Back to Journals » Diabetes, Metabolic Syndrome and Obesity » Volume 13
Renoprotective Effect of Formononetin by Suppressing Smad3 Expression in Db/Db Mice
Authors Lv J, Zhuang K, Jiang X, Huang H ![]() , Quan S
, Quan S
Received 14 July 2020
Accepted for publication 25 August 2020
Published 28 September 2020 Volume 2020:13 Pages 3313—3324
DOI https://doi.org/10.2147/DMSO.S272147
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Juei-Tang Cheng
Jiawei Lv,1 Kai Zhuang,1 Xiyu Jiang,1 Heqing Huang,1,2 Shijian Quan1
1Department of Pharmacology, School of Pharmaceutical Sciences, Guangzhou University of Chinese Medicine, Guangzhou 510006, People’s Republic of China; 2Laboratory of Pharmacology & Toxicology, School of Pharmaceutical Sciences, Sun Yat-Sen University, Guangzhou 510006, People’s Republic of China
Correspondence: Heqing Huang; Shijian Quan
Department of Pharmacology, School of Pharmaceutical Sciences, Guangzhou University of Chinese Medicine, No. 232 East Wai Huan Road, Guangzhou 510006, People’s Republic of China
Tel +86 1 392 211 9719
Email [email protected]; [email protected]
Purpose: Glomerular sclerosis and renal interstitial fibrosis are the most important pathologies in the development of kidney damage under diabetic conditions. Smad3 plays antagonistic roles in high glucose-induced renal tubular fibrosis, which is an important treatment target for diabetic nephropathy (DN). Formononetin (FMN) has multiple effects on diabetic vascular complications including DN. However, whether it plays an anti-fibrosis role by regulating smad3 is unclear. The purpose of this study was to evaluate the renoprotective effect of FMN by suppressing smad3 expression in db/db mice.
Methods: FMN was orally administered to db/db mice with a dose of 25 or 50 mg/kg/day for 8 weeks. At the end of the study, serum, urine, and kidney samples were collected for biochemical and pathological examinations. The expressions of proteins and mRNA associated with renal fibrosis were determined by biochemical, histological, immunofluorescence, and real-time PCR analysis.
Results: The results showed that FMN substantially improved the glucolipid metabolism, reduced the oxidative stress, and protected renal function in db/db mice. Meanwhile, protein and mRNA expression of smad3 and related regulatory factor of extracellular matrix deposition were significantly suppressed.
Conclusion: The present study suggested that FMN has a good renoprotective effect in DN, which plays an anti-fibrosis role in db/db mice by suppressing the expression of smad3.
Keywords: formononetin, diabetic nephropathy, db/db mice, renal fibrosis, expression of smad3, antioxidants
Introduction
Diabetic nephropathy (DN) is one of the most severe chronic microvascular complications of diabetes mellitus (DM), and is also a growing worldwide epidemic disease, which has become a primary cause of end-stage renal disease (ESRD).1 The current major pathological manifestations of DN are glomerular hypertrophy, thickened glomerular basement membrane (GBM), and increased extracellular matrix (ECM), eventually lead to glomerular sclerosis and renal interstitial fibrosis.2 The deposition of collagen fibronectin and laminin in GBM and ECM increased significantly, particularly at an early stage of the DN.3 At a late stage, type I collagen and type III collagen (COLI, COLIII) depositions were evident in the kidney.4
Smad3 protein is an important inducer of epithelial-mesenchymal transformation (EMT) in fibrosis,5–7 and EMT is the key mechanism underlying renal fibrosis. Recent studies indicate that EMT is a potential source of activated myofibroblasts that finally result in renal fibrosis.8–10 When EMT is activated, mesenchymal feature-specific markers, such as alpha-smooth muscle actin (α-SMA) and vimentin are up-regulated.11–13 The EMT of these cells leads to activated fibroblasts and enhances the pathological process of renal fibrosis. Smad3 protein can directly bind to the promoter region of collagen to up-regulate the expression of collagen and promote the process of renal fibrosis.14,15 Oxidative stress injury is the basis of EMT,16 smad3 protein reduces fibrosis and ECM deposition by inhibiting oxidative stress.17,18 Thus, renal fibrosis is the most important feature of renal pathology of DN, which was related with smad3-mediated fibrosis and oxidative stress.
Formononetin (FMN, Figure 1) is a kind of isoflavone (7-hydroxy-4-methoxy isoflavone, C16H12O4), which is found in plants, such as the family lamiaceae and Leguminosae. It is the main active ingredient of Chinese medicine Radix Astragali, Spatholobus suberectus, and Beautiful Millettia Root. Accumulating evidence indicates that FMN has the effect on regulating glycolipid metabolism and anti-AGEs activity, which is used in the treatment of diabetic vascular complications.19–25 However, the anti-fibrosis activities of FMN and their underlying mechanisms have not been reported.
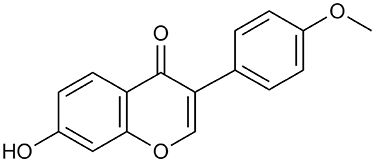 |
Figure 1 Chemical structure of FMN. |
The db/db mouse exhibits insulin resistance at around 2 weeks of age and eventually develops hyperglycemia induced by β cell failure at 4–8 weeks, which accurately reflects the pathophysiology of diabetes.26,27 In the db/db mouse at 7–8 weeks, urinary albumin excretion increased significantly,28 and the kidney began to show pathological changes of DN such as increased Mesangium Matrix, basement membrane thickening, and tubule dilation.29 This pathological change is consistent with the diabetic nephropathy III period, which is known as the early diabetic nephropathy. In the present study, we investigated the role of FMN in delaying DN and explored the potential mechanism related to smad3-mediated anti-fibrosis in a db/db mouse model.
Materials and Methods
Chemicals, Reagents, Kits, and Antibodies
Formononetin (FMN, Purity: HPLC>98%), Metformin (MET), and carboxymethyl cellulose sodium (CMC-Na) were provided by Weikeqi Biotechnology Co., Ltd. (Sichuan, China). RevertAid First Strand cDNA Synthesis Kit (#K1622) and FastStart Universal SYBR Green Master (Rox) were purchased from Thermo Fisher Scientific Co., Ltd. (USA). All test assay kits and the SOD, MDA, GSH-Px, and CAT ELISA kits were supplied by Nanjing Jiancheng Bioengineering Institute (Nanjing, China). The full wavelength enzyme marker was provided by Thermo Fisher Scientific Co., Ltd. (USA). Hematoxylin-eosin stain (H&E), Periodic Acid-Schiff stain (PAS), and Masson staining kits were provided by Servicebio Company (Wuhan, China). Goat anti-Rabbit IgG and Goat anti-Mouse IgG were provided by Servicebio Company (Wuhan, China). The FMN was dissolved with 0.5% CMC-Na. The MET was dissolved with water.
Animals
The male db/db mice (50–60 g) and wild db/+ littermates (25–30 g) in a C57BLKs/J background (8 weeks) were provided by Cavens Laboratory animals, Ltd. (Changzhou, China) [License No: SCXK (Su) 2016–0010]. Mice were housed at the Experimental Animals Center of Guangzhou University of Chinese Medicine (Guangdong, China) [License No: SCXK (Yue) 2018–0085]. All animals were fed freely under the conditions of temperature 22.0–26.0°C, humidity 50–60%, 12 hours:12 hours light dark cycle. The experimental animal protocol was approved by the Animal Ethics Committee of Guangzhou University of Chinese Medicine (No.00222199). All procedures were carried out on the basis of the Laboratory Animal Care and Use recommendations, which are intended to reduce the use of animals and minimize animal distress.
Drug Administration
The db/db mouse group is the model of spontaneous type 2 diabetes. MET is demonstrated to have a good renoprotective effect in DN, which is used as a positive control drug.30,31 After 1 week adaptation, the db/db mice (9 weeks) with non-random blood glucose levels >11.1 mmol/L were considered to be diabetes. The mice were randomly divided into five groups (n=10/group): normal control group (db/m) (NM), model group (db/db) (MD), positive control group (Met), FMN-treated low dose group (FMNL), and FMN-treated high dose group (FMNH). NM and MD were orally administered with 0.5% CMC-Na solution (50 mg/kg). FMNL and FMNH were orally administered with 25 mg/kg and 50 mg/kg FMN, respectively.32 MET were orally administered with MET (100 mg/kg). All mice were gavaged once per day, for 8 consecutive weeks.33
Sample Collection
During the experiment, weight, fasting blood glucose (FBG), food intake, and water intake were measured weekly. After 8 weeks of oral drug treatment, all mice were fasted overnight and then placed in metabolic cages (Yuyan Instruments, Inc., Shanghai, China) to collect fresh urine samples. After measuring urine volume, urine samples were collected by using EP tubes and centrifuged at 2000 rpm for 10 minutes to obtain urine samples and then stored at −20°C. Fresh blood samples were collected from retro-orbital plexus using micro-hematocrit capillaries and centrifuged at 3000 rpm for 10 minutes to obtain serum samples and then stored at −20°C. After the animals were sacrificed, their kidneys, livers, and spleens were removed and weighed. The kidney was fixed in 4% paraformaldehyde and the remaining kidney tissue was snap frozen in liquid nitrogen and then stored at −80°C for analysis.
Biochemical Analysis
The levels of FBG, BUN, Scr, TG, TC, and FINS in serum were determined by test assay kits. ISI and IRI were calculated according to the formulas: ISI=-Ln(FBG×FINS) and IRI=(FINS×FBG/22.5). BUN/Scr was calculated according to the formula: BUN/Scr=(BUN/Scr).
Urine Analysis
The levels of 24-hour urine protein and Ucr in urine were determined by test assay kits. Urine Albumin-to-Creatinine Ratio (ACR) was calculated according to the formulas: ACR=(Urine protein/Ucr).
Oxidative Stress Parameters
After blood samples were taken, the renal tissues were separated. They were homogenized with homogenizer (IKA Company, Germany) and centrifuged at 3500 rpm for 15 minutes at 4°C, and the supernatants were taken. Total protein concentration in the supernatants was separately determined by BCA protein assay kit. The SOD, MDA, GSH-Px, and CAT concentration of the pituitary in the supernatants were detected by ELISA assay kit. The results of ELISA assay kit were read by a ELISA reader (BioTeK Instruments, Inc., USA). The SOD, MDA, GSH-Px, and CAT expression levels were the ratio of their respective protein concentration to the total protein concentration.
Histological Analysis
After being fixed in 4% paraformaldehyde and embedded in paraffin, the kidney tissues were cut into 3 μm thick sections. Sections were then deparaffinized in xylene and rehydrated in ethyl alcohol (from 100 to 70%) in reverse order. All specimens were stained with hematoxylin and eosin (H&E), periodic acid schiff (PAS), and Masson staining and assessed for kidney damage and inflammation under a Eclipse E100 light microscope (Nikon, Japan) and imaged (200×). After the Masson staining procedure, the collagen fibers were bluish green in color, we analyzed and calculated the Collagen volume fraction (CVF).
Immunofluorescence
Expression of COLI, COLIII, and α-SMA in renal tissues were detected by immunofluorescence method. Renal tissues sections (3 μm) were blocked with 4% paraformaldehyde for 10 minutes and heated with EDTA antigen recovery buffer for 23 minutes. When the sections were cooled to room temperature and blocked with BSA for 30 minutes, then the immune fluorescent primary antibodies COLI, COLIII (1:300), and α-SMA (1:600) were incubated all-night at 4°C. CY3-labeled goat anti-rabbit and FITC-labeled goat anti-mouse fluorescent secondary antibodies were added dropwise and incubated for 50 minutes at room temperature, and DAPI stain was added dropwise and incubated at room temperature for 10 minutes in the dark. Three pancreas sections of each group were placed under a Eclipse E100 light microscope (Nikon, Japan).
Quantitative Real-Time PCR
Renal tissues of mice were isolated. Total RNA was extracted from the tissues by using TRIzol reagent. Total RNA samples were quantified spectrophotometrically at 260 and 280 nm, with the OD260/OD280 ratio ranging from 1.8 to 2.0. The mRNA was transcribed into cDNA according to instructions of RevertAid First Strand cDNA Synthesis Kit. The polymerase chain reaction (PCR) was performed using the Stepone plus real-time PCR device (ABI, USA) by referencing to the protocol of FastStart Universal SYBR Green Master (Rox). The cycling conditions were as follows: 95°C, 120 seconds and 45 cycles of 95°C, 10 seconds, 60°C, 34 seconds, 72°C, 30 seconds. The upstream and downstream primers (Servicebio Company, Wuhan, China) of these genes are shown in Table 1. Levels of mRNA expression were subjected to housekeeping gene glyceraldehyde-3-phosphate dehydrogenase (GAPDH). The relative transcript levels were quantified by 2−ΔΔCt value method and the final result was exhibited as a percentage of mRNA of normal.
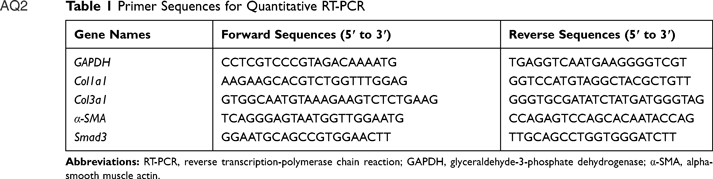 |
Table 1 Primer Sequences for Quantitative RT-PCR |
Statistical Analysis
SPSS 22.0 software (IBM Corporation, USA), was used for the statistical analyses. All data are checked by Shapiro-wilk test, P-values>0.05 are shown as the means±standard error (M±SEM). Significant differences of multiple groups were analyzed by using one-way analysis of variance (ANOVA). P-values<0.05 are shown as M (P25, P75). Significant differences of between groups were analyzed by using Mann–Whitney U-test. P<0.05 was considered to be statistically significant.
Results
Effect of FMN on Urine Volume, Food and Water Intake in db/db Mice
As shown in Table 2, compared with other groups, mice in the db/db model group showed slowed reactions, sluggish spirit, lackluster hair, and typical performance: Urine volume, food intake, and water intake have all increased (P<0.001). Compared with the model group, FMN significantly improved the symptoms (P<0.01).
 |
Table 2 Effect of FMN on Urine Volume, Food and Water Intake in db/db Mice |
Effect of FMN on Body Weight and Organ Indices in db/db Mice
As shown in Table 3, Compared with db/m, the db/db mice showed increased bodyweight and changes in the organ indices of the kidney, spleen, and liver, presenting typical obesity manifestations of type 2 diabetes. After 8 weeks of administration, FMN strongly enhanced the kidney indices (P<0.01).
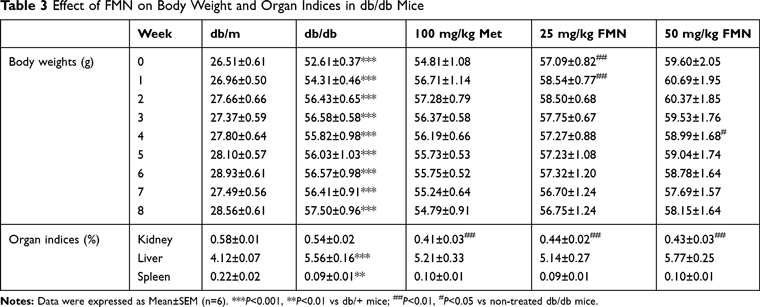 |
Table 3 Effect of FMN on Body Weight and Organ Indices in db/db Mice |
Effect of FMN on Blood Glucose, Insulin, and Blood Lipids in db/db Mice
As shown in Figure 2, the levels of FBG, FINS, IRI, TG, and TC in db/db mice were significantly higher than those in the db/m group, while the ISI level was significantly decreased (P<0.05). FMN significantly decreased the levels of FBG, FINS, IRI, TG, and TC, and increased ISI level when compared to db/db mice (P<0.05).
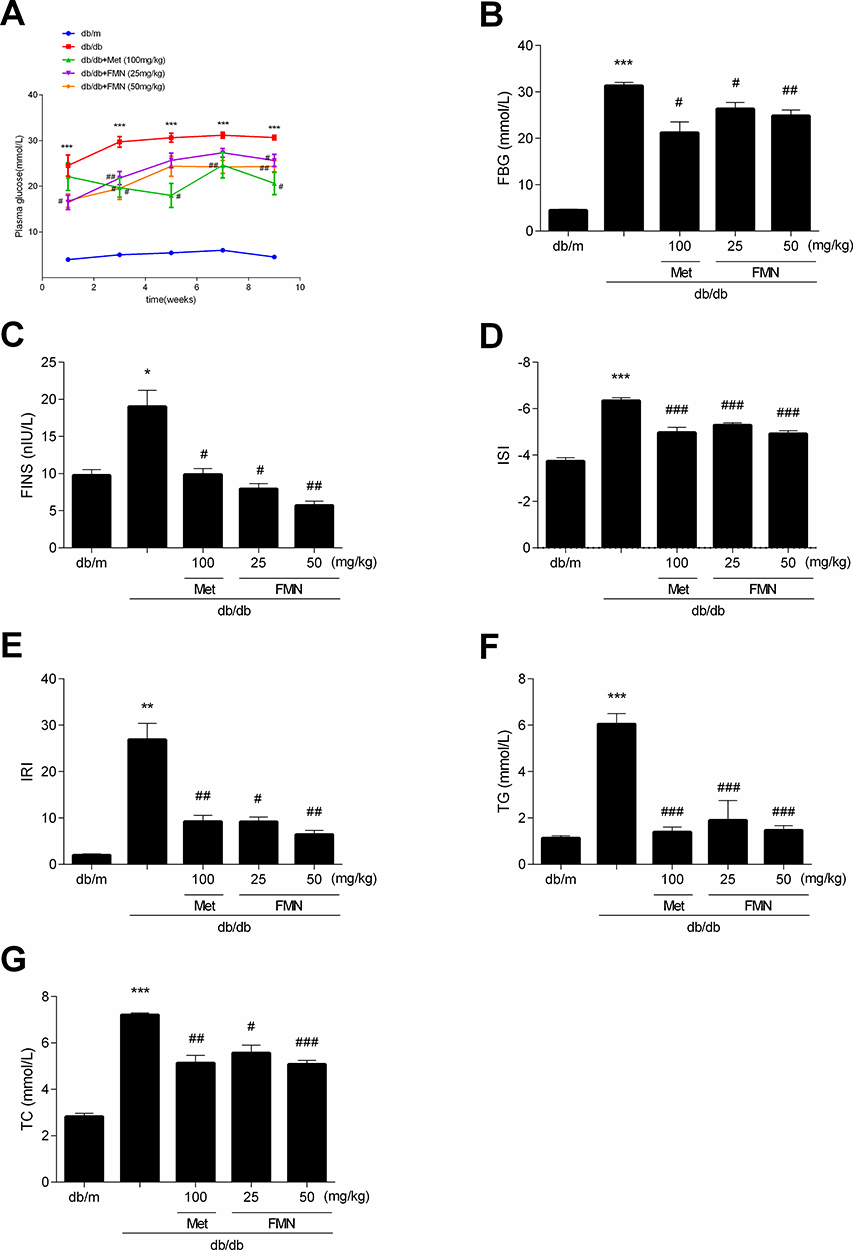 |
Figure 2 Effect of FMN on FBG (A, B), FINS (C), ISI (D), IRI (E), TG (F), and TC (G) in db/db mice. Data were expressed as Mean±SEM (n=6). ***P<0.001, **P<0.01, *P<0.05 vs db/+ mice; ###P<0.001, ##P<0.01, #P<0.05 vs non-treated db/db mice. |
Effect of FMN on Renal Function in db/db Mice
As shown in Figure 3, the levels of 24 hour urine protein, Ucr, BUN, Scr, and ACR in db/db mice were significantly higher than those in the db/m group, and BUN/Scr levels were significantly decreased (P<0.05). FMN significantly decreased the levels of 24-hour urine protein, Ucr, BUN, Scr, and ACR, and increased BUN/Scr levels when compared to db/db mice (P<0.05).
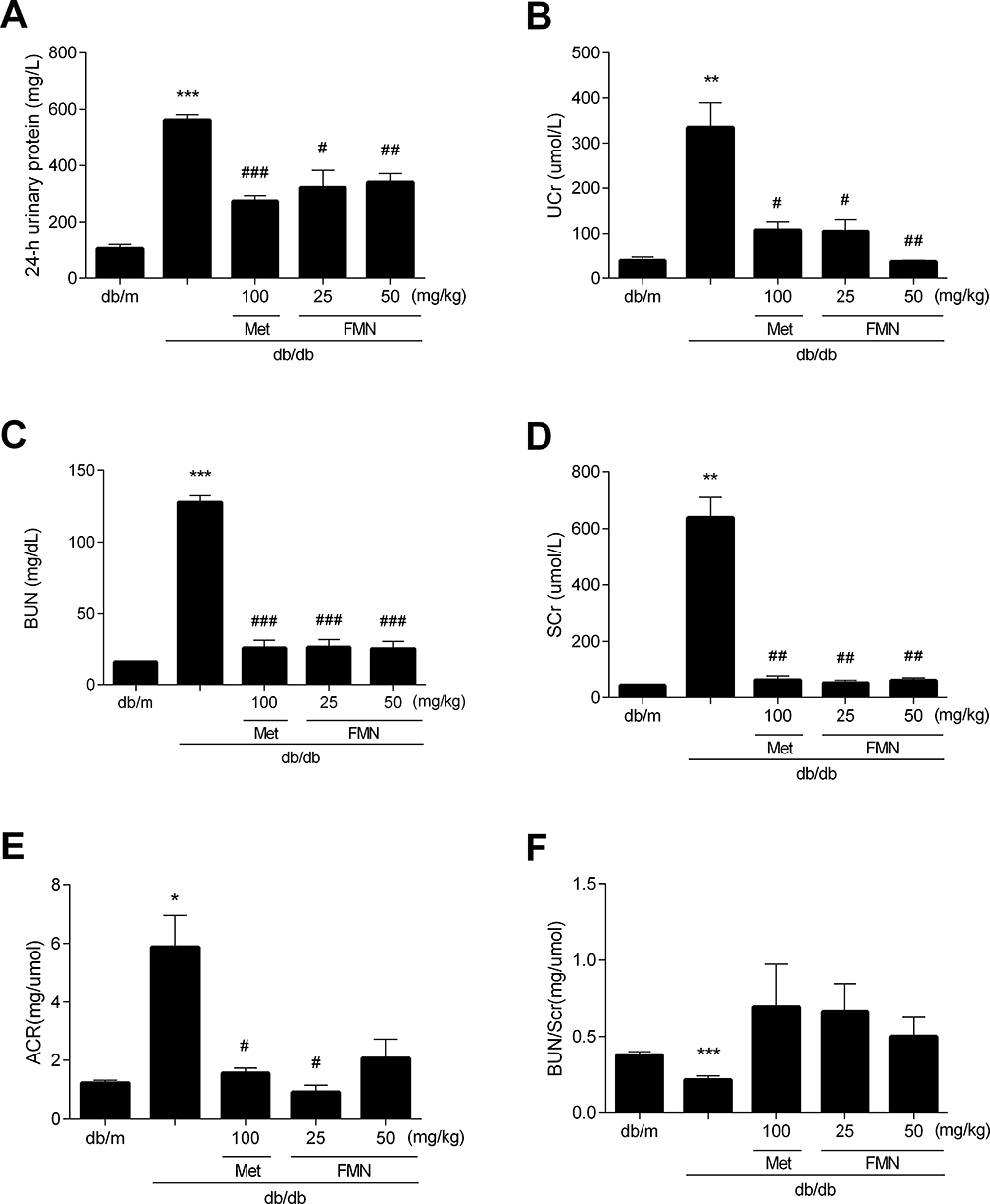 |
Figure 3 Effect of FMN on 24hUP (A), UCr (B), BUN (C), SCr (D), ACR (E), and BUN/SCr (F) in db/db mice. Data were expressed as Mean±SEM (n=6). ***P<0.001, **P<0.01, *P<0.05 vs db/+ mice; ###P<0.001, ##P<0.01, #P<0.05 vs non-treated db/db mice. |
Effect of FMN on Renal Histopathology in db/db Mice
As shown in Figure 4, H&E and PAS staining of the renal sections from the db/m group revealed a normal glomerulus surrounded by capsule, proximal, and distal convoluted tubules without any glycogen accumulated and inflammatory changes. While the db/db model group showed renal tubule atrophy, thickening of basement membrane, and glycogen accumulating. The FMN-treated groups exhibited a significantly-attenuated basement membrane and significantly-reduced glycogen accumulation. Compared with the db/m group, collagen volume fraction (CVF) of the db/db group exhibited a marked increase (P<0.05). Compared with the db/db group, significantly-reduced renal fibrosis was observed in the FMN-treated groups (P<0.05).
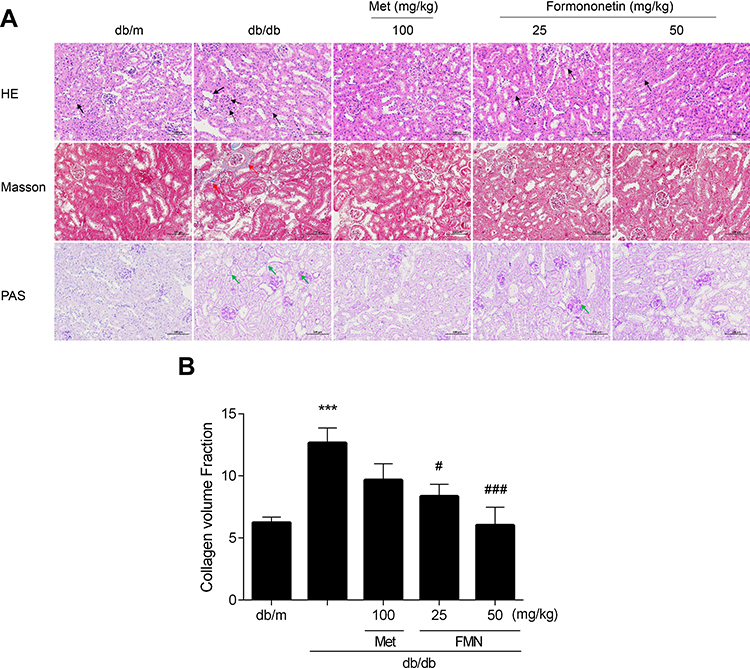 |
Figure 4 Effect of FMN on histopathologically (200X) (A), CVF (B) in db/db mice. Data were expressed as Mean±SEM (n=6). ***P<0.001 vs db/+ mice; ###P<0.001, #P<0.05 vs non-treated db/db mice. |
Effect of FMN on Renal SOD, MDA, GSH-Px, and CAT in db/db Mice
As shown in Figure 5, the level of MDA in db/db model mice was significantly higher than those in the db/m group, SOD and GSH-Px levels were significantly decreased (P<0.05). FMN significantly decreased the level of MDA, and increased SOD, GSH-Px, and CAT levels when compared to db/db mice (P<0.05).
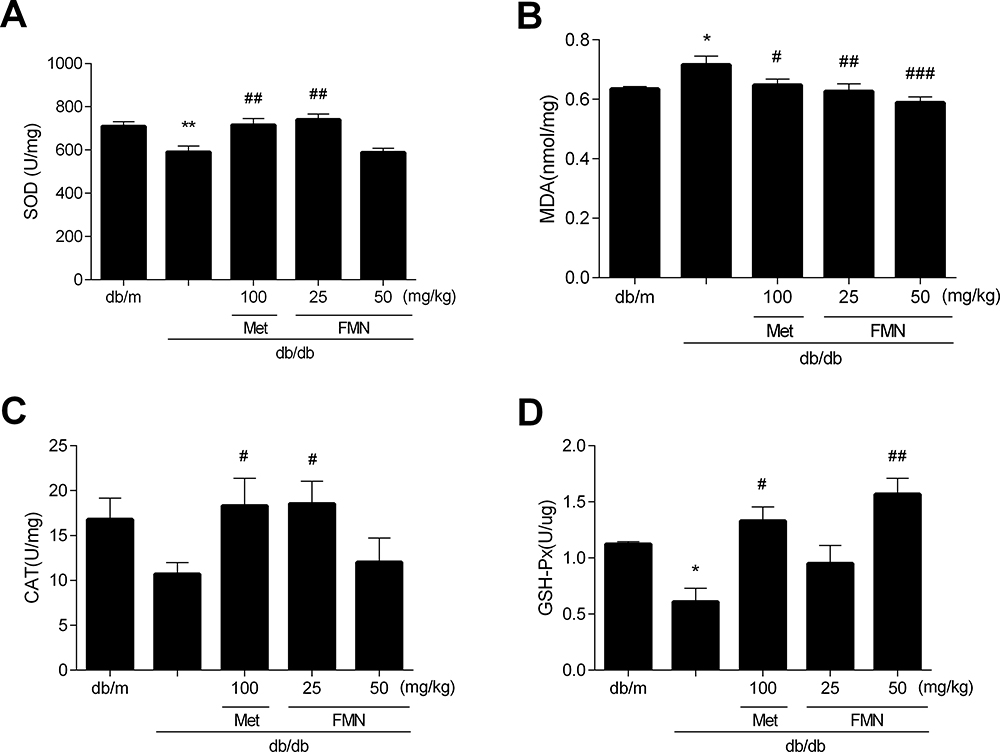 |
Figure 5 Effect of FMN on SOD (A), MDA (B), GSH-Px (C), CAT (D) in db/db mice. Data were expressed as Mean±SEM (n=6). **P<0.01, *P<0.05 vs db/+ mice; ###P<0.001, ##P<0.01, #P<0.05 vs non-treated db/db mice. |
Effect of FMN on Renal COLI, COLIII, and α-SMA Expression in db/db Mice
Renal COLI was detected by immunofluorescence labeling. Renal COLIII and α-SMA were detected by immunofluorescence double labeling. In Figure 6, the expressions of COLI, COLIII and α-SMA in the db/db model mice were significantly higher than those in the db/m group (P<0.001). FMN significantly decreased the level of COLI, COLIII, and α-SMA levels, when compared to db/db mice (P<0.01).
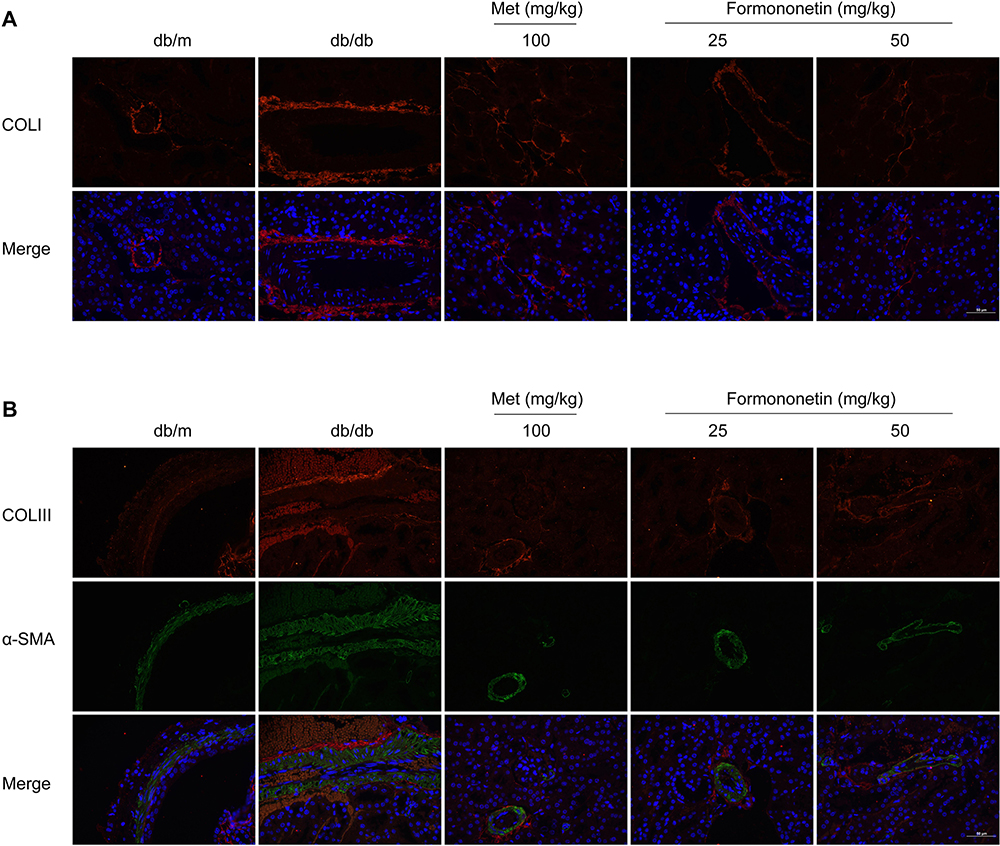 |
Figure 6 Continued. |
 |
Figure 6 The fluorescent immunogram (A, B), the expressions of COLI (C), COLIII (D), and α-SMA (E) of pancreas tissues on db/db mice between different study groups. Data were expressed as Mean±SEM (n=3). ***P<0.001 vs db/+ mice; ###P<0.001, ##P<0.01 vs non-treated db/db mice. |
Effect of FMN on Renal COL1a1, COL3a1, α-SMA, and Smad3 mRNA Expressions in db/db Mice
As shown in Figure 7, the level of COL1a1, COL3a1, α-SMA, and smad3 mRNA expressions in a db/db model mice were significantly higher than those in the db/m group (P<0.01). FMN significantly decreased the level of COL1a1, COL3a1, α-SMA, and smad3 mRNA expressions when compared to db/db mice (P<0.01).
 |
Figure 7 Effect of FMN on COL1a1 (A), COL3a1 (B), α-SMA (C), and smad3 (D) in db/db mice. Data were expressed as M(P25, P75) (n=3). **P<0.01 vs db/+ mice; ##P<0.01 vs non-treated db/db mice. |
Discussion
In the present study, db/db mice were used as experimental animal models, which have been reported under a state of high sugar status prone to kidney disease and, with an increase of blood sugar, the kidney becomes further aggravated.34 Combined with hyperglycemia and insulin resistance during the development of DN, oxidative stress, and inflammation are reported to be involved in the processes of tubular fibrosis and mesangial expansion.35
In this study, compared with NM, FBG, FINS, IRI, TG, TC, BUN, and Scr levels of MD were significantly increased, ISI level was significantly reduced of MD, and 24-hour urine protein and Ucr of urine increased of MD, which suggests that the mode of DN is successfully replicated.36 Compared with MD, oral administration of FMNH and FMNL significantly improved the abnormalities of the above indicators. Moreover, FMN significantly decreased the production and deposition of collagen and ameliorated the fibrous hyperplasia and glycogen accumulated in the kidney of db/db mice.
Oxidative stress is involved in the pathogenesis of DN, which is the basis of EMT. MDA level reflects the ability of the body to resist oxidative stress,37 and oxidative damage can be prevented by enhancing the activities of antioxidant enzymes, including CAT, GSH-Px, and SOD, which strengthen the response of the antioxidant defense system.38 In this study, Formononetin significantly decreased the level of MDA, and increased SOD,GSH-Px, and CAT levels. The results suggested that FMN has antioxidant effects.
In the process of renal fibrosis of DN, renal mesenchymal fibroblasts and renal tubular epithelial cells can be transformed into myofibroblasts, which are thought to be the primary source of ECM. α-SMA is often used as a surface marker for myofibroblasts.39 Normally, over-activation of α-SMA protein may be aggravated in renal fibrosis.40,41 Smad3 directly binds to the promoter region to activate COLI and COLIII and promotes ECM synthesis, which is involved in the process of renal fibrosis.42 In this study, FMN can suppress the mRNA expressions of smad3, COL1a1, and COL3a1, and protein expressions of smad3, COLI, and COLIII in the kidney of DN mice. The results suggested that FMN play a role in the renoprotective effect by suppressing smad3 expression.
Conclusion
In summary, FMN can significantly improve the glucolipid metabolism, reduce the oxidative stress, alleviate the renal fibrosis, and protect the renal function of DN in db/db mice. Meanwhile, protein and mRNA expression of smad3 and related regulatory factors of extracellular matrix deposition were significantly suppressed. The mechanism might be mediated by suppressing smad3 expression.
Ethics Statement
All animal care and experimental procedures complied with guidelines approved by the Animal Ethics Committee of Guangzhou University of Chinese Medicine (No.00222199). Animal studies are reported in compliance with the ARRIVE guidelines.43
Acknowledgments
This work was supported by the characteristic innovation project of Guangdong Province Office of Education [2019KTSCX023].
Disclosure
The authors report no conflicts of interest in this work.
References
1. Lovisa S, Zeisberg M, Kalluri R. Partial epithelial-to-mesenchymal transition and other new mechanisms of kidney fibrosis. Trends Endocrinol Metab. 2016;27(10):681–695. doi:10.1016/j.tem.2016.06.004
2. Marshall CB. Rethinking glomerular basement membrane thickening in diabetic nephropathy: adaptive or pathogenic? Am J Physiol Renal Physiol. 2016;311(5):F831–F843. doi:10.1152/ajprenal.00313.2016
3. Liu YW, Hao YC, Chen YJ, et al. Protective effects of sarsasapogenin against early stage of diabetic nephropathy in rats. Phytother Res. 2018;32(8):1574–1582. doi:10.1002/ptr.6088
4. Sun J, Wang Y, Cui W, et al. Role of epigenetic histone modifications in diabetic kidney disease involving renal fibrosis. J Diabetes Res. 2017;2017:7242384. doi:10.1155/2017/7242384
5. Wei Z, Cao J, Zhang X, Yin D, Xu D, Lu G. EPA attenuates epithelial-mesenchymal transition and fibrosis through the TGF-β1/Smad3/ILK pathway in renal tubular epithelial HK-2 cells by up-regulating miR-541. Int J Clin Exp Pathol. 2019;12(7):2516–2525.
6. Jin Z, Gu C, Tian F, Jia Z, Yang J. NDRG2 knockdown promotes fibrosis in renal tubular epithelial cells through TGF-β1/Smad3 pathway. Cell Tissue Res. 2017;369(3):603–610. doi:10.1007/s00441-017-2643-7
7. Feng M, Tang PM, Huang XR, et al. TGF-β mediates renal fibrosis via the Smad3-Erbb4-IR long noncoding RNA axis. Mol Ther. 2018;26(1):148–161. doi:10.1016/j.ymthe.2017.09.024
8. Peng H, Li Y, Wang C, et al. ROCK1 induces endothelial-to-mesenchymal transition in glomeruli to aggravate albuminuria in diabetic nephropathy. Sci Rep. 2016;6(1):20304. doi:10.1038/srep20304
9. Zhao L, Zhao J, Wang X, et al. Serum response factor induces endothelial-mesenchymal transition in glomerular endothelial cells to aggravate proteinuria in diabetic nephropathy. Physiol Genomics. 2016;48(10):711–718. doi:10.1152/physiolgenomics.00082.2016
10. Daehn IS. Glomerular endothelial cell stress and cross-talk with podocytes in early [corrected] diabetic kidney disease. Front Med (Lausanne). 2018;5:76. doi:10.3389/fmed.2018.00076
11. Smith BN, Bhowmick NA. Role of EMT in metastasis and therapy resistance. J Clin Med. 2016;5(2):17. doi:10.3390/jcm5020017
12. Yu CH, Suriguga GM, Liu WJ, et al. High glucose induced endothelial to mesenchymal transition in human umbilical vein endothelial cell. Exp Mol Pathol. 2017;102(3):377–383. doi:10.1016/j.yexmp.2017.03.007
13. Song S, Qiu D, Luo F, et al. Knockdown of NLRP3 alleviates high glucose or TGFB1-induced EMT in human renal tubular cells. J Mol Endocrinol. 2018;61(3):101–113. doi:10.1530/JME-18-0069
14. Zhan J, Liu M, Pan L, He L, Guo Y. Oxidative stress and TGF-β1/smads signaling are involved in Rosa roxburghii fruit extract alleviating renal fibrosis. Evid Based Complement Alternat Med. 2019;2019:4946580. doi:10.1155/2019/4946580
15. Wang T, Li Y, Chen J, Xie L, Xiao T. TGF-β1/Smad3 signaling promotes collagen synthesis in pulmonary artery smooth muscle by down-regulating miR-29b. Int J Clin Exp Pathol. 2018;11(12):5592–5601.
16. Wu Y, Wang L, Wang X, Wang Y, Zhang Q, Liu W. Renalase contributes to protection against renal fibrosis via inhibiting oxidative stress in rats. Int Urol Nephrol. 2018;50(7):1347–1354. doi:10.1007/s11255-018-1820-2
17. Wu X, Guan Y, Yan J, et al. ShenKang injection suppresses kidney fibrosis and oxidative stress via transforming growth factor-β/Smad3 signalling pathway in vivo and in vitro. J Pharm Pharmacol. 2015;67(8):1054–1065. doi:10.1111/jphp.12412
18. Montorfano I, Becerra A, Cerro R, et al. Oxidative stress mediates the conversion of endothelial cells into myofibroblasts via a TGF-β1 and TGF-β2-dependent pathway. Lab Invest. 2014;94(10):1068–1082. doi:10.1038/labinvest.2014.100
19. Qiu L, Ye H, Chen L, Hong Y, Zhong F, Zhang T. Red clover extract ameliorates dyslipidemia in streptozotocin-induced diabetic C57BL/6 mice by activating hepatic PPARα. Phytother Res. 2012;26(6):860–864. doi:10.1002/ptr.3641
20. Lai PK, Chan JY, Cheng L, et al. Isolation of anti-inflammatory fractions and compounds from the root of astragalus membranaceus. Phytother Res. 2013;27(4):581–587. doi:10.1002/ptr.4759
21. Jia WC, Liu G, Zhang CD, Zhang SP. Formononetin attenuates hydrogen peroxide (H2O2)-induced apoptosis and NF-κB activation in RGC-5 cells. Eur Rev Med Pharmacol Sci. 2014;18(15):2191–2197.
22. Oza MJ, Kulkarni YA. Formononetin treatment in type 2 diabetic rats reduces insulin resistance and hyperglycemia. Front Pharmacol. 2018;9:739. doi:10.3389/fphar.2018.00739
23. Oza MJ, Kulkarni YA. Formononetin attenuates kidney damage in type 2 diabetic rats. Life Sci. 2019;219:109–121. doi:10.1016/j.lfs.2019.01.013
24. Do MH, Hur J, Choi J, Kim Y, Park HY, Ha SK. Spatholobus suberectus ameliorates diabetes-induced renal damage by suppressing advanced glycation end products in db/db mice. Int J Mol Sci. 2018;19(9):2774. doi:10.3390/ijms19092774
25. Zhou Z, Zhou X, Dong Y, Li M, Xu Y. Formononetin ameliorates high glucose-induced endothelial dysfunction by inhibiting the JAK/STAT signaling pathway. Mol Med Rep. 2019;20(3):2893–2901. doi:10.3892/mmr.2019.10512
26. Zar Kalai F, Han J, Ksouri R, Abdelly C, Isoda H. Oral administration of nitraria retusa ethanolic extract enhances hepatic lipid metabolism in db/db mice model through the modulation of balance. Food Chem Toxicol. 2014;72:247–256. doi:10.1016/j.fct.2014.07.029
27. Senturk B, Demircan BM, Ozkan AD, et al. Diabetic wound regeneration using heparin-mimetic peptide amphiphile gel in db/db mice. Biomater Sci. 2017;5(7):1293–1303. doi:10.1039/C7BM00251C
28. Pei F, Li BY, Zhang Z, et al. Beneficial effects of phlorizin on diabetic nephropathy in diabetic db/db mice. J Diabetes Complications. 2014;28(5):596–603. doi:10.1016/j.jdiacomp.2014.04.010
29. Jia ZH, Liu ZH, Zheng JM, Zeng CH, Li LS. Combined therapy of rhein and benazepril on the treatment of diabetic nephropathy in db/db mice. Exp Clin Endocrinol Diabetes. 2007;115(9):571–576. doi:10.1055/s-2007-981469
30. Eisenreich A, Leppert U. Update on the protective renal effects of metformin in diabetic nephropathy. Curr Med Chem. 2017;24(31):3397–3412. doi:10.2174/0929867324666170404143102
31. Ravindran S, Kuruvilla V, Wilbur K, Munusamy S. Nephroprotective effects of metformin in diabetic nephropathy. J Cell Physiol. 2017;232(4):731–742. doi:10.1002/jcp.25598
32. Wang J, Wang L, Zhou J, Qin A, Chen Z. The protective effect of formononetin on cognitive impairment in streptozotocin (STZ)-induced diabetic mice. Biomed Pharmacother. 2018;106:1250–1257. doi:10.1016/j.biopha.2018.07.063
33. Nie T, Zhao S, Mao L, et al. The natural compound, formononetin, extracted from astragalus membranaceus increases adipocyte thermogenesis by modulating PPARγ activity. Br J Pharmacol. 2018;175(9):1439–1450. doi:10.1111/bph.14139
34. Einbinder Y, Ohana M, Benchetrit S, et al. Glucagon-like peptide-1 and vitamin D: anti-inflammatory response in diabetic kidney disease in db/db mice and in cultured endothelial cells. Diabetes Metab Res Rev. 2016;32(8):805–815. doi:10.1002/dmrr.2801
35. Mima A. Renal protection by sodium-glucose cotransporter 2 inhibitors and its underlying mechanisms in diabetic kidney disease. J Diabetes Complications. 2018;32(7):720–725. doi:10.1016/j.jdiacomp.2018.04.011
36. Liu Y, Dai W, Ye S. The olive constituent oleuropein exerts nephritic protective effects on diabetic nephropathy in db/db mice. Arch Physiol Biochem. 2019;1–8. doi:10.1080/13813455.2019.1691603
37. Yaribeygi H, Sathyapalan T, Atkin SL, Sahebkar A. Molecular mechanisms linking oxidative stress and diabetes mellitus. Oxid Med Cell Longev. 2020;2020:8609213. doi:10.1155/2020/8609213
38. Singh P, Prasad SM. Antioxidant enzyme responses to the oxidative stress due to chlorpyrifos, dimethoate and dieldrin stress in palak (Spinacia oleracea L.) and their toxicity alleviation by soil amendments in tropical croplands. Sci Total Environ. 2018;630:839–848. doi:10.1016/j.scitotenv.2018.02.203
39. Nikolic-Paterson DJ, Wang S, Lan HY. Macrophages promote renal fibrosis through direct and indirect mechanisms. Kidney Int Suppl. 2014;4(1):34–38. doi:10.1038/kisup.2014.7
40. Zhao JH. Mesangial cells and renal fibrosis. Adv Exp Med Biol. 2019;1165:165–194.
41. Huang S, Liu F, Niu Q, et al. GLIPR-2 overexpression in HK-2 cells promotes cell EMT and migration through ERK1/2 activation. PLoS One. 2013;8(3):e58574. doi:10.1371/journal.pone.0058574
42. Bai X, He T, Liu J, et al. Loureirin B inhibits fibroblast proliferation and extracellular matrix deposition in hypertrophic scar via TGF-β/Smad pathway. Exp Dermatol. 2015;24(5):355–360. doi:10.1111/exd.12665
43. McGrath JC, Lilley E. Implementing guidelines on reporting research using animals (ARRIVE etc.): new requirements for publication in BJP. Br J Pharmacol. 2015;172(13):3189–3193. doi:10.1111/bph.12955
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.