Back to Journals » International Journal of Women's Health » Volume 17
Relationship Between the Levels of 91 Circulating Inflammatory Proteins and Ovarian-Related Diseases: A Bidirectional Mendelian Randomization Study
Received 10 September 2025
Accepted for publication 3 December 2025
Published 9 December 2025 Volume 2025:17 Pages 5315—5327
DOI https://doi.org/10.2147/IJWH.S566694
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Everett Magann
Jiahui Qu, Liying Zhang
Department of Obstetrics and Gynecology, the Second Hospital of Harbin Medical University, Harbin, Heilongjiang, People’s Republic of China
Correspondence: Liying Zhang, Department of Obstetrics and Gynecology, the Second Hospital of Harbin Medical University, Harbin, Heilongjiang, People’s Republic of China, Email [email protected]
Background: Inflammatory proteins play a pivotal role in the pathogenesis of ovarian diseases, although the underlying mechanisms remain poorly understood. The aim of this study is to investigate the association between circulating markers of inflammation and ovarian diseases.
Methods: A two-sample bidirectional Mendelian randomization (MR) approach was employed, utilizing publicly available genetic databases to examine the relationship between 91 circulating inflammatory markers and six ovarian diseases. The primary analysis utilized the inverse variance weighting (IVW) method, with additional sensitivity analyses, including heterogeneity and pleiotropy tests, to assess the robustness of the findings.
Results: Specific inflammatory markers were found to be associated with ovarian diseases and to exhibit causal relationships. Ovarian cysts were linked to CCL4 and other proteins, showing a positive causal relationship with NT-3. Polycystic ovary syndrome (PCOS) was associated with IL-6 and other markers, with a positive causal relationship to IL-17C. Ovarian dysfunction was associated with IL-6 and other markers, while primary ovarian failure was linked to IL-33. Benign ovarian neoplasms were associated with CCL28 and other proteins, showing a positive causal relationship with FGF-19 and negative causal relationships with FGF-5 and NT-3. Malignant ovarian neoplasms were associated with CCL20 and other markers.
Discussion: This study clarifies the causal relationships between circulating inflammatory markers and ovarian diseases, providing a crucial foundation for future translational research and clinical applications.
Keywords: Mendelian randomization, circulating inflammatory protein, ovarian-related diseases, polycystic ovary syndrome, ovarian neoplasm
Introduction
The ovary is a crucial component of the female reproductive system, responsible not only for producing and releasing oocytes but also for regulating the menstrual cycle and maintaining fertility through hormone secretion. Ovarian-related diseases present a significant threat to women’s health and reproductive capacity worldwide; however, their underlying mechanisms remain incompletely understood. Ovarian cancer (OC) is the seventh most common malignancy among women globally and the eighth leading cause of female mortality.1 Additionally, PCOS, a prevalent endocrine and metabolic disorder, affects approximately 6% to 20% of women of reproductive age, with clinical symptoms typically manifesting during early adolescence.2,3 Ovarian cysts, a common benign gynecological condition, have a prevalence of 34.9% in premenopausal women and 17.0% in postmenopausal women. Moreover, premature ovarian insufficiency affects approximately 5% to 10% of women with secondary amenorrhea.4,5
Inflammation is the host’s physiological response to infection or injury; however, abnormal inflammatory responses can result in tissue damage and are central to the pathogenesis of various diseases, including sepsis, autoimmune disorders, and atherosclerotic thrombosis.6 Increasing evidence suggests a close relationship between inflammation and ovarian-related diseases. Tumor necrosis factor (TNF)-α, interleukin (IL)-1β, and IL-6 have been shown to promote the onset, growth, and progression of epithelial ovarian cancer (EOC).7,8 TNF plays a key role in the apoptosis of cells induced by both natural and synthetic environmental estrogens.9 PCOS is associated with elevated levels of IL-18, monocyte chemoattractant protein-1 (MCP-1), and macrophage inflammatory protein-1α (MIP-1α).10,11 However, these studies have primarily focused on a limited number of inflammatory proteins, neglecting the influence of other physiological factors. It remains unclear whether alterations in inflammatory factors causally contribute to the development of ovarian-related diseases. Consequently, further investigation into the pathogenesis of these diseases, particularly the role of inflammatory proteins, is crucial.
MR is a causal inference approach that employs genetic variations as instrumental variables. The fundamental principle is to utilize the random distribution of genotypes to infer causal relationships between biological exposures and disease outcomes.12 This design minimizes confounding factors and reverse causality bias, which are prevalent in traditional epidemiological studies, thus offering more robust evidence for causal effect estimation.13 Consequently, MR provides greater credibility and advantages over conventional observational studies in revealing causal relationships. This study employs a two-sample bidirectional MR approach to comprehensively investigate the genetic-based causal relationships between 91 circulating inflammatory proteins and six ovarian-related diseases, including ovarian cyst, PCOS, ovarian dysfunction, primary ovarian failure, benign neoplasm of ovary, and malignant neoplasm of ovary. The objective is to offer new theoretical foundations and practical guidance for the prediction and treatment of ovarian-related diseases.
Materials and Methods
Study Design
This study aims to investigate the causal relationship between 91 circulating inflammatory proteins and ovarian-related diseases through a two-sample bidirectional Mendelian Randomization (MR) analysis. The MR analysis is based on three core assumptions: (1) genetic variation used as instrumental variables (IVs) is strongly associated with the exposure; (2) genetic variation is independent of any known or unknown confounding factors; and (3) genetic variation affects the outcome only through the exposure, without involving any other pathways.14 We utilized publicly available genome-wide association study (GWAS) data on 91 circulating inflammatory proteins and six ovarian-related diseases for this research. First, circulating inflammatory proteins were selected as genetic instrumental variables to explore their causal association with ovarian-related diseases. Second, genetic instrumental variables associated with ovarian-related diseases were employed to analyze the causal relationship between these diseases and circulating inflammatory proteins. This study does not require ethical approval as it relies on publicly available summary data from large-scale GWAS and consortia.
Data Sources
The data for the 91 circulating inflammatory proteins were obtained from GWAS utilizing the Olink Target Inflammation panel. This study was conducted across 11 cohorts and included 14,824 participants of European descent. Summary statistics for each circulating inflammatory protein GWAS are publicly available from the GWAS catalog (registration numbers GCST90274758 to GCST90274848).6
The ovarian-related diseases studied in this paper include ovarian cyst, PCOS, ovarian dysfunction, primary ovarian failure, benign neoplasm of ovary, and malignant neoplasm of ovary. The genetic data for these diseases was obtained from the 11th round of analysis of the FinnGen biobank. The study cohorts consisted of individuals of European descent who provided informed consent, and the case diagnoses adhered strictly to the International Classification of Diseases, 10th edition (ICD-10) code standards. GWAS data for these six ovarian-related diseases are publicly available via the following link: https://r11.finngen.fi//15 Table 1 and Supplementary Table S1 provide detailed information on the included GWAS.
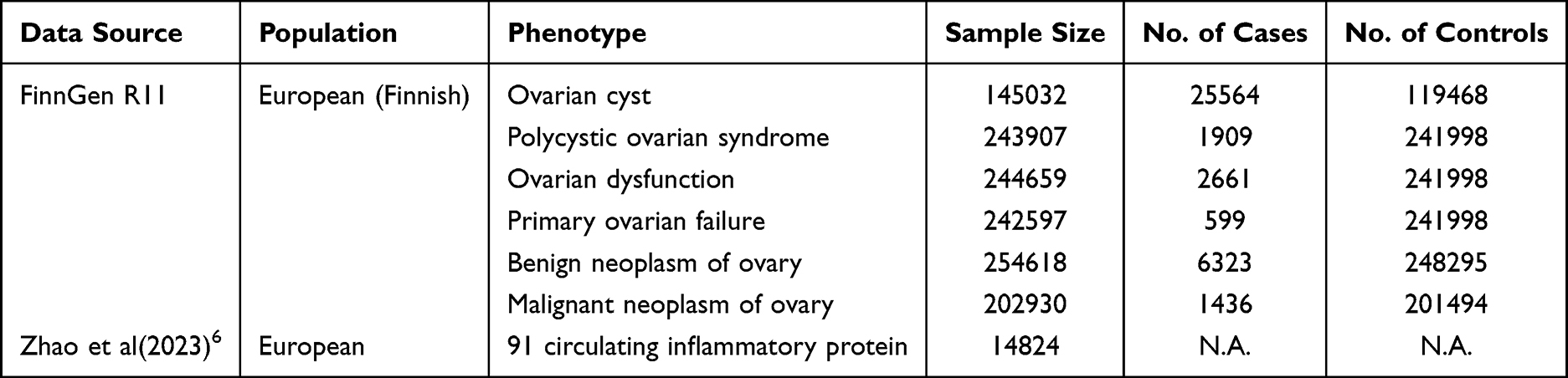 |
Table 1 Information on the GWAS Data Cohort Used to Conduct MR Analysis |
Selection of Genetic Instrumental Variables
To ensure that single nucleotide polymorphisms (SNPs) used as instrumental variables (IVs) were strongly associated with the exposure, we followed a series of steps based on the three core assumptions of MR analyses. First, we selected IVs for circulating inflammatory proteins based on previous studies, applying a screening criterion of P < 1 × 10−5.16 In the inverse MR analysis, the screening criterion for IVs associated with ovarian-related diseases was P < 5 × 10−8, due to the limited number of SNPs at this significance threshold. Next, we assessed the linkage disequilibrium (LD) among risk factor SNPs using PLINK clustering and excluded SNPs with LD coefficients (r2) > 0.001 and physical distances of less than 10,000 kb to ensure independence among SNPs and avoid genetic pleiotropy from interfering with the results.17 During the harmonization process, palindromic SNPs that were inconsistent or had intermediate allele frequencies were excluded. Finally, the strength of each SNP was assessed using the F-statistic, with SNPs having an F-statistic > 10 considered as strongly correlated.18
Statistical Analysis
This study employs a bidirectional two-sample MR analysis to investigate the causal relationship between 91 circulating inflammatory proteins and six ovarian-related diseases. In the analysis, we applied five methods—IVW, MR-Egger, weighted median, simple mode, and weighted mode—to estimate causal effects. Under the assumption that IVs are free of pleiotropy, the IVW method was selected as the primary analytical approach due to its higher statistical power and validity.19 We used Cochran’s Q test to assess the heterogeneity of SNPs in the IVW analysis. If no significant heterogeneity was detected, a fixed-effects model was used;20 if significant heterogeneity (P < 0.05) was detected, we applied the MR-PRESSO test to evaluate potential outliers, remove them from the analysis, and perform the analysis again.21 The MR-Egger intercept test was used to assess horizontal pleiotropy. A P-value greater than 0.05 indicated the absence of horizontal pleiotropy, confirming the reliability of the MR analysis results.22,23 To ensure the robustness of the results, a leave-one-out sensitivity analysis was performed to identify any SNPs that might significantly impact the results.23 MR-Steiger was used to assess the correct causal direction. If the exposure is likely to cause the outcome, “TRUE” is used; if the exposure is unlikely to cause the outcome, “FALSE” is used.24 All statistical analyses were performed using R software (version 4.4.2), with the Mendelian Randomization package (version 0.10.0) and the Two Sample MR package (version 0.5.7).
Results
The specific results of the MR analysis of the levels of 91 circulating inflammatory proteins and ovarian-related diseases are detailed in Supplementary Tables S2 and S3. Supplementary Tables S4 and S5 provide detailed information on the genetic instrumental variables for causal relationships between the levels of 91 circulating inflammatory proteins and ovarian-related diseases. In this study, the F-statistics for all SNPs used as IVs were greater than 10, indicating that these IVs had high predictive power and minimizing the potential for bias due to weak instruments.
Positive MR Results
Effects of 91 Circulating Inflammatory Proteins on Ovarian Cyst
Based on the results from the IVW method, six circulating inflammatory proteins were found to be associated with a reduced risk of ovarian cysts: C-C motif chemokine 4 (CCL4) (OR: 0.963, 95% CI: 0.931–0.995, P = 0.024), Fibroblast growth factor 19 (FGF-19) (OR: 0.948, 95% CI: 0.899–0.999, P = 0.047), Interleukin-17C (IL-17C) (OR: 0.944, 95% CI: 0.894–0.998, P = 0.042), Programmed cell death 1 ligand 1 (PD-L1) (OR: 0.875, 95% CI: 0.822–0.930, P < 0.001), SIR2-like protein 2 (SIRT2) (OR: 0.931, 95% CI: 0.871–0.994, P = 0.033), and TNF-related apoptosis-inducing ligand (TRAIL) (OR: 0.952, 95% CI: 0.920–0.986, P = 0.005). Additionally, elevated levels of two inflammatory proteins were associated with an increased risk of ovarian cysts: Interleukin-6 (IL-6) (OR: 1.138, 95% CI: 1.052–1.230, P = 0.001) and T-cell surface glycoprotein CD6 isoform (CD6) (OR: 1.051, 95% CI: 1.014–1.090, P = 0.007) (Figure 1). Cochran’s Q test and MR-Egger test results indicated no significant heterogeneity between circulating inflammatory proteins and ovarian cysts (P > 0.05). Horizontal pleiotropy was detected only for TRAIL in the MR-Egger regression intercept analysis (P < 0.05). MR-PRESSO analysis did not identify significant outliers (Supplementary Table S6). In the leave-one-out analysis, causal associations identified for FGF-19 and SIRT2 were found to be influenced by a specific instrumental variable, casting doubt on the robustness of the data, leading to their exclusion (Supplementary Figure 1A). The forest plot displayed the causal effects of individual SNPs of circulating inflammatory proteins on ovarian cysts (Supplementary Figure 2A), while the scatter plot visually represented the associations (Supplementary Figure 3A). The MR-Steiger analysis confirmed the correctness of the directionality, with no evidence of reverse causality.
 |
Figure 1 Forest plot of MR Results for causal association of 91 circulating inflammatory proteins with ovarian-related diseases. |
Effects of 91 Circulating Inflammatory Proteins on PCOS
Based on the results from the IVW method, elevated levels of three circulating inflammatory proteins were associated with a reduced risk of PCOS: C-C motif chemokine 20 (CCL20) (OR: 0.822, 95% CI: 0.678–0.996, P = 0.046), Interleukin-6 (IL-6) (OR: 0.694, 95% CI: 0.536–0.899, P = 0.006), and Interleukin-7 (IL-7) (OR: 0.760, 95% CI: 0.588–0.982, P = 0.035). Additionally, elevated levels of three inflammatory proteins were associated with an increased risk of PCOS: Interleukin-20 receptor subunit alpha (IL-20RA) (OR: 1.249, 95% CI: 1.010–1.544, P = 0.040), Monocyte chemoattractant protein-3 (CCL7) (OR: 1.214, 95% CI: 1.022–1.441, P = 0.027), and Tumor necrosis factor ligand superfamily member 14 (TNFSF14) (OR: 1.161, 95% CI: 1.006–1.341, P = 0.041) (Figure 1). Cochran’s Q test and MR-Egger test results indicated no significant heterogeneity between circulating inflammatory proteins and PCOS. In the MR-Egger regression intercept analysis, evidence of horizontal pleiotropy was found only for CCL20. The MR-PRESSO analysis detected no significant outliers (Supplementary Table S6). In the leave-one-out analysis, the causal associations identified for IL-7 and TNFSF14 were influenced by a specific instrumental variable, casting doubt on the robustness of the data, leading to their exclusion (Supplementary Figure 1B). The forest plot displayed the causal effects of individual SNPs of circulating inflammatory proteins on PCOS (Supplementary Figure 2B), while the scatter plot visually represented the associations (Supplementary Figure 3B). The MR-Steiger analysis confirmed the correctness of the directionality, with no evidence of reverse causality.
Effects of 91 Circulating Inflammatory Proteins on Ovarian Dysfunction
According to the results from the IVW method, elevated levels of three circulating inflammatory proteins were associated with a reduced risk of ovarian dysfunction: Interleukin-6 (IL-6) (OR: 0.715, 95% CI: 0.574–0.889, P = 0.003), Interleukin-7 (IL-7) (OR: 0.795, 95% CI: 0.640–0.988, P = 0.038), and SIR2-like protein 2 (SIRT2) (OR: 0.791, 95% CI: 0.658–0.951, P = 0.012). Additionally, elevated levels of three inflammatory proteins were associated with an increased risk of ovarian dysfunction: Fibroblast growth factor 5 (FGF-5) (OR: 1.095, 95% CI: 1.002–1.196, P = 0.045), Interleukin-20 receptor subunit alpha (IL-20RA) (OR: 1.218, 95% CI: 1.017–1.458, P = 0.032), and Neurotrophin-3 (NT-3) (OR: 1.184, 95% CI: 1.014–1.383, P = 0.032) (Figure 1). Cochran’s Q test and MR-Egger test results indicated no significant heterogeneity between circulating inflammatory proteins and ovarian dysfunction. The MR-Egger regression intercept analysis showed no evidence of horizontal pleiotropy between circulating inflammatory proteins and ovarian dysfunction. The MR-PRESSO analysis detected no significant outliers (Supplementary Table S6). In the leave-one-out analysis, the causal association identified for IL-7 was found to be influenced by a specific instrumental variable, casting doubt on the robustness of the data, leading to its exclusion (Supplementary Figure 1C). The forest plot displayed the causal effects of individual SNPs of circulating inflammatory proteins on ovarian dysfunction (Supplementary Figure 2C), while the scatter plot visually represented the associations (Supplementary Figure 3C). The MR-Steiger analysis confirmed the correctness of the directionality, with no evidence of reverse causality.
Effects of 91 Circulating Inflammatory Proteins on Primary Ovarian Failure
Based on the results from the IVW method, elevated levels of one circulating inflammatory protein were associated with a reduced risk of primary ovarian failure: T-cell surface glycoprotein CD6 isoform (CD6) (OR: 0.796, 95% CI: 0.643–0.984, P = 0.035). Additionally, elevated levels of one circulating inflammatory protein were associated with an increased risk of primary ovarian failure: Interleukin-33 (IL-33) (OR: 1.569, 95% CI: 1.088–2.261, P = 0.016) (Figure 1). Cochran’s Q test and MR-Egger test results indicated no significant heterogeneity between circulating inflammatory proteins and primary ovarian failure. The MR-Egger regression intercept analysis showed no evidence of horizontal pleiotropy between circulating inflammatory proteins and primary ovarian failure. The MR-PRESSO analysis detected no significant outliers (Supplementary Table S6). In the leave-one-out analysis, the causal association identified for CD6 was found to be influenced by a specific instrumental variable, casting doubt on the robustness of the data, leading to its exclusion (Supplementary Figure 1D). The forest plot displayed the causal effects of individual SNPs of circulating inflammatory proteins on primary ovarian failure (Supplementary Figure 2D), while the scatter plot visually represented the associations (Supplementary Figure 3D). The MR-Steiger analysis confirmed the correctness of the directionality, with no evidence of reverse causality.
Effects of 91 Circulating Inflammatory Proteins on Benign Neoplasm of Ovary
According to the results from the IVW method, elevated levels of three circulating inflammatory proteins were associated with a reduced risk of benign ovarian neoplasm: C-C motif chemokine 28 (CCL28) (OR: 0.887, 95% CI: 0.797–0.988, P = 0.029), Fms-related tyrosine kinase 3 ligand (Flt3L) (OR: 0.910, 95% CI: 0.829–1.000, P = 0.049), and Interleukin-13 (IL-13) (OR: 0.902, 95% CI: 0.820–0.992, P = 0.033). Additionally, elevated levels of three inflammatory proteins were associated with an increased risk of benign ovarian neoplasm: Adenosine Deaminase (ADA) (OR: 1.100, 95% CI: 1.027–1.178, P = 0.006), C-C motif chemokine 23 (CCL23) (OR: 1.086, 95% CI: 1.005–1.174, P = 0.037), and Monocyte chemoattractant protein-1 (CCL2) (OR: 1.146, 95% CI: 1.060–1.239, P = 0.001) (Figure 1). Cochran’s Q test and MR-Egger test results indicated heterogeneity for Flt3L. Although the Inverse Variance Weighted (multiplicative random effects) analysis showed significant results, it did not pass the MR-PRESSO test. No heterogeneity or evidence of horizontal pleiotropy was found for the remaining circulating inflammatory proteins. The MR-PRESSO analysis detected no significant outliers (Supplementary Table S6). The leave-one-out analysis confirmed the robustness of the results (Supplementary Figure 1E). The forest plot displayed the causal effects of individual SNPs of circulating inflammatory proteins on benign ovarian neoplasm (Supplementary Figure 2E), while the scatter plot visually represented the associations (Supplementary Figure 3E). The MR-Steiger analysis confirmed the correctness of the directionality, with no evidence of reverse causality.
Effects of 91 Circulating Inflammatory Proteins on Malignant Neoplasm of Ovary
According to the results from the IVW method, elevated levels of three circulating inflammatory proteins were associated with a reduced risk of malignant ovarian neoplasm: C-C motif chemokine 20 (CCL20) (OR: 0.778, 95% CI: 0.623–0.970, P = 0.026), C-C motif chemokine 25 (CCL25) (OR: 0.806, 95% CI: 0.704–0.923, P = 0.002), and Fms-related tyrosine kinase 3 ligand (Flt3L) (OR: 0.775, 95% CI: 0.663–0.907, P = 0.002). Additionally, elevated levels of five circulating inflammatory proteins were associated with an increased risk of malignant ovarian neoplasm: Delta and Notch-like epidermal growth factor-related receptor (DNER) (OR: 1.233, 95% CI: 1.010–1.505, P = 0.040), Interleukin-18 (IL-18) (OR: 1.220, 95% CI: 1.015–1.466, P = 0.034), Interleukin-8 (IL-8) (OR: 1.291, 95% CI: 1.030–1.618, P = 0.027), Monocyte chemoattractant protein-1 (CCL2) (OR: 1.262, 95% CI: 1.074–1.483, P = 0.005), and Neurotrophin-3 (NT-3) (OR: 1.249, 95% CI: 1.011–1.543, P = 0.039) (Figure 1). Cochran’s Q test and MR-Egger test results indicated no significant heterogeneity between circulating inflammatory proteins and malignant ovarian neoplasm. The MR-Egger regression intercept analysis showed no evidence of horizontal pleiotropy. The MR-PRESSO analysis detected no significant outliers (Supplementary Table S6). The leave-one-out analysis confirmed the robustness of the results (Supplementary Figure 1F). The forest plot displayed the causal effects of individual SNPs of circulating inflammatory proteins on malignant ovarian neoplasm (Supplementary Figure 2F), while the scatter plot visually represented the associations (Supplementary Figure 3F). The MR-Steiger analysis confirmed the correctness of the directionality, with no evidence of reverse causality.
Reverse Mendelian Randomization Results
A reverse MR analysis was conducted with six ovarian-related diseases as exposure factors and 91 circulating inflammatory proteins as outcome variables. According to the results from the IVW method, no causal relationships were found between primary ovarian insufficiency, ovarian malignancy, and circulating inflammatory proteins. Ovarian cysts were positively causally associated with Neurotrophin-3 (NT-3) (OR: 1.155, 95% CI: 1.050–1.270, P = 0.003). Polycystic ovary syndrome (PCOS) was positively causally associated with Interleukin-17C (IL-17C) (OR: 1.067, 95% CI: 1.019–1.118, P = 0.006), Natural killer cell receptor 2B4 (CD244) (OR: 1.044, 95% CI: 1.002–1.087, P = 0.039), and TNF-beta (TNFB) (OR: 1.067, 95% CI: 1.018–1.118, P = 0.007). Ovarian dysfunction was positively causally associated with C-C motif chemokine 23 (CCL23) (OR: 1.081, 95% CI: 1.003–1.164, P = 0.040). Benign ovarian tumors were positively causally associated with Fibroblast growth factor 19 (FGF-19) (OR: 1.113, 95% CI: 1.011–1.225, P = 0.029) and Interleukin-12 subunit beta (IL-12B) (OR: 1.115, 95% CI: 1.014–1.227, P = 0.025), and negatively causally associated with Fibroblast growth factor 5 (FGF-5) (OR: 0.894, 95% CI: 0.806–0.992, P = 0.034), Interleukin-20 receptor subunit alpha (IL-20RA) (OR: 0.879, 95% CI: 0.784–0.987, P = 0.029), and Neurotrophin-3 (NT-3) (OR: 0.909, 95% CI: 0.827–0.999, P = 0.048) (Figure 2). Sensitivity analyses indicated that the p-values for both Cochran’s Q test and MR-Egger intercept test were greater than 0.05, suggesting no heterogeneity or horizontal pleiotropy (Supplementary Table S7). In the leave-one-out analysis, the causal associations identified for CCL23, IL-12B, and IL-20RA may be influenced by a specific instrumental variable (IV), casting doubt on the robustness of the data; therefore, they were excluded (Supplementary Figure 4A–D). The forest plots and scatter plots of the results are shown in Supplementary Figure 4E–L.
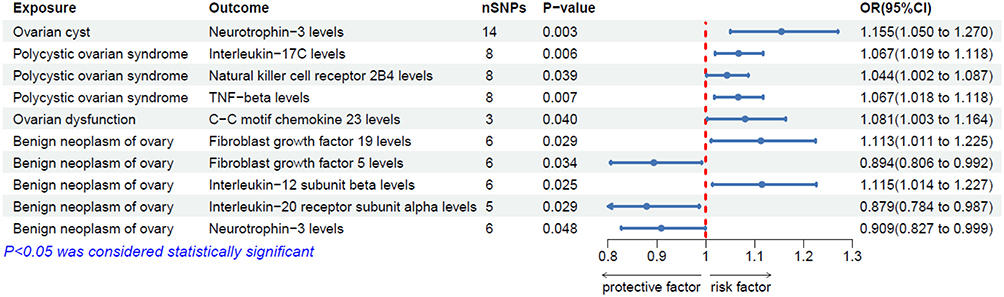 |
Figure 2 Forest plot of MR Results for causal association of ovarian-related diseases with 91 circulating inflammatory proteins. |
Discussion
In this study, we utilized a large amount of publicly available genetic data and employed a MR study design to assess the causal associations between 91 circulating inflammatory proteins and six ovarian-related diseases at the genetic level. The forward MR analysis revealed that 11 circulating inflammatory proteins were negatively associated with ovarian-related diseases, while 16 circulating inflammatory proteins were positively associated with ovarian-related diseases. The reverse MR analysis further indicated that ovarian-related diseases had positive causal relationships with five circulating inflammatory proteins and negative causal relationships with two circulating inflammatory proteins.
Ovarian malignant tumors are classified into several types, including epithelial, germ cell, sex cord-stromal, and metastatic tumors. OC is the third most common malignancy of the female reproductive system and has the highest mortality rate among gynecological cancers, posing a significant threat to women’s health.25 Inflammation plays a crucial role in the onset and progression of most cancers.26 Studies have shown that the pro-inflammatory cytokine IL-8 can promote the migration of OC cells in vitro, initiate the epithelial-mesenchymal transition (EMT), and activate the Wnt/β-catenin signaling pathway.27 Additionally, compared to healthy controls, the concentrations of IL-8 and its antibody in the serum of OC patients are significantly elevated.28 The levels of the pro-inflammatory cytokine IL-18 are also significantly increased in the tumor tissues and serum of OC patients.29 Relevant studies have shown that exogenous CCL2 not only enhances the invasion and adhesion capabilities of ovarian cancer (OC) cells but also promotes cell proliferation by activating the MAPK/ERK signaling pathway and regulating the expression of JUN, RELB, and NF-κB2.30,31 Recombinant CCL2, by downregulating the expression of TNFSF15, can promote tumor angiogenesis and accelerate the progression of OC.32 In the survival analysis of EOC, the addition of the biomarker NT-3 improves the prediction of overall survival.33 Conversely, certain inflammatory proteins can exert anti-cancer effects by enhancing immune responses. CCL20 can induce monocyte migration in vitro and, in vivo, recruit tumor-associated macrophages through interaction with its receptor CCR6.34,35 Furthermore, CCL20 can also recruit dendritic cells, thereby enhancing the immune system’s anti-cancer response.36 Similarly, FLT3L exerts an immunostimulatory effect on peripheral monocyte-derived dendritic cells and natural killer cells. Targeting mature peripheral blood cells with FLT3L could serve as a promising tool for cancer immunotherapy.37 Gene therapies based on Flt3L-related mechanisms have already advanced to the human clinical trial phase.38 Our study also identified a correlation between CCL25 and ovarian malignancies. Previous research has shown that both CCL25 and its receptor CCR9 are highly expressed in OC tissues.39 The interaction between CCL25 and CCR9 not only facilitates the migration and invasion of OC cells but also plays a crucial role in cisplatin resistance.40,41 These findings suggest that the role of inflammatory proteins is not exclusively anti-cancer or pro-cancer but rather results from a complex interplay of multiple factors. Inflammatory-related proteins are pivotal in the initiation and progression of OC. Given these observations, we propose that these proteins hold potential as biomarkers for early diagnosis and prognosis evaluation, and may serve as molecular targets for therapeutic interventions, offering new directions for future research in ovarian malignancies.
PCOS is a prevalent endocrine and metabolic disorder. Diagnosis is confirmed when two of the following three criteria are met: oligo-ovulation or anovulation, clinical or biochemical signs of hyperandrogenism, and polycystic ovarian morphology.42 CCL7, also known as monocyte chemoattractant protein-3, primarily attracts monocytes and neutrophils, playing a pivotal role in various inflammatory processes. Recent studies have shown that elevated levels of CCL7 are associated with the severity of metabolic dysfunction in PCOS.43 Moreover, CCL7 may exacerbate ovarian inflammation by recruiting inflammatory cells, potentially impairing ovarian function and disrupting hormonal balance.44 Our findings align with existing research and underscore the significant role of increased CCL7 levels in the inflammatory and metabolic features of PCOS. Additionally, our study identified a correlation between IL-6 and PCOS. IL-6 contributes to androgen excess and granulosa cell apoptosis in the ovaries, mediated by SFRP4, and interferes with insulin signaling, thereby promoting the development of PCOS induced by insulin resistance.45 Furthermore, excessive production of IL-6 may stimulate ovarian androgen synthesis, leading to irregular menstruation and ovulatory dysfunction.46 However, the relationship between IL-6 levels and PCOS remains incompletely understood.47 Meta-analysis suggests that elevated IL-6 levels are not an inherent characteristic of PCOS but may serve as an effective biomarker for monitoring treatment response in PCOS.48 The association between PCOS and chronic inflammation has been extensively studied. PCOS often leads to various health complications linked to adipose tissue accumulation, including obesity, insulin resistance, metabolic syndrome, and type 2 diabetes. In turn, obesity and hyperinsulinemia further exacerbate the chronic inflammatory state.49 Studies have demonstrated that IL-17C is closely associated with neutrophil-mediated inflammation,50 TNFB polymorphisms are linked to inflammation and immune regulatory responses,51 and CD244 modulates immune responses in NK cells, CD8+ T cells, and other immune cells by providing either stimulatory or inhibitory signals.52 These molecules may play a crucial role in the chronic inflammation observed in PCOS. Consequently, inflammation-related proteins have significant clinical implications in PCOS, serving not only as biomarkers for disease monitoring but also as potential therapeutic targets. Targeting these inflammation-related proteins with monoclonal antibodies or other inhibitors may effectively reduce inflammation in PCOS patients, thereby improving metabolic function and advancing therapeutic strategies for PCOS management.
In clinical practice, ovarian benign tumors and ovarian cysts are relatively common. Ovarian benign tumors are classified based on their tissue origin and pathological characteristics, including epithelial tumors, germ cell tumors, and sex cord-stromal tumors. Broadly speaking, ovarian cysts can also be categorized as a type of ovarian benign tumor. The pathogenesis of these conditions remains incompletely understood. Research has demonstrated that angiogenesis plays a key role in various pathological processes in the ovaries, including the formation of follicular cysts, PCOS, ovarian hyperstimulation syndrome, and both benign and malignant ovarian tumors.53 CCL23 induces endothelial cell migration by activating CCR1 on endothelial cells, thereby promoting angiogenesis.54 Additionally, CCL2-induced angiogenesis is closely linked to macrophage recruitment.55 Hormones are critical in the progression of ovarian-related diseases. ADA regulates adenosine levels by converting adenosine to inosine, and extracellular adenosine enhances the stimulatory effects of gonadotropins on granulosa and luteal cells.56,57 Furthermore, we hypothesize that activation of the inflammatory response may reduce the risk of ovarian-related tumors. IL-13 can alleviate insulin resistance by promoting the differentiation of anti-inflammatory macrophages in adipose tissue.58 CCL28 is a chemokine that is crucial for normal mucosal immune function.30 It can bind to two receptors: CCR3 and CCR10.59 CCL28 drives the migration of T cells and B cells expressing CCR10 to mucosal tissues and guides the migration of eosinophils expressing CCR3.60,61 In reverse MR analysis, we also identified causal relationships between certain inflammatory proteins and ovarian benign tumors.
Inflammation plays a crucial physiological role in folliculogenesis and ovulation, and excessive activation of the inflammatory response may impact the formation of ovarian cysts.62 CD6 is a multifunctional lymphocyte receptor that can either positively or negatively regulate lymphocyte (T, B1a, NK) function, depending on the intracellular signaling environment, and is associated with various inflammatory diseases.63 IL-6 is a pleiotropic pro-inflammatory cytokine, and its dysregulation is linked to chronic inflammation and multifactorial autoimmune diseases.64 In contrast, we hypothesize that a balanced inflammatory response may reduce the risk of its occurrence. CCL4 is a pro-inflammatory chemokine that, in conjunction with CCL3, induces the aggregation of dendritic cells, neutrophils, monocytes, macrophages, natural killer (NK) cells, and T cells at the site of inflammation.65 In various mucosal infection models, IL-17C plays a critical role in enhancing the innate immune function of epithelial cells.66 Furthermore, studies have shown that anti-PD-L1 therapy results in significant depletion of primordial/primary ovarian follicles.67 In reverse MR analysis, a correlation between ovarian cysts and NT-3 was identified. During follicular development, NT-3 promotes the transition from primordial follicles to primary follicles by upregulating several target genes associated with cytoskeletal and plasma membrane formation.68 Our study elucidates the potential mechanisms of various inflammatory proteins in ovarian benign tumors and ovarian cysts, providing a theoretical foundation for the development of novel diagnostic biomarkers. Future research will further explore the potential clinical diagnostic applications of these molecules.
In this study, we explored the relationship between two ovarian dysfunction diseases and circulating inflammatory proteins. Primary ovarian failure represents the final stage of ovarian insufficiency, typically affecting women under the age of 40 who experience amenorrhea, hypergonadotropic hypogonadism, and infertility.69 Our study found that IL-33 increases the risk of primary ovarian failure. IL-33 is a pro-inflammatory cytokine belonging to the IL-1 family. Research has shown that, with the development of early-onset ovarian insufficiency, IL-33 levels in follicular fluid (but not in serum) progressively increase.70 Supplementation with recombinant IL-33 exacerbates ovarian dysfunction by promoting tissue inflammation and fibrosis.49,71 High levels of IL-33 also enhance the invasiveness of human ovarian endometriosis stromal cells via the ST2/MAPK/MMP-9 signaling pathway, ultimately leading to ovarian dysfunction and the progression of endometriosis.72 The ovarian dysfunction discussed in this study encompasses various diseases, including PCOS and primary ovarian failure, but the research is not focused on a specific disease and should thus be considered as a general reference. Signal transduction of multiple molecules, including NT-3, plays a crucial role in regulating ovarian function, including follicle assembly, activation of primordial follicles, follicular growth and development, oocyte maturation, steroidogenesis, ovulation, and luteal formation.73 Therefore, dysregulation of the NT system may negatively impact ovarian function. In vitro studies on ovarian cells and fibroblast growth factors found no significant mitogenic effect of FGF-5 on granulosa and theca cells.74 Additionally, some studies have indicated that SIRT2 can influence apoptosis, proliferation, and secretion in ovarian granulosa cells. Knockdown of Sirt2 increases mitochondrial autophagy levels and decreases oocyte maturation rates.75 Inflammatory proteins play a critical regulatory role in ovarian development, and further research on these proteins will aid in better understanding the pathophysiological mechanisms underlying ovarian dysfunction, providing important theoretical support for early diagnosis and timely treatment.
Building on large-scale genomic studies already published, our research employed the MR method to explore the causal relationship between circulating inflammatory proteins and ovarian-related diseases. Through comprehensive sensitivity analyses, we ensured the reliability and stability of our findings. However, there are several limitations to our study. First, all exposure factors and outcome-related GWAS data used were derived from participants of European ancestry. Consequently, the conclusions of our study are limited by ethnic background and cannot be directly generalized to non-European populations. Second, despite the focus on European ancestry participants, residual confounding effects due to population stratification cannot be entirely excluded. Third, since the precise functions of most SNPs remain poorly understood, residual pleiotropic effects may still be present. Fourth, GWAS data from different institutions could introduce selection bias. Furthermore, because MR studies rely on summary-level data, they inherently exhibit a cross-sectional nature. As a result, while the study may suggest potential causal relationships, there are inherent limitations in inferring the temporal direction of these relationships, as the timing and sequence of events cannot be fully captured. Given these limitations, we encourage researchers worldwide to consider our preliminary findings and build upon them in future studies.
Conclusion
In summary, through comprehensive MR analysis, we identified the complex causal relationships at the genetic level between 91 circulating inflammatory proteins and 6 ovarian-related diseases. The forward MR analysis revealed that 27 circulating inflammatory proteins were associated with ovarian-related diseases. In contrast, the reverse MR analysis showed that ovarian-related diseases were causally linked to 7 circulating inflammatory proteins. These findings suggest that inflammatory proteins may play a significant role in the onset and progression of ovarian-related diseases through mechanisms such as regulating immune responses, influencing hormone metabolism, or participating in other biological processes. Our results not only provide potential biomarkers and therapeutic targets for the clinical prevention and treatment of ovarian-related diseases but also open new avenues for exploring the molecular mechanisms linking inflammatory proteins and ovarian function.
Abbreviations
MR, Mendelian randomization; PCOS, Polycystic ovary syndrome; IVW, Inverse Variance Weighted; OC, Ovarian cancer; EOC, Epithelial ovarian cancer; GWAS, Genome-wide association study; IVs, Instrumental variables; SNPs, Single nucleotide polymorphisms; LD, Linkage disequilibrium.
Data Sharing Statement
The original contributions of this study are included in the article and/or Supplementary Material. For further inquiries, please contact the corresponding author.
Ethical Approval
The studies involving human participants were exempted from ethical review according to the Ethical Review Measures for Life Sciences and Medical Research Involving Humans (2023), issued in China. This research was based entirely on publicly available and legally obtained databases, and all data were fully anonymized prior to analysis. No identifiable personal information, human intervention, or new collection of human biological samples was involved. Therefore, the study met the criteria for exemption from ethical review under current Chinese legislation.
Acknowledgments
All authors thank the patients and sequencers who provided samples and the publicly available databases.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Funding
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.
Disclosure
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
This paper has been uploaded to Authorea as a preprint: https://www.authorea.com/doi/full/10.22541/au.174846080.02704092/v1.
References
1. Momenimovahed Z, Tiznobaik A, Taheri S, et al. Ovarian cancer in the world: epidemiology and risk factor. Int J Women’s Health. 2019;11:287–299. doi:10.2147/IJWH.S197604
2. Escobar-Morreale HF. Polycystic ovary syndrome: definition, aetiology, diagnosis and treatment. Nat Rev Endocrinol. 2018;14(5):270–284. doi:10.1038/nrendo.2018.24
3. Azziz R, Carmina E, Chen Z, et al. Polycystic ovary syndrome. Nat Rev Dis Primers. 2016;2:16057. doi:10.1038/nrdp.2016.57
4. Pavlik EJ, Ueland FR, Miller RW, et al. Frequency and disposition of ovarian abnormalities followed with serial transvaginal ultrasonography. Obstetrics Gynecol. 2013;122(2 Pt 1):210–217. doi:10.1097/AOG.0b013e318298def5
5. Alper MM, Garner PR, Seibel MM. Premature ovarian failure. Current concepts. J Reprod Med. 1986;31(8):699–708.
6. Zhao JH, Stacey D, Eriksson N, et al. Genetics of circulating inflammatory proteins identifies drivers of immune-mediated disease risk and therapeutic targets. Nat Immunol. 2023;24(9):1540–1551. doi:10.1038/s41590-023-01588-w
7. Nowak M, Glowacka E, Szpakowski M, et al. Proinflammatory and immunosuppressive serum, ascites and cyst fluid cytokines in patients with early and advanced ovarian cancer and Benign neoplasm of ovary. Neuro Endocrinology Letters. 2010;31(3):375–383.
8. Clendenen TV, Lundin E, Zeleniuch-Jacquotte A, et al. Circulating inflammation markers and risk of epithelial ovarian cancer. Cancer Epidemiol Biomarkers Prev. 2011;20(5):799–810. doi:10.1158/1055-9965.EPI-10-1180
9. Zhang F-L, Kong L, Zhao A-H, et al. Inflammatory cytokines as key players of apoptosis induced by environmental estrogens in the ovary. Environ Res. 2021;198:111225. doi:10.1016/j.envres.2021.111225
10. Deligeoroglou E, Vrachnis N, Athanasopoulos N, et al. Mediators of chronic inflammation in polycystic ovarian syndrome. Gynecol Endocrinol. 2012;28(12):974–978. doi:10.3109/09513590.2012.683082
11. Glintborg D, Andersen M, Richelsen B, et al. Plasma monocyte chemoattractant protein-1 (MCP-1) and macrophage inflammatory protein-1alpha are increased in patients with polycystic ovary syndrome (PCOS) and associated with adiposity, but unaffected by pioglitazone treatment. Clin Endocrinol. 2009;71(5):652–658. doi:10.1111/j.1365-2265.2009.03523.x
12. Evans DM, Davey Smith G. Mendelian randomization: new applications in the coming age of hypothesis-free causality. Ann Rev Genomics Hum Genet. 2015;16(1):327–350. doi:10.1146/annurev-genom-090314-050016
13. Skrivankova VW, Richmond RC, Woolf BA, et al. Strengthening the reporting of observational studies in epidemiology using mendelian randomization: the STROBE-MR statement. JAMA. 2021;326(16):1614–1621. doi:10.1001/jama.2021.18236
14. Emdin CA, Khera AV, Kathiresan S. Mendelian randomization. JAMA. 2017;318(19):1925–1926. doi:10.1001/jama.2017.17219
15. Kurki MI, Karjalainen J, Palta P, et al. FinnGen provides genetic insights from a well-phenotyped isolated population. Nature. 2023;613(7944):508–518. doi:10.1038/s41586-022-05473-8
16. Xu J, Chen R, Yang Y, et al. Genetically predicted 91 circulating inflammatory proteins in relation to risk of urological malignancies: a Mendelian randomization study. Aging. 2024;16(12):10366–10379. doi:10.18632/aging.205934
17. Hemani G, Zheng J, Elsworth B, et al. The MR-base platform supports systematic causal inference across the human phenome. eLife. 2018;7:e34408. doi:10.7554/eLife.34408
18. Burgess S, Thompson SG, Collaboration CCG. Avoiding bias from weak instruments in Mendelian randomization studies. Int J Epidemiol. 2011;40(3):755–764. doi:10.1093/ije/dyr036
19. Burgess S, Davey Smith G, Davies NM, et al. Guidelines for performing Mendelian randomization investigations: update for summer 2023. Wellcome Open Res. 2019;4:186. doi:10.12688/wellcomeopenres.15555.1
20. Hemani G, Bowden J, Davey Smith G. Evaluating the potential role of pleiotropy in Mendelian randomization studies. Human Molecular Genetics. 2018;27(R2):R195–R208. doi:10.1093/hmg/ddy163
21. Verbanck M, Chen C-Y, Neale B, et al. Publisher Correction: detection of widespread horizontal pleiotropy in causal relationships inferred from Mendelian randomization between complex traits and diseases. Nature Genet. 2018;50(8):1196. doi:10.1038/s41588-018-0164-2
22. Burgess S, Thompson SG. Interpreting findings from Mendelian randomization using the MR-Egger method. Eur j epidemiol. 2017;32(5):377–389. doi:10.1007/s10654-017-0255-x
23. Burgess S, Bowden J, Fall T, et al. Sensitivity analyses for robust causal inference from mendelian randomization analyses with multiple genetic variants. Epidemiology. 2017;28(1):30–42. doi:10.1097/EDE.0000000000000559
24. Hemani G, Tilling K, Davey Smith G. Orienting the causal relationship between imprecisely measured traits using GWAS summary data. PLoS Genetics. 2017;13(11):e1007081. doi:10.1371/journal.pgen.1007081
25. Ferlay J, Soerjomataram I, Dikshit R, et al. Cancer incidence and mortality worldwide: sources, methods and major patterns in GLOBOCAN 2012. Inter J Cancer. 2015;136(5):E359–386. doi:10.1002/ijc.29210
26. Grivennikov SI, Greten FR, Karin M. Immunity, inflammation, and cancer. Cell. 2010;140(6):883–899. doi:10.1016/j.cell.2010.01.025
27. Wen J, Zhao Z, Huang L, et al. IL-8 promotes cell migration through regulating EMT by activating the Wnt/β-catenin pathway in ovarian cancer. J Cell Mol Med. 2020;24(2):1588–1598. doi:10.1111/jcmm.14848
28. Lokshin AE, Winans M, Landsittel D, et al. Circulating IL-8 and anti-IL-8 autoantibody in patients with ovarian cancer. Gynecologic Oncol. 2006;102(2):244–251. doi:10.1016/j.ygyno.2005.12.011
29. Le Page C, Ouellet V, Madore J, et al. From gene profiling to diagnostic markers: IL-18 and FGF-2 complement CA125 as serum-based markers in epithelial ovarian cancer. Inter J Cancer. 2006;118(7):1750–1758. doi:10.1002/ijc.21521
30. Furukawa S, Soeda S, Kiko Y, et al. MCP-1 promotes invasion and adhesion of human ovarian cancer cells. Anticancer Res. 2013;33(11):4785–4790.
31. Liu W, Wang L, Zhang J, et al. Purification of recombinant human chemokine CCL2 in E. coli and its function in ovarian cancer. 3 Biotech. 2021;11(1):8. doi:10.1007/s13205-020-02571-0
32. Deng W, Gu X, Lu Y, et al. Down-modulation of TNFSF15 in ovarian cancer by VEGF and MCP-1 is a pre-requisite for tumor neovascularization. Angiogenesis. 2012;15(1):71–85. doi:10.1007/s10456-011-9244-y
33. Dobilas A, Åkesson A, Leandersson P, et al. A multiplex biomarker assay improves the prediction of survival in epithelial ovarian cancer. Cancer Genomics Proteomics. 2023;20(3):273–280. doi:10.21873/cgp.20380
34. Korbecki J, Grochans S, Gutowska I, et al. CC chemokines in a tumor: a review of pro-cancer and anti-cancer properties of receptors CCR5, CCR6, CCR7, CCR8, CCR9, and CCR10 ligands. Int J Mol Sci. 2020;21(20):7619. doi:10.3390/ijms21207619
35. Nandi B, Shapiro M, Samur MK, et al. Stromal CCR6 drives tumor growth in a murine transplantable colon cancer through recruitment of tumor-promoting macrophages. Oncoimmunology. 2016;5(8):e1189052. doi:10.1080/2162402X.2016.1189052
36. Bonnotte B, Crittenden M, Larmonier N, et al. MIP-3alpha transfection into a rodent tumor cell line increases intratumoral dendritic cell infiltration but enhances (facilitates) tumor growth and decreases immunogenicity. J Immunol. 2004;173(8):4929–4935. doi:10.4049/jimmunol.173.8.4929
37. Matsumura N, Mandai M, Hamanishi J, et al. Immunostimulatory effect of Fms-like tyrosine kinase 3 ligand on peripheral monocyte-derived dendritic cells and natural killer cells: utilization for ovarian cancer treatment. Oncol Rep. 2008;19(2):505–515.
38. Faisal SM, Castro MG, Lowenstein PR. Combined cytotoxic and immune-stimulatory gene therapy using Ad-TK and Ad-Flt3L: translational developments from rodents to glioma patients. Mol ther. 2023;31(10):2839–2860. doi:10.1016/j.ymthe.2023.08.009
39. Singh R, Stockard CR, Grizzle WE, et al. Expression and histopathological correlation of CCR9 and CCL25 in ovarian cancer. Int J Oncol. 2011;39(2):373–381. doi:10.3892/ijo.2011.1059
40. Johnson EL, Singh R, Singh S, et al. CCL25-CCR9 interaction modulates ovarian cancer cell migration, metalloproteinase expression, and invasion. World J Surg Oncol. 2010;8(1):62. doi:10.1186/1477-7819-8-62
41. Johnson EL, Singh R, Johnson-Holiday CM, et al. CCR9 interactions support ovarian cancer cell survival and resistance to cisplatin-induced apoptosis in a PI3K-dependent and FAK-independent fashion. J Ovarian Res. 2010;3(1):15. doi:10.1186/1757-2215-3-15
42. Group R E a-S P C W. Revised 2003 consensus on diagnostic criteria and long-term health risks related to polycystic ovary syndrome (PCOS). Hum Reprod. 2004;19(1):41–47. doi:10.1093/humrep/deh098
43. De Melo Cavalcante RB, Leão LMCSM, Tavares ABW, et al. Fat distribution and its correlation with insulin resistance, androgen markers, and proinflammatory cytokines in polycystic ovary syndrome. Hormone Metab Res. 2025;57(1):25–32. doi:10.1055/a-2386-9281
44. Liu Y, Cai Y, Liu L, et al. Crucial biological functions of CCL7 in cancer. PeerJ. 2018;6:e4928. doi:10.7717/peerj.4928
45. Wang J, Gui R, Li Y, et al. SFRP4 contributes to insulin resistance-induced polycystic ovary syndrome by triggering ovarian granulosa cell hyperandrogenism and apoptosis through the nuclear β-catenin/IL-6 signaling axis. Biochim Biophys Acta Mol Cell Res. 2024;1871(7):119822. doi:10.1016/j.bbamcr.2024.119822
46. Stanczyk FZ, Mandelbaum R, Baker M, et al. Quantitation of 5α-androstanedione in normal women and women with PCOS. J Steroid Biochem Mol Biol. 2023;231:106289. doi:10.1016/j.jsbmb.2023.106289
47. Olszanecka-Glinianowicz M, Banaś M, Zahorska-Markiewicz B, et al. Is the polycystic ovary syndrome associated with chronic inflammation per se?. Eur J Obstet Gynecol Reprod Biol. 2007;133(2):197–202. doi:10.1016/j.ejogrb.2006.10.037
48. Peng Z, Sun Y, Lv X, et al. Interleukin-6 levels in women with polycystic ovary syndrome: a systematic review and meta-analysis. PLoS One. 2016;11(2):e0148531. doi:10.1371/journal.pone.0148531
49. Rudnicka E, Suchta K, Grymowicz M, et al. Chronic low grade inflammation in pathogenesis of PCOS. Int J Mol Sci. 2021;22(7):3789. doi:10.3390/ijms22073789
50. Wen Y, Chen Q, Wang H, et al. Contribution of IL-17C-mediated macrophage polarization to type 17 inflammation in neutrophilic asthma. Cell Commun Signal. 2024;22(1):557. doi:10.1186/s12964-024-01937-8
51. Laddha NC, Dwivedi M, Gani AR, et al. Tumor necrosis factor B (TNFB) genetic variants and its increased expression are associated with vitiligo susceptibility. PLoS One. 2013;8(11):e81736. doi:10.1371/journal.pone.0081736
52. Sun L, Gang X, Li Z, et al. Advances in understanding the roles of CD244 (SLAMF4) in immune regulation and associated diseases. Front Immunol. 2021;12:648182. doi:10.3389/fimmu.2021.648182
53. Abulafia O, Sherer DM. Angiogenesis of the ovary. Am J Clin Exp Obstet Gynecol. 2000;182(1 Pt 1):240–246. doi:10.1016/S0002-9378(00)70519-9
54. Hwang J, Son K-N, Kim CW, et al. Human CC chemokine CCL23, a ligand for CCR1, induces endothelial cell migration and promotes angiogenesis. Cytokine. 2005;30(5):254–263. doi:10.1016/j.cyto.2005.01.018
55. Goede V, Brogelli L, Ziche M, et al. Induction of inflammatory angiogenesis by monocyte chemoattractant protein-1. International Journal of Cancer. 1999;82(5):765–770. doi:10.1002/(SICI)1097-0215(19990827)82:5<765::AID-IJC23>3.0.CO;2-F
56. Zhulai G, Oleinik E, Shibaev M, et al. Adenosine-metabolizing enzymes, adenosine kinase and adenosine deaminase, in cancer. Biomolecules. 2022;12(3):418. doi:10.3390/biom12030418
57. Billig H, Rosberg S, Johanson C, et al. Adenosine as substrate and receptor agonist in the ovary. Steroids. 1989;54(5):523–542. doi:10.1016/0039-128X(89)90045-7
58. Duffen J, Zhang M, Masek-Hammerman K, et al. Modulation of the IL-33/IL-13 axis in obesity by IL-13Rα2. J Immunol. 2018;200(4):1347–1359. doi:10.4049/jimmunol.1701256
59. Pan J, Kunkel EJ, Gosslar U, et al. A novel chemokine ligand for CCR10 and CCR3 expressed by epithelial cells in mucosal tissues. J Immunol. 2000;165(6):2943–2949. doi:10.4049/jimmunol.165.6.2943
60. Lazarus NH, Kunkel EJ, Johnston B, et al. A common mucosal chemokine (mucosae-associated epithelial chemokine/CCL28) selectively attracts IgA plasmablasts. J Immunol. 2003;170(7):3799–3805. doi:10.4049/jimmunol.170.7.3799
61. John AE, Thomas MS, Berlin AA, et al. Temporal production of CCL28 corresponds to eosinophil accumulation and airway hyperreactivity in allergic airway inflammation. Am J Pathol. 2005;166(2):345–353. doi:10.1016/S0002-9440(10)62258-4
62. Boots CE, Jungheim ES. Inflammation and human ovarian follicular dynamics. Seminars Reprod Med. 2015;33(4):270–275. doi:10.1055/s-0035-1554928
63. Enyindah-Asonye G, Li Y, Ruth JH, et al. CD318 is a ligand for CD6. Proc Natl Acad Sci USA. 2017;114(33):E6912–E6921. doi:10.1073/pnas.1704008114
64. Kaur S, Bansal Y, Kumar R, et al. A panoramic review of IL-6: structure, pathophysiological roles and inhibitors. Bioorg Med Chem. 2020;28(5):115327. doi:10.1016/j.bmc.2020.115327
65. Menten P, Wuyts A, Van Damme J. Macrophage inflammatory protein-1. Cytokine Growth Factor Rev. 2002;13(6):455–481. doi:10.1016/S1359-6101(02)00045-X
66. Ramirez-Carrozzi V, Sambandam A, Luis E, et al. IL-17C regulates the innate immune function of epithelial cells in an autocrine manner. Nat Immunol. 2011;12(12):1159–1166. doi:10.1038/ni.2156
67. Park SJ, Kim YY, Park W, et al. Effects of cetrorelix on ovary and endometrium prior to anti-PD-L1 antibody in murine model. Tissue Eng and Regener Med. 2024;21(2):319–328. doi:10.1007/s13770-023-00617-x
68. Nilsson E, Dole G, Skinner MK. Neurotrophin NT3 promotes ovarian primordial to primary follicle transition. Reproduction. 2009;138(4):697–707. doi:10.1530/REP-09-0179
69. Jankowska K. Premature ovarian failure. Przeglad Menopauzalny. 2017;16(2):51–56. doi:10.5114/pm.2017.68592
70. Tang M, Sun X, Li P, et al. IL-33 and soluble ST2 in follicular fluid are associated with premature ovarian insufficiency. Front Endocrinol. 2024;15:1463371. doi:10.3389/fendo.2024.1463371
71. Miller JE, Monsanto SP, Ahn SH, et al. Interleukin-33 modulates inflammation in endometriosis. Sci Rep. 2017;7(1):17903. doi:10.1038/s41598-017-18224-x
72. Lin T-C, Wang K-H, Chuang K-H, et al. Interleukin-33 promotes invasiveness of human ovarian endometriotic stromal cells through the ST2/MAPK/MMP-9 pathway activated by 17β-estradiol. Taiwanese J Obstetrics Gynecol. 2021;60(4):658–664. doi:10.1016/j.tjog.2021.05.013
73. Chang H-M, Wu H-C, Sun Z-G, et al. Neurotrophins and glial cell line-derived neurotrophic factor in the ovary: physiological and pathophysiological implications. Human Reprod Update. 2019;25(2):224–242. doi:10.1093/humupd/dmy047
74. Roberts RD, Ellis RC. Mitogenic effects of fibroblast growth factors on chicken granulosa and theca cells in vitro. Biology Reprod. 1999;61(6):1387–1392. doi:10.1095/biolreprod61.6.1387
75. Fang X, Xia W, Li S, et al. SIRT2 is critical for sheep oocyte maturation through regulating function of surrounding granulosa cells. Int J Mol Sci. 2022;23(9):5013. doi:10.3390/ijms23095013
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.