Back to Journals » Drug Design, Development and Therapy » Volume 19
Relief from Chronic Unpredictable Mild Stress-Induced Constipation by Fluoxetine and Butyrate and Their Impact on Gut Microecology
Authors Li X, Suo J, Yao S, Tian B, Zhao Z, Wang Y, Li X, Jin D, Yang X ![]()
Received 30 July 2025
Accepted for publication 10 December 2025
Published 26 December 2025 Volume 2025:19 Pages 11675—11695
DOI https://doi.org/10.2147/DDDT.S552593
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Anastasios Lymperopoulos
Xuzhao Li,1,* Jing Suo,2,* Shiqin Yao,1,* Bowen Tian,1,* Zhifeng Zhao,3 Yanrong Wang,4 Xiaoliang Li,5 Dingding Jin,6 Xiaoran Yang7
1Department of Gastrointestinal Surgery, People’s Hospital of Ningxia Hui Autonomous Region, Ningxia Medical University, Yinchuan, Ningxia Hui Autonomous Region, 750002, People’s Republic of China; 2Medical Image Center, People’s Hospital of Ningxia Hui Autonomous Region, Ningxia Medical University, Yinchuan, Ningxia Hui Autonomous Region, 750002, People’s Republic of China; 3Department of Gastrointestinal Surgery, Xijing Hospital of Digestive Diseases, Air Force Medical University, Xi’an, 710032, People’s Republic of China; 4Department of Neurology, People’s Hospital of Ningxia Hui Autonomous Region, Ningxia Medical University, Yinchuan, Ningxia Hui Autonomous Region, 750002, People’s Republic of China; 5Department of Gastrointestinal Surgery, Xi’an International Medical Center Hospital, Xi’an, 710119, People’s Republic of China; 6Department of General Surgery, People’s Hospital of Ningxia Hui Autonomous Region, Ningxia Medical University, Yinchuan, Ningxia Hui Autonomous Region, 750002, People’s Republic of China; 7Department of Otolaryngology, Head and Neck surgery, People’s Hospital of Ningxia Hui Autonomous Region, Ningxia Medical University, Yinchuan, Ningxia Hui Autonomous Region, 750002, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Dingding Jin, Department of General surgery, People’s Hospital of Ningxia Hui Autonomous Region, Ningxia Medical University, 255 Zhengyuan North Street, Jinfeng District, Yinchuan, Ningxia Hui Autonomous Region, 750002, People’s Republic of China, Email [email protected] Xiaoran Yang, Department of Otolaryngology, head and neck surgery, People’s Hospital of Ningxia Hui Autonomous Region, Ningxia Medical University, 255 Zhengyuan North Street, Jinfeng District, Yinchuan, Ningxia Hui Autonomous Region, 750002, People’s Republic of China, Email [email protected]
Purpose: This study investigated fluoxetine (FLX) effects on chronic unpredictable mild stress (CUMS)-induced depression and constipation, and to explore the potential synergistic benefits of combining FLX with butyrate supplementation.
Methods: A mouse model of CUMS and functional constipation (FC) was established. Behavioral assessments, fecal parameters, colonic transit time, and histological analysis were performed. Serotonin signaling, gastrointestinal hormones, intestinal barrier function, and inflammatory markers were measured. Gut microbiota composition was assessed using 16S rRNA sequencing, and fecal short-chain fatty acid (SCFA) levels were quantified by gas chromatography-mass spectrometry.
Results: FLX significantly alleviated depressive-like behaviors (40% reduction in immobility time) and improved gastrointestinal motility (35% increase in fecal pellet output) in CUMS-FC mice. FLX restored serotonin signaling (2.5-fold increase in 5-HT levels), enhanced intestinal barrier integrity (60% upregulation of ZO-1/Occludin), and increased gut microbial diversity (Shannon index increased by 30%). FLX elevated butyrate-producing bacteria and fecal butyrate levels by 45%. Notably, the novel FLX-butyrate combination produced synergistic effects, achieving 65% improvement in gut motility, 70% reduction in pro-inflammatory cytokines (IL-1β, TNF-α, IL-6), and 80% restoration of intestinal barrier function compared to FLX alone.
Conclusion: FLX alleviates CUMS-induced depression and constipation through restoring serotonin signaling, modulating gut microbiota toward SCFA production, and improving intestinal barrier function. The novel synergistic enhancement achieved by FLX-butyrate combination demonstrates superior therapeutic efficacy, highlighting its potential as an innovative dual-targeting strategy for gut-brain axis disorders.
Keywords: fluoxetine, butyrate, CUMS, constipation, gut-brain axis
Introduction
Chronic unpredictable mild stress (CUMS) is widely recognized for inducing a variety of depression-like behaviors and gastrointestinal dysfunction, such as constipation.1,2 These symptoms are strongly linked to dysregulation within the gut–brain axis, a bidirectional communication system between the central nervous system and the gastrointestinal tract.3 Research has increasingly demonstrated the impact of this axis on both mental and digestive health.4
Fluoxetine (FLX), a well-known selective serotonin reuptake inhibitor (SSRI), is often prescribed to treat depression.5 Beyond its central nervous system effects, FLX has been observed to influence gut–brain axis comorbidities, particularly in the context of depression.6,7 The possible effects of FLX on alleviating CUMS-induced depression and constipation are of particular interest, as they highlight the therapeutic potential of this medication beyond traditional psychiatric applications.8
The composition of gut microbiota plays a crucial role in the development and management of CUMS-related symptoms.3,9 Alterations in gut microbiota composition have been associated with serotonin signaling, balanced gastrointestinal hormone levels, enhanced intestinal barrier integrity, and alleviated colonic mucosal damage.10–12 Notably, butyrate-producing bacteria contribute to these beneficial effects by enhancing fecal short-chain fatty acid (SCFA) levels, which are integral to maintaining gut health.13 Enhancing levels of butyrate-producing bacteria are known to increase the levels of serotonin in the prefrontal cortex (PFC) of mice and prevent depression from chronic stress.14
Butyrate supplementation has garnered attention for its potential to improve gut health and mitigate symptoms of depression.15,16 As a key SCFA, butyrate supports gut barrier integrity, reduces inflammation, and modulates neurotransmitter production. These beneficial effects motivate an exploration of the synergistic impact of butyrate and FLX on gut microecology and overall health during CUMS. We propose that FLX and butyrate supplementation will synergistically improve gut microecology and alleviate both depressive and gastrointestinal symptoms induced by CUMS.
This study aims to investigate the effects of FLX on CUMS-induced depression and constipation, with a particular focus on exploring the novel therapeutic potential of combining FLX with butyrate supplementation. While previous studies have examined FLX or butyrate individually for depression or gastrointestinal disorders, the synergistic effects of their combination in addressing both neurological and gastrointestinal symptoms simultaneously remain unexplored. To this end, behavioral assessments, fecal pellet output, colonic transit time, and colonic histology were analyzed to evaluate depressive and gastrointestinal symptoms in a mouse model of CUMS and functional constipation (FC). Additionally, serotonin signaling, gastrointestinal hormone levels, intestinal barrier function, and inflammatory markers were measured. Gut microbiota composition was assessed using 16S rRNA sequencing, and fecal SCFA levels were quantified. The unique contribution of this study lies in systematically investigating the synergistic effects of FLX-butyrate combination therapy, which represents a novel dual-targeting approach that addresses both the central nervous system dysfunction and gut microbiota dysbiosis characteristic of CUMS-related disorders. This integrated therapeutic strategy may offer superior efficacy compared to monotherapy, providing new insights into managing the complex gut-brain axis dysfunction in stress-related disorders with gastrointestinal comorbidities.
Materials and Methods
Experimental Materials and Animal Experiment Design
FLX (fluoxetine hydrochloride) and loperamide were obtained from Thermo Fisher Scientific (Waltham, MA, USA) and dissolved in 0.03% sodium carboxymethyl cellulose. Both FLX and loperamide were administered at 10 mg/kg body weight (BW), following the optimal dosage reported in previous research.1,17 The loperamide-induced constipation model was chosen as it reliably reproduces the gastrointestinal dysfunction commonly observed in depression patients, including delayed intestinal transit and reduced fecal water content. This model has been validated in the context of stress-induced comorbidity studies, as loperamide’s μ-opioid receptor agonist activity mimics the opioid-mediated constipation frequently seen in clinical depression with gastrointestinal symptoms.18,19
In this study, 40 specific pathogen-free (SPF) female BALB/c mice, aged 6–7 weeks and weighing 18 ± 2 g, were obtained from SPF (Beijing) Biotechnology Co., Ltd., Beijing, China, and housed in a SPF facility with a controlled environment of 23°C and a 12-hour light-dark cycle. The mice had unrestricted access to food and water. All animal experiments were conducted in accordance with the Guidelines for the Care and Use of Laboratory Animals and were approved by the Ethics Committee of the People’s Hospital of Ningxia Hui Autonomous Region (license number: 2022 NZR-196). Following a 3-day acclimation period, the mice were randomly assigned to four groups (n = 6 per group) using a random number generator: (1) control (normal saline); (2) Model group (CUMS + FC with vehicle treatment); (3) Model + FLX group (CUMS+FC with fluoxetine 10 mg/kg/day, i.g).; (4) Model + FLX + Butyrate group (CUMS+FC with fluoxetine 10 mg/kg/day plus sodium butyrate 200 mg/kg/day, i.g).
For butyrate supplementation, sodium butyrate (Sigma-Aldrich, # B5887) was freshly dissolved in sterile saline to a concentration of 20 mg/mL. The solution was administered via oral gavage at 200 mg/kg body weight once daily at 9:00 am, 30 minutes after fluoxetine administration. The dose was selected based on previous studies demonstrating optimal effects on intestinal barrier function and microbiota modulation without adverse effects. The Model and Control groups received an equivalent volume of saline (10 mL/kg). The treatment continued for 14 consecutive days, starting from the third week of CUMS exposure. All mice were maintained on standard rodent maintenance diet and had ad libitum access to food and water throughout the study. No dietary restrictions were applied as part of the CUMS protocol. Body weight was monitored twice weekly to adjust dosing accordingly. All treatments were administered for the final 2 weeks of the 4-week CUMS protocol. Behavioral tests were conducted 24 h after the last treatment. Following behavioral assessments, mice were anesthetized with isoflurane (3–4% for induction, 1.5–2% for maintenance) via inhalation. Mice were then euthanized by cervical dislocation under deep anesthesia, and tissue samples were immediately collected, snap-frozen in liquid nitrogen, and stored at −80°C for subsequent analyses. All procedures were performed in accordance with institutional guidelines for humane animal euthanasia.
CUMS Procedure
The CUMS procedure was performed on the mice to simulate the social and environmental pressures experienced by patients with depression, incorporating various stressors over 6 weeks. The stressors included 24-hour water and food deprivation, empty water bottles, 24-hour wet litter, noise (>80 dB) for 3 hours, tail-clamping for 30 minutes, 45° cage tilt for 24 hours, grouped housing, soiled cage for 24 hours, stroboscopic lighting for 24 hours, restricted access to food, 5-minute cold swimming (at 15°C), 5-minute heat exposure (at 45°C), 17-hour white noise, 1-minute tail pinch (1 cm from the end of the tail), 3-hour physical restraint, overnight illumination, 17-hour exposure to a strange object, and turning the cage upside down day and night. One or two of these stressors were applied daily, ensuring no repetition within 3 days, while control group animals were left undisturbed except for routine procedures.
Barnes Maze Test
The Barnes Maze Test was performed to assess spatial learning and memory in mice, following the established methodology.20 All behavioral assessments and data analyses were conducted by investigators blinded to the experimental groups. Spatial heat maps, generated using EthoVision XT (Noldus Information Technology, Wageningen, Netherlands), visualized the time mice spent in different areas of the maze, with warmer colors indicating longer durations. Errors were defined as nose pokes into incorrect holes before locating the target hole. Escape latency, which represented the time taken to find the target hole, was recorded. Path length, the total distance traveled by the mice within the maze, was measured using automated tracking software. Furthermore, the percentage of time spent in the target zone and the duration taken to initially approach the target hole were also calculated.
Open Field Test
The entire experimental process was conducted in a quiet and dark environment. Before the experiment, the mice were acclimated to the experimental environment for 1–2 hours. Thirty minutes post-administration, the mice were placed in a square arena (50 × 50×50 cm). The number of horizontal crawls and uprights performed by the mice within 5 minutes was recorded by an investigator blinded to the experimental groups, following previously established protocol.21 After each trial, the inner walls and the bottom of the arena were thoroughly cleaned to remove any residual odors from the previous mice.
Forced Swim Test
The forced swim test (FST) was conducted by gently placing the mice into cylindrical tanks 80 cm high with a diameter of 40 cm containing water maintained at a temperature of 23°C, according to previously described methods.22 The mice were allowed to remain in the tanks for 6 minutes. The immobility time during the final 5 minutes was recorded by assistants who were blinded to the experimental design.
Assessment of General Condition and Fecal Parameters
Throughout the experiment, BW and food intake were recorded twice a week, and daily observations of physiological and fecal status were conducted. All assessments were performed by investigators blinded to the experimental groups. The time of the first black stool (FBS) was measured by individually placing mice in clean cages and recording the time each mouse excreted its FBS. Fecal water content (FWC) was determined by measuring the wet weight of the feces immediately after defecation, then drying the feces thoroughly in an oven and measuring the dry weight. FWC was calculated as follows: FWC (%) = (wet weight − dry weight)/wet weight × 100. Gastrointestinal motility was measured as the number of fecal pellets excreted within 6 hours.
Quantitative PCR (RT-qPCR) for Inflammatory Cytokines
The mRNA expression levels of IL-1β, TNF-α, IL-6, and IL-10 in colon tissues were quantified by qRT-PCR to assess the inflammatory response. Primers were designed according to sequences obtained from BLAST and Primer Premier 5 and synthesized by Sangon Biotech Co., Ltd. (Shanghai, China). Total RNA was extracted from colon tissue by using an RNA extraction kit (Invitrogen, #15596026), followed by cDNA synthesis with a First Strand cDNA Synthesis Kit (Thermo scientific, #K1622) and a Mastercycler Gradient Thermal Cycler (Eppendorf, Hamburg, Germany). RT-qPCR was conducted with PowerUp SYBR Green PCR Master Mix (Thermo Fisher Scientific, #A25742) using a CFX96 Real-Time System (Bio-Rad, Hercules, CA, USA). GAPDH was used to determine relative mRNA levels, which were calculated using the 2−Δ Δ Ct method.
Serum Biochemical Analysis
Following anesthesia with isoflurane, blood was collected from the enucleated eyeballs of the mice. The collected blood was then centrifuged at 4°C and 1500 × g for 15 minutes using a 5804R desktop refrigerated centrifuge (Eppendorf, Hamburg, Germany). The resulting serum samples were stored at −80°C. The serum levels of Gastrin (Gas, #JN20306), Somatostatin (SS, #JN17252), Acetylcholinesterase (AChE, #JN18891), Serotonin (5-HT, #JN7486), Substance P (SP, #JN17198), Motilin (MTL, #JN19674), and Vasoactive Intestinal Peptide (VIP, #JN18994) were measured following the instructions of an enzyme-linked immunosorbent assay (ELISA) kit (Elisa Biotech Co., Ltd., Shanghai, China).
Small Intestinal Transit Rate
Whole intestines were collected to measure the distance traveled by the activated charcoal. Measurements were performed by an investigator blinded to the experimental groups. The gastrointestinal transit rate (GTR) was determined by the following equation GTR (%) = (distance charcoal traveled/total intestinal length) × 100.
Histological Analysis
Colon tissue samples were fixed using 4% paraformaldehyde, followed by embedding in paraffin, and sliced into sections with a thickness of 4µm. The fixed sections were deparaffinized with xylene, rehydrated through a series of graded ethanol solutions, and then rinsed with distilled water. For hematoxylin and eosin (H&E), the sections were stained with hematoxylin to color the nuclei blue and then stained with eosin to color the cytoplasm and extracellular matrix pink. For Alcian Blue-Periodic Acid-Schiff (AB-PAS) staining, the sections were stained with Alcian Blue to color acidic mucins blue, then oxidized with periodic acid to convert certain tissue components into aldehydes. Finally, the tissue sections were treated with Schiff reagent to stain neutral mucins magenta. All histological evaluations and scoring were conducted by investigators blinded to the experimental groups.
Immunohistochemistry (IHC) and Immunofluorescence (IF) Staining
IHC staining was conducted using paraffin-embedded colon tissue samples (5 µm thick). The samples were placed in an oven at 60°C for 4 hours. They were then deparaffinized with xylene and rehydrated with graded ethanol (100–50%). The sections were then boiled in water at 100°C for 10 minutes and washed three times with tris-buffered saline with Tween (TBST). The slides were then blocked with 3% hydrogen peroxide for 20 minutes at room temperature. After washing three times with TBST, the sections were incubated with a primary antibody for C-Kit (Abcam, #ab317843, 1: 1000) for 16 hours. The sections were washed three times with TBST and then incubated with a secondary antibody (Beijing Biosynthesis Biotechnology Co. Ltd., Beijing, China) for 30 minutes at 37°C, followed by DAB and hematoxylin staining. For IF staining, the slides were incubated with primary antibodies against ZO-1 (Abcam, #ab307799, 1: 500) and Occludin (Abcam, #ab216327, 1: 100) at 4°C overnight. Subsequently, all slides were incubated with fluorescent-conjugated goat anti-chicken IgG, followed by DAPI staining. Images were obtained using an Olympus BX53 fluorescence microscope and analyzed with ImagePro Plus 6.0 software (Rockville, MD, USA). All immunostaining quantification and image analyses were performed by investigators blinded to the experimental groups.
Determination of SCFA Content
Mouse feces (200 mg) were first added to 1 mL of ultrapure water, followed by acidification with 10 µL of concentrated hydrochloric acid and homogenized for 3 min. The homogenize was then centrifuged at 10,000 × g for 10 min. The levels of acetic acid, propionic acid, butyric acid, isobutyric acid, valeric acid, and isovaleric acid were determined by gas chromatography using an Agilent GC-7890A (Agilent Technologies Inc., Santa Clara, CA, USA). The initial temperature was 100°C for 5 min. This was then increased at 10°C/min to 250°C. The final temperature was maintained for 12 min.
Analysis of the Gut Microbiota by 16S rRNA Gene Sequencing
DNA Extraction and Sequencing
Mouse fecal DNA was extracted using the QIAamp DNA Stool Mini Kit (Qiagen, Cat# 51504) following the manufacturer’s protocol. DNA concentration and purity were assessed using a NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific) and agarose gel electrophoresis. The V4 region of the 16S rRNA gene was amplified using universal primers 515F (5’-GTGCCAGCMGCCGCGGTAA-3’) and 806R (5’-GGACTACHVGGGTWTCTAAT-3’). PCR amplification was performed in a 50 µL reaction containing 25 µL 2× Premix Taq (Takara, Cat# RR003), 1 µL of each primer (10 µM), 10 ng template DNA, and nuclease-free water. PCR conditions were: initial denaturation at 98°C for 30s; 30 cycles of 98°C for 10s, 50°C for 30s, and 72°C for 30s; and final extension at 72°C for 5 min. PCR products were purified using AMPure XP beads (Beckman Coulter, Cat# A63881) and quantified using a Qubit 3.0 Fluorometer (Thermo Fisher Scientific). Purified amplicons were pooled in equimolar concentrations and sequenced on an Illumina HiSeq 2500 platform generating 2×250 bp paired-end reads.
Bioinformatics Analysis
Raw sequencing data were processed using QIIME2 (version 2023.5). Paired-end reads were imported and quality filtered using the DADA2 plugin with the following parameters: forward and reverse reads were truncated at 240 bp and 200 bp respectively, with a maximum expected error of 2. DADA2 was used to denoise sequences, merge paired-end reads, and remove chimeric sequences, generating amplicon sequence variants (ASVs). This approach provides single-nucleotide resolution without requiring similarity-based clustering used in traditional OTU methods. Taxonomy was assigned to ASVs using the classify-sklearn naïve Bayes classifier trained on the SILVA 138 99% reference database, with a confidence threshold of 0.7. ASVs assigned to chloroplast, mitochondria, or unassigned at the kingdom level were removed from the analysis.
Data Normalization and Diversity Analysis
To account for uneven sequencing depth across samples, data were rarefied to 10,000 sequences per sample, which retained >95% of samples while maintaining adequate sequencing depth for diversity analyses. Alpha diversity metrics including Chao1 (richness), Shannon index (diversity), and Faith’s phylogenetic diversity were calculated using the diversity plugin in QIIME2. Beta diversity was assessed using both Bray-Curtis dissimilarity (abundance-based) and weighted UniFrac distance (phylogeny-based). Principal coordinate analysis (PCoA) was performed to visualize beta diversity patterns, and permutational multivariate analysis of variance (PERMANOVA) with 999 permutations was used to test for significant differences between groups.
Western Blotting
Proteins were extracted from homogenized mouse PFC using cold RIPA buffer containing 0.1% phenylmethylsulfonyl fluoride. The protein concentration was measured using a Coomassie blue-based assay. An equal concentration of proteins was separated using SDS-polyacrylamide gel electrophoresis and then transferred onto a polyvinylidene fluoride membrane. The membrane was blocked with 5% skimmed milk and then incubated at 4°C overnight specific primary antibodies for anti-mTOR (Cell signaling, #2972,1:1000), anti-p-mTOR (Cell signaling, #2971,1:1000), anti-BDNF antibody (Cell signaling, #47808, 1:1000), anti-PSD-95 (Cell signaling, #2507,1:1000), anti-Synapsin 1 (Cell signaling, #5297,1:1000), anti-GluR1 (Cell signaling, #13185,1:1000), anti-IL-1β (Cell signaling, #12242,1:1000), anti-TNF-α (Cell signaling, #3707,1:1000), anti-IL-6 (Cell signaling, #12912,1:1000), anti-β-Actin (Cell signaling, #4967,1:1000), and anti-GAPDH (Cell signaling, #2118,1:1000). After washing with TBST, the membranes were incubated with horseradish peroxidase conjugated secondary antibody (1:12,000) for 2 hours at RT. Antibody-reactive bands were visualized using enhanced chemiluminescence detection reagents (Tanon Science & Technology Co., Ltd., Shanghai, China).
Statistical Analysis
Data normality was assessed using the Shapiro–Wilk test. All normally distributed data are presented as mean ± SD. For single time-point measurements with multiple groups (eg, behavioral tests, molecular markers, histological scores), one-way ANOVA was performed followed by Newman-Keuls post-hoc test for pairwise comparisons. For measurements involving repeated observations over time (eg, body weight changes, food consumption, daily fecal pellet counts), two-way repeated measures ANOVA was used with treatment group as between-subjects factor and time as within-subjects factor, followed by Bonferroni post-hoc tests for multiple comparisons. Pearson correlation analysis was used to evaluate linear relationships between continuous variables. For microbiome analyses, multiple testing correction was performed using the Benjamini-Hochberg procedure, with a false discovery rate (FDR) threshold of 0.1. P-values of less than 0.05 were considered statistically significant for all other analyses. All statistical analyses were performed using GraphPad Prism 9.0 (GraphPad Software, San Diego, CA, USA).
Results
FLX Ameliorates CUMS-Induced Behavioral Deficits and Constipation Symptoms in Mice
We conducted comprehensive behavioral, physiological, and cognitive assessments of mice across experimental groups. Our findings revealed that the BW of mice increased and remained consistent across all groups over 7 weeks (Figure 1A). The number of fecal pellets excreted within 6 hours was used as an indicator of gastrointestinal motility. Gastrointestinal motility was significantly slower in the CUMS and FC groups compared to the control group. However, treatment with FLX increased the number of fecal pellets (Figure 1B). Exploratory behavior, self-care, and locomotor activity were higher in the control group compared to the FC and CUMS groups, with the CUMS + FC group being the most adversely affected. However, treatment with FLX seemed to alleviate some of these symptoms (Figure 1C–E). The Barnes maze heat map supported these initial findings. The CUMS + FC group had the highest number of errors in the maze and took the longest to reach the target. FLX treatment improved the performance of mice in the CUMS + FC group (Figure 1F–K). Overall, our study demonstrated that FLX could enhance defecation, behavioral indicators, and OFT parameters in CUMS-induced mice.
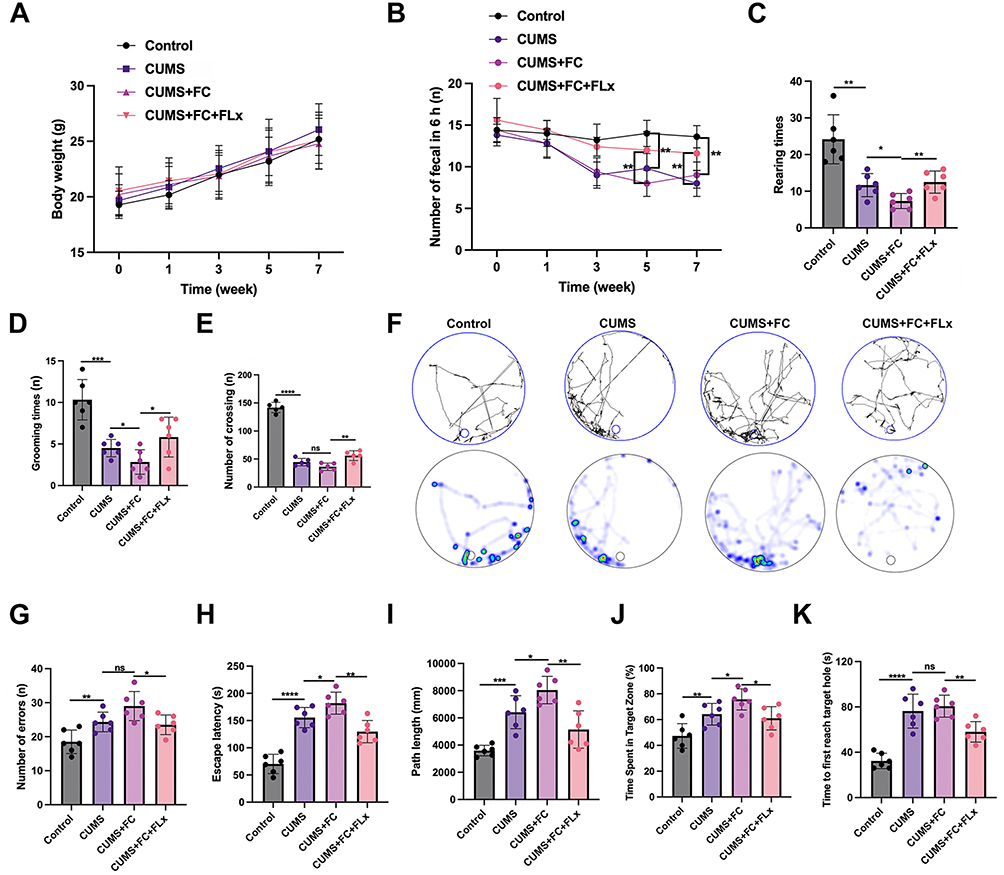 |
Figure 1 FLX ameliorates CUMS-induced behavioral deficits and constipation symptoms in mice. (A) Body weight (BW) of mice across different groups. (B) Number of fecal pellets excreted within 6 hours as an indicator of gastrointestinal motility. (C) Rearing time, representing exploratory behavior. (D) Grooming time, reflecting self-care and anxiety-related behavior. (E) Number of crossings, indicating locomotor activity. (F) Barnes maze heat map: warmer colors represent increased time spent in specific locations. (G) Number of errors made during the Barnes maze test. (H) Escape latency (seconds) in the Barnes maze. (I) Path length (millimeters) traveled during the Barnes maze test. (J) Time spent in the target zone (%) in the Barnes maze. (K) Time to first reach the target hole (seconds) in the Barnes maze. Data are presented as mean ± SD, n = 6. Statistical significance for (A) and (B) was determined by two-way repeated measures ANOVA followed by Bonferroni post-hoc tests. Statistical significance for (C–K) was determined by one-way ANOVA followed by Newman-Keuls post-hoc tests. * p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001, ns, no significance. |
FLX Improves Constipation-Related Parameters in CUMS-Induced Mice
To investigate the effects of FLX on constipation-related parameters in CUMS-induced mice in greater detail we measured constipation-related parameters. A model group of CUMS + FC mice were treated with FLX for 4 weeks and compared to untreated mice. Representative images show that the fur texture, body posture, and general condition of mice in the model group were poor compared to the control group or the model mice treated with FLX (Figure 2A). BW and food intake improved in the model upon treatment with FLX (Figure 2B and C). Gastrointestinal motility represented as the time to the first black stool defecation and the number of fecal pellets at 6 hours was substantially improved by FLX, although they did not reach control levels (Figure 2D–F). FWC, the transit rate of charcoal meal through the gastrointestinal tract, and intestinal propulsion distance significantly improved with FLX and returned to similar levels of the control (Figure 2G and H). The weight of the liver and kidney improved in response to FLX also (Figure 2I and J). Collectively, these data confirm that FLX enhances gastrointestinal health in the CUMS mouse model.
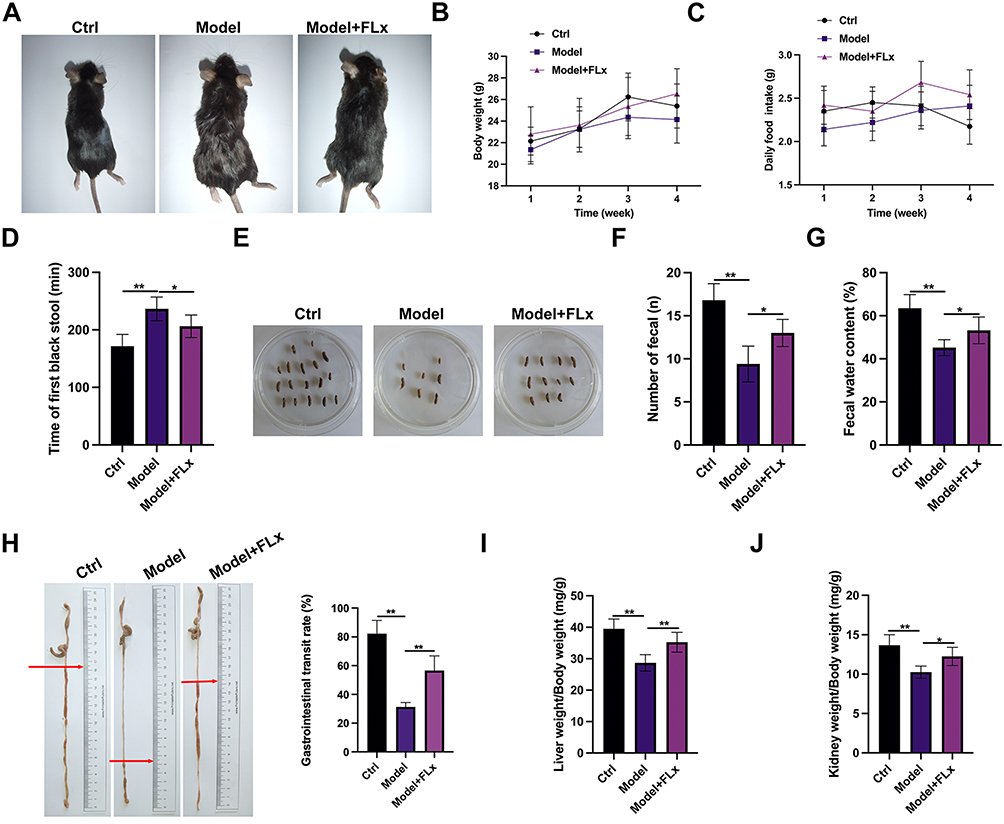 |
Figure 2 Constipation-related parameters in CUMS-induced mice treated with FLX. After CUMS modeling, mice were divided into three groups: control group (Ctrl), Model group (CUMS + FC), and FLX treatment group (Model + FLX). FLX was administered for 4 weeks, and the following parameters were measured: (A) Representative images showing the general physical condition of mice in the control group, model group, and model +FLX group. The images illustrate differences in fur texture, body posture, and overall appearance among the groups. (B) Body weight (BW): the effect on BW (g). (C) Food consumption: daily food intake (g). (D) First black stool defecation time: the time to first black stool defecation (min). (E) Representative fecal pellet output: representative images showing fecal pellets collected from each group during the 6-hour observation period in petri dishes. (F) Quantification of fecal pellet number: total number of fecal pellets excreted within 6 hours post-charcoal meal administration. (G) Fecal water content. (H) Intestinal propulsion distance: the distance the charcoal meal traveled in the intestine (cm). Gastrointestinal transit rate (GTR): the percentage of charcoal meal transit through the gastrointestinal tract. (I) Liver index: liver weight/BW ratio (mg/g). (J) Kidney index: kidney weight/BW ratio (mg/g). Statistical significance for (B) and (C) was determined by two-way repeated measures ANOVA followed by Bonferroni post-hoc tests. Statistical significance for (D–J) was determined by one-way ANOVA followed by Newman-Keuls post-hoc tests (*p < 0.05, **p < 0.01 compared to the model group). |
FLX-Induced Histological and Inflammatory Changes in Colonic Tissues of the CUMS Animal Model
To investigate whether FLX can offer a beneficial effect on the intestinal tract in mice with constipation and CUMS, we analyzed histomorphological changes in the distal colon using histological staining. As shown in Figure 3A, the model group displayed increased inflammatory cell infiltration. A greater muscularis-crypt distance was observed compared to the control group and shorter crypts with fewer goblet cells. After FLX treatment, a decrease in inflammatory cell infiltration and an increase in goblet cells were observed. These findings indicate that constipation could induce colitis in the mouse model. FLX alleviated the inflammatory response caused by constipation. AB-PAS staining was used to evaluate goblet cell abundance and mucin production in the colon of the model mice and those treated with FLX (Figure 3B). AB-PAS staining indicated that the goblet cell abundance was greater in the control than in the model. However, FLX restored mucin production in the goblet cells. FLX also ameliorated the enhanced inflammatory response instigated by CUMS and FC (Figure 3C and D). Levels of IL-1β, TNF-α, and IL-6 were elevated in the colon tissues of the model, whereas IL-10 was expressed at lower levels. These findings demonstrate that CUMS and FC have a detrimental effect on colon function that is alleviated by FLX.
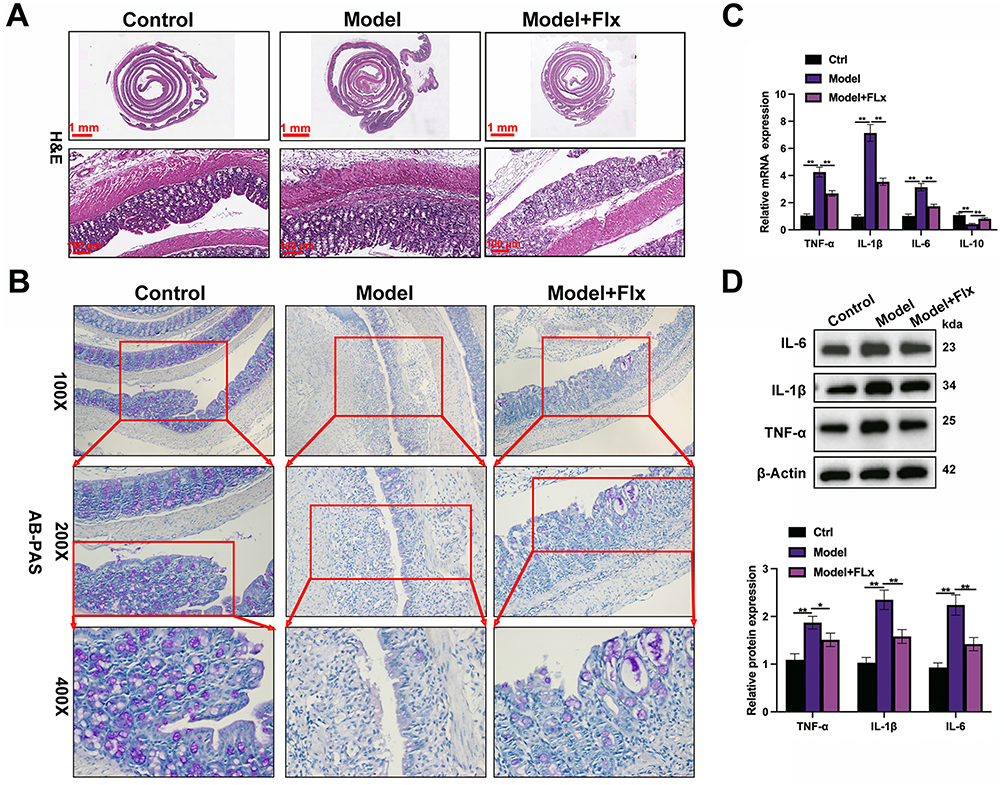 |
Figure 3 Histological and inflammatory changes in colonic tissues of CUMS-induced mice treated with FLX. FLX treatment was applied for 4 weeks after CUMS modeling. The following assays were performed to evaluate the histological and inflammatory changes in the colons of the mice: (A) H&E staining was used to assess the histopathological changes in the colon tissue. (B) AB-PAS staining was performed to evaluate goblet cell abundance and mucin production in the colon. (C) The mRNA expression levels of IL-1β, TNF-α, IL-6, and IL-10 in colon tissues were quantified by qRT-PCR to assess the inflammatory response. (D) Western blot results showing the protein expression levels of TNF-α, IL-1β, and IL-6 in colonic tissues from the Control, Model, and FLX treatment group. Data are presented as mean ± SD (n = 6 per group). Statistical significance was determined by one-way ANOVA followed by post-hoc tests (*p < 0.05, **p < 0.01 compared to the model group). |
FLX Enhances Serotonin Signaling, Gastrointestinal Hormones, Smooth Muscle Contraction, and Intestinal Barrier Function in CUMS Mice
To determine the overall impact of FLX in CUMS we measured the serum content of Gastrin (Gas), Somatostatin (SS), Acetylcholinesterase (AChE), Serotonin (5-HT), Substance P (SP), Motilin (MTL), and Vasoactive intestinal peptide (VIP) in serum samples. The levels of these markers demonstrate that gastrointestinal function is compromised in the model but is alleviated by FLX. However, FLX does not completely restore gastrointestinal function to control levels (Figure 4A). Similar results were obtained from the mRNA expression levels of serotonin transporter (SERT), 5-hydroxytryptamine receptor 4 (5-HT4), mucin-2 (MUC-2), stem cell factor (SCF), and CD117 (C-KIT) in colon tissues (Figure 4B). Levels were reduced in the CUMS model, but expression was restored with FLX. IHC reveals a lower expression of C-KIT in the colon tissues of the model with CUMS and FC (Figure 4C). The immunofluorescence staining of tight junction proteins showed reduced levels of Occludin and ZO-1 in the colon of model mice compared to control, with FLX treatment partially restoring their expression (Figure 4D). Overall, these results demonstrate that FLX can enhance serotonin signaling, gastrointestinal hormones, smooth muscle contraction, and intestinal barrier function in CUMS mice with FC.
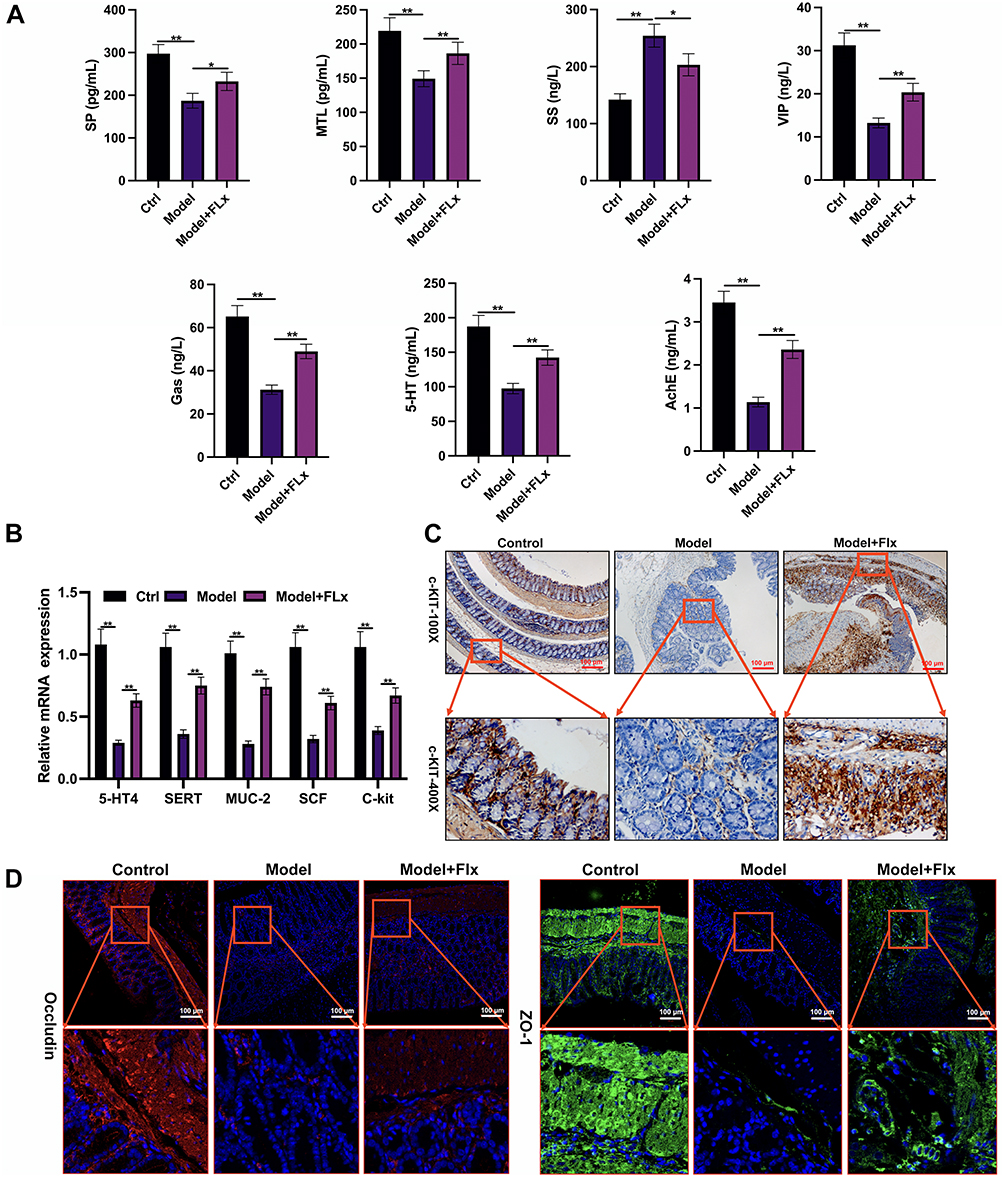 |
Figure 4 FLX enhances serotonin signaling, gastrointestinal hormones, smooth muscle contraction, and intestinal barrier function in CUMS mice. (A) Serum biochemical analysis: ELISA kits were used to quantify the levels of Gas, SS, AChE, 5-HT, SP, MTL, and VIP in serum samples from the Control group (Ctrl), Model group (CUMS+FC), and FLX treatment group (MODEL + FLX). (B) qRT-PCR analysis of gastrointestinal function markers in colon tissues. (C) Immunohistochemical staining of C-KIT in colon tissues from all groups. (D) Immunofluorescence staining of tight junction proteins ZO-1 and Occludin in colon tissues across the three groups. Data are presented as mean ± SD (n = 6 per group). Statistical significance was determined by one-way ANOVA followed by post-hoc tests (*p < 0.05, **p < 0.01 compared to the model group). |
FLX Enhances Neuroprotection and Synaptic Plasticity in the PFC of Mice
To investigate the neurological impact of CUMS and FLX we measured levels of key proteins involved in neuroprotection and synaptic plasticity in the PFC of the experimental models. Our results indicate that levels of brain-derived neurotrophic factor (BDNF), mechanistic target of rapamycin (mTOR), and phosphorylated mTOR are adversely affected by CUMS and FC in the PFC and serum of mice (Figure 5A–C) and that FLX can restore levels. However, FLX does not restore the levels of proteins in the model to those of the control. We also examined levels of the synaptic molecular marker postsynaptic density protein 95 (PSD-95), Synapsin I, and AMPA receptor subunit GluA1 (GluR1) in the PFC. We found that the synaptic molecular markers were significantly depleted in the model but synaptic function was significantly restored by FLX (Figure 5D). In summary, CUMS negatively impacts the expression of key proteins involved in neuroprotection and synaptic plasticity in the PFC, whereas FLX restores their levels and improves synaptic function.
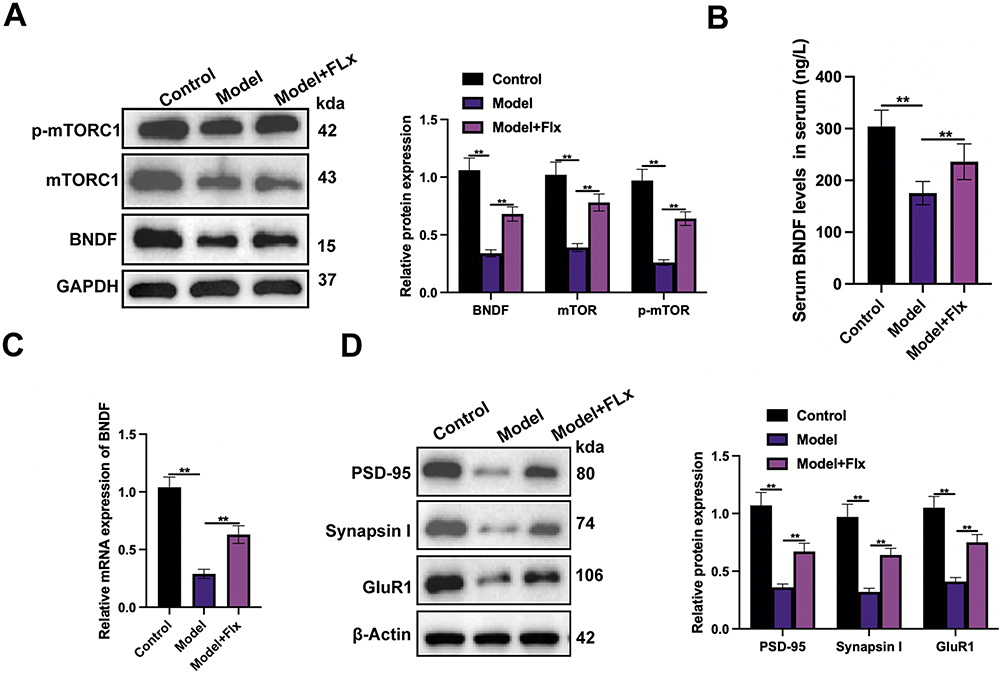 |
Figure 5 FLX treatment enhances neuroprotection and synaptic plasticity in the PFC of mice. (A) Western blot analysis of BDNF, mTOR, and p-mTOR expression levels. (B) ELISA quantification of BDNF levels in mouse serum across the three groups. (C) qRT-PCR analysis of BDNF mRNA expression levels in the PFC of mice. (D) Western blot analysis of PSD-95, Synapsin I, and GluR1 in the PFC of mice. Data are presented as mean ± SD (n = 6 per group). Statistical significance was determined by one-way ANOVA followed by post-hoc tests (**p < 0.01). |
FLX Treatment Restores Gut Microbiota Diversity Disrupted by CUMS-Induced Constipation
Having confirmed that the CUMS + FC model and FLX could impact both the neurological and intestinal health of the mice we measured the diversity of gut microbiota across the three experimental groups. The ASVs between the three groups differed considerably (Figure 6A). The number of unique ASVs was the highest in the FLX-treated model group followed by the model and then the control. The Chao1 index, representing species richness, and the Shannon index, representing species diversity, were both significantly lower in the model (Figure 6B and C). The FLX-treated model group exhibited the most significant species richness compared to the model group and the greatest species richness and evenness across the samples from the three groups (Figure 6D). PCoA based on Bray-Curtis dissimilarity, displayed the gut microbiota composition of the three groups in unique clusters (Figure 6E). These results demonstrate that FLX treatment significantly reversed CUMS+FC-induced gut microbiota dysbiosis, restoring diversity, richness, and evenness, with butyrate supplementation enhancing these restorative effects.
 |
Figure 6 FLX treatment restores gut microbiota diversity disrupted by CUMS-induced constipation. (A) Venn diagram/Petal chart based on the distribution of amplicon sequence variants (ASVs), showing the shared and unique ASVs among the Control group (Ctrl), Model group (chronic unpredictable mild stress + functional constipation), and Fluoxetine treatment group (Model + FLX). (B) The Chao1 index representing species richness in the three groups. (C) Shannon index showing species diversity in the three groups. (D) Rank abundance curve, illustrating species richness and evenness across the samples from the three groups. (E) PCoA based on Bray-Curtis dissimilarity, displaying the clustering of gut microbiota composition among the three groups. Data are presented as mean ± SD (n = 6 per group). Statistical significance was determined by one-way ANOVA followed by post-hoc tests (*p < 0.05, ***p < 0.001). |
FLX Alters the Relative Abundance of Gut Microbiota and Increases Beneficial Bacterial Taxa in the Experimental Model
We examined the taxonomical abundance of the gut bacterial species between the three experimental groups in more detail. At the phylum levels, most species were represented by Bacteroidota and Firmicutes (Figure 7A). The ratio of Bacteroidota and Firmicutes was similar in the control and FLX-treated groups. However, ANOVA of the dominant phyla revealed that the relative abundance of Bacteroidota was lower, and Firmicutes was higher in the CUMS + FC model group compared to the control and FLX-treated model groups (Figure 7B and C). Relative abundance of gut microbiota at the genus level across the three groups, revealed that Muribaculaceae and Lactobacillus were the dominant genera (Figure 7D). There was no significant difference in the abundance of Muribaculaceae between the control and FLX-treated model group (Figure 7E). However, there were significantly lower levels of Muribaculaceae in the untreated model. Additionally, although the gut abundance of Lactobacillus seemed lower in the untreated group than in the control and FLX-treated mice, there was no significant difference (Figure 7F). The phylogenic relationship of the taxa is shown in Figure 7G. The gut of the treated model mice seemed to contain a greater number of Eubacterium oxidoreducens (Figure 7H). To summarize, FLX improved overall species richness, diversity, and evenness, and increased the presence of butyrate-producing E. oxidoreducens.
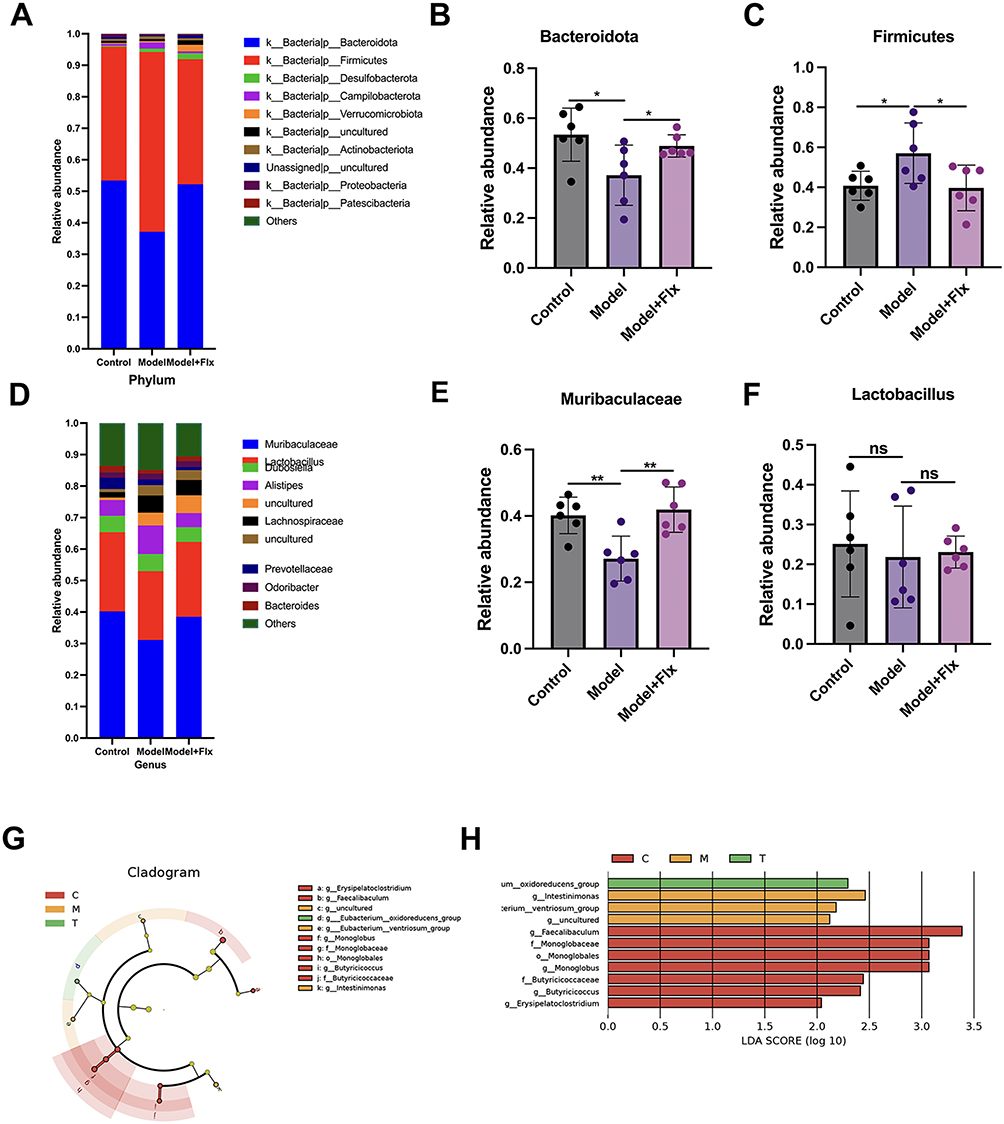 |
Figure 7 Fluoxetine (FLX) treatment alters the relative abundance of gut microbiota and highlights key bacterial taxa in the different groups. (A) Relative abundance of gut microbiota at the phylum level across the Control group (Ctrl), Model group (chronic unpredictable mild stress + functional constipation), and Fluoxetine treatment group (Model + FLX). (B) Relative abundance of Bacteroidota at the phylum level across experimental groups. (C) Relative abundance of Firmicutes at the phylum level across experimental groups. (D) Relative abundance of gut microbiota at the genus level across the three groups. (E) Relative abundance of Muribaculaceae at the genus level across experimental groups. (F) Relative abundance of Lactobacillus at the genus level across experimental groups. (G) Linear discriminant analysis effect size (LEfSE) evolutionary branching diagram, showing the phylogenetic relationship of differentially abundant taxa among the groups. (H) Linear discriminant analysis (LDA) scores, indicating the bacterial taxa that contribute most to the differences between the groups. Data are presented as mean ± SD (n = 6 per group). Statistical significance for (B, C, E, F) was determined by one-way ANOVA with Benjamini-Hochberg FDR correction for multiple testing (FDR < 0.1). For LEfSe analysis (G and H) (*p < 0.05, **p < 0.01, ns, no significance). |
FLX-Induced Fecal SCFA Levels and Their Correlation with Gut Microbiota
We conducted a further analysis of fecal SCFAs using gas chromatography-mass spectrometry (GC/MS) to determine whether they were influenced by the experimental conditions (Figure 8A–E). Levels of acetic acid, propionic acid, and butyric acid were significantly higher in the control group and in the FLX-treated group than in the model group, with the highest levels found consistently in the control group. Levels of isobutyric acid and isovaleric acid were not significantly different across the three groups, although the control group had the lowest levels of isovaleric acid. There was a significant correlation between the relative abundance of Muribaculaceae and butyric acid levels (Pearson p = 0.048) (Figure 8F). Muribaculaceae produce SCFA, including butyric acid, through the fermentation of dietary fibers and mucin glycans.23 The levels of butyric acid across the three groups reflected the levels of Muribaculaceae found in the gut (Figure 8D).
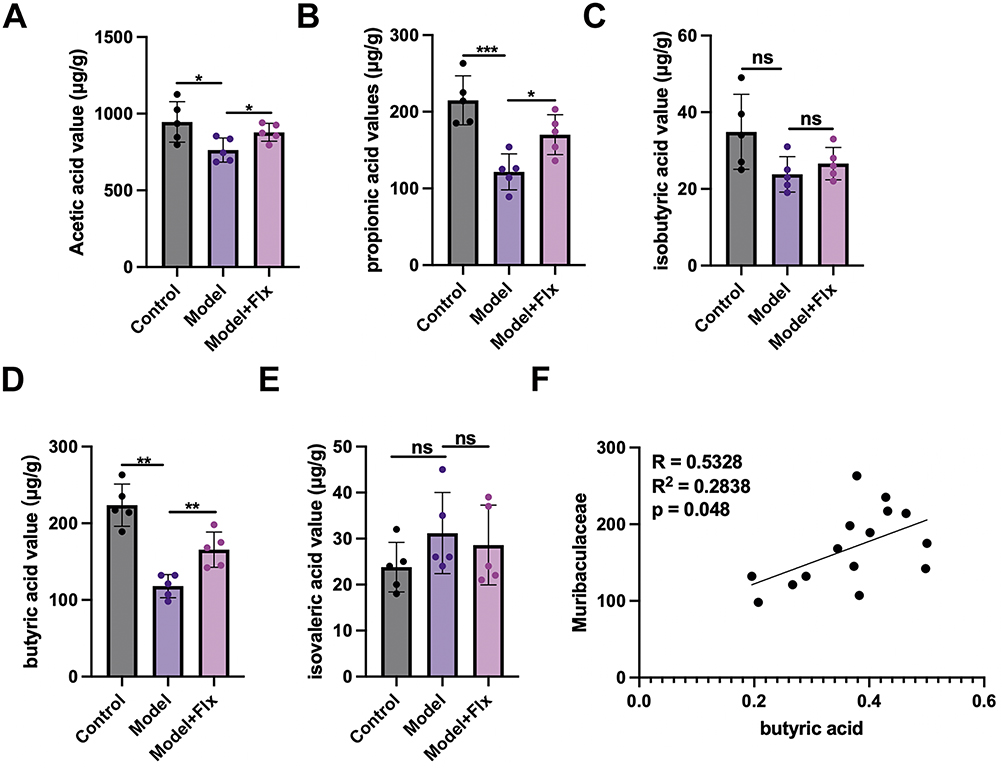 |
Figure 8 FLX-induced fecal SCFA levels and their correlation with gut microbiota. The levels of major SCFAs, including acetic acid (A), propionic acid (B), isobutyric acid (C), butyric acid (D), and isovaleric acid (E), in stool samples from the Control group (Ctrl), Model group (chronic unpredictable mild stress + functional constipation), and FLX treatment group (Model + FLX). The SCFA concentrations were measured using gas chromatography-mass spectrometry (GC/MS). (F) Pearson correlation analysis between the relative abundance of Muribaculaceae and butyric acid levels across the three groups. Data are presented as mean ± SD (n = 6 per group). Statistical significance for (A–E) was determined by one-way ANOVA followed by Newman-Keuls post-hoc tests. For (F), Pearson correlation coefficient (r) and p-value were calculated to assess the linear relationship between variables (*p < 0.05, **p < 0.01, ***p<0.001 ns, no significance). |
Butyrate Combined with Fluoxetine Improves Gut Motility, Reduces Inflammation, and Enhances Intestinal Barrier Integrity in Mice
To finalize our analysis, we repeated our experiments with the inclusion of butyrate supplementation. The improvements in fecal pellet number and GTR observed in the CUMS model with FLX treatment were further enhanced by butyrate supplementation (Figure 9A–C). Inflammation was reduced and the number of mucin-producing goblet cell density was increased in the colon by supplementation with butyrate (Figure 9D–G). Additionally, intestinal barrier integrity evaluated by tight junction proteins was further restored and gastrointestinal function was improved in the group that received FLX and butyrate (Figure 9F–I). To conclude, butyrate supplementation combined with FLX alleviates CUMS-induced depression and constipation by targeting gut–brain axis dysfunction, modulating gut microbiota, and improving intestinal barrier function.
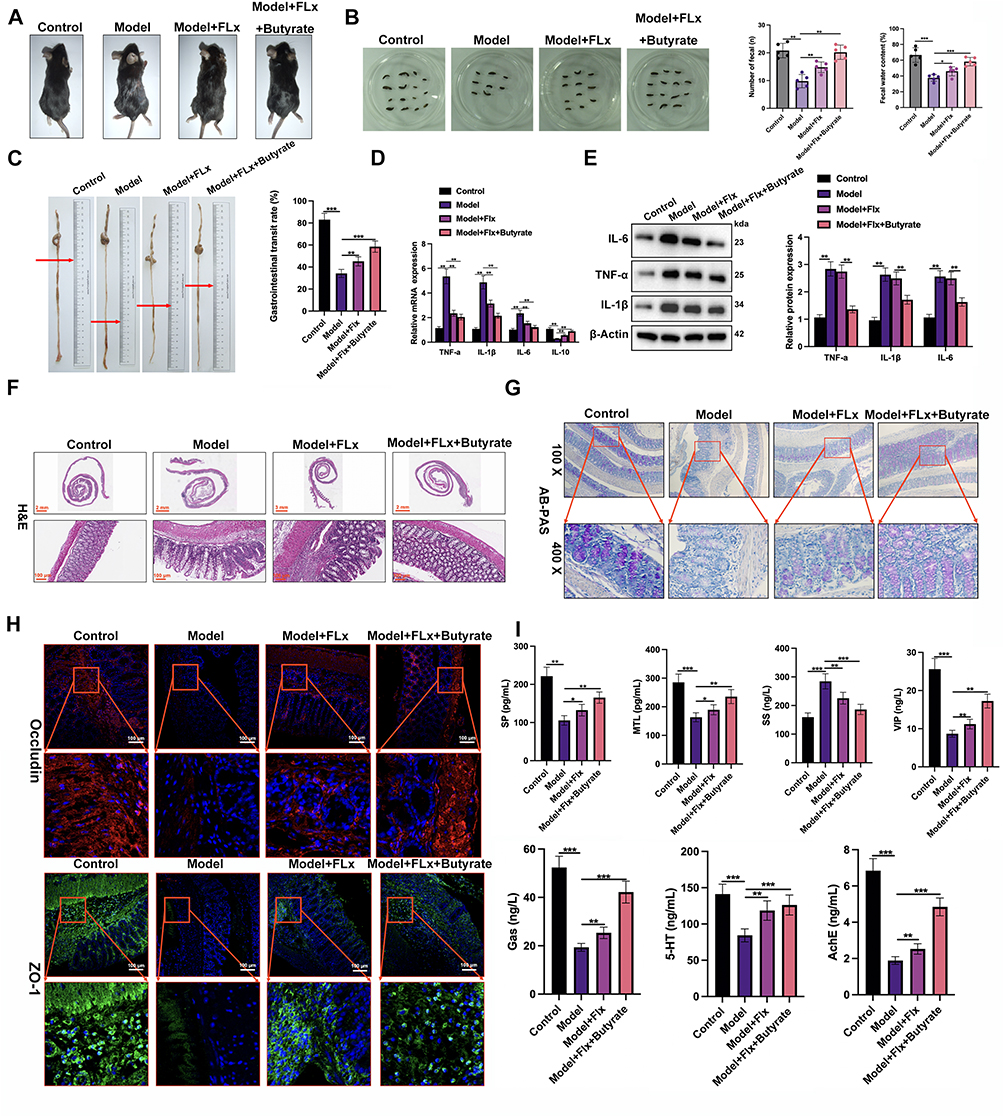 |
Figure 9 Butyrate combined with fluoxetine improves gut motility, reduces inflammation, and enhances intestinal barrier integrity in mice. (A) Representative images showing the general physical condition of mice in the Control group (Ctrl), Model group (CUMS + FC), FLX treatment group (Model + FLX), and FLX + Butyrate treatment group (Model + FLX + Butyrate). (B) Fecal output analysis: The number of fecal pellets and the wet weight of feces excreted over 6 hours were measured. (C) Gastrointestinal motility: the distance traveled by activated charcoal in the intestines (in cm) and the GTR, expressed as the percentage of the total gastrointestinal tract distance traversed by the charcoal. (D) Quantification of inflammatory cytokines in colon tissues, including IL-1β, TNF-α, IL-6, and IL-10, using qRT-PCR. (E) Western blot analysis of pro-inflammatory cytokines in colonic tissues. (F) H&E staining of colon tissue sections from all groups to assess histopathological changes. (G) AB-PAS staining to evaluate mucin production and goblet cell density in the colon across the four groups. (H) Immunofluorescence staining to detect the expression of tight junction proteins Occludin and ZO-1 in the colon tissues. (I) Serum biochemical analysis: Using ELISA kits to quantify serum levels of Gas, SS, AChE, 5-HT, SP, MTL, and VIP. Data are presented as mean ± SD (n = 6 per group). Statistical significance was determined by one-way ANOVA followed by post-hoc tests (*p < 0.05, **p < 0.01, ***p < 0.001). |
Discussion
A simple and effective way to alleviate CUMS would have far-reaching social and economic benefits, improving the well-being of individuals and communities.24 The link between depression and constipation is well-documented.25 The depression and gastrointestinal dysfunction associated with CUMS are believed to be strongly related to gut–brain axis dysregulation.26 Therefore, it is important to understand and address this axis in therapeutic interventions.
Our study demonstrated that FLX can enhance defecation, behavioral indicators, and OFT parameters in a CUMS model with constipation in mice. Similar studies have shown that antidepressants like FLX can reverse depression-induced gastrointestinal dysfunction, supporting our findings. For instance, Sharma et al highlighted the effectiveness of SSRIs in reversing CUMS-induced behavioral despair and cognitive impairments.24 CUMS induced notable histological and inflammatory changes in colonic tissues in our animal model. This aligns with findings from Alqurashi et al, who reported that CUMS-induced depression negatively impacts histological markers and inflammatory responses in the hippocampus and PFC.27 Our results showed that treatment with FLX restored mucin production and the reduction of inflammatory markers in mice colon tissue. FLX was also shown to enhance serotonin signaling, gastrointestinal hormones, smooth muscle contraction, and intestinal barrier function in our mouse model of CUMS and FC model. This is in line with previous studies that have demonstrated the role of serotonin in gastrointestinal function and its modulation by antidepressants.28,29 The restoration of SERT and 5-HT4 GPCR expression by FLX in our study is supported by these findings. Although FLX did not completely restore gastrointestinal function to control levels, the improvements were notable and consistent with existing literature.
We also demonstrate that FLX enhances neuroprotection and synaptic plasticity in the PFC of mice. Studies have shown that CUMS negatively impacts synaptic markers and neurotrophic factors, which can be restored by antidepressant treatment. For instance, Han et al found that BDNF and synaptic plasticity markers were decreased in the hippocampus and the PFC in a CUMS rat model.30 Similarly, we found that levels of BDNF, mTOR, and synaptic plasticity markers are adversely affected by CUMS and FC in the PFC and serum of mice. FLX has been shown to increase BDNF expression in the PFC, which can help restore synaptic plasticity and cognitive function.31 FLX also influences mTOR signaling, promoting protein synthesis and synaptic strength. In agreement with other studies, the CUMS-induced decrease in PSD-95 and Synapsin I levels were reversed by FLX.31
Other studies have found that CUMS can generate depression-like phenotypes in mice by differentially altering gut microbiota and triggering macroglia activation.32 We found that FLX treatment altered gut microbiota diversity in mice, increasing the relative abundance of beneficial bacterial taxa. This is supported by studies that have shown the impact of antidepressants on gut microbiota composition.33,34 However, our study uniquely demonstrates FLX’s therapeutic effects in a comorbid CUMS+FC model, revealing that FLX improves both depression-like behaviors and constipation symptoms through microbiota modulation—a finding not previously reported. Importantly, we provide mechanistic insights by linking FLX-induced microbiota changes to specific molecular pathways, including restoration of intestinal barrier proteins (ZO-1, Occludin) and activation of BDNF/mTOR signaling in gut and brain tissues. The increase in Eubacterium oxidoreducens in our study is consistent with previous studies that describe the beneficial roles of this genus in gut health.35 We also studied the effects of gut microbiota on fecal SCFA levels and found a significant correlation between the relative abundance of Muribaculaceae and butyric acid levels. This is consistent with research indicating that gut microbiota can influence SCFA production and gastrointestinal health.36 Our work extends beyond previous findings by demonstrating that combining FLX with butyrate supplementation produces enhanced therapeutic outcomes, particularly in reversing constipation-related pathology and neuroinflammation, suggesting a novel therapeutic strategy for treating depression with gastrointestinal comorbidities. The role of Muribaculaceae and other microbiota in producing SCFAs like butyric acid has been well-documented in the literature.15,23 For instance, He et al found that Astragaloside IV could modulate the gut microbiota profile to relieve slow transit constipation in mice by promoting the generation of butyric acid.15
Finally, we investigated the combined effects of butyrate supplementation and FLX. Butyrate supplementation enhanced the FLX-induced improvements in gastrointestinal function and reduction in inflammation. Other studies have demonstrated the synergistic effects of butyrate and antidepressant agents in modulating gut–brain axis dysfunction and improving intestinal barrier function.16,37–39 Palepu et al discovered that supplementing a treatment-resistant model of depression in rats with butyrate could significantly improve both antidepressant and antianxiety effects by modulating the gut–brain axis.38 While our data suggest synergistic effects between FLX and butyrate, the precise mechanistic interplay requires further investigation. The enhanced therapeutic outcomes may involve multiple pathways, including histone deacetylase (HDAC) inhibition by butyrate potentiating FLX-mediated serotonergic signaling, cross-talk between GPR41/43 receptors and serotonin receptors, or convergent effects on immune-inflammatory pathways through NF-κB suppression. Future studies employing dose-response analyses and isobolographic assessments would help formally distinguish between synergistic versus additive interactions, while investigation of receptor-level mechanisms and immune mediators could elucidate the molecular basis of this therapeutic cooperation.
Regarding translational implications, the butyrate dose used in our study (200 mg/kg/day) corresponds to approximately 16 mg/kg/day in humans using allometric scaling based on body surface area conversion factors. For a 70 kg adult, this would translate to approximately 1120 mg/day, which is within the range used in clinical studies for gastrointestinal disorders (typically 600–2000 mg/day).40,41 However, the bioavailability of oral butyrate in humans is limited due to rapid absorption and metabolism in the upper gastrointestinal tract. Clinical formulations often employ enteric-coated or slow-release preparations to enhance colonic delivery, which may be necessary for optimal therapeutic effects. Furthermore, while our study demonstrates proof-of-concept for FLX-butyrate combination therapy, careful dose-optimization studies in humans would be essential, as factors such as differences in gut microbiota composition, intestinal transit time, and hepatic metabolism between mice and humans may influence the optimal therapeutic dosing regimen.
Our study has some limitations. The CUMS model involves exposure to a variety of stressors, which can introduce variability in stress responses among individual mice, potentially affecting the consistency and reproducibility of the results. Additionally, we used only female mice without monitoring their estrous cycles, which may have introduced variability in behavioral, neurobiological, and microbiota-related outcomes. While efforts were made to minimize observer bias through blinding procedures, behavioral indicators and OFT parameters remain subject to potential subjective interpretation. Sample size variations between experiments may have affected statistical power, and future studies should include larger group sizes to enhance robustness of findings. Furthermore, the analysis of gut microbiota is based on a single time point, potentially overlooking temporal changes and dynamics in microbiota composition and function through the course of the study. Fecal microbiota transplantation (FMT) experiments using feces from treated and untreated mice could have strengthened causal inferences about the role of microbiota in mediating the observed therapeutic effects, which should be considered in future studies. The long-term effects of butyrate supplementation in combination with FLX were also not assessed. While our study demonstrates acute therapeutic benefits, it does not address questions about treatment durability, potential tolerance development, or long-term safety of the combination therapy. Future studies should extend the treatment period to 12–16 weeks with post-treatment follow-up to evaluate whether the beneficial effects persist after discontinuation and whether prolonged butyrate supplementation maintains its synergistic effects with FLX without adverse consequences. Additionally, longitudinal assessment of gut microbiota changes and metabolic parameters would help determine whether the observed microbial shifts stabilize or continue to evolve with extended treatment.
Conclusions
This study underscores the pivotal role of the gut–brain axis in CUMS-induced depression-like behaviors and gastrointestinal dysfunctions, such as constipation. Through our experimental findings, we have demonstrated that FLX has multifaceted therapeutic effects in alleviating these symptoms. FLX improved behavioral and gastrointestinal parameters in a CUMS + FC model but also induced significant histological and inflammatory changes, restoring mucin production and reducing pro-inflammatory cytokine levels. FLX also enhanced serotonin signaling, gastrointestinal hormones, intestinal barrier function, neuroprotection, and synaptic plasticity. The alteration of gut microbiota diversity by FLX, with an increase in beneficial bacterial taxa, further emphasizes the interconnectedness of the gut–brain axis. The correlation between Muribaculaceae and butyric acid levels underscores the role of gut microbiota in producing SCFAs that support gastrointestinal health. Lastly, the combined effects of butyrate supplementation and FLX demonstrated enhanced improvements in gastrointestinal function, inflammation reduction, and intestinal barrier integrity, suggesting a synergistic therapeutic potential. In conclusion, our study contributes to the growing body of evidence supporting the role of the gut–brain axis in CUMS-induced depression and gastrointestinal dysfunction. FLX with butyrate supplementation, emerges as a promising intervention to manage these interconnected health issues.
This study demonstrates that FLX effectively alleviates both depression-like behaviors and constipation symptoms in a CUMS-induced mouse model through modulation of the gut-brain axis. Our results showed that FLX treatment improved behavioral and gastrointestinal parameters, restored colonic histological architecture, reduced inflammation, and enhanced intestinal barrier function. Additionally, FLX promoted neuroprotection and synaptic plasticity in the PFC while restructuring the gut microbiota composition, particularly increasing butyrate-producing bacteria with a significant correlation between Muribaculaceae abundance and fecal butyrate levels. The combination of FLX with butyrate supplementation produced superior therapeutic outcomes compared to FLX monotherapy, with enhanced improvements in gastrointestinal function and inflammation. This dual-targeting approach addresses both central nervous system dysfunction and gut dysbiosis simultaneously, representing a novel strategy for treating CUMS-induced comorbid depression and constipation.
These findings have translational relevance given that depression with gastrointestinal comorbidities affects approximately 30% of major depressive disorder patients, often showing limited response to conventional antidepressants. Both FLX and sodium butyrate are clinically available with established safety profiles, and our dosing regimen translates to clinically feasible levels for human application. This preclinical study suggests that FLX-butyrate combination therapy may offer a more comprehensive treatment approach than current monotherapies for managing gut-brain axis dysfunction in stress-related disorders.
Abbreviations
5-HT, Serotonin; 5-HT4 GPCR, 5-hydroxytryptamine receptor 4; AB-PAS, Alcian Blue-Periodic Acid-Schiff; AChE, Acetylcholinesterase; ASV, Amplicon sequence variant; BDNF, Brain-derived neurotrophic factor; BW, Body weight; C-KIT, CD117, tyrosine-protein kinase Kit; CUMS, Chronic unpredictable mild stress; ELISA, Enzyme-linked immunosorbent assay; FC, Functional constipation; FBS, First black stool; FLX, Fluoxetine; FST, Forced swim test; FWC, Fecal water content; Gas, Gastrin; GC/MS, Gas chromatography-mass spectrometry; GTR, Gastrointestinal transit rate; H&E, Hematoxylin and eosin; ICC, Interstitial cells of Cajal; IF, Immunofluorescence; IHC, Immunohistochemistry; LDA, Linear discriminant analysis; MTL, Motilin; MUC-2, Mucin-2; mTORC1, Mechanistic target of rapamycin complex 1; OFT, Open field test; OUT, Operational taxonomic unit; PCoA, Principal coordinates analysis; PFC, Prefrontal cortex; PSD-95, Postsynaptic density protein 95; RT-qPCR, Reverse transcription quantitative polymerase chain reaction; SCF, Stem cell factor; SCFA, Short-chain fatty acid; SDS, Sodium dodecyl sulfate; SERT, Serotonin transporter; SP, Substance P; SPF, Specific pathogen-free; SS, Somatostatin; SSRI, Selective serotonin reuptake inhibitor; TBST, Tris-buffered saline with Tween; TNF-α, Tumor necrosis factor alpha; VIP, Vasoactive intestinal peptide; ZO-1, Zonula occludens-1.
Data Sharing Statement
All data generated or analyzed during this study are included in the manuscript. Additional reasonable requests for data can be directed to the corresponding author, Xiaoran Yang.
Acknowledgments
This research was supported by the Ningxia Nature Foundation (2023AAC03471), Ningxia Hui Autonomous Region Science and Technology Benefit People Project (2023CMG03017) and National Natural Science Foundation Pre-experimental Project in 2026 (2026GZRYSY001).
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
There are no conflicts to declare.
References
1. Tianzhu Z, Shihai Y, Juan D. Antidepressant-like effects of cordycepin in a mice model of chronic unpredictable mild stress. Evid Based Complement Alternat Med. 2014;2014(1):438506. doi:10.1155/2014/438506
2. Jia Z, Yang J, Cao Z, et al. Baicalin ameliorates chronic unpredictable mild stress-induced depression through the BDNF/ERK/CREB signaling pathway. Behav Brain Res. 2021;414:113463. doi:10.1016/j.bbr.2021.113463
3. Sun Y, Zhao H, Chang M, Yue T, Yuan Y, Shi Y. Prophylactic effects of Tibetan goat kefir on depression-like behaviors in chronic unpredictable stress model through the gut-brain axis. J Sci Food Agric. 2025;105(1):266–275. doi:10.1002/jsfa.13825
4. Li N, Wang Q, Wang Y, et al. Fecal microbiota transplantation from chronic unpredictable mild stress mice donors affects anxiety-like and depression-like behavior in recipient mice via the gut microbiota-inflammation-brain axis. Stress. 2019;22(5):592–602. doi:10.1080/10253890.2019.1617267
5. Szoke-Kovacs Z, More C, Szoke-Kovacs R, Mathe E, Frecska E. Selective inhibition of the serotonin transporter in the treatment of depression: sertraline, fluoxetine and citalopram. Neuropsychopharmacol Hung. 2020;22(1):4–15.
6. Irum N, Afzal T, Faraz MH, Aslam Z, Rasheed F. The role of gut microbiota in depression: an analysis of the gut-brain axis. Front Behav Neurosci. 2023;17:1185522. doi:10.3389/fnbeh.2023.1185522
7. Zhang W, Qu W, Wang H, Yan H. Antidepressants fluoxetine and amitriptyline induce alterations in intestinal microbiota and gut microbiome function in rats exposed to chronic unpredictable mild stress. Transl Psychiatry. 2021;11(1):131. doi:10.1038/s41398-021-01254-5
8. Yuan Q, Lei Y, Yu K, et al. Repetitive transcranial magnetic stimulation and fluoxetine attenuate astroglial activation and benefit behaviours in a chronic unpredictable mild stress mouse model of depression. World J Biol Psychiatry. 2024;25(2):82–94. doi:10.1080/15622975.2023.2279958
9. Sun Y, Geng W, Pan Y, Wang J, Xiao P, Wang Y. Supplementation with Lactobacillus kefiranofaciens ZW3 from Tibetan Kefir improves depression-like behavior in stressed mice by modulating the gut microbiota. Food Funct. 2019;10(2):925–937. doi:10.1039/c8fo02096e
10. Potter K, Gayle EJ, Deb S. Effect of gut microbiome on serotonin metabolism: a personalized treatment approach. Naunyn Schmiedebergs Arch Pharmacol. 2024;397(5):2589–2602. doi:10.1007/s00210-023-02762-5
11. Ding G, Yang X, Li Y, et al. Gut microbiota regulates gut homeostasis, mucosal immunity and influences immune-related diseases. Mol Cell Biochem. 2024. doi:10.1007/s11010-024-05077-y
12. Di Vincenzo F, Del Gaudio A, Petito V, Lopetuso LR, Scaldaferri F. Gut microbiota, intestinal permeability, and systemic inflammation: a narrative review. Intern Emerg Med. 2024;19(2):275–293. doi:10.1007/s11739-023-03374-w
13. LaBouyer M, Holtrop G, Horgan G, et al. Higher total faecal short-chain fatty acid concentrations correlate with increasing proportions of butyrate and decreasing proportions of branched-chain fatty acids across multiple human studies. Gut Microbiome. 2022;3:e2. doi:10.1017/gmb.2022.1
14. Tian P, Zou R, Song L, et al. Ingestion of Bifidobacterium longum subspecies infantis strain CCFM687 regulated emotional behavior and the central BDNF pathway in chronic stress-induced depressive mice through reshaping the gut microbiota. Food Funct. 2019;10(11):7588–7598. doi:10.1039/c9fo01630a
15. He Q, Han C, Huang L, et al. Astragaloside IV alleviates mouse slow transit constipation by modulating gut microbiota profile and promoting butyric acid generation. J Cell Mol Med. 2020;24(16):9349–9361. doi:10.1111/jcmm.15586
16. Cheng J, Hu H, Ju Y, et al. Gut microbiota-derived short-chain fatty acids and depression: deep insight into biological mechanisms and potential applications. Gen Psychiatr. 2024;37(1):e101374. doi:10.1136/gpsych-2023-101374
17. Tang T, Wang J, Jiang Y, et al. Bifidobacterium lactis TY-S01 prevents loperamide-induced constipation by modulating gut microbiota and its metabolites in mice. Front Nutr. 2022;9:890314. doi:10.3389/fnut.2022.890314
18. Wang P, Yu C, Li Y, Zhang X, Yao X, Zhang Y. Experimental study on the effects of ghrelin on gastric smooth muscle and posterior limb skeletal muscle in mice. Front Med Lausanne. 2025;12:1631707. doi:10.3389/fmed.2025.1631707
19. Zhan Y, Wen Y, Du LJ, et al. Effects of maren pills on the intestinal microflora and short-chain fatty acid profile in drug-induced slow transit constipation model rats. Front Pharmacol. 2022;13:804723. doi:10.3389/fphar.2022.804723
20. Barnes CA. Memory deficits associated with senescence: a neurophysiological and behavioral study in the rat. J Comp Physiol Psychol. 1979;93(1):74–104. doi:10.1037/h0077579
21. Gao X, Liang M, Fang Y, et al. Deciphering the differential effective and toxic responses of bupleuri radix following the induction of chronic unpredictable mild stress and in healthy rats based on serum metabolic profiles. Front Pharmacol. 2017;8:995. doi:10.3389/fphar.2017.00995
22. Huang X, Li W, You B, et al. Serum metabonomic study on the antidepressant-like effects of ellagic acid in a chronic unpredictable mild stress-induced mouse model. J Agric Food Chem. 2020;68(35):9546–9556. doi:10.1021/acs.jafc.0c02895
23. Zhu Y, Chen B, Zhang X, et al. Exploration of the Muribaculaceae family in the gut microbiota: diversity, metabolism, and function. Nutrients. 2024;16(16):2660. doi:10.3390/nu16162660
24. Sharma S, Chawla S, Kumar P, Ahmad R, Kumar Verma P. The chronic unpredictable mild stress (CUMS) Paradigm: bridging the gap in depression research from bench to bedside. Brain Res. 2024;1843:149123. doi:10.1016/j.brainres.2024.149123
25. Adibi P, Abdoli M, Daghaghzadeh H, et al. Relationship between depression and constipation: results from a large cross-sectional study in adults. Korean J Gastroenterol. 2022;80(2):77–84. doi:10.4166/kjg.2022.038
26. Sun J, Wang F, Hu X, et al. Clostridium butyricum attenuates chronic unpredictable mild stress-induced depressive-like behavior in mice via the gut-brain axis. J Agric Food Chem. 2018;66(31):8415–8421. doi:10.1021/acs.jafc.8b02462
27. Alqurashi GK, Hindi EA, Zayed MA, et al. The impact of chronic unpredictable mild stress-induced depression on spatial, recognition and reference memory tasks in mice: behavioral and histological study. Behav Sci. 2022;12(6). doi:10.3390/bs12060166
28. Hung LY, Alves ND, Del Colle A, et al. Intestinal epithelial serotonin as a novel target for treating disorders of gut-brain interaction and mood. Gastroenterology. 2024. doi:10.1053/j.gastro.2024.11.012
29. Koopman N, Katsavelis D, Hove AST, Brul S, Jonge WJ, Seppen J. The multifaceted role of serotonin in intestinal homeostasis. Int J Mol Sci. 2021;22(17):9487. doi:10.3390/ijms22179487
30. Han YX, Tao C, Gao XR, et al. BDNF-related imbalance of Copine 6 and synaptic plasticity markers couples with depression-like behavior and immune activation in CUMS rats. Front Neurosci. 2018;12:731. doi:10.3389/fnins.2018.00731
31. Liu XL, Luo L, Mu RH, et al. Fluoxetine regulates mTOR signalling in a region-dependent manner in depression-like mice. Sci Rep. 2015;5(1):16024. doi:10.1038/srep16024
32. Wu J, Li J, Gaurav C, et al. CUMS and dexamethasone induce depression-like phenotypes in mice by differentially altering gut microbiota and triggering macroglia activation. Gen Psychiatr. 2021;34(6):e100529. doi:10.1136/gpsych-2021-100529
33. Gamboa J, Le GH, Wong S, et al. Impact of antidepressants on the composition of the gut microbiome: a systematic review and meta-analysis of in vivo studies. J Affect Disord. 2025;369:819–833. doi:10.1016/j.jad.2024.10.042
34. Xu F, Xie Q, Kuang W, Dong Z. Interactions between antidepressants and intestinal microbiota. Neurotherapeutics. 2023;20(2):359–371. doi:10.1007/s13311-023-01362-8
35. Zhang X, Chen J, He F, Du W, Yu X. Assessing the causal effects of Eubacterium and Rumphococcus on constipation: a Mendelian randomized study. Front Microbiol. 2024;15:1376232. doi:10.3389/fmicb.2024.1376232
36. Radjabzadeh D, Bosch JA, Uitterlinden AG, et al. Gut microbiome-wide association study of depressive symptoms. Nat Commun. 2022;13(1):7128. doi:10.1038/s41467-022-34502-3
37. Xia H, Guo J, Shen J, Jiang S, Han S, Li L. Butyrate ameliorated the intestinal barrier dysfunction and attenuated acute pancreatitis in mice fed with ketogenic diet. Life Sci. 2023;334:122188. doi:10.1016/j.lfs.2023.122188
38. Palepu MSK, Gajula SNR, K M, Sonti R, Dandekar MP. SCFAs supplementation rescues anxiety- and depression-like phenotypes generated by fecal engraftment of treatment-resistant depression rats. ACS Chem Neurosci. 2024;15(5):1010–1025. doi:10.1021/acschemneuro.3c00727
39. van de Wouw M, Boehme M, Lyte JM, et al. Short-chain fatty acids: microbial metabolites that alleviate stress-induced brain-gut axis alterations. J Physiol. 2018;596(20):4923–4944. doi:10.1113/JP276431
40. Administration FD. Guidance for industry: estimating the maximum safe starting dose in initial clinical trials for therapeutics in adult healthy volunteers. 2005.
41. Di Sabatino A, Morera R, Ciccocioppo R, et al. Oral butyrate for mildly to moderately active Crohn’s disease. Aliment Pharmacol Ther. 2005;22(9):789–794. doi:10.1111/j.1365-2036.2005.02639.x
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.