Back to Journals » Drug Design, Development and Therapy » Volume 20
Shuyu Wan Potentiates PD-1 Inhibitor Efficacy in Non-Small Cell Lung Cancer: Integrated Bioinformatics and Experimental Evidence for Gut Microbiota–Tumor Immune Crosstalk
Authors Sang T, Li Y, He Q, Wu W, Fang Q, Liu Y, Yue L, Shi L, Mei L, Huang C, Huang H, Zhang H, Wan Q, Wan Y
Received 27 April 2026
Accepted for publication 30 June 2026
Published 8 July 2026 Volume 2026:20 617793
DOI https://doi.org/10.2147/DDDT.S617793
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Anastasios Lymperopoulos
Tianqing Sang,1– 3,* Yajing Li,1,3,* Qihan He,1– 3,* Wei Wu,1 Qijun Fang,1 Yinglu Liu,1 Lijun Yue,1 Lijing Shi,1 Lifang Mei,4 Chubo Huang,4 Huang Huang,4 Huajun Zhang,1 Qian Wan,1 Yigang Wan1
1Department of Traditional Chinese Medicine, Nanjing Drum Tower Hospital Affiliated Hospital of Medical School, Nanjing University, Nanjing, People’s Republic of China; 2Institute of Chinese Medicine, Nanjing University, Nanjing, People’s Republic of China; 3Department of Traditional Chinese Medicine, Nanjing Drum Tower Hospital Clinical College of Nanjing University of Chinese Medicine, Nanjing, People’s Republic of China; 4International Jingfang Institute, Nanjing University of Chinese Medicine, Nanjing, People’s Republic of China
*These authors have contributed equally to this work
Correspondence: Yigang Wan, Email [email protected] Qian Wan, Email [email protected]
Background: Therapeutic heterogeneity limits the efficacy of immune checkpoint inhibitors (ICIs) in non-small cell lung cancer (NSCLC). Shuyu Wan (SYW), a classic TCM formula, has shown potential in modulating gut microbiota (GM) and enhancing immunotherapy, yet its synergistic mechanism with PD-1 inhibitors remains unclear.
Materials and Methods: SYW components were identified by UPLC-MS. NSCLC-related targets were integrated with SYW targets for pathway enrichment, and molecular docking validated component-target binding. NSCLC syngeneic mice were treated with SYW and/or PD-1 inhibitor (RMP1-14). Tumor growth, histopathology, serum cytokines, tumor-infiltrating CD8+T cell subsets (flow cytometry), PD-1/PD-L1 expression and co-localization (immunofluorescence), GM composition (16S rRNA), and metabolomics were assessed. FMT verified the role of GM–TME crosstalk.
Results: SYW monotherapy showed no significant tumor inhibition, whereas SYW combined with PD-1 inhibitor dose-dependently suppressed NSCLC growth. The combination reduced PD-1 expression and PD-1/PD-L1 co-localization, elevated serum IL-12, IFN-γ, and TNF-α, increased total tumor-infiltrating CD8+ T cells, decreased PD-1+ and TIM-3+ exhausted subsets, and expanded IFN-γ+ and Granzyme B+ effector subsets. Concurrently, it reshaped GM (increased Bacillota, decreased Patescibacteria) and altered metabolites (L-glycine, L-proline). These effects were abolished in antibiotic treated mice and restored by FMT, suggesting GM–TME crosstalk as essential.
Conclusion: SYW acts as a microbiota-dependent immune sensitizer that potentiates PD-1 inhibitor efficacy in NSCLC by remodeling GM and enhancing effector CD8+ T cell infiltration while reducing exhaustion. GM–TME crosstalk is the potential mechanism, supporting SYW as an adjunct to PD-1 blockade in NSCLC therapy.
Keywords: Shuyu Wan, non-small cell lung cancer, PD-1 inhibitors, gut microecology, tumor immune microenvironment
Introduction
Lung cancer remains the most prevalent and lethal malignancy worldwide, with non-small cell lung cancer (NSCLC) accounting for over 85% of all cases. Its high invasiveness, recurrence rate, and therapeutic resistance present persistent challenges in clinical management.1,2 In recent years, the field of cancer therapy has undergone a revolutionary shift, progressing from the indiscriminate killing of traditional chemotherapy and radiotherapy to the precision targeting of molecular therapies, and further to the active defense of immunotherapy. Immune checkpoint inhibitors (ICI), particularly monoclonal antibodies targeting the Programmed cell death protein 1 (PD-1)/programmed death - ligand 1(PD-L1) pathway, have brought hope for long-term survival to patients with advanced NSCLC by reversing T-cell immunosuppression, restoring the recognition and killing functions of CD8+ exhausted cytotoxic T cells (Tex), and increasing the activity of CD8+ effector T cells (Teff).3–5 They are currently established as first-line treatments for NSCLC patients without driver gene mutations or with resistance to targeted therapies.6 However, the clinical benefit of ICI monotherapy remains heterogeneous in NSCLC. Even among patients with high PD-L1 expression (≥50%), objective response rates to pembrolizumab monotherapy are approximately 39–45% in the first-line setting and around 30% in previously treated patients, while response rates in broader or lower PD-L1-expressing populations are generally lower, often around 18%–27%.7–9 This heterogeneity primarily stems from the complexity of the tumor immune microenvironment (TME), including insufficient CD8+ T- cell infiltration, excessive accumulation of regulatory T cells (Tregs), and imbalanced secretion of pro-inflammatory cytokines.10,11 Therefore, improving the TME and enhancing the anti-tumor activity of T cells have become core scientific issues in overcoming ICI heterogeneity and improving treatment response rates.
The human commensal microbiota plays a central role in maintaining physiological homeostasis throughout the body and at local epithelial barriers by regulating innate and adaptive immunity. Among them, the gut microecology (GM), regarded as a “hidden immune organ,” has emerged as a research frontier due to its close association with tumor immunity.12 A 2024 study published in Cell by Jia, D. et al demonstrated that Lactobacillus johnsonii and Clostridium sporogenes synergistically produce indole-3-propionic acid (IPA), which enhances CD8+ T-cell stemness through H3K27 acetylation, reverses T-cell exhaustion in models of melanoma, breast cancer, and colorectal cancer, and thereby improves the efficacy of PD-1 inhibitors (PD-1 Inh).13 A 2025 report in Nature by a Japanese team described the isolation of a novel Ruminococcaceae strain, YB328 (Hominenteromicrobium mulieris), from the feces of NSCLC patients responding to PD-1 therapy. This strain activates intestinal CD103+CD11b− dendritic cells, promotes their migration to the tumor microenvironment, and enhances antigen presentation and CD8+ T - cell activation.14 These findings not only clarify the molecular basis by which GM influences ICIs efficacy but also provide a new strategy for optimizing immunotherapy through GM modulation.
In the comprehensive treatment of malignant tumors, the advantages of traditional Chinese medicine (TCM) in “holistic regulation” have become increasingly prominent. Shuyu Wan (SYW), a classic formula from Zhang Zhongjing’s Jin Kui Yao Lue (Eastern Han Dynasty), strengthens vital qi and consolidates the root, showing value in treating malignancies.15 Facing NSCLC immunotherapy efficacy heterogeneity, SYW’s “vital qi-strengthening” mechanism combine with gut microecology (GM)-tumor immune microenvironment (TME) crosstalk offers a breakthrough. Key evidence from its monarch medicines: SYPs inhibit HCC immune escape and enhance chemosensitization by simultaneously suppressing HIF-1αand PD-L1.15 Ginseng polysaccharides enhance NSCLC sensitivity to PD-1 inhibitors by enriching Parabacteroides, upregulating valeric acid, and reducing kynurenine/tryptophan ratio.16 Yam polysaccharides enrich Clostridium_UCG-014, reduce pro-tumor metabolites like deoxyguanosine, and improve PD-1 inhibitor efficacy in colorectal cancer.17 Consistent with reports that SYW regulates Lactobacillus, Bifidobacterium, and the HMGB1 pathway,18 it potentially modulates tumor immune responses via GM-immune microenvironment crosstalk.
However, the dynamic regulatory network of gut microbiota remodeling, metabolite changes, and TME in the context of SYW-based NSCLC immunotherapy remains unelucidated. This study intends to integrate bioinformatics and experimental verification to analyze the mechanism by which SYW enhances the anti-NSCLC efficacy of PD-1 Inh by reshaping GM and regulating GM-TME crosstalk, providing theoretical support for optimizing NSCLC immunotherapy through the integration of traditional Chinese and Western medicine.
Materials and Methods
SYW Preparations
SYW consists of Rhizoma Dioscoreae (Shanyao, SY, 50 g, Dioscorea opposita Thunb)., Ginseng Radix et Rhizoma (Renshen, RS, 10 g, Panax ginseng C.A.Mey)., Rhizoma Atractylodis macrocephalae (Baizu, BZ, 10 g, Atractylodes macrocephala Koidz)., Poria (Fulin, FL, 10 g, Poria cocos (Schw). Wolf), Radix Glycyrrhizae Praeparata cum Melle (Gancao, GC, 10 g, Glycyrrhiza uralensis Fisch. ex DC). Radix Angelicae Sinensis (Danggui, DG, 10 g, Angelica sinensis (Oliv). Diels), Rhizoma Chuanxiong (Chuanxiong, CX, 10 g, Ligusticum chuanxiong Hort)., Radix Paeoniae Alba (Baishao, BS, 10 g, Paeonia lactiflora Pall)., Radix Rehmanniae Preparata (Shudi, SD, 15 g, Rehmannia glutinosa (Gaertn). Libosch. ex Fisch. et Mey)., Asini Corii Colla (Ejiao, EJ, 10 g, Equus asinus L)., Cinnamomi Ramulus (Guizhi, GZ, 10 g, Cinnamomum cassia Presl), Ophiopogonis Radix (Maidong, MD, 15 g, Ophiopogon japonicus (L. f). Ker-Gawl)., Bupleuri Radix (Chaihu, CH, 10 g, Bupleurum chinense DC)., Saposhnikoviae Radix (Fangfeng, FF, 10 g, Saposhnikovia divaricata (Turcz). Schischk)., Armeniacae Semen Amarum (Xinren, XR, 10 g, Prunus armeniaca L. var. ansu Maxim)., Platycodonis Radix (Jiegen, JG, 10 g, Platycodon grandiflorus (Jacq). A. DC)., Ampelopsis Radix (Bailian, BL, 10 g, Ampelopsis japonica (Thunb). Makino), Medicinalis Fermentata (Shenqu, SQ, 10 g, Massa Medicata Fermentata), Sojae Semen Germinatum (Dadouhuangjuan, DDHJ, 10 g, Glycine max (L). Merr)., Zingiberis Rhizoma Exsiccatum (Ganjiang, GJ, 10 g, Zingiber officinale Rosc)., Jujubae Fructus (Dazao, DZ, 50 g, Ziziphus jujuba Mill). The medicinal materials were sourced from Pharmacy of Traditional Chinese Medicine, Affiliated Drum Tower Hospital, Medical School of Nanjing University. And they identified by Professor Huang Huang, and the voucher specimens (No. SYW-101) of the herbal pieces are deposited in the Herbarium of the Jingfang Institute, Nanjing University of Chinese Medicine. The medicinal materials were added to 8 times the amount of water and decocted twice (first decoction for 1.5 h, second for 1 h). The two decocted liquids were combined, concentrated to 1.5 g/mL, and stored at −20°C.
UPLC-MS
Take 100 μL SYW liquid into a 1.5 mL centrifuge tube, add 300 μL extraction solution (methanol:acetonitrile = 1:1) containing internal standards (eg., L-2-chlorophenylalanine, 0.02 mg/mL). Vortex for 30s, ultrasonically extract at 5°C, 40 KHz for 30 min, stand at −20°C for 30 min, then centrifuge (13000 g, 4°C) for 15 min. Dry the supernatant under nitrogen, redissolve with 100 μL methanol:acetonitrile = 1:1, vortex, ultrasonically extract at 5°C, 40 KHz for 5 min, centrifuge (13000 g, 4°C), and transfer the supernatant to an injection vial. Mix 20 μL supernatant from each sample to prepare quality control samples for monitoring analysis stability.
Analyze using UHPLC-Q Exactive HF-X (Thermo Scientific) with an ACQUITY UPLC HSS T3 column (100 mm × 2.1 mm, 1.8 μm) at 40°C, injection volume 3 μL. Mobile phase A: 95% water + 5% acetonitrile (0.1% formic acid); B: 47.5% acetonitrile + 47.5% isopropanol + 5% water (0.1% formic acid). Gradient elution: 0–1 min, 5% B; 1–15 min, 5–95% B; 15–16 min, 95% B; 16–16.1 min, 95–5% B; 16.1–20 min, 5% B. Flow rate 0.3 mL/min. Collect data in positive and negative ion modes via ESI. The ion source parameters are described in Table S2.
Filter the data, identify peaks, correct retention times, and align peaks using Progenesis QI v3.0 (Waters Corporation, Milford, USA). Eventually, a data matrix containing information such as retention time, mass - to - charge ratio, and peak intensity is obtained. The MS mass error is set to less than 10 ppm. Metabolites are identified based on the secondary mass spectrometry matching scores according to public databases (http://www.hmdb.ca/, https://metlin.scripps.edu/).
Identification of NSCLC-Associated Genes and SYW Targets
UPLC-MS-identified SYW components were matched against TCMSP (https://old.tcmsp-e.com/index.php) and filtered by oral bioavailability (OB ≥ 30%), drug-likeness (DL ≥ 0.18), and half-life (HL ≥ 4 h). Their target genes were retrieved from TCMSP, validated via Pubmed, CNKI, and Wanfang, and standardized using UniProt (https://www.uniprot.org). NSCLC-related genes were obtained from GEO. After filtering for Homo sapiens, differential expression analysis was performed with GEO 2R (p < 0.05, |log2FC| ≥ 2), heatmaps and volcano plots were generated via “ggplot2” in R. These genes were further filtered and supplemented using keywords “Non-small cell lung cancer”, “Lung adenocarcinoma”, and “Lung squamous cell carcinoma” in GeneCards (https://www.genecards.org) and OMIM (http://www.omim.org).
Prediction of Potential Mechanisms of SYW in Treating NSCLC
Potential SYW-NSCLC targets, identified by intersecting SYW compound targets and NSCLC-related genes, were visualized via a Venn diagram. The PPI network was constructed using STRING v11.0 (Homo sapiens, minimum interaction confidence score = 0.9). GO and KEGG enrichment analyses of these targets were performed using DAVID (https://david.ncifcrf.gov/) and Metascape (http://metascape.org/gp/index.html). Significant terms included biological processes (BP), molecular functions (MF), cellular components (CC), and KEGG pathways with p < 0.05.
Batch Molecular Docking Verification
Cytoscape 3.7.2 was used to construct and visualize an integrated network of SYW compounds, NSCLC targets, and enriched pathways. Core compounds and targets were identified via built-in tools analyzing topology parameters (degree, betweenness centrality, closeness centrality). 3D structures (PDB format) of target proteins (Homo sapiens) were retrieved from RCSB-PDB (http://www.RCSB.org), with dehydration and removal of excess ligands performed using PyMOL. 3D structures (SDF format) of potential active compounds were obtained from PubChem (https://pubchem.ncbi.nlm.nih.gov) and converted to mol2 format via Chem3D. Molecular docking between components and target proteins was conducted using Outoduck 1.1.2, with results visualized by PyMOL. Heatmaps for batch molecular docking were generated using R packages.
Animal Model and Experimental Design
This study was carried out in strict accordance with the ARRIVE guidelines and approved by the Ethics Committee of the Affiliated Hospital of Nanjing University School of Medicine (Approval No. 2025AE01047). All mice were housed in specific pathogen-free (SPF) environment with controlled temperature (22–25 °C), humidity (50–60%), 12 h light/12 h dark cycle, and free access to autoclaved food and water.
Forty-eight male C57BL/6 mice (4–6 weeks old) were purchased from Celine Biosciences (Nanjing) Co., Ltd. (Nanjing, China, animal license: SCXK (Su) 2022–0006). Luciferase-transfected Lewis lung carcinoma (LLC-luc) cells were obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA). Cells were authenticated by short tandem repeat (STR) profiling within 6 months before use and cultured in our laboratory as described below. Cells were maintained in RPMI 1640 medium supplemented with 10% fetal bovine serum (Gibco, Grand Island, NY, USA), 100 U/mL penicillin, and 100μg/mL streptomycin (Solarbio, Beijing, China) at 37°C in a 5% CO2 incubator (Thermo Fisher Scientific, Waltham, MA, USA). For tumor inoculation, 2×106 LLC-luc cells in 100 μL PBS were subcutaneously injected into the left axilla of each mouse. Seven days after inoculation, 30 mice were randomly divided into 5 groups (n=6 per group) using a random number table: control group, SYW, low-dose SYW+RMP1-14 group, high-dose SYW+RMP1-14 group, and RMP1-14 monotherapy group. Based on body surface area conversion, SYW was administered by intragastric gavage at 20 g/kg/d (low dose) and 40 g/kg/d (high dose) for 14 consecutive days. RMP1-14 (BE0146, Bio X Cell, Lebanon, NH, USA) was injected intraperitoneally at 5 mg/kg twice weekly for 14 days. Another 18 mice were randomly assigned to conventional (N), antibiotic treated (ABX), and fecal microbiota transplantation (FMT) groups (n=6 per group). Mice in the ABX and FMT groups were treated with a broad-spectrum antibiotic cocktail (ampicillin 1 g/L, neomycin 1 g/L, metronidazole 1 g/L, vancomycin 0.5 g/L in drinking water for 1 week to deplete gut microbiota. The FMT group received 200 μL fresh fecal suspension from healthy donor mice by intragastric gavage once daily for 3 consecutive days after antibiotic treatment. For microbiota reconstitution, fresh fecal pellets were collected daily from age- and gender-matched healthy tumor-free C57BL/6 donor mice with intact endogenous gut microbiota. Fecal suspensions were prepared immediately after collection in sterile PBS at a concentration of 100 mg/mL, homogenized thoroughly and filtered through a 70 μm sterile cell strainer to remove insoluble particulate matter. Mice in the FMT group received 200 μL of fresh fecal suspension by intragastric gavage once daily for 3 consecutive days after the 7-day antibiotic depletion. All three groups received high-dose SYW combined with RMP1-14 for 14 consecutive days.
At the end of the experiment, mice were anesthetized with 2% isoflurane inhalation until deep anesthesia was achieved and euthanized by cervical dislocation under anesthesia. Tumor tissues and intestinal contents were rapidly dissected under sterile conditions, snap-frozen in liquid nitrogen.
In vivo Bioluminescence Imaging
On the final day of treatment, mice were anesthetized with 2% isoflurane inhalation and intraperitoneally injected with D-luciferin potassium salt (150 mg/kg body weight). After 10–15 minutes of dark incubation, in vivo bioluminescence imaging was performed using a Tanon 5200 Multi In Vivo Imaging System (Tanon Science & Technology Co., Ltd., Shanghai, China) with an exposure time of 10–30 seconds. Tumor bioluminescence intensity (photons/sec/cm2/sr) was quantified using the matched Tanon image analysis software to determine relative tumor burden.
Tumor Inhibition Rate
Mice were weighed every 3 days, and tumor dimensions (long diameter L, short diameter W, in mm) were measured with vernier calipers. Tumor volume was calculated as: V =1/2 ×L×W2. At study endpoint, mice were sacrificed; tumors were dissected, weighed, and tumor inhibition rate (TIR) calculated as: TIR (%) = [(Control mean tumor weight - Experimental mean tumor weight)/Control mean tumor weight] × 100%.
Pathology of Tumor Tissue
Tumor tissues were gently rinsed with pre-cooled PBS to remove residual blood and impurities, then trimmed of surrounding fat/connective tissue and cut into 3–5 mm3 pieces. These were fixed in 4% paraformaldehyde for 24–48 hours, followed by dehydration in gradient ethanol (70%–100%, 1–2 hours each), xylene treatment (twice, 30 minutes each), and paraffin infiltration (three times, 1 hour each at 56–58°C) before embedding into blocks. 4–5μm sections were mounted on poly-L-lysine-coated slides and baked at 60°C for 30 minutes to fix. After dewaxing in xylene (twice, 5 minutes each) and hydration through gradient ethanol (100%→70%, 2 minutes each) to distilled water, sections were stained with hematoxylin for 5–8 minutes, rinsed, differentiated with 1% hydrochloric acid ethanol, and blued in water for 10–15 minutes. Subsequent eosin staining (1–2 minutes), gradient ethanol dehydration, xylene clearing, and neutral gum mounting were performed. Tissue structure, cell morphology, nuclear chromatin, atypia, and pathological mitoses were observed under an optical microscope.
Immunofluorescence
Tumor paraffin sections (5 μm) were deparaffinized, rehydrated, and subjected to antigen retrieval in citrate buffer (pH 6.0). After permeabilization with 0.1% Triton X-100 and blocking with 5% BSA containing 10% goat serum, sections were incubated overnight at 4°C with the following primary antibodies: anti-PD-1 (#135208, BioLegend, San Diego, CA, USA) and Alexa Fluor 647-conjugated anti-PD-L1 (#50625, Cell Signaling Technology, Danvers, MA, USA). After washing, sections were incubated with Alexa Fluor 488-conjugated anti-rat IgG (green) to visualize PD-1 signals, followed by DAPI counterstaining of nuclei (blue). Images were acquired using a Leica DMi8 fluorescence microscope. The mean fluorescence intensity (MFI) of PD-1 and PD-L1 was quantified via ImageJ. Co-localization was assessed by line-scan intensity profiling of the two fluorescent signals across representative regions.
Elisa
Tumor tissues were homogenized in ice-cold lysis buffer with protease inhibitors. After centrifugation (12,000 rpm, 15 min, 4°C), supernatants were assayed for IL-2, IFN-γ, and TNF -α using ELISA kits (CSB-E04584m, CSB-E04578m, CSB-E04741m, Cusabio, Wuhan, China). Samples, standards, and blanks (100 μL/well) were incubated at 37°C for 2 h. Following 5 washes per step, plates were sequentially incubated with biotinylated antibody (1 h) and enzyme conjugate (30 min) at 37°C. Substrate (90 μL) was added, incubated for 15–20 min in the dark, and reactions were stopped with 50 μL stop solution. OD450 was measured immediately, and concentrations were calculated via standard curves for statistical analysis.
Flow Cytometric Analysis of Tumor-Infiltrating CD8⁺ T Cell Subsets
Fresh tumor tissues were minced and digested with collagenase IV and DNase I at 37°C for 30–60 min. Single-cell suspensions were filtered through a 70-μm strainer, centrifuged, and resuspended in 2% FBS-PBS. Cells were stained with Fixable Viability Dye eFluor 780 (65–0865-14, Thermo Fisher Scientific eBioscience, San Diego, CA, USA) to exclude dead cells. After blocking Fc receptors, surface staining was performed with the following fluorophore-conjugated antibodies (all from Thermo Fisher Scientific eBioscience, San Diego, CA, USA): CD45 mAb (30-F11, PerCP-Cyanine5.5, 45–0451-80), CD3 mAb (17A2, Brilliant Violet 421, 40–0032-82), CD8a mAb (53–6.7, FITC, 11–0081-81), PD-1 (CD279) mAb (RMP1-30, APC, 17–9981-80), TIM-3 (CD366) mAb (RMT3-23, PE, 12–5870-81). For intracellular staining, cells were stimulated with PMA/ionomycin/brefeldin A, fixed/permeabilized, then stained with IFN-γ mAb (XM G1.2, APC, 17–7311-81), Granzyme B mAb (NGZB, PE, 12–8898-80). Isotype controls were set in parallel. Gating strategy (BD FACSAria™, FlowJo, BD Biosciences, Franklin Lakes, NJ, USA): FSC/SSC gated mononuclear cells; FSC-A/FSC-H excluded doublets Viability dye excluded dead cells; selected CD45+ immune cells, Gated CD3+ T cells from CD45+ population, Gated CD8+ T cells from CD3+ population, Quantified CD8+PD-1+, CD8+TIM-3+, CD8+IFN-γ+, CD8+Granzyme B+ subsets.
16S rRNA Gene Sequencing of Gut Microbiota
Intestinal contents of the mice were collected after administrating with a corresponding dose of agents for gut microbiota analyses. Stool DNA was extracted using PowerSoil DNA Isolation Kit (MOBIO, Carlsbad, CA,USA) according to the manufacturer’s instructions. The 16S rDNA V3-V4 region was amplified by PCR using the 343F and 798R primers (343F: TACGGRAGGCAGCAG; 798R: AGGGTATCTAATCCT); then we performed the quantification, qualification, and purification of the PCR product. Sequencing was performed on the Illumina MiSeq (Illumina Inc., San Diego, CA,USA) based on the previous study. Raw reads were filtered using FLASH (Version 1.2.11) with a Phred quality score threshold of 20, and chimeric sequences were removed using UCHIME. Operational taxonomic units (OTUs) were picked at 97% sequence similarity, and then the taxonomical identification was aligned using the Silva database (Release 132). Moreover, the RDP classifier was applied to assign 16S rRNA sequences at a given taxonomic rank. Alpha diversity was calculated using QIIME2, and beta diversity was visualized using principal coordinate analysis (PCoA). Differential taxa were identified using LEfSe with LDA score > 2.0 and p < 0.05. All samples were processed in a single batch to minimize batch effects.
Non-Targeted Metabolomics of Gut Microbiota
Mouse intestinal content samples (200 mg) were thawed on ice, added with 1 mL pre - cooled methanol/water (8:2), vortexed for 30s, and ice - bathed for ultrasonic treatment (40 kHz) for 15 min. After centrifuging at 13,000 rpm for 15 min at 4°C, 800 μL of supernatant was taken, dried by nitrogen blow, re - dissolved in 100 μL acetonitrile/water (1:1), vortexed for 1 min, then centrifuged at 13,000 rpm for 10 min at 4°C. The supernatant was collected for testing. For UPLC, an ACQUITY HSS T3 column (100 mm×2.1 mm, 1.8 μm) was used at 40°C. The mobile phase A (0.1% formic acid in water) - B (acetonitrile) had a gradient elution: 0–1 min 5% B, 1–12 min 5–95% B, 12–14 min 95% B, 14–14.1 min back to 5% B, 14.1–16 min at 5% B. The flow rate was 0.3 mL/min, and injection volume was 5 μL. Mass spectrometry used an ESI source (positive/negative ion mode) with capillary voltage 3.5/2.5 kV, cone voltage 35 V, desolvation gas at 500°C, 800 L/h, scanning m/z 100–1500 (2 Hz). One QC sample of equal - volume mixture was inserted per 10 samples to assess stability. Data processed by XCMS (peak extraction, correction, alignment) was used to identify metabolites with Compound Discoverer by matching HMDB, METLIN, and a self - built database (error <5 ppm). After log2 - transformation and normalization, PCA and OPLS - DA were used to screen differential metabolites (VIP > 1.0, P < 0.05), and a cluster heatmap was drawn. MetaboAnalyst 5.0 was used for metabolic pathway enrichment analysis (P < 0.05) and graphing.
Statistical Analysis
All statistical analyses were performed using GraphPad Prism 9.0 (GraphPad Software, Inc., USA) and R software (v4.2.1). Data are presented as the mean ± standard deviation (SD) unless otherwise specified, with sample sizes (n) indicated in the figure legends. For longitudinal data, mixed-effects models with Tukey’s post-hoc test were used to account for repeated measurements from the same animal. For single-time-point comparisons (, one-way ANOVA with Tukey’s post-hoc test was used for normally distributed data, and the Kruskal–Wallis test for non-normally distributed data. Statistical significance was set at p < 0.05.
Results
Bioinformatics Identifies PD-1/PD-L1 Pathway and Gut Microbiota as Key Targets of SYW in NSCLC Intervention
Phytochemical Profiling and Target Prediction of SYW Active Components
Phytochemical analysis of SYW was performed using UPLC-MS combined with database searches against HMDB and METLIN. Under both positive and negative ion modes, the peaks exhibited good shape and relatively uniform distribution. A total of 356 components were identified (Table S1), and 30 active components were further confirmed by accurate molecular weight determination and characteristic peak comparison (Figure 1A–B). From these, 10 core bioactive components were selected as representative quality markers of SYW, including signature steroidal saponins of SY (dioscin, pseudoprotodioscin), ginsenosides from RS (Rb1, Rg1, Re), and characteristic components from Saposhnikoviae Radix, Chuanxiong Rhizoma and Glycyrrhizae Radix et Rhizoma. Their chemical structures are displayed in Figure 1C.
 |
Figure 1 Identification of active components and target prediction of SYW by UPLC-MS combined with network pharmacology. (A) Total ion current (TIC) chromatogram of SYW acquired under positive ion mode. (B) TIC chromatogram of SYW acquired under negative ion mode. (C) Chemical structures of 10 representative bioactive components in SYW. (D) Distribution network of active components from various herbs in SYW and their corresponding targets. |
These 10 core components included dioscin and pseudoprotodioscin (SY, the monarch herb of SYW), ginsenosides Rb1, Rg1 and Re (signature triterpenoid saponins of RS), cimifugin (FF), senkyunolide F (CX), isoliquiritigenin (GC), gallic acid (a ubiquitous polyphenolic marker), and panaxynol (a characteristic polyacetylene constituent of RS).
By integrating UPLC-MS results with TCMSP database searches, followed by ADME screening and UniProt normalization, 185 compounds corresponding to each herb in SYW were finally obtained: SY (16), RS (20), BZ (5), FL (14), GC (78), CX (6), BS (10), SD (2), MD (1), CH (14), FF (9), XR (12), JG (7), BL (7), and DZ (23) (17 components were shared by different herbs), together with their 264 corresponding targets (Figure 1D).
Mining of Potential Targets for SYW Mediated NSCLC Intervention
Dataset GSE10072 was retrieved from the GEO database, which contains 22,283 genes from 58 fresh NSCLC tissue samples and 49 healthy lung tissue samples profiled using the Affymetrix GeneChip Human Genome U133 Array. Differential gene analysis between the two groups was performed using GEO2R with a threshold of p < 0.05, yielding 11,964 differentially expressed genes. These results indicated remarkable differences in gene expression between NSCLC and normal lung tissues (Figure 2A–C).
 |
Figure 2 Analysis of overlapping targets between SYW and NSCLC. (A) UMAP clustering plot of GSE10072 dataset. (B) Volcano plot of differentially expressed genes between NSCLC and healthy tissues (|log FC| > 2, p < 0.05). (C) Heatmap analysis of differential genes. (D) Venn diagram of SYW-NSCLC overlapping genes. (E) PPI network of SYW - NSCLC overlapping genes. |
After further screening with |log FC| > 2 and integration with GeneCards, OMIM databases and literature retrieval, 5,803 NSCLC-related genes were obtained. A total of 200 overlapping targets between SYW and NSCLC were identified using a Venn diagram (Figure 2D). A PPI network of these overlapping targets was constructed using Cytoscape, showing that TP53, AKT1, STAT3, BCL-2, IL-6 and others were the core hub genes in the intersection (Figure 2E).
Pathway Prediction and Molecular Docking Validation of SYW-NSCLC Interactions
Enrichment analysis of SYW-NSCLC overlapping targets indicated that SYW intervention in NSCLC mainly affected biological processes including “response to molecule of bacterial origin” and “positive regulation of programmed cell death”; molecular functions including “protein kinase binding” and “cytokine activity”; and cellular components including “protein kinase complex”.
Key involved pathways included “PD-L1 expression and PD-1 checkpoint pathway in cancer”, “T cell receptor signaling pathway”, “Chemokine signaling pathway”, and “Escherichia coli, Salmonella, and Shigellosis infection” (Figure 3A and B).
 |
Figure 3 Key pathways of SYW in intervening NSCLC and batch molecular docking validation. (A) GO enrichment analysis of SYW-NSCLC overlapping targets. (B) KEGG pathway enrichment analysis of SYW-NSCLC overlapping targets. Red: Immune pathways; Blue: Intestinal infection pathways. (C) SYW-NSCLC overlapping targets enriched in the PD-1/PD-L1 pathway and corresponding SYW components. (D) Heatmap of batch molecular docking between core components of SYW and key targets in the PD-1/PD-L1 pathway. |
Network analysis of PD-1/PD-L1 pathway-related targets and SYW components was performed, and core targets were screened using topological criteria (degree ≥ 2, BC ≥ 0.2, CC ≥ 0.5). Batch molecular docking of core components [A13 (quercetin), JG5 (luteolin), A4 (kaempferol), FF6 (wogonin), GC9 (naringenin)] with key targets (MAPK14, RELA, AKT1, STAT1, STAT3) showed that all complexes possessed negative binding energies. Notably, component-target pairs matched by database exhibited binding energies ≤ −7 kcal/mol, indicating strong binding affinity (Figure 3C and D).
In vivo Validation Demonstrates that SYW Synergizes with PD-1 Inh to Inhibit NSCLC, Ameliorate TME, and Reconstruct the Gut Microecology
SYW Synergizes with PD-1 Inh to Enhance the Inhibition of NSCLC in vivo
All experimental mice survived until the end of the study, with no evident systemic toxicity or adverse events observed. In vivo bioluminescence imaging showed the lowest signal intensity in the H-SYW+RMP1-14 group, followed by the L-SYW+RMP1-14 group, while SYW monotherapy showed no significant difference from the control group (Figure 4A and B). The same trend was observed for tumor volume, ex vivo tumor weight and tumor inhibition rate (Figure 4C, E and F). Mixed-effects model analysis revealed a significant treatment × time interaction (F(24, 120) = 8.45, p < 0.05), indicating that tumor growth trajectories differed significantly among groups.
 |
Figure 4 SYW synergizes with PD-1 Inh to the in vivo inhibitory effect on NSCLC. (A) In vivo bioluminescence imaging of tumor burden in NSCLC syngeneic mice (n=6). (B) Quantitative statistical analysis of tumor bioluminescence signals to reflect tumor burden in NSCLC syngeneic mice (n=6). (C) Tumor volume growth curves (n=6). (D) Body weight changes (n=6). (E) Ex vivo tumor weight (n=6). (F) Tumor inhibition rate (n=6). (G) HE staining of tumor tissues (n=6). *p < 0.05 vs. control group, Δp < 0.05 vs. SYW group, %p < 0.05 vs. L-SYW+RMP1-14 group, &p < 0.05 vs. H-SYW+RMP1-14 group, #p< 0.05 vs. RMP1-14 group. |
Histopathological analysis revealed that H-SYW+RMP1-14 tumors showed sparse cellularity, prominent apoptotic features and increased inflammatory infiltration, which were distinct from the control and monotherapy groups (Figure 4G). Body weight in the H-SYW+RMP1-14 group remained stable or slightly increased, indicating favorable tolerability and no obvious systemic toxicity (Figure 4D).
These findings indicate that SYW synergizes with PD-1 blockade to suppress NSCLC growth in a dose-dependent manner with an excellent safety profile.
Effects of SYW Combined with PD-1 Inh on PD-1/PD-L1 Expression and Co-Localization in Tumor Tissues
Immunofluorescence co-staining was performed to detect PD-1 and PD-L1 expression in tumor tissues. Compared with the control group, the combination groups showed significantly decreased PD-1 expression and increased PD-L1 expression in a dose-dependent manner (Figure 5A–C). Line-scan analysis revealed that co-localization of PD-1 and PD-L1 was markedly reduced in the combination groups, especially in the H - SYW + RMP1 - 14 group (Figure 5D–H). No obvious changes in PD-1/PD-L1 expression or distribution were observed in the SYW monotherapy group.
 |
Figure 5 SYW synergizes with PD-1 Inh modulates PD-1/PD-L1 expression and co-localization in tumor tissues. (A) Immunofluorescence staining of PD-1/PD-L1 in tumor tissues (PD-1 red, PD-L1 green), n=6. (B) Quantitative analysis of PD-1 mean fluorescence intensity (MFI) across all groups, n=6. (C) Quantitative analysis of PD-L1 MFI across all groups, n=6. (D) Line-scan PD-1/PD-L1 signal distribution, Ctrl group; (E) Line-scan PD-1/PD-L1 signal distribution, SYW group. (F) Line-scan PD-1/PD-L1 signal distribution, L-SYW+RMP1-14 group. (G) Line-scan PD-1/PD-L1 signal distribution, H-SYW+RMP1-14 group. (H) Line-scan PD-1/PD-L1 signal distribution, RMP1-14 group. Statistical annotations were identical to those in *p < 0.05 vs. control group, Δp < 0.05 vs. SYW group, %p < 0.05 vs. L-SYW+RMP1-14 group, &p < 0.05 vs. H-SYW+RMP1-14 group, #p< 0.05 vs. RMP1-14 group. |
Effects of SYW Combined with PD-1 Inh on Intratumoral Cytokines and Tumor-Infiltrating CD8+ T Cell Subsets
To evaluate local immune activation status in the tumor microenvironment, we measured the concentrations of IL-2, IFN-γ and TNF-α in tumor tissue homogenates by ELISA. The results showed that all three effector cytokines were dose-dependently upregulated in the combination groups compared with the control and RMP1-14 monotherapy groups, reflecting enhanced local anti-tumor immune activity (Figure 6A–C).
 |
Figure 6 SYW synergizes with PD-1 Inh promotes effector CD8+ T cell activation via gut immunity to synergize with PD-1 inhibitor against NSCLC. (A) ELISA detection of IFN-γ (n=6). (B) ELISA detection of TNF-α(n=6). (C) ELISA detection of IL-2 (n=6). (D) Gating strategy and quantification of tumor-infiltrating CD8⁺ T cells (n=6), (i) Gating strategy: lymphocytes were gated on FSC/SSC to exclude debris, followed by singlet exclusion, viable cell selection, and sequential gating for CD45⁺ and CD3⁺. (ii) Representative CD8⁺ plot (gated from i) for control group. (iii) Representative CD8⁺ plot (gated from i) for L-SYW+RMP1-14 group. (iv) Representative CD8⁺ plot (gated from i) for H-SYW+RMP1-14 group. (v) Representative CD8⁺ plot (gated from i) for RMP1-14 group. (vi) Quantitative summary of CD8⁺ T cell proportions across groups. (E) Quantification of PD-1⁺ CD8⁺ T cell (n=6). (F)Quantification of TIM-3⁺ CD8⁺ T cell (n=6). (G) Quantification of IFN-γ⁺ CD8⁺ T cell (n=6). (H) Quantification of Granzyme B⁺ CD8⁺ T cell proportions (n=6). *p < 0.05 vs. control group, %p < 0.05 vs. L-SYW+RMP1-14 group, &p < 0.05 vs. H-SYW+RMP1-14 group, #p< 0.05 vs. RMP1-14 group. |
Flow cytometric analysis showed that the proportion of tumor-infiltrating CD8+ T cells was significantly increased in the combination groups, with the highest level observed in the H-SYW+RMP1-14 group (Figure 6D). In addition, the H-SYW+RMP1-14 group exhibited the lowest percentages of PD-1+ and TIM-3+ CD8+ T cells, and the highest percentages of IFN-γ+ and Granzyme B+ CD8+ T cells among all groups (Figure 6E–H).
SYW Synergizes with PD-1 Inh to Modulate the Structure of Gut Microbiota in vivo
16S rRNA sequencing of intestinal contents from NSCLC syngeneic mice showed that α-diversity (Sobs, ACE indices) was significantly higher in the H-SYW+RMP1-14 group than in the control and RMP1-14 groups, indicating that SYW regulates gut microbiota abundance and community balance (Figure 7A). β-diversity analysis revealed distinct clustering of H-SYW+RMP1-14 samples from control and RMP1-14 groups in PC1-PC2 and PC1-PC3 dimensions, reflecting overall remodeling of the microbiota community structure (Figure 7B). Violin plots of Gini and NCI indices confirmed significant differences in microbiota diversity distribution and interspecific niche interactions between the H-SYW+RMP1-14 group and the control group (Figure 7C).
 |
Figure 7 SYW synergizes with PD-1 Inh to reshape the gut microbiota structure of NSCLC mice. (A) Alpha diversity analysis, (B) Beta diversity analysis; (C) Violin plots of Gini index and NCI index, (D) Differences in microbiota composition at the phylum level, (E) Cladogram showing characteristic branches of intergroup differences, (F) COG functional classification of differential microbiota. n=6. *p < 0.05, **p < 0.01. |
At the phylum level, H-SYW+RMP1-14 significantly altered the relative abundances of Patescibacteria, Bacillota, Thermodesulfobacterdiota and Elusimcrobiota compared with the control and RMP1-14 groups, indicating targeted regulation of core microbiota (Figure 7D). Cladogram analysis identified unique evolutionary branches in the H-SYW+RMP1-14 group, suggesting driven adjustments in microbiota evolutionary structure (Figure 7E).
PICRUSt2-based COG classification showed distinct microbiota distribution in the H-SYW+RMP1-14 group in pathways related to transcription (COG1309, 1609, 0583, 1476, 1846), ABC transport (COG1132, 1131, 1136, 0488), carbohydrate, amino acid and inorganic ion transport and metabolism (COG1753, 0295, 0524, 1879, 1904, 0366, 0601). These findings indicate that SYW combined with PD-1 Inh may exert synergistic effects by reshaping microbiota structure to modulate gut microecological functions and metabolism (Table S3 and Figure 7F).
SYW Synergizes with PD-1 Inh to Regulate the Metabolites of Gut Microbiota in vivo
Untargeted metabolomic analysis of intestinal metabolites in NSCLC mice showed clear separation of H-SYW+RMP1-14 samples from control and RMP1-14 groups in 2D/3D PCA plots, and OPLS-DA confirmed intergroup differences in metabolite profiles (Figure 8A and 8C). Quantitative analysis identified numerous differential metabolites between groups (upregulated in red, downregulated in blue) (Figure 8B). Venn analysis focused on 42 common differential metabolites among H-SYW+RMP1-14, control and RMP1-14 groups (Figure 8D).
 |
Figure 8 SYW synergizes with PD-1 Inh to regulate the gut microbiota metabolite profile of NSCLC mice. (A) 2D PCA analysis of metabolites, (B) 3D PCA analysis of metabolites, (C) OPLS-DA analysis of metabolites; (D) Venn diagram of intergroup metabolite sets, (E) Hierarchical clustering analysis of differential metabolites, (F) KEGG pathway enrichment analysis of differential metabolites. (G) Network visualization of differential metabolites. n=6. |
Hierarchical clustering showed that H-SYW+RMP1-14 decreased carbohydrates (D-Gal Alpha 1->6D-Gal Alpha, fucose), phenols/flavonoids (phloretin, coniferaldehyde) and certain terpenoids (deoxyloganin), while increasing amino acids (L-glycine, 4-hydroxyproline), vitamins (panthenol, 5-methyltetrahydrofolate), peptides (carnosine, S-sulfanylglutathione), triterpenoids/steroids (oleanolic acid, stigmasterol) and specific phenols/flavonoids (isoliquiritigenin, formononetin, epigallocatechin) (Figure 8E).
KEGG enrichment highlighted pathways including plant secondary metabolite biosynthesis, amino acid metabolism and ABC transporters (Figure 8F). Network visualization identified the top 5 differential metabolites (L-glycine, L-proline, 10-formyltetrahydrofolate, anthranilic acid and 5-methyltetrahydrofolate) and pathways (ABC transporters, biosynthesis of phenylpropanoids, flavonoid biosynthesis, arginine and proline metabolism, D-amino acid metabolism) (Figure 8G).
Collectively, the combination therapy systematically reshapes the gut microbiota metabolic profile in NSCLC mice.
FMT Confirms GM-TME Crosstalk as Key Mediator of SYW-PD-1 Inh Synergism in NSCLC
Gut Microecology Mediates the Synergistic Anti-NSCLC Effect of SYW and PD-1 Inh in vivo
All mice survived during the experiment, and no significant difference in body weight was observed among the three groups (Figure 9C). In vivo bioluminescence imaging and quantitative analysis showed that tumor burden was the lowest in the N group, the highest in the ABX group, and significantly restored in the FMT group (Figure 9A and B).
 |
Figure 9 Gut microecology mediates the synergistic anti-NSCLC effect of SYW synergizes with PD-1 inhibitor in vivo. (A) In vivo bioluminescence imaging of tumor burden in each group, (B) Quantitative analysis of bioluminescence intensity (n=6), (C) Body weight changes of mice during the study (n=6), (D) Tumor volume growth curves (n=6), (E) Ex vivo tumor weight at the experimental endpoint (n=6), (F) Tumor inhibition rate calculated based on tumor weight (n=6), (G) HE staining of tumor tissues (scale bar = 200 μm, n=3). *p < 0.05 vs. N group, #p < 0.05 vs. FMT group, &p < 0.05 vs. ABX group, Δp < 0.05. |
Mixed-effects model analysis revealed a significant treatment × time interaction (F(12, 60) = 6.72, p < 0.05), indicating that tumor growth trajectories differed significantly among the N, ABX, and FMT groups. Tumor growth curves showed that the N group had the slowest growth, the ABX group grew rapidly, and the FMT group restored the inhibitory effect (Figure 9D). At the end of the experiment, tumor weight was the highest and inhibition rate was the lowest in the ABX group; both indices were significantly reversed in the FMT group and close to those in the N group (Figure 9E and F).
HE staining showed that tumors in the N and FMT groups had sparse cell arrangement and obvious apoptotic features, while the ABX group showed dense tumor cells (Figure 9G). These results indicate that the synergistic anti-tumor effect of SYW and PD-1 Inh relies on gut microbiota (GM): effective in the N group, lost in the ABX group, and restored by FMT.
Gut Microbiota Regulates PD-1/PD-L1 Expression and Co-Localization in Tumor Tissues
Immunofluorescence staining showed that PD-1 expression was the lowest in the N group, significantly increased in the ABX group, and decreased again in the FMT group (Figure 10A and B). PD-L1 expression was the highest in the N group, decreased in the ABX group, and did not recover obviously in the FMT group, which may be related to antibiotic use during antibiotic treated processing (Figure 10C).
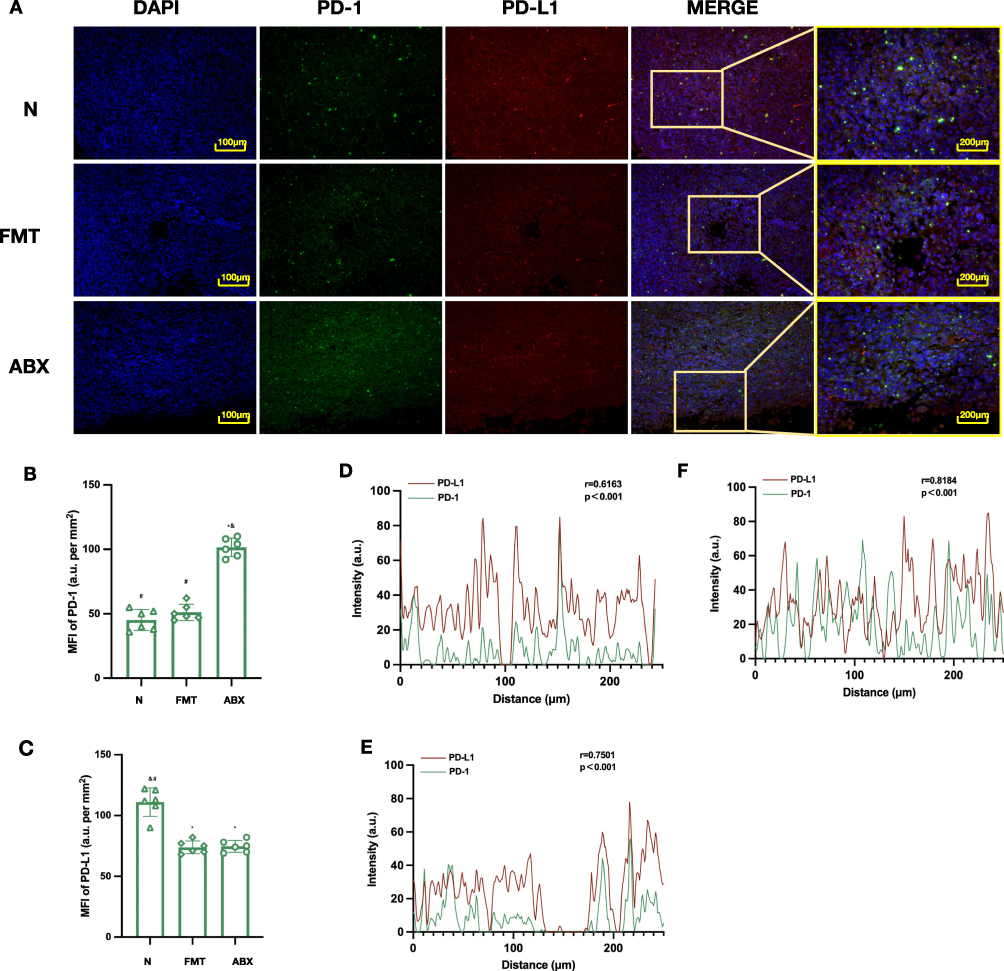 |
Figure 10 Effects of gut microbiota on PD-1/PD-L1 expression and co-localization in tumor tissues of mice treated with SYW+PD-1 inhibitor. (A) Representative immunofluorescence co-staining images of PD-1 (green), PD-L1 (red), and DAPI (blue) in tumor tissues (scale bar = 100 μm). (B) Quantitative analysis of PD-1 MFI across all groups, n=6. (C) Quantitative analysis of PD-L1 MFI across all groups, n=6. (D) Line-scan PD-1/PD-L1 signal distribution, N group. (E) Line-scan PD-1/PD-L1 signal distribution, FMT group. (F) Line-scan PD-1/PD-L1 signal distribution, ABX group. *p < 0.05 vs. N group, #p < 0.05 vs. FMT group, &p < 0.05 vs. ABX group. |
Line-scan analysis showed the strongest co-localization of PD-1 and PD-L1 in the ABX group, weaker in the FMT group, and the weakest in the N group (Figure 10D and F). These results suggest that GM is required for SYW+PD-1 Inh to regulate PD-1/PD-L1 distribution.
Gut Microbiota Mediates Immune Activation Induced by SYW Combined with PD-1 Inh
ELISA results showed that the levels of IFN-γ, TNF-α and IL-2 in tumor tissue homogenates were the highest in the N group, the lowest in the ABX group, and restored in the FMT group (Figure 11A–C). Flow cytometry showed that tumor-infiltrating CD8+ T cells were most abundant in the N group, reduced in the ABX group, and recovered in the FMT group (Figure 11D). The ABX group exhibited the highest proportion of PD-1+ and TIM-3+ exhausted CD8+ T cells, and the lowest proportion of IFN-γ+ and Granzyme B+ effector CD8+ T cells. The N group showed the opposite trend, and the FMT group restored a phenotype similar to that in the N group (Figure 11E–H).
 |
Figure 11 Effects of gut microbiota on serum cytokines and tumor-infiltrating CD8+ T cell subsets in mice treated with SYW+PD-1 inhibitor. (A) ELISA detection of IFN-γ (n=6). (B) ELISA detection of TNF-α(n=6). (C) ELISA detection of IL-2 (n=6). (D) Gating strategy and quantification of tumor-infiltrating CD8⁺ T cells (n=6). (i) Representative CD8⁺ plot for N group. (ii) Representative CD8⁺ plot for FMT group. (iii) Representative CD8⁺ plot for ABX group. (iv) Quantitative summary of CD8⁺ T cell proportions across groups. (E) Quantification of PD-1⁺ CD8⁺ T cell (n=6). (F)Quantification of TIM-3⁺ CD8⁺ T cell (n=6). (G) Quantification of IFN-γ⁺ CD8⁺ T cell (n=6). (H) Quantification of Granzyme B⁺ CD8⁺ T cell proportions (n=6). *p < 0.05 vs. N group, #p < 0.05 vs. FMT group, &p < 0.05 vs. ABX group. |
Discussion
This study adopted a stepwise “mechanism mining-in vivo efficacy validation-pathway confirmation” strategy to investigate the synergistic anti-NSCLC effect of SYW combined with a PD-1 inhibitor. Integrating bioinformatics, multi-omics analyses, and functional validation, we uncovered that SYW, despite lacking direct antitumor activity, dose-dependently potentiates PD-1 blockade efficacy by remodeling the gut microbiota, altering immune-related metabolites, and enhancing effector CD8+T cell infiltration to ameliorate the TME. FMT experiments further established that an intact gut microbiota is indispensable for this synergy, identifying GM-TME crosstalk as the core mediator of SYW-ICI synergy in NSCLC. The comprehensive regulatory mechanism is summarized and visually presented in Figure 12.
 |
Figure 12 SYW enhances PD-1 inhibitor efficacy against NSCLC by regulating GM-TME crosstalk. |
The pathological complexity of NSCLC renders single-target interventions insufficient, whereas the multi-component nature of TCM formulas offers distinct advantages. Using UPLC-MS and database mining, we identified 185 SYW-derived compounds and 264 corresponding targets. Integration with GEO NSCLC transcriptomic data yielded 200 overlapping targets, which were enriched in immune-related pathways including “PD-L1 expression and PD-1 checkpoint pathway in cancer”, “T cell receptor signaling pathway”, and “Chemokine signaling pathway”. Molecular docking further confirmed strong binding affinities (≤ −7 kcal/mol)19 between core SYW components and key targets. These in silico findings not only substantiate the multi-target potential of SYW but also point toward the gut microbiota as a pivotal hub bridging TCM components and host immunity.
Notably, our in vivo data revealed a striking pattern: SYW monotherapy did not significantly differ from the control group across all measured parameters—tumor bioluminescence, volume, weight, and histopathology—whereas its combination with PD-1 inhibitor produced dose-dependent tumor suppression. This dissociation between monotherapy inefficacy and combination potency is mechanistically informative. It indicates that SYW does not function as a direct cytotoxic agent against NSCLC cells, but rather acts as an immune modulator that creates a permissive milieu for PD-1 blockade. Immunofluorescence analysis further clarified that the enhanced anti-tumor effect of the combination was not achieved by directly augmenting PD-1 inhibition. Instead, SYW plus PD-1 inhibitor reduced PD-1 expression and increased PD-L1 expression in tumor tissues, while markedly decreasing PD-1/PD-L1 co-localization. This pattern is consistent with increased infiltration of effector T cells, which themselves express PD-1, yet the net outcome of reduced co-localization reflects a functional disruption of the immunosuppressive axis. Therefore, the synergistic benefit arises from immune microenvironment remodeling—mediated by GM-derived signals—rather than from intensified checkpoint blockade.
Persistent antigen stimulation drives CD8+ T cells into a state of exhaustion (Tex), characterized by co-expression of multiple inhibitory receptors and metabolic dysfunction, which substantially limits the efficacy of PD-1/PD-L1 blockade.20 Moreover, ICI-based combinations frequently provoke immune-related adverse events (irAEs), underscoring the need for safer synergistic strategies.21 Our flow cytometric analysis demonstrated that the combination therapy significantly increased tumor-infiltrating CD8+ T cells, with a preferential expansion of functional effector subsets (IFN-γ+ and Granzyme B+) and a concomitant reduction in exhausted subsets (PD-1+ and TIM-3+). Concurrently, serum levels of IL-12, IFN-γ and TNF-α were elevated. IL-12 bridges innate and adaptive immunity by promoting Th1 differentiation and enhancing the cytotoxic activity of CD8+T cells and NK cells.22 IFN-γ signaling is intimately linked to ICI responsiveness and MHC-I-mediated antigen presentation.23,24 TNF-α induces tumor cell apoptosis via TNFR1-mediated death receptor signaling,25 while disrupting tumor vasculature to promote CD8+ T cell infiltration and activating NF-κB to enhance dendritic cell maturation and M1 macrophage polarization.26 Collectively, these data indicate that SYW synergizes with PD-1 blockade not by directly intensifying checkpoint inhibition, but by systemically reshaping the host immune landscape to favor effector T cell infiltration and function.
A growing body of evidence supports the concept of GM-TME crosstalk in modulating tumor immunity.27 Based on bioinformatics predictions and in vivo efficacy verification, 16S rRNA sequencing of intestinal contents revealed distinct alterations in the composition and dominant bacterial taxa of the gut microbiota in the H-SYW+PD-1 Inh group. The most notable changes were an increase in Bacillota abundance and a decrease in Patescibacteria abundance. Bacillota, a major phylum of Gram-positive bacteria,28 produces metabolites with diverse immunomodulatory and anti-tumor activities. For instance, cyclic peptides such as bacillistatins 1 and 2, halobacillin, and mixirins A-C can directly inhibit tumor cell proliferation by interfering with the cell cycle and inducing apoptosis,29,30 while 7-O-succinyl macrolactin A modulates TNF-α and IL-6 secretion and regulates mTOR and NF-κB activation.31 Short-chain fatty acids derived from Bacillota maintain intestinal barrier integrity and restrain excessive immune responses; disruptions in their abundance may contribute to immune escape or autoimmunity.32 Patescibacteria is a recently defined, largely uncultured phylum often referred to as “microbial dark matter”.33,34 Its subordinate clade Saccharibacteria can inhibit phage receptor expression in host bacteria, thereby reducing phage susceptibility and indirectly stabilizing the microbial community. This “host protection” effect suggests a potential role for Patescibacteria in mucosal immune regulation via microbiota-TME crosstalk.35
PICRUSt2 functional prediction of differential bacteria indicated that the H-SYW+RMP1-14 group displayed distinct distributions in functional pathways related to transcription, transport, signal transduction, and amino acid and carbohydrate metabolism. These differences were primarily associated with transcriptional regulators, ABC transporters, and the transport and metabolism of carbohydrates, amino acids, and inorganic ions, consistent with the trends observed in untargeted metabolomics. KEGG pathway analysis combined with network topology evaluation highlighted several differential metabolites in the combination group—including L-glycine, L-proline, 10-formyltetrahydrofolate, anthranilic acid, and 5-methyltetrahydrofolate—and differential pathways such as ABC transporters, biosynthesis of phenylpropanoids, flavonoid biosynthesis, arginine and proline metabolism, and D-amino acid metabolism. These metabolites and pathways are intimately linked to tumor immunity. L-glycine promotes CD8+ T cell infiltration by modulating T cell metabolism,36 whereas excessive accumulation of L-proline can drive collagen deposition and tumor stromal fibrosis, thereby impeding CD8+ T cell access.37 Folate derivatives (10-formyltetrahydrofolate and 5-methyltetrahydrofolate) serve as one-carbon donors that support purine and pyrimidine synthesis, fueling tumor proliferation.38 Anthranilic acid participates in the kynurenine pathway of tryptophan metabolism; alterations in its levels may reduce IDO1-mediated immunosuppressive metabolites and influence T cell exhaustion.39 ABC transporters constitute a superfamily of proteins that hydrolyze ATP to transport diverse biomolecules; in cancer, they affect drug resistance, invasion, and the immune microenvironment by regulating metabolite flux across membranes.40,41 For example, ABCC6 may enhance the invasive phenotype of HepG2 cells by promoting epithelial-mesenchymal transition,42 and ABCC5 expression is negatively correlated with K-Ras mutations in lung cancer cells upon PD-L1 inhibition.43 Furthermore, D-amino acid modifications have been exploited to screen CD24/Siglec-10 blocking peptides, which represent the first peptides capable of simultaneously blocking PD-1/PD-L1 interaction, thereby enhancing macrophage- and M-MDSC-mediated phagocytosis of tumor cells and improving CD8+T cell activity.44 Collectively, these metabolites constitute a microbiota-derived metabolic network that communicates with the TME and serves as an important medium through which SYW remodels the immune landscape. FMT experiments provided causal evidence for the role of gut microbiota in SYW-PD-1 inhibitor synergy. In ABX mice, the tumor-inhibiting effect of SYW+PD-1 Inh was substantially diminished, accompanied by reduced Teff cell infiltration and attenuated tumor apoptosis. In contrast, transplantation of fecal microbiota from healthy donors restored therapeutic efficacy to levels comparable with those observed in conventional mice receiving the same combination treatment. Nevertheless, our work is limited to CD8+ T cell-focused immune profiling and preclinical murine models. The specific keystone bacterial taxa and downstream effector metabolites driving the immune-sensitizing activity remain to be fully elucidated; follow-up studies will further dissect these microbial and metabolic mediators, alongside expanded characterization of the tumor immune landscape and clinical translational validation. Taken together, and that GM–TME crosstalk is a potential core mechanism underlying this synergy, providing preclinical evidence supporting SYW as a promising microbiota-targeted adjunct to PD-1 blockade in NSCLC.
Conclusion
Through multi-component and multi-target regulation, SYW enriches beneficial gut microbiota (elevated Bacillota, reduced Patescibacteria) and modulates characteristic metabolites such as L-glycine and L-proline, thereby shaping a gut microecological milieu that enhances PD-1 pathway blockade and effector CD8+T cell infiltration to ameliorate the tumor immune microenvironment. Critically, SYW itself lacks direct anti-NSCLC activity and instead functions as a microbiota-dependent immune sensitizer that potentiates PD-1 inhibitor efficacy. FMT experiments confirm that an intact gut microbiota is indispensable for this synergistic effect. Employing a progressive “mechanism exploration - efficacy verification - pathway confirmation” strategy, this study is the first to identify GM-TME crosstalk as the potential core mediator of SYW-ICI synergy in NSCLC. These findings provide pharmacological evidence for SYW-based combination immunotherapy and offer a novel integrative TCM-Western medicine approach to address immune resistance in NSCLC.
Abbreviations
ABX, Antibiotics; ADME, Absorption, Distribution, Metabolism and Excretion; BP, Biological processes; CC, Cellular components; DC, Dendritic cell; DL, Drug-likeness; FMT, Fecal Microbiota Transplantation; GEO, Gene Expression Omnibus; GO, Gene Ontology; GM, Gut microecology; ABX, antibiotic treated; HL, Half-Life; IPA, Indole-3-propionic acid; IL-2, Interleukin-2; IFN-γ, Interferon-γ; ICI, Immune checkpoint inhibitors; IrAEs, Immune-related adverse events; KEGG, Kyoto Encyclopedia of Genes and Genomes; LLC, Lewis lung cancer; NSCLC, Non-small cell lung cancer; MF, Molecular functions; OB, Oral bioavailability; PD-1, Programmed cell death protein 1; PD-L1, Programmed death-ligand 1;PD-1 Inh, PD-1 inhibitors; SYW, Shuyu Wan; TME, Tumor immune microenvironment; Teff, Effector T cells; Tex, Exhausted cytotoxic T cells; Tregs, Regulatory T cells; TCM, Traditional Chinese medicine; TCMSP, Traditional Chinese Medicine Systems Pharmacology Database and Analysis Platform; TIR, Tumor inhibition rate; TNF-α,Tumor necrosis factor-α; UPLC-MS, Ultra-Performance Liquid Chromatography-Mass Spectrometry.
Data Sharing Statement
All data are presented in the manuscript and attachments.
Ethics Statement
Animal experiments were approved by the Animal Ethics Committee of the Affiliated Hospital of Nanjing University School of Medicine (Approval No. 2025AE01047). All procedures adhered to NIH laboratory animal care guidelines.
Funding
This research was supported by the Natural Science Foundation of Jiangsu Province (Youth Project, Grant number:BK20250243; General Program, Grant number: BK20231123), the National Natural Science Foundation of China (Youth Project, Grant number:82505305; General Program, Grant number: 82374364), the Jiangsu Provincial Program for Cultivating Leading Talents in Traditional Chinese Medicine (Grant number: SLJ0301), the Nanjing Medical Science and Technology Development Project (Grant number: ZKX24015), the Clinical Research Special Fund of Nanjing Drum Tower Hospital (Grant number: 2024-LCYJ-ZXY-01), the Nanjing Famous TCM Doctor Studio Construction Project (Grant number: CZXM2024021), Entrusted Development Project by Lunan Hopeful Pharmaceutical Co., Ltd. and Nanjing Drum Tower Hospital (GL20252960),Jiangsu Province Traditional Chinese Medicine Climbing Plan Project (PDJH2026033) and the Youth Project of Open Research Fund for Integrated Chinese and Western Medicine Collaboration of Institute of Chinese Medicine, Nanjing University (Grant number: ICM2024030).
Disclosure
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
1. Siegel RL, Kratzer TB, Giaquinto AN, Sung H, Jemal A. Cancer statistics, 2025. CA Cancer J Clin. 2025;75(1):10–24. doi:10.3322/caac.21871
2. Herbst RS, Morgensztern D, Boshoff C. The biology and management of non-small cell lung cancer. Nature. 2018;553(7689):446–454. doi:10.1038/nature25183
3. Geng Q, Xu J, Du C, et al. Small molecules targeting immune checkpoint proteins for cancer immunotherapy: a patent and literature review (2020–2024). Expert Opin Ther Pat. 2025;35(4):409–440. doi:10.1080/13543776.2025.2462849
4. Zhao L, Li T, Zhou Y, Wang P, Luo L. Monoclonal antibody targeting CEACAM1 enhanced the response to anti-PD1 immunotherapy in non-small cell lung cancer. Int Immunopharmacol. 2024;143:113395. doi:10.1016/j.intimp.2024.113395
5. Zhou H, Zhang H, Shi M, Wang J, Huang Z, Shi J. A robust signature associated with patient prognosis and tumor immune microenvironment based on immune-related genes in lung squamous cell carcinoma. Int Immunopharmacol. 2020;88:106856. doi:10.1016/j.intimp.2020.106856
6. Riely GJ, Wood DE, Ettinger DS, et al. Non-Small cell lung cancer, version 4.2024, NCCN clinical practice guidelines in oncology. J Natl Compr Canc Netw. 2024;22(4):249–274. doi:10.6004/jnccn.2204.0023
7. Dang TO, Ogunniyi A, Barbee MS, Drilon A. Pembrolizumab for the treatment of PD-L1 positive advanced or metastatic non-small cell lung cancer. Expert Rev Anticancer Ther. 2016;16(1):13–20. doi:10.1586/14737140.2016.1123626
8. Haslam A, Prasad V. Estimation of the percentage of US patients with cancer who are eligible for and respond to checkpoint inhibitor immunotherapy drugs. JAMA Network Open. 2019;2(5):e192535. doi:10.1001/jamanetworkopen.2019.2535
9. Mok TSK, Wu YL, Kudaba I, et al. Pembrolizumab versus chemotherapy for previously untreated, PD-L1-expressing, locally advanced or metastatic non-small-cell lung cancer (KEYNOTE-042): a randomised, open-label, controlled, Phase 3 trial. Lancet. 2019;393(10183):1819–1830. doi:10.1016/s0140-6736(18)32409-7
10. Murata D, Azuma K, Murotani K, et al. Characterization of pre- and on-treatment soluble immune mediators and the tumor microenvironment in NSCLC patients receiving PD-1/L1 inhibitor monotherapy. Cancer Immunol Immunother. 2024;73(11):214. doi:10.1007/s00262-024-03781-8
11. Lin A, Zheng K, Jiang A, et al. The evolving landscape of immunotoxicity: charting mechanisms and future strategies for immune checkpoint inhibitor adverse events. Med Res. 2025;1(3):322–358. doi:10.1002/mdr2.70019
12. Gazzaniga FS, Kasper DL. The gut microbiome and cancer response to immune checkpoint inhibitors. J Clin Invest. 2025;135(3). doi:10.1172/jci184321
13. Jia D, Wang Q, Qi Y, et al. Microbial metabolite enhances immunotherapy efficacy by modulating T cell stemness in pan-cancer. Cell. 2024;187(7):1651–1665.e21. doi:10.1016/j.cell.2024.02.022
14. Lin NY, Fukuoka S, Koyama S, et al. Microbiota-driven antitumour immunity mediated by dendritic cell migration. Nature. 2025. doi:10.1038/s41586-025-09249-8
15. Deng Z, Teng YJ, Zhou Q, et al. Shuyu pills inhibit immune escape and enhance chemosensitization in hepatocellular carcinoma. World J Gastrointest Oncol. 2021;13(11):1725–1740. doi:10.4251/wjgo.v13.i11.1725
16. Huang J, Liu D, Wang Y, et al. Ginseng polysaccharides alter the gut microbiota and kynurenine/tryptophan ratio, potentiating the antitumour effect of antiprogrammed cell death 1/programmed cell death ligand 1 (anti-PD-1/PD-L1) immunotherapy. Gut. 2022;71(4):734–745. doi:10.1136/gutjnl-2020-321031
17. Zhang G, Pan J, Xu X, et al. Chinese yam polysaccharide enhances anti-PD-1 immunotherapy in colorectal cancer through alterations in the gut microbiota and metabolites. Int J Biol Macromol. 2025;310(Pt 2):143323. doi:10.1016/j.ijbiomac.2025.143323
18. Chen Y, Huang M, Wang H, Sun G, Li P, Guo Z. Effects of modified shuyuwan on gut microbiota and HMGB1 pathway-related proteins in Alzheimer’s mice. Pharmacol Clin Chin Mater Med. 2023;39(3):15–19. doi:10.13412/j.cnki.zyyl.20230202.001
19. Paggi JM, Pandit A, Dror RO. The art and science of molecular docking. Annu Rev Biochem. 2024;93:389–410. doi:10.1146/annurev-biochem-030222-120000
20. Andrews LP, Butler SC, Cui J, et al. LAG-3 and PD-1 synergize on CD8(+) T cells to drive T cell exhaustion and hinder autocrine IFN-γ-dependent anti-tumor immunity. Cell. 2024;187(16):4355–4372.e22. doi:10.1016/j.cell.2024.07.016
21. Blum SM, Rouhani SJ, Sullivan RJ. Effects of immune-related adverse events (irAEs) and their treatment on antitumor immune responses. Immunol Rev. 2023;318(1):167–178. doi:10.1111/imr.13262
22. Allen GM, Frankel NW, Reddy NR, et al. Synthetic cytokine circuits that drive T cells into immune-excluded tumors. Science. 2022;378(6625):eaba1624. doi:10.1126/science.aba1624
23. Grasso CS, Tsoi J, Onyshchenko M, et al. Conserved interferon-γ signaling drives clinical response to immune checkpoint blockade therapy in melanoma. Cancer Cell. 2020;38(4):500–515.e3. doi:10.1016/j.ccell.2020.08.005
24. Ren J, Li N, Pei S, et al. Histone methyltransferase WHSC1 loss dampens MHC-I antigen presentation pathway to impair IFN-γ-stimulated antitumor immunity. J Clin Invest. 2022;132(8). doi:10.1172/jci153167
25. Wang L, Du F, Wang X. TNF-α induces two distinct caspase-8 activation pathways. Cell. 2008;133(4):693–703. doi:10.1016/j.cell.2008.03.036
26. Huyghe J, Priem D, Bertrand MJM. Cell death checkpoints in the TNF pathway. Trends Immunol. 2023;44(8):628–643. doi:10.1016/j.it.2023.05.007
27. Fernandes MR, Aggarwal P, Costa RGF, Cole AM, Trinchieri G. Targeting the gut microbiota for cancer therapy. Nat Rev Cancer. 2022;22(12):703–722. doi:10.1038/s41568-022-00513-x
28. Chukwudulue UM, Barger N, Dubovis M, Luzzatto Knaan T. Natural products and pharmacological properties of symbiotic bacillota (Firmicutes) of marine macroalgae. Mar Drugs. 2023;21(11). doi:10.3390/md21110569
29. Pettit GR, Knight JC, Herald DL, et al. Antineoplastic agents. 570. Isolation and structure elucidation of bacillistatins 1 and 2 from a marine bacillus silvestris. J Nat Prod. 2009;72(3):366–371. doi:10.1021/np800603u
30. Zhang HL, Hua HM, Pei YH, et al. Three new cytotoxic cyclic acylpeptides from marine bacillus sp. Chem Pharm Bull. 2004;52(8):1029–1030. doi:10.1248/cpb.52.1029
31. Park S, Regmi SC, Park SY, et al. Protective effect of 7-O-succinyl macrolactin A against intestinal inflammation is mediated through PI3-kinase/Akt/mTOR and NF-κB signaling pathways. Eur J Pharmacol. 2014;735:184–192. doi:10.1016/j.ejphar.2014.04.024
32. Xiao S, Chen N, Chai Z, et al. Secondary metabolites from marine-derived bacillus: a comprehensive review of origins, structures, and bioactivities. Mar Drugs. 2022;20(9). doi:10.3390/md20090567
33. Wang Y, Gallagher LA, Andrade PA, et al. Genetic manipulation of Patescibacteria provides mechanistic insights into microbial dark matter and the epibiotic lifestyle. Cell. 2023;186(22):4803–4817.e13. doi:10.1016/j.cell.2023.08.017
34. Srinivas P, Peterson SB, Gallagher LA, Wang Y, Mougous JD. Beyond genomics in Patescibacteria: a trove of unexplored biology packed into ultrasmall bacteria. Proc Natl Acad Sci U S A. 2024;121(51):e2419369121. doi:10.1073/pnas.2419369121
35. Zhong Q, Liao B, Liu J, et al. Episymbiotic Saccharibacteria TM7x modulates the susceptibility of its host bacteria to phage infection and promotes their coexistence. Proc Natl Acad Sci U S A. 2024;121(16):e2319790121. doi:10.1073/pnas.2319790121
36. Tong H, Jiang Z, Song L, et al. Dual impacts of serine/glycine-free diet in enhancing antitumor immunity and promoting evasion via PD-L1 lactylation. Cell Metab. 2024;36(12):2493–2510.e9. doi:10.1016/j.cmet.2024.10.019
37. Tharp KM, Kersten K, Maller O, et al. Tumor-associated macrophages restrict CD8(+) T cell function through collagen deposition and metabolic reprogramming of the breast cancer microenvironment. Nat Cancer. 2024;5(7):1045–1062. doi:10.1038/s43018-024-00775-4
38. Abdelaziz OA, El Husseiny WM, Selim KB, Eisa HM. Dihydrofolate reductase inhibition effect of 5-substituted pyrido[2,3-d]pyrimidines: synthesis, antitumor activity and molecular modeling study. Bioorg Chem. 2019;90:103076. doi:10.1016/j.bioorg.2019.103076
39. Gouasmi R, Ferraro-Peyret C, Nancey S, et al. The kynurenine pathway and cancer: why keep it simple when you can make it complicated. Cancers. 2022;14(11). doi:10.3390/cancers14112793
40. Muriithi W, Macharia LW, Heming CP, et al. ABC transporters and the hallmarks of cancer: roles in cancer aggressiveness beyond multidrug resistance. Cancer Biol Med. 2020;17(2):253–269. doi:10.20892/j.issn.2095-3941.2019.0284
41. Gerard L, Duvivier L, Fourrez M, et al. Identification of two novel heterodimeric ABC transporters in melanoma: ABCB5β/B6 and ABCB5β/B9. J Biol Chem. 2024;300(2):105594. doi:10.1016/j.jbc.2023.105594
42. Matera I, Miglionico R, Abruzzese V, et al. A regulator role for the ATP-Binding cassette subfamily C member 6 transporter in HepG2 cells: effect on the dynamics of cell-cell and cell-matrix interactions. Int J Mol Sci. 2023;24(22):16391. doi:10.3390/ijms242216391
43. Unver N, Tavukcuoglu E, Esendagli G. Tailored modulation of stemness and drug resistance marker characteristics in K-Ras mutant lung cancer cells via PD-L1 gene suppression. Life Sci. 2022;311(Pt B):121171. doi:10.1016/j.lfs.2022.121171
44. Shen W, Shi P, Dong Q, et al. Discovery of a novel dual-targeting D-peptide to block CD24/Siglec-10 and PD-1/PD-L1 interaction and synergize with radiotherapy for cancer immunotherapy. J Immunother Cancer. 2023;11(6). doi:10.1136/jitc-2023-007068
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.