Back to Journals » International Journal of Nanomedicine » Volume 20
Targeting Metal Ion Homeostasis for Regulated Cell Death-Amplified Tumor Nanomedicine
Authors Chen Q, Li K, Li J ![]() , Liu X, Li J, Xu L, Han Y, Zou T, Wang X, Yao Y
, Liu X, Li J, Xu L, Han Y, Zou T, Wang X, Yao Y ![]() , Mao Y
, Mao Y
Received 1 October 2025
Accepted for publication 11 December 2025
Published 24 December 2025 Volume 2025:20 Pages 15627—15654
DOI https://doi.org/10.2147/IJN.S571617
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Sachin Mali
Qiqing Chen,1,* Kun Li,2,* Jinzhuo Li,3 Xue Liu,4 Jiaxin Li,5 Lianuang Xu,5 Yabing Han,5 Ting Zou,5 Xingrong Wang,5 Yangcheng Yao,6 Yuhang Mao5
1Department of Ultrasound, Hainan General Hospital, Hainan Affiliated Hospital of Hainan Medical University, Haikou, 570311, People’s Republic of China; 2Department of Thoracic Surgery, Shanghai Pulmonary Hospital, Tongji University School of Medicine, Shanghai, 200433, People’s Republic of China; 3Faculty of Health Sciences, University of Macau, Macau, SAR, 999078, People’s Republic of China; 4Department of Biological Chemical Engineering, College of Chemical Engineering and Materials Science, Tianjin University of Science and Technology, Tianjin, 300222, People’s Republic of China; 5School of Medicine, Ankang University, Ankang, 725000, People’s Republic of China; 6Center for Reproductive Medicine, Guangdong Women and Children Hospital, Jinan University, Guangzhou, 511400, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yangcheng Yao, Email [email protected] Yuhang Mao, Email [email protected]
Abstract: Amidst escalating global health challenges, neoplastic diseases remain a predominant cause of morbidity and mortality, exerting complex and far-reaching effects on human health and societal well-being. The advent of precision medicine has ushered in an era of tailored therapeutic strategies, leveraging individual genetic profiles, tumor microenvironmental features, and exogenous factors to redefine oncology care. Central to these advances is the understanding of cell death, a fundamental biological process encompassing both programmed and non-programmed forms. Programmed cell death is orchestrated through sophisticated genetic and molecular mechanisms. Emerging evidence underscores the role of metal ion dyshomeostasis, particularly of iron, copper, zinc, sodium, magnesium, manganese, and calcium, in disrupting intracellular signaling and metabolic equilibrium, thereby inducing lethal cascades in malignant cells. Concurrently, innovations in nanomedicine have enabled precise modulation of ion fluxes within tumors, enhancing therapeutic specificity while minimizing systemic toxicity. This confluence of ion-mediated cell death mechanisms and nanotechnology not only exemplifies a transformative approach in cancer treatment but also aligns seamlessly with the tenets of precision medicine, offering novel pathways for therapeutic innovation and clinical translation.
Keywords: regulated cell death, metal ion-induced cell death, tumor nanomedicine, precision cancer therapy
Introduction
The growing incidence of neoplasms worldwide presents a major public health challenge, profoundly affecting human health and quality of life in multifaceted ways.1 According to the latest 2025 statistics from the American Cancer Society, it is projected that there will be 2,041,910 new cancer cases and 618,120 cancer deaths in the United States by 2025.2 The array of cancer treatment modalities includes surgery, radiotherapy, chemotherapy, targeted therapy, and immunotherapy—each plays a unique role in clinical practice, though none alone achieves universal efficacy for all tumor types.3 Currently, the most optimal approach to cancer treatment is precision medicine, which is tailored to the patient’s genetic profile, specific cancer traits, and environmental influences.4 This approach holds promise for personalized cancer treatment; however, its translation to clinical practice is hindered by two categories of challenges: (1) scientific and technical challenges, especially difficulty in identifying specific metal ion targets for heterogeneous tumors, limitations in nanocarrier stability and targeting efficiency; (2) clinical and practical challenges in high production costs of nanomedicines, lack of large-scale clinical data on long-term safety.
Cell death is primarily categorized into two distinct types: Programmed Cell Death (PCD) and Non-Programmed Cell Death (NPCD). These categories differ significantly in their biological implications, induction mechanisms, and morphological characteristics. PCD is a complex process meticulously regulated by genes, encompassing various forms such as apoptosis, autophagy, pyroptosis, programmed necrosis, ferroptosis, and cuproptosis.5 In contrast, NPCD is a passive process of cell death triggered by severe damage or external stimuli, without genetic regulation. Each mode of cell death involves unique signaling cascades and molecular mechanisms, exhibiting distinct morphological and biochemical features. Emerging research indicates that metal ion overload disrupts cellular homeostasis, thereby influencing the regulation of cell death.6 Common intracellular metal ions include calcium (Ca2+), ferric ions (Fe3+), copper ions (Cu2+), zinc ions (Zn2+), sodium ions (Na+), manganese ions (Mn2+), magnesium ions (Mg2+), among others.7 Under normal physiological conditions, the concentrations of these metal ions are dynamically balanced through the regulation of various transport proteins and storage reservoirs. However, factors such as radiation, tumors, or accidental injuries can lead to ion overload, causing signaling disruption and toxicity, ultimately resulting in cell death.8 Metal ions are chosen as the focus of this review for three key reasons: (1) They are essential for cellular metabolism, and tumor cells exhibit distinct metal ion dependencies that can be targeted selectively; (2) Metal ion dyshomeostasis directly induces irreversible cell death without relying on traditional apoptotic pathways, bypassing drug resistance; (3) Their concentration and distribution can be precisely modulated by nanomedicines, enabling spatiotemporal control over therapeutic effects.
Gaining a deeper understanding and modulating the intricate regulatory networks of metal ion-induced cell death may offer novel therapeutic strategies and hope for cancer treatment.9 Moreover, the advent of nanomedicine has provided innovative tools and approaches for precision cancer treatment. By integrating nanotechnology with disciplines like biology and pharmacy, it has enhanced the targeting and specificity of cancer therapies, significantly reducing toxic side effects and broadening the prospects for precision cancer treatment.10 In recent years, nanomedicine has become a critical tool for metal ion-based tumor therapy. Key advancements include: the development of pH/ROS-responsive nanocarriers for targeted metal ion delivery, the design of multifunctional nanoplatforms that synergize metal ion-induced cell death with photodynamic/sonodynamic therapy, and the application of metal-based nanozymes to regulate intracellular ion metabolism. These progressions lay the groundwork for translating metal ion-targeted strategies into clinical practice. Nanomedicine, through precise regulation of metal ion metabolism, has emerged as a crucial bridge for the integration of cell death mechanisms with cancer therapy, offering the potential to greatly improve the efficiency and personalization of cancer treatments.11 Therefore, this review will focus on recent advancements in the regulatory networks of cell death induced by metal ions during tumor progression, with a special emphasis on how nanomedicines target metal ion metabolism pathways to amplify regulated cell death effects (enhancing metal ion accumulation, synergizing with other therapeutic modalities) and their potential in precision cancer therapy (Figure 1).12
 |
Figure 1 Metal ion-mediated regulated cell death (RCD) represents an emerging frontier in cancer therapeutics, encompassing mechanisms such as ferroptosis, cuproptosis, and manganese death et al. These processes are driven by respective metal-dependent oxidative stress, protein aggregation, and metabolic disruption. In nanomedicine, tailored nanoparticles enable targeted delivery of metal ions and potentiate synergistic antitumor effects when combined with adjuvant modalities including sonodynamic therapy (SDT), photodynamic therapy (PDT), gas therapy, and starvation therapy (ST) et al. By precisely modulating ion homeostasis and amplifying reactive oxygen species generation, nanomaterial-based strategies offer a promising approach to induce metal-related RCD, highlighting their essential role in advancing precision oncology treatments. |
Cell Death and Tumor Progression
Cell Death Mode
For clarity, core concepts are defined herein: (1) Regulated Cell Death (RCD): A genetically or molecularly controlled cell death process that maintains tissue homeostasis or eliminates abnormal cells, encompassing both Programmed Cell Death (PCD) and some regulated necrotic pathways; (2) Programmed Cell Death (PCD): A subclass of RCD orchestrated by specific genes, including apoptosis, autophagy, pyroptosis, and metal ion-induced death; (3) Non-Programmed Cell Death (NPCD): A passive, unregulated cell death process triggered by severe external damage, typically manifested as necrosis. This review focuses on PCD mediated by metal ions, as it is the primary target of nanomedicines. Cell death can be broadly categorized into two main types: Programmed Cell Death (PCD) and Non-Programmed Cell Death (NPCD).13 PCD is a genetically regulated and orderly process of cell death that is essential for development, maintaining physiological balance, and eliminating abnormal cells. The common forms of PCD include apoptosis, autophagy, pyroptosis, and cell death induced by metal ions.14 In contrast, NPCD is a passive process initiated by uncontrollable external factors such as mechanical damage, infection, or toxic agents. This type of cell death usually accompanies inflammation and leads to substantial damage to adjacent tissues, with necrosis being a typical example.15 The distinctions between these two forms of cell death in terms of their initiating mechanisms, regulatory processes, and biological significance lay the groundwork for comprehending cell injury and disease mechanisms.
The defining feature of NPCD is its lack of regulation by gene-encoded signals. It is typically triggered by unpredictable external factors and is often accompanied by inflammatory responses, causing harm to surrounding tissues.16 For instance, mechanical cell death resulting from physical injuries like cutting, crushing, or rupture leads to the direct loss of cell integrity, subsequent leakage of cellular contents, and ultimately cell death.17 Necrosis caused by conditions such as ischemia, hypoxia, toxic exposure, or infection usually results in cell swelling, cytoplasmic dissolution, and eventual membrane rupture, leading to the release of cellular contents and activation of immune responses.18 Necrosis serves as a significant driver in tumor development and can drastically affect the efficacy of anti-tumor therapies.19 When intracellular components like DNA, ATP, and HMGB1 leak out, they activate immune cells and pro-inflammatory signaling pathways, which may induce chronic inflammation and create a cancer-promoting environment within the tumor microenvironment.20 Moreover, bioactive factors released from necrotic cells, including reactive oxygen species and pro-inflammatory cytokines, can activate oncogenic signaling in normal tissues, increasing the mutation rate in healthy cells and fostering tumor growth.21 Under certain circumstances, molecules such as HMGB1 and heat shock proteins released from necrotic cells might suppress anti-tumor immune responses, causing tumor-associated macrophages to adopt an immunosuppressive phenotype, thereby facilitating tumor growth and metastasis.22
On the other hand, the hallmark of PCD is its status as a highly regulated, gene-encoded process that typically does not provoke significant inflammation and can occur under both physiological and certain pathological conditions.23 Apoptosis generally occurs through two main pathways: the extrinsic pathway (activated by death receptors on the cell surface, such as Fas/FasL and Caspase-8) and the intrinsic pathway (activated by intracellular stress signals, particularly the release of cytochrome c from mitochondria, which activates Caspase-9). Both pathways ultimately activate Caspase-3, leading to systematic disintegration of the cell.24 Autophagy-induced cell death typically occurs due to excessive autophagy, involving processes such as mTOR inhibition, autophagosome formation, and the accumulation of LC3-II. While the primary function of autophagy is to clear damaged organelles, excessive autophagy can lead to cell death, especially under conditions of energy deprivation or excessive stress.25 Pyroptosis, induced by the activation of the inflammasome NLRP3, results in cell death through Caspase-1 cleavage, which further activates Gasdermin D. Gasdermin D forms pores in the cell membrane, causing leakage of cellular contents and triggering a strong inflammatory response.26 Metal ion-induced cell death primarily results from disruptions in metal ion metabolism, involving oxidative stress and lipid peroxidation.27 Different types of PCD have diverse effects on tumor biology. They can suppress tumor growth by activating cell death mechanisms, but due to adaptive changes in tumor cells, they may also promote tumor growth and metastasis. By modulating these cell death pathways, new targets and strategies for cancer treatment can be explored. The differences and relationships between the four major types of PCD are summarized in Table 1.
 |
Table 1 Types and Characteristics of Cancer Cell Death |
PCD and Tumor Therapy
PCD plays a multifaceted role in tumor therapy, influencing tumor cell survival, drug resistance, metastasis, and immune evasion. As the understanding of PCD mechanisms deepens, more studies are targeting these pathways as novel therapeutic targets for cancer treatment. In cancer therapy, inducing apoptosis is a common and essential strategy, especially in chemotherapy, radiotherapy, and immunotherapy. Most chemotherapy drugs, such as cisplatin, paclitaxel, and cyclophosphamide, kill tumor cells by activating the apoptosis pathway. Drug-induced DNA damage or oxidative stress activates key molecules, such as p53, caspases, and members of the Bcl-2 family, initiating apoptosis (Figure 2A)28 However, many tumor cells evade apoptosis by altering key signaling molecules in the apoptosis pathway, such as p53 mutations, upregulation of Bcl-2 family members, and inhibition of caspase activity, leading to resistance to chemotherapy.29 Liu et al found that the Bcl-2 family inhibitor ABT-737 enhanced tumor cell apoptosis by targeting the inhibition of Bcl-2 and Bcl-xL expression. This inhibitor has shown promise in clinical trials.30
 |
Figure 2 (A): The apoptotic signaling pathway of DNA damage or oxidative stress involves mitochondrial membrane permeability changes,cytoplasmic release of cytochrome c, activation of the Caspase cascade, and initiation of programmed cell death. Adapted from Ref 28 (B): Formation of autophagosomes through encapsulation of damaged organelles or protein aggregates by autophagic membrane via a circular structure. Adapted from Ref 31 (C): A tumor immunotherapy strategy based on a nanodrug delivery system integrates multiple mechanisms including drug loading, targeted delivery, immune activation, and tumor microenvironment regulation. Adapted from Ref 32 (D): Schematic representation of a nanoparticle-based ferroptosis-targeted therapy strategy encompassing nanocarrier design, molecular mechanisms, and in vitro/in vivo validation models. Adapted from Ref 33. |
Autophagy shows a dual role in tumor development. In the early stages of tumor development, autophagy helps eliminate damaged cellular components, thus suppressing tumor formation. As the tumor progresses, cancer cells may utilize autophagy to obtain energy and metabolites, which helps them survive in nutrient-deprived and hypoxic conditions (Figure 2B).31 Paulo et al suggested that autophagy inhibitors such as chloroquine and hydroxychloroquine, by inhibiting the acidification of auto phagolysosomes and preventing autophagosome degradation, enhance the cytotoxic effects of chemotherapy drugs on tumor cells.34 Additionally, He et al found that upregulation of autophagy using rapamycin could restore certain cellular functions in multiple myeloma cells, improving the prognosis of patients with multiple myeloma during bleomycin treatment. Thus, regulating autophagy levels offers a promising strategy for tumor therapy (Figure 2C).32
Pyroptosis, through the release of pro-inflammatory cytokines such as IL-1β and IL-18, activates local immune responses that help clear tumor cells. Compared to apoptosis, pyroptosis activates the immune system more strongly, promoting tumor immune surveillance.35 Recent studies have found that activating the pyroptosis pathway may enhance the efficacy of cancer therapy by strengthening the immune response. Activation of the NLRP3 inflammasome or Gasdermin D can promote tumor cell death while inducing a strong local immune response, aiding in tumor clearance.36 Moreover, Zhang et al conducted bioinformatics analysis on several glioma public databases and identified four key pyroptosis-related genes (CASP4, CASP9, GSDMC, IL1A), constructing a risk score model. Their study revealed that these genes significantly correlate with the diagnostic prognosis and immune microenvironment status in glioma patients.37 As a consequence, pyroptosis not only boosts cell death, but also plays a significant part in immunotherapy for cancer.38
The link between metal ion-induced cell death and treatment for tumors is extensive and complex (Table 2). Cell death can be mediated by a variety of metal ions, including calcium, iron, copper, and zinc. These pathways provide novel targets for the therapy of cancers, especially those that do not respond to traditional treatments.39 For example, ferroptosis increases intracellular iron buildup and peroxide production, which results in tumor cell death. Cancer cell death can result from increasing tumor cells’ susceptibility to ferroptosis by targeting GPX4 or iron metabolism pathways (Figure 2D).33,40 A novel anti-tumor approach might be developed by focusing on copper metabolism pathways or by directly triggering cuproptosis. Under specific circumstances, copper ions could lead to copper overload, which activates the cuproptosis pathway to stops the proliferation of tumor cells.41 Tumor therapy that targets metal ion-induced cell death pathways provides a novel treatment option. This novel therapeutic method has showed substantial promise in treating cancers with a poor prognosis and treatment resistance by controlling metal ion metabolism to kill tumor cells. As a result, this article blends it with nanomedicine, concentrating on recent advances in research into the regulation of metal ion-induced cell death in precision cancer therapy.42
 |
Table 2 Metal Ion Cell Death |
Ferroptosis-Inspired Tumor Nanomedicine
Overview of Ferroptosis
Ferroptosis, initially described by Dixon et al in 2012, is a type of iron-dependent programmed cell death caused by an excessive accumulation of lipid peroxides on the cell membrane.43 Distinct mitochondrial morphological alterations obviously, including reduced mitochondrial volume, increased membrane density, and decreased mitochondrial cristae, separate it from apoptosis, necrosis, autophagy, and other forms of programmed cell death.40 The accumulation of reactive oxygen species (ROS) and the overproduction of lipid peroxides are core features of ferroptosis. This process involves the interplay between the ferroptosis execution system (which includes the synthesis and peroxidation of phospholipid polyunsaturated fatty acids (PUFA-PL), iron metabolism, and mitochondrial metabolism) and the ferroptosis defense system, comprising the glutathione peroxidase 4 (GPX4)-reduced glutathione (GSH) system, ferroptosis suppressor protein 1 (FSP1)-coenzyme QH2 (CoQH2) system, dihydroorotate dehydrogenase (DHODH)-CoQH2 system, and guanosine triphosphate cyclohydrolase 1 (GCH1) -tetrahydrobiopterin (BH4) system. When the activity of the ferroptosis execution system significantly exceeds that of the defense system, ferroptosis occurs.44,45 As a unique mechanism of cell death, targeting tumor cell ferroptosis offers a novel therapeutic opportunity for cancer treatment.46,47 In recent years, significant progress has been made in understanding the role of ferroptosis in tumor biology and its potential in cancer therapy.
Mechanism of Ferroptosis
The core mechanism of ferroptosis lies in the disruption of the balance between the ferroptosis execution system and the ferroptosis defense system. The execution system involves lipid metabolism and peroxidation, iron metabolism, and mitochondrial metabolism, while the defense system primarily includes the GPX4-GSH system, FSP1-CoQH2 system, DHODH-CoQH2 system, and GCH1-BH4 system. Polyunsaturated fatty acids (PUFAs), major components of cell membranes, are esterified and incorporated into phospholipids (PUFA-PLs) through the action of long-chain acyl-CoA synthetase 4 (ACSL4).48 These PUFA-PLs are oxidized by iron ions to generate phospholipid hydroperoxides (PLOOH), with lipoxygenases (LOXs) playing a role in this process under the regulation of iron ions. The accumulation of PLOOH leads to irreversible damage to the cell membrane, triggering ferroptosis.49 Iron ions are the most critical trigger of ferroptosis. Iron is ingested through the diet and initially exists as ferric iron, which is reduced to ferrous iron in the duodenum. Fe2+ is then transported into intestinal epithelial cells via divalent metal transporter 1 (DMT1).50 Once inside the cell, iron can either be stored by ferritin or released into the bloodstream via ferroportin. Fe2+ reacts with hydrogen peroxide (H2O2) in the Fenton reaction, producing highly reactive hydroxyl radicals (·OH), which oxidize the PUFAs in cell membranes, accelerating the formation of lipid peroxides.51 Mitochondria, as the main source of ROS and ATP within cells, also drive mitochondrial lipid peroxidation and ferroptosis from various functional aspects.52,53
Nanomedicine of Ferroptosis in Cancer Therapy
The progression of nanomedicine has facilitated the creation of precise therapeutic modalities for the induction of ferroptosis, particularly within the realm of cancer management. Principal therapeutic strategies encompass the targeted delivery of ferrous ions, enhancement of lipid peroxidation processes,suppression of GPX4 activity, and the implementation of synergistic combination therapies. These methodologies exhibit substantial potential for augmenting the effectiveness of cancer treatments through the strategic exploitation of ferroptosis mechanisms to selectively eradicate neoplastic cells. Nanomedicines amplify ferroptosis through three key mechanisms: (1) Targeted iron delivery: pH-sensitive nanocarriers release Fe2+ specifically in the acidic tumor microenvironment, increasing the labile iron pool and enhancing Fenton reactions to generate hydroxyl radicals; (2) GSH depletion: Nanoparticles deplete intracellular glutathione, inhibiting GPX4 activity and amplifying lipid peroxidation; (3) Synergy with other therapies: Photodynamic therapy (PDT)-responsive nanoplatforms generate reactive oxygen species (ROS) while delivering iron, creating a “double amplification” effect on ferroptosis.32
Cancer Drug Resistance and Ferroptosis
Tumor resistance refers to the decreased or lost sensitivity of cancer cells to anticancer drugs, leading to suboptimal or ineffective treatment outcomes. This resistance can be classified into primary resistance (present at the start of treatment) and acquired resistance (developed during treatment).54 Numerous studies have indicated that inducing ferroptosis can overcome both primary and acquired resistance in tumor cells.55,56 Olaparib, a PARP inhibitor, has been approved by the FDA for treating ovarian cancer patients with BRCA mutations. However, patients without BRCA mutations show some degree of resistance to this drug.57 Xie et al conducted in vivo and in vitro studies and found that β-caryophyllene, a novel ferroptosis inducer, combined with cetuximab, induces iron-dependent ROS accumulation, GSH depletion, lipid peroxidation, upregulation of HO-1 and transferrin, and downregulation of ferroptosis negative regulators (GPX4, SLC7A11, FTH1, glutaminase, and SLC40A1). This combination treatment enhanced drug sensitivity in KRAS-mutant colorectal cancer patients by inducing ferroptosis and inhibiting epithelial-mesenchymal transition (EMT) (Figure 3A).58,59
 |
Figure 3 (A) Schematic representation of the cell death mechanism induced by β-elemene and cetuximab, involving iron ion metabolism regulation and lipid peroxide accumulation. Adapted from Ref 58 (B):Depiction of a novel ferroptosis-based combination therapy strategy via nanomaterial-mediated chemodynamic-immunotherapy synergy.Adapted from Ref 60a. |
Therefore, based on the ability of ferroptosis inducers to enhance the sensitivity of cancer therapy, the further use of nanocarriers can enhance the targeted delivery of chemotherapy drugs (such as cisplatin, paclitaxel, etc.) to tumors. By simultaneously promoting ferroptosis induction, the anti-tumor effect is enhanced, achieving a 1+1 > 2 effect. Nanocarriers can carry both chemotherapy drugs and ferroptosis inducers to exert a dual strike on tumor cells. Yu et al designed butyrate-modified nanoparticles that interact with the multifunctional ligand butyrate and monocarboxylate transporter 1 (MCT-1) to increase the uptake of sorafenib and salvianolic acid, thereby inducing ferroptosis to treat hepatocellular carcinoma. Butyrate-modified nanoparticles effectively and continuously enhance the trans-epithelial transport in the intestine, promote drug accumulation in the liver, and enhance the drug uptake in liver cancer cells (Figure 3B).60,61
Immunotherapy and Ferroptosis
Nanomedicine can combine immunotherapy (such as immune checkpoint inhibitors like PD-1/PD-L1 antibodies) with ferroptosis inducers to enhance the anti-tumor effects of immune cells and break the tumor’s immune escape mechanisms through ferroptosis.32,62 Nanomaterials can also release immunostimulatory factors in the tumor microenvironment while inhibiting ferroptosis, thereby activating anti-tumor immune responses.63–65 Chih et al synthesized zero-valent iron nanoparticles (ZVI-NP), which can induce mitochondrial dysfunction, intracellular oxidative stress, and lipid peroxidation, leading to ferroptosis in lung cancer cells. Additionally, ZVI-NP converts pro-tumor M2 macrophages to anti-tumor M1 macrophages, reduces the number of regulatory T cells, downregulates PD-1 and CTLA4 in CD8+ T cells, and enhances their cytotoxic activity against cancer cells. ZVI-NP demonstrates the potential for an integrated anti-cancer strategy by synergistically inducing ferroptosis in cancer cells and reprogramming the immune-suppressive microenvironment (Figure 4A).66 Wang et al constructed a BSA-FA-functionalized iron-based metal-organic framework (TPL@TFBF), which induces ferroptosis and pyroptosis in tumor cells, releasing large amounts of damage-associated molecular patterns (DAMPs), thereby triggering immunogenicity and stimulating a strong systemic anti-tumor immune response. This approach showed excellent efficacy in inhibiting melanoma lung metastasis in vivo (Figure 4B).67 Guo et al first upregulated miR-21-3p to promote IFN-γ-mediated ferroptosis through enhanced lipid peroxidation. They constructed miR-21-3p-loaded gold nanoparticles, which systemically delivered the treatment, increasing the efficacy of anti-PD-1 antibody therapy for melanoma. These nanoparticles showed no significant side effects in preclinical mouse models.65,68
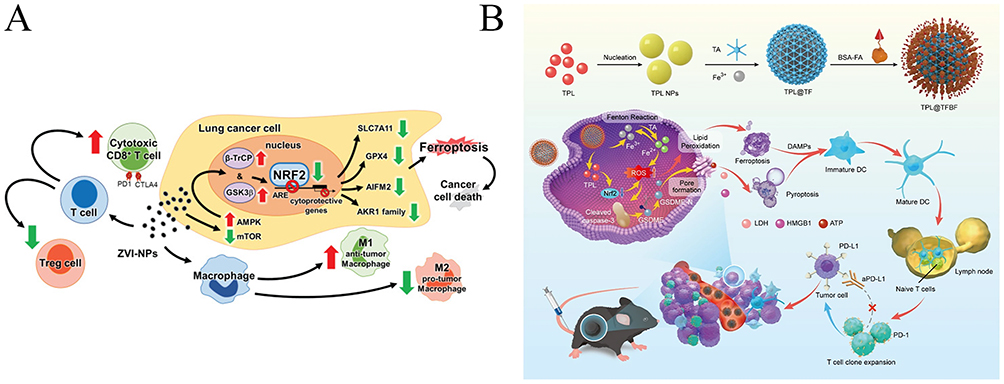 |
Figure 4 (A): Visualization of the dynamic interactions between lung cancer cells and immune cells in the tumor niche. Green arrows indicate “promotion/activation” effects, while red arrows indicate “inhibition/downregulation” effects. Adapted from Ref 66 (B) Schematic of a nanoparticle-based ferroptosis-inducing drug delivery system and its therapeutic mechanism in tumor treatment. Adapted from Ref 67. |
Multimodal Dynamic Therapy Combined with Ferroptosis
Multimodal Dynamic Therapy includes Photodynamic Therapy (PDT), Sonodynamic Therapy (SDT), and Radiodynamic Therapy (RDT), which induce tumor cell death through the ferroptosis mechanism within cells using different nanomaterials.69 These therapies have significant potential in treating refractory tumors, especially deep-seated ones, by enhancing tumor-killing effects and overcoming drug resistance.70
PDT is a treatment method that uses photosensitizers and specific wavelengths of light to treat cancer and other diseases. The basic principle involves activating a photosensitizer (a chemical substance that absorbs light and produces reactive oxygen species, ROS) under light exposure at a specific wavelength. These ROS can damage cell structures and induce cell death, particularly in cancer cells (Figure 5A).71 Zhou et al reported a microwave-induced copper-cysteine (Cu-Cy) nanoparticle PDT, which, through cell viability assays (Cell Titer-Blue) and live/dead cell tests, demonstrated that under microwave radiation, Cu-Cy could effectively destroy HCT15 colorectal cancer cells. Ferroptosis inhibitors could alleviate this process. Cu-Cy nanoparticles show promising clinical applications in microwave dynamic therapy, effectively inhibiting tumor cell proliferation by inducing ferroptosis, providing a potential solution to overcome cancer resistance (Figure 5B).72 Wang et al synthesized a chemical therapy agent (HA-Fc-Mal) and constructed hyaluronic acid-based chemical/chemical dynamic (CT/CDT) nanoparticles (HCM@DOX) through self-assembly of HA-Fc-Mal and doxorubicin (DOX). This formulation could induce both ferroptosis and apoptosis, thus offering an effective treatment for triple-negative breast cancer. The tumor inhibition rate reached 81.87%, with significant suppression of lung metastasis and excellent biosafety (Figure 5C).73–75
 |
Figure 5 (A): Design principles of a nanodiagnosis and treatment system integrating ferrodynamic mechanisms with multimodal therapeutic actions through combination with photodynamic therapy (PDT).Adapted from Ref 71 (B) Molecular mechanism of copper ion carrier (Cu-Cy)-mediated regulation of tumor angiogenesis through ferroptosis pathway modulation.Adapted from Ref 72 (C) Illustration of the combined tumor treatment process involving nanoparticle-induced apoptosis/ferroptosis with concurrent photothermal therapy. Adapted from Ref 74 (D) Diagram of an integrated anticancer therapeutic platform combining nanotechnology and ferroptosis mechanisms, spanning from drug design to targeted delivery and efficacy monitoring. Adapted from Ref 76 (E) Formation mechanisms of tumor hypoxic microenvironment and associated therapeutic challenges, with potential intervention strategies for multimodal combination therapies including oxygen-generating radiotherapy (O2-RDT).Adapted from Ref 77. |
SDT is a newly developed cancer treatment method that combines the advantages of ultrasound technology and PDT. It activates sonosensitizers via ultrasound radiation to induce the production of ROS thereby inhibiting and killing tumor cells.78,79 The basic principle involves using low-intensity ultrasound to activate a sonosensitizer (usually a nanomaterial), which reacts within tumor cells to generate a large amount of ROS.80 These ROS can trigger oxidative stress reactions within the cells, leading to ferroptosis in tumor cells.81 Compared to traditional radiotherapy and chemotherapy, SDT offers higher penetration, does not produce radiation, and can target tumors, reducing damage to normal tissues.82 By using nanomaterials for drug delivery, sonosensitizers can be better transported to the tumor site (Figure 5D).76,83 The high surface area and excellent targeting properties of nanoparticle carriers enable the drug to accumulate in tumor tissues and exert therapeutic effects.84 Yang et al synthesized copper-doped molybdenum-based polyoxometalate nanozymes (CP) using a one-step method at room temperature, integrating enzymatic catalysis and sonocatalytic therapy to enhance cancer treatment efficacy. Their study demonstrated that CP exhibits multiple enzyme-like activities, including peroxidase, catalase, and glutathione peroxidase. Additionally, its semiconductor properties were optimized through oxygen vacancy modulation, improving sonocatalytic efficiency. Under ultrasound irradiation, CP generated various ROS, synergistically depleted GSH, disrupted tumor redox homeostasis, and induced ferroptosis, ultimately effectively inhibiting tumor growth.85 Moreover, ferroptosis, an emerging form of programmed cell death, has gained increasing attention for its synergistic effects with SDT. Through nano-catalysis, SDT can enhance oxidative stress, deplete GSH, effectively induce ferroptosis, and improve tumor-killing efficacy.32,86,87
Additionally, RDT utilizes X-rays or γ-rays to activate radiosensitizers, which, in the presence of oxygen or water, generate ROS, thereby inducing oxidative stress damage to tumor cells and ultimately leading to apoptosis or other forms of programmed cell death (Figure 5E).77 In recent years, studies have shown that RDT can induce ferroptosis through mechanisms mediated by nanomaterials, such as oxidative stress-induced depletion of GSH and inhibition of GPX4.88 The application of RDT combined with nanotechnology holds great promise, particularly in the treatment of difficult-to-treat deep-seated solid tumors, such as gliomas, pancreatic cancer, and liver cancer, demonstrating superior therapeutic effects.89 Jiang et al developed a simplified radiodynamic therapy (RDT) strategy based on CsI (Na)@MgO nanoparticles and 5-ALA. The combination of CsI(Na)@MgO and 5-ALA in in vitro experiments enhanced radiation-induced ROS, increasing damage to mitochondria, DNA, and lipids, thereby reducing cell proliferation and clonogenicity.90
Cuproptosis-Based Anti-Tumor Nanotherapy
Overview of Cuproptosis
Cuproptosis is a recently identified form of regulatory cell death dependent on copper, with potential links to cancer development and progression.91 It is a recently identified form of regulatory cell death dependent on copper, with potential links to cancer development and progression.92 Copper death was first described by the Todd team in 2022 as a unique form of programmed cell death that is distinct from apoptosis, ferroptosis, and necroptosis.41,93 It is closely associated with mitochondrial function and the lipoic acid (LA) pathway. By attaching itself to lipoic acid-modified mitochondrial proteins and causing their aggregation, a decrease in iron-sulfur (Fe-S) clusters, and ultimately protein toxicity stress and cell death.94,95 The research of cuproptosis provides fresh insights into tumor biology and prospective possibilities for generating newer treatment approaches.
Mechanism of Cuproptosis
Cuproptosis is the excessive buildup of copper ions, which impairs mitochondrial activity and cellular metabolism, eventually leading to cell death. This process mainly includes the accumulation of copper ions, their interaction with mitochondria, the aggregation of dihydrolipoamide S-acetyltransferase (DLAT), oxidative stress, and cell damage and death.96,97 In the human body, copper is a necessary trace metal that catalyzes and supports a number of biological functions, such as antioxidant defense, mitochondrial respiration, and the creation of bioactive chemicals.98 The concentration of copper within cells must be tightly regulated, as excessive copper accumulation can lead to cell toxicity, especially when copper ions cannot be effectively removed from the body.99 Copper ions predominantly target mitochondria, where they bind to DLAT within the mitochondria. This binding reduces the levels of iron-sulfur (Fe-S) clusters, affecting the mitochondrial respiratory chain and ATP production, thereby leading to mitochondrial dysfunction.100 Iron-sulfur clusters are integral components of various key enzymes, and their reduction impacts cellular metabolism and antioxidant capacity, leading to oxidative stress, the production of reactive oxygen species (ROS), and the oxidation of cellular lipids, proteins, and DNA, which results in cell damage.101 Overall, the main features of cuproptosis include mitochondrial dysfunction, protein aggregation, and oxidative stress.102 Extensive research has confirmed that cuproptosis is associated with various diseases, including cancer, neurodegenerative diseases, cardiovascular diseases, and metabolic disorders. A better understanding of cuproptosis mechanisms may provide new targets and therapeutic solutions for these disorders.
Nanomedicine of Cuproptosis in Cancer Therapy
Cuproptosis, a novel type of programmed cell death, has been linked to the formation and progression of cancer. Many malignancies have much greater copper contents than normal tissues, and excessive copper buildup may be connected to tumor growth, metastasis, and medication resistance.103 Current research focuses on medications that regulate cuproptosis, with the goal of treating cancer by reducing copper levels or causing cuproptosis directly. These medications can destroy tumor cells by boosting copper ion levels or strengthening copper’s cytotoxic effects.104 Li et al undertook a thorough examination of the expression variations, gene mutations, and methylation alterations of cuproptosis-related genes (CRGs) across malignancies. They calculated the cuproptosis score (CS) for individual samples using single-sample gene set enrichment analysis (ssGSEA) and investigated the relationship between CS and copy number variations, clinical features, immune-related genes, tumor mutation burden (TMB), microsatellite instability (MSI), and tumor immune dysfunction and exclusion (TIDE). The findings revealed that CRGs were strongly expressed in a variety of malignancies, with CDKN2A having the highest mutation frequency and being associated with a poor prognosis in the majority of cancers. Additionally, CS was significantly negatively correlated with tumor microenvironment scores in over ten types of tumors and positively correlated with PD-L1 in eleven types of tumors, suggesting that cuproptosis might be involved in tumor immune evasion.105
Moreover, current treatments targeting cuproptosis mainly rely on copper ion carriers to deliver copper (Cu) into cancer cells, thereby inducing cell death. However, existing copper ion carriers are typically small molecules with short blood half-lives, making it challenging to transport sufficient copper into cancer cells (Figure 6A).106,107 Xing et al designed a ROS-sensitive polymer (PHPM) for co-delivery of elesclomol (ES) and copper, forming nanoparticles (NP@ESCu). After entering cancer cells, ES and copper are rapidly released in response to excessive intracellular ROS. The synergistic effect of ES and copper not only induces cuproptosis to kill cancer cells but also triggers an immune response, providing a novel strategy for future cancer therapy (Figure 6B).32,108 Ning et al constructed a platelet vesicle (PV)-coated copper(I) oxide (Cu2O)/TBP-2 copper death sensitization system (PTC) for multi-induction of tumor copper death. PTC was prepared by a physical extrusion method, combining the AIE photosensitizer (TBP-2), Cu2O, and PV together. PTC rapidly degrades under acidic conditions and hydrogen peroxide (H2O2) in tumor cells, releasing copper ions. Under light exposure, TBP-2 quickly enters the cell membrane and generates hydroxyl radicals, depleting glutathione and inhibiting copper efflux. The accumulated copper leads to the aggregation of acylated proteins and the loss of iron-sulfur proteins, triggering protein toxicity stress and ultimately causing copper-induced cell death. Nanomedicines amplify cuproptosis by overcoming the limitations of small-molecule copper carriers: ROS-sensitive nanoparticles accumulate in tumors, release copper ions and elesclomol in response to high intracellular ROS, and downregulate copper efflux proteins to maintain high intracellular copper levels, this “retention-amplification” mechanism significantly enhances cuproptosis efficacy. PTC treatment targets and induces copper death in tumor cells both in vitro and in vivo, significantly inhibiting lung metastasis of breast cancer, increasing the number of central memory T cells in peripheral blood, and preventing tumor recurrence.109 Wang et al developed nanoparticles, PCD@Cu, by self-assembling ROS-responsive prodrug PEG-TK-DOX and GSH-responsive prodrug PEG-DTPA-SS-CPT, which are coordinated with Cu2+. In TNBC cells, increased ROS and GSH levels disrupted the PCD@Cu structure, leading to the release of Cu2+, DOX, and CPT, while depleting GSH. DOX and CPT induce apoptosis accompanied by immunogenic cell death in TNBC cells. Meanwhile, PCD@Cu downregulated the expression of copper-transporting ATPase 2 (ATP7B), resulting in significant copper accumulation in TNBC cells, further promoting copper death in tumor cells. In vitro and in vivo experiments confirmed that PCD@Cu induces apoptosis and copper death in TNBC, activates the immune system, and demonstrates strong antitumor capabilities.110
 |
Figure 6 (A): Illustration of the PEG-TK-DOX nanodrug delivery system composed of polyethylene glycol, thymidine kinase, and doxorubicin for enhanced tumor-targeted drug accumulation and efficacy. Adapted from Ref 107 (B) Schematic diagram of the therapeutic mechanism of ROS-sensitive linker-based nanoparticles for tumor treatment via drug release, ROS elevation, and immune activation. Adapted from Ref 32 (C) Comprehensive depiction of tumor therapy enhancement mechanisms through metabolic regulation, oxidative stress induction, and immunogenic cell death-mediated immune activation. Adapted from Ref 111 (D) Diagram of the research strategy for copper ion-delivering nanoparticles in cancer therapy, demonstrating cuproptosis/ferroptosis induction and MRI-monitored in vivo tracking. Adapted from Ref 112. |
Combining ferroptosis inducers with copper ion carriers to simultaneously target ferroptosis and cuproptosis in cancer cells represents an innovative cancer treatment strategy. Li et al reported the design of a core-shell structured nanoparticle CuP/Er, which is used for the synergistic delivery of copper (Cu) and erlotinib (Er) to cancer cells, achieving synergistic copper death and ferroptosis. CuP/Er promotes T-cell proliferation and infiltration, and when combined with immune checkpoint inhibitors, it effectively reactivates T-cells, mediating tumor regression in mouse colon adenocarcinoma and triple-negative breast cancer, while preventing tumor metastasis (Figure 6C).111 Huang et al developed hollow mesoporous copper-iron sulfide (HMCIS) nanoparticles loaded with disulfiram (DSF), modified with polyethylene glycol (PEG) and folic acid (FA) (ie, DSF@HMCIS-PEG-FA), which can rapidly release DSF, H2S, Cu2+, and Fe2⁺ in the acidic tumor microenvironment (TME). The released H2S enhances hydrogen peroxide (H2O2) levels and acidity within tumor cells, accelerating Fenton (Fe2+) and Fenton-like (Cu2+) reactions, thereby achieving a potent ferroptosis effect in tumors. Ferroptosis in tumor cells manifests as severe mitochondrial damage and glutathione (GSH) depletion, leading to excess intracellular copper ions, which then trigger copper death. Copper death disrupts mitochondria, damages iron-sulfur (Fe-S) proteins and increases oxidative stress within the cell by releasing free Fe3⁺. These interconnected processes form a self-amplifying ferroptosis-copper death cycle, exhibiting strong anticancer capabilities, which have been validated in cancer cells and tumor-bearing mice (Figure 6D).112,113
Zinc Death-Promoted Tumor Nano-Suppression
Overview of Zinc Death
Zinc death, also known as zinc-induced cell death, is a newly discovered form of programmed cell death associated with abnormal accumulation and imbalance of zinc ions (Zn2+) within cells. Although zinc plays crucial physiological roles in the body, its excess or deficiency can lead to various pathological conditions, including cell death.114,115
As an essential trace element, zinc participates in numerous biochemical processes. Zinc acts as a cofactor for various enzymes, playing a role in protein synthesis, DNA repair, and antioxidant responses. Zinc finger proteins bind to DNA to regulate gene expression. Zinc ions regulate cell signaling pathways, which affect cell proliferation, differentiation, and death.116,117 Zinc death has been linked to a variety of diseases, including neurological disorders, cardiovascular disease, and cancer.
Mechanisms of Zinc Death
Zinc death is a cell death process caused by intracellular zinc ion overload, which can occur through different mechanisms, including exogenous zinc exposure or endogenous zinc homeostasis imbalance.118 Zinc ion buildup affects mitochondrial activity, resulting in loss of membrane potential and decreased energy generation. This, in turn, enhances oxidative stress, which causes excessive ROS creation and interferes with protein folding and stability, resulting in protein aggregation or denaturation. Loss of cell membrane integrity causes leakage of cellular contents and cell death.119,120 The molecular pathways involved include the ROS/NF-κB, STING, MAPK, and Hedgehog pathways.121,122 These mechanisms are linked and collectively determine the cell’s fate during zinc excess. Understanding these systems can aid in the development of new therapeutic techniques to treat diseases caused by zinc imbalances.123
Nanomedicine of Zinc Death in Cancer Therapy
Excessive zinc ions impair intracellular redox equilibrium, causing protein aggregation, mitochondrial malfunction, and cell membrane damage, ultimately resulting in cell death. These findings provide a foundation for applying zinc death in cancer treatment. Additionally, zinc death interacts with other forms of cell death such as ferroptosis and apoptosis. Excessive zinc ions may influence lipid peroxidation and GSH levels, thereby modulating ferroptosis pathways and affecting cancer cell survival (Figure 7A).123–125 As an emerging form of cell death, zinc death offers new perspectives and directions for cancer therapy (Figure 7B).126,127
 |
Figure 7 (A) Systematic elucidation of molecular mechanisms underlying ZnO NPs-Lut complex-mediated ferroptosis through triple-pathway coordination: iron homeostasis disruption, antioxidant defense system impairment, and lipid peroxide accumulation.Adapted from Ref 123 (B) Nanodrug delivery platform leveraging synergistic interactions between zinc oxide nanomaterials and iron ions for multi-pathway induction of tumor cell apoptosis.Adapted from Ref 127 (C) Demonstration of nanomaterial-based tumor therapeutic strategy (ZnO NPs) targeting intracellular/extracellular biochemical reactions and zinc-specific cell death regulatory mechanisms.Adapted from Ref 128 (D) Schematic overview of the cytotoxic cascade initiated by nanoparticles, demonstrating multilevel disruption of cellular homeostasis leading to programmed cell death.Adapted from Ref 129. |
Jan et al assessed the 10-year survival rate of 284 consecutive, unscreened kidney cancer patients in relation to selenium and zinc levels in blood and serum, considering variables such as age at diagnosis, sex, smoking status, type of surgery, and histopathological results. Their study combined selenium and zinc levels and observed a risk ratio of 12.4 for kidney cancer mortality, suggesting that these levels could serve as prognostic indicators for kidney cancer patient survival.130 Achuth et al described the synthesis and characterization of zinc oxide nanoparticles (ZnO-NPs) and their physical properties, with particle sizes ranging from 15 nm to 55 nm. They evaluated the potential of ZnO-NPs as a therapeutic agent for ovarian cancer using established human ovarian cancer cell lines. Their findings showed that ZnO-NPs caused acute oxidative stress and protein toxicity stress in ovarian cancer cells, resulting in cell death via apoptosis. Furthermore, the cytotoxic effects of ZnO-NPs were marginally increased as the nanoparticle size dropped (Figure 7C).128,131 Tabrizi et al devised a folic acid-conjugated ZnO-decorated bovine serum albumin/silicon nanoparticles (FZBS-NP) delivery system to explore its antioxidant activity and pro-apoptotic potential in human pancreatic, breast, lung, and colon cancer cell lines. Their research indicated that FZBS-NP exhibited excellent antioxidant activity and had the potential to selectively trigger apoptosis in human colon and breast cancer cells while sparing normal cells, making it a powerful and safe anti-cancer chemical.132 Kavipriya et al studied the antibacterial, antibiofilm, and cytotoxic characteristics of biocompatible zinc oxide (ZnONP) nanoparticles produced from Gymnema sylvestre. The study indicated that at a concentration of 250 μg/mL, ZnONPs significantly inhibited biofilm formation, resulting in lower density and structural alterations. ZnONPs’ cytotoxicity on breast cancer cells was dose-dependent, with an IC50 value of 19.4 µg/mL. AO/EB staining revealed early and late apoptosis of breast cancer cells under fluorescence microscopy. Hemolytic activity tests confirmed the biocompatibility of ZnONPs (Figure 7D).129,133–135
Calcicoptosis-Elevated Cancer Therapeutic Strategy
Overview of Calcicoptosis
Calcicoptosis is a type of cell death caused by a disruption in intracellular calcium ion (Ca2+) homeostasis.It involves a series of events that begin with unusually high cytosolic Ca2+ concentrations and end with either intentional or accidental cell death (Figure 8A).136 Ca2+, a ubiquitous second messenger, regulates critical physiological processes such as gene expression, synaptic plasticity, and excitation-contraction coupling through tightly regulated spatiotemporal dynamics under physiological conditions.137 Intracellular resting concentrations are around 100 nM and extracellular levels are 1–2 mM. Plasma membrane Ca2+-ATPases (PMCAs), Na+/Ca2+ exchangers (NCXs),endoplasmic reticulum Ca2+ storage,and mitochondrial Ca2+ buffering mechanisms work together to maintain homeostasis. When exposed to pathological stimuli such as oxidative stress, ischemia-reperfusion injury,neurotoxic agents, or inflammatory factors, dysregulation of the the mineral calcium regulating network can cause various types of cell death, including apoptosis provoked by the mitochondrial the mineral calcium overload, necrosis set off by calpain activation;the autophagy process linked to lysosomal membranes permeabilization, or iron metabolism involving lipid peroxidation. Current research suggests that abnormal calcium signaling is intimately related to pathological processes such as neuronal loss in Alzheimer’s disease, cardiac injury caused by ischemia-reperfusion, and tumor treatment resistance (Figure 8B).138 Understanding its molecular mechanisms has been a critical focus of cell suicide research.139
 |
Figure 8 (A) Mechanism and effects of calcium electroporation in cancer therapy.Ref 136 (B) Mechanism of oxytosis. Oxytosis is a calcium regulated oxidative glutamate cytotoxicity resulted due to calcium accumulation into the cells.Ref 138. |
Mechanisms of Calcicoptosis
Calcicoptosis is a type of dying cell that occurs when intracellular Ca2+ homeostasis is disrupted. The process is primarily triggered by Ca2+ excess,which leads to cascade reactions. When inside of cells Ca2+ concentrations rise abnormally, the mitochondrial permeability transition pore (mPTP) opens irreversibly, causing the collapse of the membrane potential of mitochondrial cells,impaired the production of ATP, and the dissolution of cytochrome c. This activates the intrinsic apoptotic pathway.140,141 Ca2+-dependent proteases,including calpain,and nucleases are activated, resulting in cytoskeletal protein breakdown and destruction of DNA. Oxidative Ca2+ overload leads to increased ROS generation,causing oxygen consumption and additional damage to structures within cells. Through the PERK/CHOP pathway, this process also creates a positive feedback loop with ER stress, which eventually results in apoptosis, necroptosis, or ferroptosis, among other types of cell death.142,143 Numerous pathways of signaling, including the calpain-caspase cascade, ROS/NF-κB, oxidative apoptosis, and stress in the ER responses, work in concert to accomplish this complex process.144
Nanomedicine of Calcicoptosis in Cancer Therapy
Research on anti-tumor therapies based on calcicoporosis has advanced significantly in recent years. The fundamental idea behind these tactics is to use the distinct calcium homeostasis imbalance of tumor cells to specifically create intracellular Ca2+ overload using nanodrug methods of distribution, which sets off a series of cell death mechanisms.145 Theoretically, preferential the death of tumor cells is possible because tumor cells are more susceptible to Ca2+ excess than normal cells because of aberrant expression of calcium-regulating proteins and their endoplasmic reticulum malfunction. Researchers have produced biodegradable calcium-based nanoparticles that can quickly disintegrate in the cancerous microenvironment (acidic pH, high reductivity) and release Ca.2+146 Dong et al developed a nanoscale NCOF-based nanoagent, CaCO3@COF-BODIPY-2I@GAG. This nanoagent contains CaCO3 particulates and is surface-functionalized with a photosensitizer (PS) BODIPY-2I and glycosaminoglycan (GAG) targeting the CD44 receptors on digestive tract tumor cells. Photo-triggered production of 1O2 not only damages tumor cells but also causes the mitochondrial malfunction and Ca2+ overload, resulting in increased antitumor efficacy through synergistic PDT and Ca2+ overload therapy.147 Huang’s group created a calcium phosphate (CaP)-based oxygen self-supplied nanosystem to combat tumor hypoxia and trigger tumor-specific cascade catalysis.148
This nanoplatform is made up of mixed CaP-based nanoparticles that have been co-loaded with catalase and the photosensitizer DVDMS, with calcium phosphide acting as both a drug carrier and a release function. Smart drug delivery methods,including Ca2+-sensitive polymeric carriers, allow for targeted medication release at tumor locations. Jiang et al presented a 3D-printed bone-filling scaffold capable of loading both therapeutic medicines and bone regeneration factors for the treatment of bone cancers. The scaffold was created by layering polydopamine-hybridized ZIF-8 and polydopamine-modified hydroxyapatite (HAP) onto a gelatin-based scaffold. As a drug carrier platform, the scaffold was loaded with the bone regeneration factor BMP-2 and the anticancer medication cisplatin, which efficiently stimulated osteogenic differentiation and reduced tumour growth. Furthermore, multimodal synergistic therapeutic systems, which include photodynamic therapy, ferroptosis inducers, and other techniques, can improve therapeutic success. These nanosystems can not only directly trigger calcium-dependent cell death, but also co-deliver chemotherapeutic drugs,resulting in a calcium overload-chemotherapy combination treatment that greatly improves anticancer results.149,150
Sodium Overload-Induced Cancer Cell Nano-Killing
Sodium homeostasis is essential for cellular life activities. Na+-K+ ATPase and ion channels regulate intracellular sodium concentration at the concentration of 10–15 mM and extracellular saline concentration with 145 mM, creating a critical gradient in concentration. The resulting gradient is not only necessary for maintaining membrane potential and action potential conduction, but it also helps to regulate cell volume, pH balance, and nutrition transportation, among other fundamental physiological activities. More recent studies have discovered that disrupting this delicate equilibrium resulting to intracellular Na+ overload might cause a novel form of programmed cell death called “sodium death”. Abnormal sodium channel activation or Na+/K+-ATPase malfunction leads to an increase in Na+ influx,causing membrane depolarization,osmotic imbalance,and cellular edema.151,152 Research suggests that Na+ overload can cause secondary Ca2+ retention by hindering the Na+/Ca+ exchanger (NCX), resulting in mitochondrial membrane potential collapse, explosive reactive oxygen species (ROS) increase (up to 5–8 times baseline level), and ATP depletion (over 70% reduction). Activation of stress pathways, including JNK/p38 MAPK, NF-κB, and the NLRP3 inflammasome, leads to a combination of apoptosis and the death of tissue.153,154
Wan et aldeveloped a chemical called necrocide 1 (NC1) that causes necrotic cell death via sodium overload, a mechanism known as necrosis by sodium overload (NECSO). NC1 targets TRPM4, a non-selective monovalent cation channel. It enhances Na+ influx,which causes cell necrosis. Luo et al used a dissociated retinal neuron culture model from C57B6 mice to examine the role of sodium ions in retinal cell death. They used excitotoxic agonists (glutamate, NMDA, KA) in combination with the concentration of ions modulation and blocking agents.Their findings revealed that removing extracellular sodium totally prevented cell death,but removing extracellular calcium only partially decreased it. The deadly calcium influx was mostly mediated by the reverse activity of the sodium-calcium exchanger.155,156 Intracellular sodium chloride overflow was required for pathogenic calcium influx, which then caused amacrine cell death,indicating that sodium overload plays a critical role in excitotoxicity-induced retinal neuron death.Sodium overload-induced cell death is important in cardiac ischemia-reperfusion injury, neuronal death in stroke,and the anticancer effects of certain chemotherapy medicines. Its distinct ion-dependent mechanism suggests a promising approach for developing novel therapeutic methods against NCX inhibitors and others. However, the precise molecular regulatory network of sodium death, particular biomarkers,and its implications in other pathophysiological processes have yet to be fully defined157 (Figure 9).
 |
Figure 9 Flowchart summarizing the mechanism of sodium-induced cell death (NECSO). Pathological stimuli increase intracellular Ca2+ levels, activating the TRPM4 channel and resulting n significant Na+ influx. The subsequent osmotic imbalance leads to cell swelling, rupture, and ultimately sodium-induced necrotic cell death. |
Manganese-Associated Cancer Cell Death
Overview of Manganese-Associated Cell Death
Manganese-associated cell death occurs when the homeostasis of manganese ions (Mn2+) is disrupted. Manganese, a vital trace element for the human body, performs critical physiological processes under normal conditions, including regulating SOD activity, bone growth, and neurotransmitter production.158 Nevertheless, abnormally high intracellular manganese ion concentrations trigger a cascade of pathological processes that eventually lead to cell death. This type of cell death has distinct molecular properties, such as mitochondrial malfunction, increased oxidative stress,and aberrant protease activity.159,160 From a pathological point of view, manganese-associated cell death has been related to occupational manganese toxicity, neurological disorders,and the development and progression of some malignancies. In recent years, with more research into metal ion homeostasis, the processes of manganese-associated cell death have emerged as a new emphasis in cell death research, demonstrating tremendous usefulness, particularly in the exploration of neuroprotective techniques and the development of innovative anticancer medicines.161,162
Mechanisms of Manganese-Associated Cell Death
Manganese-associated cell death is caused by a combination of critical pathogenic events. Mn2+ competitively binds to electron transfer chain complexes II and III in the mitochondria, limiting oxidative phosphorylation and impairing ATP generation. Mn2+ opens the mitochondrial transmembrane transition pore (mPTP), causing membrane potential collapse and cytochrome C release.163 This activates caspase-dependent apoptosis pathways. Mn2+ catalyzes the formation of huge amounts of hydroxyl radicals via Fenton-like processes, which damage essential biomolecules such as lipids that make up membranes, proteins, and DNA. Notably, Mn2+ inhibits iron transport proteins, leading to iron buildup and cross-activation with ferroptosis pathways. Furthermore, Mn2+ indirectly activates calpain by boosting intracellular calcium ion levels, which leads to breakdown of cytoskeletal proteins. This process can be especially significant for neurodegenerative illnesses.164
Nanomedicine of Manganese-Associated Cell Death in Cancer Therapy
Significant advancement has recently been made in tumor nanotechnology in medicine using manganese-associated cell death pathways. Manganese-based nanotechnology can release Mn2+ in the TME, preferentially killing tumor cells by oxidative stress and mitochondrial damage. Li et al created a near-infrared light-triggered titanium nanomicelle (PPIR780-ZMS) that produced ROS and caused immune-stimulating cell death, activated the cGAS-STING pathway, increased T cell infiltration and anticancer immunological responses, and effectively suppressed primary tumors and lung metastases.165,166 Mn2+ has been shown to activate the STING signaling pathway, leading to increased type I interferon release and improved antitumor resistance. Luo et al created a pH-responsive nanoplatform (CM NPs) loaded with curcumin, CaCO3, and MnO2 and coated with tumor cell membranes. This platform releases Ca2+ and Mn2+ in the acidic tumor microenvironment, inducing ROS production,immune-stimulating cell death, alleviating a lack of oxygen and triggering the cGAS-STING the path. The aforementioned system enhances anticancer immune responses by modulating critical ion concentrations in tumors, promoting macrophage polarization, and dendritic cell maturation.167 It also synergizes with αPD-1 immunotherapy for better therapeutic effects (Figure 10A).168 Ma et al created a temperature-sensitive composite hydrogel system, PF-127@MnCl2/alginate microspheres (ALG-MS)@MgCl2, that can release Mn2+ quickly and sustainably to manage the tumor microenvironment. In vitro and in vivo investigations confirmed that this hydrogel significantly improved the efficacy of PD-L1 immune checkpoint inhibitor therapy by activating CD8+ T cells and natural killer cells, converting immunized cold tumors into immune hot tumors, and enhancing tumor responsiveness to immunotherapy. Regarding transport techniques, targeting ligand changes and the development of pH/ROS-responsive carriers have considerably improved manganese-based nanomedicines’ tumor-targeting efficacy and safety. Yu et al used CGKRK-modified lipid nanoparticles (C-SMNPs) loaded with shikonin (SHK) and Mn2+ to attract tumor cells, induce necrosis, activate the cGAS-STING route, enhance type I interferon responses, promote dendritic cell maturation, and increase CD8+ T cell infiltration. C-SMNPs combination treatment demonstrated more lasting and significant tumor growth inhibition than typical immune checkpoint blockade (ICB) therapy by altering the immune milieu and increasing tumor immunogenicity, paving the way for new cancer treatment options. These pioneering investigations give critical insights for the development of novel metal ion-regulated anticancer treatments (Figure 10B).169,170
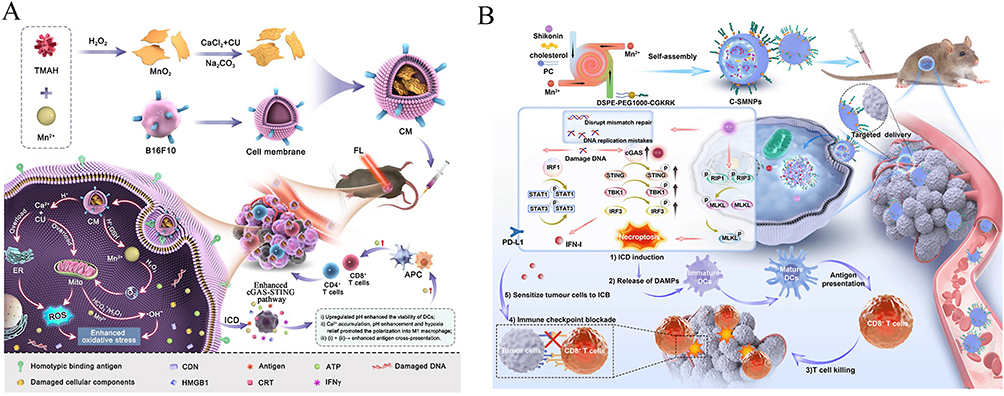 |
Figure 10 (A):Schematic diagram of the multifunctional calcium−manganese nanomodulator provides antitumor treatment and improved immunotherapy via reprogramming of the tumor microenvironment. Adapted from Ref 168 (B):Schematic illustration of nanomedicine C-SMNPs delivering SHK and manganese to restore antitumor immunity and sensitize tumor cells to ICB by inducing necroptotic ICD and boosting the cGAS-STING-mediated immune response. Adapted from Ref 170. |
Magnesium-Induced Cell Death
Magnesium ions play a central role in triggering distinct death pathways in cancer cells. Elevated intracellular Mg2+ levels directly disrupt mitochondrial function, leading to membrane depolarization, calcium dysregulation, and excessive reactive oxygen species production. This ionic imbalance activates various proteases and endonucleases, precipitating DNA fragmentation and irreversible metabolic collapse. Furthermore, Mg2+ potentiates apoptotic signaling by facilitating caspase activation and can also induce necrotic-like death under sustained stress conditions. Its critical involvement stems from the modulation of key ATP-dependent processes and the destabilization of cellular ion homeostasis, making magnesium a decisive factor in orchestrating cancer cell death.
Li et al proposed a novel therapeutic strategy using a biodegradable magnesium-based implant to combat bone tumors (Figure 11A).171 The implant, presumably composed of magnesium alloys, utilizes a multifunctional coating to enhance corrosion resistance and enable responsive ion release. Under the acidic TME, the implant degrades, releasing Mg2+ and elevating local pH. The sudden surge of Mg2+ disrupts mitochondrial function, induces calcium dysregulation, and generates excessive ROS, leading to metabolic collapse. Simultaneously, the alkaline environment triggers autophagic cell death through pH-dependent pathways. Notably, the released Mg2+ activate the cGAS-STING pathway by promoting cytosolic DNA sensing, which further amplifies autophagic flux and synergizes with alkaline metabolic stress to eradicate tumor cells. The innovations of this nanomaterial lie in its multimodal synergy through dual induction of autophagic death and metabolic collapse, pH-responsive biodegradation targeting tumor specificity, activation of innate immunity via STING-dependent signaling, and dual functionality as both a tumor-ablation agent and a bone-repair scaffold. This nanomaterial exhibited several advantages, including high biocompatibility, biodegradability into endogenous ions, and avoidance of conventional chemotherapy resistance. However, limitations involve potential premature corrosion in physiological environments, challenges in controlling precise ion release kinetics, and the need for further validation in large animal models to assess long-term safety and immune modulation efficacy. This approach represents a paradigm shift in implant-based oncology therapy by harnessing bioresponsive materials to manipulate metabolic and immune pathways.
 |
Figure 11 (A):Biodegradable Mg drives cGAS-STING-dependent tumor suppression via autophagy-metabolism-immuneaxis. Ref 171 (B) Schematic illustration of the construction of pH-sensitive MgNF@PEG/PMMVP by self-assembling amphiphilic PEG-b-PMMVP on the surface of MgNFs and the mechanism of synergistic tumor burst-like and H2 therapy. Ref 172. |
In addition, Liu et al developed pH-sensitive polymer-coated magnesium nanoflowers (MgNFs) with a diameter of ~100-250 nm for cancer therapy (Figure 11B).172 In the acidic TME, the polymer coating degrades, exposing MgNFs to water and triggering a rapid reaction that produces hydrogen gas (H2) bubbles. These bubbles induce dual mechanical and biochemical cell death. Firstly, they cause transient cavitation and mechanical rupture of lysosomes, leading to necrosis. Secondly, the released H2 disrupts mitochondrial function, reduces ATP production, and generates high oxidative stress, resulting in DNA damage and apoptosis. It’s the first synthesis of size- and shape-controlled Mg nanoparticles with NIR-II photoacoustic and bubble-enhanced ultrasound imaging capabilities for precise guidance, and a drug-free, synergistic therapeutic strategy combining mechanical disruption and H2-mediated biochemical effects. Advantages encompass excellent biodegradability, high biocompatibility, and minimal side effects. However, it also has potential premature degradation in physiological environments and challenges in controlling precise H2 release kinetics for optimal therapeutic outcomes. This approach offers a promising, non-invasive magnesium-based paradigm for cancer theranostics.
Summary and Prospects
The integration of metal ion homeostasis regulation with nanomedicine has ushered in a transformative era for cancer therapy. By leveraging the unique biological roles of iron, copper, and zinc ions in driving PCD pathways, researchers have unlocked novel strategies to selectively eradicate malignant cells while minimizing off-target toxicity. This review synthesizes the rapid advancements in understanding metal ion-induced cell death mechanisms and their synergy with nanotechnology, highlighting how nanocarriers enhance therapeutic precision, enable controlled drug release, and facilitate multimodal treatments. Despite these breakthroughs, significant challenges persist in translating these innovations into clinical practice. Here, we critically evaluate the current landscape and propose forward-looking directions to propel this field toward impactful clinical translation.
Current Achievements and Mechanistic Insights
Nanomedicine has revolutionized the delivery of metal ions and their modulators by addressing longstanding limitations of conventional therapies. Iron-based nanoparticles, copper-loaded frameworks, and zinc oxide nanostructures exemplify how nanotechnology improves bioavailability, targets the TME, and mitigates systemic toxicity. For example, pH-responsive nanocarriers selectively release iron ions in acidic TMEs, inducing ferroptosis through lipid peroxidation cascades. Similarly, copper-delivering nanoparticles exploit mitochondrial vulnerabilities in cancer cells, triggering cuproptosis via protein aggregation and oxidative stress. Moreover, zinc oxide nanoparticles disrupt redox balance and synergize with the apoptosis or ferroptosis pathways. These approaches are further amplified by combining metal ion therapies with immunotherapy, radiotherapy, or SDT, creating multimodal regimens that overcome drug resistance and enhance immune activation. A core achievement of this field is the establishment of “nanomedicine-metal ion-RCD” regulatory networks. Unlike traditional small-molecule modulators, nanomedicines enable spatiotemporal control over metal ion delivery and metabolism, directly addressing the key challenge of “specifically amplifying RCD in tumors without off-target toxicity”-this forms the central theme of this review.
A key advancement lies in the design of “smart” nanoplatforms capable of real-time adaptation to TME cues. For example, ROS-sensitive polymers codelivering copper and elesomol dynamically respond to intracellular oxidative stress, ensuring spatiotemporal control over cuproptosis induction. Similarly, Janus-structured nanoparticles integrate imaging and therapeutic functions, enabling simultaneous ferroptosis induction and immune microenvironment remodelling. These innovations underscore the potential of nanotechnology to bridge mechanistic discovery and clinical application.
Persistent Challenges
Despite progress, critical hurdles remain. First, preclinical challenges: Nanocarrier biocompatibility issues, such as Fe3O4 nanoparticles are easily cleared by the reticuloendothelial system, reducing tumor accumulation; off-target metal ion toxicity, such as excessive Mn2+ may induce neurotoxicity. Second, translational challenges: High production costs of complex nanoplatforms, such as C-SMNPs require multi-step modification; lack of patient-derived organoid (PDO) validation for Mg2+-based implants; insufficient long-term safety data for metal ion nanomedicines.
Future Directions
To address these challenges, future research should prioritize the following interdisciplinary strategies. Current studies focus on single-metal systems, yet natural biological processes often rely on metal crosstalk. For example, iron and copper synergistically amplify ROS production via Fenton and Fenton-like reactions. Designing nanoparticles that codeliver iron and copper ions, or their chelators, could exploit this synergy to induce “panmetaloptosis,” a hybrid cell death modality. Similarly, zinc ions, which modulate ferroptosis through GPX4 inhibition, could be integrated into dual-action nanocarriers to enhance lipid peroxidation. Such systems would require precise stoichiometric control and stimuli-responsive release mechanisms to avoid metal overload toxicity.
Next-generation nanoplatforms should incorporate biosensors to autonomously adjust metal ion release on the basis of real-time TME biomarkers. For example, CRISPR-engineered “logic-gated” nanoparticles can activate copper release only in cells with specific genetic signatures. Additionally, integrating wearable or implantable devices with nanotherapies could enable closed-loop monitoring of systemic metal ion levels, ensuring therapeutic efficacy while preventing toxicity. Machine learning algorithms can accelerate the discovery of optimal nanocarrier compositions by predicting structure‒activity relationships. Training models on datasets linking nanoparticle size, surface charge, and metal-loading capacity to tumor penetration depth or immune activation could yield novel architectures, such as fractal-shaped zinc oxide particles, for increased ROS generation. AI could also identify patient-specific metal ion vulnerabilities via multiomics data, enabling precision nanomedicine tailored to individual metabolic profiles.
Emerging evidence suggests that metal ions influence epigenetic regulation. Copper, for example, modulates histone acetylation, whereas zinc finger proteins are critical for DNA repair. NPs delivering metal ions alongside epigenetic modulators could reprogram cancer cells to become “primed” for metal-induced PCD. This approach might also reverse resistance by downregulating efflux pumps or upregulating death receptors. Traditional xenograft models poorly mimic the heterogeneity of the human TME. Patient-derived organoids (PDOs) or humanized murine models engineered with patient-specific immune cells could be used to better evaluate nanotherapy efficacy. Additionally, 3D bioprinted tumor models incorporating stromal and immune components would allow high-throughput screening of metal ion‒nanocarrier interactions under physiologically relevant conditions.
Nanoplatforms combining metal ion delivery with diagnostic imaging would enable real-time visualization of treatment response. For example, ferroptosis-inducing iron oxide nanoparticles could be used as contrast agents, allowing clinicians to monitor lipid peroxide accumulation by imaging. This theranostic approach would facilitate adaptive dosing regimens and early detection of resistance. Metal ion imbalances profoundly affect immune cell function. Zinc, for example, regulates T-cell activation, whereas copper modulates PD-L1 expression. NPs engineered to selectively deliver zinc ions to tumor-associated macrophages (TAMs) or copper chelators to regulatory T cells (Tregs) can reprogram immunosuppressive niches into immunostimulatory environments. Combining this with checkpoint inhibitors might amplify abscopal effects, turning “cold” tumors “hot”.
Concluding Remarks
The convergence of metal ion biology and nanomedicine holds unprecedented potential for redefining cancer treatment. By transcending traditional monotherapies and embracing complexity through multimetal synergies, AI-driven design, and immune-metabolic crosstalk, researchers can develop robust, adaptive platforms capable of overcoming cancer evolution. However, success hinges on addressing translational gaps, particularly in biocompatibility and personalized dosing. As we venture into this new frontier, interdisciplinary collaboration among chemists, biologists, and clinicians will be paramount to harness the full therapeutic spectrum of metal ions, ultimately delivering safer, smarter, and more effective nanomedicines to patients worldwide.
Data Sharing Statement
No datasets were generated or analysed during the current study.
Funding
This work was supported by the National Natural Science Foundation of China (82160337, 82200999), Shaanxi Peovincial Youth Science Foundation (2023-JC-QN-0233), and Scientific and technological project of Ankang University (2023AYQDZR12).
Disclosure
The authors declare that they have no conflicts of interest.
References
1. Alabi BR, Liu S, Stoyanova T. Current and emerging therapies for neuroendocrine prostate cancer. Pharmacol Ther. 2022;238:108255. doi:10.1016/j.pharmthera.2022.108255
2. Siegel RL, Kratzer TB, Giaquinto AN, et al. Cancer statistics, 2025. CA Cancer J Clin. 2025;75(1):10–45. doi:10.3322/caac.21871
3. Wang Y, Yan Q, Fan C, et al. Overview and countermeasures of cancer burden in China. Sci China Life Sci. 2023;66(11):2515–2526. doi:10.1007/s11427-022-2240-6
4. Sun L, Liu H, Ye Y, et al. Smart nanoparticles for cancer therapy. Signal Transduct Target Ther. 2023;8(1):418. doi:10.1038/s41392-023-01642-x
5. Yuan J, Ofengeim D. A guide to cell death pathways. Nat Rev Mol Cell Biol. 2024;25(5):379–395. doi:10.1038/s41580-023-00689-6
6. Lai Y, Gao FF, Ge RT, et al. Metal ions overloading and cell death. Cell Biol Toxicol. 2024;40(1):72. doi:10.1007/s10565-024-09910-4
7. Price EE, Boyd JM. Genetic regulation of metal ion homeostasis in staphylococcus aureus. Trends Microbiol. 2020;28(10):821–831. doi:10.1016/j.tim.2020.04.004
8. Hu H, Xu Q, Mo Z, et al. New anti-cancer explorations based on metal ions. J Nanobiotechnol. 2022;20(1):457. doi:10.1186/s12951-022-01661-w
9. Yan H, Talty R, Aladelokun O, et al. Ferroptosis in colorectal cancer: a future target? Br J Cancer. 2023;128(8):1439–1451. doi:10.1038/s41416-023-02149-6
10. Bhatia SN, Chen X, Dobrovolskaia MA, et al. Cancer nanomedicine. Nat Rev Cancer. 2022;22(10):550–556. doi:10.1038/s41568-022-00496-9
11. Wei G, Wang Y, Yang G, et al. Recent progress in nanomedicine for enhanced cancer chemotherapy. Theranostics. 2021;11(13):6370–6392. doi:10.7150/thno.57828
12. Zheng S, Li X, Guo S. Nanomedicine hitchhiking on bacteria for treating tumors. BMEMat. 2024;2(4).
13. Newton K, Strasser A, Kayagaki N, et al. Cell death. Cell. 2024;187(2):235–256. doi:10.1016/j.cell.2023.11.044
14. Kari S, Subramanian K, Altomonte IA, et al. Programmed cell death detection methods: a systematic review and a categorical comparison. Apoptosis. 2022;27(7–8):482–508. doi:10.1007/s10495-022-01735-y
15. Yee PP, Wei Y, Kim SY, et al. Neutrophil-induced ferroptosis promotes tumor necrosis in glioblastoma progression. Nat Commun. 2020;11(1):5424. doi:10.1038/s41467-020-19193-y
16. Ren K, Pei J, Guo Y, et al. Regulated necrosis pathways: a potential target for ischemic stroke. Burns Trauma. 2023;11:tkad016. doi:10.1093/burnst/tkad016
17. Hänggi K, Ruffell B. Cell death, therapeutics, and the immune response in cancer. Trends Cancer. 2023;9(5):381–396. doi:10.1016/j.trecan.2023.02.001
18. Balkwill F. Tumour necrosis factor and cancer. Nat Rev Cancer. 2009;9(5):361–371. doi:10.1038/nrc2628
19. Delre DP, Amgalan D, Linkermann A, et al. Fundamental mechanisms of regulated cell death and implications for heart disease. Physiol Rev. 2019;99(4):1765–1817. doi:10.1152/physrev.00022.2018
20. Zhang C, Liu N. Ferroptosis, necroptosis, and pyroptosis in the occurrence and development of ovarian cancer. Front Immunol. 2022;13:920059. doi:10.3389/fimmu.2022.920059
21. D’arcy MS. Cell death: a review of the major forms of apoptosis, necrosis and autophagy. Cell Biol Int. 2019;43(6):582–592. doi:10.1002/cbin.11137
22. Hadian K, Stockwell BR. The therapeutic potential of targeting regulated non-apoptotic cell death. Nat Rev Drug Discov. 2023;22(9):723–742. doi:10.1038/s41573-023-00749-8
23. Nguyen TT, Wei S, Nguyen TH, et al. Mitochondria-associated programmed cell death as a therapeutic target for age-related disease. Exp Mol Med. 2023;55(8):1595–1619. doi:10.1038/s12276-023-01046-5
24. Liu L, Zhou L, Wang LL, et al. Programmed cell death in asthma: apoptosis, autophagy, pyroptosis, ferroptosis, and necroptosis. J Inflamm Res. 2023;16:2727–2754. doi:10.2147/JIR.S417801
25. Debnath J, Gammoh N, Ryan KM. Autophagy and autophagy-related pathways in cancer. Nat Rev Mol Cell Biol. 2023;24(8):560–575. doi:10.1038/s41580-023-00585-z
26. Kesavardhana S, Malireddi RKS, Kanneganti TD. Caspases in cell death, inflammation, and pyroptosis. Annu Rev Immunol. 2020;38:567–595. doi:10.1146/annurev-immunol-073119-095439
27. Li Y, Du Y, Zhou Y, et al. Iron and copper: critical executioners of ferroptosis, cuproptosis and other forms of cell death. Cell Commun Signal. 2023;21(1):327. doi:10.1186/s12964-023-01267-1
28. Jan R, Chaudhry GE. Understanding apoptosis and apoptotic pathways targeted cancer therapeutics. Adv Pharm Bull. 2019;9(2):205–218. doi:10.15171/apb.2019.024
29. Lopez A, Reyna DE, Gitego N, et al. Co-targeting of BAX and BCL-XL proteins broadly overcomes resistance to apoptosis in cancer. Nat Commun. 2022;13(1):1199. doi:10.1038/s41467-022-28741-7
30. Liu X, Fan Z, Zhang L, et al. Bcl-2 inhibitor uploaded upconversion nanophotosensitizers to overcome the photodynamic therapy resistance of cancer through adjuvant intervention strategy. Biomaterials. 2017;144:73–83. doi:10.1016/j.biomaterials.2017.08.010
31. Škubník J, Svobodová Pavlíčková V, Ruml T, et al. Autophagy in cancer resistance to paclitaxel: development of combination strategies. Biomed Pharmacother. 2023;161:114458. doi:10.1016/j.biopha.2023.114458
32. Zhou L, Chen Y, Xie D, et al. Regulated cell death‐amplified sonodynamic anti‐tumor immune nanotherapeutics. BMEMat. 2024;2(4). doi:10.1002/bmm2.12079
33. Cao Z, Liu X, Zhang W, et al. Biomimetic macrophage membrane-camouflaged nanoparticles induce ferroptosis by promoting mitochondrial damage in glioblastoma. ACS Nano. 2023;17(23):23746–23760. doi:10.1021/acsnano.3c07555
34. Ferreira PMP, Sousa RWR, ferreira JRO, et al. Chloroquine and hydroxychloroquine in antitumor therapies based on autophagy-related mechanisms. Pharmacol Res. 2021;168:105582. doi:10.1016/j.phrs.2021.105582
35. Wei X, Xie F, Zhou X, et al. Role of pyroptosis in inflammation and cancer. Cell Mol Immunol. 2022;19(9):971–992. doi:10.1038/s41423-022-00905-x
36. Hsu SK, Li CY, Lin IL, et al. Inflammation-related pyroptosis, a novel programmed cell death pathway, and its crosstalk with immune therapy in cancer treatment. Theranostics. 2021;11(18):8813–8835. doi:10.7150/thno.62521
37. Zhang Y, Zhang C, Yang Y, et al. Pyroptosis-related gene signature predicts prognosis and indicates immune microenvironment infiltration in glioma. Front Cell Dev Biol. 2022;10:862493. doi:10.3389/fcell.2022.862493
38. Zhang G, Wang N, Ma Y, et al. Metal coordination‐driven assembly of stimulator of interferon genes‐activating nanoparticles for tumor chemo‐immunotherapy. BMEMat. 2024;2(2).
39. Luan M, Feng Z, Zhu W, et al. Mechanism of metal ion-induced cell death in gastrointestinal cancer. Biomed Pharmacother. 2024;174:116574. doi:10.1016/j.biopha.2024.116574
40. Lei G, Zhuang L, Gan B. Targeting ferroptosis as a vulnerability in cancer. Nat Rev Cancer. 2022;22(7):381–396. doi:10.1038/s41568-022-00459-0
41. Tang D, Kroemer G, Kang R. Targeting cuproplasia and cuproptosis in cancer. Nat Rev Clin Oncol. 2024;21(5):370–388. doi:10.1038/s41571-024-00876-0
42. Wu Q, Zhang H, Liu H, Dong Y. External physical field‐driven nanocatalytic cancer therapy. BME Mat. 2023;1(1).
43. Dixon SJ, Lemberg KM, Lamprecht MR, et al. Ferroptosis: an iron-dependent form of nonapoptotic cell death. Cell. 2012;149(5):1060–1072. doi:10.1016/j.cell.2012.03.042
44. Zhang C, Liu X, Jin S, et al. Ferroptosis in cancer therapy: a novel approach to reversing drug resistance. Mol Cancer. 2022;21(1):47. doi:10.1186/s12943-022-01530-y
45. Zhao L, Zhou X, Xie F, et al. Ferroptosis in cancer and cancer immunotherapy. Cancer Commun. 2022;42(2):88–116. doi:10.1002/cac2.12250
46. Sun S, Shen J, Jiang J, et al. Targeting ferroptosis opens new avenues for the development of novel therapeutics. Signal Transduct Target Ther. 2023;8(1):372. doi:10.1038/s41392-023-01606-1
47. Liang C, Zhang X, Yang M, et al. Recent progress in ferroptosis inducers for cancer therapy. Adv Mater. 2019;31(51):e1904197. doi:10.1002/adma.201904197
48. Mortensen MS, Ruiz J, Watts JL. Polyunsaturated fatty acids drive lipid peroxidation during ferroptosis. Cells. 2023;12(5):804. doi:10.3390/cells12050804
49. Rochette L, Dogon G, Rigal E, et al. Lipid peroxidation and iron metabolism: two corner stones in the homeostasis control of ferroptosis. Int J Mol Sci. 2022;24(1):449. doi:10.3390/ijms24010449
50. Pei Z, Qin Y, Fu X, et al. Inhibition of ferroptosis and iron accumulation alleviates pulmonary fibrosis in a bleomycin model. Redox Biol. 2022;57:102509. doi:10.1016/j.redox.2022.102509
51. Li J, Jia B, Cheng Y, et al. Targeting molecular mediators of ferroptosis and oxidative stress for neurological disorders. Oxid Med Cell Longev. 2022;2022:3999083. doi:10.1155/2022/3999083
52. Gao M, Yi J, Zhu J, et al. Role of mitochondria in ferroptosis. Mol Cell. 2019;73(2):354–63.e3. doi:10.1016/j.molcel.2018.10.042
53. Bock FJ, Tait SWG. Mitochondria as multifaceted regulators of cell death. Nat Rev Mol Cell Biol. 2020;21(2):85–100. doi:10.1038/s41580-019-0173-8
54. Xing P, Wang S, Cao Y, et al. Treatment strategies and drug resistance mechanisms in adenocarcinoma of different organs. Drug Resist Updat. 2023;71:101002. doi:10.1016/j.drup.2023.101002
55. Friedmann angeli JP, Krysko DV, Conrad M. Ferroptosis at the crossroads of cancer-acquired drug resistance and immune evasion. Nat Rev Cancer. 2019;19(7):405–414. doi:10.1038/s41568-019-0149-1
56. Wang Y, Wu X, Ren Z, et al. Overcoming cancer chemotherapy resistance by the induction of ferroptosis. Drug Resist Updat. 2023;66:100916. doi:10.1016/j.drup.2022.100916
57. Raycoquard I, Pautier P, Pignata S, et al. Olaparib plus bevacizumab as first-line maintenance in ovarian cancer. N Engl J Med. 2019;381(25):2416–2428. doi:10.1056/NEJMoa1911361
58. Chen P, Li X, Zhang R, et al. Combinative treatment of β-elemene and cetuximab is sensitive to KRAS mutant colorectal cancer cells by inducing ferroptosis and inhibiting epithelial-mesenchymal transformation. Theranostics. 2020;10(11):5107–5119. doi:10.7150/thno.44705
59. Zuo Y, Ying Y, Huang Z, et al. Reactive oxygen species targeted biomaterials for spinal cord injury therapy. Brain‐X. 2023;1(3). doi:10.1002/brx2.32
60. Sha X, Wang C, Liu Y, et al. Multifunctional glycyrrhizic acid-loaded nanoplatform combining ferroptosis induction and HMGB1 blockade for enhanced tumor immunotherapy. J Nanobiotechnol. 2025;23(1). doi:10.1186/s12951-025-03307-z
61. Yu Y, Shen X, Xiao X, et al. Butyrate modification promotes intestinal absorption and hepatic cancer cells targeting of ferroptosis inducer loaded nanoparticle for enhanced hepatocellular carcinoma therapy. Small. 2023;19(36):e2301149. doi:10.1002/smll.202301149
62. Liu H, Yu H, Li J. Biomedical materials benefit health. BMEMat. 2023;1(1):e12013.
63. You K, Wang Q, Osman MS, et al. Advanced strategies for combinational immunotherapy of cancer based on polymeric nanomedicines. BMEMat. 2024;2(2).
64. Chen Y, Shangguan S, Lin Z, et al. Nanomaterials for enhanced X‐ray‐triggered cancer therapy: progress and prospects. BMEMat. 2024.
65. Shen J, Zhou Y, Yin L. Nano/genetically engineered cells for immunotherapy. BMEMat. 2024;3(1).
66. Hsieh CH, Hsieh HC, Shih FS, et al. An innovative NRF2 nano-modulator induces lung cancer ferroptosis and elicits an immunostimulatory tumor microenvironment. Theranostics. 2021;11(14):7072–7091. doi:10.7150/thno.57803
67. Wang S, Guo Q, Xu R, et al. Combination of ferroptosis and pyroptosis dual induction by triptolide nano-MOFs for immunotherapy of Melanoma. J Nanobiotechnol. 2023;21(1):383. doi:10.1186/s12951-023-02146-0
68. Guo W, Wu Z, Chen J, et al. Nanoparticle delivery of miR-21-3p sensitizes melanoma to anti-PD-1 immunotherapy by promoting ferroptosis. J Immunother Cancer. 2022;10(6):e004381. doi:10.1136/jitc-2021-004381
69. Sandbhor P, Palkar P, Bhat S, et al. Nanomedicine as a multimodal therapeutic paradigm against cancer: on the way forward in advancing precision therapy. Nanoscale. 2024;16(13):6330–6364. doi:10.1039/D3NR06131K
70. Feng Z, Wang Z, Xiang X, et al. Progress in nanomedicine for sonodynamic immunotherapy of tumors. EngMedicine. 2024;1(2):100027. doi:10.1016/j.engmed.2024.100027
71. Wang H, Qiao C, Guan Q, et al. Nanoparticle-mediated synergistic anticancer effect of ferroptosis and photodynamic therapy: novel insights and perspectives. Asian J Pharm Sci. 2023;18(4):100829. doi:10.1016/j.ajps.2023.100829
72. Zhou H, Liu Z, Zhang Z, et al. Copper-cysteamine nanoparticle-mediated microwave dynamic therapy improves cancer treatment with induction of ferroptosis. Bioact Mater. 2023;24:322–330. doi:10.1016/j.bioactmat.2022.12.023
73. Wang N, Zhang Q, Wang Z, et al. A chemo/chemodynamic nanoparticle based on hyaluronic acid induces ferroptosis and apoptosis for triple-negative breast cancer therapy. Carbohydr Polym. 2024;329:121795. doi:10.1016/j.carbpol.2024.121795
74. Wu Y, Tian J, Yang J, et al. Bufotalin-loaded biomimetic prussian blue nanoparticles for colorectal cancer chemo-photothermal ferroptosis therapy. Nanomedicine. 2024;19(2):109–125. doi:10.2217/nnm-2023-0293
75. Sun X, Zhang S, Jiang S, et al. Combined positron emission tomography‐guided modified black phosphorus nanosheet‐based photothermal therapy and anti programmed cell death protein ligand 1 therapy. Radiology. 2024;2(2):103–112.
76. Tian Y, He X, Yuan Y, et al. TME-responsive nanoplatform with glutathione depletion for enhanced tumor-specific mild photothermal/gene/ferroptosis synergistic therapy. Int J Nanomed. 2024;19:9145–9160. doi:10.2147/IJN.S475698
77. Li J, Lu W, Yang Y, et al. Hybrid Nanomaterials for Cancer Immunotherapy. Adv Sci. 2023;10(6):e2204932. doi:10.1002/advs.202204932
78. Bunevicius A, Pikis S, Padilla F, et al. Sonodynamic therapy for gliomas. J Neurooncol. 2022;156(1):1–10. doi:10.1007/s11060-021-03807-6
79. Gao Z. Ultrasound‐enabled delivery of drugs to the brain: thinking outside the blood–brain barrier. Brain‐X. 2024;2(3). doi:10.1002/brx2.73
80. Xu M, Zhou L, Zheng L, et al. Sonodynamic therapy-derived multimodal synergistic cancer therapy. Cancer Lett. 2021;497:229–242. doi:10.1016/j.canlet.2020.10.037
81. Niu B, Liao K, Zhou Y, et al. Application of glutathione depletion in cancer therapy: enhanced ROS-based therapy, ferroptosis, and chemotherapy. Biomaterials. 2021;277:121110. doi:10.1016/j.biomaterials.2021.121110
82. Su W, Wang H, Pan J, et al. Advances in Sonodynamic Therapy: focus on Ferroptosis. J Med Chem. 2025;68:5976–5992. doi:10.1021/acs.jmedchem.4c02603
83. Zhao J, Bian E, Zhang R, et al. Self-assembled aza-boron-dipyrromethene-based h(2)s prodrug for synergistic ferroptosis-enabled gas and sonodynamic tumor therapies. Adv Sci. 2024;11(30):e2309542. doi:10.1002/advs.202309542
84. He M, Dan Y, Chen M, et al. Biocompatible polymer-modified nanoplatform for ferroptosis-enhanced combination cancer therapy. Macromol Biosci. 2023;23(11):e2300215. doi:10.1002/mabi.202300215
85. Yang H, Li X, Wang Q, et al. Multienzyme-like polyoxometalate for oxygen-independent sonocatalytic enhanced cancer therapy. J Colloid Interface Sci. 2025;681:319–330. doi:10.1016/j.jcis.2024.11.168
86. Wu H, Cheng K, Sun Z, et al. Trends of mapping knowledge structure and themes of cancer sonodynamic therapy: a text-mining study. Quant Imaging Med Surg. 2024;14(12):8734–8757. doi:10.21037/qims-24-128
87. Zhou L, Lyu J, Liu F, et al. Immunogenic panoptosis‐initiated cancer sono‐immune reediting nanotherapy by iteratively boosting cancer immunity cycle. Adv. Mater. 2023;36(2).
88. Yang Z, Qiao C, Jia Q, et al. Redox dyshomeostasis modulation of the tumor intracellular environment through a metabolic intervention strategy for enhanced photodynamic therapy. Theranostics. 2022;12(14):6143–6154. doi:10.7150/thno.75837
89. Yang Z, Ren X, Li L, et al. Trojan-horse inspired nanoblaster: x-ray triggered spot attack on radio-resistant cancer through radiodynamic therapy. Biomaterials. 2025;313:122814. doi:10.1016/j.biomaterials.2024.122814
90. Jiang F, Lee C, Zhang W, et al. Radiodynamic therapy with CsI(na)@MgO nanoparticles and 5-aminolevulinic acid. J Nanobiotechnol. 2022;20(1):330. doi:10.1186/s12951-022-01537-z
91. Wang Y, Zhang L, Zhou F. Cuproptosis: a new form of programmed cell death. Cell Mol Immunol. 2022;19(8):867–868. doi:10.1038/s41423-022-00866-1
92. Festa RA, Thiele DJ. Copper: an essential metal in biology. Curr Biol. 2011;21(21):R877–83. doi:10.1016/j.cub.2011.09.040
93. Tsvetkov P, Coy S, Petrova B, et al. Copper induces cell death by targeting lipoylated TCA cycle proteins. Science. 2022;375(6586):1254–1261. doi:10.1126/science.abf0529
94. Chen L, Min J, Wang F. Copper homeostasis and cuproptosis in health and disease. Signal Transduct Target Ther. 2022;7(1):378. doi:10.1038/s41392-022-01229-y
95. Yang L, Yang P, Lip GY, et al. Copper homeostasis and cuproptosis in cardiovascular disease therapeutics. Trends Pharmacol Sci. 2023;44(9):573–585. doi:10.1016/j.tips.2023.07.004
96. Xiong C, Ling H, Hao Q, et al. Cuproptosis: p53-regulated metabolic cell death? Cell Death Differ. 2023;30(4):876–884. doi:10.1038/s41418-023-01125-0
97. Tian Z, Jiang S, Zhou J, et al. Copper homeostasis and cuproptosis in mitochondria. Life Sci. 2023;334:122223. doi:10.1016/j.lfs.2023.122223
98. Chen X, Cai Q, Liang R, et al. Copper homeostasis and copper-induced cell death in the pathogenesis of cardiovascular disease and therapeutic strategies. Cell Death Dis. 2023;14(2):105. doi:10.1038/s41419-023-05639-w
99. Jiang Y, Huo Z, Qi X, et al. Copper-induced tumor cell death mechanisms and antitumor theragnostic applications of copper complexes. Nanomedicine. 2022;17(5):303–324. doi:10.2217/nnm-2021-0374
100. Cobine PA, Moore SA, Leary SC. Getting out what you put in: copper in mitochondria and its impacts on human disease. Biochim Biophys Acta Mol Cell Res. 2021;1868(1):118867. doi:10.1016/j.bbamcr.2020.118867
101. Feng Y, Yang Z, Wang J, et al. Cuproptosis: unveiling a new frontier in cancer biology and therapeutics. Cell Commun Signal. 2024;22(1):249. doi:10.1186/s12964-024-01625-7
102. Qu J, Wang Y, Wang Q. Cuproptosis: potential new direction in diabetes research and treatment. Front Endocrinol. 2024;15:1344729. doi:10.3389/fendo.2024.1344729
103. Tong X, Tang R, Xiao M, et al. Targeting cell death pathways for cancer therapy: recent developments in necroptosis, pyroptosis, ferroptosis, and cuproptosis research. J Hematol Oncol. 2022;15(1):174. doi:10.1186/s13045-022-01392-3
104. Liu X, Luo B, Wu X, et al. Cuproptosis and cuproptosis-related genes: emerging potential therapeutic targets in breast cancer. Biochim Biophys Acta Rev Cancer. 2023;1878(6):189013. doi:10.1016/j.bbcan.2023.189013
105. Qin Y, Liu Y, Xiang X, et al. Cuproptosis correlates with immunosuppressive tumor microenvironment based on pan-cancer multiomics and single-cell sequencing analysis. Mol Cancer. 2023;22(1):59. doi:10.1186/s12943-023-01752-8
106. Mao C, Wang M, Zhuang L, et al. Metabolic cell death in cancer: ferroptosis, cuproptosis, disulfidptosis, and beyond. Protein Cell. 2024;15(9):642–660. doi:10.1093/procel/pwae003
107. Wu H, Zhang Z, Cao Y, et al. A self‐amplifying ros‐responsive nanoplatform for simultaneous cuproptosis and cancer immunotherapy. Adv. Sci. 2024;11(23).
108. Guo B, Yang F, Zhang L, et al. Cuproptosis Induced by ROS responsive nanoparticles with elesclomol and copper combined with αpd-l1 for enhanced cancer immunotherapy. Adv Mater. 2023;35(22):e2212267. doi:10.1002/adma.202212267
109. Ning S, Lyu M, Zhu D, et al. Type-I AIE photosensitizer loaded biomimetic system boosting cuproptosis to inhibit breast cancer metastasis and rechallenge. ACS Nano. 2023;17(11):10206–10217. doi:10.1021/acsnano.3c00326
110. Wang N, Liu Y, Peng D, et al. Copper-based composites nanoparticles improve triple-negative breast cancer treatment with induction of apoptosis-cuproptosis and immune activation. Adv Healthc Mater. 2024;13(28):e2401646. doi:10.1002/adhm.202401646
111. Li Y, Liu J, Chen Y, et al. Nanoparticles synergize ferroptosis and cuproptosis to potentiate cancer immunotherapy. Adv Sci. 2024;11(23):e2310309. doi:10.1002/advs.202310309
112. Wang J, Li J, Liu J, et al. Interplay of ferroptosis and cuproptosis in cancer: dissecting metal-driven mechanisms for therapeutic potentials. Cancers. 2024;16(3).
113. Huang L, Zhu J, Wu G, et al. A strategy of “adding fuel to the flames” enables a self-accelerating cycle of ferroptosis-cuproptosis for potent antitumor therapy. Biomaterials. 2024;311:122701. doi:10.1016/j.biomaterials.2024.122701
114. Park MK, Choi BY, Kho AR, et al. The protective role of glutathione on zinc-induced neuron death after brain injuries. Int J Mol Sci. 2023;24(3).
115. Li Z, Liu Y, Wei R, et al. The important role of zinc in neurological diseases. Biomolecules. 2022;13(1):28. doi:10.3390/biom13010028
116. Chen B, Yu P, Chan WN, et al. Cellular zinc metabolism and zinc signaling: from biological functions to diseases and therapeutic targets. Signal Transduct Target Ther. 2024;9(1):6. doi:10.1038/s41392-023-01679-y
117. Costa MI, Sarmento-Ribeiro AB, Gonçalves AC. Zinc: from biological functions to therapeutic potential. Int J Mol Sci. 2023;24(5):4822. doi:10.3390/ijms24054822
118. Fan L, Cui Y, Liu Z, et al. Zinc and selenium status in coronavirus disease 2019. Biometals. 2023;36(5):929–941. doi:10.1007/s10534-023-00501-0
119. Yang Y, Wang P, Guo J, et al. Zinc overload induces damage to H9c2 cardiomyocyte through mitochondrial dysfunction and ROS-mediated mitophagy. Cardiovasc Toxicol. 2023;23(11–12):388–405. doi:10.1007/s12012-023-09811-8
120. Saravanan R, Balasubramanian V, Swaroop Balamurugan SS, et al. Zinc transporter LIV1: a promising cell surface target for triple negative breast cancer. J Cell Physiol. 2022;237(11):4132–4156. doi:10.1002/jcp.30880
121. Sun X, Xu S, Liu T, et al. Zinc supplementation alleviates oxidative stress to inhibit chronic gastritis via the ROS/NF-κB pathway in a mouse model. Food Funct. 2024;15(13):7136–7147. doi:10.1039/D4FO01142B
122. Sales CBS, Dias RB, De Faro Valverde L, et al. Hedgehog components are overexpressed in a series of liver cancer cases. Sci Rep. 2024;14(1):19507. doi:10.1038/s41598-024-70220-0
123. Wang W, Li Z, Lyu C, et al. Mechanism of a novel complex: zinc oxide nanoparticles-luteolin to promote ferroptosis in human acute myeloid leukemia cells in vitro. Int J Nanomed. 2025;20:4035–4050. doi:10.2147/IJN.S509007
124. Yan Y, Huang W, Lu X, et al. Zinc oxide nanoparticles induces cell death and consequently leading to incomplete neural tube closure through oxidative stress during embryogenesis. Cell Biol Toxicol. 2024;40(1):51. doi:10.1007/s10565-024-09894-1
125. Cai J, Wang X, Wang Z, et al. ZC3H13-mediated m6a modification ameliorates acute myocardial infarction through preventing inflammation, oxidative stress and ferroptosis by targeting lncRNA93358. Inflammation. 2024;48:1270–1284. doi:10.1007/s10753-024-02116-0
126. Cui M, Xu B, Wang L. Recent advances in multi‐metallic‐based nanozymes for enhanced catalytic cancer therapy. BMEMat. 2023;2(1). doi:10.1002/bmm2.12065
127. Hu Z, Song X, Ding L, et al. Engineering Fe/Mn-doped zinc oxide nanosonosensitizers for ultrasound-activated and multiple ferroptosis-augmented nanodynamic tumor suppression. Mater Today Bio. 2022;16.
128. Lin L-S, Wang J-F, Song J, et al. Cooperation of endogenous and exogenous reactive oxygen species induced by zinc peroxide nanoparticles to enhance oxidative stress-based cancer therapy. Theranostics. 2019;9(24):7200–7209. doi:10.7150/thno.39831
129. Mejia-Mendez JL, Reza-Zaldívar EE, Sanchez-Martinez A, et al. Exploring the cytotoxic and antioxidant properties of lanthanide-doped ZnO nanoparticles: a study with machine learning interpretation. J Nanobiotechnol. 2024;22(1). doi:10.1186/s12951-024-02957-9
130. Złowocka-Perłowska E, Baszuk P, Marciniak W, et al. Blood and serum se and zn levels and 10-year survival of patients after a diagnosis of kidney cancer. Biomedicines. 2024;12(8):1775. doi:10.3390/biomedicines12081775
131. Padmanabhan A, Kaushik M, Niranjan R, et al. Zinc Oxide nanoparticles induce oxidative and proteotoxic stress in ovarian cancer cells and trigger apoptosis Independent of p53-mutation status. Appl Surf Sci. 2019;487:807–818. doi:10.1016/j.apsusc.2019.05.099
132. Sadeghzadeh F, Golestani P, Beyramabdi P, et al. The anticancer impact of folate-linked ZnO-decorated bovine serum albumin/silibinin nanoparticles on human pancreatic, breast, lung, and colon cancers. J Biomater Sci Polym Ed. 2024;35(12):1845–1862. doi:10.1080/09205063.2024.2356967
133. Wiesmann N, Kluenker M, Demuth P, et al. Zinc overload mediated by zinc oxide nanoparticles as innovative anti-tumor agent. J Trace Elem Med Biol. 2019;51:226–234. doi:10.1016/j.jtemb.2018.08.002
134. Li P, Wang C, Qiu J, et al. Inhibitory effect of zinc oxide nanorod arrays on breast cancer cells profiled through real‐time cytokines screening by a single‐cell microfluidic platform. BMEMat. 2023;1(3).
135. Ghaemi B, Mashinchian O, Mousavi T, et al. Harnessing the cancer radiation therapy by lanthanide-doped zinc oxide based theranostic nanoparticles. ACS Appl. Mater. Interfaces. 2016;8(5):3123–3134. doi:10.1021/acsami.5b10056
136. Gielecińska A, Kciuk M, Kontek R. The impact of calcium overload on cellular processes: exploring calcicoptosis and its therapeutic potential in cancer. Int J Mol Sci. 2024;25(24):13727. doi:10.3390/ijms252413727
137. Dalal PJ, Muller WA, Sullivan DP. Endothelial cell calcium signaling during barrier function and inflammation. Am J Pathol. 2020;190(3):535–542. doi:10.1016/j.ajpath.2019.11.004
138. Pandrangi SL, Chittineedi P, Manthari RK, et al. Impact of oxytosis on the cross‐talk of mTORC with mitochondrial proteins in drug‐resistant cancer stem cells. J Cell Physiol. 2024;239(12). doi:10.1002/jcp.31421
139. Khandagale A, Holmlund T, Entesarian M, et al. Severe congenital neutropenia‐associated JAGN1 mutations unleash a calpain‐dependent cell death programme in myeloid cells. Br. J. Haematol. 2020;192(1):200–211. doi:10.1111/bjh.17137
140. Patron M, Sprenger H-G, Langer T. m-AAA proteases, mitochondrial calcium homeostasis and neurodegeneration. Cell Res. 2018;28(3):296–306. doi:10.1038/cr.2018.17
141. Britti E, Ros J, Esteras N, et al. Tau inhibits mitochondrial calcium efflux and makes neurons vulnerable to calcium-induced cell death. Cell Calcium. 2020;86.
142. Laurino S, Russi S, Sabato C, et al. The inhibition of SLC8A1 promotes Ca2+-dependent cell death in Gastric Cancer. Biomed. Pharmacother. 2025;182.
143. Casazza AA, Capraro M, Pedrazzi M, et al. Temperature-dependent olive pomace extraction for obtaining bioactive compounds preventing the death of murine cortical neurons. Int J Mol Sci. 2024;25(2):907. doi:10.3390/ijms25020907
144. Calvillo-Rodríguez KM, Mendoza-Reveles R, Gómez-Morales L, et al. PKHB1, a thrombospondin-1 peptide mimic, induces anti-tumor effect through immunogenic cell death induction in breast cancer cells. OncoImmunology. 2022;11(1). doi:10.1080/2162402X.2022.2054305
145. Liu J, Xie Y, Ma J, et al. New Ca2+ based anticancer nanomaterials trigger multiple cell death targeting Ca2+ homeostasis for cancer therapy. Chem. Biol. Interact 2024;393.
146. An J, Liu M, Zhao L, et al. Boosting tumor immunotherapy by bioactive nanoparticles via ca2+ interference mediated tme reprogramming and specific PD‐L1 depletion. Adv. Funct. Mater. 2022;32(24). doi:10.1002/adfm.202201275
147. Guan Q, Zhou LL, Lv FH, et al. A glycosylated covalent organic framework equipped with BODIPY and CaCO 3 for synergistic tumor therapy. Angew. Chem. Int. Ed. 2020;59(41):18042–18047. doi:10.1002/anie.202008055
148. Qi C, Musetti S, Fu L-H, et al. Biomolecule-assisted green synthesis of nanostructured calcium phosphates and their biomedical applications. Chem. Soc. Rev. 2019;48(10):2698–2737. doi:10.1039/C8CS00489G
149. Fu LH, Wan Y, Li C, et al. Biodegradable calcium phosphate nanotheranostics with tumor‐specific activatable cascade catalytic reactions‐augmented photodynamic therapy. Adv. Funct. Mater. 2021;31(14). doi:10.1002/adfm.202009848
150. Jiang Y, Pan X, M YAO, et al. Bioinspired adhesive and tumor microenvironment responsive nanoMOFs assembled 3D-printed scaffold for anti-tumor therapy and bone regeneration. Nano Today. 2021;39.
151. Effects of water-sodium balance and regulation of electrolytes associated with antidiabetic drugs. Eur Rev Med Pharmacol Sci. 2023;27(12):5784–5794. doi:10.26355/eurrev_202306_32817
152. Costa TGF, Oliveira MM, Toledo MM, et al. Effect of Fe3+ on Na,K-ATPase: unexpected activation of ATP hydrolysis. Biochimica et Biophysica Acta (BBA) - Biomembranes. 2022;1864(4):183868. doi:10.1016/j.bbamem.2022.183868
153. Donck LV, Borgers M, Verdonck F. Inhibition of sodium and calcium overload pathology in the myocardium: a new cytoprotective principle. Cardiovasc Res. 1993;27(3):349–357. doi:10.1093/cvr/27.3.349
154. Ulshöfer R, Bros H, Hauser AE, et al. Preventing axonal sodium overload or mitochondrial calcium uptake protects axonal mitochondria from oxidative stress‐induced alterations. Oxid Med Cell Longev. 2022;2022(1). doi:10.1155/2022/6125711
155. Fu W, Wang J, Li T, et al. Persistent activation of TRPM4 triggers necrotic cell death characterized by sodium overload. Nat Chem Bio. 2025;21(8):1238–1249.
156. Luo X, Baba A, Matsuda T, et al. Susceptibilities to and mechanisms of excitotoxic cell death of adult mouse inner retinal neurons in dissociated culture. Investigat Opthalmol Vis Sci. 2004;45(12).
157. Jiang J, Hu S, Hu K, et al. Novel impact of metal ion-induced cell death on diabetic cardiomyopathy pathogenesis and therapy. Apoptosis. 2025;30(5–6):1152–1181. doi:10.1007/s10495-025-02090-4
158. Manganese: its Role in Disease and Health. Met Ions Life Sci. 2019;19.
159. Roth JA, Horbinski C, Higgins D, Lein P, Garrick MD. Mechanisms of manganese-induced rat pheochromocytoma (PC12) cell death and cell differentiation. Neurotoxicology. 2002;23(2):147–157. doi:10.1016/S0161-813X(01)00077-8
160. Banerjee K, Das S, Majumder S, et al. Modulation of cell death in human colorectal and breast cancer cells through a manganese chelate by involving GSH with intracellular p53 status. Mol Cell Biochem. 2016;427(1–2):35–58. doi:10.1007/s11010-016-2896-6
161. Porte Alcon S, Gorojod RM, Kotler ML. Regulated necrosis orchestrates microglial cell death in manganese-induced toxicity. Neuroscience. 2018;393:206–225. doi:10.1016/j.neuroscience.2018.10.006
162. Zhang S, Kang L, Dai X, et al. Manganese induces tumor cell ferroptosis through type-I IFN dependent inhibition of mitochondrial dihydroorotate dehydrogenase. Free Radic Biol Med. 2022;193:202–212. doi:10.1016/j.freeradbiomed.2022.10.004
163. Li Z, Chu Z, Yang J, et al. Immunogenic cell death augmented by manganese zinc sulfide nanoparticles for metastatic melanoma immunotherapy. ACS Nano. 2022;16(9):15471–15483. doi:10.1021/acsnano.2c08013
164. Dehghani F, Xu B, Liu W, et al. Inhibition of calpain prevents manganese-induced cell injury and alpha-synuclein oligomerization in organotypic brain slice cultures. PLoS One. 2015;10(3).
165. Peng P, Cao J, Cheng W, et al. Manganese dioxide-based in situ vaccine boosts antitumor immunity via simultaneous activation of immunogenic cell death and the STING pathway. Acta Biomater. 2025;194:467–482. doi:10.1016/j.actbio.2025.01.029
166. Han X, Leng C, Zhao S, et al. Development and verification of a manganese metabolism- and immune-related genes signature for prediction of prognosis and immune landscape in gastric cancer. Front Immunol. 2024;15.
167. Vechalapu SK, Kumar R, Sachan SK, et al. Copper and manganese complexes of pyridinecarboxaldimine induce oxidative cell death in cancer cells. ACS Appl. Bio Mater. 2024;7(10):6696–6705. doi:10.1021/acsabm.4c00854
168. Luo G, Li X, Lin J, et al. Multifunctional calcium–manganese nanomodulator provides antitumor treatment and improved immunotherapy via reprogramming of the tumor microenvironment. ACS Nano. 2023;17(16):15449–15465. doi:10.1021/acsnano.3c01215
169. Ma L, Wang X, Wu Y, et al. Controlled release of manganese and magnesium ions by microsphere-encapsulated hydrogel enhances cancer immunotherapy. J Control Release. 2024;372:682–698. doi:10.1016/j.jconrel.2024.06.067
170. Yu S, Li J, Zhang J, et al. Nanosized shikonin disrupts tumor-cell mismatch repair and synergizes with manganese to sensitize squamous carcinoma to immunotherapy. ACS Nano. 2025;19(14):13889–13905. doi:10.1021/acsnano.4c17090
171. Li M, Wei T, Liu H, et al. Revolutionizing bone tumor therapy: biodegradable magnesium implant triggers cGAS‐STING‐dependent synergy of autophagic cell death and alkaline metabolic collapse. Adv. Funct. Mater. 2025.
172. Liu L, Wu Y, Ye J, et al. Synthesis of magnesium nanoparticle for NIR-II-photoacoustic-imaging-guided synergistic burst-like and H2 cancer therapy. Chem. 2022;8(11):2990–3007. doi:10.1016/j.chempr.2022.07.001
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.