Back to Journals » International Journal of Nanomedicine » Volume 21
Recent Advances in Nanodelivery Systems Based on Extracellular and Intracellular Reprogramming Strategies for Enhanced Therapy of Atherosclerosis
Authors Liu Y, Li Y, Wang Q, Wu M ![]() , Zhang Z, Liang X, Yang X, He X, Tang J, Li C, Yuan P, Li T
, Zhang Z, Liang X, Yang X, He X, Tang J, Li C, Yuan P, Li T
Received 10 June 2025
Accepted for publication 6 January 2026
Published 7 February 2026 Volume 2026:21 541504
DOI https://doi.org/10.2147/IJN.S541504
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Dr Yan Shen
Yilin Liu,1,* Yao Li,1,2,* Qian Wang,3,* Mengxi Wu,1,4 Zongquan Zhang,1 Xiaoya Liang,1 Xi Yang,1 Xinghui He,1 Jun Tang,5 Chunhong Li,1 Ping Yuan,6 Tao Li7
1Department of Pharmacy Sciences, School of Pharmacy, Southwest Medical University, Luzhou, Sichuan, 646000, People’s Republic of China; 2Science and Technology Department, Southwest Medical University, Luzhou, Sichuan, 646000, People’s Republic of China; 3Classical Teaching and Research Department, College of Integrated Chinese and Western Medicine, the Affiliated Traditional Chinese Medicine Hospital of Southwest Medical University, Luzhou, 646000, People’s Republic of China; 4Chinese Pharmacy Laboratory, School of Pharmacy, Southwest Medical University, Luzhou, Sichuan, 646000, People’s Republic of China; 5Analysis and Testing Center, School of Pharmacy, Southwest Medical University, Luzhou, Sichuan, 646000, People’s Republic of China; 6Department of Neurology, the Affiliated Hospital of Southwest Medical University, Luzhou, Sichuan, 646000, People’s Republic of China; 7Key Laboratory of Medical Electrophysiology of Ministry of Education, Institute of Cardiovascular Research, Southwest Medical University, Luzhou, 646000, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Ping Yuan, Email [email protected] Tao Li, Email [email protected]
Abstract: Atherosclerosis (AS) represents a chronic immunoinflammatory disorder characterized by lipid metabolism dysregulation, predominantly affecting medium and large arteries. During the initial phase of the disease, endothelial cells (ECs) undergo mesenchymal transformation, subsequently releasing various adhesion molecules and chemokines that facilitate the recruitment of monocytes and neutrophils (NEs) to developing plaque sites. Under sustained inflammatory stimulation, NEs transform into neutrophil extracellular traps (NETs), monocytes differentiate into pro-inflammatory macrophages, and vascular smooth muscle cells (VSMCs) exhibit phenotypic switching, and adopt macrophage-like characteristics. These inflammatory cells release excessive inflammatory factors and reactive oxygen species (ROS), thereby amplifying the inflammatory cascade. Concurrently, these activated macrophages and VSMCs internalize oxidized low-density lipoprotein (ox-LDL) particles, promoting foam cell’s generation and plaque’s formation. Given the critical role of the activation of various inflammatory cells and the accumulation of inflammatory mediators in progression of the disease, nanodelivery systems (NDSs) have shown remarkable promise in treating AS, attributed to their excellent cell-specific targeting capabilities and regulation capabilities for inflammatory mediator. This paper comprehensively reviews the progress of NDSs based on intracellular and extracellular reprogramming strategies for the therapy of AS, emphasizing their role in reversing EC’s phenotypes, monocytes’ reprogramming, macrophages’ repolarization, inhibition of Nes’ recruitment, suppression of VSMCs’ proliferation, modulation of inflammatory mediators, reprogramming of lipid’s metabolism, and scavenging of ROS to enhance therapeutic efficacy. We further elaborate on the advantages, challenges, and opportunities of NDSs in enhancing intracellular and extracellular reprogramming strategies, aiming to deepen these strategies and their potential applications in the therapy of AS.
Keywords: atherosclerosis, nanodelivery systems, microenvironment, reprogramming strategies
Introduction
Atherosclerosis (AS), a chronic inflammatory process affecting arterial vessels, serves as the principal driver of diverse cardiovascular pathologies,1 characterized by the accumulation of lipid plaques on the arterial wall and accompanying endothelial dysfunction.2 Due to its elevated morbidity and mortality rates, AS imposes a significant burden on both individuals and society, underscoring the critical need for effective therapeutic interventions.
In clinical practice, the primary therapeutic strategies for AS remain surgical plaque removal, implantation of stents, and pharmacological control.3 Statins remain the first-line pharmacological intervention, demonstrating efficacy in reducing low-density lipoprotein (LDL) through competitively inhibiting HMG-CoA reductase this intervention effectively suppresses de novo cholesterol biosynthesis.4,5 Nevertheless, substantial therapeutic limitations persist, primarily attributable to complex pathophysiological disturbances across multiple cellular and molecular pathways that drive its pathogenesis. These abnormalities include the infiltration of excessive neutrophils (NEs),6 phenotypic switching of macrophages,7 the dysfunction of endothelial cells (ECs),8 dysregulated cellular expansion of vascular smooth muscle cells (VSMCs),9 the accumulation of lipid, and the overproduction of reactive oxygen species (ROS).10–12 These dysregulated processes synergistically drive the development of a pathologically sophisticated atherosclerotic microenvironment (ASM) (Figure 1). Consequently, reprogramming strategies for the ASM including remodeling cellular phenotypes, inhibiting the proliferation of pathological cells, modulating inflammatory responses, and clearing lipids and ROS holds substantial therapeutic promise for the disease.
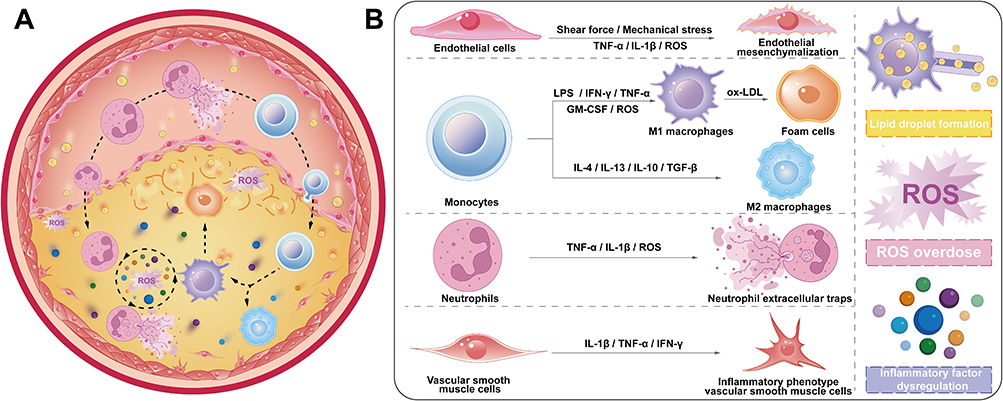 |
Figure 1 The complex inflammatory microenvironment of atherosclerosis. (A) Multiple cell types undergo changes during the atherosclerotic process, involving a vicious cycle driven by inflammatory factors, lipids, and ROS. (B) The inflammatory vicious cycle in ASM specifically includes: interstitialization of ECs, polarization of macrophages, transformation of NEs into NETs, phenotypic transformation of VSMCs, formation of lipid droplet, excessive ROS, and disruption of inflammatory factors. |
Endothelial dysfunction, as a key driver of AS, is mainly attributed to the transformation of endothelial cells (ECs) into inflammatory endothelial cells and mesenchymal endothelial cells, which jointly promote the subsequent recruitment of large numbers of neutrophils (NEs) and the generation of neutrophil extracellular traps (NETs). The latter further exacerbates endothelial dysfunction, forming a self-perpetuating vicious cycle.13,14 Consequently, reprogramming the endothelial cell phenotype or reversing the inflammatory state of endothelial cells represents a promising strategy for restoring endothelial barrier function.15 The overexpression of ICAM-1 on ECs, as well as p-selectin on NEs,16 enables their targeted interaction with ligand-modified nanodelivery systems (NDSs), allowing for specific therapeutic actions on these cells. Beyond neutrophil’s recruitment, significant monocyte’s infiltration occurs at inflammatory sites, followed by their polarization into pro-inflammatory M1 macrophages. These macrophages release substantial quantities of inflammatory factors and ROS. Furthermore, M1 Macrophage engulfment of oxidized low-density lipoprotein (ox-LDL) and conversion to foam cells, thereby promoting plaque formation.17 Accordingly, reprogramming monocytes or converting the M1 macrophage phenotype to the M2 is a promising strategy. At plaques site, inflammatory macrophages overexpress scavenger receptors (eg, SR-A, CD36), CD44, and other receptors on their surfaces.18,19 Nanoparticles (NPs), through ligand modifications such as hyaluronic acid (HA) and folic acid, can enhance specifically targeting ability to macrophages.20,21 Notably, VSMCs demonstrate proliferation and migration patterns resembling tumor cells, while the phagocytic uptake of lipids facilitates their transformation into foam cells, exacerbate endothelial damage and plaque’s formation.9 Thus, inhibiting abnormal proliferation of VSMCs or inducing their apoptosis is crucial for enhancing therapeutic efficacy. Previous studies have demonstrated that various nanomaterials, including iron oxide nanoparticles (Fe3O4 NPs),22 silver nanoparticles (Ag NPs),23 etc, exhibit excellent capabilities in inducing apoptosis. These nanomaterials with appropriate modifying, can specifically target pathological cells such as inflammatory macrophages,24 and VSMCs25 at lesion sites, thereby inducing apoptosis and improving treatment of the disease. To sum up, inflammation within the ASM leads to morphological and functional alterations in key cells. Leveraging the precise targeting ability of NDSs could achieve better reprogramming strategies of ASM (Table 1).
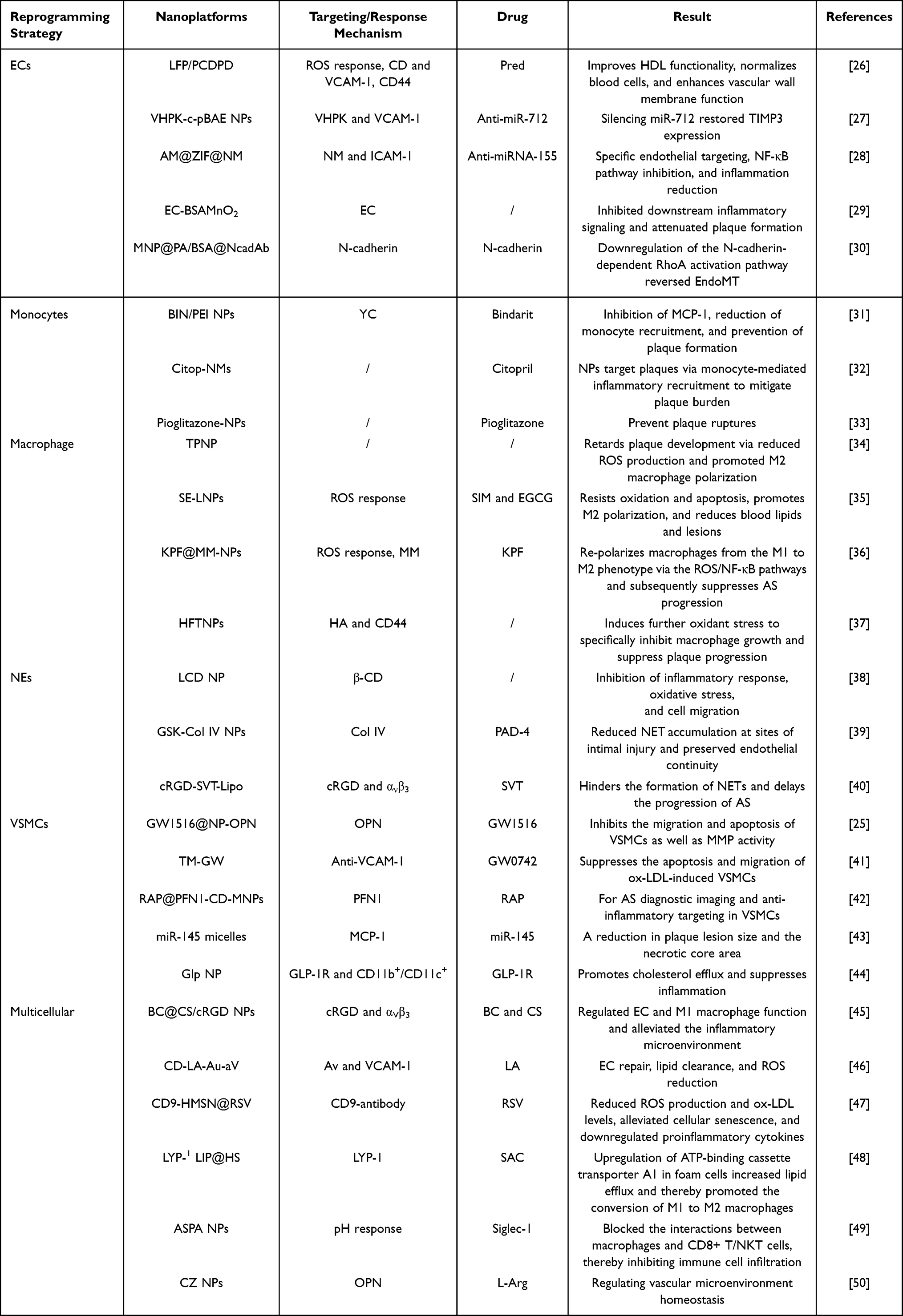 |
Table 1 Nanodelivery Systems Based on Intracellular Reprogramming Strategies for Enhancing the Therapy of Atherosclerosis |
Meanwhile, M1 macrophages and NEs release substantial amounts of inflammatory factors, which exacerbate the inflammatory response in the disease.51 Anti-inflammatory drugs, such as colchicine and inflammatory pathway inhibitors, can significantly suppress the expression of inflammatory factors. However, their clinical application is limited by the non-targeting biodistribution and systemic side effects.52–54 Various optimized NDSs can target transport of anti-inflammatory drugs to the plaque site, thereby effectively modulating the inflammatory response. In addition, various inflammatory cells at lesion sites release considerable ROS and oxidize LDL to ox-LDL, which further contributes to endothelial damage, as well as the formation of foam cells.55,56 Although natural enzymes such as superoxide dismutase (SOD) and catalase (CAT) can scavenge ROS at lesion sites, their poor stability and susceptibility to degradation have severely restricted their therapeutic potential.11,57 Metal-organic framework (MOF),58 silicon dioxide (SiO2),59 Prussian blue (PB) NPs, selenium (Se) NPs, and cerium oxide (CeO2) have enzyme-like activity and demonstrate excellent ROS-clearing ability, bridging the gap of NDSs-based efficient reprogramming strategies of ROS.60,61 More importantly, reprogramming lipid’s metabolism is of great research value for the treatment of AS, as increased dyslipidemia leads to the passage of lipids across the damaged endothelial barrier, and their uptake by inflammatory macrophages, driving the development of the disease. Statins could effectively inhibit cholesterol synthesis, nevertheless, the significant oral first-pass effect and high plasma protein binding rate limit their application.62,63 Optimized NDSs enable lipid’s reprogramming strategies by targeting the delivery of statins to the lesion site. In addition, soybean polyunsaturated phosphatidylcholine (PL) NPs and NPs modified with liver X receptor (LXR) ligands can reprogram lipid metabolism by regulating the expression of ATP-binding cassette transporters ABCA1 and ABCG1 or by improving the lipid composition of high-density lipoprotein (HDL) to enhance the capacity of cholesterol efflux (CEC).64–67 To sum up, NDSs significantly enhance extracellular-based reprogramming strategies for the therapy of AS through their anti-inflammatory properties, ROS-scavenging capabilities, and superior lipid-clearing potential (Table 2).
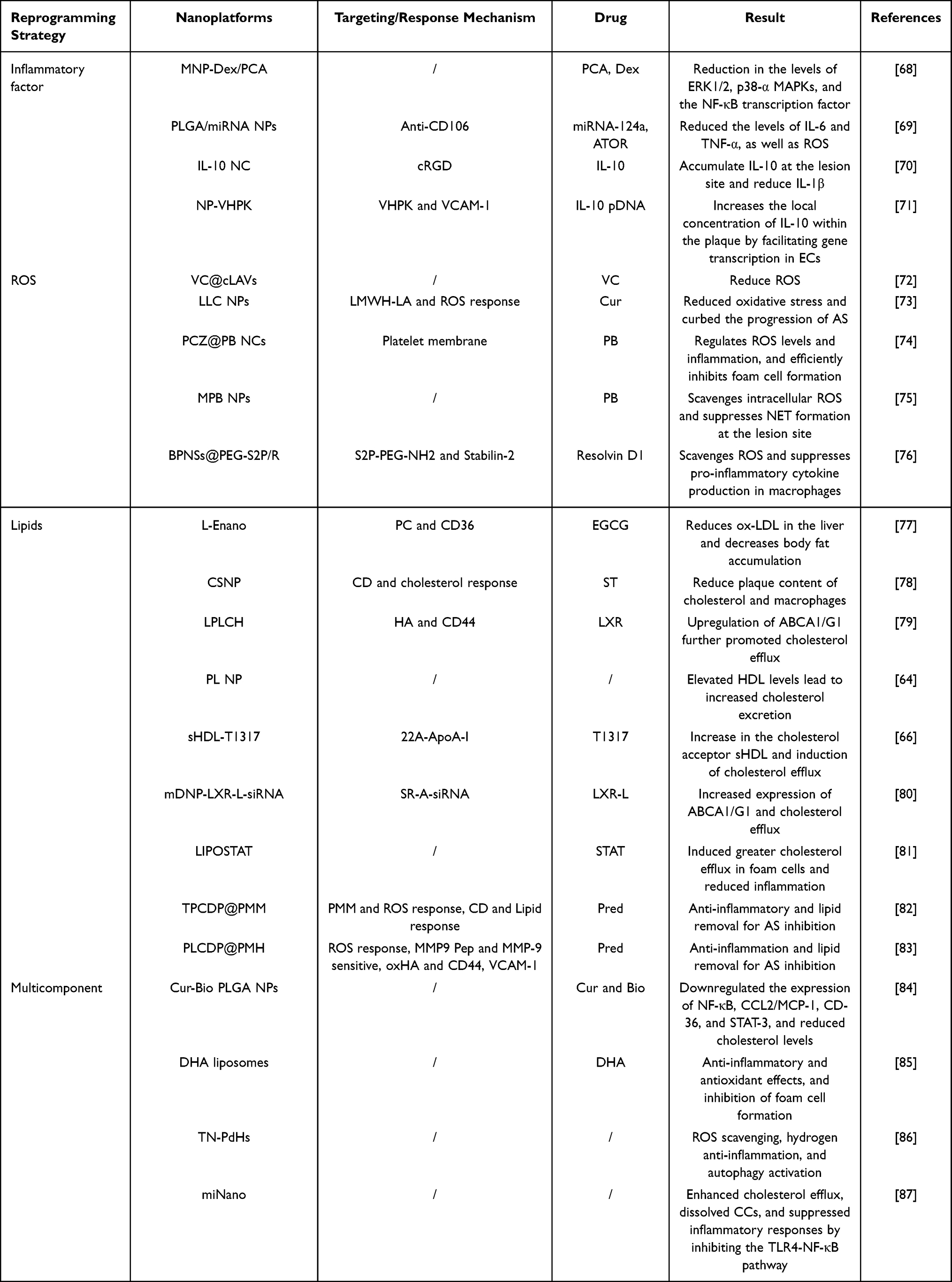 |
Table 2 Nanodelivery Systems Based on Extracellular Reprogramming Strategies for Enhancing the Therapy of Atherosclerosis |
This paper provides the first comprehensive review of progress over the past five years on NDSs based on intracellular and extracellular reprogramming strategies to improve the treatment efficacy of AS. It delves into intracellular reprogramming strategies, including reversing the phenotype of ECs and inflammatory states, the reprogramming of monocyte, the repolarization of macrophage, inhibiting the recruitment of NEs, and suppressing the proliferation of VSMCs, as well as extracellular reprogramming strategies, such as regulating inflammatory factors, reprogramming lipid’s metabolism, and scavenging ROS. The review covers multiple aspects, including cellular phenotypes and molecular pathways, emphasizing the advantages of these strategies over traditional delivery systems in the treatment of AS. This review provides new insights and perspectives on improving ASM and advancing its treatment through intracellular and extracellular reprogramming strategies.
Intracellular-Based Reprogramming Strategies
ECs-Based Reprogramming Strategies
Within the ASM, vascular shear and mechanical enlargement directly contribute to ECs damage.88,89 Moreover, elevated levels of ox-LDL, ROS, and inflammatory factors at plaque sites contribute to mesenchymal transition of ECs.90 NDSs have demonstrated the ability to reprogram the cells by reducing ROS production, enhancing endogenous antioxidant enzyme defenses, clearing lipids, mitigating shear stress, and increasing endothelial nitric oxide synthase (eNOS) activity. These mechanisms collectively inhibit or reverse endothelial mesenchymalization and restore endothelial barrier integrity.
Vascular cell adhesion molecule 1 (VCAM-1) is overexpressed on ECs,91 and it offers a wide range of perspectives for enhancing the targeting function of NDSs to ECs. Here, Xu et al prepared a ROS-responsive multifunctional NDSs (LFP/PCDPD) based on cyclodextrin loaded with a lipid-specific AIE fluorescent probe (LFP) and anti-inflammatory drug prednisolone (Pred) in its hydrophobic lumen, connected the hydrophilic and hydrophobic portions via poly(2-(methylthio) ethyl methacrylate (PMEMA), and modified the surface with dextran.26 The NDSs effectively targeted ECs via VCAM-1 and CD44 receptors, and subsequently achieved ECs reprogramming strategy by scavenging ROS and lipids to effectively inhibit endothelial injury. Similarly, Dosta et al prepared a novel NDSs (VHPK-c-pBAE) by using targeted poly (β-amino ester) pBAE NPs, loaded with anti-microrna-712 (anti-miR-712) antibody and modified by VHPK peptide that could specifically target to ECs.27 The NDSs selectively bound VCAM-1 on ECs, downregulating miR-712 while preserving tissue inhibitor of metalloproteinase 3 expression in inflamed endothelium. This dual action improved ECs function and therapeutic efficacy.
More importantly, by taking advantage of the recruiting effect of VCAM-1 overexpressed on the surface of ECs to NEs, the membrane of NEs could be used as a targeting material for ECs. Herein, Liu et al prepared a biomimetic NDSs (AM@ZIF@NM) using NEs membrane-coated zeolite imidazoline framework 8 (ZIF-8) to deliver microRNA-155 (anti-miRNA-155) antisense oligonucleotides (ASO).28 The formulation could be recruited by VCAM-1 overexpressed on the surface of ECs, thereby specifically targeted the cells. Furthermore, it down-regulated the effects of miR-155 and the expression of its downstream target genes, C-C motif chemokine ligand (CCL2) and ICAM-1, effectively alleviating disease. It was shown that biomimetic NPs prepared through the plasma membrane of ECs had excellent targeting effects. Here, GAO et al designed a high-performance integrin-activating biomimetic (EC-BSAMnO2) featuring reductively cleavable MnO2, and core was fully encapsulated within ECs plasma membranes for enhanced biocompatibility.29 The NPs effectively inhibited endothelial injury by reacting with considerable metabolites (H2O2 and H+) in the plaque and decomposing in situ to Mn2+, thereby activating integrin β3 on the surface of ECs and inhibiting downstream integrin-coupled inflammatory signaling pathway.
Endothelial-mesenchymal transition (EndoMT) is a newly recognized process of cellular transdifferentiation in which ECs loosen their intercellular junctions, and endothelial junctions are reduced, while permeability is increased.92 Growing evidence supports EndoMT inhibition as a valuable therapeutic paradigm in inflammatory disease treatment. Here, Liu et al prepared a multifunctional NDSs (MNP@PA/BSA@NcadAb) using an N-cadherin antibody to modify melanin NPs.30 By specifically targeting N-cadherin overexpressed on the surface, the NDSs modulated the downstream signaling of N-cadherin to reverse their EndoMT. Moreover, senescent ECs disrupt vascular endothelial integrity, accelerating vascular aging and increasing susceptibility to AS. Here, Xiao et al investigated that mesenchymal stem cell (MSCs)-derived extracellular vesicles (sEVs) enhanced AS therapeutic efficacy by preventing and attenuating senescence of ECs.93 Results indicated MSCs-sEVs effectively suppressed senescent markers components while reversing oxidative damage, functional deficits and migratory capacity in aging ECs. It achieved reprogramming effects on senescent ECs, and stimulated angiogenesis via miR-146a/Src. MSCs-sEVs was expected to be used as an overlay material for enhancing the therapeutic effects for AS.
Monocyte-Based Reprogramming Strategies
The inflammatory properties of AS accelerate the production of bone marrow monocytes, and there is extravasation of circulating monocytes to sites of endovascular inflammation. The EndoMT leads to overexpression of P-selectin and E-selectin on the cell membrane, together with the release of adhesion factors, which recruit monocytes to accumulate at the AS lesion plaque.94 Subsequently, Exposure to ox-LDL, ROS, and pro-inflammatory cytokines drives monocyte differentiation into inflammatory macrophages, amplifying the inflammatory cascade through positive feedback mechanisms.95 In addition, anti-inflammatory cytokine-driven polarization of M2 macrophages promotes plaque microenvironment remodeling through resolution of inflammation and tissue repair mechanisms. Accordingly, targeted monocyte reprogramming emerges as a viable therapeutic paradigm for AS intervention.
Harnessing the inherent inflammatory chemotactic properties of monocytes for AS plaque targeting is a precise delivery strategy based on physiological mechanisms. Here, Yi et al prepared a positively charged NPs (BIN/PEI NPs) by self-assembly of MCP-1 synthesis inhibitor (bindarit) with polyethyleneimine, and further filled the BIN/PEI NPs into yeast-derived microcapsules (YCs) to obtain a biomimetic orally consumable NDSs (BIN/YCs).31 Upon oral administration of the NPs, they were distributed into the peripheral blood monocytes and transported to plaques via monocytes, thereby inhibiting release MCP-1, in turn inhibiting monocyte aggregation and chemotaxis, downregulating the inflammatory cascade response, and effectively preventing plaque formation. Furthermore, the monocyte-mediated delivery approach capitalizes on the marked increase in ROS that occurs during monocyte-to-macrophage differentiation within AS lesions, herein, Fu et al engineered citopril, an ROS-responsive prodrug, through thioketal-mediated conjugation of citral and captopril. This compound was subsequently self-assembled with amphiphilic polymers (PEG-DSPP) into nanomicelles (Citop-NMs).32 After intravenous injection, it was internalized into circulating monocytes and effectively accumulated within the plaque through inflammatory recruitment properties. Elevated levels of intracellular ROS triggered the pharmacological activation of Citopril, effectively reducing NLRP3-dependent IL-1β secretion, modulating the inflammatory response and preventing lipid deposition in plaques, and reducing the proportion of foam cells. Beyond suppressing monocyte recruitment, redirecting monocyte differentiation toward an M2 macrophage phenotype represents a promising therapeutic approach to stabilize vulnerable plaques. Nakashiro et al engineered pioglitazone-loaded nanoparticles (pioglitazone-NPs) using biodegradable poly (lactic-co-glycolic acid) (PLGA) polymers.33 The NPs enhanced therapeutic outcomes by downregulating inducers of extracellular matrix metalloproteinases (MMP), reducing pro-inflammatory cytokine production in bone marrow-derived monocytes, and promoting their polarization into anti-inflammatory M2 macrophages.
Macrophage-Based Reprogramming Strategies
Macrophages serve as key immunomodulatory cells that critically orchestrate atherosclerotic plaque progression through their phenotypic plasticity and inflammatory mediator secretion.96 M1 macrophages drive inflammatory pathogenesis through robust production of ROS and pro-inflammatory cytokines, while their M2 counterparts promote tissue homeostasis via anti-inflammatory mediators and reparative functions.97 In early lesions, inflammatory macrophages have ability to remove apoptotic cells to avoid disease progression. During lesion progression, intimal macrophages actively engulf ox-LDL via scavenger receptors, transforming into lipid-laden foam cells that ultimately contribute to necrotic core formation through apoptotic cell death and defective efferocytosis.98 Consequently, remodeling the inflammatory macrophage phenotype or induce apoptosis using NDSs is expected to improve the efficacy of AS.
Polarizing M1 macrophages into M2 macrophages has the potential to reprogram the pro-inflammatory into an anti-inflammatory response, thereby improving the inflammatory microenvironment.36,99 Studies demonstrate that combined ROS elimination and pro-inflammatory factor reduction reliably reprograms macrophages toward tissue-repairing M2 polarization.100 Here, Wu et al prepared tannic acid (TA)-poloxamer NPs (TPNP) by self-assembly of TA and poloxamer to achieve macrophage reprogramming to M2 type.34 It has been demonstrated that epigallocatechin gallate (EGCG) can inhibit inflammatory factors, NF-κB inflammatory pathway and glycolysis by scavenging ROS. Wan et al developed a co-delivery system comprising simvastatin and EGCG using liposomal NPs (SE-LNPs) prepared from distearoylphosphatidylcholine (DSPC).35 The NPs were observed to reprogram M1 macrophages into M2 macrophages through superior ROS scavenging and anti-inflammatory capabilities. To optimize NDSs accumulation at inflammatory sites and enhance M1 macrophage specificity, Zhao et al devised a ROS-responsive bionic NDSs (KPF@MM-NPs) by enclosing dextran-g-PBMEO NPs loaded with the natural drug kaempferol (KPF) through macrophage membranes (MM).36 The ROS-responsive NDSs demonstrated targeted plaque accumulation through oxidative stress-dependent activation, enabling selective macrophage delivery in lesions. This resulted in a reduction in key pro-inflammatory factors through regulate of the NF-κB pathway, thereby enabling macrophage reprogramming strategies.
The process of endocytosis, proliferation, accumulation, and absorb of ox-LDL by inflammatory macrophages at the lesion site exacerbates plaque development. Triggering oxidative stress, mitochondrial damage, and DNA destruction through the Fenton reaction, and ultimately activating the macrophage apoptotic pathway, can effectively arrest disease progression. Based on this, Mu et al used dopamine-modified hyaluronic acid (HD) loaded with stabilized iron (III)-TA NPs (HFTNPs) to specifically induce apoptosis in inflammatory macrophages.37 The NDSs demonstrated selective targeting of inflammatory macrophages via HA-CD44 receptor interactions. This targeting strategy simultaneously elevated intracellular ROS levels, amplified iron-mediated Fenton reactions, and triggered significant apoptosis in pro-inflammatory macrophages. Consequently, this triple-action mechanism effectively suppressed inflammatory cytokine secretion and attenuated atherosclerotic plaque development.
NEs-Based Reprogramming Strategies
In the ASM, NEs undergo activation via ROS and pro-inflammatory cytokines, and their activation synergistically interacts with the calcium-dependent enzyme peptidyl arginine deiminase 4 (PAD-4) through histone citrullination. This interaction triggers cellular disintegration, resulting in the cellular contents release of cytoplasmic component,101,102 and this cascade culminates in NETs formation, which accelerate plaque expansion.103 Concurrently, NETs-derived histones and granular proteins directly induce VSMCs and ECs lysis, while neutrophil elastase-mediated extracellular matrix degradation synergistically compromises plaque structural integrity.104 These findings position NDS-mediated NEs reprogramming strategies as a novel approach to address both ECs dysfunction and plaque vulnerability in AS, potentially overcoming current treatment limitations.
Due to recruitment of NEs is a typical feature of AS, inhibition of NEs recruitment by targeting NDSs is expected to block subsequent NETs-mediated endothelial damage and inflammatory responses. Here, Guo et al constructed a cyclodextrin-derived intrinsically bioactive NPs (LCD NPs),38 which enhanced the therapeutic efficacy by inhibiting the recruitment of NEs and inflammatory macrophages, thereby preventing subsequent pro-inflammatory events. Moreover, the areas of the basement membrane that have undergone ECs detachment are characterized by a high concentration of type IV collagen. Here, Molinaro et al constructed targeted NDSs (GSK-Col IV NPs) that limit NET formation by utilizing poly(D,L-propenolactone) (PLA) as the central part and polyethylene glycolated phospholipid layer as the shell of type IV collagen-targeted NPs (Col-IV NPs) for the delivery of the PAD-4 enzyme inhibitor GSK484.39 The NDSs effectively inhibited NEs infiltration and suppressed NETs-driven inflammatory response. Additionally, they reduced thrombus formation induced by NETs-associated superficial erosion while promoting endothelial repair. Notably, NEs elastase that catalyzes the formation of NETs, enhances immune cell recruitment, stimulates interferon-α release. More importantly, it degrades elastin and other matrix proteins in motile AS plaques, thereby accelerating the formation of vulnerable plaques.105,106 Here, Yin et al constructed a plaque coupled targeting and NEs hitchhiking liposomes (cRGD-SVT-Lipo) by loading the NEs elastase-competitive inhibitor sivastigmine (SVT) into cRGD peptide-modified liposomes.40 The NDSs used cRGD peptide to specifically recognize with integrin ανβ3 on the surface of NEs and being internalized into them. After reaching the plaque site with the inflammatory chemotactic effect of NEs, they effectively inhibited the elastase activity of intra- and extracellular NEs, reduced NETs formation, and ultimately alleviated the development of AS.
VSMCs-Based Reprogramming Strategies
VSMCs undergo significant phenotypic changes switching from contractile to synthetic phenotype in the inflammatory environment of AS, and the phenotype is distinguished by strengthened migration, multiplication, and secretion.9,107 This is demonstrated by VSMCs migrating from the arterial wall’s medial layer, proliferating to form a fibrous protective cap abundant in extracellular matrix. This encapsulation process surrounds the plaque while simultaneously promoting expansion of the necrotic core.108 Secondly, under the stimulation of ROS and inflammatory factors, VSMCs can be transformed into transformed into macrophage-like cells, which further release pro-inflammatory factors, extracellular matrix, and adhesion molecules, thereby recruiting a large number of inflammatory cells to accumulate at plaques, exacerbating the development of inflammation, and inducing plaque instability and calcification.109,110 Consequently, reprogramming of the VSMCs phenotype holds promising future for optimizing the therapeutic effectiveness of AS.
VSMCs exhibit remarkable plasticity, transitioning from a quiescent, contractile state to a migratory, proliferative, and synthetic phenotype. It has been demonstrated that the peroxisome proliferators-activated receptors (PPARδ) agonist GW1516 inhibits VSMCs migration and apoptosis. Here, Huang et al developed a GW1516-loaded NDSs (GW1516@NP-OPN) modifying with anti osteopontin (OPN),25 and they effectively suppressed VSMC migration and apoptosis, resulting in a significant reduction of lesion size in vivo.111 Similarly, Gang et al delivered the PPARδ agonist GW0742 using anti-VCAM-1-modified nanocolloid,41 which effectively inhibited ox-LDL-induced apoptosis and migration of VSMCs. Furthermore, Profilin-1 (PFN1) is plentiful in VSMCs and is involved the reorganization of cytoskeletal polymerization.112,113 Here, Zhang et al prepared a VSMCs-targeting NDSs (RAP@PFN1-CD-MNPs) by conjugating paramagnetic Fe3O4 NPs with pH-sensitive cyclodextrins (CDs) and coupled them with an antibody to PFN1 loaded with the anti-inflammatory drug (rapamycin) (Figure 2).42 The NDSs utilized the pH sensitivity of cyclodextrins to achieve responsive release in the plaque region and successfully inhibited the proliferation of vascular ECs. Moreover, RAP@PFN1-CD-MNPs could function as a biocompatible MRI contrast agent for AS lesion detection.
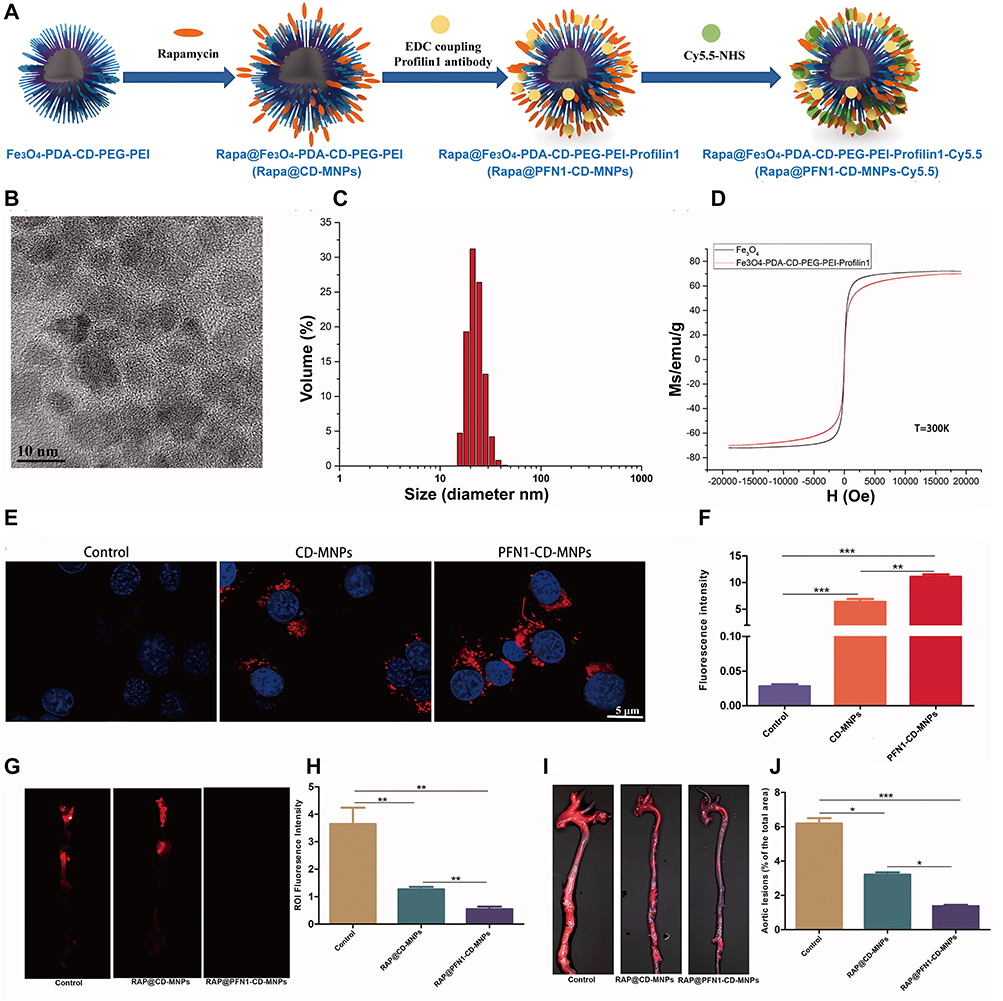 |
Figure 2 Preparation of RAP@PFN1-CD-MNPs and its mechanism of enhancing the therapy of AS by reprogramming strategy of ECs (A) Schematic diagram of synthesis principle of [email protected] NPs. (B) TEM images of NPs (scale bar = 10 nm). (C) Magnetic properties of NPs. (D) FTIR spectra of NPs. Uptake of NPs in VSMCs. Fluorescence imaging (E) and quantification (scale bar = 5 μm) (F) of the ability of PFN1-CD-MNPs to bind to VSMCs in vitro. NIRF of aorta at 2 months after treatment (G) and quantification (H). General ORO staining (I) and quantification (J). Animals in the control group were administered saline, *P < 0.05, **P < 0.01, ***P < 0.001. Reproducted from Zhang S, Xu W, Gao P, Chen W, Zhou Q. Construction of dual nanomedicines for the imaging and alleviation of atherosclerosis. Artif Cells Nanomed Biotechnol. Dec 2020;48(1):169–179.42 © 2019 The Author(s). Published by Informa UK Limited, trading as Taylor & Francis Group. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0/). |
Given the phenotypic transformation of VSMCs and its high degree of plasticity, the use of targeted NDSs to modulate its phenotype or intervene in its differentiation is expected to attenuate plaque development. Herein, Chen et al used MCP-1 peptide-modified nucleic acid micelles (miR-145 micelles) to target and meddle in the phenotypic transition of VSMCs.43 This micelle specifically binds to C-C chemokine receptor-2, which was upregulated in VSMCs surface, and enhances the expression of cardiac myosin, calponin, and α-smooth muscle actin by upregulating contractile genes. At the same time, Krüppel-like factor-4/5 and ETS domain-containing protein-1 were downregulated, thereby inhibiting VSMCs-derived plaque propagating cell types, maintaining the normal contractile state of VSMCs, and attenuating the development of AS. Recent studies have shown that an extensive majority of macrophages and foam cells in AS stem from VSMCs differentiation, which is positively correlated with markers of dedifferentiation and Glp1r expression (eg, increased Klf4).114 Maiseyeu et al prepared a theranostics NPs (GLP NP) by utilizing synthesized protease-resistant and “activatable” NPs conveying the GLP-1R agonist liraglutide (GlpNP) and incorporated gadolinium chelates (MRI imaging agents).44 The NPs were capable of effectively preventing the transition of VSMCs to macrophage-like phenotype.
Multiple Cells-Based Reprogramming Strategies
AS progression involves a sophisticated interplay of diverse cell populations and complex signaling networks.115,116 Under inflammatory conditions, elevated levels of ROS, inflammatory factors, and injury signals induce ECs damage. This damage triggers the secretion of substantial amounts of adhesion factors, which recruit large numbers of monocytes and NEs. Monocytes differentiate into a significant population of inflammatory macrophages, while NEs are stimulated to form NETs. These NETs not only aggravate endothelial dysfunction but also potentiate inflammatory cascades, thereby accelerating AS progression.117–120 In summary, in the ASM, various cells interact through complex cross-talk mechanisms, forming a network that collectively drives inflammation and plaque progression. Consequently, reprogramming cells and the associated network of contacts in ASM has significant implications for AS therapy.
In the initial phase of AS, ECs injury triggers the recruitment of pro-inflammatory macrophages that release abundant cytokines, creating a vicious cycle of endothelial dysfunction. Therefore, simultaneous targeting and reprogramming of ECs and macrophages can achieve suppression of inflammation while repairing endothelial damage. Here, our group prepared a pH-responsive dual-targeting NDDS (BC@CS/cRGD NPs) by combining baicalin (BC) with chondroitin sulphate (CS) through an amidation reaction followed by conjugation with the cRGD peptide targeting moiety.45 This NDSs could effectively target ECs and inflammatory macrophages through the specific binding of the cRGD peptide to the αVβ3 protein receptor, thereby effectively inhibiting inflammation and repairing endothelial damage. In addition, removal of large numbers of inflammatory macrophages from AS plaques, inhibition of foam cell formation, and repair of ECs are important for the treatment of the disease. Herein, Li et al constructed a bimodal propellant NDSs CD-LA-Au-aV sensitive to ROS and near-infrared lasers by covalent binding and self-assembly of β-cyclodextrin (β-CD) with L-arginine (L-Arg), followed by the immobilization of gold NPs.46 The NDSs could remove inflammatory macrophages through the excellent photothermal conversion properties of Au, thereby inhibiting inflammation along with foam cell formation, while promoting cholesterol removal from foam cells and reducing ROS to achieve endothelial repair. To sum up, the NDSs could reprogram multiple cells and components of ASM in multiple ways, thereby synergistically and comprehensively enhancing the therapeutic efficacy in a non-pharmacological strategy.
Macrophages and foam cells play pivotal roles in AS plaque formation and the associated inflammatory cascade. Therapeutic strategies that simultaneously modulate macrophage polarization and enhance foam cells CEC may provide a comprehensive approach for mitigating AS progression. An et al developed an endogenous H2S-based gas therapy utilizing multienzyme-mimicking nanozymes (LYP−1 LIP@HS) using hollow mesoporous prussian blue (HMPB) loaded with the H2S donor S-allyl-L-cysteine (SAC), and to enhance the targeting specificity and achieve controlled release kinetics of the HS NP, further the LYP-1 peptide-functionalized lipid membrane was modified on its surface.48 The NDSs unlocked the polymerase-like activity of HMPB after cavitation induced by low-intensity focused ultrasound (LIFU), while promoting the release of the endogenous H2S donor SAC. Notably, the enzyme-catalysed generation of endogenous H2S plays multiple roles by upregulating ATP-binding cassette transporter A1 in foam cells, enhancing cholesterol efflux, and facilitating pro-inflammatory M1 polarization to anti-inflammatory M2 phenotype, thereby effectively delaying disease. In addition, the crosstalk between T cells and macrophages constitutes a pivotal regulatory axis driving inflammatory pathogenesis in this disease, and inhibition of antigen presentation from macrophages to T cells not only modulates the number and type of T cell subsets, but also the levels of inflammatory factors in ASM, which in turn remodels the macrophage phenotype. Here, Zhou et al designed a pH-responsive and charge reversible NPs (ASPA NPs) through synthesizing short hairpin RNA targeting mouse sialic-acid-binding immunoglobulin-like lectin-1 (Siglec-1) loaded and polyethyleneimine-acetylsalicylic acid (PEI-ASA) coated Au-PEI NPs (Figure 3).49 The NPs blocked CD8+T/NKT cell-macrophage interactions by silencing Siglec-1, thereby inhibiting CD8+T/NKT cell infiltration and inhibited Th 17 cell differentiation, as well as activating intracellular macrophage peroxisome proliferator-activated receptor α (PPAR-α) and γ (PPAR-γ) to facilitate cholesterol efflux, thus attenuating foam cell formation. In addition, reduced lipid antigen presentation by macrophages led to decreased IL-6 and IL-1β levels in lesions. To sum up, the NDSs multifaceted and remodeling of multiple cells in ASM effectively delayed AS.
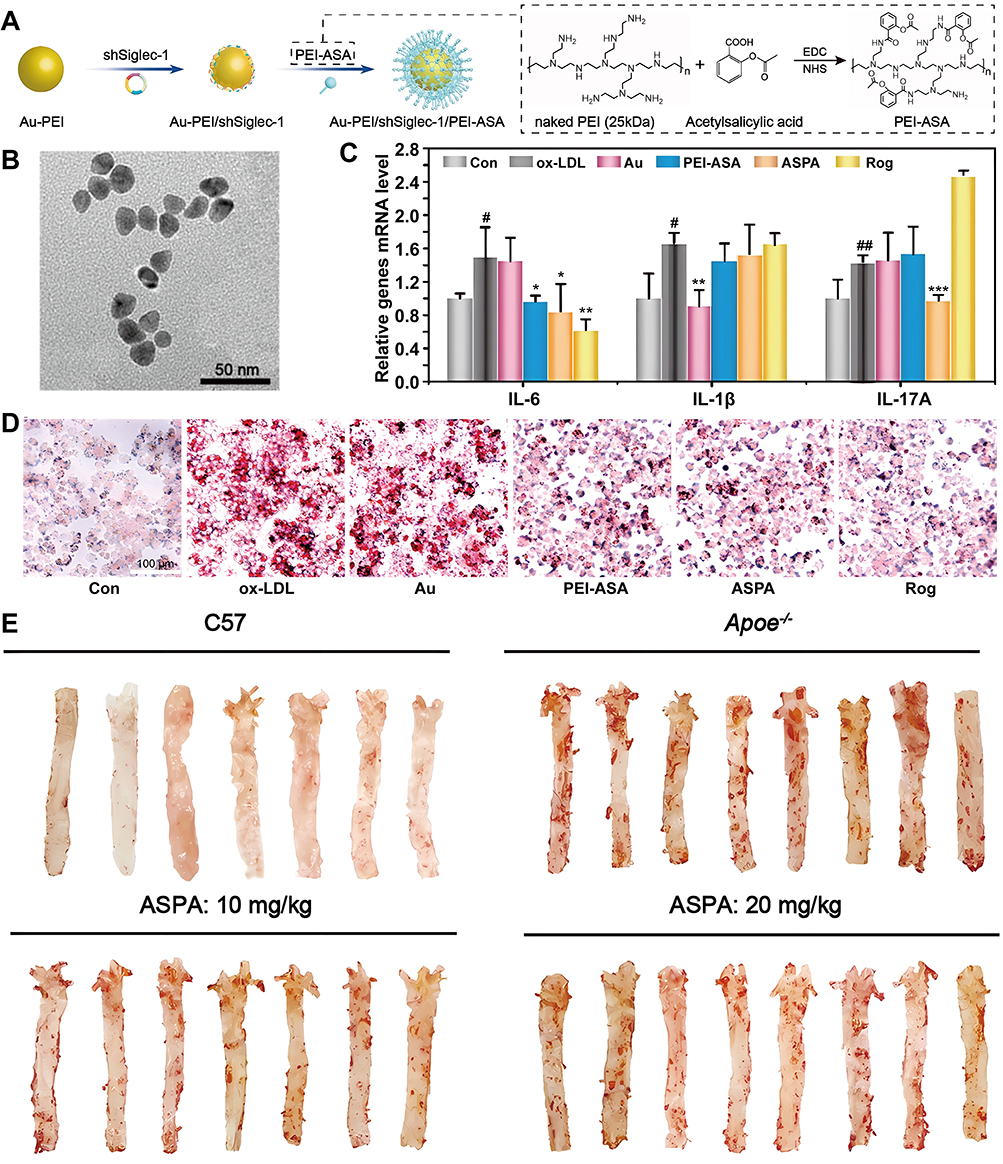 |
Figure 3 Preparation of ASPA NPs and their mechanism of action through remodeling of various cells to enhance the therapy of AS. (A) Schematic diagram of the construction of ASPA NPs. (B) TEM image of Au-PEI NPs (scale bar = 50 nm). (C) Inflammatory factor expression levels in RAW264.7 cells after different treatments. (D) ASPA NPs inhibits the formation of foam cells (scale bar = 100 μm). (E) Representative photographs of ORO-stained enface aortas from mice after different treatments. Data are shown as mean ± SD. #P < 0.05, ##P < 0.01, ox-LDL group vs Con group; test group vs ox-LDL group, *P < 0.05, **P < 0.01, ***P < 0.001. Reproducted from Zhou Y, Wang S, Liang X et al. Turning Hot into Cold: Immune Microenvironment Reshaping for Atherosclerosis Attenuation Based on pH-Responsive shSiglec-1 Delivery System. ACS Nano. Jul 26 2022;16(7):10,517–10,533.49 Copyright 2022, American Chemical Society. |
Accumulating evidence demonstrates that cellular senescence promotes AS progression across all disease stages and exacerbates cardiovascular pathogenesis. Emerging evidence indicates that CD9 critically regulates both cellular senescence induction and atherosclerotic plaque development via activation of the PI3K-AKT-mTOR-p53 signaling cascade. Here, Pham et al prepared a multifunctional NDSs (CD9-HMSN@RSV) utilizing CD9 antibody-modified, HA-coated mesoporous silica NPs, to deliver the anti-aging drug Rosuvastatin (RSV).47 This NDSs could effectively target senescent foam-like macrophages and senescent ECs. In vitro and in vivo studies explanation that these NDSs could productively remove senescent macrophages and ECs, further reduce ROS levels and inhibiting HDL oxidation, TNF-α and IL-6 production. In addition, inhibiting endothelial cell senescence while reprogramming the macrophage phenotype is expected to yield better therapeutic results. Herein, Peng et al developed osteopontin (OPN)-functionalized nanoliposomes, designated CZALO, which encapsulated both L-Arg and cerium-zirconium oxide nanoparticles (CZ NPs).50 On the one hand, the NDSs could achieve anti-inflammatory and antioxidant effects by reprogramming the macrophage phenotype through multi-enzyme mimetic activity, including SOD, CAT, peroxidase, and glutathione peroxidase. On the other hand, nitric oxide synthase overexpressed in macrophages selectively catalyze the in-situ conversion of L-Arg to NO, which modulates the release of senescence-associated secretory phenotype (SASP) factors from ECs, enhances lysosomal activity, overcomes cell-cycle blockade, and mitigates DNA damage in aged ECs. To sum up, the NDSs-mediated antioxidant and anti-aging therapeutic “killing two birds with one stone” strategy showed good anti-AS efficacy in vitro and in vivo.
Extracellular-Based Reprogramming Strategies
Inflammatory Response-Based Reprogramming Strategies
Clinical and epidemiological studies confirm that inflammation is pivotal in the development of AS. In its early stages, ox-LDL causes endothelial damage and subsequent mobilization and infiltration of monocytic and T cells. Circulating monocytes undergo macrophage differentiation, acquiring enhanced expression of innate immune receptors such as scavenger receptors and Toll-like receptors. Toll-like receptors further transmit activation signals, triggering the secretion of pro-inflammatory cytokines, matrix-degrading proteases, and vascular mediators that collectively establish a pro-inflammatory niche.51 During initial inflammatory responses, the dynamic interplay of inflammatory mediators orchestrates immune cell differentiation pathways and phenotypic commitment through concentration-dependent signaling gradients. For example, pro-inflammatory cytokines including TNF-α and IL-1β drive both endothelial-to-mesenchymal transition and monocyte polarization toward pro-inflammatory macrophage phenotypes.121 Subsequently, these activated immune cells and stromal cells secrete substantial quantities of inflammatory factors to exacerbate the inflammatory response and plaque growth.122 More importantly, inflammatory factors can also exacerbate plaque development by advancing apoptosis and necrotic tissue formation. In contrast, various anti-inflammatory factors can not only attenuate the inflammatory response induced by pro-inflammatory factors, but also induce the shift of immune cells to an anti-inflammatory phenotype, IL-10 and TGF-β can reverse mesenchymalisation of ECs and achieve macrophage repolarization.123 These immune cells with anti-inflammatory phenotypes can also secrete various anti-inflammatory factors to improve the inflammatory microenvironment and inhibit plaque growth.124 Therefore, the use of multifunctional NDSs to remodel the inflammatory response at the AS plaque site not only improves the inflammatory microenvironment, but also reprograms various immune cells into anti-inflammatory phenotypes to achieve enhanced therapeutic effects.
Although natural polyphenols exhibit potent anti-inflammatory properties, their clinical application is significantly constrained by poor bioavailability and chemical instability. NDSs with optimized physicochemical characteristics may effectively address these limitations. Here, Anghelache et al loaded protocatechuic acid (PCA) into dextran-modified iron oxide shells to obtain a core-shell magnetic NPs (MNP-Dex/PCA).68 The magnetic NPs enabled disease visualization to ensure localized release at the plaque site, which in turn significantly reduced the release of multiple pro-inflammatory factors by down-regulating the levels of ERK1/2 and p38-alpha mitogen-activated protein kinases (MAPKs), coupled with suppression of NF-κB transcriptional activity. Interestingly, the combination of statins and other bioactive drugs into a single polymeric nanocarrier promises enhanced synergistic therapeutic outcomes through complementary pathways. Here, Leal et al prepared a pleiotropic nanoplatform (PLGA/miRNA NPs) by co-loading atorvastatin (ATOR) with miRNA-124a into PLGA NPs.69 The nanoplatform was able to regulate inflammation-related genes simultaneously, and statins’ pleiotropic anti-inflammatory properties enabled targeted modulation of inflamed vascular endothelium, representing a therapeutic breakthrough in endothelial dysfunction management, utilizing complementary pathways to achieve synergistic therapeutic outcomes.
Increasing evidence suggests that delivery of anti-inflammatory factors such as IL-10 to ASM is expected to reprogram the inflammatory response and achieve enhanced therapeutic effects. To enhance IL-10 accumulation at plaque sites, Kim et al prepared plaque-localized NDS (IL10-NC) using cRGD-conjugated pluronic-based nanocarriers (NC) to deliver IL-10.70 Compared with free IL-10, IL-10-NC significantly improves its pharmacokinetic profile, thereby effectively increasing IL-10 accumulation at plaque sites, enabling an inflammatory response reprogramming strategy. The development from exogenous cytokine delivery to endogenous gene therapy can achieve the extension from short-term effects to long-term modulation. Herein, Distasio et al engineered a DNA NPs (NP-VHPK) utilizing VHPK peptide-conjugated branched poly (β-amino ester) for targeted delivery of IL-10 encoding plasmid DNA.71 The NPs achieves ECs-specific targeting via VHPK peptide-VCAM-1 molecular recognition, followed by robust IL-10 transgene expression post-internalization that potently attenuates inflammatory signaling.
ROS-Based Reprogramming Strategies
Numerous studies have shown that ROS, consisting of H2O2, O2·-, OH-, etc, is indispensable for the development of AS.125 Elevated ROS levels trigger NF-κB pathway activation, amplifying pro-inflammatory signaling cascades. This oxidative stress-mediated response enhances leukocyte-endothelial adhesion through upregulated cell adhesion molecules, ultimately inducing endothelial dysfunction and vascular injury.8,126 Therefore, therapeutic modulation of the inflammatory milieu through ROS scavenging may represent a promising strategy for attenuating disease progression.95,127
Contemporary antioxidant therapeutics primarily utilize ascorbic acid (VC), polyunsaturated fatty acids, and various polyphenolic compounds as core therapeutic agents.128–130 However, there are three major limitations of these antioxidants: firstly, short biological half-life and susceptibility to rapid hepatic and renal clearance; secondly, lack of plaque site targeting and inefficient delivery; and most importantly, limited ROS scavenging ability of single antioxidants.131,132 Here, Lu et al constructed a novel natural antioxidant (VC@cLAVs) by loading VC into cross-linked vesicles of lipoic acid (LA).72 This synergistic formulation substantially prolonged the plasma circulation time of the natural antioxidant while enhancing its redox activity. The coupled LA/DHLA and VC/DHA cycling systems created an amplified antioxidant effect, demonstrating superior ROS neutralization capacity. However, the NDSs still have the problem of insufficient targeting, in order upgrade the therapeutic strategy from “passive removal” to “active regulation”, Luo et al prepared a ROS-sensitive NPs (LLC NPs) by using heparin-lipoic acid coupling (LMWH-LA) as an amphiphilic shell for encapsulating curcumin (Cur) (Figure 4).73 The NPs target P-selectin on ECs at the plaque site, competitively blocking the migration of inflammatory cells to ECs and reducing ROS production from cellular sources. Subsequently, the NPs were oxidatively triggered to convert from hydrophilic to hydrophobic, accelerating the release of Cur, thereby removing excess ROS from the plaque.
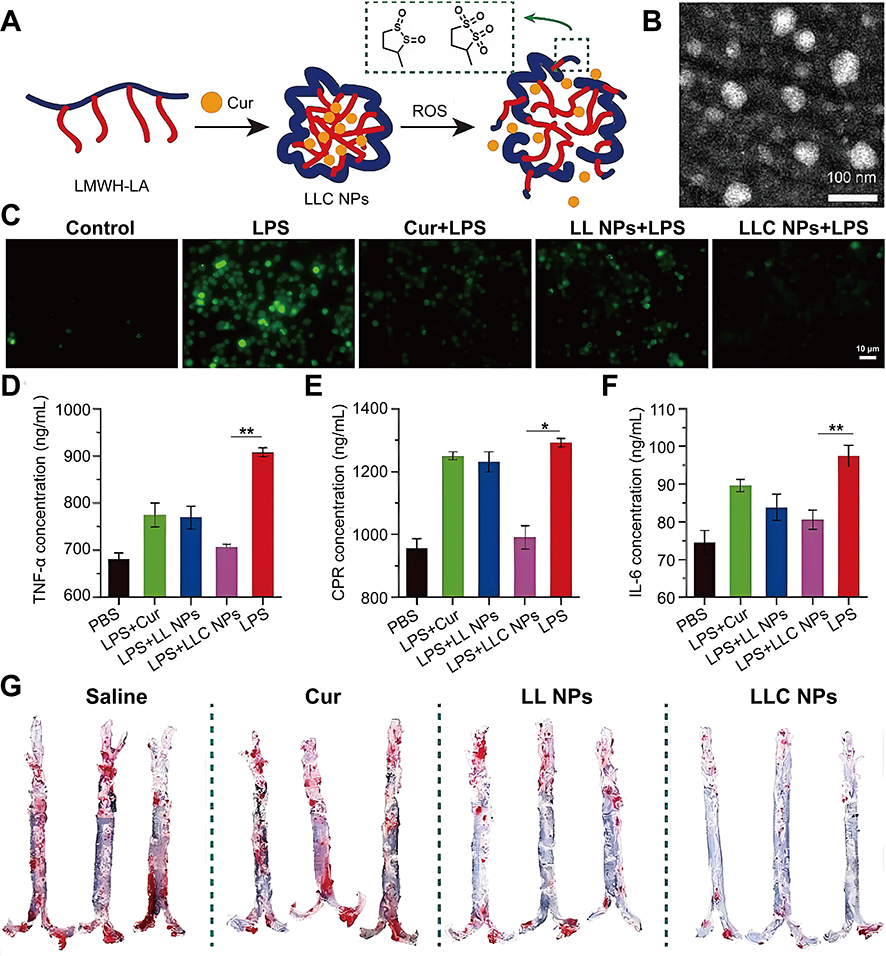 |
Figure 4 Preparation process of LLC NPs and their enhancement of the therapy of AS by scavenging ROS. (A) Schematic diagram of self-assembly and ROS response of LLC NPs (B) TEM images of LLC NPs (scale bar = 100 nm). (C) Flow cytometry was used to quantify the levels of intracellular ROS in RAW264.7 cells following various treatments (scale bar = 10 μm). (D–F) Inflammatory cytokines TNF-α (D), IL-6 (E), CRP (F) secreted by RAW264.7 cells treated with different formulation. (G) Image of ORO-stained enface aortas. Data are shown as mean ± SD, n = 5. *P < 0.05, **p < 0.01. Reproduction from Luo X, Zhang M, Dai W et al. Targeted nanoparticles triggered by plaque microenvironment for atherosclerosis treatment through cascade effects of reactive oxygen species scavenging and anti-inflammation. J Nanobiotechnology. Jul 27 2024;22(1):440. © The Author(s) 2024. This article is licensed under a Creative Commons Attribution 4.0 International License.73 |
Notably, a variety of inorganic nanomaterials possess CAT and SOD-like enzyme that can productively scavenge ROS, including CeO2, manganese oxide (MnOx), PB analogues and black phosphorus nanosheets (BPNSs).133 Among them, CeO2 NPs mimic the dual enzyme activities of SOD and CAT through the Ce3+/Ce4+ redox cycle.134 In addition, MnOx catalyzes ROS’s catabolism using its Mn2+/Mn3+/Mn4+ interconversion.135,136 Similarly, PB analogues scavenging ROS by redox of iron ions Fe2+/Fe3+.137 Functionalizing nanozymes as drug loading and delivery platforms is not only conducive to the improvement of their multi-enzyme activity to reduce toxic side effects, but also to enhance ROS’s scavenging.138 Here, Fu et al innovatively constructed a biomimetic delivery system (PCZ@PB NCs) by self-assembling zoledronic acid with cerium ions into cerium-like-MOF nanomaterials (CZ NCs) and encapsulating them through platelet membranes.74 This study not only enhanced the multi-enzyme activity of CeO2, the formulation demonstrated selective biodistribution to atherosclerotic plaques, where it effectively attenuated local oxidative stress through ROS downregulation. To optimize plaque targeting and therapeutic performance of nanozymatic delivery systems Wu et al prepared a NEs hitchhiking nanoplatform (MPB NPs) by using bacterial mimetic membranes (mycoderm) encapsulating Prussian blue nanoparticles (PB NPs).75 The NPs can be specifically recognized and efficiently internalized by NEs. Using the inflammatory chemotaxis mechanism, they can accurately accumulate at the AS plaque site. In addition, the NPs have the dual function of effectively scavenging intra and extracellular ROS and inhibiting NET formation. This multimodal action ultimately contributes to their anti-atherosclerotic efficacy. In addition, Tao et al obtained a type of nano-NDSs (BPNSs@PEG-S2P/R) by preparing a two-dimensional BPNSs and modifying its surface with ROS-responsive targeting materials for the delivery of Resolvin D1.76 These nanosheets exhibit high ROS’s scavenging rates due to their excellent specific surface area. Importantly, their layered structure facilitates their uptake by macrophages, allowing them to accumulate within plaques of AS, thereby effectively reducing expression levels of inflammatory factor and achieving the reprogramming of inflammatory macrophage.
Lipids-Based Reprogramming Strategies
Lipids are divided into three main categories: total cholesterol, HDL, and LDL. AS lesions typically manifest lipids metabolic dysregulation, exhibiting elevated LDL concentrations, reduced HDL levels, and overall cholesterol homeostasis disruption.139 When LDL penetrates damaged ECs, it is oxidized in response to ROS to form ox-LDL, which has the characteristics of a high degree of AS. Modified LDL particles undergo extensive macrophage internalization primarily through scavenger receptor-mediated endocytosis, such as CD36, LOX-1, and SR-A. This receptor-dependent uptake drives foam cell transformation through intracellular cholesterol ester accumulation, while simultaneously activating NF-κB dependent proinflammatory signaling cascades-collectively accelerating plaque development. Incorporating lipids reprogramming strategies into the design of NDSs is therefore essential to reduce lipid deposition, particularly the retention of LDL in the arterial wall. This can be achieved by a dual mechanism: reducing cellular uptake of cholesterol and enhancing cellular efflux of cholesterol. This approach has great therapeutic potential to improve the treatment of AS.
Studies show that statins have ability to decrease lipid uptake by M1 macrophages and increase the antioxidant capacity of LDL. Here, Rakshit et al prepared a simvastatin (STAT) nano-liposomal formulation (LIPOSTAT).81 When administered intravenously, it reduced serum cholesterol and LDL receptor expression in hepatocytes. It was found that oxidized phosphatidylcholines (PCs) on the surface of LDL promote its binding to macrophage CD36 receptors. To this end, Zhang et al prepared an NDSs (L-Enano) and successfully synthesized EGCG-loaded NPs (Enano) by using EGCG, PC, (+)α-tocopheryl acetate, and surfactants as the main raw materials and modified the surface of the nanocapsules with KOdiA-PC (1-(palmitoyl)-2-(5-keto-6-octen-diene) PC (KOdiA-PC)) to modify it.77 The NDSs competitively blocked oxLDL internalization in macrophages via KOdiA-PC’s high-affinity binding to the CD36 scavenger receptor, effectively disrupting foam cell formation.
Studies have shown that facilitating reverse cholesterol transport from macrophages can inhibit foam cell formation. Ligands for LXR or LXR agonists can upregulate the ATP-binding cassette transporter proteins ABCA1 and ABCG1 on macrophages, thereby enhancing cholesterol excretion.140,141 To this end, studies have utilized PLGA-b-PEG NPs to target delivery of LXR to macrophages on ABCA1 and ABCG1 as way to increase cholesterol efflux.142 Second, when excess LDL penetrates the arterial wall, it causes cholesterol crystals (CCs) to accumulate within the plaque and induces inflammation in a various cells through multiple pathways.143 Kim et al engineered core-shell NPs through host-guest complexation of simvastatin within methyl-β-cyclodextrin cavities, enveloped by a phospholipid bilayer shell, creating a cargo-switching NDSs (CSNP) capable of controlled drug release.78 The CSNP demonstrated excellent extracellular and intracellular CCs and cholesterol scavenging ability, effectively reducing cholesterol levels in AS. To further enhance the plaque targeting of NDSs, Chen et al constructed a target-responsive nanoplatform, LPLCH by using a covalent cholesterol group (PLC) of poly-ε-lysine coupled to a pH-responsive imine bond, loaded with an LXR agonist, and surface-modified with oxidized hyaluronic acid (oxHA).79 The nanoplatform could first be activated by the interaction between HA and CD44 targeting foam cells, and triggering a surface charge shift during lysosomal immune escape at acidic pH 5.5, effectively removing CCs from lysosomes by carrying them. Moreover, the NDSs could promote lipid efflux by targeting up-regulation of ABCA1 and ABCG1 at the site of plaques, they synergistically achieved dual-track reverse cholesterol transporter. In addition, HDL can interact with ABCA1, ABCG1, and scavenger receptor B1 (SR-BI), to facilitate macrophage-specific cholesterol efflux from foam cells through enhanced reverse cholesterol transport (RCT) pathways, promoting lipid clearance from plaque microenvironments.139,144 Here, Kudinov et al utilized PL NPs containing PL to reprogram HDL and improve the CEC.64 Likewise, Guo et al prepared a cholesterol efflux-promoting NDSs (sHDL-T1317) using phospholipid recombinant apoA-1 peptide (22A)-derived HDL NPs (sHDL) delivering the LXR agonist T1317.66 The NDSs significantly enhanced ABCA1 and ABCG1 transporter expression in macrophages and increased cholesterol efflux. More importantly, inhibiting cholesterol uptake while promoting cholesterol efflux would enable a better lipid reprogramming strategy. Here, a multifunctional NDSs (mDNP-LXR-L-siRNA) was prepared by optimized mannose-functionalized dendritic NDSs (mDNP) co-coating with reduce LDL uptake SR-A siRNA and LXR-L.80 This delivery strategy achieved aortic cholesterol-lowering function through a synergistic multi-pathway approach.
Multiple Components-Based Reprogramming Strategies
In ASM, multiple extracellular components, including inflammatory factors, ROS, and lipids, interact in a complex manner, creating a vicious cycle that exacerbates the inflammatory response and plaque formation.145–148 Consequently, a simultaneous reprogramming strategy targeting multiple components holds promise for increasing the AS therapy.
The deterioration of AS is characterized by overexpressed ROS and lipid deposition.149 Consequently, simultaneous modulation of ROS and lipid levels at the disease site is critical for improving efficacy. Here, Ma et al developed a therapeutic nanomicelles (TPCDP@PMM) for lipid and inflammation reprogramming.82 In addition, they first bridged LXR-L with β-cyclodextrin (β-CD) and loaded Pred onto β-CD via host-guest interactions to construct LCDP complexes, then encapsulated LCDPs and Π-conjugated polymer-based photoacoustic probes with ROS-responsive polymers (poly (2-methylthioethanol methacrylate)) to form PLCDP@PM NPs. Finally, oxHA was utilized to encapsulate the NPs and cross-link the MMP-9-sensitive peptide (MMP9 Pep) on their surface to obtain multifunctional NDSs (PLCDP@PMH).83 The NDSs effectively inhibited inflammatory response and plaque formation through M1 macrophage reprogramming strategy that synergized with LXR-mediated lipid reprogramming strategy, showing excellent therapeutic effects.
The phytochemical curcumin mediates cholesterol clearance from lipid-laden macrophages through SCAR-B1/SR-B1 receptor upregulation, while concurrently attenuating pro-inflammatory signaling via NF-κB and STAT-3 pathway suppression.150,151 Pillai et al obtained NDSs (Cur-Bio PLGA NPs) using polyethylene glycol delivery of Cur and biopiperidine.84 The NDSs significantly increased the bioavailability of Cur through the bioaugmentation of bioperine, effectively down-regulating inflammatory pathway-related genes to achieve the inflammatory reprogramming strategy. Meanwhile, it upregulated SCAR-B1 expression and inhibited the c-Jun N-terminal kinase pathway (JNK) to remove cholesterol accumulated in macrophages and reduced foam cells formation. Docosahexaenoic acid (DHA), which demonstrates potent anti-inflammatory effects. Here, Chong et al developed an injectable liposomal DHA for the treatment of AS.85 The NDSs could be effectively taken up by macrophages after reaching the plaque site, exerting anti-inflammatory and antioxidant effects. Moreover, the liposomal DHA formulation reduction in macrophage infiltration, decrease in plaque lipid content, increase in fibrillar collagen, and improving the stability of plaques.
Accumulating evidence indicates that molecular hydrogen (H2) can scavenging ROS and suppressing NF-κB mediated production of proinflammatory cytokines. Herein, Hu et al designed a distinct tetrapod needle-like palladium-hydrogen nanozyme (TN-PdHs) by adding Fe during Pd nano-enzyme synthesis and inserting H2 into the Pd nano-enzyme lattice.86 Interestingly, the specific spiky morphology of TN-PdHs triggers a strong autophagic response in macrophages, synergizing excellent ROS scavenging and anti-inflammatory capabilities to mitigate AS progression. Studies have shown that CCs exacerbate disease progression in AS by inducing inflammation and promoting plaque destabilization; therefore, depletion of CCs is expected to achieve dual regulation of inflammation and lipids. Herein, Luo et al prepared a phospholipid-based HDL-like nanoparticle (miNano) (Figure 5).87 The preparation enhanced cholesterol efflux by inhibiting the TLR4-NF-kB pathway, dissolved CCs in AS plaques, prevented foam cells formation, and concurrently inhibited inflammatory responses for effective disease treatment.
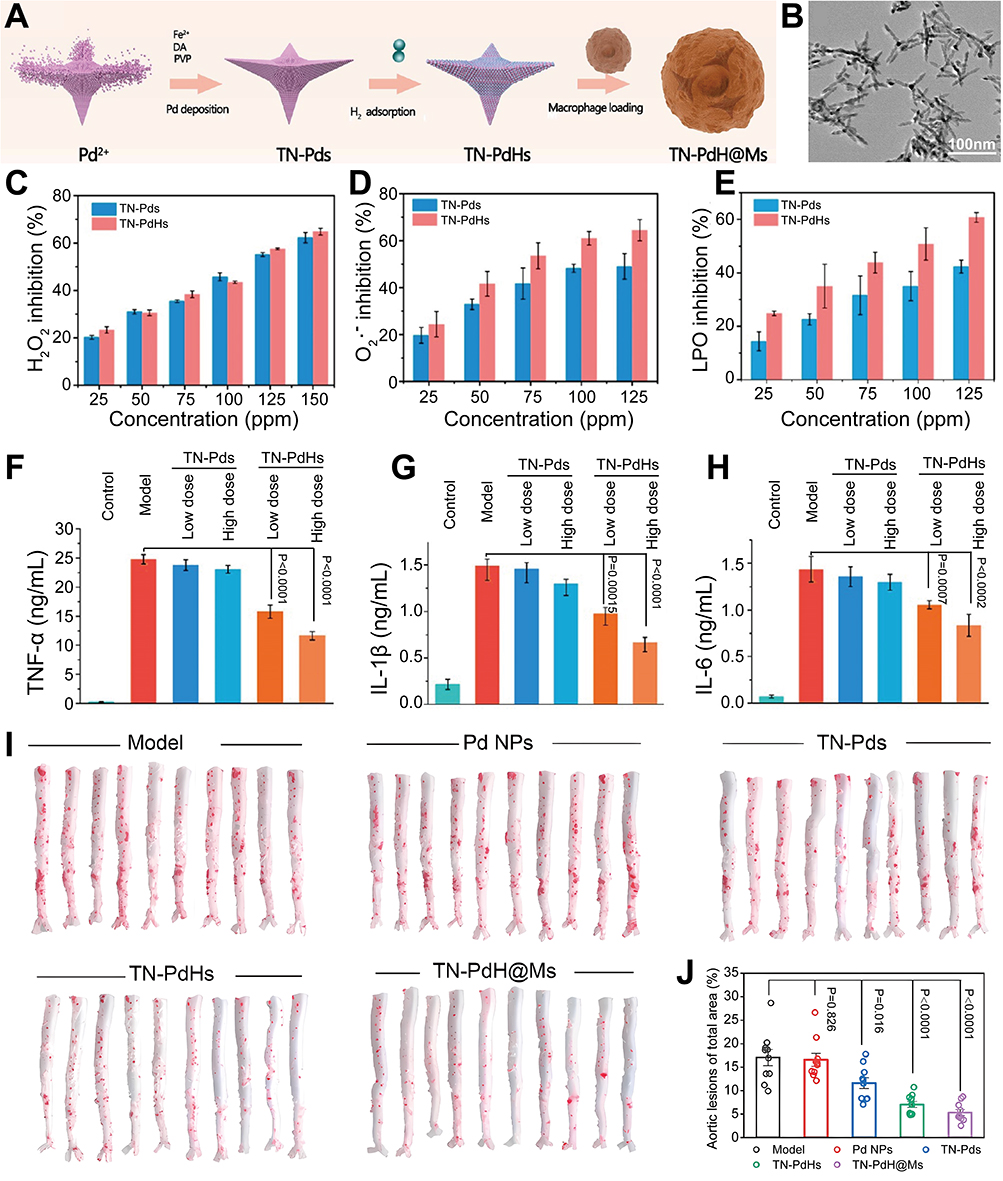 |
Figure 5 Preparation of TN-PdH@Ms and its enhancement of AS therapy by scavenging ROS and modulating inflammatory factors. (A) The preparation process of TN-PdH@Ms. (B) TEM image of TN-Pds (scale bar = 100 nm). (C) H2O2-scavenging performance of TN-PdHs. (D) O2•−-scavenging capability of TN-Pds and TN-PdHs. (E) LPO inhibition performance of TN-Pds and TN-PdHs. Inflammatory factor levels of (F) TNF-α, (G) IL-1β, and (H) IL-6 in LPS-induced RAW264.7 cells after varied treatments, including control, LPS, LPS + low dose TN-Pds (25 ppm), LPS + high dose TN-Pds (50 ppm), LPS + low dose TN-PdHs (25 ppm), and LPS + high dose TN-PdHs (50 ppm). (I) Representative photographs of enface ORO-stained aortas from each mouse after varied treatments. (J) Quantitative analysis of the lesion areas in the tested aortas. Reproduction form Hu R, Dai C, Dong C et al. Living Macrophage-Delivered Tetrapod PdH Nanoenzyme for Targeted Atherosclerosis Management by ROS Scavenging, Hydrogen Anti-inflammation, and Autophagy Activation. ACS Nano. Oct 25 2022;16(10):15,959–15,976.86 Copyright 2022, American Chemical Society. |
Discussion
The activation of various inflammatory cells and the accumulation of inflammatory mediators in ASM play an important role in disease progression and plaque stability. Undeniably, intracellular and extracellular reprogramming strategy solves the challenges of multidrug resistance and inferior specificity in AS treatment to some extent.152 NDSs offer substantial advantages for intracellular and extracellular reprogramming caused by their superior drug-loading capacity, target-specific modifications, ability to protect enzymatic stabilization, and intrinsic properties such as lipid-lowering and ROS scavenging capabilities. By reprogramming the interactions among various cells and components within the ASM, NDSs can further optimize the ASM and significantly enhance the therapeutic efficacy of AS.153
However, significant challenges remain for the practical application of NDSs in AS therapy. Plaque formation is primarily driven by a complex interplay of immune and inflammatory responses. ASM is a highly complex and dynamic process: it involves cell-cell, cell-molecule, and molecule-molecule interactions.154 It should be emphasized that NPs are merely discrete carrier units with particle sizes ranging from 1 to 100 nm, while NDSs integrate surface modifications, targeting ligands, and controlled-release modules on this basis to form a fully functional delivery system. The two are related as “components” and “systems”.155 Current research predominantly focuses on isolated aspects of NDSs’ effects on the ASM, often overlooking the complex network of interactions within the microenvironment. This narrow focus frequently results in suboptimal therapeutic outcomes.156 Furthermore, enzymes, inflammatory factors, and extracellular matrix components in the ASM can alter the surface properties of NPs, including surface zeta potential, hydrophilicity or hydrophobicity, functional groups, and ligand morphology, thereby affecting their binding efficiency to target cells.157 Additionally, after intravenous injection, NPs must traverse the endothelial barrier. Larger NPs tend to accumulate in the bloodstream, imposing additional challenges for AS treatment.158 In addition, compositional and structural changes in the ECM, such as alterations in collagen and glycosaminoglycans, increase the viscosity of the microenvironment, thereby reducing the targeting efficiency of NPs. The physicochemical properties of ASM, such as a weakly acidic environment and a low concentration of oxygen, also affect the stability and release rate of the drug.159 During systemic circulation, specific enzymes, such as MMP, degrade the carrier materials of NPs, including poly (lactic acid) (PLA), poly (glycolic acid) (PGA), and collagen. Proteolytic cleavage of the delivery vehicle triggers untimely payload liberation, reducing therapeutic in simulated physiological conditions.160 Furthermore, premature release of NPs loaded with apoptosis-inducing or antiproliferative drugs can result in insufficient cell specificity, causing unintended damage to healthy cells. Therefore, delivery strategies must be optimized to improve the safety and specificity of NDSs.161 This includes exploring more specific biomarkers such as angiogenesis targets, apoptosis/necrosis targets, etc. Developing superior targeted ligands for existing biomarkers, such as VCAM-1 antibodies and RGD peptides. Avoiding the clearance by immune system through invisible coatings and biomimetic nanoparticles. Reducing interference from blood rheology through shape’s optimization, promoting drug’s accumulation at plaque sites through charge’s regulation, microenvironment-responsive design, and targeted ligand’s modification. At the same time, NPs exhibit more complex biotoxicity and toxicological mechanisms compared to traditional drug formulations. Certain NPs, such as metal-based NPs (eg, Ag NPs, Fe3O4NPs, and zinc NPs [Zn NPs]), can damage the endothelial barrier, trigger inflammatory responses, and exacerbate the progression of AS. Therefore, when designing NDSs, it is important to consider whether NPs exacerbate the risk of AS.162
Although NPs could enhance therapeutic efficacy, difficulties with their safety and efficacy assessment technologies have made it difficult to move them into clinical translation.163,164 For example, the complex in vivo behavior of nanomaterials, including poor stability, unpredictable biodistribution, accelerated blood clearance, and low permeability through physiological barriers, often leads to diminished efficacy in human’s trials compared to animal’s studies. More importantly, the immunostimulatory and immunosuppressive properties of NPs pose a series of unique challenges. Their interaction with the immune system can sometimes cause adverse reactions, such as inflammation triggered by lipid or surfactant components, which may impair therapeutic efficacy. Conversely, immunosuppressive NPs may inadvertently suppress local immune responses, compromising the body’s ability to fight infection. A comprehensive assessment of immunological safety is critical for successful clinical translation. In addition, insufficient understanding of the pathogenesis of severe cardiovascular diseases, the classification of macrophage’s phenotype in the microenvironment, and the actual clinical relevance of specific cell’s states further limits the application of NDSs based on intracellular reprogramming strategies in clinical practice. Pharmacological and pathological differences in nanoparticle’s behavior, both within cells and between animals’ models and clinical patients, contribute to low clinical translation rates.165 Finally, the preparation process for nanomaterials is complex and significantly more expensive than conventional treatment’s regimens, which often results in poor patient’s compliance, posing another obstacle to the clinical translation of nanomedicine. Consequently, further research is needed to optimize the performance and conduct comprehensive risk assessments of NDSs. This includes optimizing the targeting of nanoparticles in complex biological environments, establishing standardized methods of toxicity assessment, and addressing factors that limit large-scale production and regulatory approval.
Studies have shown that abnormalities in glycolytic flux during the development of AS accelerate its progression, and promote endothelial dysfunction.166,167 Glycolysis-mediated has been established as a key regulator of phenotypic switching of macrophage in AS.168 Enhanced glycolytic activity also drives the proliferation and migration of VSMCs. Consequently, reducing ECs’s dysfunction by inhibiting glycolysis and inhibiting polarization of macrophage and proliferation of VSMCs may stand for a creative therapeutic approach for AS. In addition, dysregulated lipid’s metabolism serves as a pathogenic cornerstone of AS, and it has implications for the design of NDSs’ targeting therapies.169 To sum up, the complex metabolic lattice in ASM requires further investigation to advance the integration of metabolic reprogramming strategies with AS. More importantly, the metalloproteinase system, comprising MMP and their endogenous tissue inhibitors (TIMPs), exhibits dualistic regulation of advanced plaque pathophysiology. On one hand, their beneficial effects include promoting VSMCs’ growth and survival, thereby contributing to plaque’s stabilization. On the other hand, excessive MMP’s activity and persistent monocyte/macrophage’s accumulation lead to the degradation of extracellular matrix, which promotes plaque’s destabilization and rupture.170 Accordingly, the controlled regulation of MMP is important for the treatment of AS. Other studies have showed that CXCR1 and CXCR2 chemokine receptors are overexpressed on the surface of various cell types. These cell surface receptors critically mediate the trafficking of leukocyte and NEs to inflamed tissues through chemotactic signaling cascades, thereby amplifying pathological inflammation and accelerating pathogenesis of the disease.171 Therefore, targeting and inhibiting chemokine receptors on cell’s surfaces by optimized NDSs to remodel the cellular phenotype and inhibit inflammatory cell’s recruitment has potential research value for the treatment of AS. Previously, we prepared NDSs with hitchhiking strategy of a circulating monocyte for the treatment of myocardial ischemia/reperfusion injury (MI/RI).172 Our results demonstrated that these NDSs could efficiently target lesion sites by leveraging the inflammatory homing and plasticity properties of monocytes. Furthermore, the NDSs’ reprogrammed MNs into anti-inflammatory macrophages, thereby exerting therapeutic effects. This approach provides valuable insights for designing reprogramming strategies targeting monocytes in AS, given the similarities in the inflammatory microenvironments of these two diseases. In summary, several critical considerations remain for NDSs-based modulation of ASM, including optimization of its physicochemical characteristics, biological complexity of heterogeneous plaque components, and risk-benefit assessment of potential off-target effects and long-term biocompatibility. However, the enhancement of drug delivery efficiency and the improvement of the inflammatory microenvironment in AS through NDSs represent a highly promising research direction. Our ongoing research focuses on pioneering next-generation NDSs’ engineered to precisely target and remodel the ASM. These efforts aim to develop multifunctional platforms for direct plaque’s regression, and synergistic adjuvants to enhance conventional therapies, ultimately advancing precision medicine for the management of AS.
Conclusion
This review summarizes recent advances in NDSs for enhancing the treatment of AS based on intracellular and extracellular reprogramming strategies. Among intracellular reprogramming approaches, we highlight various NDSs specifically targeting plaque-resident endothelial cells, monocytes, inflammatory macrophages, NEs, and VSMCs within plaque sites. These strategies aim to reverse endothelial-mesenchymal transition and inflammatory states, reprogram the differentiation pathways of monocyte, promote the repolarization of inflammatory macrophages into anti-inflammatory macrophages, suppress infiltration of NEs and NETs-driven inflammatory responses, and inhibit proliferation and migration of VSMCs. In extracellular reprogramming strategies, we focused on regulating inflammatory mediators, ROS, and lipids within the ASM using various NDSs. This achieves extracellular reprogramming by reshaping pro-inflammatory responses into anti-inflammatory ones, scavenging ROS, and remodeling the fate of lipid. More importantly, we examined how optimized multifunctional NDSs simultaneously reprogram multiple cell types or components within plaques of AS to achieve enhanced therapeutic outcomes. Finally, we explored the advantages and potential of NDSs based on intracellular and extracellular reprogramming strategies for improving treatment efficacy of the disease, including restoring endothelial barriers, suppressing inflammation, clearing lipids, and enhancing plaque’s stability. Additionally, we address challenges and critical issues in the clinical translation of novel NDSs. In summary, future research should prioritize elucidating the relationship between structure and function of NDSs while deepening understanding of its microenvironment of the disease. This will guide the design of superior NDSs capable of regulating multiple inflammatory cells and mediators at the disease site, thereby comprehensively improving its microenvironment. This review aims to provide guidance for designing NDSs based on intracellular and extracellular reprogramming strategies.
Funding
This work was supported by the Sichuan Provincial Science and Technology Project (grant number (2025ZNSFSC0710); the Luzhou Municipal People’s Government-Southwest Medical University Joint Scientific Research Project (grant number 2025LZXNYDJC16, 2024LZXNYDJ070, 2023LZXNYDJ003); the Project of Application and Basic Research of Southwest Medical University [2024YYJC091].
Disclosure
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper. The images used in this manuscript that are related to other articles have been authorized by the authors and publishers of those articles, and comply with correct ethics statement.
References
1. Libby P. The changing landscape of atherosclerosis. Nature. 2021;592(7855):524–26. doi:10.1038/s41586-021-03392-8
2. Hou P, Fang J, Liu Z, et al. Macrophage polarization and metabolism in atherosclerosis. Cell Death Dis. 2023;14(10):691. doi:10.1038/s41419-023-06206-z
3. Wilkins JT, Gidding SS, Robinson JG. Can atherosclerosis be cured? Curr Opin Lipidol. 2019;30(6):477–484. doi:10.1097/mol.0000000000000644
4. Younis MA, Tawfeek HM, Abdellatif AAH, Abdel-Aleem JA, Harashima H. Clinical translation of nanomedicines: challenges, opportunities, and keys. Adv Drug Deliv Rev. 2022;181:114083. doi:10.1016/j.addr.2021.114083
5. Nesti L, Mengozzi A, Natali A. Statins, LDL cholesterol control, cardiovascular disease prevention, and atherosclerosis progression: a clinical perspective. Am J Cardiovasc Drugs. 2020;20(5):405–412. doi:10.1007/s40256-019-00391-z
6. Hidalgo A, Libby P, Soehnlein O, Aramburu IV, Papayannopoulos V, Silvestre-Roig C. Neutrophil extracellular traps: from physiology to pathology. Cardiovasc Res. 2022;118(13):2737–2753. doi:10.1093/cvr/cvab329
7. Tabas I, Bornfeldt KE. Intracellular and intercellular aspects of macrophage immunometabolism in atherosclerosis. Circ Res. 2020;126(9):1209–1227. doi:10.1161/circresaha.119.315939
8. Zheng D, Liu J, Piao H, Zhu Z, Wei R, Liu K. ROS-triggered endothelial cell death mechanisms: focus on pyroptosis, parthanatos, and ferroptosis. Front Immunol. 2022;13:1039241. doi:10.3389/fimmu.2022.1039241
9. Miano JM, Fisher EA, Majesky MW. Fate and state of vascular smooth muscle cells in atherosclerosis. Circulation. 2021;143(21):2110–2116. doi:10.1161/circulationaha.120.049922
10. Gaggini M, Gorini F, Vassalle C. Lipids in atherosclerosis: pathophysiology and the role of calculated lipid indices in assessing cardiovascular risk in patients with hyperlipidemia. Int J Mol Sci. 2022;24(1). doi:10.3390/ijms24010075
11. Batty M, Bennett MR, Yu E. The role of oxidative stress in atherosclerosis. Cells. 2022;11(23). doi:10.3390/cells11233843
12. Cheng L, Yue H, Zhang H, et al. The influence of microenvironment stiffness on endothelial cell fate: implication for occurrence and progression of atherosclerosis. Life Sci. 2023;334:122233. doi:10.1016/j.lfs.2023.122233
13. Demkow U. Molecular mechanisms of neutrophil extracellular trap (NETs) degradation. Int J Mol Sci. 2023;24(5). doi:10.3390/ijms24054896
14. Yang X, Ma Y, Chen X, Zhu J, Xue W, Ning K. Mechanisms of neutrophil extracellular trap in chronic inflammation of endothelium in atherosclerosis. Life Sci. 2023;328:121867. doi:10.1016/j.lfs.2023.121867
15. Xu S, Ilyas I, Little PJ, et al. Endothelial dysfunction in atherosclerotic cardiovascular diseases and beyond: from mechanism to pharmacotherapies. Pharmacol Rev. 2021;73(3):924–967. doi:10.1124/pharmrev.120.000096
16. Singh V, Kaur R, Kumari P, Pasricha C, Singh R. ICAM-1 and VCAM-1: gatekeepers in various inflammatory and cardiovascular disorders. Clin Chim Acta. 2023;548:117487. doi:10.1016/j.cca.2023.117487
17. Yang S, Yuan HQ, Hao YM, et al. Macrophage polarization in atherosclerosis. Clin Chim Acta. 2020;501:142–146. doi:10.1016/j.cca.2019.10.034
18. Manta CP, Leibing T, Friedrich M, et al. Targeting of scavenger receptors stabilin-1 and stabilin-2 ameliorates atherosclerosis by a plasma proteome switch mediating monocyte/macrophage suppression. Circulation. 2022;146(23):1783–1799. doi:10.1161/circulationaha.121.058615
19. Mineo C. Lipoprotein receptor signalling in atherosclerosis. Cardiovasc Res. 2020;116(7):1254–1274. doi:10.1093/cvr/cvz338
20. Iaconisi GN, Lunetti P, Gallo N, et al. Hyaluronic acid: a powerful biomolecule with wide-ranging applications-a comprehensive review. Int J Mol Sci. 2023;24(12). doi:10.3390/ijms241210296
21. Wen X, Zeng X, Liu J, et al. In vivo comparative study of radioiodinated folate receptor targeting albumin probes for atherosclerosis plaque imaging. Bioconjug Chem. 2023;34(12):2387–2397. doi:10.1021/acs.bioconjchem.3c00486
22. Segers FME, Ruder AV, Westra MM, et al. Magnetic resonance imaging contrast-enhancement with superparamagnetic iron oxide nanoparticles amplifies macrophage foam cell apoptosis in human and murine atherosclerosis. Cardiovasc Res. 2023;118(17):3346–3359. doi:10.1093/cvr/cvac032
23. Tang C, Wang H, Guo L, et al. CpG-conjugated silver nanoparticles as a multifunctional nanomedicine to promote macrophage efferocytosis and repolarization for atherosclerosis therapy. ACS Appl Mater Interfaces. 2023. doi:10.1021/acsami.3c11227
24. Li F, Zhang H. Targeting Macrophage Epsins to Reverse Atherosclerosis. Circ Res. 2023;132(1):7–9. doi:10.1161/circresaha.122.322273
25. Huang X, Zhang Y, Zhang W, et al. Osteopontin-targeted and PPARδ-agonist-loaded nanoparticles efficiently reduce atherosclerosis in apolipoprotein E−/− mice. ACS Omega. 2022;7(33):28767–28778. doi:10.1021/acsomega.2c00575
26. Xu H, She P, Ma B, Zhao Z, Li G, Wang Y. ROS responsive nanoparticles loaded with lipid-specific AIEgen for atherosclerosis-targeted diagnosis and bifunctional therapy. Biomaterials. 2022;288:121734. doi:10.1016/j.biomaterials.2022.121734
27. Dosta P, Tamargo I, Ramos V, et al. Delivery of anti-microRNA-712 to inflamed endothelial cells using Poly(beta-amino ester) nanoparticles conjugated with VCAM-1 targeting peptide. Adv Healthc Mater. 2021;10(15):e2001894. doi:10.1002/adhm.202001894
28. Liu Y, He M, Yuan Y, et al. Neutrophil-membrane-coated biomineralized metal-organic framework nanoparticles for atherosclerosis treatment by targeting gene silencing. ACS Nano. 2023;17(8):7721–7732. doi:10.1021/acsnano.3c00288
29. Gao W, Yang H, Liu X, et al. Reductively dissociable biomimetic nanoparticles for control of integrin-coupled inflammatory signaling to retard atherogenesis. Chem Commun. 2019;55(77):11535–11538. doi:10.1039/c9cc06039a
30. Liu J, Yu X, Braucht A, Smith S, Wang C. N-Cadherin targeted melanin nanoparticles reverse the endothelial-mesenchymal transition in vascular endothelial cells to potentially slow the progression of atherosclerosis and cancer. ACS Nano. 2024;18(11):8229–8247. doi:10.1021/acsnano.3c12281
31. Yin L, Peng C, Tang Y, et al. Biomimetic oral targeted delivery of bindarit for immunotherapy of atherosclerosis. Biomater Sci. 2020;8(13):3640–3648. doi:10.1039/d0bm00418a
32. Fu C, Tao Y, Li Z, et al. Circulating monocyte differentiation-activated nanoprodrugs for reprogramming macrophage immunity in atherosclerotic plaques. Nano Today. 2024;56:102304. doi:10.1016/j.nantod.2024.102304
33. Nakashiro S, Matoba T, Umezu R, et al. Pioglitazone-incorporated nanoparticles prevent plaque destabilization and rupture by regulating monocyte/macrophage differentiation in ApoE-/- mice. Arterioscler Thromb Vasc Biol. 2016;36(3):491–500. doi:10.1161/atvbaha.115.307057
34. Wu H, Sheng J, Wang Z, et al. Tannic acid-poloxamer self-assembled nanoparticles for advanced atherosclerosis therapy by regulation of macrophage polarization. J Mater Chem B. 2024;12(19):4708–4716. doi:10.1039/d3tb01157g
35. Wan J, Yang J, Lei W, et al. Anti-oxidative, anti-apoptotic, and M2 polarized DSPC liposome nanoparticles for selective treatment of atherosclerosis. Int J Nanomed. 2023;18:579–594. doi:10.2147/ijn.S384675
36. Zhao J, Ling L, Zhu W, et al. M1/M2 re-polarization of kaempferol biomimetic NPs in anti-inflammatory therapy of atherosclerosis. J Control Release. 2023;353:1068–1083. doi:10.1016/j.jconrel.2022.12.041
37. Mu D, Wang W, Li J, et al. Ultrasmall Fe(III)-tannic acid nanoparticles to prevent progression of atherosclerotic plaques. ACS Appl Mater Interfaces. 2021;13(29):33915–33925. doi:10.1021/acsami.1c09480
38. Guo J, Li D, Tao H, et al. Cyclodextrin-derived intrinsically bioactive nanoparticles for treatment of acute and chronic inflammatory diseases. Adv Mater. 2019;31(46):e1904607. doi:10.1002/adma.201904607
39. Molinaro R, Yu M, Sausen G, et al. Targeted delivery of protein arginine deiminase-4 inhibitors to limit arterial intimal NETosis and preserve endothelial integrity. Cardiovasc Res. 2021;117(13):2652–2663. doi:10.1093/cvr/cvab074
40. Shi Y, Dong M, Wu Y, et al. An elastase-inhibiting, plaque-targeting and neutrophil-hitchhiking liposome against atherosclerosis. Acta Biomater. 2024;173:470–481. doi:10.1016/j.actbio.2023.11.020
41. Wei G, Hao L, Li X, et al. VCAM-1-targeted and PPARdelta-agonist-loaded nanomicelles enhanced suppressing effects on apoptosis and migration of oxidized low-density lipoprotein-induced vascular smooth muscle cells. Biosci Rep. 2020;40(5). doi:10.1042/BSR20200559
42. Zhang S, Xu W, Gao P, Chen W, Zhou Q. Construction of dual nanomedicines for the imaging and alleviation of atherosclerosis. Artif Cells Nanomed Biotechnol. 2020;48(1):169–179. doi:10.1080/21691401.2019.1699823
43. Chin DD, Poon C, Wang J, et al. miR-145 micelles mitigate atherosclerosis by modulating vascular smooth muscle cell phenotype. Biomaterials. 2021;273:120810. doi:10.1016/j.biomaterials.2021.120810
44. Maiseyeu A, Di L, Ravodina A, et al. Plaque-targeted, proteolysis-resistant, activatable and MRI-visible nano-GLP-1 receptor agonist targets smooth muscle cell differentiation in atherosclerosis. Theranostics. 2022;12(6):2741–2757. doi:10.7150/thno.66456
45. Deng Y, Liu L, Li Y, et al. pH-sensitive nano-drug delivery systems dual-target endothelial cells and macrophages for enhanced treatment of atherosclerosis. Drug Deliv Transl Res. 2025. doi:10.1007/s13346-025-01791-2
46. Wu Z, Wu R, Li X, et al. Multi-pathway microenvironment regulation for atherosclerosis therapy based on beta-cyclodextrin/L-Arginine/Au nanomotors with dual-mode propulsion. Small. 2022;18(9):e2104120. doi:10.1002/smll.202104120
47. Pham LM, Kim EC, Ou W, et al. Targeting and clearance of senescent foamy macrophages and senescent endothelial cells by antibody-functionalized mesoporous silica nanoparticles for alleviating aorta atherosclerosis. Biomaterials. 2021;269:120677. doi:10.1016/j.biomaterials.2021.120677
48. An H, Qiu X, Wang X, et al. LIFU-unlocked endogenous H2S generation for enhancing atherosclerosis-specific gas-enzymatic therapy. Biomaterials. 2025;315:122972. doi:10.1016/j.biomaterials.2024.122972
49. Zhou Y, Wang S, Liang X, et al. Turning hot into cold: immune microenvironment reshaping for atherosclerosis attenuation based on pH-responsive shSiglec-1 delivery system. ACS Nano. 2022;16(7):10517–10533. doi:10.1021/acsnano.2c01778
50. Peng Y, Feng W, Huang H, Chen Y, Yang S. Macrophage-targeting antisenescence nanomedicine enables in-Situ NO induction for Gaseous and antioxidative atherosclerosis intervention. Bioact Mater. 2025;48:294–312. doi:10.1016/j.bioactmat.2025.02.025
51. Kong P, Cui ZY, Huang XF, Zhang DD, Guo RJ, Han M. Inflammation and atherosclerosis: signaling pathways and therapeutic intervention. Signal Transduct Target Ther. 2022;7(1):131. doi:10.1038/s41392-022-00955-7
52. Schwarz N, Fernando S, Chen YC, et al. Colchicine exerts anti-atherosclerotic and -plaque-stabilizing effects targeting foam cell formation. FASEB J. 2023;37(4):e22846. doi:10.1096/fj.202201469R
53. Deftereos SG, Beerkens FJ, Shah B, et al. Colchicine in cardiovascular disease: in-depth review. Circulation. 2022;145(1):61–78. doi:10.1161/circulationaha.121.056171
54. Tardif JC, Kouz S, Waters DD, et al. Efficacy and safety of low-dose colchicine after myocardial infarction. N Engl J Med. 2019;381(26):2497–2505. doi:10.1056/NEJMoa1912388
55. Singh S, Changkija S, Mudgal R, Ravichandiran V. Bioactive components to inhibit foam cell formation in atherosclerosis. Mol Biol Rep. 2022;49(3):2487–2501. doi:10.1007/s11033-021-07039-9
56. Galindo CL, Khan S, Zhang X, Yeh YS, Liu Z, Razani B. Lipid-laden foam cells in the pathology of atherosclerosis: shedding light on new therapeutic targets. Expert Opin Ther Targets. 2023;27(12):1231–1245. doi:10.1080/14728222.2023.2288272
57. Zhao H, Zhang R, Yan X, Fan K. Superoxide dismutase nanozymes: an emerging star for anti-oxidation. J Mater Chem B. 2021;9(35):6939–6957. doi:10.1039/d1tb00720c
58. Yao X, Chen D, Zhao B, et al. Acid-degradable hydrogen-generating metal-organic framework for overcoming cancer resistance/metastasis and off-target side effects. Adv Sci. 2022;9(10):e2101965. doi:10.1002/advs.202101965
59. Kirla H, Henry DJ, Jansen S, Thompson PL, Hamzah J. Use of silica nanoparticles for drug delivery in cardiovascular disease. Clin Ther. 2023;45(11):1060–1068. doi:10.1016/j.clinthera.2023.08.017
60. Wang Y, Liang Z, Liang Z, Lv W, Chen M, Zhao Y. Advancements of Prussian blue-based nanoplatforms in biomedical fields: progress and perspectives. J Control Release. 2022;351:752–778. doi:10.1016/j.jconrel.2022.10.007
61. Dabravolski SA, Sukhorukov VN, Melnichenko AA, Khotina VA, Orekhov AN. The role of selenium in atherosclerosis development, progression, prevention and treatment. Biomedicines. 2023;11(7). doi:10.3390/biomedicines11072010
62. Tsushima Y, Hatipoglu B. Statin Intolerance: A Review And Update. Endocr Pract. 2023;29(7):566–571. doi:10.1016/j.eprac.2023.03.004
63. Vell MS, Loomba R, Krishnan A, et al. Association of statin use with risk of liver disease, hepatocellular carcinoma, and liver-related mortality. JAMA Network Open. 2023;6(6):e2320222. doi:10.1001/jamanetworkopen.2023.20222
64. Kudinov VA, Torkhovskaya TI, Zakharova TS, et al. High-density lipoprotein remodeling by phospholipid nanoparticles improves cholesterol efflux capacity and protects from atherosclerosis. Biomed Pharmacother. 2021;141:111900. doi:10.1016/j.biopha.2021.111900
65. Prilepskii AY, Serov NS, Kladko DV, Vinogradov VV. Nanoparticle-based approaches towards the treatment of atherosclerosis. Pharmaceutics. 2020;12(11). doi:10.3390/pharmaceutics12111056
66. Guo Y, Yuan W, Yu B, et al. Synthetic high-density lipoprotein-mediated targeted delivery of liver x receptors agonist promotes atherosclerosis regression. EBioMedicine. 2018;28:225–233. doi:10.1016/j.ebiom.2017.12.021
67. He H, Yuan Q, Bie J, et al. Development of mannose functionalized dendrimeric nanoparticles for targeted delivery to macrophages: use of this platform to modulate atherosclerosis. Transl Res. 2018;193:13–30. doi:10.1016/j.trsl.2017.10.008
68. Anghelache M, Turtoi M, Petrovici AR, Fifere A, Pinteala M, Calin M. Development of dextran-coated magnetic nanoparticles loaded with protocatechuic acid for vascular inflammation therapy. Pharmaceutics. 2021;13(9). doi:10.3390/pharmaceutics13091414
69. Leal BH, Velasco B, Cambon A, et al. Combined therapeutics for atherosclerosis treatment using polymeric nanovectors. Pharmaceutics. 2022;14(2). doi:10.3390/pharmaceutics14020258
70. Kim M, Sahu A, Hwang Y, et al. Targeted delivery of anti-inflammatory cytokine by nanocarrier reduces atherosclerosis in Apo E−/− mice. Biomaterials. 2020;226:119550. doi:10.1016/j.biomaterials.2019.119550
71. Distasio N, Dierick F, Ebrahimian T, Tabrizian M, Lehoux S. Design and development of Branched Poly(ss-aminoester) nanoparticles for Interleukin-10 gene delivery in a mouse model of atherosclerosis. Acta Biomater. 2022;143:356–371. doi:10.1016/j.actbio.2022.02.043
72. Lu X, He Z, Xiao X, Wei X, Song X, Zhang S. Natural antioxidant-based nanodrug for atherosclerosis treatment. Small. 2023;19(49):e2303459. doi:10.1002/smll.202303459
73. Luo X, Zhang M, Dai W, et al. Targeted nanoparticles triggered by plaque microenvironment for atherosclerosis treatment through cascade effects of reactive oxygen species scavenging and anti-inflammation. J Nanobiotechnol. 2024;22(1):440. doi:10.1186/s12951-024-02652-9
74. Fu X, Yu X, Jiang J, et al. Small molecule-assisted assembly of multifunctional ceria nanozymes for synergistic treatment of atherosclerosis. Nat Commun. 2022;13(1):6528. doi:10.1038/s41467-022-34248-y
75. Wu M, Chen M, Zhao Y, et al. Neutrophil hitchhiking-mediated delivery of ROS-scavenging biomimetic nanoparticles for enhanced treatment of atherosclerosis. Small Methods. 2025:e2402019. doi:10.1002/smtd.202402019.
76. He Z, Chen W, Hu K, et al. Resolvin D1 delivery to lesional macrophages using antioxidative black phosphorus nanosheets for atherosclerosis treatment. Nature Nanotechnol. 2024;19(9):1386–1398. doi:10.1038/s41565-024-01687-1
77. Zhang J, Nie S, Zu Y, et al. Anti-atherogenic effects of CD36-targeted epigallocatechin gallate-loaded nanoparticles. J Control Release. 2019;303:263–273. doi:10.1016/j.jconrel.2019.04.018
78. Kim H, Kumar S, Kang D-W, Jo H, Park J-H. Affinity-driven design of cargo-switching nanoparticles to leverage a cholesterol-rich microenvironment for atherosclerosis therapy. ACS Nano. 2020;14(6):6519–6531. doi:10.1021/acsnano.9b08216
79. Chen Z, Zhu Q, Li D, et al. Targeting nanoplatform for atherosclerosis inhibition and degradation via a dual-track reverse cholesterol transport strategy. Small. 2024;20(7):e2306457. doi:10.1002/smll.202306457
80. He H, Wang J, Yannie PJ, Korzun WJ, Yang H, Ghosh S. Nanoparticle-based “Two-pronged” approach to regress atherosclerosis by simultaneous modulation of cholesterol influx and efflux. Biomaterials. 2020;260:120333. doi:10.1016/j.biomaterials.2020.120333
81. Rakshit M, Darwitan A, Muktabar A, et al. Anti-inflammatory potential of simvastatin loaded nanoliposomes in 2D and 3D foam cell models. Nanomedicine. 2021;37:102434. doi:10.1016/j.nano.2021.102434
82. Ma B, Xu H, Zhuang W, Wang Y, Li G, Wang Y. ROS responsive nanoplatform with two-photon AIE imaging for atherosclerosis diagnosis and “two-pronged” therapy. Small. 2020;16(45):e2003253. doi:10.1002/smll.202003253
83. Ma B, Xiao Y, Lv Q, Li G, Wang Y, Fu G. Targeting theranostics of atherosclerosis by dual-responsive nanoplatform via photoacoustic imaging and three-in-one integrated lipid management. Adv Mater. 2023;35(5):e2206129. doi:10.1002/adma.202206129
84. Pillai SC, Borah A, MNT L, Kawano H, Hasegawa K, Kumar DS. Co-delivery of curcumin and bioperine via PLGA nanoparticles to prevent atherosclerotic foam cell formation. Pharmaceutics. 2021;13(9). doi:10.3390/pharmaceutics13091420
85. Chong SY, Wang X, van Bloois L, et al. Injectable liposomal docosahexaenoic acid alleviates atherosclerosis progression and enhances plaque stability. J Control Release. 2023;360:344–364. doi:10.1016/j.jconrel.2023.06.035
86. Hu R, Dai C, Dong C, et al. Living macrophage-delivered tetrapod pdh nanoenzyme for targeted atherosclerosis management by ROS scavenging, hydrogen anti-inflammation, and autophagy activation. ACS Nano. 2022;16(10):15959–15976. doi:10.1021/acsnano.2c03422
87. Luo Y, Guo Y, Wang H, et al. Phospholipid nanoparticles: therapeutic potentials against atherosclerosis via reducing cholesterol crystals and inhibiting inflammation. eBioMedicine. 2021;74. doi:10.1016/j.ebiom.2021.103725
88. Zha D, Wang S, Monaghan-Nichols P, Qian Y, Sampath V, Fu M. Mechanisms of endothelial cell membrane repair: progress and perspectives. Cells. 2023;12(22). doi:10.3390/cells12222648
89. Ammendolia DA, Bement WM, Brumell JH. Plasma membrane integrity: implications for health and disease. BMC Biol. 2021;19(1):71. doi:10.1186/s12915-021-00972-y
90. Thangasparan S, Kamisah Y, Ugusman A, Mohamad Anuar NN, Ibrahim N. Unravelling the mechanisms of oxidised low-density lipoprotein in cardiovascular health: current evidence from in vitro and in vivo studies. Int J Mol Sci. 2024;25(24). doi:10.3390/ijms252413292
91. Wadey RM, Connolly KD, Mathew D, Walters G, Rees DA, James PE. Inflammatory adipocyte-derived extracellular vesicles promote leukocyte attachment to vascular endothelial cells. Atherosclerosis. 2019;283:19–27. doi:10.1016/j.atherosclerosis.2019.01.013
92. Alvandi Z, Bischoff J. Endothelial-mesenchymal transition in cardiovascular disease. Arterioscler Thromb Vasc Biol. 2021;41(9):2357–2369. doi:10.1161/atvbaha.121.313788
93. Xiao X, Xu M, Yu H, et al. Mesenchymal stem cell-derived small extracellular vesicles mitigate oxidative stress-induced senescence in endothelial cells via regulation of miR-146a/Src. Signal Transduct Target Ther. 2021;6(1):354. doi:10.1038/s41392-021-00765-3
94. Medrano-Bosch M, Simón-Codina B, Jiménez W, Edelman ER, Melgar-Lesmes P. Monocyte-endothelial cell interactions in vascular and tissue remodeling. Front Immunol. 2023;14:1196033. doi:10.3389/fimmu.2023.1196033
95. Hu X, Zhao P, Lu Y, Liu Y. ROS-based nanoparticles for atherosclerosis treatment. Materials. 2021;14(22). doi:10.3390/ma14226921
96. Barrett TJ. Macrophages in atherosclerosis regression. Arterioscler Thromb Vasc Biol. 2020;40(1):20–33. doi:10.1161/atvbaha.119.312802
97. Luo M, Zhao F, Cheng H, Su M, Wang Y. Macrophage polarization: an important role in inflammatory diseases. Front Immunol. 2024;15:1352946. doi:10.3389/fimmu.2024.1352946
98. Vassiliou E, Farias-Pereira R. Impact of lipid metabolism on macrophage polarization: implications for inflammation and tumor immunity. Int J Mol Sci. 2023;24(15). doi:10.3390/ijms241512032
99. Qiao Y, Wang C, Kou J, et al. MicroRNA-23a suppresses the apoptosis of inflammatory macrophages and foam cells in atherogenesis by targeting HSP90. Gene. 2020;729:144319. doi:10.1016/j.gene.2019.144319
100. Wang Y, Shi R, Zhai R, et al. Matrix stiffness regulates macrophage polarization in atherosclerosis. Pharmacol Res. 2022;179:106236. doi:10.1016/j.phrs.2022.106236
101. Tsourouktsoglou TD, Warnatsch A, Ioannou M, Hoving D, Wang Q, Papayannopoulos V. Histones, DNA, and citrullination promote neutrophil extracellular trap inflammation by regulating the localization and activation of TLR4. Cell Rep. 2020;31(5):107602. doi:10.1016/j.celrep.2020.107602
102. Döring Y, Soehnlein O, Weber C. Neutrophil extracellular traps in atherosclerosis and atherothrombosis. Circ Res. 2017;120(4):736–743. doi:10.1161/circresaha.116.309692
103. Hofbauer TM, Ondracek AS, Lang IM. Neutrophil Extracellular Traps in Atherosclerosis and Thrombosis. In: von Eckardstein A, Binder CJ, editors. Prevention and Treatment of Atherosclerosis: Improving State-of-the-Art Management and Search for Novel Targets. Springer Copyright 2020, The Author(s); 2022:405–425.
104. Silvestre-Roig C, Fridlender ZG, Glogauer M, Scapini P. Neutrophil diversity in health and disease. Trends Immunol. 2019;40(7):565–583. doi:10.1016/j.it.2019.04.012
105. Warnatsch A, Ioannou M, Wang Q, Papayannopoulos V. Inflammation. Neutrophil extracellular traps license macrophages for cytokine production in atherosclerosis. Science. 2015;349(6245):316–320. doi:10.1126/science.aaa8064
106. Dollery CM, Owen CA, Sukhova GK, Krettek A, Shapiro SD, Libby P. Neutrophil elastase in human atherosclerotic plaques: production by macrophages. Circulation. 2003;107(22):2829–2836. doi:10.1161/01.Cir.0000072792.65250.4a
107. Durham AL, Speer MY, Scatena M, Giachelli CM, Shanahan CM. Role of smooth muscle cells in vascular calcification: implications in atherosclerosis and arterial stiffness. Cardiovasc Res. 2018;114(4):590–600. doi:10.1093/cvr/cvy010
108. Grootaert MOJ, Bennett MR. Vascular smooth muscle cells in atherosclerosis: time for a re-assessment. Cardiovasc Res. 2021;117(11):2326–2339. doi:10.1093/cvr/cvab046
109. Pan H, Xue C, Auerbach BJ, et al. Single-cell genomics reveals a novel cell state during smooth muscle cell phenotypic switching and potential therapeutic targets for atherosclerosis in mouse and human. Circulation. 2020;142(21):2060–2075. doi:10.1161/circulationaha.120.048378
110. Wang C, He Y, Tang J, et al. Chondroitin sulfate functionalized nanozymes inhibit the inflammation feedback loop for enhanced atherosclerosis therapy by regulating intercellular crosstalk. Int J Biol Macromol. 2024;282(Pt 3):136918. doi:10.1016/j.ijbiomac.2024.136918
111. Dai Y, Sha X, Song X, et al. Targeted therapy of atherosclerosis vulnerable plaque by ros-scavenging nanoparticles and MR/Fluorescence dual-modality imaging tracing. Int J Nanomed. 2022;17:5413–5429. doi:10.2147/IJN.S371873
112. Paszek E, Żmudka K, Plens K, Legutko J, Rajs T, Zajdel W. Evaluation of profilin 1 as a biomarker in myocardial infarction. Eur Rev Med Pharmacol Sci. 2020;24(15):8112–8116. doi:10.26355/eurrev_202008_22497
113. Allen A, Gau D, Roy P. The role of profilin-1 in cardiovascular diseases. J Cell Sci. 2021;134(9). doi:10.1242/jcs.249060
114. Wang Y, Dubland JA, Allahverdian S, et al. Smooth muscle cells contribute the majority of foam cells in ApoE (Apolipoprotein E)-deficient mouse atherosclerosis. Arterioscler Thromb Vasc Biol. 2019;39(5):876–887. doi:10.1161/atvbaha.119.312434
115. Döring Y, van der Vorst EPC, Weber C. Targeting immune cell recruitment in atherosclerosis. Nat Rev Cardiol. 2024;21(11):824–840. doi:10.1038/s41569-024-01023-z
116. Qin W, Gan F, Liang R, et al. Identification of monocyte-associated genes related to the instability of atherosclerosis plaque. Oxid Med Cell Longev. 2022;2022:3972272. doi:10.1155/2022/3972272
117. Yu L, Xu L, Chu H, et al. Macrophage-to-endothelial cell crosstalk by the cholesterol metabolite 27HC promotes atherosclerosis in male mice. Nat Commun. 2023;14(1):4101. doi:10.1038/s41467-023-39586-z
118. Chai H, Qu H, He S, et al. Zedoarondiol inhibits atherosclerosis by regulating monocyte migration and adhesion via CXCL12/CXCR4 pathway. Pharmacol Res. 2022;182:106328. doi:10.1016/j.phrs.2022.106328
119. Libby P. Inflammation and the pathogenesis of atherosclerosis. Vascul Pharmacol. 2024;154:107255. doi:10.1016/j.vph.2023.107255
120. Ren X, Manzanares LD, Piccolo EB, et al. Macrophage-endothelial cell crosstalk orchestrates neutrophil recruitment in inflamed mucosa. J Clin Invest. 2023;133(15). doi:10.1172/jci170733
121. Tedgui A, Mallat Z. Cytokines in atherosclerosis: pathogenic and regulatory pathways. Physiol Rev. 2006;86(2):515–581. doi:10.1152/physrev.00024.2005
122. Tyrrell DJ, Goldstein DR. Ageing and atherosclerosis: vascular intrinsic and extrinsic factors and potential role of IL-6. Nat Rev Cardiol. 2021;18(1):58–68. doi:10.1038/s41569-020-0431-7
123. Weber C, Habenicht AJR, Von hundelshausen P. Novel mechanisms and therapeutic targets in atherosclerosis: inflammation and beyond. Eur Heart J. 2023;44(29):2672–2681. doi:10.1093/eurheartj/ehad304
124. Pedro-Botet J, Climent E, Benaiges D. Atherosclerosis and inflammation. New therapeutic approaches. Med Clin. 2020;155(6):256–262.Arteriosclerosis e inflamación. Nuevos enfoques terapéuticos. doi:10.1016/j.medcli.2020.04.024
125. Ji H, Peng R, Jin L, et al. Recent advances in ROS-sensitive nano-formulations for atherosclerosis applications. Pharmaceutics. 2021;13(9). doi:10.3390/pharmaceutics13091452
126. Wu C, Mao J, Wang X, et al. Advances in treatment strategies based on scavenging reactive oxygen species of nanoparticles for atherosclerosis. J Nanobiotechnol. 2023;21(1):271. doi:10.1186/s12951-023-02058-z
127. Wang W, Kang PM. Oxidative stress and antioxidant treatments in cardiovascular diseases. Antioxidants. 2020;9(12). doi:10.3390/antiox9121292
128. Langlois M, Duprez D, Delanghe J, De Buyzere M, Clement DL. Serum vitamin C concentration is low in peripheral arterial disease and is associated with inflammation and severity of atherosclerosis. Circulation. 2001;103(14):1863–1868. doi:10.1161/01.cir.103.14.1863
129. Hodis HN, Mack WJ, LaBree L, et al. Alpha-tocopherol supplementation in healthy individuals reduces low-density lipoprotein oxidation but not atherosclerosis: the Vitamin E Atherosclerosis Prevention Study (VEAPS). Circulation. 2002;106(12):1453–1459. doi:10.1161/01.cir.0000029092.99946.08
130. Wang Q, Duan Y, Jing H, et al. Inhibition of atherosclerosis progression by modular micelles. J Control Release. 2023;354:294–304. doi:10.1016/j.jconrel.2023.01.020
131. Wang Y, Li L, Zhao W, et al. Targeted therapy of atherosclerosis by a broad-spectrum reactive oxygen species scavenging nanoparticle with intrinsic anti-inflammatory activity. ACS Nano. 2018;12(9):8943–8960. doi:10.1021/acsnano.8b02037
132. Kornfeld OS, Hwang S, Disatnik MH, Chen CH, Qvit N, Mochly-Rosen D. Mitochondrial reactive oxygen species at the heart of the matter: new therapeutic approaches for cardiovascular diseases. Circ Res. 2015;116(11):1783–1799. doi:10.1161/circresaha.116.305432
133. Zhou S, Cai H, He X, Tang Z, Lu S. Enzyme-mimetic antioxidant nanomaterials for ROS scavenging: design, classification, and biological applications. Coord Chem Rev. 2024;500:215536. doi:10.1016/j.ccr.2023.215536
134. Wang S, Zhang J, Li W, et al. Hyaluronic acid-guided assembly of ceria nanozymes as plaque-targeting ROS scavengers for anti-atherosclerotic therapy. Carbohydr Polym. 2022;296:119940. doi:10.1016/j.carbpol.2022.119940
135. Jia M, Ren W, Liu Y, et al. Messenger nanozyme for reprogramming the microenvironment of rheumatoid arthritis. ACS Appl Mater Interfaces. 2023;15(1):338–353. doi:10.1021/acsami.2c16458
136. Sun W, Xu Y, Yao Y, et al. Self-oxygenation mesoporous MnO2 nanoparticles with ultra-high drug loading capacity for targeted arteriosclerosis therapy. J Nanobiotechnol. 2022;20(1):88. doi:10.1186/s12951-022-01296-x
137. Xu M, Ran D, Hu J, et al. Multifunctional Prussian blue nanozymes alleviate atherosclerosis through inhibiting the inflammation feedback loop. J Mater Chem B. 2025;13(4):1459–1473. doi:10.1039/d4tb01926a
138. Wu X, Zhang Y, Xing P, Zhu M. Surface functionalization strategies of ROS-scavenging nanozymes for synergistic therapy and efficient delivery. J Mater Chem B. 2025;13(26):7653–7667. doi:10.1039/d5tb00877h
139. Kornmueller K, Vidakovic I, Prassl R. Artificial high density lipoprotein nanoparticles in cardiovascular research. Molecules. 2019;24(15). doi:10.3390/molecules24152829
140. Matsuo M. ABCA1 and ABCG1 as potential therapeutic targets for the prevention of atherosclerosis. J Pharmacol Sci. 2022;148(2):197–203. doi:10.1016/j.jphs.2021.11.005
141. Yang TM, Miao M, Yu WQ, et al. Targeting macrophages in atherosclerosis using nanocarriers loaded with liver X receptor agonists: a narrow review. Front Mol Biosci. 2023;10:1147699. doi:10.3389/fmolb.2023.1147699
142. Zhang XQ, Even-Or O, Xu X, et al. Nanoparticles containing a liver X receptor agonist inhibit inflammation and atherosclerosis. Adv Healthc Mater. 2015;4(2):228–236. doi:10.1002/adhm.201400337
143. Baumer Y, Mehta NN, Dey AK, Powell-Wiley TM, Boisvert WA. Cholesterol crystals and atherosclerosis. Eur Heart J. 2020;41(24):2236–2239. doi:10.1093/eurheartj/ehaa505
144. Chen J, Zhang X, Millican R, Creutzmann JE, Martin S, Jun H-W. High density lipoprotein mimicking nanoparticles for atherosclerosis. Nano Convergence. 2020;7(1). doi:10.1186/s40580-019-0214-1
145. Doran AC. Inflammation resolution: implications for atherosclerosis. Circ Res. 2022;130(1):130–148. doi:10.1161/circresaha.121.319822
146. Ait-Oufella H, Libby P. Inflammation and atherosclerosis: prospects for clinical trials. Arterioscler Thromb Vasc Biol. 2024;44(9):1899–1905. doi:10.1161/atvbaha.124.320155
147. Biros E, Reznik JE, Moran CS. Role of inflammatory cytokines in genesis and treatment of atherosclerosis. Trends Cardiovasc Med. 2022;32(3):138–142. doi:10.1016/j.tcm.2021.02.001
148. Fang F, Ni Y, Yu H, et al. Inflammatory endothelium-targeted and cathepsin responsive nanoparticles are effective against atherosclerosis. Theranostics. 2022;12(9):4200–4220. doi:10.7150/thno.70896
149. Deroissart J, Porsch F, Koller T, Binder CJ. Anti-inflammatory and Immunomodulatory Therapies in Atherosclerosis. Handb Exp Pharmacol. 2022;270:359–404. doi:10.1007/164_2021_505
150. Laurindo LF, de Carvalho GM, de Oliveira Zanuso B, et al. Curcumin-based nanomedicines in the treatment of inflammatory and immunomodulated diseases: an evidence-based comprehensive review. Pharmaceutics. 2023;15(1). doi:10.3390/pharmaceutics15010229
151. Lv F, Fang H, Huang L, et al. Curcumin equipped nanozyme-like metal-organic framework platform for the targeted atherosclerosis treatment with lipid regulation and enhanced magnetic resonance imaging capability. Adv Sci. 2024;11(26):e2309062. doi:10.1002/advs.202309062
152. Xing Y, Lin X. Challenges and advances in the management of inflammation in atherosclerosis. J Adv Res. 2024. doi:10.1016/j.jare.2024.06.016
153. Lin L, Chen L, Yan J, et al. Advances of nanoparticle-mediated diagnostic and theranostic strategies for atherosclerosis. Front Bioeng Biotechnol. 2023;11:1268428. doi:10.3389/fbioe.2023.1268428
154. Perrotta I. Atherosclerosis: from molecular biology to therapeutic perspective 2.0. Int J Mol Sci. 2022;23(23). doi:10.3390/ijms232315158
155. Wilczewska AZ, Niemirowicz K, Markiewicz KH, Car H. Nanoparticles as drug delivery systems. Pharmacol Rep. 2012;64(5):1020–1037. doi:10.1016/s1734-1140(12)70901-5
156. Shoaran M, Maffia P. Tackling inflammation in atherosclerosis. Nat Rev Cardiol. 2024;21(7):442. doi:10.1038/s41569-024-01007-z
157. Xu M, Qi Y, Liu G, Song Y, Jiang X, Du B. Size-dependent in vivo transport of nanoparticles: implications for delivery, targeting, and clearance. ACS Nano. 2023;17(21):20825–20849. doi:10.1021/acsnano.3c05853
158. Chen Y, De Koker S, De Geest BG. Engineering strategies for lymph node targeted immune activation. Acc Chem Res. 2020;53(10):2055–2067. doi:10.1021/acs.accounts.0c00260
159. Zhang S, Liu Y, Cao Y, et al. Targeting the microenvironment of vulnerable atherosclerotic plaques: an emerging diagnosis and therapy strategy for atherosclerosis. Adv Mater. 2022;34(29):e2110660. doi:10.1002/adma.202110660
160. Najahi-Missaoui W, Arnold RD, Cummings BS. Safe nanoparticles: are we there yet? Int J Mol Sci. 2020;22(1). doi:10.3390/ijms22010385
161. Mao Y, Ren J, Yang L. Advances of nanomedicine in treatment of atherosclerosis and thrombosis. Environ Res. 2023;238(Pt 2):116637. doi:10.1016/j.envres.2023.116637
162. Reddy ARN, Lonkala S. In vitro evaluation of copper oxide nanoparticle-induced cytotoxicity and oxidative stress using human embryonic kidney cells. Toxicol Ind Health. 2019;35(2):159–164. doi:10.1177/0748233718819371
163. Shreffler JW, Pullan JE, Dailey KM, Mallik S, Brooks AE. Overcoming hurdles in nanoparticle clinical translation: the influence of experimental design and surface modification. Int J Mol Sci. 2019;20(23). doi:10.3390/ijms20236056
164. Yang X, Hu J, Gao Q, et al. Advances in nano-delivery systems based on diagnosis and theranostics strategy for atherosclerosis. J Drug Target. 2024:1–16. doi:10.1080/1061186x.2024.2433560.
165. Li X, Peng X, Zoulikha M, et al. Multifunctional nanoparticle-mediated combining therapy for human diseases. Signal Transduct Target Ther. 2024;9(1):1. doi:10.1038/s41392-023-01668-1
166. Li L, Wang M, Ma Q, Ye J, Sun G. Role of glycolysis in the development of atherosclerosis. Am J Physiol Cell Physiol. 2022;323(2):C617–c629. doi:10.1152/ajpcell.00218.2022
167. Wang R, Wang M, Ye J, Sun G, Sun X. Mechanism overview and target mining of atherosclerosis: endothelial cell injury in atherosclerosis is regulated by glycolysis (Review). Int J Mol Med. 2021;47(1):65–76. doi:10.3892/ijmm.2020.4798
168. Li L, Mou J, Han Y, et al. Calenduloside e modulates macrophage polarization via KLF2-regulated glycolysis, contributing to attenuates atherosclerosis. Int Immunopharmacol. 2023;117:109730. doi:10.1016/j.intimp.2023.109730
169. Zhang Z, Liang X, Yang X, Liu Y, Zhou X, Li C. Advances in nanodelivery systems based on metabolism reprogramming strategies for enhanced tumor therapy. ACS Appl Mater Interfaces. 2024;16(6):6689–6708. doi:10.1021/acsami.3c15686
170. Johnson JL. Metalloproteinases in atherosclerosis. Eur J Pharmacol. 2017;816:93–106. doi:10.1016/j.ejphar.2017.09.007
171. Dhayni K, Zibara K, Issa H, Kamel S, Bennis Y. Targeting CXCR1 and CXCR2 receptors in cardiovascular diseases. Pharmacol Ther. 2022;237:108257. doi:10.1016/j.pharmthera.2022.108257
172. Liu Y, Zhou M, Xu M, et al. Reprogramming monocytes into M2 macrophages as living drug depots to enhance treatment of myocardial ischemia-reperfusion injury. J Control Release. 2024;374:639–652. doi:10.1016/j.jconrel.2024.08.045
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.