Back to Journals » Journal of Experimental Pharmacology » Volume 17
Repeated Levonorgestrel Administration Impairs Cognitive Function via Glutamatergic Transmission and CaMKII–PKA–BDNF Signaling in the Hippocampus of Female Wistar Rats
Authors Abdulrahim HA, Omeiza NA, Akorede GJ, Iyapo O ![]() , Fatoki JO, Benneth BA, Oyewo EB, Oyewole AL, Alagbonsi AI
, Fatoki JO, Benneth BA, Oyewo EB, Oyewole AL, Alagbonsi AI ![]() , Adekunle AS
, Adekunle AS
Received 8 August 2025
Accepted for publication 9 December 2025
Published 18 December 2025 Volume 2025:17 Pages 841—859
DOI https://doi.org/10.2147/JEP.S559477
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Abdelwahab Omri
Halimat Amin Abdulrahim,1,2 Noah Adavize Omeiza,3,4 Ganiu Jimoh Akorede,5 Oluwadamilare Iyapo,6 John Olabode Fatoki,1,7 Ben-Azu Benneth,8 Emmanuel Bukoye Oyewo,1 Aboyeji Lukuman Oyewole,9 Abdullateef Isiaka Alagbonsi,10 Adeniran Sanmi Adekunle1
1Department of Biochemistry, Faculty of Basic Medical Sciences, College of Health Sciences, Ladoke Akintola University of Technology, Ogbomoso, Oyo State, Nigeria; 2Department of Medical Biochemistry, Faculty of Basic Medical Sciences, College of Health Sciences, University of Ilorin, Ilorin, Kwara State, Nigeria; 3Department of Pharmacology and Therapeutics, Faculty of Basic Clinical Sciences, University of Medical Sciences, Ondo, Ondo State, Nigeria; 4Department of Pharmacology and Therapeutics, College of Medicine, University of Ibadan, Ibadan, Oyo State, Nigeria; 5Department of Veterinary Pharmacology and Toxicology, Faculty of Veterinary Medicine, University of Ilorin, Ilorin, Kwara State, Nigeria; 6Department of Morbid Anatomy, Eko University of Medicine and Health Sciences, Ijanikin, Lagos State, Nigeria; 7Department of Medical Biochemistry, Faculty of Basic Medical Sciences, College of Health Sciences, Osun State University, Osogbo, Nigeria; 8DELSU Joint Canada-Israel Neuroscience and Biopsychiatry Laboratory, Department of Pharmacology, Faculty of Basic Medical Sciences, Delta State University, Abraka, Delta State, Nigeria; 9Department of Physiology, Faculty of Basic Medical Sciences, College of Health Sciences, University of Ilorin, Ilorin, Kwara State, Nigeria; 10Department of Physiology, School of Medicine and Pharmacy, College of Medicine and Health Sciences, University of Rwanda, Huye, Southern Province, Rwanda
Correspondence: Abdullateef Isiaka Alagbonsi, Department of Physiology, School of Medicine and Pharmacy, College of Medicine and Health Sciences, University of Rwanda, Huye, Southern Province, Rwanda, Tel +250786092444, Email [email protected]
Background: Levonorgestrel (LNG), a synthetic progestin widely used in emergency contraception, is increasingly taken frequently and often without medical supervision. With rising concerns that repeated exposure to such hormones may adversely affect brain function, this study investigated whether chronic LNG administration impairs cognitive-like behavior and alters key neurochemical pathways in female Wistar rats.
Methods: Experimental rats were assigned to three groups receiving normal saline (control) or LNG (4 or 8 μg/kg) every alternate day for 60 days. Cognitive performance was assessed using the Morris water maze (MWM) and novel object recognition (NOR) tests. Hippocampal tissues were subsequently analyzed for glutamatergic markers and downstream signaling molecules involved in learning and memory.
Results: Chronic LNG exposure (4 and 8 μg/kg) impaired both spatial and non-spatial memory, evidenced by prolonged escape latency and reduced path efficiency in the MWM, along with a decreased discrimination index in the NOR test. Neurochemically, LNG significantly reduced hippocampal levels of glutamate, N-methyl-D-aspartate (NMDA) and α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptor ligands, protein kinase A (PKA), calcium/calmodulin-dependent kinase II (CaMKII), and brain-derived neurotrophic factor (BDNF), with the 8 μg/kg dose exerting more pronounced effects.
Conclusion: Repeated LNG administration leads to notable cognitive deficits, likely mediated by impairments in glutamatergic signaling and downstream molecular pathways essential for synaptic plasticity. These findings underscore potential neurocognitive risks associated with prolonged LNG exposure.
Keywords: cognitive dysfunction, glutamatergic pathway, hippocampus, levonorgestrel, memory, neuropsychological tests
Introduction
Oral contraceptives (OCs), including levonorgestrel (LNG), have been widely used for decades for pregnancy prevention and the management of several gynecological conditions.1 However, reports of frequent and unsupervised use of LNG, particularly among adolescents, students, and women aged 15 to 49, are becoming increasingly common. Many individuals take LNG repeatedly without medical guidance to suppress endogenous progesterone activity for purposes such as preventing ovulation, reducing menstrual flow, and alleviating cramps. Although effective for these indications, chronic or unsupervised exposure may predispose users to metabolic disturbances and potential neurological consequences.2–5
LNG is a synthetic progestin that binds progesterone receptors (PR-A and PR-B), forming a hormone–receptor complex that inhibits ovulation and prevents pregnancy.6 Unlike endogenous progesterone, LNG does not fully replicate the neuromodulatory actions of physiological hormones, which include regulating neurochemical signals essential for sexual behavior and broader brain functions.6,7 In addition to PRs, LNG can interact with estrogen, androgen, glucocorticoid, and mineralocorticoid receptors.8,9 Notably, glucocorticoid receptors have been implicated in stress-related cognitive decline, suggesting a possible link between exogenous progestin exposure and altered cognitive outcomes.
PR-A and PR-B are widely distributed in brain regions critical for cognition, such as the hippocampus, prefrontal cortex, and amygdala.10 Many neurotransmitters involved in cognitive function, like learning and memory - including acetylcholine, glutamate (Glu), γ-aminobutyric acid (GABA), dopamine, and norepinephrine - are modulated by neurosteroids or progesterone.11,12 Consequently, altering progesterone signaling through synthetic progestins that exert both genomic13 and non-genomic14 actions may influence several non-reproductive brain functions, particularly learning and memory.
Within the domain of cognition, the hippocampus plays a central role in the acquisition, consolidation, and retrieval of information.15,16 It expresses high densities of PRs17 and is enriched with glutamatergic synapses essential for long-term potentiation (LTP), a cellular substrate of memory formation.18–20 Glutamate primarily activates ionotropic N-methyl-D-aspartate (NMDA) and α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptors, triggering calcium influx and stimulating downstream signaling molecules such as calcium/calmodulin-dependent kinase II (CaMKII) and cyclic adenosine monophosphate–dependent protein kinase A (PKA).18 These kinases regulate the transcription factor, cAMP response element–binding protein (CREB), which modulates gene expression and subsequently drives synthesis of proteins—including brain-derived neurotrophic factor (BDNF)—that support synaptic strengthening and memory consolidation.21 Disruption at any point within this pathway (glutamate release, receptor activation, kinase signaling, CREB regulation, or BDNF expression) may impair synaptic plasticity and manifest as cognitive dysfunction.
Importantly, recent findings indicate that these molecular vulnerabilities do not operate in isolation. High exposure to LNG, alongside other systemic physiological stressors, may further modulate neuronal function by altering microglial activity and increasing proinflammatory mediators.22,23 Such inflammatory influences have been shown to converge on glutamatergic signaling and neurotrophic pathways, potentially amplifying disruptions in LTP-related cognitive processes.24,25 Although neuroinflammatory mechanisms were not the primary focus of the present study, their growing recognition in previous reports highlights the multifaceted ways hormonal disturbances can shape cognitive function. Together, these observations reveal the need to consider both direct molecular effects of LNG on synaptic plasticity and the broader physiological context that may exacerbate or modify its impact on brain function.
Despite these insights, the impact of repeated LNG exposure on hippocampal glutamatergic transmission and downstream plasticity-related molecules such as CaMKII, PKA, CREB, and BDNF remains poorly understood. Given the increasing and often unsupervised use of LNG, elucidating its potential neurocognitive implications is both scientifically important and of substantial public health relevance. This study tested the hypothesis that repeated LNG usage would cause a decline in cognitive function via derangement of neurochemicals.
Materials and Methods
Laboratory Animals
This experimental study, conducted between January and March 2019, used 15 mature female Wistar rats (Average weight: 180 g) procured from the Central Research Animal House, Ladoke Akintola University of Technology, Ogbomoso, Oyo State, Nigeria. The experiment was conducted at the Central Research Animal House, University of Ilorin, Nigeria. The animals were bred and housed in the polystyrene cages [n = 5/cage (L: 42 cm × B: 34 cm × H: 25 cm)] under standardized conditions of temperature (25 ± 1 °C), with a photoperiod of 12-h light/dark cycle. They were allowed to have free access to standard pelletized rodent feed and water and acclimatized for two weeks before the commencement of the study. All protocols involving the use of experimental animals were properly adhered to, especially according to the criteria specified in the National Institutes of Health (NIH) (1986) “Guide for the Care and Use of Laboratory Animals”. The study protocols were also approved by the Ethical Research Committee of the University of Ilorin, Ilorin, Nigeria, with the following number: UERC/ASN/2019/1806.
Chemical and Drug Preparation
LNG (4 and 8 µg/kg) (Shuangwei Pharmaceutical Company Limited, Ninghai, China) was dissolved in normal saline (vehicle) and administered intragastrically in a volume of 1.0 mL per 100 g body weight as previously described.26 The selected doses were adapted and modified from the work of Braden et al.27 We exclusively utilized chemicals of analytical grade sourced from British Drug House in Poole, Dorset, UK. Details concerning the analytical kits, their respective manufacturers, catalogue numbers, as well as information on the sensitivity of the products, can be found in Table 1.
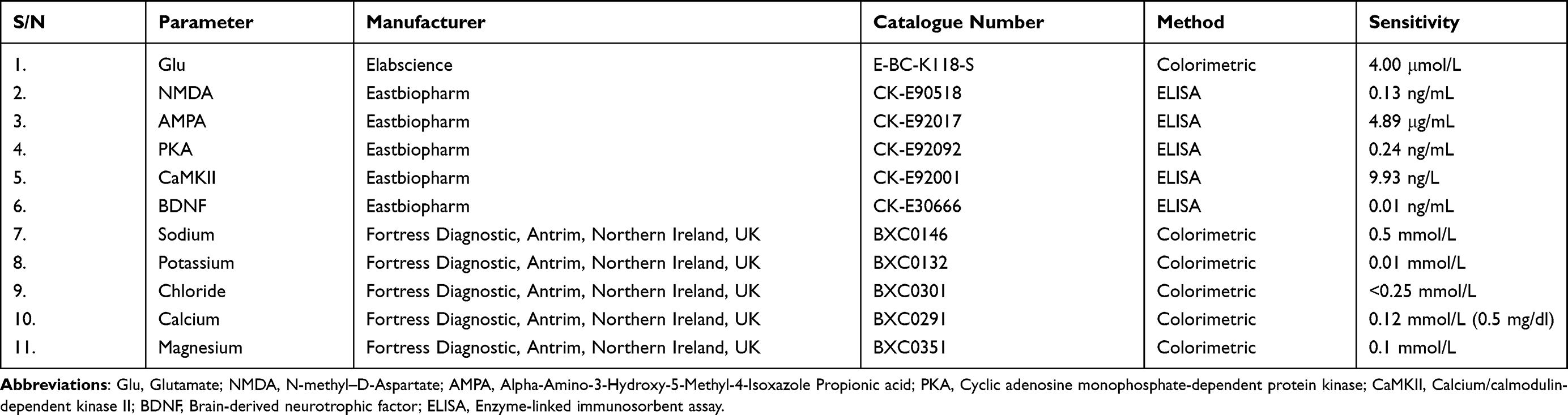 |
Table 1 Product Information on Biochemical Assays |
Experimental Design
To evaluate the effect of LNG on cognitive function, rats were randomly divided into three groups (n = 5/group) based on the sampling method described by Charan and Kantharia.28 Group 1 (Control or vehicle; n = 5) received 10 mL/kg normal saline, group 2 (n = 5) received 4 µg/kg LNG, while group 3 (n = 5) received 8 µg/kg LNG. All treatments were given by oral gavage three times per week at 48-hour intervals for 60 days, between 7:00 and 8:00 AM. To ensure unbiased data collection, the experimenter responsible for administering treatments was not involved in behavioral assessments or biochemical analyses.
Assessment of Learning and Memory
Neurobehavioral assessments were conducted in a quiet room between 9:00 AM and 12:00 PM by trained experimenters who were unaware of the treatment protocol. Rats were transported to the testing room in their home cages and allowed to acclimatize for 60 min before the commencement of behavioral tests.29 In the Morris water maze (MWM) and novel object recognition (NOR) tests, rats were examined for behavioral characterization that mimicked learning ability and memory potentials of the animals (Figure 1).
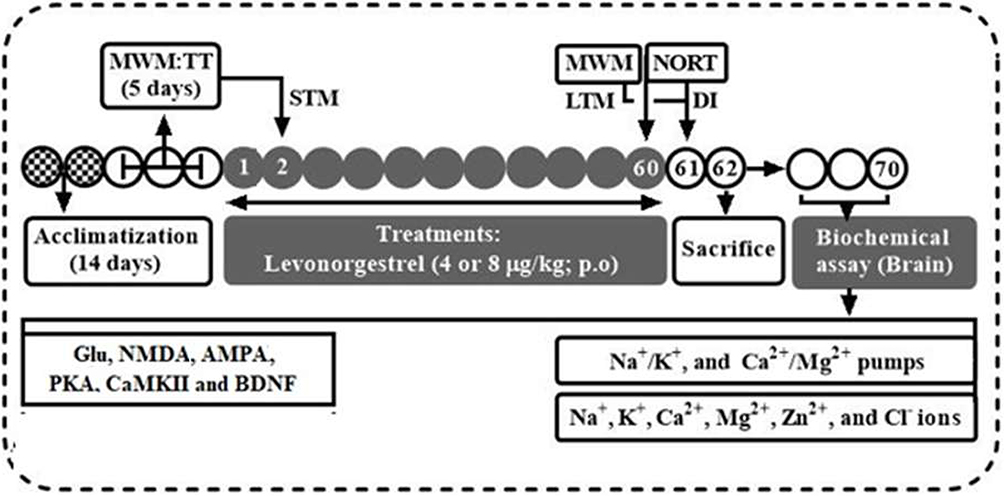 |
Figure 1 Experimental timeline and design. |
Morris Water Maze
The MWM navigation task is a tool for testing spatial and related aspects of rodent learning and memory. As previously described by Omeiza et al,26 rats were tested in a circle-shaped pool (60 cm height × 65 cm diameter) filled with water (40 cm depth) that was made opaque with peak milk at a regulated temperature (25 ± 1 °C). The pool was divided into four quadrants with boundaries marked north (N), east (E), south (S), and west (W), and a circular white platform (10 cm diameter, 28 cm height) was submerged approximately 2 cm below the water surface in the center of the pool’s “southwest” (SW) quadrant. Rats were allowed to swim until they found a mounted platform that served as an escape route within a maximum swimming time of 60 seconds, and then remained on the platform for another 30 seconds to explore the platform quadrant (SW), before being removed from the pool, dried, and placed in a holding cage for the next 30 minutes before another trial. Meanwhile, rats that were unable to locate the platform after 60 seconds of swimming were guided to it by the experimenter and allowed to stay on it for 30 seconds. Each trial was videotaped using a video camera (Webcam) connected to a computer system mounted directly above the apparatus and ANY-Maze video tracking system software from Stoelting Co. (Version 6.3; Wood Dale, IL), as previously reported by Omeiza et al.26
Each rat received a training session consisting of three trials daily for 5 days before LNG administration. A probing test was carried out by removing the platform and allowing the rats to swim freely in the pool for 60 seconds; the time spent in the target quadrant that previously contained the hidden platform was recorded as the reference memory (a day before the first dose). This time spent in the target quadrant indicated the degree of relative consolidation of memory that had taken place after training.30 The time taken to locate the hidden platform (escape latency) was taken as short-term memory (STM), which was measured within six hours after the first dose of LNG. The long-term memory (LTM) was recorded 24 hours after the last dose (on day 61).31 Path efficiency, a parameter for assessing navigation efficiency to reach the invisible platform, was also determined. It was calculated by dividing the straight-line distance between the animal’s first and last positions in the test by the total distance travelled during the test. Path efficiency with a value of 1 indicates perfect efficiency (the animal moved in a straight line), while values less than 1 indicate decreasing efficiency.
Novel Object Recognition Test
Novel object recognition test (NORT) is used to evaluate rats’ non-spatial memory function, especially memory recognition processes, as previously described by Leger et al32 in rodent models of CNS impairments. The experiment was based on rats’ natural tendency to spend more time exploring a novel object than a familiar one. Briefly, the apparatus is a wooden box (60 cm × 60 cm × 50 cm) with a sawdust-covered bottom, and the test is separated into three phases: habituation, training, and testing. In the habituation phase that took place on day 60, the rats were exposed to an open field with no objects; the purpose of this phase is to acclimate the rats to the open field. Three hours later, rats were exposed to the second phase, in which identical objects were put in the open field for exploration. Twenty-four hours after the last LNG treatments (day 61), rats were subjected to the third phase, in which a new object would replace one of the two identical objects. The sniffing time and number were recorded with a 10-minute cutoff time. To avoid any odor signals, the box and items were cleaned with 70% alcohol before and after each behavioral session. Exploratory behavior is defined as pointing the nose at a distance of 20 mm towards an object. The operation was video coded and assessed by an investigator who was not aware of the treatments using a Webcam mounted directly above the apparatus and ANY-Maze video tracking system software from Stoelting Co. (Version 6.3; Wood Dale, IL).26 The percent recognition index was used as a memory parameter using the following formula: [(Tnovel − Tfamiliar)/(Tnovel + Tfamiliar)] × 100, where T represents the length of time spent in exploring the novel object versus the familiar item.
Sample Collection and Preparation
On day 62 of the experiment, rats were sacrificed under cervical decapitation and transcardially perfused with 50 mL of phosphate buffer (0.1 M, pH 7.4). To remove hippocampal issues, sagittal blunt dissection was used. Afterwards, the hippocampal tissues were rinsed with phosphate buffer before being homogenized in aqueous potassium phosphate buffer (0.1 M, pH 7.4) in a 1:4 ratio w/v using a mechanized homogenizer (LANKAI FSH 2A, CHINA), centrifuged at 4000 rpm for 30 min at 4 °C to residue, and the resulting supernatants on the top were aspirated into plain bottles and stored at - 20 °C before neurochemical assays.
Determination of Neuro-Biochemical Parameters
To establish the effect of LNG on memory formation or the propagation of the consolidating signaling memory pathway, analytical kits were used to determine the concentrations of the neurochemicals (Glutamate, NMDA, AMPA, PKA, CaMKII, and BDNF) in the hippocampal tissue supernatants following the procedures described in the manufacturer’s protocols (Table 1). Furthermore, the activities of membrane-bound carrier pumps in the hippocampus (Na+/K+ and Ca2+/Mg2+ ATPases) were measured using a malachite Green-based reagent (Biomol No: AK-111, Enzo Life Sciences), as previously described by Gache et al.33 Regarding the determination of electromolecules (such as Na+, K+, Ca2+, Mg2+, Zn2+, and Cl–), the colorimetric method was used to determine the concentrations of Na+, K+, Mg2+, Ca2+, and Zn2+ as previously reported by Dorup et al34 and Cl–, as previously described.35
Determination of Total Protein and Weight Gain
The weight of the animals was measured before and during the experimental period. Weight gain was determined as the difference between the initial and the final weight of the animal. To precipitate the protein, 100 mg (wet weight) of hippocampal tissue homogenate was mixed with 10% Trichloroacetic acid. The sample was centrifuged at 3000 rpm for 5 minutes. The upper layer was removed. In 1N sodium hydroxide (NaOH), the precipitate was dissolved again. This was then mixed well with the addition of 5 mL of reagent C and allowed to sit uninterrupted for 10 minutes. This was then stirred with 0.5 mL of Folin phenol reagent and set aside for 30 minutes. 1% BSA and 1N NaOH were used as the standard and blank, respectively. The generated blue colour was measured with an ultraviolet-visible double-beam spectrophotometer (Elico) at 650 nm, which automatically quantifies the protein content in the sample.
Data Analysis
GraphPad Prism 8.0 software was employed for the analysis of behavioral tests, biochemical parameters, and histopathological quantifications. Data were expressed as mean ± SEM, and statistical significance (p < 0.05) was determined by one-way analysis of variance, followed by Tukey’s multiple comparison post-hoc test.
Results
LNG Exposure Impairs Cognitive-Like Behaviors in Female Wistar Rats
Given the increasing concern that prolonged exposure to synthetic progestins may adversely affect hippocampal-dependent cognition,22 we first assessed the impact of repeated LNG administration on spatial and non-spatial memory in female Wistar rats. ANOVA revealed significant treatment effects across all assessed cognitive-like behavioral parameters, including escape latency and path efficiency in the MWM, and the discrimination index in the NORT.
In the MWM, rats treated with 4 or 8 µg/kg LNG showed significantly higher (p < 0.05) escape latency compared to controls, indicating impaired acquisition of both STM [F(2, 12) = 11.76, p = 0.0015] (Figure 2A) and LTM [F(2, 12) = 188.5, p < 0.0001] (Figure 2B). These increases in escape latency reflect a reduced ability to efficiently learn and recall the platform’s location. To further evaluate spatial learning efficiency, path efficiency was analyzed and rats exposed to 4 µg/kg LNG, but not 8 µg/kg, displayed a significantly reduced (p < 0.05) path efficiency compared to control animals [F(2, 12) = 7.773, p = 0.0068] (Figure 2C). Reduced path efficiency indicates difficulty navigating directly toward the hidden platform, reflecting diminished spatial learning accuracy. In the NORT, both LNG doses (4 and 8 µg/kg) significantly reduced (p < 0.05) the discrimination index [F(2, 12) = 9.171, p = 0.0038] (Figure 2D), demonstrating impaired ability to distinguish between familiar and novel objects. This suggests a clear deficit in non-spatial memory following repeated LNG exposure. Taken together, these findings suggest that chronic LNG administration disrupts both spatial and non-spatial cognitive processes.
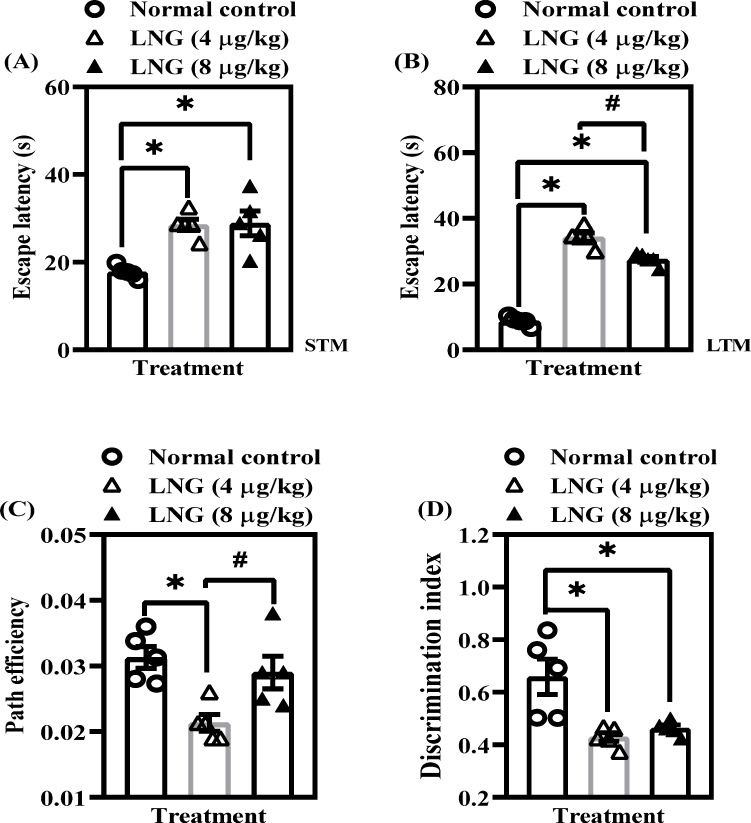 |
Figure 2 LNG exposure impairs cognitive-like behaviors in female Wistar rats. (A) Escape latency – short-term memory [STM]; (B) Escape latency – long-term memory [LTM]; (C) Path length efficiency; (D) Discrimination index. Bars = mean ± SEM (n = 5/group). *p<0.05, comparing the control vs LNG (4 and 8 µg/kg) groups; #p < 0.05, comparing both LNG 4 µg/kg and LNG 8 µg/kg groups. |
LNG Insult Impairs Neurochemical Pathways Responsible for Cognitive Function in Female Wistar Rats
Considering that hippocampal-dependent memory relies heavily on glutamatergic transmission and downstream signaling cascades,18,20 we examined key neurochemical markers involved in synaptic plasticity in rats exposed to LNG. ANOVA results revealed that LNG treatment significantly (p < 0.05) altered multiple biomarkers essential for learning and memory. As shown in Figure 3A, hippocampal glutamate concentration was significantly reduced by LNG, with the effect being more pronounced at 8 µg/kg [F(2, 12) = 158.5, p < 0.0001]. Similarly, both doses of LNG (4 and 8 µg/kg) markedly decreased NMDA [F(2, 12) = 46.06, p < 0.0001] (Figure 3B) and AMPA agonist levels [F(2, 12) = 516.5, p < 0.0001] (Figure 3C), with the higher dose again exerting a significantly greater suppressive effect. These findings indicate that LNG compromises excitatory synaptic transmission in a dose-dependent manner, potentially impairing LTP and memory encoding.
 |
Figure 3 LNG insult impairs neurochemical pathways responsible for cognitive function in female Wistar rats. (A) Glutamate; (B) N-methyl-D aspartate [NMDA]; (C) Alpha-amino-3-hydroxy-5-methyl-4-Isoxazole propionic acid [AMPA]; (D) Protein kinase A [PKA]; (E) calcium/calmodulin kinase 2 [CaMKII]; (F) Brain-derived neurotrophic factor [BDNF]. Bars = mean ± SEM (n = 5/group); *p < 0.05, comparing the control vs LNG (4 and 8 µg/kg) groups; #p < 0.05, comparing both LNG 4 µg/kg and LNG 8 µg/kg groups. |
In line with these observations, PKA activity, a key mediator of cAMP-dependent signaling, was also significantly decreased, with the reduction being more pronounced at 8 µg/kg [F(2, 12) = 361.4, p < 0.0001] (Figure 3D). Similarly, the CaMKII activity was significantly reduced at both doses [F(2, 12) = 73.22, p < 0.0001] (Figure 3E), indicating impaired calcium/calmodulin-dependent kinase activation critical for memory consolidation. Besides, repeated LNG exposure led to a pronounced decline in BDNF levels [F(2, 12) = 114.4, p < 0.0001] (Figure 3F), suggesting a reduction in neurotrophic support necessary for neuronal maintenance and synaptic plasticity. Collectively, these neurochemical alterations demonstrate that repeated LNG exposure disrupts glutamatergic neurotransmission and downstream signaling pathways within the hippocampus in a dose-dependent fashion, with effects more pronounced at 8 µg/kg. These disruptions likely underlie the cognitive deficits observed in the behavioral tests (Figure 2).
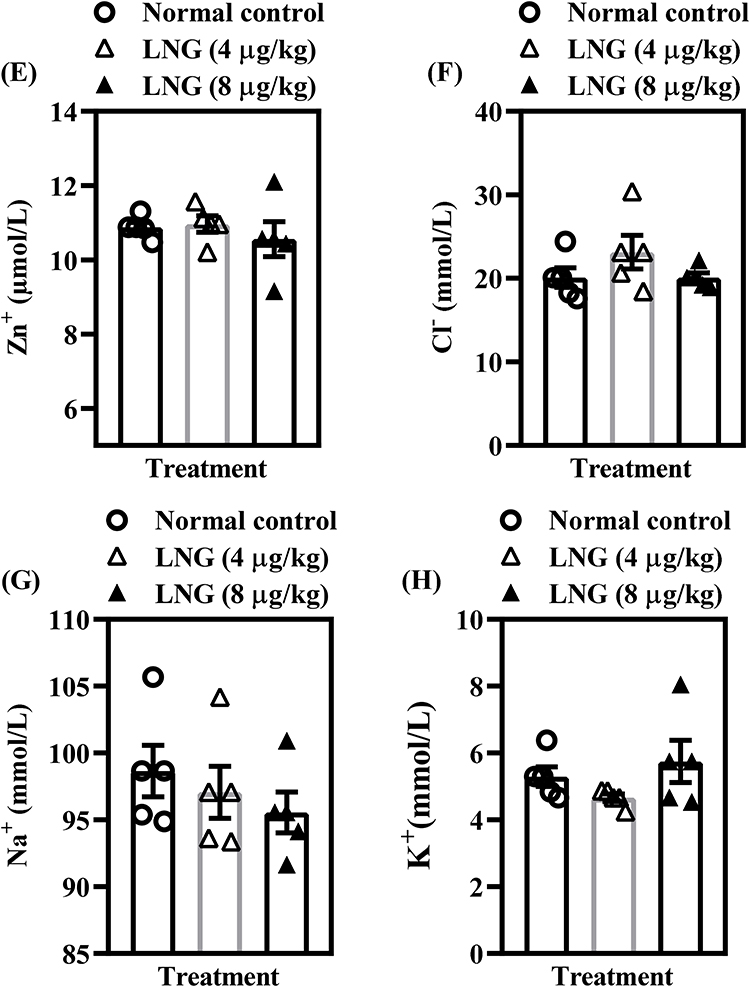
LNG Maintains Baseline Electromolecular Homeostasis but Enhances Ca²⁺/Mg²⁺-ATPase Activity at Higher Dose in Female Wistar Rats
To determine whether repeated LNG exposure affects the ionic and electrophysiological components critical for neuronal excitability and cognitive processing, several electromolecules were assessed (Figure 4). Statistical analysis showed that neither 4 µg/kg nor 8 µg/kg LNG produced significant alterations (p > 0.05) in Na⁺/K⁺-ATPase activity [F(2, 12) = 1.896, p = 0.1925] (Figure 4A) or in the concentrations of Ca²⁺ [F(2, 12) = 3.556, p = 0.0613] (Figure 4C), Mg²⁺ [F(2, 12) = 1.864, p = 0.1972] (Figure 4D), Zn⁺ [F(2, 12) = 0.4748, p = 0.6332] (Figure 4E), Cl− [F(2, 12) = 1.628, p = 0.2369] (Figure 4F), Na⁺ [F(2, 12) = 0.7326, p = 0.5010] (Figure 4G), or K⁺ levels [F(2, 12) = 1.793, p = 0.2083] (Figure 4H). These findings suggest that basic ionic homeostasis and membrane pump function remain largely preserved under the conditions of LNG exposure used in this study. However, Ca²⁺/Mg²⁺-ATPase activity was significantly increased by the higher dose of LNG (8 µg/kg) [F(2, 12) = 12.66, p = 0.0011] (Figure 4B), indicating a dose-dependent modulation of ATPase-driven calcium and magnesium transport. Since Ca²⁺/Mg²⁺-ATPases are involved in maintaining intracellular calcium balance—an essential factor in synaptic signaling—this isolated elevation may reflect a compensatory response to disruptions observed in upstream glutamatergic and kinase pathways. Overall, the current findings suggest that LNG’s impact on cognitive function is not primarily mediated through widespread ionic imbalance but may instead stem from targeted disruptions in receptor-mediated signaling and neuroplasticity pathways described earlier.
Figure 4 Continued. Figure 4 LNG maintains baseline electromolecular homeostasis but enhances Ca²⁺/Mg²⁺-ATPase activity at higher dose in female Wistar rats. (A) Na+/K+ ATPase; (B) Ca2+/Mg2+ ATPase; (C) Ca2+; (D) Mg2+; (E) Zn2+; (F) Cl−, (G) Na+; and (H) K+ levels. Bars = mean ± SEM (n = 5/group). *p < 0.05, comparing the control vs LNG (4 and 8 µg/kg) groups.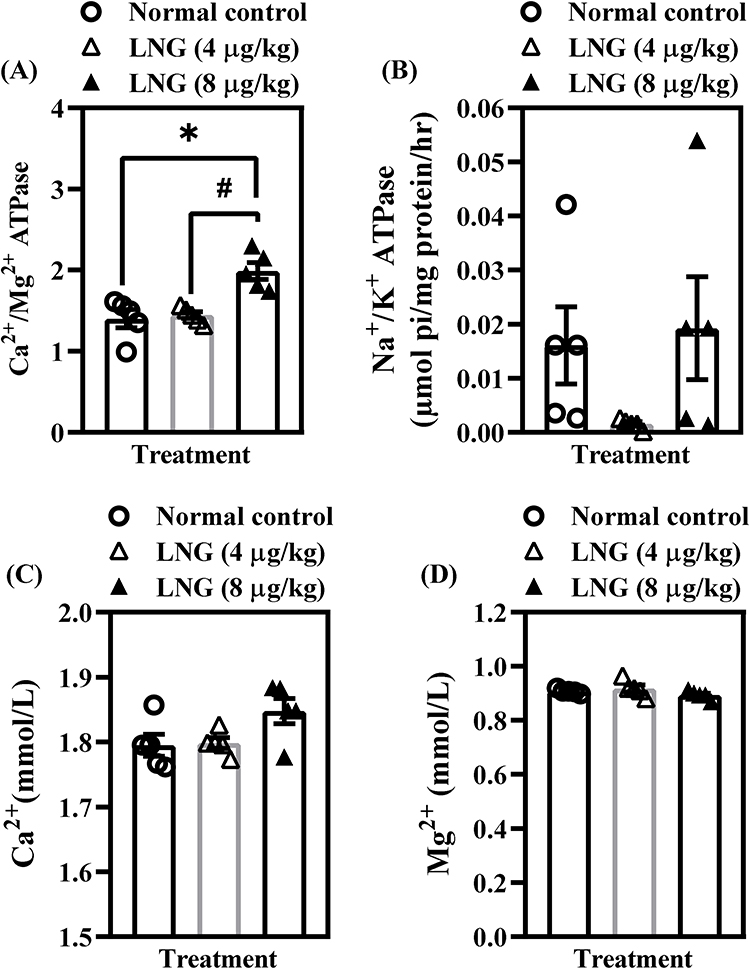
LNG Treatment Increases Percentage Weight Gain and Total Protein in Female Wistar Rats
Figure 5 shows the impact of repeated LNG exposure on percentage weight gain and total protein levels in female Wistar rats. Treatment with 4 µg/kg LNG resulted in a significant increase (p < 0.05) in percentage weight gain compared with the control group [F(2, 12) = 7.325, p = 0.0083] (Figure 5A). However, the higher dose (8 µg/kg) did not produce a similar effect, suggesting a dose-sensitive or non-linear influence of LNG on body weight regulation. In contrast, both 4 and 8 µg/kg LNG significantly elevated total protein levels (p < 0.05) relative to controls [F(2, 12) = 89.32, p < 0.0001] (Figure 5B). Increased total protein concentration may reflect enhanced protein synthesis, altered liver function, or a systemic metabolic response to LNG exposure.
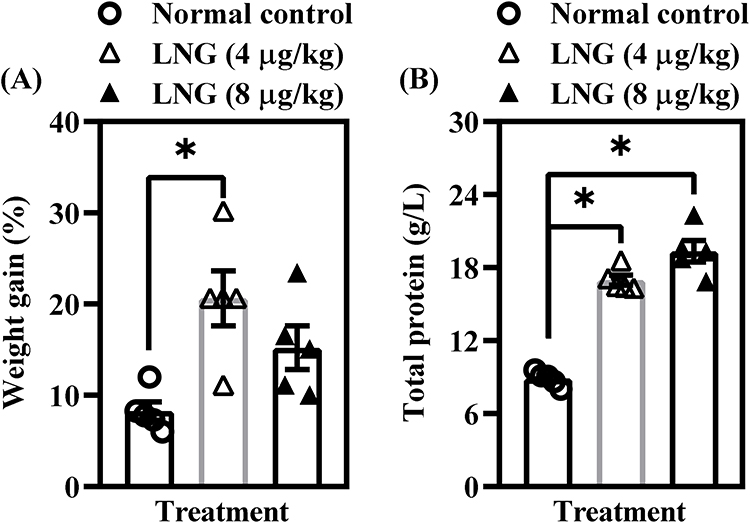 |
Figure 5 LNG treatment increases percentage weight gain (A) and total protein (B) in female Wistar rats. Bars = mean ± SEM (n = 5/group). *p < 0.05, comparing the control vs LNG (4 and 8 µg/kg) groups. |
Progesterone Receptor (Pgr (PR, NR3C3) Is Highly Expressed in Hippocampal Principal Neurons, Supporting the Vulnerability of This Region to LNG Exposure
To strengthen the mechanistic basis for the behavioral and neurochemical deficits observed in this study, we examined the distribution of the progesterone receptor gene (Pgr) within hippocampal principal neurons. Because LNG exerts its biological effects primarily through PRs, the presence and regional enrichment of Pgr in hippocampal circuitry would provide critical evidence for the hippocampus as a direct target of LNG action.
Using the data from the HippoSeq RNA-seq database,36 we evaluated Pgr expression across major hippocampal neuronal subtypes. As shown in Figure 6A, Pgr is expressed broadly within dentate gyrus granule cells, mossy cells, and CA1–CA3 pyramidal neurons. Notably, expression levels were highest in dorsal CA1 (dCA1) and ventral CA3 (vCA3) pyramidal neurons (Figure 6B), a pattern reinforced by the heatmap distribution (Figure 6C). These regions are critically involved in spatial learning, episodic memory encoding, and pattern completion,37 functions that were significantly impaired in LNG-treated rats. Figure 6D-F further show pairwise scatterplots comparing global transcriptional signatures across hippocampal neuron groups. The strong linear clustering and high correlation coefficients (0.86–0.93) confirm the reliability of the dataset and support the validity of the observed Pgr expression patterns, especially in CA1 and CA3. In sum, these findings demonstrate that Pgr is robustly expressed throughout hippocampal principal neurons, with particularly high enrichment in subfields essential for memory processing. This receptor distribution provides a strong anatomical and molecular rationale for the cognitive impairments and synaptic signaling disruptions observed following chronic LNG treatment in our study, suggesting that hippocampal circuits are likely direct sites of LNG-induced neurobiological alterations.
Figure 6 Continued. Figure 6 Pgr gene expression in the hippocampus. (A) Overview of the hippocampal ventro-dorsal axis (illustration by Noah A. Omeiza with BioRender). (B–F) Analysis of Pgr (progesterone receptor; PR/NR3C3) expression in mouse hippocampal neurons using HippoSeq, a publicly available RNA-seq database of hippocampal principal cells.36 (B) Bar plot showing Pgr expression (FPKM) across major hippocampal principal neuron types, including dentate gyrus granule cells (dg), mossy cells (mc), and CA1–CA3 pyramidal neuron subpopulations. Pgr is consistently expressed across all classes and is highest in dCA1 and vCA3 neurons. (C) Heatmap representation of Pgr expression across the same neuronal groups, with warmer colors indicating higher expression. Expression is enriched particularly in dCA1 and vCA3 pyramidal neurons. (D–F) Pairwise scatterplots comparing gene expression patterns across hippocampal neuron classes, with correlation coefficients (CC) ranging from 0.86 to 0.93. These high correlations indicate strong transcriptional similarity across hippocampal principal neurons and consistent Pgr co-expression patterns. Abbreviations: PGR=PR, progesterone receptor; FPKM, Fragments Per Kilobase of Exon Per Million Reads Mapped.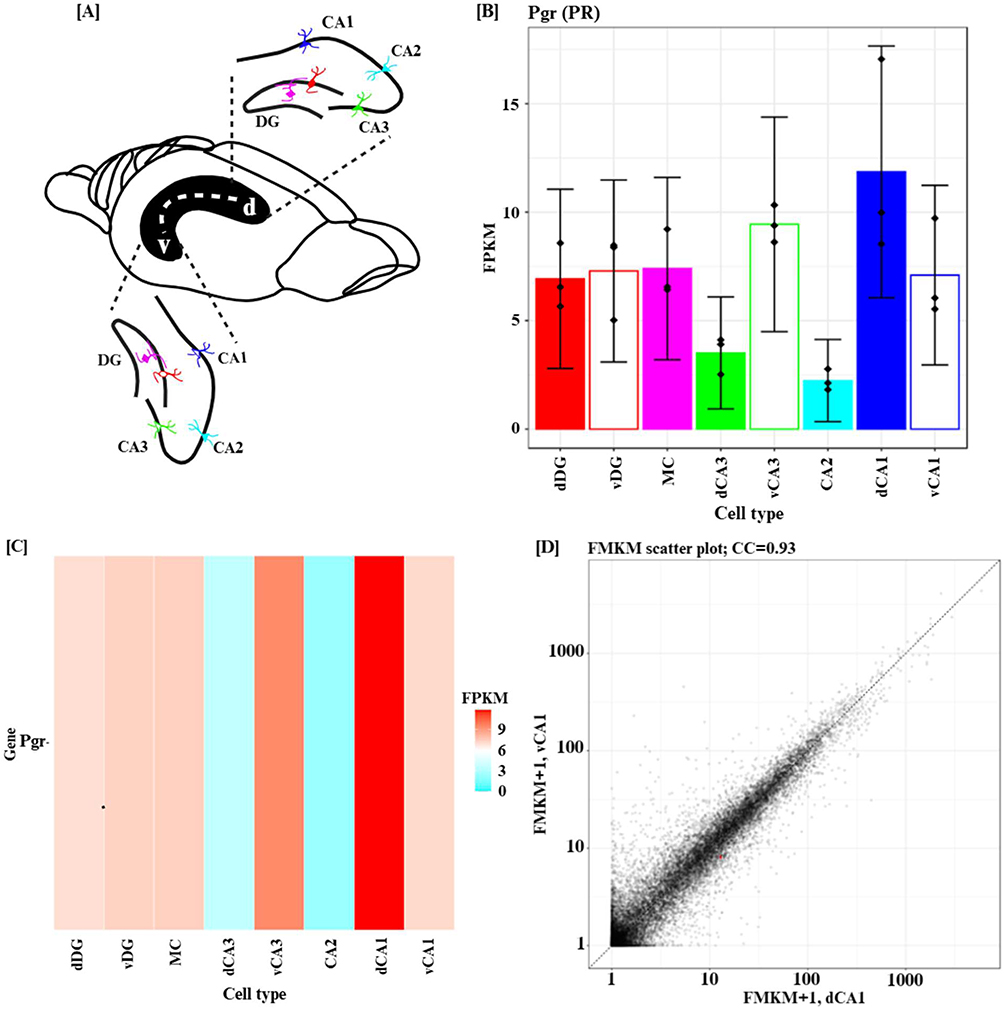

Discussion
This study investigated the impact of repeated LNG exposure on hippocampal-dependent cognitive function and key neurochemical pathways involved in synaptic plasticity. The findings demonstrate that chronic administration of LNG impairs both cognitive functions, disrupts glutamatergic neurotransmission, and alters intracellular signaling cascades central to learning and memory. Together, these results reveal a multifaceted influence of LNG on cognitive and neurochemical processes, providing experimental evidence that prolonged exposure to synthetic progestins may pose neurocognitive risks. The hippocampus is highly sensitive to fluctuations in steroid hormone levels, and progestins such as LNG can exert both genomic and non-genomic effects via progesterone and androgen receptors.8,9 These hormonal interactions can modulate synaptic strength, receptor trafficking, and neurotrophic factor expression. Therefore, the observed derangements in the neurochemical signaling in this study likely reflect both direct receptor-mediated actions and secondary consequences of altered hormonal balance within hippocampal circuits.
In the current study, LNG-treated rats showed prolonged escape latency and reduced path efficiency in the MWM, a well-established model for assessing hippocampal-dependent learning and memory in rodents.25,38 These impairments suggest a decline in both spatial learning and LTM consolidation, consistent with previous evidence that disruption of synaptic plasticity processes, particularly LTP, impedes the conversion of STM to LTM.26,39 The deficits observed in the NOR test further support this conclusion. The NOR test is a classical test for assessing recognition sensitivity based on the natural tendency of rodents to explore novel stimuli.29,40 The reduced discrimination index in LNG-exposed rats suggests a diminished ability to encode or retrieve novel information, aligning with earlier findings that synthetic contraceptive hormones, such as ethinyl estradiol (EE) and LNG, impair cognition by reducing tyrosine hydroxylase and BDNF expression in the locus coeruleus and hippocampus.1 Interestingly, our observations differ from reports showing that LNG enhances spatial memory compared with other synthetic progestins such as norethindrone acetate (NETA) and medroxyprogesterone acetate (MPA), which may be attributed to LNG’s stronger affinity for progesterone and androgen receptors.27,41 Differences in dose, frequency, or route of administration likely account for these discrepancies, highlighting the importance of evaluating LNG’s effects in chronic exposure conditions.
Because synaptic plasticity strongly depends on glutamatergic transmission, we next examined Glu and related biochemical markers. Glu is the principal excitatory neurotransmitter in the brain, essential for LTP induction and memory processes. In our report, the higher dose of LNG significantly reduced total Glu concentration in hippocampal tissue. Studies by Dolphin et al42 and Errington et al43 first demonstrated that increases in extracellular Glu accompany LTP induction, and subsequent work confirmed that lower Glu availability impairs synaptic plasticity.44 Our findings are consistent with these observations and support a presynaptic mechanism whereby LNG reduces Glu supply, which in turn diminishes excitatory drive to hippocampal circuits. This reduction also mirrors the findings of Gibbs et al45 who reported decreased Glu levels in the brains of chicks exposed to neurotoxic compounds. Moreover, Porcu et al46 showed that long-term LNG—but not EE—modified GABAA receptor activity and emotional behavior, suggesting that LNG may enhance GABAergic transmission through increased glutamate decarboxylase activity.47 Given that glutamate decarboxylase converts Glu to GABA,48 reduced Glu levels in our study may reflect LNG-induced shifts in the Glu–GABA balance.
Beyond presynaptic mechanisms, we also evaluated postsynaptic components of glutamatergic neurotransmission by assessing total ligand concentrations for NMDA and AMPA receptors. Although our assay measured ligands, not receptor proteins, reduced NMDA and AMPA ligand levels indicate diminished activation potential at these receptor sites. Earlier studies emphasize that NMDA receptor–dependent LTP relies on both presynaptic Glu release and postsynaptic receptor activation.49,50 NMDA receptor hypofunction is a known mechanism underlying cognitive deficits in Alzheimer’s disease, schizophrenia, and other neurological conditions,29,51,52 Reduced ligand availability, which directly limits receptor engagement, therefore compromises the induction of LTP and delays synaptic impulses in glutamatergic networks.53,54 AMPA receptor activation is equally essential, as AMPAR recruitment and trafficking contribute to early-phase LTP.55,56 Thus, the reductions in NMDA and AMPA ligands observed here are consistent with impaired synaptic plasticity and align with reports showing that chemical agents such as doxorubicin suppress NMDA/AMPA activity and induce memory deficits.57
Given the observed impairments at the receptor ligand level, we investigated whether downstream signaling pathways were similarly affected. LTP induction critically depends on the activation of CaMKII, the phosphorylation of AMPA receptor subunits, and the cAMP–PKA–CREB–BDNF cascade, which promotes gene transcription necessary for long-term synaptic strengthening.58,59 CaMKII phosphorylation of GluA1 subunits increases AMPA receptor conductance,60,61 and PKA-mediated CREB activation induces synthesis of transmembrane AMPAR regulatory proteins and neurotrophic factors such as BDNF.62,63 BDNF is particularly important, as it enhances CaMKII signaling and regulates AMPAR trafficking through interactions with α-stargazin.64 In this study, both doses of LNG significantly reduced CaMKII, PKA, and BDNF levels in the hippocampus. These findings align with previous studies showing that inhibition of CaMKII or BDNF is sufficient to disrupt LTP and impair learning.57 Reduced PKA levels likely reflect diminished cAMP generation, possibly due to decreased adenylyl cyclase activity, which would limit CREB phosphorylation and gene transcription.65 BDNF reductions may result from LNG-mediated transcriptional repression.66 Because BDNF is central to synaptic plasticity, its depletion is linked to cognitive dysfunction in Alzheimer’s disease, schizophrenia, and developmental cognitive impairments.67,68 The behavioral deficits observed in MWM and NOR tasks in our study are therefore consistent with disruption of the CaMKII–PKA–CREB–BDNF axis.35,69 In agreement with prior work, inhibition of the cAMP–PKA–CREB–BDNF pathway culminates in memory impairment.70–72
We further assessed whether ionic components critical for membrane excitability were altered. Ionic fluxes, particularly Na⁺, K⁺, Ca²⁺, Mg²⁺, Zn²⁺, and Cl−, serve essential roles in action potential generation, neurotransmitter release, and receptor activation.73,74 Sodium influx initiates action potentials and triggers presynaptic Glu release,75 whereas postsynaptic ligand–receptor interactions induce additional influx of Na⁺ and Ca²⁺ and efflux of K⁺, leading to depolarization and signal transmission.76 Disturbances in Ca²⁺ homeostasis are often associated with cognitive disorders characterized by impaired cAMP–PKA–CREB–BDNF signaling.77,78 In our study, however, LNG did not significantly alter most ion concentrations or Na⁺/K⁺-ATPase activity, suggesting that basic ionic gradients remained intact. Only Ca²⁺/Mg²⁺-ATPase activity increased at the higher LNG dose, potentially reflecting a compensatory response to reduced calcium entry via NMDA receptor channels. Nevertheless, ionic stability indicates that cognitive deficits were driven primarily by neurotransmitters and signaling disruptions rather than membrane-level ionic disturbances. Additionally, computational interrogation of hippocampal transcriptomics using the HippoSeq platform revealed that Pgr expression is abundant across hippocampal principal neurons, particularly in dCA1 and vCA3, regions implicated in spatial navigation, episodic memory, and pattern completion. The high receptor density in these subfields suggests that LNG can act directly on hippocampal circuits, disrupting neurochemical homeostasis and synaptic plasticity. This receptor distribution provides an anatomical basis for the behavioral and neurochemical impairments observed.36,37
Despite the valuable insights gained, several limitations should be acknowledged. First, the sample size of n = 5/group was relatively small, which may limit the generalizability and statistical power of the results. Larger cohorts would strengthen the reliability of behavioral and biochemical observations. Second, the study assessed outcomes only after repeated LNG exposure without including intermediate time-point evaluations, making it difficult to determine whether the observed neurochemical changes emerge progressively or acutely. Third, histological and Western blot analyses were not conducted to confirm receptor or protein-level alterations within hippocampal tissue. Incorporating such assays would provide direct evidence of receptor expression and pathway engagement. Fourth, electrophysiological assessments—such as local field potential (LFP) recordings to measure LTP in the perforant path (EC–DG), mossy fiber tract (DG–CA3), and Schaffer collateral (CA3–CA1) pathways—were not conducted. Such experiments, combined with single-cell recordings of principal neurons expressing Pgr, would clarify how LNG modulates synaptic strength and neuronal excitability. Incorporating these complementary approaches in future research would provide a more comprehensive understanding of LNG’s mechanistic effects on hippocampal circuitry and synaptic plasticity.
Conclusion
In summary, repeated LNG exposure in female Wistar rats significantly impairs hippocampal-dependent cognitive performance, evidenced by prolonged escape latency and reduced recognition memory. These behavioral deficits correspond with marked neurochemical alterations, including reduced glutamate availability, decreased NMDA and AMPA receptor ligand concentrations, and suppression of the CaMKII–PKA–CREB–BDNF signaling cascade—pathways essential for synaptic plasticity and memory formation. Although ionic homeostasis remained largely intact, the selective increase in Ca²⁺/Mg²⁺-ATPase activity at higher LNG doses indicates a compensatory adjustment aimed at preserving calcium balance. Together, these findings suggest that LNG-induced cognitive dysfunction arises primarily from disrupted glutamatergic and neurotrophic signaling rather than from global ionic imbalance. The data thus provide a mechanistic framework linking chronic LNG exposure to impaired hippocampal plasticity and cognitive decline, highlighting the need for further investigation into the neurocognitive safety of prolonged synthetic progestin use.
Abbreviations
AMPA, Alpha-Amino-3-Hydroxy-5-Methyl-4-Isoxazole Propionic acid; AMPAR, AMPA receptor; BDNF, brain-derived neurotrophic peptide; Ca2+, calcium; CaMKII, calcium/calmodulin-dependent kinase II; cAMP, cyclic adenosine monophosphate; Cl−, chloride; CNS, central nervous system; CREB, cAMP responsive element binding protein; EE, ethinyl estradiol; ELISA, enzyme-linked immunosorbent assay; GABA, gamma-aminobutyric acid; glu, glutamate; K+, potassium; LNG, levonorgestrel; Mg+, magnesium; MPA, medroxyprogesterone acetate; MWM, morris water maze; LTM, long-term memory; LTP, long-term potentiation; Na+, sodium; NaOH, sodium hydroxide; NETA; norethindrone acetate; NMDA, N-methyl-D-aspartate; NMDAR, NMDA receptor; NOR, novel object recognition; OCs, Oral contraceptives; PKA, cyclic adenosine monophosphate-dependent protein kinase A; PRs, progesterone receptors; STM, short-term memory; SW, southwest; Zn2+, zinc.
Data Sharing Statement
Data will be made available upon reasonable request from the corresponding author.
Ethical Considerations
All protocols involving the use of experimental animals were properly adhered to, especially according to the criteria specified in the National Institutes of Health (NIH) (1986) “Guide for the Care and Use of Laboratory Animals”. The study protocols were also approved by the Ethical Research Committee of the University of Ilorin, Ilorin, Nigeria, with the following number: UERC/ASN/2019/1806.
Acknowledgments
The authors wish to thank Staffs of the Central Research Laboratory (University of Ilorin, Ilorin, NG), Mr. Sarafudeen O. Owolabi (Omega Super Global Trading Services Limited, Ilorin, NG), BOLAB Medical and Scientific Laboratory, Ogbomoso and Mr. Fowotade A. Adekunle (Department of Morbid Anatomy, University of Ilorin Teaching Hospital, Ilorin, NG) for their excellent technical assistance during the study.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Disclosure
The authors declare no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
References
1. Simone J, Bogue EA, Bhatti DL, et al. Ethinyl estradiol and levonorgestrel alter cognition and anxiety in rats concurrent with a decrease in tyrosine hydroxylase expression in the locus coeruleus and brain-derived neurotrophic factor expression in the hippocampus. Psychoneuroendocrinology. 2015;62:265–278. doi:10.1016/j.psyneuen.2015.08.015
2. Hakizimana JC, Alagbonsi AI. Modulation of lactose synthesis and orexinergic‐glucose pathway by sex steroid hormones. Physiol Rep. 2025;13(22). doi:10.14814/phy2.70661
3. Kampire M, Hakizimana J, Mucumbitsi J, Alagbonsi A. Pathophysiological consequences associated with hormonal contraceptives use in Sub-Saharan Africa: a scoping review. Open Access J Contracept. 2025;16:171–187. doi:10.2147/OAJC.S563680
4. Apter D. Contraception options: aspects unique to adolescent and young adult. Best Pract Res Clin Obstet Gynaecol. 2018;48:115–127. doi:10.1016/j.bpobgyn.2017.09.010
5. Zou Y, Lu Q, Zheng D, et al. Prenatal levonorgestrel exposure induces autism-like behavior in offspring through ERβ suppression in the amygdala. Mol Autism. 2017;8(1):46. doi:10.1186/s13229-017-0159-3
6. Schumacher M, Ghoumari A, Guennoun R, et al. Progestins and antiprogestins: mechanisms of action, neuroprotection and myelination. In: Hormones and the Brain. Berlin Heidelberg: Springer; 2005:111–154. doi:10.1007/3-540-26940-1_8
7. Wise PM. Neuroendocrine modulation and repercussions of female reproductive aging. Recent Prog Horm Res. 2002;57(1):235–256. doi:10.1210/rp.57.1.235
8. Djebaili M, Hoffman SW, Stein DG. Allopregnanolone and progesterone decrease cell death and cognitive deficits after a contusion of the rat pre-frontal cortex. Neuroscience. 2004;123(2):349–359. doi:10.1016/j.neuroscience.2003.09.023
9. Komane M, Avenant C, Louw-du Toit R, Africander DJ, Hapgood JP. Differential off-target glucocorticoid activity of progestins used in endocrine therapy. Steroids. 2022;182:108998. doi:10.1016/j.steroids.2022.108998
10. Rossetti MF, Cambiasso MJ, Holschbach MA, Cabrera R. Oestrogens and progestagens: synthesis and action in the brain. J Neuroendocrinol. 2016;28(7). doi:10.1111/jne.12402
11. Barth C, Villringer A, Sacher J, Hattori A, Ando H. Sex hormones affect neurotransmitters and shape the adult female brain during hormonal transition periods. Front Neurosci. 2015;9:9. doi:10.3389/fnins.2015.00037
12. Bernal A, Paolieri D. The influence of estradiol and progesterone on neurocognition during three phases of the menstrual cycle: modulating factors. Behav Brain Res. 2022;417:113593. doi:10.1016/j.bbr.2021.113593
13. Slater EP, Hesse H, Beato M. Regulation of Transcription by Steroid Hormones a. Ann N Y Acad Sci. 1994;733(1):103–112. doi:10.1111/j.1749-6632.1994.tb17260.x
14. Barabás K, Godó S, Lengyel F, Ernszt D, Pál J, Ábrahám IM. Rapid non-classical effects of steroids on the membrane receptor dynamics and downstream signaling in neurons. Horm Behav. 2018;104:183–191. doi:10.1016/j.yhbeh.2018.05.008
15. Wang H, Xu X, Xu X, Gao J, Zhang T. Enriched environment and social isolation affect cognition ability via altering excitatory and inhibitory synaptic density in mice hippocampus. Neurochem Res. 2020;45(10):2417–2432. doi:10.1007/s11064-020-03102-2
16. Thomas SA. Neuromodulatory signaling in hippocampus‐dependent memory retrieval. Hippocampus. 2015;25(4):415–431. doi:10.1002/hipo.22394
17. Joshi S, Williams CL, Kapur J. Limbic progesterone receptors regulate spatial memory. Sci Rep. 2023;13(1):2164. doi:10.1038/s41598-023-29100-2
18. Sheng M, Kim E. The postsynaptic organization of synapses. Cold Spring Harb Perspect Biol. 2011;3(12):a005678–a005678. doi:10.1101/cshperspect.a005678
19. Wang H, Ardiles AO, Yang S, et al. Metabotropic glutamate receptors induce a form of LTP controlled by translation and arc signaling in the hippocampus. J Neurosci. 2016;36(5):1723–1729. doi:10.1523/JNEUROSCI.0878-15.2016
20. Flemming D, Przysucha M, Hübner U. Cognitive maps to visualise clinical cases in handovers. Methods Inf Med. 2015;54(05):412–423. doi:10.3414/ME15-02-0001
21. Barros LA, Tufik S, Andersen ML. The role of progesterone in memory: an overview of three decades. Neurosci Biobehav Rev. 2015;49:193–204. doi:10.1016/j.neubiorev.2014.11.015
22. Amedu NO, Ifelola B. Impact of ethinylestradiol and levonorgestrel on memory, neurochemical indicators, and neuron density in the hippocampus of wistar rats. J Clin Basic Res. 2025;9(1):14–19.
23. Glumac S, Kardum G, Sodic L, Supe-Domic D, Karanovic N. Effects of dexamethasone on early cognitive decline after cardiac surgery. Eur J Anaesthesiol. 2017;34(11):776–784. doi:10.1097/EJA.0000000000000647
24. de Bartolomeis A, Manchia M, Marmo F, Vellucci L, Iasevoli F, Barone A. Glycine signaling in the framework of dopamine-glutamate interaction and postsynaptic density. implications for treatment-resistant schizophrenia. Front Psychiatry. 2020;11:11. doi:10.3389/fpsyt.2020.00369
25. Omeiza NA, Bakre A, Ben-Azu B, et al. Mechanisms underpinning Carpolobia lutea G. Don ethanol extract’s neurorestorative and antipsychotic-like activities in an NMDA receptor antagonist model of schizophrenia. J Ethnopharmacol. 2023;301:115767. doi:10.1016/j.jep.2022.115767
26. Omeiza NA, Abdulrahim HA, Alagbonsi AI, et al. Melatonin salvages lead‐induced neuro‐cognitive shutdown, anxiety, and depressive‐like symptoms via oxido‐inflammatory and cholinergic mechanisms. Brain Behav. 2021;11(8). doi:10.1002/brb3.2227
27. Braden BB, Andrews MG, Acosta JI, Mennenga SE, Lavery C, Bimonte-Nelson HA. A comparison of progestins within three classes: differential effects on learning and memory in the aging surgically menopausal rat. Behav Brain Res. 2017;322:258–268. doi:10.1016/j.bbr.2016.06.053
28. Charan J, Kantharia ND. How to calculate sample size in animal studies? J Pharmacol Pharmacother. 2013;4(4):303–306. doi:10.4103/0976-500X.119726
29. Omeiza NA, Bakre AG, Abdulrahim HA, et al. Pretreatment with Carpolobia lutea ethanol extract prevents schizophrenia-like behavior in mice models of psychosis. J Ethnopharmacol. 2022;295:115432. doi:10.1016/j.jep.2022.115432
30. Morris R. Developments of a water-maze procedure for studying spatial learning in the rat. J Neurosci Method. 1984;11(1):47–60. doi:10.1016/0165-0270(84)90007-4
31. Nalla S, Ganta S, Bukke SPN, Bandaru N, Onohuean H, Alagbonsi AI. Kaempferide and Norbergenin avert aluminium chloride-induced amyloid β accumulation and neurocognitive shutdown via oxidative and apoptotic mechanisms. Int J Immunopathol Pharmacol. 2025;39. doi:10.1177/03946320251343687
32. Leger M, Quiedeville A, Bouet V, et al. Object recognition test in mice. Nat Protoc. 2013;8(12):2531–2537. doi:10.1038/nprot.2013.155
33. Martínez Gache SA, Recoulat Angelini AA, Sabeckis ML, González Flecha FL. Improving the stability of the malachite green method for the determination of phosphate using Pluronic F68. Anal Biochem. 2020;597:113681. doi:10.1016/j.ab.2020.113681
34. Dørup I, Skajaa K, Clausen T. A simple and rapid method for the determination of the concentrations of magnesium, sodium, potassium and sodium, potassium pumps in human skeletal muscle. Clin Sci. 1988;74(3):241–248. doi:10.1042/cs0740241
35. Munns R, Wallace PA, Teakle NL, Colmer TD. Measuring soluble ion concentrations (Na+, K+, Cl−) in salt-treated plants. In. 2010;371–382. doi:10.1007/978-1-60761-702-0_23
36. Cembrowski MS, Wang L, Sugino K, Shields BC, Spruston N. Hipposeq: a comprehensive RNA-seq database of gene expression in hippocampal principal neurons. Elife. 2016;5. doi: 10.7554/eLife.14997
37. Dimsdale-Zucker HR, Ritchey M, Ekstrom AD, Yonelinas AP, Ranganath C. CA1 and CA3 differentially support spontaneous retrieval of episodic contexts within human hippocampal subfields. Nat Commun. 2018;9(1):294. doi:10.1038/s41467-017-02752-1
38. Bromley-Brits K, Deng Y, Song W. Morris water maze test for learning and memory deficits in alzheimer’s disease model mice. J Visualized Exp. 2011;(53). doi:10.3791/2920
39. Zhang L, Zhang HQ, Liang XY, Zhang HF, Zhang T, Liu FE. Melatonin ameliorates cognitive impairment induced by sleep deprivation in rats: role of oxidative stress, BDNF and CaMKII. Behav Brain Res. 2013;256:72–81. doi:10.1016/j.bbr.2013.07.051
40. Cohen SJ, Stackman RW. Assessing rodent hippocampal involvement in the novel object recognition task. A review. Behav Brain Res. 2015;285:105–117. doi:10.1016/j.bbr.2014.08.002
41. Schindler AE, Campagnoli C, Druckmann R, et al. Reprint of classification and pharmacology of progestins. Maturitas. 2008;61(1–2):171–180. doi:10.1016/j.maturitas.2008.11.013
42. Dolphin AC, Errington ML, Bliss TVP. Long-term potentiation of the perforant path in vivo is associated with increased glutamate release. Nature. 1982;297(5866):496–497. doi:10.1038/297496a0
43. Errington ML, Lynch MA, Bliss TVP. Long-term potentiation in the dentate gyrus: induction and increased glutamate release are blocked by d(−)aminophosphonovalerate. Neuroscience. 1987;20(1):279–284. doi:10.1016/0306-4522(87)90019-4
44. Errington ML, Galley PT, Bliss TVP. Long-term potentiation in the dentate gyrus of the anaesthetized rat is accompanied by an increase in extracellular glutamate: real-time measurements using a novel dialysis electrode. Philos Trans R Soc Lond B Biol Sci. 2003;358(1432):675–687. doi:10.1098/rstb.2002.1251
45. Gibbs ME, hertz L, Ng KT. Inhibition of short-term memory formation in the chick by blockade of extracellular glutamate uptake. Neurobiol Learn Mem. 2004;81(2):115–119. doi:10.1016/j.nlm.2003.10.002
46. Porcu P, Mostallino MC, Sogliano C, Santoru F, Berretti R, Concas A. Long-term administration with levonorgestrel decreases allopregnanolone levels and alters GABAA receptor subunit expression and anxiety-like behavior. Pharmacol Biochem Behav. 2012;102(2):366–372. doi:10.1016/j.pbb.2012.05.011
47. Brambilla P, Perez J, Barale F, Schettini G, Soares JC. GABAergic dysfunction in mood disorders. Mol Psychiatry. 2003;8(8):721–737. doi:10.1038/sj.mp.4001362
48. Buddhala C, Hsu CC, Wu JY. A novel mechanism for GABA synthesis and packaging into synaptic vesicles. Neurochem Int. 2009;55(1–3):9–12. doi:10.1016/j.neuint.2009.01.020
49. Kessels HW, Malinow R. Synaptic AMPA receptor plasticity and behavior. Neuron. 2009;61(3):340–350. doi:10.1016/j.neuron.2009.01.015
50. Bliss TV, Collingridge GL. Expression of NMDA receptor-dependent LTP in the hippocampus: bridging the divide. Mol Brain. 2013;6(1):5. doi:10.1186/1756-6606-6-5
51. Danysz W, Parsons CG. Alzheimer’s disease, β‐amyloid, glutamate, NMDA receptors and memantine – searching for the connections. Br J Pharmacol. 2012;167(2):324–352. doi:10.1111/j.1476-5381.2012.02057.x
52. Uruaka CI, Ben-Azu B, Omeiza NA, et al. Inhibition of oxidative stress, neuroinflammatory cytokines, and protein expressions contributes to the antipsychotic effects of geraniol in mice with ketamine-induced schizophrenia. J Neuroimmune Pharmacol. 2025;20(1):70. doi:10.1007/s11481-025-10226-1
53. Mirsaeed-Ghazi F, Sharifzadeh M, Ashrafi-Kooshk MR, et al. Astaxanthin decreases spatial memory and glutamate transport impairment induced by fluoride. Iran J Pharm Res. 2021;20(4):238–254. doi:10.22037/ijpr.2021.114919.15107
54. Butterfield DA, Pocernich CB. The glutamatergic system and alzheimer Disease. CNS Drugs. 2003;17(9):641–652. doi:10.2165/00023210-200317090-00004
55. Luo Y, Yu Y, Zhang M, He H, Fan N. Chronic administration of ketamine induces cognitive deterioration by restraining synaptic signaling. Mol Psychiatry. 2021;26(9):4702–4718. doi:10.1038/s41380-020-0793-6
56. Citri A, Malenka RC. Synaptic plasticity: multiple forms, functions, and mechanisms. Neuropsychopharmacology. 2008;33(1):18–41. doi:10.1038/sj.npp.1301559
57. Alhowail AH, Pinky PD, Eggert M, et al. Doxorubicin induces dysregulation of AMPA receptor and impairs hippocampal synaptic plasticity leading to learning and memory deficits. Heliyon. 2021;7(7):e07456. doi:10.1016/j.heliyon.2021.e07456
58. Lee SJR, Escobedo-Lozoya Y, Szatmari EM, Yasuda R. Activation of CaMKII in single dendritic spines during long-term potentiation. Nature. 2009;458(7236):299–304. doi:10.1038/nature07842
59. Murakoshi H, Wang H, Yasuda R. Local, persistent activation of Rho GTPases during plasticity of single dendritic spines. Nature. 2011;472(7341):100–104. doi:10.1038/nature09823
60. Kristensen AS, Jenkins MA, Banke TG, et al. Mechanism of Ca2+/calmodulin-dependent kinase II regulation of AMPA receptor gating. Nat Neurosci. 2011;14(6):727–735. doi:10.1038/nn.2804
61. Coombs ID, Cull-Candy SG. Single-channel mechanisms underlying the function, diversity and plasticity of AMPA receptors. Neuropharmacology. 2021;198:108781. doi:10.1016/j.neuropharm.2021.108781
62. Jha NK, SK J, Kar R, Nand P, Swati K, Goswami VK. Nuclear factor‐kappa β as a therapeutic target for alzheimer’s disease. J Neurochem. 2019;150(2):113–137. doi:10.1111/jnc.14687
63. Ramírez D, Saba J, Carniglia L, Durand D, Lasaga M, Caruso C. Melanocortin 4 receptor activates ERK-cFos pathway to increase brain-derived neurotrophic factor expression in rat astrocytes and hypothalamus. Mol Cell Endocrinol. 2015;411:28–37. doi:10.1016/j.mce.2015.04.008
64. Poo M-M. Poo M ming. Neurotrophins as synaptic modulators. Nat Rev Neurosci. 2001;2(1):24–32. doi:10.1038/35049004
65. Wand G, Levine M, Zweifel L, Schwindinger W, Abel T. The cAMP–protein kinase a signal transduction pathway modulates ethanol consumption and sedative effects of ethanol. J Neurosci. 2001;21(14):5297–5303. doi:10.1523/JNEUROSCI.21-14-05297.2001
66. Kadoya Y, Zen K, Tamaki N, et al. Early effects of transcatheter aortic valve replacement on cardiac sympathetic nervous function assessed by 123I-metaiodobenzylguanidine scintigraphy in patients with severe aortic valve stenosis. Eur J Nucl Med Mol Imaging. 2020;47(7):1657–1667. doi:10.1007/s00259-019-04523-0
67. Morris KA, Gold PE. Age-related impairments in memory and in CREB and pCREB expression in hippocampus and amygdala following inhibitory avoidance training. Mech Ageing Dev. 2012;133(5):291–299. doi:10.1016/j.mad.2012.03.004
68. Buhusi M, Etheredge C, Granholm AC, Buhusi CV. Increased hippocampal ProBDNF contributes to memory impairments in aged mice. Front Aging Neurosci. 2017;9:9. doi:10.3389/fnagi.2017.00284
69. Roceri M, Hendriks W, Racagni G, Ellenbroek BA, Riva MA. Early maternal deprivation reduces the expression of BDNF and NMDA receptor subunits in rat hippocampus. Mol Psychiatry. 2002;7(6):609–616. doi:10.1038/sj.mp.4001036
70. Zhong Y, Chen J, Li L, et al. PKA-CREB-BDNF signaling pathway mediates propofol-induced long-term learning and memory impairment in hippocampus of rats. Brain Res. 2018;1691:64–74. doi:10.1016/j.brainres.2018.04.022
71. Li Y, Zhang W, R SHI, et al. Prenatal caffeine damaged learning and memory in rat offspring mediated by ARs/PKA/CREB/BDNF pathway. Physiol Res. 2018:975–983. doi:10.33549/physiolres.933906
72. Luo Y, Kuang S, Li H, Ran D, Yang J. cAMP/PKA-CREB-BDNF signaling pathway in hippocampus mediates cyclooxygenase 2-induced learning/memory deficits of rats subjected to chronic unpredictable mild stress. Oncotarget. 2017;8(22):35558–35572. doi:10.18632/oncotarget.16009
73. Hu R, Gong X, Duan Y, et al. Neurotoxicological effects and the impairment of spatial recognition memory in mice caused by exposure to TiO2 nanoparticles. Biomaterials. 2010;31(31):8043–8050. doi:10.1016/j.biomaterials.2010.07.011
74. Abhilash M, Alex M, Mathews VV, Nair RH. Chronic effect of aspartame on ionic homeostasis and monoamine neurotransmitters in the rat brain. Int J Toxicol. 2014;33(4):332–341. doi:10.1177/1091581814537087
75. Patri M. Synaptic transmission and amino acid neurotransmitters. In: Neurochemical Basis of Brain Function and Dysfunction. IntechOpen; 2019. doi10.5772/intechopen.82121
76. Kew JNC, Kemp JA. Ionotropic and metabotropic glutamate receptor structure and pharmacology. Psychopharmacology. 2005;179(1):4–29. doi:10.1007/s00213-005-2200-z
77. Bhat A, Tan V, Heng B, et al. Roflumilast, a cAMP-specific phosphodiesterase-4 inhibitor, reduces oxidative stress and improves synapse functions in human cortical neurons exposed to the excitotoxin quinolinic acid. ACS Chem Neurosci. 2020;11(24):4405–4415. doi:10.1021/acschemneuro.0c00636
78. Liu H, Han L, Xia X, Xiang X. α-Linolenic acid alleviates aluminium chloride-induced toxicity in PC12 cells by activation of PKA-CREB-BDNF signaling pathway. Oil Crop Sci. 2022;7(2):63–70. doi:10.1016/j.ocsci.2022.02.008
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.