Back to Journals » International Journal of Nanomedicine » Volume 21
Targeting the Epithelial Alarmin Axis with Biomedical Nanoparticles: A New Frontier in Allergic Asthma Therapy
Authors Yang Y ![]()
Received 13 December 2025
Accepted for publication 23 March 2026
Published 31 March 2026 Volume 2026:21 588680
DOI https://doi.org/10.2147/IJN.S588680
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr RDK Misra
Yifei Yang
School of Basic Medicine, Chongqing Medical University, Chongqing, People’s Republic of China
Correspondence: Yifei Yang, School of Basic Medicine, Chongqing Medical University, Chongqing, People’s Republic of China, Tel +86-182-1773-5013, Email [email protected]
Abstract: Allergic asthma is a chronic inflammatory airway disease driven by type 2 immune responses, whose pathogenesis correlates with the release of epithelial alarm proteins—thymic stromal lymphopoietin (TSLP), interleukin-25 (IL-25), and interleukin-33 (IL-33)—by airway epithelial cells following barrier injury. This paper systematically reviews the cutting-edge applications of nanoparticles (NPs) in targeting the epithelial alarmin signaling axis and its downstream immune cells, including dendritic cells, macrophages, Th2 cells, and regulatory T cells. By systematically reviewing research progress on nanoparticles in allergic asthma treatment, this review provides crucial theoretical support and technical frameworks for developing precise, efficient, and less-side-effect asthma therapies. It offers forward-looking guidance for advancing asthma treatment from laboratory to clinical translation.
Keywords: nanoparticles, allergic asthma, epithelial alarmins, immunomodulation, MSC-exosomes, DNAzyme nanocapsules
Introduction
Asthma, a chronic noncommunicable disease characterized by airway inflammation, is gradually posing a global threat. According to projections from the 2021 Global Burden of Disease (GBD) study, the number of asthma patients worldwide has reached 260 million and is projected to rise to 275 million by 2050.1 Its clinical symptoms primarily manifest as chest tightness, shortness of breath, and recurrent wheezing, which can be fatal in severe cases. However, given the complex pathophysiology of this heterogeneous disease, existing treatment regimens can only alleviate rather than cure patients’ symptoms. Current clinical asthma management strategies primarily rely on combination therapy with corticosteroids and bronchodilators. Symptoms are controlled through anti-inflammatory responses induced by inhaled corticosteroids (ICS), long-acting beta-agonists (LABA), and leukotriene antagonists. Among these, ICS use is universally recommended in the Global Initiative for Asthma (GINA) guidelines as the first-line treatment for all patients with persistent asthma.2 In fact, frequent or excessive use of steroids may lead to certain systemic side effects and adverse reactions.3–5 Therefore, further elucidating the pathogenesis of asthma and developing more precise and effective treatment approaches have become the two primary objectives of current asthma research.
Allergic asthma is the most common asthma phenotype across all studies. Its pathogenesis is primarily driven by a type 2 immune response triggered by inhaled airborne allergens (such as pollen and fungal spores), which produces type 2 helper T (Th2) cytokines leading to airway edema, excessive mucus secretion, and bronchial hyperresponsiveness.6 Over the past two decades, translational research has identified numerous therapeutic targets for asthma associated with type 2 immune responses, expanding treatment options for asthma management. Furthermore, the recently proposed epithelial barrier hypothesis suggests that many allergens can cause lesions in affected skin and mucosal tissues by disrupting the epithelial barrier, triggering various allergic inflammatory diseases such as asthma.7 Extensive research demonstrates that in allergic asthma, airway epithelial cells (AECs) respond to compromised barriers by releasing three epithelial cytokines known as “alarm proteins,” thereby mediating a complex Type 2 inflammatory cascade. This discovery not only clarifies part of the mechanism by which allergens initiate Type 2 immune responses but also provides novel therapeutic targets for asthma treatment.
To date, biologic agents such as human monoclonal antibodies (eg., omalizumab, mepolizumab) have been used to treat moderate-to-severe refractory asthma.8 However, the significant variation in clinical response to treatment and its limitations have compelled contemporary researchers to seek new avenues for asthma management.9,10 Today, with the widespread application of nanomedicine and the rapid growth of the nanomedicine market, biomedical nanoparticles (NPs) have garnered significant attention in drug delivery and immunomodulation due to their multifunctionality, high biocompatibility, and potential for advanced therapeutic applications. This paper will focus on discussing the role of epithelial alarmins in type 2 immune responses and the progress of biomedical NPs in their mediation of allergic asthma.
Alarmins
Epithelial alarmins are a class of endogenous danger signaling molecules rapidly released by epithelial cells upon sensing danger signals such as tissue injury, infection, or stress. They constitute an important component of damage-associated molecular patterns (DAMPs). Typical epithelial alarmins include TSLP, IL-25, and IL-33. Produced by healthy activated immune cells and secreted via the endoplasmic reticulum-Golgi pathway or non-classical routes (such as non-programmed cell death), they play crucial roles as initiators and participants in various physiological and pathophysiological processes, including inflammation, infection, and tumor immunity.11,12 Under steady-state conditions, they are typically confined within cells. However, upon cellular injury, necrosis, or pathogen stimulation, they are actively secreted or passively released into the extracellular environment. There, they bind to receptors to activate downstream inflammatory signaling pathways, recruiting and activating immune effector cells such as dendritic cells, eosinophils, and type 2 innate lymphoid cells, thereby amplifying the local inflammatory response. In allergic asthma, epithelial alarmins serve as crucial mediators, providing distinct tissue-specific signals to innate and adaptive cell populations, making them key therapeutic targets for intervention.
TSLP
TSLP is the earliest clearly identified epithelial alarm protein, holding significant importance in the field of immunology. Its discovery not only revealed a novel mechanism for epithelial-immune cell communication but also established its pivotal role within the core regulatory network of type 2 immune responses. TSLP is an IL-7-like lymphocyte growth factor primarily produced in response to pathogenic stimuli in lung and intestinal epithelial cells and skin keratinocytes. It promotes the adaptive immune system by recruiting and activating antigen-presenting cells such as dendritic cells (DCs) and innate lymphoid cells (ILCs), thereby inducing complex type 2 immune responses at barrier surfaces.13,14 The functional receptor for TSLP is a heterodimeric complex composed of TSLPR (thymic stromal lymphopoietin receptor) and IL-7Rα.15 TSLP binds specifically to its receptor, thereby activating downstream signaling pathways. This process constitutes the key molecular basis for its role as a core regulator in type 2 immune responses. Consequently, TSLP is also recognized as a primary modulator of type 2 immune responses.
In allergic asthma, DCs serve as the primary target cell type for TSLP. Under TSLP induction, DCs express OX40 ligand (OX40L), triggering naive CD4+ T cells to produce IL-4, IL-5, and IL-13. This process induces Th2 cell polarization, thereby establishing the Th2-polarized inflammatory environment characteristic of the airway microenvironment in allergic asthma.16 The latter primarily manifests as elevated levels of cytokines (IL-4, IL-5, IL-13), chemokines (CCL5, CCL11, CXCL2, CXCL12), and growth factors (transforming growth factor-beta (TGF-β); basic fibroblast growth factor (bFGF); vascular endothelial growth factor (VEGF); platelet-derived growth factor-beta (PDGF-β)).17 These cytokines, chemokines, and growth factors are stored by eosinophils recruited to the site of inflammation and are rapidly released at the inflammatory site to coordinate the Type 2 immune response in an exacerbating or modulating manner. Furthermore, these soluble mediators profoundly affect airway structural cells, directly leading to excessive mucus production, fibrosis, and airway smooth muscle changes, which in turn directly contribute to the manifestation of asthma symptoms.18,19
Alveolar macrophages (AM) are also key cellular targets for TSLP-mediated Type 2 immune responses. It is well established that macrophages constitute the most abundant immune cells in the lung. Upon exposure to prototypical cytokines or TLR agonists, they differentiate into two functionally distinct subsets: classically activated (M1) macrophages and alternatively activated (M2) macrophages, forming a crucial component of the innate immune system. Among these, M2 macrophages are closely associated with asthma pathogenesis. Induced by Th2 cytokines such as IL-4 and IL-13, they express high levels of C-type lectin receptors, chemokines, cytokines, macrophage signaling proteins, and transcription factors that promote airway inflammation, fibrosis, and airway hyperresponsiveness (AHR).20 Existing research indicates that TSLP can amplify M2 macrophage differentiation. Through TSLP/TSLPR signaling, it participates in M2 cell polarization in mice, inducing M2 macrophages to produce the Th2-inducing cytokine OX40L and the Th2-recruiting chemokines thymus and activation-regulated chemokine (TARC) and macrophage-derived chemokine (MDC), thereby exacerbating allergic inflammatory symptoms.21 Moreover, TSLP can upregulate AMFR (also known as gp78, an endoplasmic reticulum-resident E3 ubiquitin ligase) expression in AMs. By fine-tuning TSLP-STAT5 signaling within AMs, it promotes Th2 responses, thereby driving the progression of allergic airway inflammation.22
In summary, TSLP serves not only as a key upstream alarm molecule initiating allergic asthma, but its cascade amplification effect within signaling pathways constitutes the core scientific rationale for therapeutic intervention. TSLP exerts its effects through three primary pathways. First, it directly activates DC-mediated Th2 cell polarization. Second, it drives AM differentiation toward the M2 phenotype. Third, it promotes Th2 immune responses via the TSLP-STAT5 signaling axis. This multi-target regulatory property explains why nanoparticle targeting TSLP demonstrates more pronounced therapeutic advantages compared to other single-target strategies.
IL-25
IL-25 is an upstream protein in immune responses and a key member of the epithelial alerting protein family. Also known as IL-17E, IL-25 was initially defined as a cytokine produced by Th2 cells and is a member of the IL-17 cytokine family.23 Its receptor is a heterodimeric complex composed of two subunits, IL-17RA and IL-17RB, which is widely expressed across multiple cell types. Its biological effects are distinctly different from those of other members of the IL-17 family.24 In vivo studies indicate that IL-25 infusion in mice primarily induces a Th2-type immune response across multiple tissues, characterized by enhanced expression of Th2 cytokines, elevated serum immunoglobulin (Ig)E, IgG1, and IgA concentrations, and increased blood eosinophil counts23 Moreover, studies on the sources of IL-25 indicate that, in addition to Th2 cells, alveolar macrophages (AM), innate lymphoid cells 2 (ILC2), eosinophils, and basophils are all potential sources of IL-25.25 These findings reveal the intricate mechanisms by which IL-25 mediates Type 2 immune responses. Therefore, elucidating the role of IL-25 in allergic asthma will aid in identifying therapeutic intervention targets (Table 1).
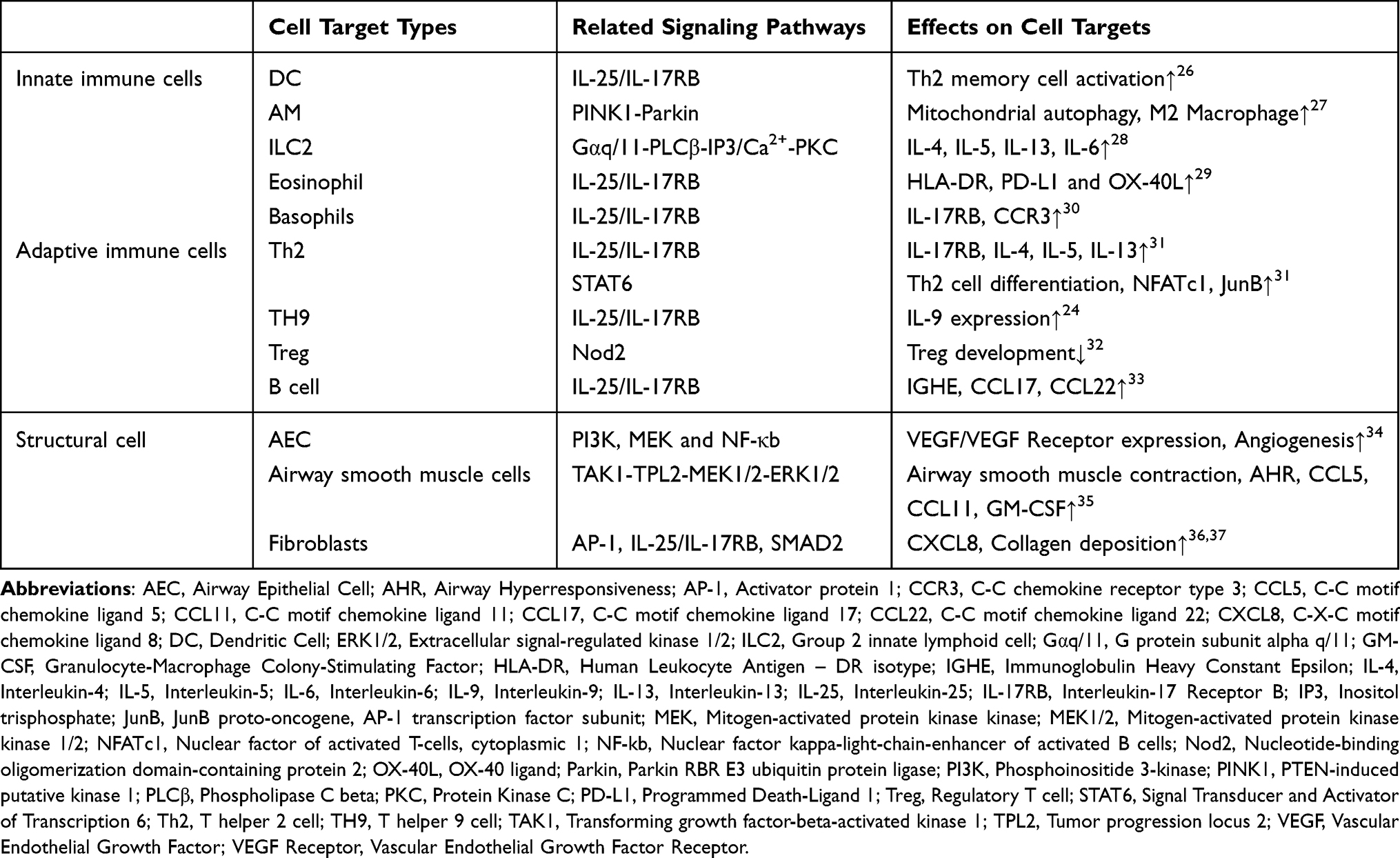 |
Table 1 Mechanisms of IL-25 in Allergic Asthma |
In allergic asthma, the mechanism by which IL-25 triggers Type 2 immune responses largely overlaps with that of TSLP. Specifically, it initiates the recruitment of innate and adaptive immune cells such as eosinophils by inducing excessive production of Th2 cytokines, thereby forming an inflammatory microenvironment that leads to asthma symptoms.31
However, among the three typical epithelial alarmin proteins, IL-25 particularly enhances the type 2 immune response and further exacerbates allergic diseases. This effect is attributed to the dependence of the type 2 response on the IL-25 receptor. Petersen et al identified a novel neutrophil population expressing IL-25 receptors, termed type 2 myeloid (T2M) cells. These cells serve as the primary source of type 2 cytokines following pulmonary IL-25 administration.38 Their chronic allergic asthma mouse model demonstrated that when the IL-25-specific receptor is absent, type 2 cytokines associated with T2M cells are significantly reduced, proving that the IL-25 receptor plays a crucial role in mediating innate and adaptive type 2 immune responses in the lungs. Moreover, another experiment surprisingly revealed that mouse T2M cells exhibited resistance to high-dose steroid treatment, and a similar IL-4/IL-13-producing granulocyte population was identified in the peripheral blood of asthma patients. These data establish IL-25 and its receptor as novel targets for both innate and adaptive immune responses in allergic asthma.
Moreover, IL-25 is a major driver of airway remodeling in asthma. As previously mentioned, inhaled airborne allergens damage AECs and trigger the release of epithelial alarm proteins, thereby driving downstream inflammatory processes. The anti-inflammatory repair processes resulting from these abnormal inflammatory responses lead to structural changes in both large and small airways. These changes include epithelial dysfunction, goblet cell hyperplasia and metaplasia, thickening and fibrosis of the subepithelial matrix, increased airway smooth muscle mass, and enhanced angiogenesis—collectively termed airway remodeling.39 As one of the core characteristics of asthma, airway remodeling not only leads to clinically observable airflow limitation and worsening respiratory symptoms in patients but also triggers multiple adverse consequences such as reduced lung function and diminished responsiveness to medications.
Research indicates that a high IL-25 pattern appears to be associated with poor asthma control.40 Therefore, the driving mechanism of IL-25 is particularly crucial during airway remodeling. Within AEC itself, IL-25 release induces pro-allergic chemokine production, increased goblet cell and mucus secretion, overall epithelial cell proliferation, and AHR.31 Research data from Yao et al indicate that IL-25 promotes airway fibrotic remodeling by inducing an epithelial-mesenchymal transition (EMT)-like pro-fibrotic phenotypic shift in bronchial epithelial cells through an autocrine-dependent mechanism.41 Furthermore, mounting evidence indicates that IL-25 plays a pivotal role in epithelial-mesenchymal crosstalk and local tissue remodeling. Research by Gregory et al demonstrates that IL-25 directly acts on human mesenchymal cells to increase collagen deposition around airways. It enhances bronchial neovascularization by activating the PI3K/Akt and Erk/MAPK pathways, thereby increasing endothelial cell expression of VEGF.37 Weathington et al demonstrated that IL-25 can maintain a receptor-activated phenotype in human monocytes and macrophages through synergistic stimulation with IL-4. Furthermore, IL-25 induces the expression of specific receptors via feedforward autocrine signaling, thereby establishing a positive feedback loop system capable of perpetuating inflammation.42
A deep analysis of IL-25’s functional network reveals that its fundamental distinction from other alarmins lies in its receptor dependency and feedback signaling mechanisms. The IL-25/IL-17A-IL-17RB signaling axis is not only essential for initiating Type 2 immune responses but also self-amplifies inflammation through positive feedback loops. This characteristic necessitates that nanoparticle designs targeting IL-25 prioritize signal interruption strategies over simple ligand blockade. These findings are unique among epithelial alarmin molecules, establishing IL-25 as a potential therapeutic target for cytokine-based therapies.
IL-33
IL-33 is another key alarmin within the epithelial alarmin family. As a dual-function protein derived from the endothelium, IL-33 exhibits multiple characteristics as both a transcriptional repressor and a cytokine.43 Its gene is located on chromosome 9p24.1, comprising one non-coding exon (exon 1) and seven coding exons (exons 2–8).44 IL-33 exerts its biological functions through its specific receptor ST2 (IL-33R). The ST2 receptor is widely expressed on the surface of immune cells and serves as a key molecular switch for IL-33-mediated Type 2 immune responses. Current research indicates that among the 61 asthma susceptibility loci identified in recent genome-wide association studies, variants near the IL-33 cytokine and its receptor ST2 gene carry potential significance.45,46 Experimental data indicate that asthma-associated variants at the IL-33 locus can mediate allele-specific regulation of activity and IL-33 expression, thereby elevating IL-33 protein levels. The latter is constitutively and abundantly expressed in numerous human tissues. As a member of the IL-1 cytokine family, it plays a crucial role in many inflammatory processes and diseases.
Unlike the TSLP-DC-OX40L-Th2 axis described above, in allergic asthma, IL-33 operates through a distinct mechanism, directly influencing both innate and adaptive immune cells without requiring DC-mediated Th2 polarization. This divergence in signaling pathways—with IL-33 primarily activating ILC2s while TSLP engages DCs—illustrates the complementary therapeutic targets presented by different alarmins and justifies the development of alarmin-specific nanoparticle strategies. Currently, IL-33 primarily promotes the progression of Type 2 immune responses through two pathways: by interacting with group II ILC2, basophils, and natural killer (NK) cells to induce Th2 cytokine secretion; and by directly inducing Th2 cytokine production in CD4+ T cells.
In the innate immune pathway, pulmonary ILC2s, which are abundant in mucosal tissues and barrier surfaces, serve as the primary targets for IL-33.47,48 Research indicates that the IL-33/ST2 axis is a key factor driving the extrusion and transport of bone marrow-derived ILC2 lineage cells to the lungs. It effectively induces rapid expansion of ILC2s in the lungs, thereby enhancing airway constriction capacity.49,50 Its significance in allergic airway inflammation has been demonstrated in numerous experiments.51–55
In the adaptive immune pathway, IL-33 induces distinct types of effector CD4+ T cells, influencing various aspects of the Type 2 immune response through Type 2 cytokine production, airway eosinophilia, mucus hyperplasia, and IgE antibody production.56
Moreover, IL-33 participates in regulating adaptive immune responses and maintaining homeostasis. Research by Chen et al indicates that IL-33 directly activates regulatory T cells (Tregs) by enhancing the expression of their surface receptor ST2. This process causes Tregs to lose their ability to suppress effector T cells and instead produce Th2 cytokines, thereby compromising the immune tolerance established in the lungs following prior antigen exposure. Consequently, IL-33 promotes the progression of airway inflammation.57 IL-33 exhibits the broadest spectrum of actions among the three alarmins, capable of both activating ILC2 and regulating adaptive immunity (Th2, Treg). This dual regulatory capacity makes it the most promising therapeutic target. Notably, IL-33’s bidirectional regulation of Treg cells—activating ST2+ Tregs while suppressing conventional Tregs—reveals the complexity of immune regulatory networks. These findings indicate that IL-33 is a central mediator in triggering Type 2 immune responses.
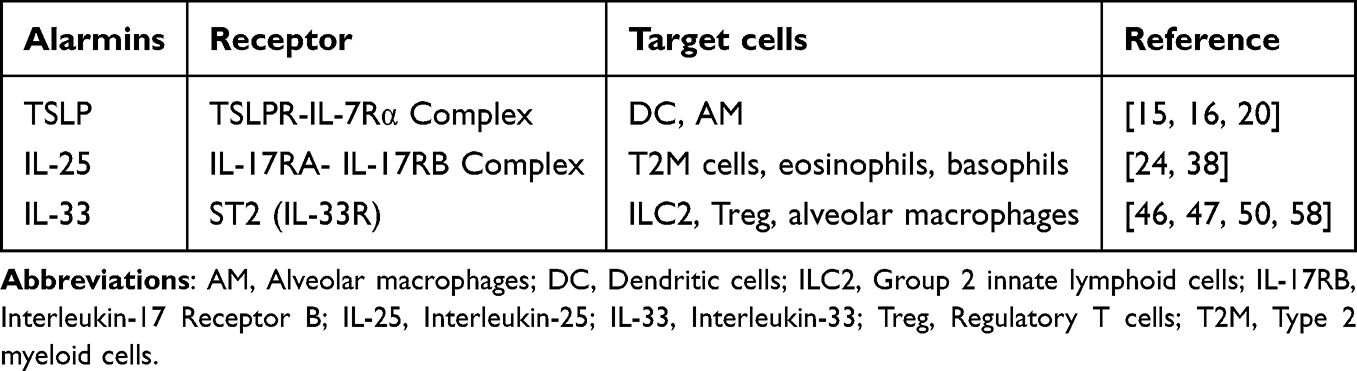 |
Table 2 Alarmins and Their Target Cells |
Synergistic Effects of TSLP, IL-25, and IL-33
As described above, TSLP, IL-25, and IL-33 are potent inducers of airway inflammation, receptors and target cells were detailed in Table 2. Although their cellular functions overlap, in allergic asthma, these three molecules drive Type 2 immune responses through distinct tissue-specific signaling pathways, thereby synergistically promoting the development of airway inflammation. Studies indicate that in a mouse model of innate allergic airway inflammation induced by Alternaria extract (Alt-Ext), TSLPR and ST2 signaling pathways mutually enhance the expression and release of their respective ligands in the lungs.58 In TSLP-DC-activated human Th2 memory cells, IL-25 maintains Th2 transcription factor expression independently of IL-4, enhancing Th2 memory cell polarization and cytokine production.26 In an ovalbumin (OVA)-induced mouse model, TSLP, IL-25, and IL-33 act synergistically to promote cell migration and ILC2 activation.59 Although studies by Vannella et al have found that the combined use of antibodies targeting TSLP, IL-25, and IL-33 does not further reduce established pulmonary inflammation or fibrosis in certain circumstances, indicating partial redundancy among these three molecules in maintaining type 2 pathology.60 During the early inflammatory phase preceding chronicity, combined targeting of these mediators is essential for alleviating subsequent allergic airway manifestations. The central role of epithelial alarmins in initiating type 2 immune responses and the corresponding nanoparticle-based intervention strategies are schematically summarized in Figure 1, which serves as a visual roadmap for the following sections.
 |
Figure 1 Schematic overview of the epithelial alarmin axis and nanoparticle-based intervention strategies in allergic asthma. |
Application of Biomedical Nanoparticles in Targeting Epithelial Alarmin-Producing Cells
NPs, ie., functional NPs applied in the biomedical field, represent one of the most promising frontier research hotspots in recent years. Their unique physicochemical properties enable precise control over the biodistribution of drug payloads and the rate of drug release, facilitating site-specific delivery, deposition, and cellular uptake. This achieves targeted drug delivery, thereby enhancing drug efficacy and therapeutic outcomes.61,62 Therefore, biomedical nanoparticles are frequently developed as drug delivery systems to overcome the limitations of traditional delivery methods. Currently, with the continuous advancement of nanomedicine technology, the structural design of nanoparticle drug delivery systems has become increasingly complex. They exhibit high diversity in chemical composition, size, morphology, electrostatic charge, hydrophobicity, surface chemistry, and other properties, aiming to leverage biomaterials and biomedical engineering innovations to overcome biological barriers and patient heterogeneity.63 Among these, NPs with immunomodulatory properties represent emerging tools for developing anti-infective therapies, inducing immune tolerance, and regulating inflammation, offering significant therapeutic potential. Since the introduction of the epithelial barrier hypothesis, the pathophysiological mechanisms of allergic asthma have been further refined. Immunomodulatory NPs targeting epithelial alarmin cells have consequently emerged as a promising new generation of asthma treatment strategies.
Innate Immune Cells
Dendritic Cells
DCs are key players in Type 2 immune responses. As professional antigen-presenting cells (APCs) bridging innate and adaptive immunity, DCs bear significant responsibility in initiating and regulating the dynamic process of Type 2 immune responses. Extensive research analyses indicate that most airborne allergens can directly or indirectly activate DC sensitization via epithelial alarmins, amplifying Th2 cell-mediated airway inflammation during the effector phase of allergic asthma.64–66 These findings have positioned tolerance-inducing nanoparticles (tNPs) that induce DC differentiation into tolerogenic DCs as promising candidates for allergic asthma treatment. It is well established that DCs differentiate into either immunogenic DCs or tolerogenic DCs based on the regulation of different microenvironmental signals. The former phagocytose exogenous antigens and present them to naive CD4+ T cells to initiate a Th2 immune response, while the latter maintain immune tolerance, preventing excessive immune reactions to self-antigens or harmless antigens. Based on this, tNPs that can precisely target DCs and induce tolerogenic DC differentiation, while exhibiting good stability, biocompatibility, and immunomodulatory properties, have become a hotspot for development in asthma treatment strategies.
tNPs, which carry allergens or self-antigens, can target APCs and deliver tolerance-coordinating signals to promote antigen-specific immune responses, demonstrating significant potential in inducing tolerance-inducing immunity. Over the past decades, researchers have explored three distinct delivery strategies for tNPs. Among these, tNPs simultaneously carrying both allergens and immunomodulators have been demonstrated to induce antigen-specific tolerance by delivering allergens to APCs, thereby inducing the differentiation of tolerogenic APCs.67 This form of treatment, also known as allergen-specific immunotherapy (AIT), has yielded encouraging results in treating inflammatory conditions such as allergies and autoimmune diseases, representing a potential curative approach for asthma.68
In recent years, Zhao et al have developed biodegradable tNPs loaded with the allergen OVA and the immunomodulator rapamycin.69 These tNPs can be loaded onto microneedle tips via centrifugation to form sustained-release microneedles for transdermal delivery of tNPs. The PLGA@OVA-Rapa formulation exhibits sustained release within the skin for over 96 hours, inducing the generation of allergen-specific Treg cells. This reduces pulmonary inflammation, mucus, and collagen accumulation, ultimately providing effective relief from allergic symptoms. Research indicates that these nanoscale needles can overcome the skin barrier to deliver drugs non-invasively and painlessly into the epidermis and dermis, significantly enhancing the transdermal delivery efficiency of biopharmaceuticals.70 Following successful co-delivery of OVA and rapamycin to the dorsal skin of mice via microneedles, expression of DC co-stimulatory molecules was significantly downregulated, effectively inducing antigen-specific immune tolerance and alleviating asthma symptoms.
Additionally, sublingual delivery represents a promising therapeutic strategy. Sadeghi et al engineered Exo-Ova composite tNPs decorated with DC-specific aptamers, combining the allergen OVA with mesenchymal stem cell-derived exosomes (MSC-exos) for targeted delivery to DCs. Their therapeutic efficacy was evaluated via sublingual immunotherapy in a mouse model of allergic asthma.71 Experimental results demonstrate that this complex precisely delivers allergens to dendritic cells within the sublingual mucosa, significantly reducing the allergen dose required for sublingual immunotherapy and effectively minimizing adverse reactions associated with AIT. Moreover, these tNPs exhibit outstanding immunomodulatory capabilities. MSC-exos, a subtype of naturally occurring extracellular vesicular nanoparticles, possess inherent stability, biocompatibility, minimal immunogenicity, and leverage the potent immunomodulatory and regenerative properties of MSCs. They play a crucial role in diverse therapeutic fields including drug delivery and immune regulation.72 Extensive research has demonstrated its immunomodulatory capacity in inflammatory airway diseases.73–76 Undoubtedly, MSC-exos are emerging as a highly promising therapeutic vehicle for allergic asthma. Administration results in mouse models further corroborate this finding. Data indicate that tNPs can enhance the therapeutic efficacy of sublingual immunotherapy, significantly inducing Treg cell immune responses to alleviate airway and pulmonary inflammation.77 This offers a novel strategy for applying MSC-exos in the immunotherapy of allergic asthma.
DCs, serving as a bridge between innate and adaptive immunity, act as both early respo410nders to alarmins and key initiators of Th2 polarization in TSLP-driven asthma inflammation. Current DC-targeted NP strategies have evolved into three major technical pathways: transdermal microneedle sustained-release systems, sublingual aptamer-targeted delivery, and exosome-based drug-loading platforms. The core rationale behind these designs is to precisely modulate the expression of DC co-stimulatory molecules, redirecting immune responses from pathogenic Th2 to tolerogenic Treg pathways, thereby achieving a paradigm shift from “anti-inflammatory” to “tolerogenic” effects.
Alveolar Macrophages
AM are the most abundant immune cells in the lungs. As mentioned earlier, AM exposed to prototypical cytokines or TLR agonists can differentiate into two functionally distinct subsets: pro-inflammatory M1 macrophages and anti-inflammatory M2 macrophages. The former exhibit potent antibacterial and anticancer activity, aiding in pathogen clearance during infections. The latter participate in Th2 responses, parasite clearance, and inflammation suppression, promoting tissue repair, wound healing, angiogenesis, and fibrosis. Both are alternately activated during inflammatory processes, maintaining homeostasis through a balanced ratio. Epithelial alarmin proteins like TSLP activate AMs through signaling pathways and induce their polarization toward M2 macrophages, thereby activating Th2 cells and driving the development of allergic asthma. Consequently, targeting signaling pathways that promote M2 macrophage polarization to remodel AM polarization and restore pulmonary homeostasis may represent a potential strategy for immunomodulatory NP-based therapies against allergic asthma. Increasing research supports the feasibility of this approach. Recently, Pei et al developed a biomimetic NP (EM-PLGA@Dnmt3aos smart silencer) that leverages the “homing” properties of AMs to precisely deliver small nucleic acid drugs to lung and airway inflammatory tissues by coating M2 macrophage exosome membranes to modify siRNA-loaded poly(lactic-co-glycolic acid) (PLGA) NPs, enhancing PLGA targeting to pulmonary M2 macrophages. This enables efficient delivery of therapeutic Dnmt3aossmart silencer to suppress M2 polarization, effectively treating allergic airway inflammation in mice.78 Wu et al developed cationic liposomes loaded with Mbd2 siRNA that specifically target AMs via macrophage phagocytosis. By reducing Mbd2 expression in AMs, this approach eliminates M2 macrophage polarization, significantly reversing OVA-induced allergic asthma symptoms in mice, including airway inflammation and excessive mucus secretion.79 Xin et al purified extracellular vesicles containing Emu-miR-10a-5p from the scolex of Echinococcus multilocularis, which can interfere with leukemic factors and JAK1-STAT3 signaling in the lungs of allergic asthma mice. Through a cross-species approach, it targets and binds to leukemic pathogenic factors to inhibit M2 macrophage polarization, thereby alleviating OVA-induced allergic asthma.80 These findings demonstrate that inhibiting M2 macrophage polarization through diverse mechanisms suggests that developing NP delivery strategies targeting M2 macrophages represents a promising therapeutic approach for asthma. Notably, distinct NPs exhibit markedly different mechanisms for inhibiting M2 polarization, encompassing focused epigenetic regulation, gene targeting, and cross-species regulatory pathways. Intervention through these diverse pathways consistently demonstrates superior efficacy compared to conventional asthma medications while providing a foundation for mechanistic research.
Adaptive Immune Cells
Th2 Cells
Th2 cells are central participants in the pathogenesis of allergic asthma and the ultimate targets for epithelial alarmins in inducing inflammation. They mediate IgE production and inflammatory cell recruitment by secreting multiple cytokines, triggering immediate allergic reactions and exacerbating airway inflammation.81 It is well established that IgE serves as the primary initiator of immediate allergic reactions. Upon allergen exposure, IgE cross-links with its specific allergen, rapidly activating bound mast cells and basophils to release inflammatory mediators such as histamine and leukotrienes. This triggers immediate-phase asthma responses including bronchospasm and mucosal edema. Concurrently, inflammatory cells such as eosinophils—targeted by Th2 cytokines—accumulate in the airways. These cells release inflammatory mediators like granule proteins, damaging AECs and disrupting airway structure. This triggers late-phase asthma responses including AHR and airway remodeling, transforming asthma from acute episodes into a chronic, difficult-to-treat condition requiring long-term management. Therefore, drug delivery and immunomodulation targeting Th2 cell differentiation and function have become primary strategies for asthma treatment in nanomedicine. Based on different mechanisms of action, NPs can be categorized into the following two types.
First are NPs that suppress Th2 cell differentiation. In allergic asthma, an imbalance in the Th1/Th2 cell ratio has long been recognized as the root cause of airway inflammation. Therefore, shifting the immune response from the disease-promoting Th2 type to the non-pathogenic Th1 type—thereby bringing the immune system into a more balanced state—may help alleviate asthma symptoms. In recent years, novel NP delivery systems designed to modulate the Th1/Th2 response balance have proliferated.
Cell membrane-coated nanoparticles (CNPs) are one such example. As the name suggests, these are bio-inspired nanoparticles synthesized by modifying cell membranes from natural sources. They possess dual characteristics of both the original nanoparticles and the biological interface of cell membranes, demonstrating therapeutic potential in multiple fields including drug delivery, immunotherapy, and tumor targeting.82 In allergic asthma, Jin et al designed a bioinspired nanoparticle delivery system, PM@Ber-NPs, coated with platelet membranes. They loaded the natural herbal compound berberine into the nanomaterial PLGA and administered it intranasally to target delivery to the airways and inflammatory regions of the lungs, thereby suppressing the inflammatory response.83 Berberine is a benzoisoquinoline alkaloid extracted from the Chinese medicinal plant Berberis, which effectively suppresses T-selectin-like protein (TSLP) expression. It downregulates Th2 cells and their cytokine levels, thereby inhibiting asthma progression.84 However, its low water solubility and bioavailability significantly limit its therapeutic efficacy. To harness its anti-inflammatory effects, researchers developed a nanoscale membrane vesicle platform derived from platelets. This platform shields the encapsulated mimetic NPs from phagocytic uptake by macrophages, enabling precise targeting and retention at inflammatory sites.85 Studies demonstrate that these bio-inspired NPs exhibit enhanced cellular uptake and targeted retention within inflammatory microenvironments. This cell membrane-coated biomimetic drug delivery system enhances the stability and active targeting capabilities of NPs by coating extracellular vesicles onto existing biocompatible NPs like PLGA. This approach significantly improves drug efficacy in airway inflammatory diseases such as allergic asthma, offering a promising platform for advancing asthma treatment.
Beyond this, novel nanocarrier systems for delivering bioactive proteins and peptides represent an exciting avenue of research. Song et al proposed a novel cathepsin B-activated nanocarrier MPP-Trp for asthma immunotherapy. This involved obtaining the cathepsin B-reactive peptide fragment Fmoc-IALLIPF-GFLG-W via solid-phase synthesis and combining it with the anti-inflammatory agent tryptophan (Trp), and hydrophilic-modified copolymer polyethylene glycol (PEG). This PEG, which exhibits excellent biocompatibility, water dispersibility, and stability, demonstrates remarkable sensitivity and selectivity toward cathepsin B, a key enzyme in asthma-related inflammation.86 Research indicates that MPP-Trp exerts immunomodulatory effects on Th1/Th2 cell populations, significantly alleviating OVA-induced allergic asthma by shifting the immune response toward Th1 dominance. This approach may offer a safer, more sustainable solution for asthma management.
Rochman et al encapsulated nucleoside-modified allergen-encoding mRNA vaccines for allergy prevention and treatment within lipid nanoparticles (LNPs) exhibiting adjuvant properties, forming allergen-specific mRNA-LNPs. This enabled mRNA to shape CD4⁺ and CD8⁺ T cell responses and induce allergen-specific IgG1 and IgG2 antibodies at lower doses, block Th2 cell activation, and generate anti-allergic environments to prevent allergic manifestations such as experimental asthma. This demonstrates a promising approach for preventing and treating allergic diseases.87
Next are NPs that suppress Th2 cytokine expression. In allergic asthma, the transcription factor GATA-binding protein 3 (GATA-3) is a key regulator of Th2-specific cytokine transcription. It directly binds to the promoter and enhancer regions of the IL-4, IL-5, and IL-13 genes to initiate their transcription, thereby inducing cytokine expression.88 It is evident that the expression of these Th2 cytokines is the direct cause of allergic airway inflammation. Consequently, NP delivery systems targeting GATA-3 expression to suppress Th2 cytokine production have emerged. In recent years, Gavitt et al attempted to cross-link matrix metalloproteinase-9 (MMP-9)-specific peptide substrates with DNAzyme-functionalized nucleic acid nanocapsules (DNAzyme-NAN) to achieve GATA-3-specific gene regulation in allergic asthma.89 Previously, their group had demonstrated that DNAzyme-NAN is a promising therapeutic nucleic acid delivery vehicle capable of achieving up to 60% GATA-3 mRNA knockdown in MCF-7 cells without the use of toxic transfection agents.90 Based on this, they hypothesized that increased MMP-9 during airway inflammation could enhance the degradation specificity of the peptide-crosslinked GATA-3 DNAzyme-NAN, enabling its rapid breakdown in the lungs to specifically knock down GATA-3. Experimental results demonstrate that peptide-conjugated GATA3 DNAzyme-NANs effectively deliver GATA-3-specific DNA enzymes to immune cells and regulate eosinophil levels in asthmatic mice, potentially reducing the severity of asthma symptoms in human patients. Furthermore, Jürgens et al engineered transferrin-conjugated lipid nanoparticles (Tf-LNPs) for Th2 cell targeting in allergic asthma. Leveraging the abundant expression of transferrin receptors (TfRs) on Th2 cell surfaces, they achieved specific delivery of therapeutic siRNA to Th2 cells, enhancing Tf-dependent cellular uptake and therapeutic efficacy of GATA-3.91 Tf-LNP enhances the efficiency and recovery rate of GATA-3 knockdown in the NP delivery system. In summary, by precisely regulating Th2 cell differentiation and function, nanomedicine has opened new avenues for treating allergic asthma. These innovative therapeutic approaches not only improve drug targeting and efficacy but also lay the foundation for developing safer, more effective asthma management strategies.
Regulatory T Cells
Treg cells are a subset of T cells with immunosuppressive functions that play a crucial role in maintaining immune homeostasis and tolerance to harmless antigens. In allergic asthma, Treg cells precisely downregulate Th2 cells through three mechanisms: directly inhibiting Th2 cell activation, secreting anti-inflammatory factors to neutralize Th2 effects, and depleting essential cytokines for Th2 cells, thereby blocking the Th2-mediated Type 2 immune response.92–95 However, extensive research indicates that both the number and function of Treg cells are significantly suppressed by epithelial alarmins.32,57,96 To this end, researchers customized nanoparticles capable of reducing epithelial alarmin expression while promoting Treg cell differentiation and survival, thereby restoring immune homeostasis and tolerance within the body. In recent years, Liu et al designed tNPs surface-modified with apolipoprotein B peptide sequences or mannan oligosaccharides. By targeting scavenger or mannose receptors, these nanoparticles selectively delivered the allergen OVA to naturally tolerant APCs—hepatic sinusoidal endothelial cells—thereby inducing Treg cell production.97 In vitro and in vivo experiments demonstrated that these tNPs effectively enhanced the uptake of OVA by hepatic sinusoidal endothelial cells and the production of anti-inflammatory cytokines IL-10 and transforming growth factor TGF-β. Through tissue infiltration of Treg cells, they significantly suppressed OVA-induced allergic airway inflammation.97 Furthermore, Li et al encapsulated IL-10 within AM-membrane-coated PLGA particles. Leveraging the biological properties of the AM membrane, this approach enabled efficient delivery of the therapeutic cytokine to the lungs, substantially increasing IL-10 bioavailability. This intervention improved the Th2/Treg response balance and alleviated airway inflammation.98 These findings suggest that using nanoparticles to induce Treg cell generation represents a promising strategy for asthma management. Liver-targeted tNPs and AM-membrane-coated NPs may form complementary strategies. The former induces Tregs by enhancing antigen presentation capacity in hepatic sinusoidal endothelial cells, while the latter leverages the innate homing properties of macrophages to improve pulmonary targeting efficiency, potentially enabling synergistic regulation of the “liver-lung axis.”
In summary, the application of biomedical nanoparticles in epithelial alarmin cell targeting demonstrates a multi-level precision treatment strategy: at the innate immune level, DC-targeted tNPs reshape the Th1/Th2 balance, while AM-membrane-modified bionic NPs target macrophage polarization pathways; At the adaptive immune level, they employ dual pathways of inhibiting Th2 cell differentiation and blocking cytokine expression, while simultaneously restoring immune tolerance through Treg cell induction and functional recovery. These NPs not only address the issues of low bioavailability and poor targeting inherent in traditional drugs but also achieve multi-targeted intervention from the cellular to the molecular level through their unique physicochemical properties, offering novel solutions to overcome drug resistance and side effects in asthma treatment. Additionally, beyond the aforementioned NPs specifically targeting the epithelial alarmin signaling pathway, various NPs have emerged in recent years that suppress allergic airway inflammation through different mechanisms (Table 3). These nanocarriers validate the broad application potential of NPs in asthma treatment from multiple perspectives.
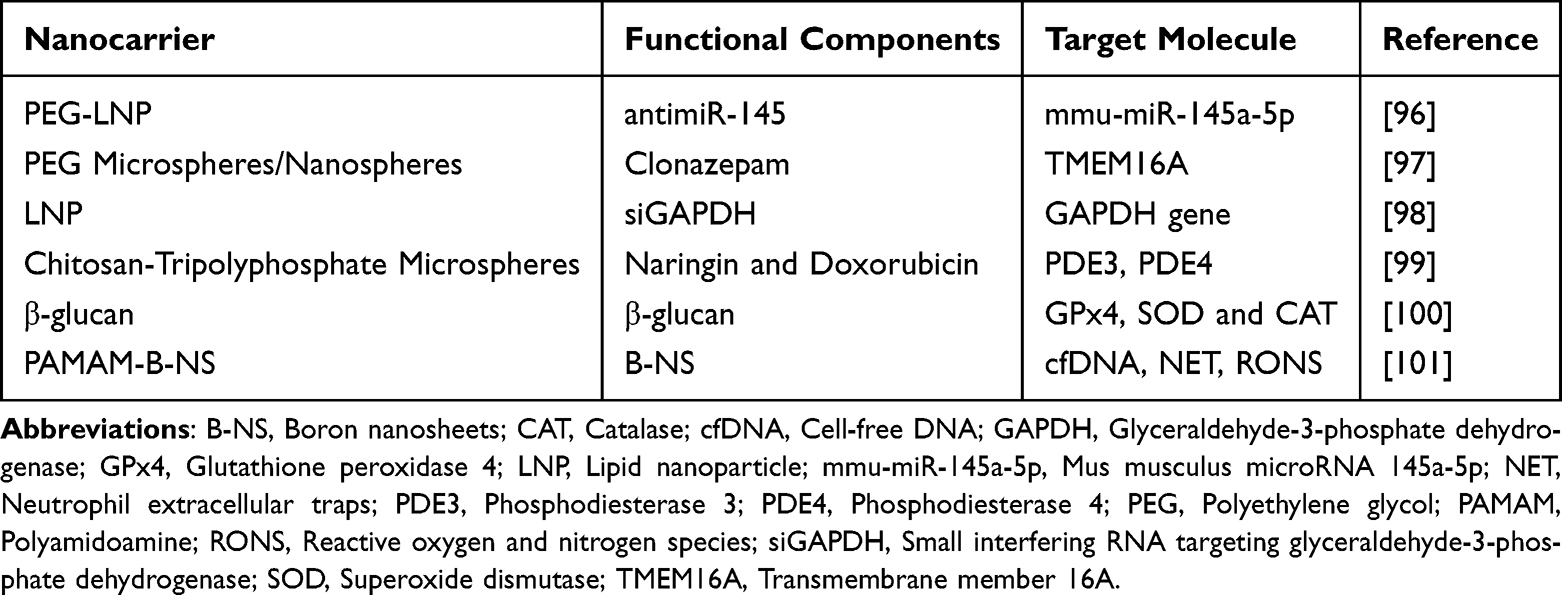 |
Table 3 Details of NPs Suppressing Allergic Airway Inflammation |
Application of Biomedical Nanoparticles in Conventional Asthma Therapies
Beyond immunomodulation, delivering conventional asthma medications such as steroids using biomedical nanoparticles is also a common approach to asthma management in the field of nanomedicine.99 Budesonide (BUD) is a commonly used ICS medication for asthma that effectively suppresses type 2 inflammatory responses and repairs airway epithelium, reducing the release of epithelial alarmin. The SMART (Single Maintenance and Reliever Therapy) regimen combining budesonide with the long-acting bronchodilator formoterol is currently prioritized in asthma management guidelines by the National Asthma Education and Prevention Program Coordinating Committee and the GINA for sustained maintenance and rapid relief of asthma symptoms.100,101 However, inhalers have drawbacks such as medical-legal issues, high drug costs, and poor patient compliance.102,103 Therefore, there is an urgent need for novel therapeutic interventions to enhance the bioavailability of budesonide in the lungs, thereby improving its therapeutic efficacy to control symptoms and exacerbations in asthma patients. To address this, Zuo et al developed BUD-LNPs by incorporating budesonide into LNP systems via thin-film hydration. This approach leverages the LNP’s ability to encapsulate hydrophobic substances, thereby improving budesonide’s bioavailability, preventing premature degradation, and ensuring controlled-release delivery. The study demonstrated that BUD-LNPs exhibit excellent biocompatibility both in vitro and in vivo, along with superior therapeutic efficacy compared to free BUD. This approach holds great promise for the development of novel drugs for future asthma treatment.104 Moreover, the hydrophilic/hydrophobic surface properties of LNP enhance drug delivery by prolonging retention time and boosting systemic absorption. In an OVA-induced allergic asthma mouse model, Liu et al designed a highly hydrophilic LN loaded with BUD.105 Mechanistic studies indicate that this LNP prolongs the pulmonary residence time of BUD by reducing alveolar epithelial transport and avoiding clearance by alveolar macrophages, thereby decreasing dosing frequency and significantly alleviating asthma symptoms.
Additionally, numerous natural products such as curcumin and quercetin exhibit anti-inflammatory effects. However, they typically suffer from poor water solubility, rapid metabolism, and low bioavailability, hindering their optimal therapeutic efficacy. Research indicates that curcumin nanoformulations can significantly enhance its bioavailability and drug stability.106
In the field of traditional asthma treatment, biomedical nanoparticles have significantly enhanced the therapeutic efficacy of existing drugs through innovative delivery strategies. Addressing issues such as low bioavailability, poor patient compliance, unstable formulations, unclear targeting, and inappropriate delivery timing, researchers have developed nanoparticle drug delivery systems. These systems improve the bioavailability of hydrophobic drugs while enabling controlled-release administration to prevent premature degradation. These studies confirm that by rationally designing nanoparticle surface properties and drug-loading mechanisms, the delivery efficiency and clinical efficacy of traditional asthma medications can be improved, providing crucial technical support for developing next-generation asthma treatment solutions.
Beyond these preclinical and formulation-based advances, several nanoparticle-based therapies have entered clinical trials for asthma and allergic diseases, marking a critical step toward clinical translation. These include AZD-1419 (TLR9 agonist-conjugated nanoparticles; NCT02898662) for moderate-to-severe asthma, which completed Phase II evaluation with time to first loss of asthma control as the primary endpoint; Inflamax TSLP-siRNA (siRNA-loaded lipid nanoparticles; NCT05243550), a Phase I/IIa study assessing safety and TSLP knockdown in mild-to-moderate asthma; and AllerT (liposomal allergen peptides; NCT04072796), a Phase IIb trial in house dust mite-allergic patients that met its primary endpoint of improved Combined Symptom and Medication Score. These ongoing studies provide critical proof-of-concept for the translational potential of the strategies discussed throughout this review and highlight the growing maturity of nanomedicine approaches for respiratory diseases.107–109
Conclusion and Prospect
This review has systematically examined the emerging frontier of biomedical nanoparticles designed to intercept the epithelial alarmin axis in allergic asthma. The reviewed studies demonstrate quantifiable therapeutic advances that transcend generic claims: DNAzyme-functionalized nucleic acid nanocapsules achieve up to 60% GATA-3 mRNA knockdown in immune cells, offering a precision gene-regulation platform inaccessible to antibody-based biologics; MSC-exosome-based tolerogenic nanoparticles reduce the required allergen dose for sublingual immunotherapy by approximately 5-fold while maintaining efficacy, addressing a critical safety limitation of conventional allergen-specific immunotherapy; and budesonide-loaded lipid nanoparticles prolong pulmonary residence time from 4 hours to 24 hours by avoiding macrophage clearance, transforming a standard-of-care corticosteroid into a sustained-release formulation with reduced dosing frequency.
These advances must be contextualized within the evolving therapeutic landscape. Compared to biologic agents requiring subcutaneous administration every 2–4 weeks at an annual cost of $20,000–40,000 per patient with variable response rates of 50–70%, NP-based approaches offer the potential for inhaled or transdermal delivery, targeting of intracellular transcription factors, and enhanced patient adherence through reduced dosing frequency. With approximately 26 million patients globally suffering from severe asthma inadequately controlled by current therapies, even modest improvements in efficacy or accessibility could yield substantial clinical and economic impact. The path forward requires scalable GMP manufacturing, biomarker-driven patient stratification, and rigorous head-to-head trials against standard-of-care biologics to establish the clinical superiority and cost-effectiveness that will ultimately determine whether these platforms fulfill their transformative potential.
Abbreviations
AECs, Airway epithelial cells; AHR, Airway hyperresponsiveness; AIT, Allergen-specific immunotherapy; AM, Alveolar macrophages; APCs, Antigen-presenting cells; bFGF, Basic fibroblast growth factor; BUD, Budesonide; CNPs, Cell membrane-coated nanoparticles; DAMPs, Damage-associated molecular patterns; DCs, Dendritic cells; DNAzyme-NAN, DNAzyme-functionalized nucleic acid nanocapsules; EMT, Epithelial-mesenchymal transition; GATA-3, GATA-binding protein 3; GBD, Global Burden of Disease; GINA, Global Initiative for Asthma; ICS, Inhaled corticosteroids; ILCs, Innate lymphoid cells; IL-25, Interleukin-25; IL-33, Interleukin-33; LNP, Lipid nanoparticle; LABA, Long-acting beta-agonists; MDC, Macrophage-derived chemokine; MSC-exos, Mesenchymal stem cell-derived exosomes; MMP-9, Metalloproteinase-9; NPs, Nanoparticles; NK, Natural killer; OVA, Ovalbumin; OX40L, OX40 ligand; PDGF-β, Platelet-derived growth factor-beta; PEG, Polyethylene glycol; TARC, Thymus and activation-regulated chemokine; TGF-β, Transforming growth factor-beta; Tregs, Regulatory T cells; TSLP, Thymic stromal lymphopoietin; tNPs, Tolerance-inducing nanoparticles; TfRs, Transferrin receptors; Tf-LNPs, Transferrin-conjugated lipid nanoparticles; Trp, Tryptophan; TSLP, T-selectin-like protein; Th2, Type 2 helper T; T2M, Type 2 myeloid; VEGF, Vascular endothelial growth factor.
Acknowledgments
Neither ChatGPT nor other generative language models were used for the ideation or writing process.
Disclosure
The author reports no conflicts of interest in this work.
References
1. Oh J, Kim S, Kim MS, et al. Global, regional, and national burden of asthma and atopic dermatitis, 1990–2021, and projections to 2050: a systematic analysis of the Global Burden of Disease Study 2021. Lancet Respir Med. 2025;13(5):425–16. doi:10.1016/S2213-2600(25)00003-7
2. 2025 Severe Asthma Guide. Global Initiative for Asthma - GINA. Available from: https://ginasthma.org/severe-asthma/.
3. Mortimer KJ, Cruz AA, Sepúlveda-Pachón IT, Jorga A, Vroling H, Williams C. Global herpes zoster burden in adults with asthma: a systematic review and meta-analysis. Eur Respir J. 2024;64(2):2400462. doi:10.1183/13993003.00462-2024
4. See KC. Impact of inhaled and intranasal corticosteroids on glucose metabolism and diabetes mellitus: a mini review. World J Diabetes. 2023;14(8):1202–1211. doi:10.4239/wjd.v14.i8.1202
5. Persaud PN, Tran AP, Messner D, et al. Perception of burden of oral and inhaled corticosteroid adverse effects on asthma-specific quality of life. Ann Allergy Asthma Immunol. 2023;131(6):745–751.e11. doi:10.1016/j.anai.2023.08.595
6. León B, Ballesteros-Tato A. Modulating Th2 Cell Immunity for the Treatment of Asthma. Front Immunol. 2021;12:637948. doi:10.3389/fimmu.2021.637948
7. Akdis CA. Does the epithelial barrier hypothesis explain the increase in allergy, autoimmunity and other chronic conditions? Nat Rev Immunol. 2021;21(11):739–751. doi:10.1038/s41577-021-00538-7
8. Howell I, Howell A, Pavord ID. Type 2 inflammation and biological therapies in asthma: targeted medicine taking flight. J Exp Med. 2023;220(7):e20221212. doi:10.1084/jem.20221212
9. Hansen S, Søndergaard MB, von Bülow A, et al. Clinical Response and Remission in Patients With Severe Asthma Treated With Biologic Therapies. CHEST. 2024;165(2):253–266. doi:10.1016/j.chest.2023.10.046
10. Mukherjee M, Forero DF, Tran S, et al. Suboptimal treatment response to anti-IL-5 monoclonal antibodies in severe eosinophilic asthmatics with airway autoimmune phenomena. Eur Respir J. 2020;56(4):2000117. doi:10.1183/13993003.00117-2020
11. Duchesne M, Okoye I, Lacy P. Epithelial cell alarmin cytokines: frontline mediators of the asthma inflammatory response. Front Immunol. 2022;13:975914. doi:10.3389/fimmu.2022.975914
12. Stanbery AG, Smita S, von Moltke J, Tait Wojno ED, Ziegler SF. TSLP, IL-33, and IL-25: not just for allergy and helminth infection. J Allergy Clin Immunol. 2022;150(6):1302–1313. doi:10.1016/j.jaci.2022.07.003
13. Sims JE, Williams DE, Morrissey PJ, et al. Molecular Cloning and Biological Characterization of a Novel Murine Lymphoid Growth Factor. J Exp Med. 2000;192(5):671–680. doi:10.1084/jem.192.5.671
14. Soumelis V, Reche PA, Kanzler H, et al. Human epithelial cells trigger dendritic cell mediated allergic inflammation by producing TSLP. Nat Immunol. 2002;3(7):673–680. doi:10.1038/ni805
15. Braile M, Fiorelli A, Sorriento D, et al. Human Lung-Resident Macrophages Express and Are Targets of Thymic Stromal Lymphopoietin in the Tumor Microenvironment. Cells. 2021;10(8):2012. doi:10.3390/cells10082012
16. Ito T, Wang YH, Duramad O, et al. TSLP-activated dendritic cells induce an inflammatory T helper type 2 cell response through OX40 ligand. J Exp Med. 2005;202(9):1213–1223. doi:10.1084/jem.20051135
17. Davoine F, Lacy P. Eosinophil Cytokines, Chemokines, and Growth Factors: emerging Roles in Immunity. Front Immunol. 2014;5:570. doi:10.3389/fimmu.2014.00570
18. Sahnoon L, Bajbouj K, Mahboub B, Hamoudi R, Hamid Q. Targeting IL-13 and IL-4 in Asthma: therapeutic Implications on Airway Remodeling in Severe Asthma. Clin Rev Allergy Immunol. 2025;68(1):44. doi:10.1007/s12016-025-09045-2
19. Drake MG, Scott GD, Blum ED, et al. Eosinophils increase airway sensory nerve density in mice and in human asthma. Sci Transl Med. 2018;10(457):eaar8477. doi:10.1126/scitranslmed.aar8477
20. Abdelaziz MH, Abdelwahab SF, Wan J, et al. Alternatively activated macrophages; a double-edged sword in allergic asthma. J Transl Med. 2020;18:58. doi:10.1186/s12967-020-02251-w
21. Deng R, Chen X, Zhang Y, et al. Short ragweed pollen promotes M2 macrophage polarization via TSLP/TSLPR/OX40L signaling in allergic inflammation. Mucosal Immunol. 2019;12(5):1141–1149. doi:10.1038/s41385-019-0187-8
22. Zhang H, Wei R, Yang X, et al. AMFR drives allergic asthma development by promoting alveolar macrophage–derived GM-CSF production. J Exp Med. 2022;219(5):e20211828. doi:10.1084/jem.20211828
23. Fort MM, Cheung J, Yen D, et al. IL-25 Induces IL-4, IL-5, and IL-13 and Th2-Associated Pathologies In Vivo. Immunity. 2001;15(6):985–995. doi:10.1016/S1074-7613(01)00243-6
24. Angkasekwinai P, Chang SH, Thapa M, Watarai H, Dong C. Regulation of IL-9 expression by IL-25 signaling. Nat Immunol. 2010;11(3):250–256. doi:10.1038/ni.1846
25. Yao X, Sun Y, Wang W, Sun Y. Interleukin (IL)-25: pleiotropic roles in asthma. Respirology. 2016;21(4):638–647. doi:10.1111/resp.12707
26. Wang YH, Angkasekwinai P, Lu N, et al. IL-25 augments type 2 immune responses by enhancing the expansion and functions of TSLP-DC–activated Th2 memory cells. J Exp Med. 2007;204(8):1837–1847. doi:10.1084/jem.20070406
27. Tsai ML, Tsai YG, Lin YC, et al. IL-25 Induced ROS-Mediated M2 Macrophage Polarization via AMPK-Associated Mitophagy. Int J Mol Sci. 2021;23(1):3. doi:10.3390/ijms23010003
28. Ualiyeva S, Lemire E, Aviles EC, et al. Tuft cell-produced cysteinyl leukotrienes and IL-25 synergistically initiate lung type 2 inflammation. Sci Immunol. 2021;6(66):eabj0474. doi:10.1126/sciimmunol.abj0474
29. Peng B, Sun L, Zhang M, et al. Role of IL-25 on Eosinophils in the Initiation of Th2 Responses in Allergic Asthma. Front Immunol. 2022;13:842500. doi:10.3389/fimmu.2022.842500
30. Salter BM, Oliveria JP, Nusca G, et al. IL-25 and IL-33 induce Type 2 inflammation in basophils from subjects with allergic asthma. Respir Res. 2016;17:5. doi:10.1186/s12931-016-0321-z
31. Angkasekwinai P, Park H, Wang YH, et al. Interleukin 25 promotes the initiation of proallergic type 2 responses. J Exp Med. 2007;204(7):1509–1517. doi:10.1084/jem.20061675
32. Duan W, Mehta AK, Magalhaes JG, et al. Innate Signals from Nod2 Block Respiratory Tolerance and Program Th2 Driven Allergic Inflammation. J Allergy Clin Immunol. 2010;126(6):1284–93.e10. doi:10.1016/j.jaci.2010.09.021
33. Abdu S, Xia J, Yuan H, et al. IL-25 Enhances B Cell Responses in Type 2 Inflammation Through IL-17RB Receptor. Allergy. 2025;80(4):965–975. doi:10.1111/all.16472
34. Corrigan CJ, Wang W, Meng Q, et al. T-helper cell type 2 (Th2) memory T cell-potentiating cytokine IL-25 has the potential to promote angiogenesis in asthma. Proc Natl Acad Sci U S A. 2011;108(4):1579–1584. doi:10.1073/pnas.1014241108
35. Huang ZX, Qiu ZE, Chen L, et al. Cellular mechanism underlying the facilitation of contractile response induced by IL-25 in mouse tracheal smooth muscle. Am J Physiol Lung Cell Mol Physiol. 2022;323(1):L27–L36. doi:10.1152/ajplung.00468.2021
36. Létuvé S, Lajoie-Kadoch S, Audusseau S, et al. IL-17E upregulates the expression of proinflammatory cytokines in lung fibroblasts. J Allergy Clin Immunol. 2006;117(3):590–596. doi:10.1016/j.jaci.2005.10.025
37. Gregory LG, Jones CP, Walker SA, et al. IL-25 drives remodelling in allergic airways disease induced by house dust mite. Thorax. 2013;68(1):82–90. doi:10.1136/thoraxjnl-2012-202003
38. Petersen BC, Budelsky AL, Baptist AP, Schaller MA, Lukacs NW. IL-25 induces type 2 cytokine production in a novel, steroid resistant IL-17RB+ myeloid population that exacerbates asthmatic pathology. Nat Med. 2012;18(5):751–758. doi:10.1038/nm.2735
39. Hough KP, Curtiss ML, Blain TJ, et al. Airway Remodeling in Asthma. Front Med. 2020;7:191. doi:10.3389/fmed.2020.00191
40. Seys SF, Grabowski M, Adriaensen W, et al. Sputum cytokine mapping reveals an ‘IL-5, IL-17A, IL-25-high’ pattern associated with poorly controlled asthma. Clin Exp Allergy. 2013;43(9):1009–1017. doi:10.1111/cea.12125
41. Hellings PW, Steelant B. Epithelial barriers in allergy and asthma. J Allergy Clin Immunol. 2020;145(6):1499–1509. doi:10.1016/j.jaci.2020.04.010
42. Weathington NM, Kanth SM, Gong Q, et al. IL-4 Induces IL17Rb Gene Transcription in Monocytic Cells with Coordinate Autocrine IL-25 Signaling. Am J Respir Cell Mol Biol. 2017;57(3):346–354. doi:10.1165/rcmb.2016-0316OC
43. Carriere V, Roussel L, Ortega N, et al. IL-33, the IL-1-like cytokine ligand for ST2 receptor, is a chromatin-associated nuclear factor in vivo. Proc Natl Acad Sci. 2007;104(1):282–287. doi:10.1073/pnas.0606854104
44. Schmitz J, Owyang A, Oldham E, et al. IL-33, an Interleukin-1-like Cytokine that Signals via the IL-1 Receptor-Related Protein ST2 and Induces T Helper Type 2-Associated Cytokines. Immunity. 2005;23(5):479–490. doi:10.1016/j.immuni.2005.09.015
45. Pividori M, Schoettler N, Nicolae DL, Ober C, Im HK. Shared and Distinct Genetic Risk Factors for Childhood Onset and Adult Onset Asthma: genome- and Transcriptome-wide Studies. Lancet Respir Med. 2019;7(6):509–522. doi:10.1016/S2213-2600(19)30055-4
46. Aneas I, Decker DC, Howard CL, et al. Asthma-associated genetic variants induce IL33 differential expression through an enhancer-blocking regulatory region. Nat Commun. 2021;12:6115. doi:10.1038/s41467-021-26347-z
47. TYF Halim, A MacLaren, MT Romanish, et al. Retinoic-Acid-Receptor-Related Orphan Nuclear Receptor Alpha Is Required for Natural Helper Cell Development and Allergic Inflammation. Immunity. 2012;37(3):463–474. doi:10.1016/j.immuni.2012.06.012
48. Spits H, Cupedo T. Innate Lymphoid Cells: emerging Insights in Development, Lineage Relationships, and Function. Annu Rev Immunol. 2012;30(30):2012):647–675. doi:10.1146/annurev-immunol-020711-075053
49. Stier MT, Zhang J, Goleniewska K, et al. IL-33 promotes the egress of group 2 innate lymphoid cells from the bone marrow. J Exp Med. 2018;215(1):263–281. doi:10.1084/jem.20170449
50. JL Barlow, S Peel, J Fox, et al. IL-33 is more potent than IL-25 in provoking IL-13–producing nuocytes (type 2 innate lymphoid cells) and airway contraction. J Allergy Clin Immunol. 2013;132(4):933–941. doi:10.1016/j.jaci.2013.05.012
51. Boberg E, Johansson K, Malmhäll C, Calvén J, Weidner J, Rådinger M. Interplay Between the IL-33/ST2 Axis and Bone Marrow ILC2s in Protease Allergen-Induced IL-5-Dependent Eosinophilia. Front Immunol. 2020;11:1058. doi:10.3389/fimmu.2020.01058
52. Cai T, Qiu J, Ji Y, et al. IL-17-producing-ST2+ILC2 plays a pathogenic role in lung inflammation. J Allergy Clin Immunol. 2019;143(1):229–244.e9. doi:10.1016/j.jaci.2018.03.007
53. Xiao Q, He J, Lei A, et al. PPARγ enhances ILC2 function during allergic airway inflammation via transcription regulation of ST2. Mucosal Immunol. 2021;14(2):468–478. doi:10.1038/s41385-020-00339-6
54. Steer CA, Mathä L, Shim H, Takei F. Lung group 2 innate lymphoid cells are trained by endogenous IL-33 in the neonatal period. JCI Insight. 2020;5(14):e135961. doi:10.1172/jci.insight.135961
55. de Kleer IM, Kool M, de Bruijn MJW, et al. Perinatal Activation of the Interleukin-33 Pathway Promotes Type 2 Immunity in the Developing Lung. Immunity. 2016;45(6):1285–1298. doi:10.1016/j.immuni.2016.10.031
56. Drake LY, Kita H. IL-33: biological Properties, Functions and Roles in Airway Disease. Immunol Rev. 2017;278(1):173–184. doi:10.1111/imr.12552
57. Chen CC, Kobayashi T, Iijima K, Hsu FC, Kita H. IL-33 dysregulates regulatory T (Treg) cells and impairs established immunological tolerance in the lungs. J Allergy Clin Immunol. 2017;140(5):1351–1363.e7. doi:10.1016/j.jaci.2017.01.015
58. Toki S, Goleniewska K, Zhang J, et al. TSLP and IL-33 reciprocally promote each other’s lung protein expression and ILC2 receptor expression to enhance innate type-2 airway inflammation. Allergy. 2020;75(7):1606–1617. doi:10.1111/all.14196
59. An G, Wang W, Zhang X, et al. Combined blockade of IL-25, IL-33 and TSLP mediates amplified inhibition of airway inflammation and remodelling in a murine model of asthma. Respirology. 2020;25(6):603–612. doi:10.1111/resp.13711
60. Vannella KM, Ramalingam TR, Borthwick LA, et al. Combinatorial targeting of TSLP, IL-25, and IL-33 in type 2 cytokine-driven inflammation and fibrosis. Sci Transl Med. 2016;8(337):337ra65. doi:10.1126/scitranslmed.aaf1938
61. Blank F, Fytianos K, Seydoux E, et al. Interaction of biomedical nanoparticles with the pulmonary immune system. J Nanobiotechnology. 2017;15(1):6. doi:10.1186/s12951-016-0242-5
62. Zuo X, Guo X, Gu Y, et al. Recent Advances in Nanomaterials for Asthma Treatment. Int J Mol Sci. 2022;23(22):14427. doi:10.3390/ijms232214427
63. Mitchell MJ, Billingsley MM, Haley RM, Wechsler ME, Peppas NA, Langer R. Engineering precision nanoparticles for drug delivery. Nat Rev Drug Discov. 2021;20(2):101–124. doi:10.1038/s41573-020-0090-8
64. Hammad H, Lambrecht BN. Dendritic cells and epithelial cells: linking innate and adaptive immunity in asthma. Nat Rev Immunol. 2008;8(3):193–204. doi:10.1038/nri2275
65. van Rijt LS, Jung S, Kleinjan A, et al. In vivo depletion of lung CD11c+ dendritic cells during allergen challenge abrogates the characteristic features of asthma. J Exp Med. 2005;201(6):981–991. doi:10.1084/jem.20042311
66. Aono Y, Suzuki Y, Horiguchi R, et al. CD109 on Dendritic Cells Regulates Airway Hyperreactivity and Eosinophilic Airway Inflammation. Am J Respir Cell Mol Biol. 2023;68(2):201–212. doi:10.1165/rcmb.2022-0109OC
67. Maldonado RA, LaMothe RA, Ferrari JD, et al. Polymeric synthetic nanoparticles for the induction of antigen-specific immunological tolerance. Proc Natl Acad Sci U S A. 2015;112(2):E156–E165. doi:10.1073/pnas.1408686111
68. Kenison JE, Stevens NA, Quintana FJ. Therapeutic induction of antigen-specific immune tolerance. Nat Rev Immunol. 2024;24(5):338–357. doi:10.1038/s41577-023-00970-x
69. Zhao J, He P, Jiang M, et al. Transdermally delivered tolerogenic nanoparticles induced effective immune tolerance for asthma treatment. J Control Release off J Control Release Soc. 2024;366:637–649. doi:10.1016/j.jconrel.2024.01.018
70. Karve T, Dandekar A, Agrahari V, Melissa Peet M, Banga AK, Doncel GF. Long-acting transdermal drug delivery formulations: current developments and innovative pharmaceutical approaches. Adv Drug Deliv Rev. 2024;210:115326. doi:10.1016/j.addr.2024.115326
71. Sadeghi M, Dehnavi S, Khodadadi A, et al. Immunomodulatory features of MSC-derived exosomes decorated with DC-specific aptamer for improving sublingual immunotherapy in allergic mouse model. Stem Cell Res Ther. 2024;15:481. doi:10.1186/s13287-024-04099-z
72. Kumar MA, Baba SK, Sadida HQ, et al. Extracellular vesicles as tools and targets in therapy for diseases. Signal Transduct Target Ther. 2024;9:27. doi:10.1038/s41392-024-01735-1
73. de Castro LL, Xisto DG, Kitoko JZ, et al. Human adipose tissue mesenchymal stromal cells and their extracellular vesicles act differentially on lung mechanics and inflammation in experimental allergic asthma. Stem Cell Res Ther. 2017;8(1):151. doi:10.1186/s13287-017-0600-8
74. Gholami M, Ghorban K, Sadeghi M, Dadmanesh M, Rouzbahani NH, Dehnavi S. Mesenchymal stem cells and allergic airway inflammation; a therapeutic approach to induce immunoregulatory responses. Int Immunopharmacol. 2023;120:110367. doi:10.1016/j.intimp.2023.110367
75. Dong B, Wang C, Zhang J, et al. Exosomes from human umbilical cord mesenchymal stem cells attenuate the inflammation of severe steroid-resistant asthma by reshaping macrophage polarization. Stem Cell Res Ther. 2021;12(1):204. doi:10.1186/s13287-021-02244-6
76. Abbaszadeh H, Ghorbani F, Abbaspour-Aghdam S, et al. Chronic obstructive pulmonary disease and asthma: mesenchymal stem cells and their extracellular vesicles as potential therapeutic tools. Stem Cell Res Ther. 2022;13:262. doi:10.1186/s13287-022-02938-5
77. Rao SP, Ge XN, Sriramarao P. Regulation of Eosinophil Recruitment and Activation by Galectins in Allergic Asthma. Front Med Lausanne. 2017;4:68. doi:10.3389/fmed.2017.00068
78. Pei W, Li X, Bi R, et al. Exosome membrane-modified M2 macrophages targeted nanomedicine: treatment for allergic asthma. J Control Release. 2021;338:253–267. doi:10.1016/j.jconrel.2021.08.024
79. Wu GR, Zhou M, Wang Y, et al. Blockade of Mbd2 by siRNA-loaded liposomes protects mice against OVA-induced allergic airway inflammation via repressing M2 macrophage production. Front Immunol. 2022;13:930103. doi:10.3389/fimmu.2022.930103
80. Xin Y, Wen R, Song D, et al. Emu-miR-10a-5p in Echinococcus multilocularis-derived-extracellular vesicles alleviates airway inflammation in mice with allergic asthma by inhibiting macrophage M2a polarization through LIF-mediated JAK1–STAT3 signaling. Front Immunol. 2025;16:1577349. doi:10.3389/fimmu.2025.1577349
81. Sikazwe CT, Laing IA, Imrie A, et al. Nasal Cytokine Profiles of Patients Hospitalised with Respiratory Wheeze Associated with Rhinovirus C. Viruses. 2019;11(11):1038. doi:10.3390/v11111038
82. Fang RH, Gao W, Zhang L. Targeting drugs to tumours using cell membrane-coated nanoparticles. Nat Rev Clin Oncol. 2023;20(1):33–48. doi:10.1038/s41571-022-00699-x
83. Jin H, Li J, Zhang M, et al. Berberine-Loaded Biomimetic Nanoparticles Attenuate Inflammation of Experimental Allergic Asthma via Enhancing IL-12 Expression. Front Pharmacol. 2021;12:724525. doi:10.3389/fphar.2021.724525
84. Tew XN, Lau NJ, Chellappan DK, et al. Immunological axis of berberine in managing inflammation underlying chronic respiratory inflammatory diseases. Chem Biol Interact. 2020;317:108947. doi:10.1016/j.cbi.2020.108947
85. Gao C, Huang Q, Liu C, et al. Treatment of atherosclerosis by macrophage-biomimetic nanoparticles via targeted pharmacotherapy and sequestration of proinflammatory cytokines. Nat Commun. 2020;11(1):2622. doi:10.1038/s41467-020-16439-7
86. Song T, Yao L, Zhu A, et al. Cathepsin B-Activatable Bioactive Peptide Nanocarrier for High-Efficiency Immunotherapy of Asthma. Int J Nanomed. 2024;19:8059–8070. doi:10.2147/IJN.S455633
87. Rochman Y, Kotliar M, Klingler AM, et al. Allergen-Specific mRNA-Lipid Nanoparticle Therapy for Prevention and Treatment of Experimental Allergy in Mice. J Clin Invest. 2025:e194080. doi:10.1172/JCI194080.
88. Zhu J, Yamane H, Cote-Sierra J, Guo L, Paul WE. GATA-3 promotes Th2 responses through three different mechanisms: induction of Th2 cytokine production, selective growth of Th2 cells and inhibition of Th1 cell-specific factors. Cell Res. 2006;16(1):3–10. doi:10.1038/sj.cr.7310002
89. Gavitt TD, Hartmann AK, Sawant SS, Mara AB, Szczepanek SM, Rouge JL. A GATA3 Targeting Nucleic Acid Nanocapsule for In Vivo Gene Regulation in Asthma. ACS Nano. 2021;15(7):11192–11201. doi:10.1021/acsnano.0c07781
90. Hartmann AK, Cairns-Gibson DF, Santiana JJ, Tolentino MQ, Barber HM, Rouge JL. Enzymatically Ligated DNA-Surfactants: unmasking Hydrophobically Modified DNA for Intracellular Gene Regulation. Chembiochem Eur J Chem Biol. 2018. doi:10.1002/cbic.201800302
91. Jürgens DC, Müller JT, Nguyen A, Merkel OM. Tailoring lipid nanoparticles for T-cell targeting in allergic asthma: insights into efficacy and specificity. Eur J Pharm Biopharm. 2024;198:114242. doi:10.1016/j.ejpb.2024.114242
92. McGee HS, Yagita H, Shao Z, Agrawal DK. Programmed Death-1 antibody blocks therapeutic effects of T-regulatory cells in cockroach antigen-induced allergic asthma. Am J Respir Cell Mol Biol. 2010;43(4):432–442. doi:10.1165/rcmb.2009-0258OC
93. Turner JA, Stephen-Victor E, Wang S, et al. Regulatory T Cell-Derived TGF-β1 Controls Multiple Checkpoints Governing Allergy and Autoimmunity. Immunity. 2020;53(6):1202–1214.e6. doi:10.1016/j.immuni.2020.10.002
94. Coomes SM, Kannan Y, Pelly VS, et al. CD4+ Th2 cells are directly regulated by IL-10 during allergic airway inflammation. Mucosal Immunol. 2017;10(1):150–161. doi:10.1038/mi.2016.47
95. Chinen T, Kannan AK, Levine AG, et al. An essential role for the IL-2 receptor in Treg cell function. Nat Immunol. 2016;17(11):1322–1333. doi:10.1038/ni.3540
96. Khumalo J, Kirstein F, Hadebe S, Brombacher F. IL-4Rα signaling in CD4+CD25+FoxP3+ T regulatory cells restrains airway inflammation via limiting local tissue IL-33. JCI Insight. 2020;5(20):e136206. doi:10.1172/jci.insight.136206
97. Liu Q, Wang X, Liu X, et al. Use of Polymeric Nanoparticle Platform Targeting the Liver to Induce Treg-Mediated Antigen-Specific Immune Tolerance in a Pulmonary Allergen Sensitization Model. ACS Nano. 2019;13(4):4778–4794. doi:10.1021/acsnano.9b01444
98. Li J-D, Yin J. Interleukin-10-alveolar macrophage cell membrane-coated nanoparticles alleviate airway inflammation and regulate Th17/regulatory T cell balance in a mouse model. Front Immunol. 2023;14:1186393. doi:10.3389/fimmu.2023.1186393
99. Tessier B, Tsapis N, Fattal E, Moine L. Emerging nanoparticle platforms to improve the administration of glucocorticoids. J Control Release off J Control Release Soc. 2023;358:273–292. doi:10.1016/j.jconrel.2023.04.039
100. Expert Panel Working Group of the National Heart, Lung, and Blood Institute (NHLBI) administered and coordinated National Asthma Education and Prevention Program Coordinating Committee (NAEPPCC), Cloutier MM, Baptist AP, et al. 2020 Focused Updates to the Asthma Management Guidelines: a Report from the National Asthma Education and Prevention Program Coordinating Committee Expert Panel Working Group. J Allergy Clin Immunol. 2020;146(6):1217–1270. doi:10.1016/j.jaci.2020.10.003.
101. Reddel HK, Bacharier LB, Bateman ED, et al. Global Initiative for Asthma Strategy 2021: executive Summary and Rationale for Key Changes. Am J Respir Crit Care Med. 2022;205(1):17–35. doi:10.1164/rccm.202109-2205PP
102. Janson C, Melén E, de Fine Licht S, et al. Prescribing patterns of budesonide/formoterol maintenance and reliever therapy in patients with asthma in Sweden. Allergy. 2023;78(8):2311–2314. doi:10.1111/all.15710
103. Krings JG, Sekhar TC, Chen V, et al. Beginning to Address an Implementation Gap in Asthma: clinicians’ Views of Prescribing Reliever Budesonide-Formoterol Inhalers and SMART in the United States. J Allergy Clin Immunol Pract. 2023;11(9):2767–2777. doi:10.1016/j.jaip.2023.05.023
104. Zuo X, Gu Y, Guo X, et al. Preparation of Budesonide-Loaded Liposomal Nanoparticles for Pulmonary Delivery and Their Therapeutic Effect in OVA-Induced Asthma in Mice. Int J Nanomed. 2024;19:673–688. doi:10.2147/IJN.S441345
105. Liu X, Zhang L, Li S, et al. Harnessing Surface Hydrophilicity of Inhalable Nanoparticles for Precision Delivery of Glucagon-like Peptide-1 Receptor Agonists or Anti-Asthmatic Therapeutics. ACS Nano. 2025;19(24):22357–22375. doi:10.1021/acsnano.5c05745
106. Quispe C, Cruz-Martins N, Manca ML, et al. Nano-Derived Therapeutic Formulations with Curcumin in Inflammation-Related Diseases. Oxid Med Cell Longev. 2021;2021:3149223. doi:10.1155/2021/3149223
107. Shinde V, Bhikha S, Hoosain Z, et al. Efficacy of NVX-CoV2373 Covid-19 Vaccine against the B.1.351 Variant. N Engl J Med. 2021;384(20):1899–1909. doi:10.1056/NEJMoa2103055
108. Stuart ASV, Shaw RH, Liu X, et al. Immunogenicity, safety, and reactogenicity of heterologous COVID-19 primary vaccination incorporating mRNA, viral-vector, and protein-adjuvant vaccines in the UK (Com-COV2): a single-blind, randomised, Phase 2, non-inferiority trial. Lancet Lond Engl. 2022;399(10319):36–49. doi:10.1016/S0140-6736(21)02718-5
109. Shaw RH, Greenland M, Stuart ASV, et al. Persistence of immune response in heterologous COVID vaccination schedules in the Com-COV2 study - A single-blind, randomised trial incorporating mRNA, viral-vector and protein-adjuvant vaccines. J Infect. 2023;86(6):574–583. doi:10.1016/j.jinf.2023.03.027
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Comparative in vitro Study on Immunomodulatory Effects of Nano-Hydroxpatite Powders on Human Immune Cells
Grubczak K, Kretowska-Grunwald A, Szalaj U, Starosz A, Swieszkowski W, Moniuszko M
International Journal of Nanomedicine 2026, 21:555182
Published Date: 6 May 2026