Back to Journals » Clinical Ophthalmology » Volume 13
Steady-State Pattern Electroretinogram and Frequency Doubling Technology in Adult Dyslexic Readers
Authors Schiavi C ![]() , Finzi A, Cellini M
, Finzi A, Cellini M
Received 14 September 2019
Accepted for publication 27 November 2019
Published 11 December 2019 Volume 2019:13 Pages 2451—2459
DOI https://doi.org/10.2147/OPTH.S229898
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Scott Fraser
Costantino Schiavi, Alessandro Finzi, Mauro Cellini
Department of Experimental, Diagnostic, and Specialty Medicine, Ophthalmology Service, University of Bologna, Bologna 40138, Italy
Correspondence: Mauro Cellini
Policlinico Sant’Orsola-Malpighi, Ophthalmology Service, University of Bologna, Via Pelagio Palagi 9, Bologna 40138, Italy
Tel/Fax +39 0516362835
Email [email protected]
Purpose: Dyslexia is a reading disorder with neurological deficit of the magnocellular pathway. The aim of our study was to evaluate the functionality of the magnocellular–Y (M–Y) retinal ganglion cells in adult dyslexic subjects using steady-state pattern electroretinogram and frequency doubling perimetry.
Methods: Ten patients with dyslexia (7 females and 3 males), mean age 28.7 ± 5.9 years, and 10 subjects without dyslexia (6 females and 4 males), mean age 27.8 ± 4.1 years, were enrolled in the study and underwent both steady-state pattern-electroretinogram examination and frequency doubling perimetry.
Results: There was a significant difference in the amplitude of the steady-state pattern electroretinogram of the dyslexic group and the healthy controls (0.610±0.110 μV vs 1.250±0.296 μV; p=0.0001). Furthermore, in the dyslexic group we found a significant difference between the right eye and the left eye (0.671±0.11 μV vs 0.559±0.15 μV; p=0.001). With frequency doubling perimetry, the pattern standard deviation index increased in dyslexic eyes compared to healthy controls (4.40±0.81 dB vs 2.99±0.35 dB; p=0.0001) and in the left eye versus the right eye of the dyslexic group (4.43±1.10 dB vs 3.66±0.96 dB; p=0.031). There was a correlation between the reduction in the wave amplitude of the pattern electroretinogram and the simultaneous increase in the pattern standard deviation values (r=0.80; p=0.001). This correlation was also found to be present in the left eye (r=0.93; p<0.001) and the right eye (r=0.81; p=0.005) of dyslexic subjects.
Conclusion: Our study shows that there was an alteration of the activity of M–Y retinal ganglion cells, especially in the left eye. It confirms that in dyslexia there is a deficit of visual attention with damage not only of the magnocellular-dorsal pathway but also of the M-Y retinal ganglion cells.
Keywords: steady-state pattern-electroretinogram, frequency doubling technology perimetry, retino-geniculate pathways, dyslexia
Introduction
Dyslexia is a condition characterized by impairment of reading skills in subjects without alterations of visual acuity and with normal intelligence. It affects boys and girls equally and it is usually first observed during childhood. The etiology of dyslexia is still under discussion, although an alteration of the magnocellular pathway, in particular the magnocellular-dorsal (M–D) pathway, is hypothesized.1–8 Previous autopsy studies have shown that in dyslexics the cells of the magnocellular pathway in the ventral layers of the lateral geniculate nucleus (LGN) were smaller than those of healthy controls.4
The human LGN contains three distinct retinal pathways:9 the parvocellular (P) pathway originating from the midget retinal ganglion cells,10–12 the koniocellular pathway receiving afferents from the retinal ganglion cells driven by short-wavelength photoreceptors,13–15 and the magnocellular (M) pathway, derived from the parasol retinal ganglion cells (RGCs),10,12,16,17 which contains two functional cell classes that are similar to cat X and cat Y geniculate cells. About 5% of the magnocellular cells of LGN display a nonlinear Y-type response,18 hence the term M–Y cells.
Selective electrofunctional investigation of the M pathway requires the use of electrophysiological methods based on the recording of pattern visual evoked potentials (PVEP) and appropriately modified stimuli with regard to contrast, spatial frequency, temporal frequency, and movement.19–24
PVEP studies appear to confirm the results of functional magnetic resonance imaging (fMRI).25–28 The M pathway and especially the M–Y ganglion cell involved in the analysis of visual motion can also be studied using frequency doubling illusion (FDI). FDI is based on a doubling illusion created by counterphase flickering of a low spatial frequency sinusoidal grating at a high temporal frequency.29
This type of psychophysical examination has been proposed as a sensitive test for detecting early functional changes in M–Y ganglion cells, mainly in glaucoma and ocular hypertension,18,30,31 but it has rarely been used in amblyopia32 and dyslexia.33,34 From an electrophysiological point of view, the study of M–Y cells in the M pathway can be performed using the steady-state pattern electroretinogram (SS-PERG).35 This examination was initially proposed in patients suffering from simple chronic open-angle glaucoma and ocular hypertension since it was able to detect early alterations in the bioelectric response of retinal M–Y retinal ganglion cells. The purpose of our study was to use a SS-PERG with a stimulus that creates a doubling illusion similar to that achieved using frequency doubling technology (FDT), in order to selectively investigate the activity of the M–Y retinal ganglion cells in adult dyslexic subjects.
Materials and Methods
Ten patients with dyslexia (7 women and 3 men), mean age 28.7 ± 5.9 years, and 10 patients without dyslexia (6 women and 4 men), mean age 27.8 ± 4.1 years, were enrolled in the study. All patients underwent a complete ophthalmological evaluation, including measurement of visual acuity, slit-lamp examination of the anterior and posterior segment, and random-dot stereopsis test. Ophthalmologic evaluation excluded a refractive defect higher than ± 2 diopters of spherical equivalent (SE). All subjects had normal binocular vision with random-dot stereopsis, absence of retinal and optic nerve diseases, and transparent dioptric media.
Patients enrolled in the study were diagnosed with dyslexia by the Neuropsychiatric Center of the National Health Service in Bologna according to the diagnostic criteria for learning disabilities and with a test for reading abilities.36,37
All patients also underwent SS-PERG and FDT perimetry examination.
The study was approved by the Local Ethics Committee of the University of Bologna and adhered to tenets of the Declaration of Helsinki. Written informed consent was obtained from all participants.
The Steady-State Pattern-Electroretinogram was recorded using the RetimaxPlus system (CSO Instruments, Florence, Italy). The patient sat on a chair at a distance of 57 cm from the television screen (resolution 1024x768; size 34 inches) and fixed binocularly on a red cross at the center of the screen, which subtended a visual angle of 48.89 degrees. The generated potential was measured with HK-LOOP ocular electrodes; the reference electrode was located near the outer canthus and the ground electrode was placed on the ear lobe. The interelectrode resistance was less than 5 kOhm. All subjects had undilated pupils, measuring between 3 and 4 mm, with an appropriate correction for the working distance, and they were allowed to blink freely.
PERG stimulus was first presented as a full-screen black-and-white vertical bar pattern (contrast: 20%; spatial frequency: 0.3 cycles per degree/cpd; temporal frequency: 15 Hz). The number of samples acquired, mediated and processed with Discrete Fourier Transform (DFT) was 300 and the acquisition time was 133 ms.
The pattern presentation (approximately 4 mins) was preceded by an unmodulated uniform field (approximately 1 min) of the same mean luminance (blank), which was used to evaluate the background noise level. The noise level was 0.08 ± 0.03 μV in both normal and dyslexic patients.
Because SS-PERG was recorded in response to relatively fast alternating gratings, the response waveforms were sinusoidal-like with a frequency corresponding to the reversal rate. Packets were automatically evaluated in the frequency domain by DFT to isolate the component at the reversal rate (30 Hz), and the amplitude in microvolts was displayed as a function of time.
The FDT perimetry was performed using the full-threshold program N-30 of the Humphrey FDT perimeter (Carl Zeiss Meditec, Dublin, CA) which tested 19 different points within the central 30 degrees of the visual field. Each target was displayed as a square of 10 x 10 degrees where a grid of black and white bars was projected. Furthermore, the 0.25 cycles/degree sinusoidal grid undergone counterphase flicker at 18 Hz to create the illusion of doubling (FDI).18 The dyslexic and the normal reader subjects underwent three different sessions of visual field tests at intervals of 4 ± 1 days to become familiar with the procedure because none of the study subjects had previous experience with FDT.
Mean defect (MD) and pattern standard deviation (PSD) were evaluated and were considered for the statistical analysis.
Statistical Analysis
For the statistical analysis of the data, we used the MedCalc 10.9.1 statistical program (MedCalc Software, Ostend, Belgium). MD and PSD of FDT and the amplitude of SS-PERG were analyzed using the Mann–Whitney U-test to assess group differences, Wilcoxon’s test to evaluate within-subject comparisons, and the Spearman's correlation test, considering p< 0.05 as significant.
Results
The mean visual acuity (BCVA) and the spherical equivalent (SE) in the control and dyslexic groups are reported in Table 1. There was no significant difference in BCVA (p = 0.980) and SE (p = 0.312) of two groups.
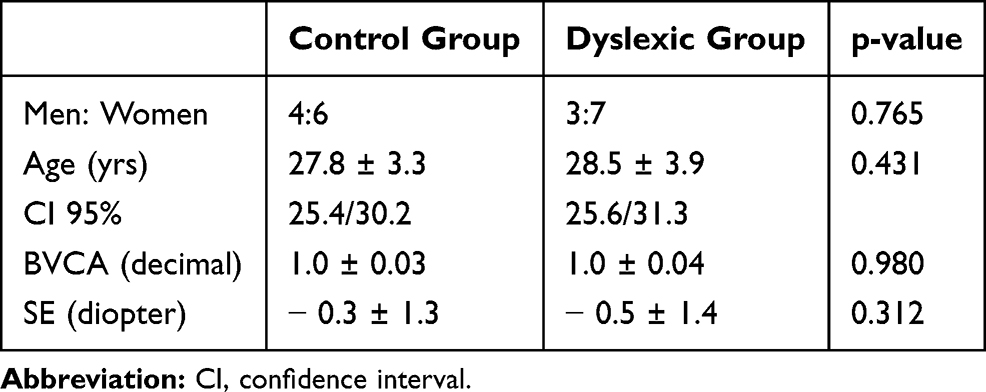 |
Table 1 Best Corrected Visual Acuity (BCVA) Values (Mean Values and Standard Deviation) and Diopter Spherical Equivalent (SE) Values (Mean Values and Standard Deviation) |
In regard to the amplitude of the SS-PERG waveform, there was a significant difference between the control and dyslexic group (p = 0.0001) (Table 2). Furthermore, in the dyslexic group we found a significant difference between the right eye (RE) and the left eye (LE) (p = 0.001) but no difference was found in the control group (p<0.596) (Table 3; Figure 1).
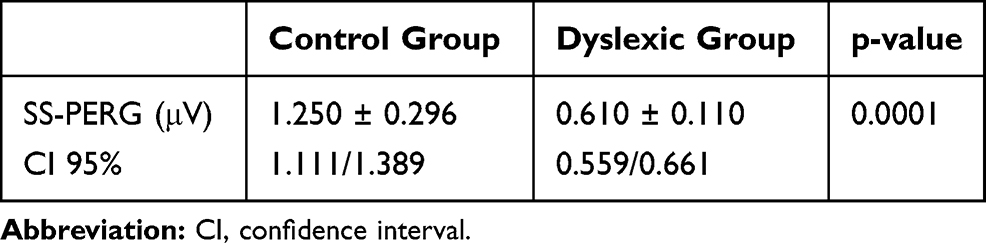 |
Table 2 Steady-State Pattern Electroretinogram (SS-PERG) Amplitude Values (Mean Values and Standard Deviation) |
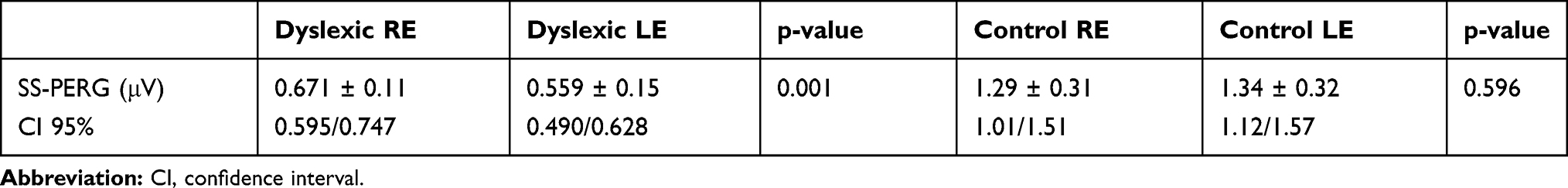 |
Table 3 Steady-State Pattern Electroretinogram (SS-PERG) Amplitude Values of the Right Eye (RE) and Left eye (LE) in Dyslexics and Control Subjects |
 |
Figure 1 Steady-state pattern electroretinogram waveforms recorded in the right and left eye of control normal subject (top) and dyslexic subject (bottom). It can be observed a reduction of waveform amplitude in both eyes of the dyslexic subject (bottom) against the normal subject (top). Moreover, the dyslexics have a waveform amplitude decrease in the LE compared to the RE while in the normal subjects we have a slight increase in waveform amplitude in the LE than in the RE. Abbreviations: RE, right eye; LE, left eye. |
With regard to FDT parameters, the MD was similar in the healthy group and the dyslexic group (p = 0.056). The PSD was significantly higher in dyslexic subjects compared to normal subjects (p = 0.0001) (Table 4). Furthermore, in the dyslexic group, we found that the PSD of LE was significantly higher compared to RE (p = 0.031) (Table 5). For PSD, in the control group, there was no statistically significant difference between RE and LE (p <0.791) (Table 5).
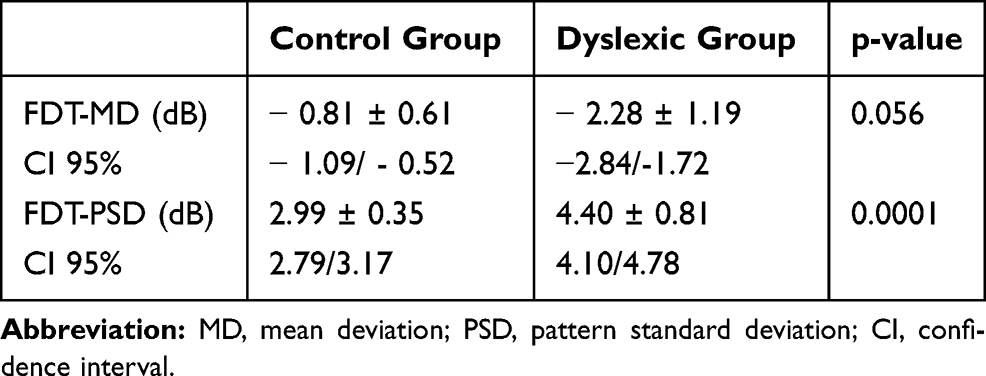 |
Table 4 Frequency Doubling Technology (FDT) Perimetry Values (Mean Values and Standard Deviation) |
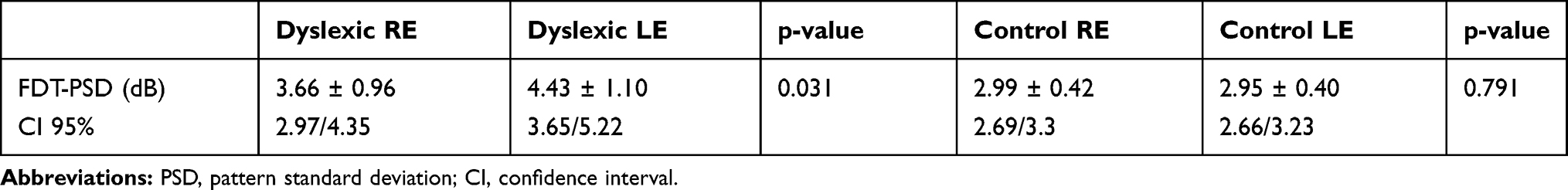 |
Table 5 Frequency Doubling Technology (FDT) Index Values of the Right Eye (RE) and Left eye (LE) in Dyslexics and Control Subjects |
Statistical analysis of the SS-PERG among all groups shows a statistically significant reduction of the wave amplitude in dyslexics compared to the control group, both for the RE (p = 0.0003) and for the LE (p = 0.0001). Also, when analyzing the FDT data we found an increase in PSD values in dyslexic subjects compared to normal subjects both in the RE (p <0.0002) and in the LE (p <0.0002) (Table 6).
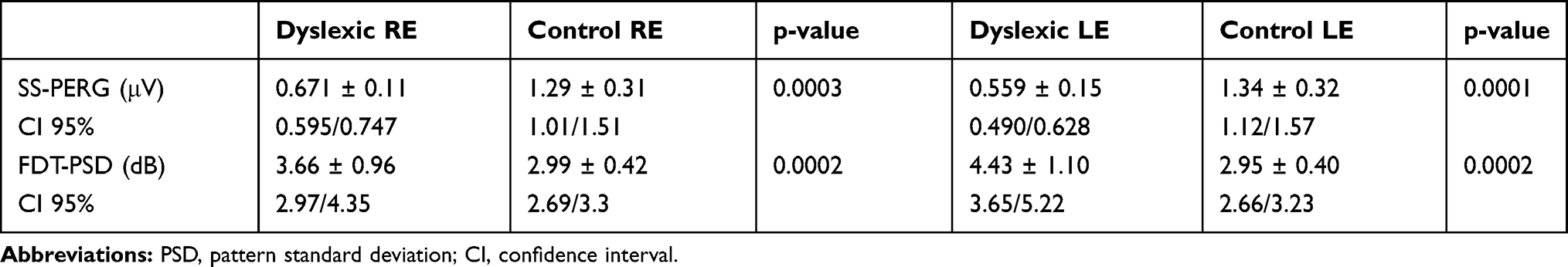 |
Table 6 Intragroup Steady-State Pattern Electroretinogram (SS-PERG) and Frequency Doubling Technology (FDT) Perimetry Index Values of the Right Eye (RE) and Left eye (LE) in Dyslexic Subjects and Control Subjects |
In dyslexic subjects, Spearman correlation test showed a significant correlation between the reduction in the SS-PERG wave amplitude and the simultaneous increase in PSD index values (r = 0.80; p = 0.001) (Table 7, Figure 2). Furthermore, this correlation was also found to be present in the LE (r = 0.93; p < 0.001) and the RE (r = 0.81; p = 0.005) of these subjects (Table 7, Figures 3–4). The same significant correlation between SS-PERG wave amplitude and PSD index values was found not only in the case of dyslexic subjects but also in the control group (Table 7)
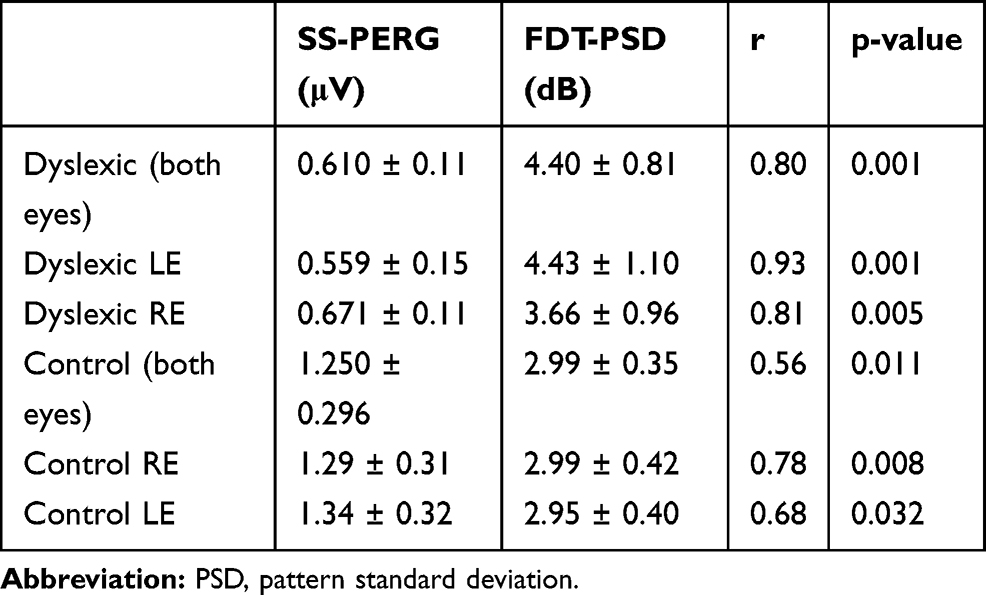 |
Table 7 Spearman’s Correlation Test Between Steady-State Pattern Electroretinogram (SS-PERG) and Frequency Doubling Technology (FDT) Index Values in the Right Eye (RE) and in the Left Eye (LE) of Dyslexic Subjects and of Control Subjects |
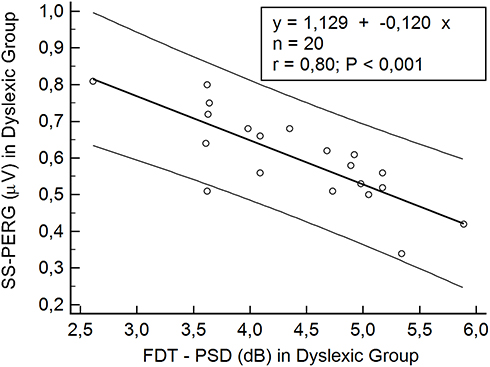 |
Figure 2 Scatterplot of Spearman’s correlation test between steady-state pattern electroretinogram (SS-PERG) and frequency doubling technology (FDT) in the dyslexic group. Abbreviation: PSD, pattern standard deviation. |
 |
Figure 3 Scatterplot of Spearman’s correlation test between steady-state pattern electroretinogram (SS-PERG) and frequency doubling technology (FDT) in the left eye of the dyslexic group. Abbreviation: PSD, pattern standard deviation. |
 |
Figure 4 Scatterplot of Spearman’s correlation test between steady-state pattern electroretinogram (SS-PERG) and frequency doubling technology (FDT) in the right eye of dyslexic group. Abbreviation: PSD, pattern standard deviation. |
Discussion
Dyslexia is a reading disorder afflicting 5–17% of the school-age population38–40 and characterized by difficulty in accessing and manipulating the phonemic units of written language.41,42 In recent decades, the most established hypothesis to explain developmental dyslexia was based on the presence of an auditory-phonological processing deficit;43–49 however, recent studies would seem to show how the absence of development of fluent reading could be attributed both to a deficit in visual attention50–55 and an oculomotor deficit.56–58
Studies of the postmortem brains of known dyslexic subjects have shown several alterations: the normal asymmetry of the planum temporale, favoring the left side, tends to be absent. Furthermore, in the posterior parietal cortex, we have an anomalous symmetry with small aberrant “brain warts” clustered around the temporo-parietal junction.59,60
Neuroradiological studies using fMRI have shown that visual attention and the ability to control eye movements and identify the position of objects in space are associated with the dorsal cortical visual pathway or occipito-parietal pathway, which appears to be altered in dyslexics.27,28,61 Furthermore, there is evidence of alterations in the functionality of the ventral cortical or occipito-temporal visual pathways, which receive information from both the M and P pathways, therefore, specializing in identifying the details relating to the shape and color of objects.61–64 Moreover, in dyslexic subjects, a disconnection between the dorsal visual pathway and the ventral visual pathway in the middle frontal left gyrus has been found.65
One of the main hypotheses to explain visual deficits in dyslexia is based on the presence of a deficit in the transmission of visual stimuli along the M pathway.4,5
To validate the deficit theory of the M pathway and to confirm what was found with the fRMN, several authors used both psychophysical66–68 and electrophysiological methods.23,69,70
The use of psychophysical methods,66–68 above all FDT,18,29–31,33 which allows selective stimulation of M-Y ganglion cells,18,30,71,72 has provided definite information on the M pathway.
By using FDT perimetry in the dyslexic group, we found a significant increase in the PSD index values compared with healthy controls, confirming what was found in previous studies.33,34 In the same group of patients, we found an insignificant increase in the MD index value. These data do not agree with other studies in which the difference between dyslexic and healthy subjects was statistically significant.33,34 We believe that this discrepancy could be due to the fact that the subjects enrolled in our study were young adults and not children. We cannot forget that dyslexia over time can improve when new reading strategies are learned.73
In our investigation, we analyzed not only the MD index but also the PSD index because this index reflects the roughness (focal-cluster alteration) of the visual field.74,75 Moreover, we found that in LE of dyslexic patients the PSD values were significantly more altered than in the RE, confirming the observations of a previous study.33,34 These data confirm that dyslexics present an alteration in retinal sensitivity in the LE. This event causes an alteration in the flow of the M pathway and, consequently, a slight neuronal disorder in the right temporal-parietal area, which is essential for the development of visual attention.76,77
Regarding electrophysiological investigations, mainly visual evoked potentials (VEPs) were used to study the M pathway by applying standard methods, which gave contradictory results.23 Recently, using the pattern visual evoked potentials (PVEP) and modifying the stimulation parameters appropriately in order to isolate the response of the MD pathway, it has been observed that for 60-arc-minute stimuli in dyslexic subjects the reduction of contrast from 100% to 25% resulted in a significant reduction in amplitude with an increase in P100 wave latency while for 15-arc-minute stimuli only latency was significantly increased.23,78 These data confirmed the observations of Romani and coworkers, who used stimuli with a high temporal frequency of 8 Hz with fixed contrast at 50% and spatial frequencies of the stimulus of 0.50 cpd (large stimulus) and 2 cpd (small stimulus). In this case, only low spatial frequency stimuli (large stimuli) and high temporal frequency determined a decrease in amplitude and an increase in latency of the N95 wave in dyslexic subjects.19
Other studies have used different electrophysiological techniques, such as fixed spatial frequencies with high temporal frequencies and luminance variation,21,23 low spatial frequency and high temporal frequency stimuli (7.5 Hz),20,23 and, finally, recording of VEPs with the use of moving stimuli at low contrast,79 as well as at low contrast and radial motion full field and radial motion periphery.69 In all cases, they showed a significant increase in latencies and a reduction in the amplitudes of the waveform in dyslexic patients.24
To study the activity of only M–Y retinal ganglion cells we used the SS-PERG with a stimulus similar to that used in the FDT perimetry.32 Both transient-PERG and SS-PERG are commonly used to investigate RGCs activity, but these two examinations differ mainly in the temporal frequency of the pattern stimulus. In detail, by using a temporal frequency of 4 Hz, a transient response will be obtained; by increasing the frequency to 8 Hz, a steady-state response will be recorded80,81 and we can study the ON pathway of the spiking retinal ganglion cells (RGCs).82
Now, the question is whether the Y-cells, first identified in cats, even exist in the primate’s retina. Indeed, this is a controversial topic in the literature, but recent studies have identified Y-like RGCs in primates.71,83–85 Experimental studies, with the use of microelectrodes, have shown that when the retina is stimulated with gratings at high contrast and high spatial frequency from each parasol (M) RGCs a doubled frequency is recorded and this response is characteristic of Y-cells.83
In our study, we found a significant reduction in the amplitude of SS-PERG in the dyslexic group compared to the healthy controls. In dyslexic subjects, we found a significant difference between the LE and RE, with a greater amplitude reduction in the LE. A significant correlation between the amplitude of the SS-PERG and the pattern standard deviation index of FDT perimetry. The correlation between the amplitude of the SS-PERG waveform and the FDT perimetric index confirms what was found by Maddess and coworkers in glaucoma and suspect glaucoma.35
Our electrophysiological data could therefore confirm not only the results of a previous study using the FDT technique33,34 but also the hypothesis that in dyslexic subjects the damage would be located not only in M–D pathway but even in the RGCs.
The small number of adult dyslexic subjects included in the study is a limitation of our research. In this first study, we only enrolled adult patients without any intellectual deficiency since the technique used for the SS-PERG required high patient compliance and visual attention. Additional studies with larger groups are needed to validate our preliminary results.
Conclusion
Previous electrophysiological studies have demonstrated with the use of PVEP in dyslexic patients there is an alteration of the M–D pathway. In our study, we found that in dyslexia there is an alteration of the activity of M–Y retinal ganglion cells, especially in the left eye. These data confirm that in dyslexia there is a “minineglect” on the left eye that justifies the onset of a deficit of visual attention.
Acknowledgments
This work was done in memory of Maria Mottes.
We thank Paola Fiorini, Cristina Tura and Giulia Cecchin, the orthoptists of the Electrophysiology Service.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Lovegrove W, Bowling A, Badcock D, Blackwood M. Specific reading disability: differences in contrast sensitivity as a function of spatial frequency. Science. 1980;210:439–440. doi:10.1126/science.7433985
2. Slaghuis W, Lovegrove W. Spatial frequency mediated visible persistence and specific reading disability. Perception. 1985;13:527–534. doi:10.1068/p130527
3. Williams M, Molinet K, LeCluyse K. Visual masking as a measure of temporal processing in normal and disabled readers. Clin Vis Sci. 1989;4(2):137–144.
4. Livingstone MS, Rosen GD, Drislane FW, Galaburda AM. Physiological and anatomical evidence for a magnocellular defect in developmental dyslexia. Proc Natl Acad Sci. 1991;88:7943–7947. doi:10.1073/pnas.88.18.7943
5. Cornelissen P, Richardson A, Mason A, Fowler S, Stein F. Contrast sensitivity and coherent motion detection measured at photopic luminance levels in dyslexic and controls. Vis Res. 1995;35:1483–1494. doi:10.1016/0042-6989(95)98728-R
6. Eden GF, VanMeter JW, Rumsey JM, Maisog JM, Woods RP, Zeffiro TA. Abnormal processing of visual motion in dyslexia revealed by functional brain imaging. Nature. 1996;382:66–69. doi:10.1038/382066a0
7. Demb JB, Boynton GM, Best M, Heeger DJ. Psychophysical evidence for a magnocellular pathway deficit in dyslexia. Vis Res. 1998b;38:1555–1559. doi:10.1016/S0042-6989(98)00075-3
8. Vidyasagar TR, Pammer K. Impaired visual search in dyslexia relates to the role of the magnocellular pathway in attention. Neuroreport. 1999;10:1–5. doi:10.1097/00001756-199904260-00024
9. Hess RF, Thompson B, Gole GA, Mullen KT. The amblyopic deficit and its relationship to geniculo-cortical processing streams. J Neurophysiol. 2010;104(1):475–483. doi:10.1152/jn.01060.2009
10. Lee BB, Pokorny J, Smith VC, Martin PR, Valberg A. Luminance and chromatic modulation sensitivity of macaque ganglion cells and human observers. J Opt Soc Am A. 1990;7:2223–2236. doi:10.1364/JOSAA.7.002223
11. Merigan WH, Katz LM, Maunsell JH. The effects of parvocellular lateral geniculate lesions on the acuity and contrast sensitivity of macaque monkeys. J Neurosci. 1991;11:994–1001. doi:10.1523/JNEUROSCI.11-04-00994.1991
12. Derrington A, Lennie P. Spatial and temporal contrast sensitivities of neurons in lateral geniculate nucleus of macaque. J Physiol. 1984;357:219–240. doi:10.1113/jphysiol.1984.sp015498
13. Chatterjee S, Callaway EM. Parallel colour-opponent pathways to primary visual cortex. Nature. 2003;426:668–671. doi:10.1038/nature02167
14. Dacey DM, Packer OS. Colour coding in the primate retina: diverse cell types and cone-specific circuitry. Curr Opin Neurobiol. 2003;13:421–427. doi:10.1016/S0959-4388(03)00103-X
15. Martin PR, White AJ, Goodchild AK, Wilder HD, Sefton AE. Evidence that blue-on cells are part of the third geniculocortical pathway in primates. Eur J Neurosci. 1997;9:1536–1541. doi:10.1111/j.1460-9568.1997.tb01509.x
16. Kaplan E, Shapley RM. X and Y cells in the lateral geniculate nucleus of macaque monkeys. J Physiol. 1982;330:125–143. doi:10.1113/jphysiol.1982.sp014333
17. Solomon SG, White AJ, Martin PR. Temporal contrast sensitivity in the lateral geniculate nucleus of a New World monkey, the marmoset Callithrix jacchus. J Physiol. 1999;517:907–917. doi:10.1111/j.1469-7793.1999.0907s.x
18. Maddess T, Goldberg I, Dobinson J, Wine S, AH W, AC J. Testing for glaucoma with the spatial frequency doubling illusion. Vis Res. 1999;39:4258–4273. doi:10.1016/S0042-6989(99)00135-2
19. Romani A, Conte S, Callieco R, et al. Visual evoked potential abnormalities in dyslexic children. Funct Neurol. 2001;16:219–229.
20. Kobayashi T, Inagaki M, Yamazaki H, Kita Y, Kaga M, Oka A. Relationship between magnocellular function and reading skills in children: a study using visual evoked potentials. No To Hattatsu. 2014;46:424–428.
21. Brannan JR, Solan HA, Ficarra AP, Ong E. Effect of luminance in visual evoked potential amplitudes in normal and disabled readers. Optom Vis Sci. 1998;75:279–283. doi:10.1097/00006324-199804000-00025
22. Schulte‑Körne G, Bartling J, Deimel W, Remschmidt H. Motion‑onset VEPs in dyslexia. Evidence for visual perceptual deficit. Neuroreport. 2004;15:
23. Shandiz HJ, Heyrani M, Sobhani-Rad D, et al. Pattern visual evoked potentials in dyslexic children. J Ophthalmic Vis Res. 2017;12(4):
24. Kubova Z, Kuba M, Kreml.cˇek J, et al. Comparison of visual information processing in school-age dyslexics and normal readers via motion-onset visual evoked potentials. Vis Res. 2015;111:97–104. doi:10.1016/j.visres.2015.03.027
25. Corbetta M. Frontoparietal cortical networks for directing attention and the eye to visual locations: identical,independent or overlapping neural systems? Proc Natl Acad Sci U S A. 1998;95:831–838. doi:10.1073/pnas.95.3.831
26. Corbetta M, Akbudak E, Conturo TE, Snyder AZ, Ollinger JM, Drury HA. A common network of functional areas for attention and eye movements. Neuron. 1998;21:761–773. doi:10.1016/S0896-6273(00)80593-0
27. Peyrin C, Démonet JF, N’Guyen-Morel MA, LeBas JF, Valdois S. Superior parietal lobule dysfunction in a homogeneous group of dyslexic children with a visual attention span disorder. Brain Lang. 2011;118:128–138. doi:10.1016/j.bandl.2010.06.005
28. Lobier MA, Peyrin C, Pichat C, LeBas JF, Valdois S. Visual processing of multiple elements in the dyslexic brain: evidence for a superior parietal dysfunction. Front Hum Neurosci. 2014;8:479. doi:10.3389/fnhum.2014.00479
29. Kelly DH. Frequency doubling in visual responses. J Opt Soc Am. 1966;56:1628–1633. doi:10.1364/JOSA.56.001628
30. Johnson CA, Samuels SJ. Screening for glaucomatous visual field loss with frequency-doubling perimetry. Invest Ophthalmol Vis Sci. 1997;38:413–425.
31. Medeiros FA, Sample PA, Weinreb RN. Frequency doubling technology perimetry abnormalities as predictors of glaucomatous visual field loss. Am J Ophthalmol. 2004;137:863–871. doi:10.1016/j.ajo.2003.12.009
32. Schiavi C, Tassi F, Finzi A, Strobbe E, Cellini M. Steady-state pattern electroretinogram and frequency doubling technology in anisometropic amblyopia. Clin Ophthalmol. 2016;10:2061–2068. doi:10.2147/OPTH
33. Pammer K, Wheatley C. Isolating the M(y)-cell response in dyslexia using the spatial frequency doubling illusion. Vis Res. 2001;41:2139–2147. doi:10.1016/S0042-6989(01)00092-X
34. Avellis FO, Dassò A, Gandolfi S, Carta A. Magnocellular deficit in dyslexia: a preliminary analysis of possible patterns of visual field deficit tested with frequency doubling illusion. Transl Biomed. 2016;7(4):1–6.
35. Maddess T, James AC, Goldberg I, Wine S, Dobinson J. Comparing a parallel PERG, automated perimetry, and frequency-doubling thresholds. Invest Ophthalmol Vis Sci. 2000;41:3827–3832.
36. American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders.
37. Cornoldi C, Candela M. Prove di lettura e scrittura MT 16-19. Batteria per la verifica degli apprendimenti e la diagnosi di dislessia e disgrafia. Ed Centro Studi Erickson; 2015.
38. Siegel LS. Perspective on dyslexia. Pediatr Child Health. 2006;11(9):581–587. doi:10.1093/pch/11.9.581
39. Shaywitz SE. Dyslexia. N Engl J Med. 1998;338:307–312. doi:10.1056/NEJM199801293380507
40. Sun Z, Zou L, Zhang J, Mo S, Shao S, Zhong R. Prevalence and associated risk factors of dyslexic children in a middle‑sized city of China: a cross‑sectional study. PLoS One. 2013;8:e56688. doi:10.1371/journal.pone.0056688
41. Snowling MJ. Dyslexia.
42. Shaywitz S. Overcoming Dyslexia: A New and Complete Science-Based Program for Reading Problems at Any Level. New York: Alfred A. Knopf; 2003.
43. Snowling MJ. From language to reading and dyslexia. Dyslexia. 2001;7:37–46. doi:10.1002/(ISSN)1099-0909
44. Goswami U. Why theories about developmental dyslexia require developmental designs. Trends Cogn Sci. 2003;7:534–540.
45. Goswami U. A temporal sampling frame work for developmental dyslexia. Trends Cogn Sci. 2011;15:3–10. doi:10.1016/j.tics.2010.10.001
46. Ramus F. Developmental dyslexia: specific phonological deficitor general sensorimotor dysfunction? Curr Opin Neurobiol. 2003;13:212–218. doi:10.1016/S0959-4388(03)00035-7
47. Ramus F, Szenkovits G. What phonological deficit? Q J Exp Psychol. 2008;61:129–141. doi:10.1080/17470210701508822
48. Gabrieli JD. Dyslexia:anew synergy between education and cognitive neuroscience. Science. 2009;325:280–283. doi:10.1126/science.1171999
49. Shamma SA, Micheyl C. Behind the scenes of auditory perception. Curr Opin Neurobiol. 2010;2010(20):361–366. doi:10.1016/j.conb.2010.03.009
50. Hari R, Renvall H. Impaired processing of rapid stimulus sequences in dyslexia. Trends Cogn Sci. 2001;5:525–532. doi:10.1016/S1364-6613(00)01801-5
51. Facoetti A. Reading and selective spatial attention: evidence from behavioral studies in dyslexic children. In: Tobias HD, editor. Trends Dyslexia Research. NewYork, NY: Nova Science Publishers; 2004:35–71.
52. Facoetti A. Spatial attention disorders in developmental dyslexia: towards the prevention of reading acquisition deficits. In: Steinand J, Kapoula Z, editors. Visual Aspect of Dyslexia. Oxford: Oxford University Press; 2012:123–136.
53. Tainturier MJ. The cognitive deficits responsible for developmental dyslexia: review of evidence for a selective visual attentional disorder. Dyslexia. 2004;10:339–363. doi:10.1002/dys.284
54. Vidyasagar TR, Pammer K. Dyslexia:adeficit in visuo-spatial attention, not in phonological processing. Trends Cogn Sci. 2010;14:57–63.
55. Gori S, Facoetti A. Perceptual learning as a possible new approach for remediation and prevention of developmental dyslexia. Vis Res. 2014;99:78–87.
56. Pavlidis GT. Do eye movements hold the key to dyslexia? Neuropsychologia. 1981;19:57–64. doi:10.1016/0028-3932(81)90044-0
57. Bucci MP, Brémond-Gignac D, Kapoula Z. Latency of saccades and vergence eye movements in dyslexic children. Exp Brain Res. 2008a;188:1–12. doi:10.1007/s00221-008-1345-5
58. Bucci MP, Brémond-Gignac D, Kapoula Z. Poor binocular coordination of saccades in dyslexic children. Graefes Arch Clin Exp Ophthalmol. 2008b;246:417–428. doi:10.1007/s00417-007-0723-1
59. Galaburda AM, Kemper TL. Cytoarchitectonic abnormalities in dyslexia. Ann Neurol. 1979;6:94–100. doi:10.1002/ana.410060203
60. Galaburda AM, Lemay M, Kemper TM, Geschwind N. asymmetries in the brain may underlie cerebral dominance. Science. 1978;199:852–856. doi:10.1126/science.341314
61. Lawton T. Improving dorsal stream function in dyslexics by training figure/ground motion discrimination improves attention, reading fluency, and working memory. Front Hum Neurosci. 2016;10:397. doi:10.3389/fnhum.2016.00397
62. McCandliss BD, Cohen L, Dehaene S. The visual word form area: expertise for reading in the fusiform gyrus. Trends Cogn Sci. 2003;7:293–299.
63. Shaywitz SE, Shaywitz BA. Dyslexia: specific reading disability. Biol Psychiatry. 2005;57:1301–1309. doi:10.1016/j.biopsych.2005.01.043
64. Richlan F, Kronbichler M, Wimmer H. Functional abnormalities in the dyslexic brain: a quantitative meta-analysis of neuroimaging studies. Hum Brain Mapp. 2009;30:3299–3308.
65. Zhou W, Xia Z, Bi Y, Shu H. Altered connectivity of the dorsal and ventral visual regions in dyslexic children: a resting-state fMRI study. Front Hum Neurosci. 2015;10(9):495.
66. McLean GM, Stuart GW, Coltheart V, Castles A. Visual temporal processing in dyslexia and the magnocellular deficit theory: the need for speed? J Exp Psychol Hum Percept Perform. 2011;37(6):1957–1975. doi:10.1037/a0024668
67. Talcott JB, Hansen PC, Assoku EL, Stein JF. Visual motion sensitivity in dyslexia: evidence for temporal and energy integration deficits. Neuropsychologia. 2000;38(7):935–943. doi:10.1016/S0028-3932(00)00020-8
68. Skottum BC. The magnocellular deficit theory of dyslexia: the evidence from contrast sensitivity. Vis Res. 2000;40(1):111–127. doi:10.1016/S0042-6989(99)00170-4
69. Kuba M, Szanyi J, Gayer D, Kremlacek J, Kubova Z. Electrophysiological testing of dyslexia. Acta Med (Hradec Kralove). 2001;44(4):131–134. doi:10.14712/18059694.2019.99
70. Kubova Z, Kuba M, Kremlacek J, et al. Difficulties of motion-onset VEP interpretation in school-age children. Doc Ophthalmologica. 2014;128(2):121–129. doi:10.1007/s10633-014-9429-y
71. Petrusca D, Grivich MI, Sher A, et al. Identification and characterization of a Y-like primate retinal ganglion cell type. J Neurosci. 2007;27(41):11019–11027. doi:10.1523/JNEUROSCI.2836-07.2007
72. Patel N. The use of frequency doubling technology to determine magnocellular pathway deficiencies. J Behav Optom. 2004;15(2):31–36.
73. Werth R. Rapid improvement of reading performance in children with dyslexia by altering the reading strategy: a novel approach to diagnoses and therapy of reading deficiencies. Restor Neurol Neurosci. 2018;36:679–691. doi:10.3233/RNN-180829
74. Sousa MC, Biteli LG, Dorairaj S, Maslin JS, Leite MT, Prata TS. Suitability of the visual field index according to glaucoma severity. J Curr Glaucoma Pract. 2015;9(3):65–68. doi:10.5005/jp-journals-10008-1186
75. Lutaka NA, Grochowski RA, Kasahara N. Correlation between visual field index and other functional and structural measures in glaucoma patients and suspects. J Ophthalmic Vis Res. 2017;12(1):53–57. doi:10.4103/jovr.jovr_98_16
76. Sireteanu R, Goertz R, Bachert I, Wandert T. Children with developmental dyslexia show a left visual “minineglect”. Vis Res. 2005;45:3075–3082. doi:10.1016/j.visres.2005.07.030
77. Mather DS, Milford TM, McRae L. Does dyslexia develop from left eye dominance. Percept Mot Skills. 2015;121(2):569–601. doi:10.2466/15.10.PMS.121c21x5
78. Heravian J, Sobhani-Rad D, Lari S, et al. Pattern visual evoked potentials in dyslexic versus normal children. J Ophthalmic Vis Res. 2015;10(3):274–278. doi:10.4103/2008-322X.170361
79. Kubova Z, Kuba M, Peregrin J, Novakova V. Visual evoked potential evidence for magnocellular system deficit in dyslexia. Physiol Res. 1996;45(1):87–89.
80. Bach M, Brigell MG, Hawlina M, et al. ISCEV standard for clinical pattern electroretinography (PERG): 2012 update. Doc Ophthalmol. 2013;126(1):1–7. doi:10.1007/s10633-012-9353-y
81. Luo X, Frishman LJ. Retinal pathway origins of the pattern electroretinogram (PERG). Invest Ophthalmol Vis Sci. 2011;52(12):8571–8584. doi:10.1167/iovs.11-8376
82. Porciatti V, Ventura LM. Physiological significance of steady-state PERG losses in glaucoma: clues from simulation of abnormalities in normal subjects. J Glaucoma. 2009;18(7):535–542. doi:10.1097/IJG.0b013e318193c2e1
83. Crook JD, Peterson BB, Packer OS, Robinson FR, Troy JB, Dacey DM. Y-cell receptive field and collicular projection of parasol ganglion cells in macaque monkey retina. J Neurosci. 2008a;28:11277–11291. doi:10.1523/JNEUROSCI.2982-08.2008
84. Crook JD, Peterson BB, Packer OS, et al. The smooth monostratified ganglion cell: evidence for spatial diversity in the Y-cell pathway to the lateral geniculate nucleus and superior colliculus in the macaque monkey. J Neurosci. 2008b;28:12654–12671. doi:10.1523/JNEUROSCI.2986-08.2008
85. Rosemberg A, Talebi V. The primate retina contains distinct types of Y-like ganglion cells. J Neurosci. 2009;29(16):5048–5050. doi:10.1523/JNEUROSCI.0423-09.2009
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.