Back to Journals » OncoTargets and Therapy » Volume 12
rs187960998 polymorphism in miR-211 prevent development of human colon cancer by deregulation of 3’UTR in CHD5
Authors Zhu L, Wang R, Zhang L, Zuo C, Zhang R, Zhao S
Received 20 July 2018
Accepted for publication 16 November 2018
Published 3 January 2019 Volume 2019:12 Pages 405—412
DOI https://doi.org/10.2147/OTT.S180935
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Sanjeev K. Srivastava
Limei Zhu,* Ran Wang,* Li Zhang, Chunlei Zuo, Rui Zhang, Shaolin Zhao
Department of Clinical Laboratory, The First People’s Hospital of Lianyungang, Lianyungang, Jiangsu, China
*These authors contributed equally to this work
Background: Previous research indicated that overexpression of miRNA-211 could promote colorectal cancer cell growth by targeting tumor suppressive gene Chromodomain-helicase-DNA-binding protein 5 (CHD5) in human colon cancer (CC). Moreover, the function of the single-nucleotide polymorphism (SNP) located in the mature region of miR-211 has not been investigated. In this study, we found that SNP of rs187960998 in miR-211 was involved in the occurrence of CC by acting as a tumor suppressor by mal-regulation of its target gene CHD5.
Materials and methods: The genotype of total 685 CC patients was detected by real-time PCR, the proliferation of CC cell lines with different genotypes of miR-211 was determined by Cell Counting Kit-8, cell invasion evaluated by transwell and the activity of the CHD5 promoter in CC cell lines transfected with different miR-211 was determined by luciferase assay. The expression of CHD5 in CC patients was determined by the immunohistochemistry, and the relapse-free survival rate was analyzed by Kaplan–Meier analysis.
Results: C/T SNP of miR-211 could inhibit CC cell proliferation and invasion by upregulation of CHD5. And SNP in rs187960998 of miR-211 was associated with tumor size, metastasis and tumor differentiation in CC patients. Patients with CC genotype have significantly low CHD5 expression than the T-carrier, while no significant expression difference in miR-211 expression among different genotype subsets. Patients with CC genotype have significantly shorter postsurgery survival rate compared to the T-carrier.
Conclusion: rs187960998 in miR-211 was highly associated with a decreased risk of CC in the Chinese population by deregulating a tumor suppressive gene CHD5.
Keywords: miR-211, colon cancer, CHD5, SNP, survival
Introduction
Risk factors for colon cancer (CC) include lifestyle, older age and inherited genetic disorders.1 Other risk factors include western-style diet, smoking, alcohol, lack of physical activity, etc. Also, some of the genetic studies revealed that the single-nucleotide polymorphism (SNP) was seriously related to CC occurrence providing a reasonable explanation for individual differences of the CC occurrence.1
Activation of oncogenes and inactivation and tumor suppression genes was one of the classical hallmarks of cancer. The related mechanisms were always a core object in cancer research. Previous research indicated that Chromodomain-helicase-DNA-binding protein 5 (CHD5) which is located on 1p36 in a region of frequent deletion in neuroblastomas. CHD5 is the fifth member of a family of chromatin remodeling proteins, and it probably functions by forming a nucleosome remodeling and deacetylation complex that regulates transcription of particular genes. Existing evidence have partially explained the reasons for downregulation of CHD5 in human cancer, including aberrant methylation and regulation of miRNA. Similarly, in human CC, CHD5 was also downregulated because of CpG island hypermethylation and upregulation of miRNAs such as miR-211.2–4
miRNAs are small non-coding RNA molecules that regulate gene expression by binding to partially complementary recognition sequences of a certain mRNAs.5,6 The binding affinity between miRNA and 3′UTR of target genes was essential in the effect of miRNA. SNP in both 3′UTR and mature miRNA might have effects on the binding affinity. Unlike the somatic mutations, SNP is a genetical factor which can well interpret the individual difference in the occurrence of certain cancer. Besides the SNP in 3′UTR which we previously reported,7 another novel class of functional polymorphisms termed miRSNPs/polymorphisms were reported and defined as a polymorphism present at or near miRNA-binding sites of functional genes that can affect gene expression by interfering with a miRNA function.8 In this study, we found an SNP (rs187960998) in the mature form of miR-211 which might have an effect on the occurrence and clinical characteristic of CC in the angle of individual difference. Therefore, we carried out the study to investigate the association between the candidate SNP in miR-211 and susceptibility to CC and its precursor in Chinese based on the crucial roles previously reported. Meanwhile, we also investigated whether the risk-associated polymorphism contributes toward CC patients’ outcome and the specific mechanism involved.
Materials and methods
Clinical sample information
The case–control study consists of 685 CC patients and 618 cancer-free controls. All the participants were selected from The First People’s Hospital of Lianyungang between January 2012 and November 2016. Patients with other previous history of cancers and chemotherapy were excluded. The cancer-free control subjects from the same geographic area showed no evidence of a genetic relationship with the cases. This study was approved by the Ethics Review Board of the first people’s Hospital of Lianyungang, and every patient gave written informed consent form. The study was carried out in accordance with the principles of the Declaration of Helsinki. The clinical features of all the cases and controls were presented in Table 1.
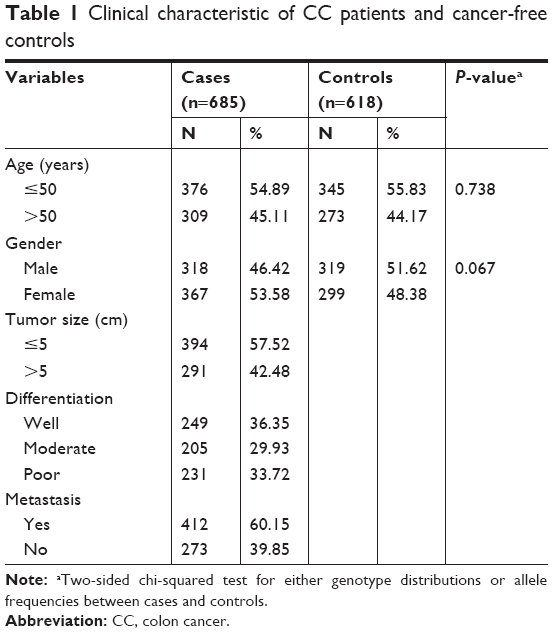 | Table 1 Clinical characteristic of CC patients and cancer-free controls |
Cell lines and cell culture
CC cell lines including Hct-116 and SW-480 were purchased from American Type Culture Collection. All cells were cultured in DMEM purchased from Thermo Fisher (Carlsbad, CA, USA) supplemented with 10% FBS (Thermo Fisher) and maintained in humidified 5% CO2 at 37°C.
Construction of luciferase-based reporter plasmids
The total fragment of the CHD5 3′UTR was amplified. The PCR production was sub-cloned into the pGL3-promoterless luciferase-based plasmid (Promega, Madison, WI, USA). The construction containing both CC and TT genotype of miR-211 was also synthesized and cloned into a pSilence 2.1-U6 plasmid in Genscript Co. (Nanjing, China).
Dual-luciferase reporter assay
The cells were harvested 48 hours after the miRNA treatment, and the activity of firefly luciferase from each group was measured and normalized to Renilla activities according to the instruction of the manufacturer (Promega) with a Fluoroskan microplate reader (Thermo Fisher Scientific, Waltham, MA, USA). The experiment was repeated thrice independently.
Cell proliferation assays
Cell proliferation was determined by using a Cell Counting Kit-8 kit according to the manufacturer’s instructions (Dojin Laboratories, Kumamoto, Japan). Briefly, the control and infected cells were seeded at a density of 1×103 cells/well in 96-well plates. Cell Counting Kit-8 was added to each well containing 100 μL of the culture medium, and the plate was incubated at 37°C for 5 days. The viability of cells was evaluated by measuring the absorbance at 450 nm, using a microplate reader (Thermo Fisher).
Genotype
Genomic DNA was extracted from peripheral mononuclear blood cells by using QIAamp DNA blood mini kits (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. Genotyping was performed with the TaqMan SNP Genotyping Assay. The PCR reactions were carried out in a total volume of 5 μL containing TaqMan Universal Master Mix, SNP Genotyping AssayMix, DNase-free water and genomic DNA. The 384-well ABI 7900HT Real-Time PCR System was applied (ABI, St. Louis, MO, USA).
Western blot
For Western blotting, proteins were extracted from the cultured cells using radioimmunoprecipitation assay buffer containing phenylmethanesulfonylfluoride (Beyotime, Nantong, China). An equal amount of proteins (100 μg) were separated with 7.5% SDS-PAGE and transferred to polyvinylidene fluoride membrane. Primary polyclonal antibodies targeting CHD5 (ab192373) and GAPDH were purchased from Abcam (Cambridge, MA, USA). The secondary antibodies were anti-rabbit or anti-mouse HRP-linked and purchased from Santa Cruz Biotechnology (Dallas, TX, USA). The blots were developed using ECL reagent (Millipore Sigma, Burlington, MA, USA).
Cell invasion assay
For transwell assay, the chamber was treated with Matrigel (Corning, CA, USA), 100 μL cell suspension (5×104) with serum-free medium was seeded to the chamber for 48 hours, cells were stained with crystal violet staining solution (Beyotime). Migrated cells were counted by using Image-pro Plus 6.0, while cell numbers of the normal control group were normalized to 1.
Immunohistochemistry
Sections were pretreated according to the previous publication.7 The section was incubated with primary mouse anti-human antibodies for CHD5 (ab192373), the sections were stained with 3,3′-Diaminobenzidine according to the manufacturer’s protocols and mounted and photographed using a digitalized microscope camera (Nikon, Tokyo, Japan).
Statistical analysis
All experiments were performed in triplicate and repeated at least three times independently. Data were presented as mean ± SD. The association between rs187960998 genotypes and the risk of CC was evaluated by calculating the ORs and their 95% CIs using univariate and multivariate logistic regression analysis. Differences between two independent groups were tested with the Student’s t-test. All statistical analyses were carried out using SPSS version 18.0 and presented with GraphPad Prism software. Kaplan–Meier survival curves were generated, and the log-rank test was done. The significance of various variables for survival was analyzed by the Cox proportional hazards model in a multivariate analysis. The results were considered to be statistically significant at P<0.05.
Results
rs187960998 clinical significance in human CC
The genotype frequencies in 685 CC cases and 618 healthy controls were investigated and characteristics are listed in Table 1. As listed in Table 2, logistic regression analysis results revealed that the CT genotype and TT genotype presented a significantly decreased risk compared with CC genotype, respectively. Besides, the T-carrier also indicated a decreased risk (P<0.0001). All ORs were adjusted for age and gender.
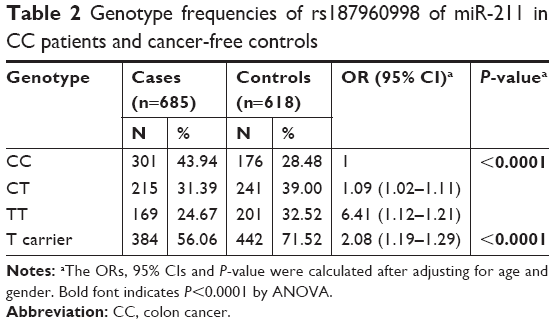 | Table 2 Genotype frequencies of rs187960998 of miR-211 in CC patients and cancer-free controls |
Stratified analysis of the correlation between miR-211 SNP and CC
Next, we conducted the stratified analysis to investigate the correlation between the SNP rs187960998 in miR-211 with the different clinical characteristics of CC (Table 3). We found a significant association between rs187960998 genotypes (C>T) with the smaller tumor size (<5 cm), non-metastasis and well-differentiation degree. Furthermore, T-carrier was also statistically related to smaller tumor size, non-metastasis and better differentiation.
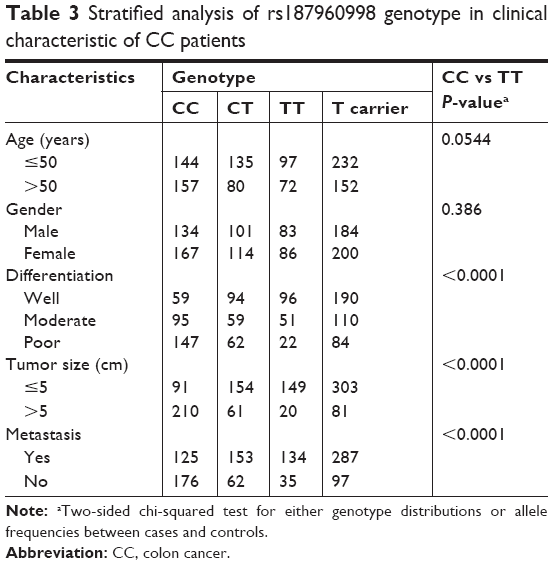 | Table 3 Stratified analysis of rs187960998 genotype in clinical characteristic of CC patients |
miR-211 SNP rs187960998 (C>T) regulates CHD5-related cell proliferation and invasion in vitro
Since the SNP rs187960998 was located in the binding site of miR-211 on 3′UTR of CHD5, and the previous study revealed that the expression of CHD5 could be regulated by miR-211.4 Therefore, to test whether this SNP can impact on the expression of CHD5 and cause related biological change, we first detected the cell proliferation by treated cells with the 3′UTR regulated CHD5 as well as miR-211 with different genotypes including CC and TT. We found that cell proliferation of both cell lines (Hct-116 and SW-480) can be inhibited by the overexpression of CHD5. However, the cell proliferation can be restored with the transfection of miR-211. However, the proliferation restoration capability was stronger in CC type of miR-211 than TT in both Hct-116 and SW-480 (Figure 1A).
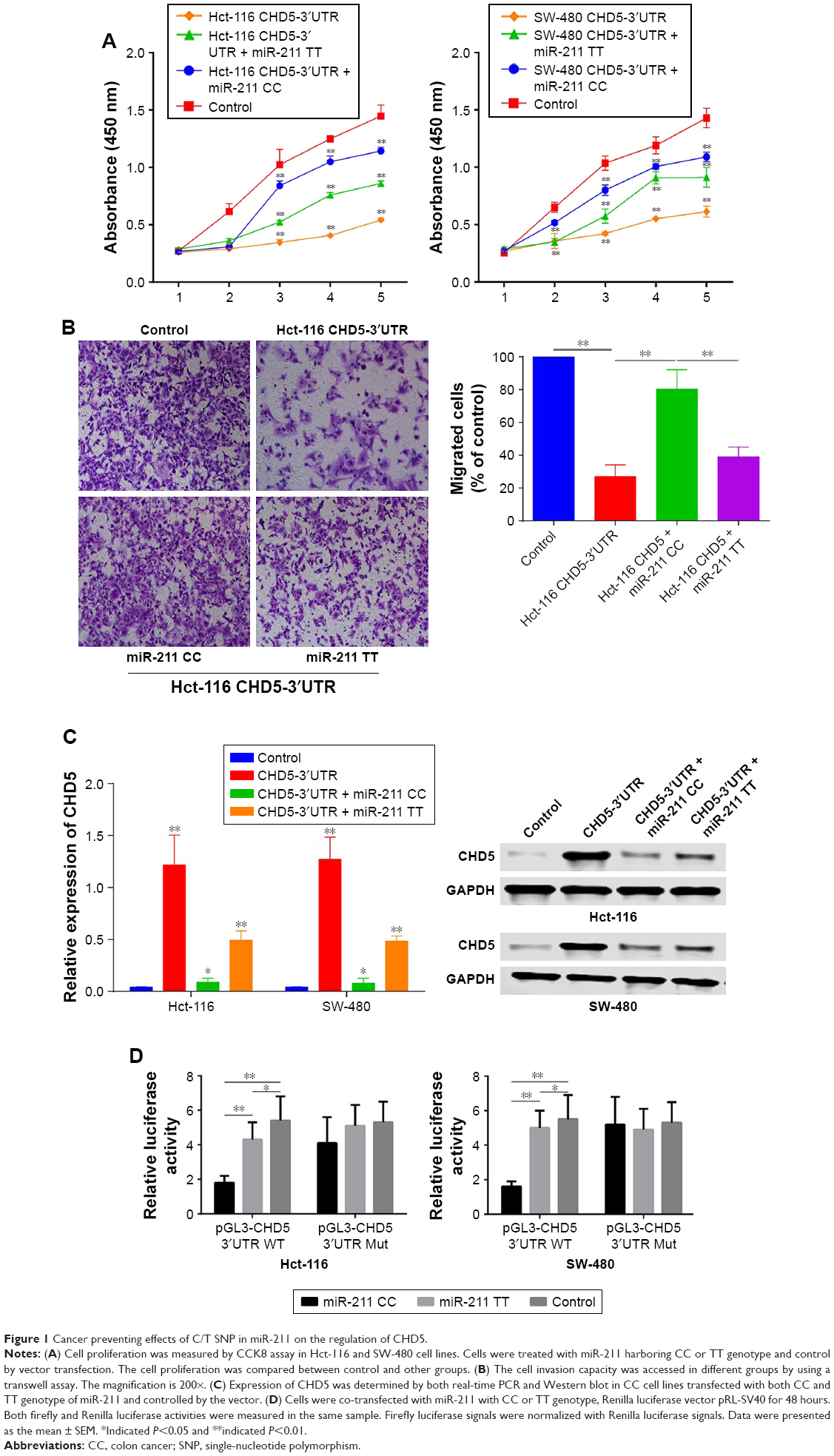 | Figure 1 Cancer preventing effects of C/T SNP in miR-211 on the regulation of CHD5. |
Next, the roles of the SNP on cell invasion were evaluated by a transwell assay, the CHD5 overexpression can significantly attenuate the invasion capacity of Hct-116 cells, but can be almost entirely restored by miR-211 CC and only restored partially by miR-211 TT (Figure 1B).
We think the effects of rs187960998 can be demonstrated by the expression levels of CHD5 in different genotype groups. Real-time PCR and Western blot revealed that the CHD5 expression in the CHD5-3′UTR group could be silenced by both miR-211 CC and TT. However, the expression of CHD5 in miR-211 TT was significantly higher than miR-211 CC, and the transcription and protein expression in CC group was also dramatically decreased (Figure 1C).
Lastly, we constructed pGL3 vectors contained the 3′UTR region of CHD5 and then co-transfected it with miR-211 with different genotypes. As it was presented in Figure 1D, we found that the overexpression of miR-211 with the CC genotype could enhance the suppression effect by miR-211 wild type but not mutants which also indicated that C/T SNP was vital in the regulation of miR-211 on CHD5.
C>T SNP was associated with high expression of CHD5 and longer postsurgery survival in human CC
We also confirmed the expression of CHD5 in clinical samples with different genotypes of rs187960998. CHD5 expression was detected in human CC by immunohistochemistry (IHC). Due to its characters as a tumor suppressive gene, there are only medium and low staining in varying CC patients, IHC staining consistency in miR-211 CC group was significantly different to CT/TT group (medium 5.1%, low 94.9% for CC group; and medium 42.6% and low 57.3% for CT/TT group, P<0.001) (Figure 2A and B). Real-time PCR further confirmed such difference in CHD5 transcription which CHD5 expression was much lower in CC group compared to CT/TT group. However, there is no significant difference in miR-211 expression between these two groups (Figure 2C and D). Among of total 685 CC patients, we have 206 patients with survival follow-up data, and these persons can be further divided into CC (n=93) and CT/TT (n=113) subgroups. The 5-year survival rate between the two groups was compared to each other. In the CC group, the 5-year survival rate was only 4.35%, which was significantly lower than in the CA/AA group with the survival rate of 38.6% (95% CI: 1.375–2.720; HR =2.149, P=0.0002) (Figure 2E).
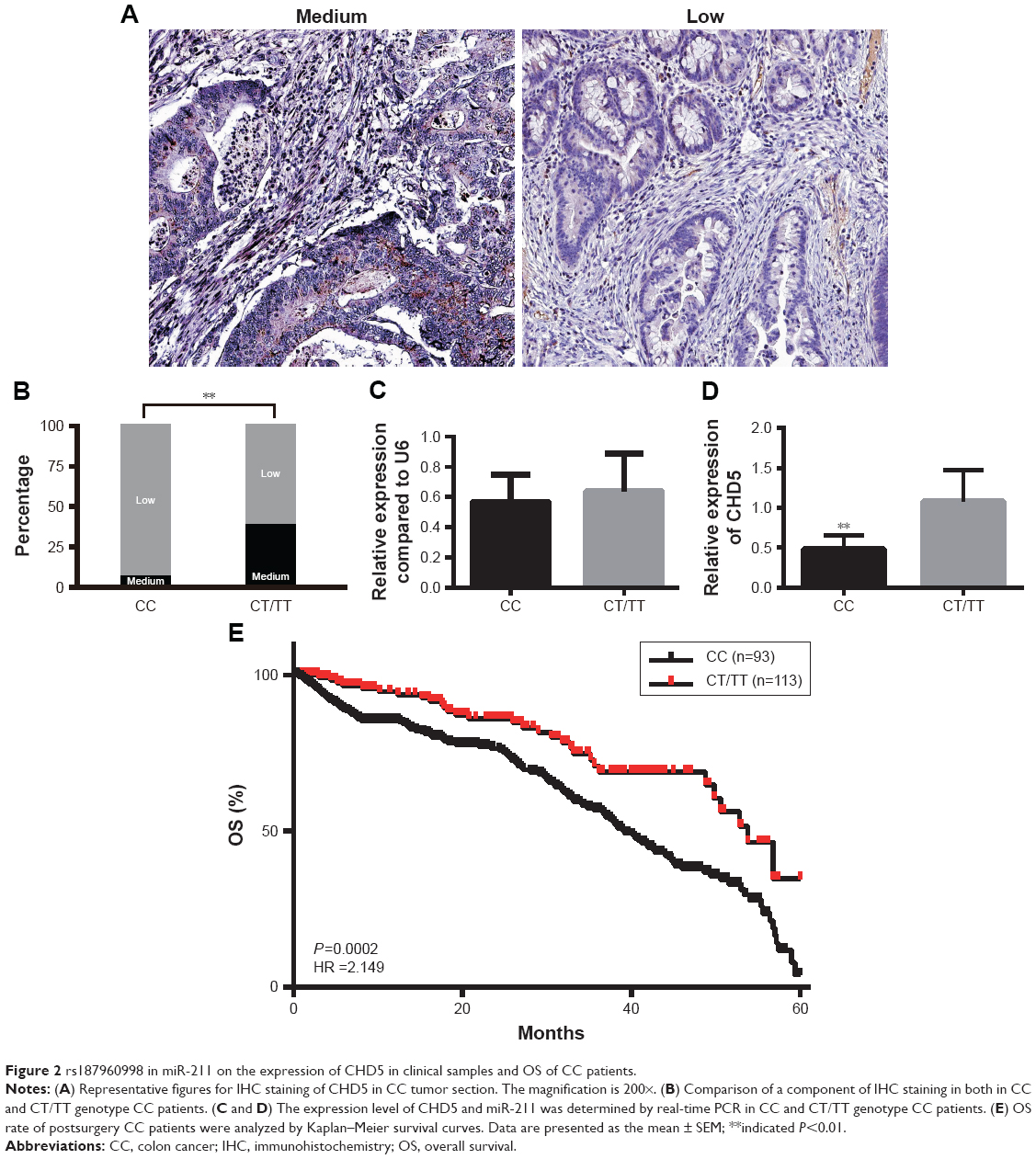 | Figure 2 rs187960998 in miR-211 on the expression of CHD5 in clinical samples and OS of CC patients. |
Discussion
Emerging data have shown that the miRNAs’ aberrant expression was closely related to the carcinogenesis by targeting their potentially regulated genes and these miRNAs potentially serve as biomarkers for prediction and prognosis in various cancers.9–11 It was also proved that the miRNA SNP especially the SNP located in the mature form could also cause an abnormal function of the miRNA in regulating their targeting genes. For example, researchers have found that a genetic variant in miR-27a contributes to CC susceptibility through affecting miR-27a and target gene: zinc finger and BTB domain containing 10 (ZBTB10).12 miRNA SNPs can affect processing and binding ability of mature miRNAs to the 3′UTR of the targeting genes. Functional SNPs of miRNA-146a,13 miRNA-49914 and miRNA-196a215 have been found associated with human disease susceptibility, including breast cancer, lung cancer and Crohn’s disease. Our results were a new evidence for functional miRNA SNP contributing to CC development.
The report concerning the roles of miR-211 in cancer was contradictory. Some studies suggested that miR-211 was an oncogenic miRNA. The survival analysis indicated that high expression of miR-211 was closely associated with a shorter survival time in post-operation CC patients. And the high expression of miR-211 was identified as risk factors for CC prognosis.16 In addition, miRNA-211 is a direct negative regulator of CDC25B expression in triple-negative breast cancer (TNBC) cells, alters other related target proteins CCNB1 and FOXM1 and then inhibits breast cancer cells growth, migration and invasion, leading to G2/M arrest.17 The transcriptional loss of miR-211 increase in CDC25B expression facilitates increased genomic instability at an early stage of TNBC development.17 Moreover, miR-211 functional analyses suggested the role of RRM2 as a target of miR-211 in the modulation of gemcitabine sensitivity.18 On the other hand, some studies implied miR-211 as a tumor suppressor. miR-211 is associated with the invasion of melanoma cells by direct posttranscriptional regulation on KCNMA1.19 In addition, miR-211 directly targets the proximal Chop/Gadd153 promoter, where it increases histone methylation and represses Chop expression and provides an opportunity for the cell to re-establish homeostasis prior to apoptosis.20
The CHD5 gene is repressed in all types of adenomas, either epigenetically or by chromosomal deletion. CHD5 activity is regulated by DNA methylation and repressive histone modifications. CHD5 likely acts as a tumor-suppressor gene in early colorectal carcinogenesis.21 For all CHDs, CpG island hypermethylation was only observed at the CHD5 promoter in human cancer cell lines and primary tumors, particularly gliomas and colon and breast carcinomas.3,22 RT-qPCR analyses correlated CHD5 loss of expression with hypermethylation of the promoter, and restoration of CHD5 mRNA levels upon treatment with a DNA demethylating agent. These results underpin the epigenetic inactivation of the chromatin remodeling factor CHD5 as one contributor to the aberrant structural changes of chromatin throughout the genome of the cancer cell.
Conclusion
We reported the first evidence that the SNP rs187960998 located in miR-211 might be a protective factor to prevent the binding on 3′UTR of CHD5 which might suppress tumor growth in CC.
Disclosure
The authors report no conflicts of interest in this work.
References
Koelzer VH, Zlobec I, Lugli A. Tumor budding in colorectal cancer – ready for diagnostic practice? Hum Pathol. 2016;47(1):4–19. | ||
Levy C, Khaled M, Iliopoulos D, et al. Intronic miR-211 assumes the tumor suppressive function of its host gene in melanoma. Mol Cell. 2010;40(5):841–849. | ||
Mulero-Navarro S, Esteller M. Chromatin remodeling factor CHD5 is silenced by promoter CpG island hypermethylation in human cancer. Epigenetics. 2008;3(4):210–215. | ||
Cai C, Ashktorab H, Pang X, et al. MicroRNA-211 expression promotes colorectal cancer cell growth in vitro and in vivo by targeting tumor suppressor CHD5. PLoS One. 2012;7(1):e29750. | ||
Polytarchou C, Hommes DW, Palumbo T, et al. MicroRNA214 is associated with progression of ulcerative colitis, and inhibition reduces development of colitis and colitis-associated cancer in mice. Gastroenterology. 2015;149(4):981–992. | ||
Nie H, Li J, Yang XM, et al. Mineralocorticoid receptor suppresses cancer progression and the Warburg effect by modulating the miR-338-3p-PKLR axis in hepatocellular carcinoma. Hepatology. 2015;62(4):1145–1159. | ||
Wang X, Li T, Li M, Cao N, Han J. The functional SOCS3 RS115785973 variant regulated by miR-4308 promotes gastric cancer development in Chinese population. Cell Physiol Biochem. 2016;38(5):1796–1802. | ||
Chen Z, Xu L, Ye X, Shen S, Li Z, Niu X, Lu S. Polymorphisms of microRNA sequences or binding sites and lung cancer: a meta-analysis and systematic review. PLoS One. 2013;8(4):e61008. | ||
Tang J, Zhuo H, Zhang X, et al. A novel biomarker Linc00974 interacting with KRT19 promotes proliferation and metastasis in hepatocellular carcinoma. Cell Death Dis. 2014;5:e1549. | ||
Schickel R, Boyerinas B, Park SM, Peter ME. MicroRNAs: key players in the immune system, differentiation, tumorigenesis and cell death. Oncogene. 2008;27(45):5959–5974. | ||
Park H, Lee MJ, Jeong JY, et al. Dysregulated microRNA expression in adenocarcinoma of the uterine cervix: clinical impact of miR-363-3p. Gynecol Oncol. 2014;135(3):565–572. | ||
Sun Q, Gu H, Zeng Y, et al. Hsa-mir-27a genetic variant contributes to gastric cancer susceptibility through affecting miR-27a and target gene expression. Cancer Sci. 2010;101(10):2241–2247. | ||
Gao LB, Bai P, Pan XM, et al. The association between two polymorphisms in pre-miRNAs and breast cancer risk: a meta-analysis. Breast Cancer Res Treat. 2011;125(2):571–574. | ||
Qiu F, Yang L, Ling X, et al. Sequence variation in mature microRNA-499 confers unfavorable prognosis of lung cancer patients treated with platinum-based chemotherapy. Clin Cancer Res. 2015;21(7):1602–1613. | ||
Brest P, Lapaquette P, Souidi M, et al. A synonymous variant in IRGM alters a binding site for miR-196 and causes deregulation of IRGM-dependent xenophagy in Crohn’s disease. Nat Genet. 2011;43(3):242–245. | ||
Cai K, Shen F, Cui JH, Yu Y, Pan HQ. Expression of miR-221 in colon cancer correlates with prognosis. Int J Clin Exp Med. 2015;8(2):2794–2798. | ||
Song GQ, Zhao Y. MicroRNA-211, a direct negative regulator of CDC25B expression, inhibits triple-negative breast cancer cells’ growth and migration. Tumour Biol. 2015;36(7):5001–5009. | ||
Maftouh M, Avan A, Funel N, et al. miR-211 modulates gemcitabine activity through downregulation of ribonucleotide reductase and inhibits the invasive behavior of pancreatic cancer cells. Nucleosides Nucleotides Nucleic Acids. 2014;33(4-6):384–393. | ||
Mazar J, Deyoung K, Khaitan D, et al. The regulation of miRNA-211 expression and its role in melanoma cell invasiveness. PLoS One. 2010;5(11):e13779. | ||
Chitnis NS, Pytel D, Bobrovnikova-Marjon E, et al. miR-211 is a prosurvival microRNA that regulates chop expression in a PERK-dependent manner. Mol Cell. 2012;48(3):353–364. | ||
Kolla V, Zhuang T, Higashi M, Naraparaju K, Brodeur GM. Role of CHD5 in human cancers: 10 years later. Cancer Res. 2014;74(3):652–658. | ||
Fatemi M, Paul TA, Brodeur GM, Shokrani B, Brim H, Ashktorab H. Epigenetic silencing of CHD5, a novel tumor-suppressor gene, occurs in early colorectal cancer stages. Cancer. 2014;120(2):172–180. |
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.