.plipres.2012.11.002
44. Sinha RA, Yen PM. Thyroid hormone-mediated autophagy and mitochondrial turnover in NAFLD. Cell Biosci. 2016;6(1). doi:10.1186/s13578-016-0113-7
45. Bonde Y, Breuer O, Lütjohann D, Sjöberg S, Angelin B, Rudling M. Thyroid hormone reduces PCSK9 and stimulates bile acid synthesis in humans. J Lipid Res. 2014;55(11):2408–2415. doi:10.1194/jlr.M051664
46. Adams AC, Astapova I, Fisher FM, et al. Thyroid hormone regulates hepatic expression of fibroblast growth factor 21 in a PPARalpha-dependent manner. J Biol Chem. 2010;285(19):14078–14082. doi:10.1074/jbc.C110.107375
47. Feng X, Jiang Y, Meltzer P, Yen PM. Thyroid hormone regulation of hepatic genes in vivo detected by complementary DNA microarray. Mol Endocrinol. 2000;14(7):947–955. doi:10.1210/mend.14.7.0470
48. Lagouge M, Argmann C, Gerhart-Hines Z, et al. Resveratrol improves mitochondrial function and protects against metabolic disease by activating SIRT1 and PGC-1alpha. Cell. 2006;127(6):1109–1122. doi:10.1016/j.cell.2006.11.013
49. Singh BK, Sinha RA, Zhou J, et al. FoxO1 deacetylation regulates thyroid hormone-induced transcription of key hepatic gluconeogenic genes. J Biol Chem. 2013;288(42):30365–30372. doi:10.1074/jbc.M113.504845
50. Perra A, Simbula G, Simbula M, et al. Thyroid hormone (T3) and TRbeta agonist GC-1 inhibit/reverse nonalcoholic fatty liver in rats. FASEB J. 2008;22(8):2981–2989. doi:10.1096/fj.08-108464
51. Alonso-Merino E, Martín Orozco R, Ruíz-Llorente L, et al. Thyroid hormones inhibit TGF-β signaling and attenuate fibrotic responses. Proc Natl Acad Sci. 2016;113(24). doi:10.1073/pnas.1506113113
52. Sinha RA, Bruinstroop E, Singh BK, Yen PM. Nonalcoholic fatty liver disease and hypercholesterolemia: roles of thyroid hormones, metabolites, and agonists. Thyroid. 2019;29(9):1173–1191. doi:10.1089/thy.2018.0664
53. Zhou J, Waskowicz LR, Lim A, et al. A Liver-Specific Thyromimetic, VK2809, Decreases Hepatosteatosis in Glycogen Storage Disease Type Ia. Thyroid. 2019;29(8):1158–1167. doi:10.1089/thy.2019.0007
54. Erion MD, Cable EE, Ito BR, et al. Targeting thyroid hormone receptor-beta agonists to the liver reduces cholesterol and triglycerides and improves the therapeutic index. Proc Natl Acad Sci U S A. 2007;104(39):15490–15495. doi:10.1073/pnas.0702759104
55. Meruvu S, Ayers SD, Winnier G, Webb P. Thyroid hormone analogues: where do we stand in 2013? Thyroid. 2013;23(11):1333–1344. doi:10.1089/thy.2012.0458
56. Loomba R, Neutel J, Mohseni R, et al. VK2809, a novel liver-directed thyroid receptor beta agonist, significantly reduces liver fat with both low and high doses in patients with non-alcoholic fatty liver disease: a phase 2 randomized, placebo-controlled trial. J Hepatol. 2019;701(1):E150–E151. doi:10.1016/S0618-8278(19)30266-X
57. Esler WP, Bence KK. Metabolic targets in nonalcoholic fatty liver disease. Cell Mol Gastroenterol Hepatol. 2019;8(2):247–267. doi:10.1016/j.jcmgh.2019.04.007
58. Alkhouri N. Results from the 52-week phase 2b VOYAGE trial of VK2809 in patients with BiopsyConfirmed non-alcoholic steatohepatitis and fibrosis: a randomized, placebo-controlled trial. Gastroenterol Hepatol. 2024;20(12 Suppl 11):13–14.
59. Chiellini G, Apriletti JW, Yoshihara HA, Baxter JD, Ribeiro RC, Scanlan TS. A high-affinity subtype-selective agonist ligand for the thyroid hormone receptor. Chem Biol. 1998;5(6):299–306. doi:10.1016/S1074-5521(98)90168-5
60. Vatner DF, Weismann D, Beddow SA, et al. Thyroid hormone receptor-β agonists prevent hepatic steatosis in fat-fed rats but impair insulin sensitivity via discrete pathways. Am J Physiol Endocrinol Metab. 2013;305(1):E89–E100. doi:10.1152/ajpendo.00573.2012
61. Baxter JD, Webb P. Thyroid hormone mimetics: potential applications in atherosclerosis, obesity and type 2 diabetes. Nat Rev Drug Discov. 2009;8(4):308–320. doi:10.1038/nrd2830
62. Berkenstam A, Kristensen J, Mellström K, et al. The thyroid hormone mimetic compound KB2115 lowers plasma LDL cholesterol and stimulates bile acid synthesis without cardiac effects in humans. Proc Natl Acad Sci U S A. 2008;105(2):663–667. doi:10.1073/pnas.0705286104
63. Sjouke B, Langslet G, Ceska R, et al. Eprotirome in patients with familial hypercholesterolaemia (the AKKA trial): a randomised, double-blind, placebo-controlled Phase 3 study. Lancet Diabetes Endocrinol. 2014;2(6):455–463. doi:10.1016/S2213-8587(14)70006-3
64. Taub R, Chiang E, Chabot-Blanchet M, et al. Lipid lowering in healthy volunteers treated with multiple doses of MGL-3196, a liver-targeted thyroid hormone receptor-β agonist. Atherosclerosis. 2013;230(2):373–380. doi:10.1016/j.atherosclerosis.2013.07.056
65. Harrison SA, Bashir MR, Guy CD, et al. Resmetirom (MGL-3196) for the treatment of non-alcoholic steatohepatitis: a multicentre, randomised, double-blind, placebo-controlled, phase 2 trial. Lancet. 2019;394(10213):2012–2024. doi:10.1016/S0140-6736(19)32517-6
66. Nielsen MJ, Leeming DJ, Goodman Z, et al. Comparison of ADAPT, FIB-4 and APRI as non-invasive predictors of liver fibrosis and NASH within the CENTAUR screening population. J Hepatol. 2021;75(6):1292–1300. doi:10.1016/j.jhep.2021.08.016
67. Younossi ZM, Stepanova M, Taub RA, Barbone JM, Harrison SA. Hepatic fat reduction due to resmetirom in patients with nonalcoholic steatohepatitis is associated with improvement of quality of life. Clin Gastroenterol Hepatol. 2022;20(6):1354–1361. doi:10.1016/j.cgh.2021.07.039
68. Harrison SA, Bedossa P, Guy CD, et al. A phase 3, randomized, controlled trial of resmetirom in NASH with liver fibrosis. N Engl J Med. 2024;390(6):497–509. doi:10.1056/NEJMoa2309000
69. Harrison SA, Taub R, Neff GW, et al. Resmetirom for nonalcoholic fatty liver disease: a randomized, double-blind, placebo-controlled phase 3 trial. Nat Med. 2023;29(11):2919–2928. doi:10.1038/s41591-023-02603-1
70. Harrison SA, Ratziu V, Anstee QM, et al. Design of the phase 3 MAESTRO clinical program to evaluate resmetirom for the treatment of nonalcoholic steatohepatitis. Aliment Pharmacol Ther. 2024;59(1):51–63. doi:10.1111/apt.17734
71. Younossi ZM, Stepanova M, Racila A, et al. Health-related quality of life (HRQL) assessments in a 52-week, double-blind, randomized, placebo-controlled phase III study of resmetirom (MGL-3196) in patients with metabolic dysfunction–associated steatohepatitis (MASH) and fibrosis. Hepatology. 2025;81(4):1318–1327. doi:10.1097/HEP.0000000000001084
72. Runfola M, Sestito S, Bellusci L, et al. Design, synthesis and biological evaluation of novel TRβ selective agonists sustained by ADME-toxicity analysis. Eur J Med Chem. 2020;188:112006. doi:10.1016/j.ejmech.2019.112006
73. Gasparrini M, Giampieri F, Alvarez SJM, et al. AMPK as a new attractive therapeutic target for disease prevention: the role of dietary compounds AMPK and disease prevention. Curr Drug Targets. 2016;17(8):865–889. doi:10.2174/1573399811666150615150235
74. Caddeo A, Kowalik MA, Serra M, et al. TG68, a novel thyroid hormone receptor-β agonist for the treatment of NAFLD. Int J Mol Sci. 2021;22(23):13105. doi:10.3390/ijms222313105
75. Caddeo A, Serra M, Sedda F, et al. Potential use of TG68 - A novel thyromimetic - for the treatment of non-alcoholic fatty liver (NAFLD)-associated hepatocarcinogenesis. Front Oncol. 2023;13:1127517. doi:10.3389/fonc.2023.1127517
76. Perra A, Kowalik MA, Cabras L, et al. Potential role of two novel agonists of thyroid hormone receptor‐β on liver regeneration. Cell Prolif. 2020;53(5):e12808. doi:10.1111/cpr.12808
77. Ascletis Pharma Inc. DDI study of ASC41 in HV and the PK, safety and tolerability in subjects with NAFLD. Available from: https://www.clinicaltrials.gov/study/NCT04845646?term=gannex&viewType=Table&rank=7.
78. Ascletis Pharma Inc. A study of the relative bioavailability of ASC41 in healthy subjects. Available from: https://www.clinicaltrials.gov/study/NCT04527250?term=ascletis&viewType=Table&page=4&rank=32.
79. Ascletis Pharma Inc. Study to evaluate the safety, tolerability, and efficacy of ASC41 tablets in adult patients with NASH. Available from: https://www.clinicaltrials.gov/study/NCT05462353?term=gannex&viewType=Table&rank=2#study-record-dates.
80. Alkhouri N. Thyromimetics as emerging therapeutic agents for nonalcoholic steatohepatitis: rationale for the development of resmetirom (MGL-3196). Expert Opin Investig Drugs. 2020;29(2):99–101. doi:10.1080/13543784.2020.1708899
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.