Back to Journals » Drug Design, Development and Therapy » Volume 19
PSORI-CM02 Restores Epidermal Differentiation in Psoriasis via the Gut Microbiota–Sphingolipid Axis
Authors Wu Y, Pan S, Yin C, Kong Y, Huo W, Wang Q, Wu J, Li L, Wei J, Lu C, Han L, Lu Y
Received 9 May 2025
Accepted for publication 20 September 2025
Published 7 November 2025 Volume 2025:19 Pages 9993—10010
DOI https://doi.org/10.2147/DDDT.S539436
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Anastasios Lymperopoulos
Yuan Wu,1,2,* Simin Pan,2,* Chubo Yin,2 Yunqi Kong,2 Wanhua Huo,2 Qiuyue Wang,2 Jingjing Wu,1– 3 Li Li,1– 3 Jianan Wei,1– 3 Chuanjian Lu,1– 5 Ling Han,1– 5 Yue Lu1– 5
1The Second Affiliated Hospital of Guangzhou University of Chinese Medicine (Guangdong Provincial Hospital of Chinese Medicine), Guangzhou, People’s Republic of China; 2The Second Clinical Medical College, Guangzhou University of Chinese Medicine, Guangzhou, People’s Republic of China; 3Guangdong-Hong Kong-Macau Joint Laboratory on Chinese Medicine and Immune Disease Research, The Second Affiliated Hospital of Guangzhou University of Chinese Medicine, Guangzhou, People’s Republic of China; 4Guangdong Provincial Key Laboratory of Clinical Research on Traditional Chinese Medicine Syndrome, The Second Affiliated Hospital of Guangzhou University of Chinese Medicine, Guangzhou, People’s Republic of China; 5State Key Laboratory of Dampness Syndrome of Chinese Medicine, The Second Affiliated Hospital of Guangzhou University of Chinese Medicine, Guangzhou, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Ling Han, Email [email protected] Yue Lu, Email [email protected]
Background: Psoriasis is linked to gut dysbiosis and disturbed sphingolipid metabolism. PSORI-CM02 improves epidermal differentiation, yet its impact on the microbiota–sphingolipid axis remains unknown.
Methods: Transcriptomics of patient keratinocytes, Carmofur inhibition in IMQ mice, and multi-omics (metabolomics, metagenomics) of skin, lymph nodes and gut were combined. SPF, PGF and GF mice underwent FMT to test microbiota dependency.
Results: Psoriatic lesions showed sphingolipid pathway enrichment. Carmofur enhanced differentiation. PSORI-CM02 lowered PASI, spleen index, and tissue levels of ceramide, S1P, C1P and sphingomyelin while restoring Flg, Krt10 and Krt14. It reduced Turicibacter, Bacteroides, Bifidobacterium and Acetobacter. PSORI-CM02-derived microbiota reproduced therapeutic effects in all FMT settings.
Conclusion: PSORI-CM02 reshapes gut microbiota, normalizes sphingolipid metabolism and improves epidermal differentiation to treat psoriasis.
Keywords: psoriasis, PSORI-CM02, sphingolipid regulation, fecal microbiome transplantation, germ-free
Introduction
Psoriasis is a chronic, polygenic inflammatory dermatosis driven by keratinocyte hyperproliferation and defective differentiation, producing erythematous, scaly plaques that affect 2–3% of the global population.1,2 The use of immunosuppressants, hormones, and biological drugs to alleviate the clinical symptoms of psoriasis may lead to drug withdrawal or side effects because of the lifelong nature of the disease.3,4 Furthermore, biological therapy is often designated for severe cases of psoriasis due to its substantial expense and may not be viable for prolonged therapy.
The gut microbiota, comprising trillions of microorganisms, modulates systemic immunity and skin homeostasis.5 Psoriatic patients exhibit compositional and functional dysbiosis,6,7 and case reports demonstrate that fecal microbiota transplantation (FMT) can improve skin lesions without adverse events.8 Yet the mechanistic pathways linking gut microbes to psoriasis remain undefined. Recent evidence implicates perturbed sphingolipid metabolism—centered on ceramide and its bioactive metabolites S1P, C1P and sphingomyelin—in barrier dysfunction, inflammation and keratinocyte hyperproliferation.9,10 Although mammalian sphingolipid turnover has been well characterized, microbial sphingolipid biosynthesis by genera such as Bacteroides, Bifidobacterium, Turicibacter and Acetobacter is only beginning to be recognized.11,12 The contribution of these microbial sphingolipids to psoriatic pathophysiology remains largely unexplored.12
Although the significance of sphingolipids turnover in mammals for regulating diverse cellular functions is widely acknowledged, the metabolic pathways of microbial sphingolipids remain poorly comprehended. Sphingolipid production has been described in Bacteroides, Bifidobacterium, Turicibacter and Acetobacter.13–15 The intestinal microbiota constitutes an exceedingly intricate ecological system with a substantial capacity to affect the well-being of the host; however, the magnitude of the influence that sphingolipids exert on the pathophysiological mechanisms in psoriasis has yet to be completely elucidated.
PSORI-CM02, derived from the Yinxieling tablet created by national medical master Xuan Guowei, consists of Radix Paeoniae Rubra, Rhizoma Curcumas, Saracandra Glabra, and Rhizoma Smilacis Glabrae. It has found extensive application in clinical settings for alleviating manifestations and reducing the reappearance in patients with psoriasis, with few side effects.16,17 A previous study demonstrated that PSORI-CM02 effectively ameliorated psoriasis-like scales induced by imiquimod (IMQ) in mice by regulating keratinocyte apoptosis.18 At present, no existing studies have verified whether PSORI-CM02 manages psoriasis by modulating the “microbiota-sphingolipid metabolism” axis, thereby potentially impacting keratinocyte differentiation. The objective of this research is to delineate the mechanisms that connect sphingolipid catabolism with epidermal stratification and to assess the regulatory impact of PSORI-CM02 on the intestinal microbiome and sphingolipid metabolism. This will be accomplished by examining the skin-gut axis through a multi-omics analytical approach and conducting fecal microbiota transplantation studies.
Methods and Materials
Tissue Sample Collection and Transcriptomic Analysis
The study included nine psoriasis patients from the Second Affiliated Hospital of Guangzhou University of Chinese Medicine, as well as nine healthy control subjects. This study was approved by the Ethics Committee of the Second Affiliated Hospital of Guangzhou University of Chinese Medicine (B2013-132-01). Baseline information is provided in the Supplementary Table 1. Skin tissues were collected, and keratinocytes were isolated and cultured in vitro, followed by transcriptomic analysis.
Following the isolation of RNA samples, evaluation of their purity, accurate measurement, and confirmation of RNA integrity, the process of transcriptome sequencing was initiated with the assembly of libraries and a subsequent quality check. Upon successful quality assessment of the libraries, they were consolidated based on their effective concentrations and the desired sequencing data yield, prior to undergoing Illumina PE150 sequencing. The subsequent analytical steps encompassed alignment to a reference genome, quantification of gene expression, analysis of differential gene expression, and functional enrichment analysis. Genes were considered to be differentially expressed if the Log2|fold change| was greater than or equal to 2 and the adjusted P value was less than 0.05.
PSORI-CM02 Formulation
The PSORI-CM02 formulation was obtained from the Second Affiliated Hospital of Guangzhou University of Chinese Medicine. It comprises five components: Smilax glabra Roxb., Sarcandra glabra (Thunb). Nakai, radix of Paeonialactiflora Pall, radix of Curcuma phaeocaulis Val., and Prunus mume (Sieh). Sieb. et Zucc., with a weight ratio of 5:5:3:2:2. Standard procedures were followed for the preparation of all herbal decoctions,19 adhering to the guidelines outlined in the 2010 Edition of the China Pharmacopoeia. In our previous study, ultra-high-performance liquid chromatography/mass spectrometry analysis was used for quality assurance. Fourteen small-molecule compounds, gallic acid, 5-hydroxymethylfurfural, astilbin, paeoniflorin, and zederone, were successfully identified.19
Grouping and Administration of Animals
Male BALB/c mice (18–22 g) were procured from the Animal Center of Guangdong Medicine Science. Before experimentation, the mice were acclimatized in a specific pathogen-free environment. Mice had unrestricted access to food and water throughout the study. All animal procedures were approved by the Animal Ethics Committee of the Second Affiliated Hospital of Guangzhou University of Chinese Medicine (Number 2021054), and were conducted in accordance with the guidelines of the Institutional Animal Care and Use Committee regarding Animal Welfare and Ethics.
The skin of IMQ-treated mice demonstrated typical psoriasis-like characteristics, including epidermal hyperplasia, immunocyte infiltration, and transformed microcirculation. Therefore, we applied IMQ to mouse skin to mimic psoriatic pathophysiology. IMQ was consecutively applied to the shaved backs of the mice for 7 days.
To investigate the relationship between sphingolipid metabolism and epidermal differentiation (Experiment 1), we applied Carmofur, inhibitor of an acid ceramidase that is key involved in the breakdown of ceramides20 to block the sphingolipid metabolic pathway. Male BALB/c mice were randomly allocated into three groups (n = 8 per group): control, model, and model + carmofur (MCE, NJ, USA, HY-B0182; 43 mg/kg/day,21 intraperitoneal injection for 7 days). The exposed skin was topically treated with 5% imiquimod cream (IMQ; Sichuan Mingxin, Chengdu, China) at a dosage of 62.5 mg per day for 7 days. Mice in the control group received Vaseline in equivalent amounts periodically. Detailed grouping and administration were shown in Figure 1A.
 |
Figure 1 Study design for the whole experiment. (A) Investigating how sphingolipid metabolism regulates epidermal differentiation. (B) The impact of PSORI-CM02 on epidermal differentiation, sphingolipid metabolism, and microbial community structure in psoriasis. (C) Through fecal microbiota transplantation experiments, to verify whether PSORI-CM02 improves epidermal differentiation in psoriasis by restoring microbial community structure and regulating sphingolipid metabolism. Abbreviations: SPF, Specific pathogen-free; IMQ, imiquimod; MTX, methotrexate; PSORI-CM02_M, PSORI-CM02 at medium dose (1.42 g/kg); PSORI-CM02_H, PSORI-CM02 at high dose (2.84 g/kg); PGF, pseudo germ-free; GF, germ-free; FMT, fecal microbiota transplantation; ABX, antibiotics. |
To demonstrate the regulatory effects of PSORI-CM02 on sphingolipid metabolism and the gut microbiota (Experiment 2), the animals were randomly allocated into five groups (n = 8 per group): control, model, methotrexate (MTX, 0.62 mg/kg; Shanghai Pharmaceutical Group Co. Ltd., Shanghai, China), medium-dose PSORI-CM02 (PSORI-CM02_M, 1.42 g/kg), and high-dose PSORI-CM02 (PSORI-CM02_H, 2.84 g/kg). MTX and both PSORI-CM02 preparations were administered by intragastric gavage at a standardized volume of 0.1 mL/10 g body weight. Control and model groups received an equivalent volume of saline (0.1 mL/10 g) by the same route and schedule. On day 7, feces were collected for metagenomic and metabolomic analyses, and on day 8, the back skin tissue, lymph nodes, and spleen were collected for further analysis. Detailed grouping and administration were shown in Figure 1B. Fresh feces in model and high-dose PSORI-CM02_H group were collected for subsequent FMT.
The efficacy of fecal microbiota transplantation in treating psoriasis was assessed utilizing specific-pathogen-free (SPF), pseudo germ-free (PGF), and germ-free (GF) BALB/c mice models (Figure 1C, Experiment 3). GF mice (produced by Jingtuo Biotechnology Co., Ltd) were strictly fed in a sterile environment. PGF mice were pretreated with antibiotics (ABX) (50mg/kg Vancomycin, 100mg/kg neomycin sulfate, 100mg/kg metronidazole, and 100mg/kg ampicillin) for a week. The SPF, PGF, and GF mice were divided into three groups respectively (n = 8 per group): control, model, and PSORI-CM02-FMT (received feces from PSORI-CM02_H group, 200 uL per day for 7 days) groups. On day 8, the back skin tissue, feces were collected for further analysis.
Real-Time Quantitative Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
Total RNA from skin tissues was extracted using TRIzol® Reagent (Life Technologies, Waltham, MA, USA, 15596026). Subsequently, DNA contamination was removed, and RNA was reverse-transcribed into cDNA using the Color Reverse Transcription Kit (EZBioscience, CA, USA, A0010CGQ).
Finally, the cDNA was combined with primers and 2×SYBR Green qPCR Master Mix (EZBioscience, CA, USA, A0001) and amplified using a PCR amplifier (Thermo Fisher Scientific, ViiA7, Waltham, MA, USA). The primer sequences are listed in Supplementary Table 2.
Histopathology Analysis
A segment of skin was preserved in 4% paraformaldehyde and sliced to a thickness of 4 μm. The slices were stained with hematoxylin and eosin (HE). Histopathological alterations were evaluated under a microscope.
Immunohistochemistry Analysis
For immunohistochemistry, paraffin slices were deparaffinized and hydrated. The sections were incubated with an anti-cytokeratin 14 antibody (Abcam, Cambridge, MA, USA, ab181595), and a secondary antibody. Detection was performed microscopically using the DAB reagent. Immunohistochemical intensities were quantified using the ImageJ software.
Detection of Sphingolipids in Skin Tissue
Sphingolipids production in tissues were quantified using an enzyme-linked immunosorbent assay (ELISA) kit (Jiangsu Meimian Industrial Co., Ltd, Jiangsu, China; Ceramide, MM-44342M1; S1P, MM-44778M1; SM, MM-45646M1; C1P, MM-927247O1) following the manufacturer’s instructions.
Metabolic Analysis of Skin, Lymph Nodes, and Fecal Sample
The metabolomic analysis was conducted using a Vanquish UHPLC system (Thermo Fisher Scientific, Waltham, MA, USA). Principal component analysis (PCA) was used to determine the metabolomic profile of each group. Partial least squares discriminant analysis (PLS-DA) was used to establish a model between metabolic product intensity and classification. Orthogonal partial least-squares discrimination analysis (OPLS-DA) was used to investigate discrepancies among the groups and identify discrepant metabolic products associated with IMQ-stimulated psoriasis.
Gut Microbiota Analysis
Metagenomic analysis was applied in Experiment 2. DNA obtained from the feces was used for metagenomic analysis. To investigate species diversity, α-diversity was assessed using 3 indicators: Shannon, Simpson, and Invsimpson; β-diversity was used to detect differences in species variety among samples and was determined by PCA or principal coordinates analysis (PCoA) analysis. Functional prediction of the gut microbiota was performed using the Kyoto Encyclopedia of Genes and Genomes (KEGG) database and the DIAMOND software. Differences in relative abundance of gut microbiota between groups at the genus level were evaluated using linear discriminant analysis effect size (LEfSe) with LEfSe software (LDA score >3.0, p<0.05).
16S rDNA amplicon sequencing was applied in Experiment 3 and utilized the Illumina NovaSeq 6000 sequencing platform to amplify and sequence the V3-V4 variable regions. Analysis of the reads includes assembly, filtering, noise reduction, species annotation, and abundance analysis, which can reveal the microbial composition of the samples, identify differences between samples, and screen for differential microbial communities.
Statistical Analyses
All data were subjected to statistical analysis using SPSS software (version 26.0; IBM, Armonk, NY, USA) and visualized using GraphPad Prism software (version 9.5; San Diego, CA, USA). First, normality tests were performed on all data. If the data were not normally distributed, nonparametric tests were performed. Conversely, if the data were normally distributed, significant differences between groups were assessed using the Kruskal–Wallis test or analysis of variance (ANOVA). P<0.05 indicates a significant difference.
Results
Sphingolipid Metabolism Modulates Epidermal Differentiation in Psoriasis
Skin tissues from nine patients and healthy control were collected respectively, and keratinocytes were isolated and cultured in vitro. Transcriptomic analysis indicated that there were 3808 differential expressed genes, with 2455 down-regulated and 1353 up-regulated genes (Figure 2A). Functional analysis indicated that differential expressed genes were enriched in epidermal cell differentiation and sphingolipid biosynthesis process (Figure 2B).
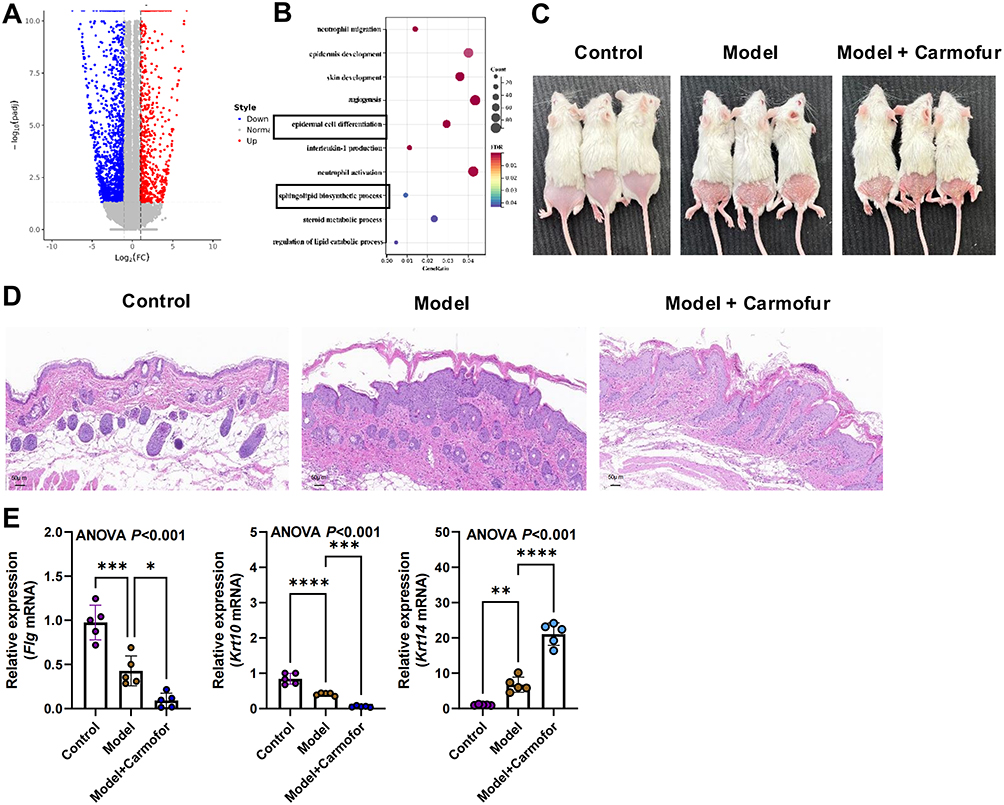 |
Figure 2 Sphingolipid metabolism regulates epidermal differentiation in psoriasis. (A) Volcano plot of the differential expressed genes between psoriasis patients and healthy control. (B) Functional enrichment analysis of the differential expressed genes. (C) Skin images in three groups. (D) HE staining (magnification ×200, scale bar = 50 μm). (E) Relative expression of Flg, Krt 10, and Krt 14 in skin tissue detected by RT-PCR (biological replicates n = 5 per group). Data are presented as mean ± standard deviation. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, ns, not significant, compared with the model group. Abbreviations: HE, hematoxylin and eosin; ANOVA, analysis of variance; ELISA, enzyme-linked immunosorbent assay; RT-PCR, Real-time quantitative reverse transcription-polymerase chain reaction. |
Since ceramide is a key metabolite in sphingolipid metabolism, this study applied Carmofur (the inhibitor of an enzyme that breakdown ceramide) to psoriasis mice to further investigate the relationship between the sphingolipid metabolic pathway and epidermal differentiation in psoriasis. Figure 2C illustrates the skin lesions of mice in each group, showing psoriasis-like symptoms such as scaling, erythema, and thickening on the dorsal skin of mice after topical application of imiquimod and intraperitoneal injection of Carmofur. The HE pathology results (Figure 2D) indicate that epidermal thickening was more pronounced in the skin of mice treated with Carmofur compared to those treated with imiquimod alone. Furthermore, RT-PCR analysis of epidermal differentiation-related molecules showed that Carmofur intervention promotes the expression of Krt14 while inhibiting the mRNA expression of Flg and Krt10 (Figure 2E), suggesting that inhibition of sphingolipid metabolic pathway can promote epidermal differentiation.
PSORI-CM02 Improves Epidermal Differentiation in Psoriasis Mice
We conducted further research to evaluate the therapeutic efficacy of PSORI-CM02 in the treatment of psoriasis. Compared to the control group, IMQ stimulation resulted in noticeable erythema and scaling of the skin, and these physical changes were significantly improved in the PSORI-CM02- and MTX-treated groups (Figure 3). The psoriatic area and severity index (PASI) scores were calculated by summing the three individual scores (range 0–12). PSORI-CM02-treated groups showed a greater overall improvement compared to the MTX group when compared to the model group (Figure 4A). HE staining of the model group skin revealed epidermal thickening, infiltration of inflammatory cells within the stratum corneum, a reduced number of cells in the granular layer, irregularly shaped rete ridges, and dilated capillaries in the dermal papillae, indicating excessive proliferation and abnormal differentiation of the epidermis. However, the typical pathological alterations in the PSORI-CM02-treated groups were notably alleviated compared to those in the model group (Figure 3, Figure 4B). Collectively, these results suggested that PSORI-CM02 efficiently alleviated IMQ-induced psoriasis-like lesions. Additionally, after IMQ administration, the spleen of mice in the model group significantly increased in size, whereas pretreatment with PSORI-CM02_H (high dose) significantly decreased the spleen index (Figure 4C and D). Immunohistochemical analysis revealed that IMQ treatment induced an elevated level of cytokeratin 14 in the skin tissue, whereas the PSORI-CM02- and MTX-treated groups exhibited significantly reduced cytokeratin 14 expression, indicating an improvement in cell differentiation (Figure 4E). RT-PCR analysis of mRNA expression for epidermal differentiation-related molecules showed that in the model group, Flg and Krt10 expression significantly decreased, while Krt14 expression significantly increased. Treatment with PSORI-CM02 reversed these trends (Figure 4F). These findings suggest that PSORI-CM02 improves epidermal differentiation in psoriasis.
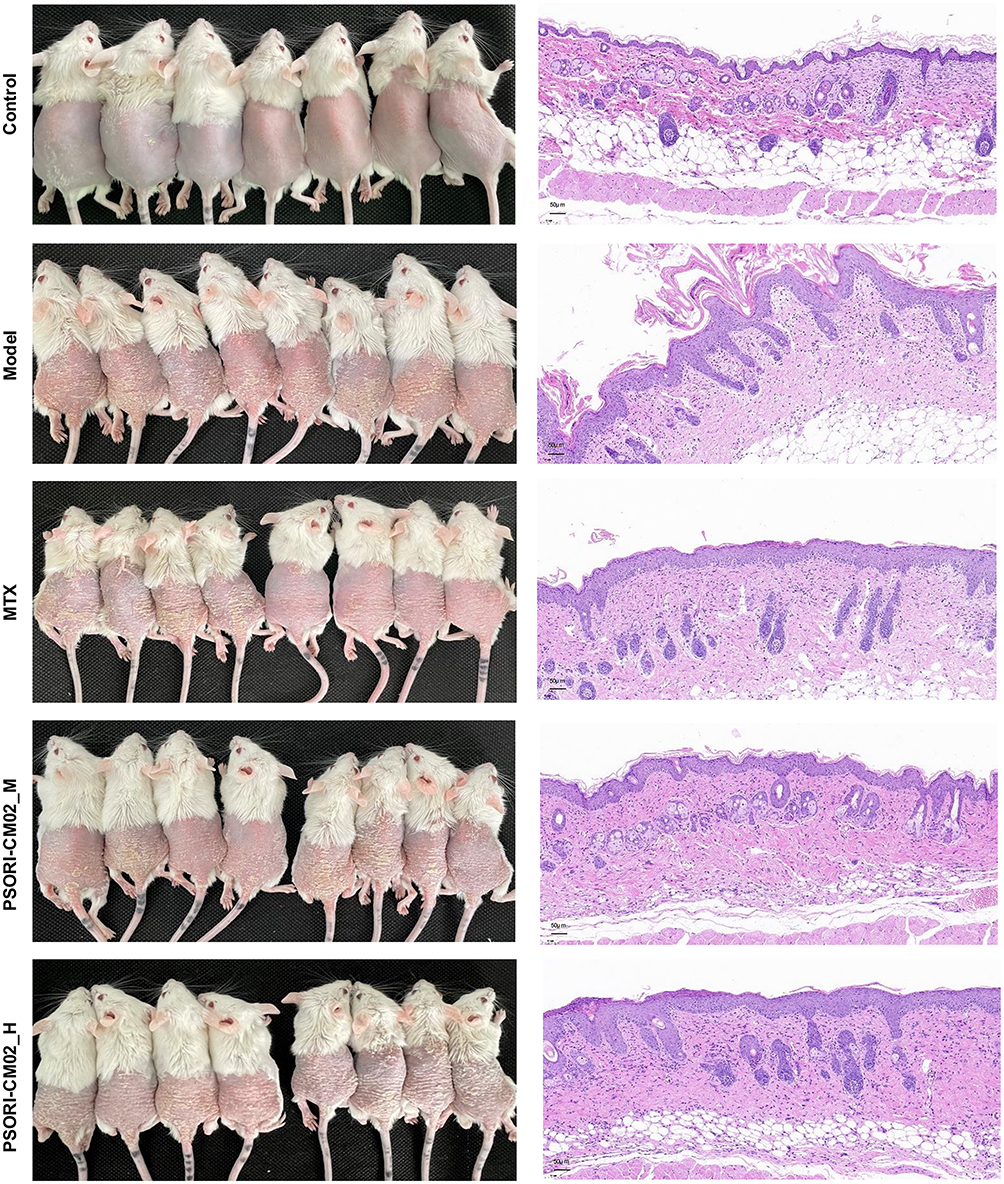 |
Figure 3 Skin images (n = 8 per group) and HE staining (magnification ×200, scale bar = 50 μm). |
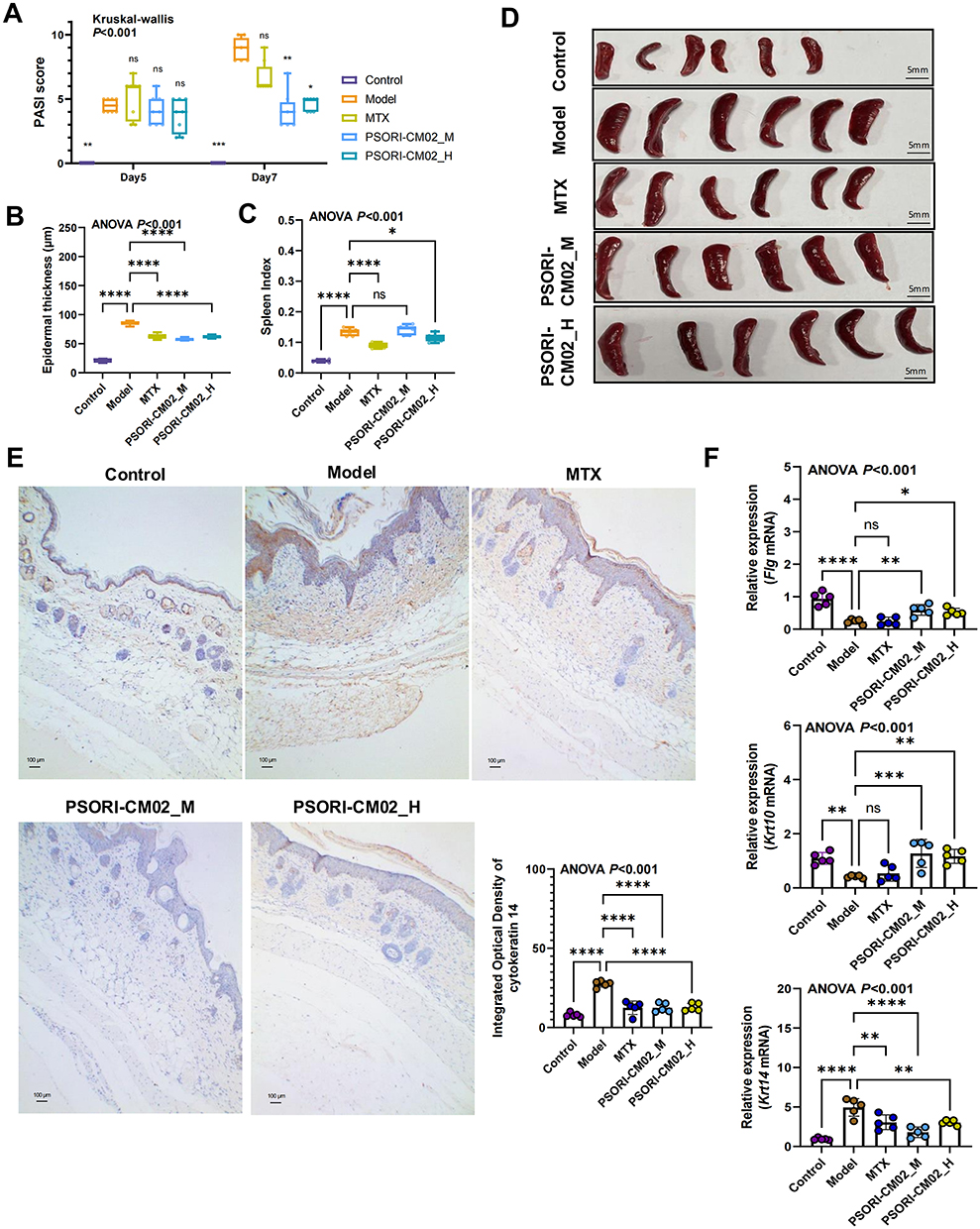 |
Figure 4 PSORI-CM02 improves epidermal differentiation in psoriasis mice. (A) PASI scores for each group (n = 8 per group). (B) Epidermal thickness for each group (n = 5 per group). (C) Spleen index for each group (n = 6 per group). (D) Spleen imagery for each group (n = 6 per group, scale bar = 50 mm). (E) Immunohistochemical examination of cytokeratin 14 (magnification ×100, scale bar = 100 μm). (F) Relative expression of Flg, Krt 10, and Krt 14 in skin tissue detected by RT-PCR (biological replicates n = 5 per group). Data are presented as mean ± standard deviation. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, ns, not significant, compared with the model group. Abbreviations: HE, hematoxylin and eosin; ANOVA, analysis of variance; RT-PCR, Real-time quantitative reverse transcription-polymerase chain reaction; MTX, methotrexate; PSORI-CM02_M, PSORI-CM02 at medium dose (1.42 g/kg); PSORI-CM02_H, PSORI-CM02 at high dose (2.84 g/kg); PASI, psoriatic area and severity index. |
For further metagenomic and metabolomic analyses, PSORI-CM02_M and PSORI-CM02_H were integrated into one PSORI-CM02 group to investigate the underlying mechanisms by which PSORI-CM02 acts on metabolites and the microbiome.
Multi-Tissue Metabolomics Elucidates the Regulatory Effects of PSORI-CM02 on Sphingolipid Metabolism
To elucidate the sphingolipid metabolism in psoriatic and PSORI-CM02-treated mice, the untargeted metabolic analysis of the feces, skins and lymph nodes were performed. In fecal metabolomics, the PCA score plot showed distinct metabolites in the three groups (Supplementary Figure 1A). The PLS-DA and OPLS-DA models further confirmed the separation among all the groups (Supplementary Figure 1B and C). Permutation tests were conducted to verify the suitability of the PLS-DA and OPLS-DA models, and R2 and Q2 were used to assess the fitting and prediction ability. The results indicated that the model did not overfit. Six differential expressed sphingolipids were identified, among which three, including N-acetyldihydrosphingosine, N-acetylsphingosine, and N-stearoylsphinganine were significantly modulated by PSORI-CM02 (Figure 5A). Similarly, six differentially expressed sphingolipids were identified in the skin of psoriasis mice, five of which, including Ganglioside GM2 (d18:1/18:0), C16-Dihydroceramide, Sphingosine 1-phosphate, D-Erythro-sphingosine 1-phosphate, and 3-Sulfodeoxycholic acid could be modulated by PSORI-CM02 (Figure 5B). In addition, all differentially expressed sphingolipids identified in the lymph nodes were significantly modulated by PSORI-CM02 (Figure 5C). We further assessed the sphingolipid content in mouse skin tissues using ELISA. The results showed that the levels of ceramide, S1P, C1P, and SM were elevated in psoriasis mice (Figure 5D). However, treatment with PSORI-CM02 reduced their concentrations.
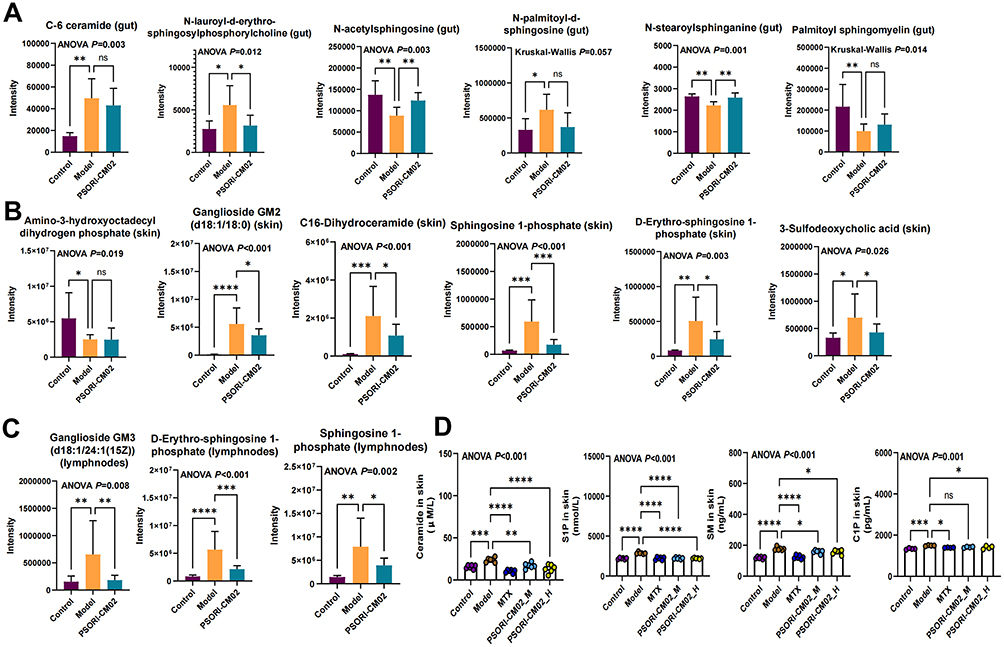 |
Figure 5 Multi-tissue metabolomics elucidates the regulatory effects of PSORI-CM02 on sphingolipid metabolism. (A) Sphingolipids in gut. (B) Sphingolipids in skin. (C) Sphingolipids in lymph nodes. (D) Validation of sphingolipid concentration in skin by ELISA (biological replicates n=5-6 per group). Data are presented as mean ± standard deviation. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, ns, not significant, compared with the model group. Abbreviations: ANOVA, analysis of variance; S1P, sphingosine-1-phosphate; C1P, ceramide-1-phosphate; SM, sphingomyelin; MTX, methotrexate; PSORI-CM02_M, PSORI-CM02 at medium dose (1.42 g/kg); PSORI-CM02_H, PSORI-CM02 at high dose (2.84 g/kg); ELISA, enzyme-linked immunosorbent assay. |
Gut Microbiota Analysis Reveals Alterations of Sphingolipid Metabolism-Related Microbiota in Psoriasis Mice
α-diversity was used to assess the complexity of the bacterial community.22 As shown in Figure 6A, the α diversity remained unchanged among the three groups, suggesting that PSORI-CM02 had no significant effect on α-diversity of the microbiota. β-diversity was used to compare the microbiome variety among different samples. PCA analysis (PCA1 = 32.25%, PCA2 = 17.51%) showed clear separation between the control and model groups, whereas samples from the PSORI-CM02 group were widely spread between the control and model groups, suggesting that PSORI-CM02 interrupted the intestinal microbiome of psoriasis to a physiological state (Figure 6B).
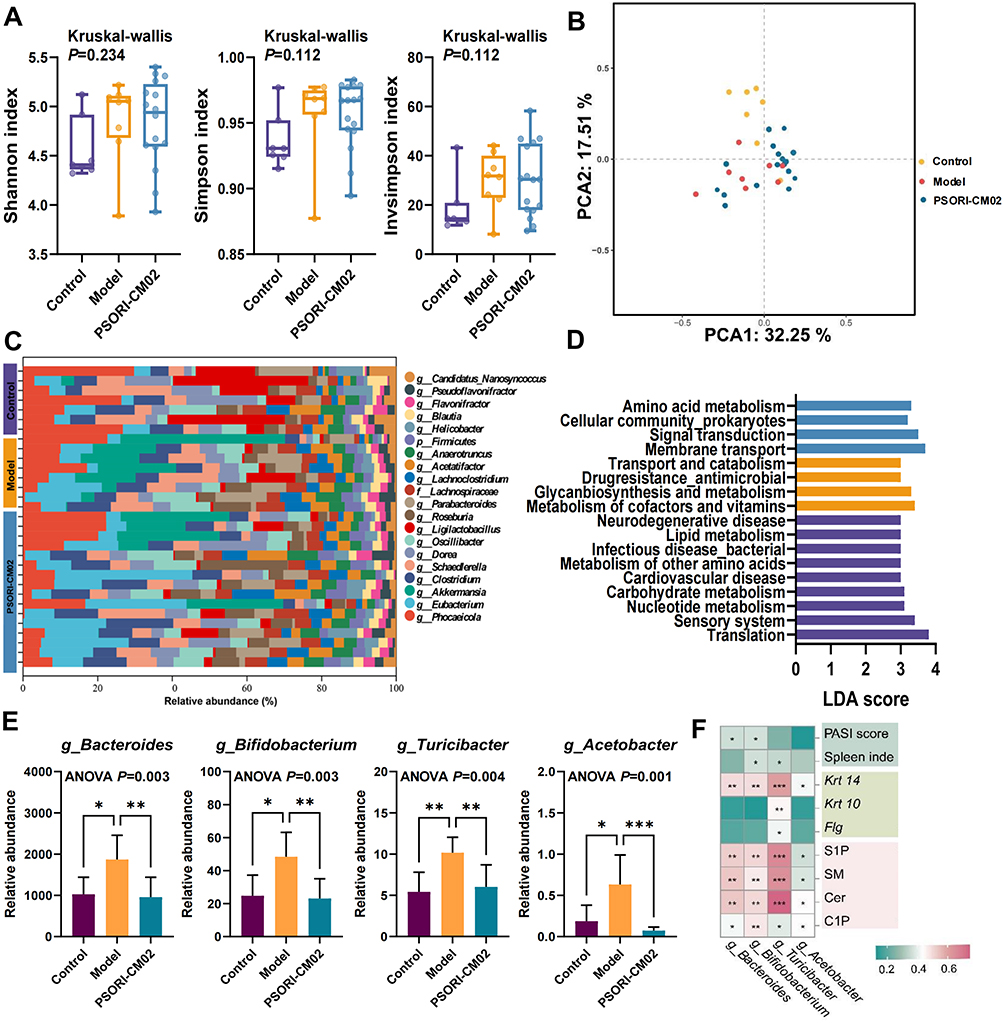 |
Figure 6 Gut metagenomics analysis. (A) α-diversity. (B) PCA of gut metagenomics. (C) Relative abundance of the top 20 microbiota at the genus level. (D) LEfSe analysis of the control, model, and PSORI-CM02 groups at the KEGG pathway level B. (E) The relative abundance of sphingolipid metabolism-related microbiome species at the genus level. (F) Spearman correlation analysis. Data are presented as mean ± standard deviation. *p < 0.05, **p < 0.01, ***p < 0.001, compared to the model group. Abbreviations: ANOVA, analysis of variance; PCA, principal component analysis; LEfSe, linear discriminant analysis effect size; LDA, linear discriminant analysis; KEGG, Kyoto Encyclopedia of Genes and Genomes; MTX, methotrexate. |
The relative abundances of the top 20 microbiota at the genus level in the three groups are shown in Figure 6C. Species structure analyses indicated that the microbiome structure of psoriatic mice was notably altered at the genus level by treatment with IMQ and that PSORI-CM02 restored the microbiota structure in psoriatic mice. In particular, Akkermansia was more abundant in the model group.
Differences in the pathway enrichment of the microbiome among the three groups were investigated. For pathways at KEGG level B, as shown in Figure 6D, the protective effect of PSORI-CM02 on the microbiome was observed mainly in amino acid metabolism, cellular community prokaryotes, signal transduction, and membrane transport and catabolism.
As for sphingolipid metabolism-related microbial communities, the relative abundance of Bacteroides, Bifidobacterium, Turicibacter, and Acetobacter in genus level were increased, whereas PSORI-CM02 treatment decreased the tendency (Figure 6E). Spearman correlation analysis between the pharmacodynamic indicators, epidermal differentiation, sphingolipids and microbiome was performed. The results indicated that relative abundance of the four sphingolipid-related microbiota were closely associated with PSORI-CM02 pharmacodynamic and epidermal differentiation (Figure 6F). Therefore, we proceeded to validate these findings by conducting FMT experiments on SPF, PGF, and GF psoriasis mice.
PSORI-CM02-FMT Restores the Intestinal Balance, Then Modifies Sphingolipid Biosynthesis, and Thereby Regulates Epidermal Proliferation
FMTs from the PSORI-CM02 treated group to IMQ-induced psoriatic mice were performed to investigated the bioactivities of PSORI-CM02-FMT on psoriatic lesion, epidermal differentiation, and sphingolipids. All the SPF, PGF and GF mice receiving PSORI-CM02-FMT demonstrated improved psoriatic lesions (Figure 7A) and decreased PASI scores (Figure 7B). HE staining suggested that PSORI-CM02-FMT alleviated epidermal thickening, reduced infiltration of inflammatory cells, improved irregularly shaped rete ridges and dilated capillaries (Figure 7C). In addition, PSORI-CM02-FMT up-regulated relative mRNA expression of Flg and Krt 10, while decreased the Krt 14 expression (Figure 7D). These results indicated PSORI-CM02-FMT inhibited excessive proliferation and abnormal differentiation of the epidermis.
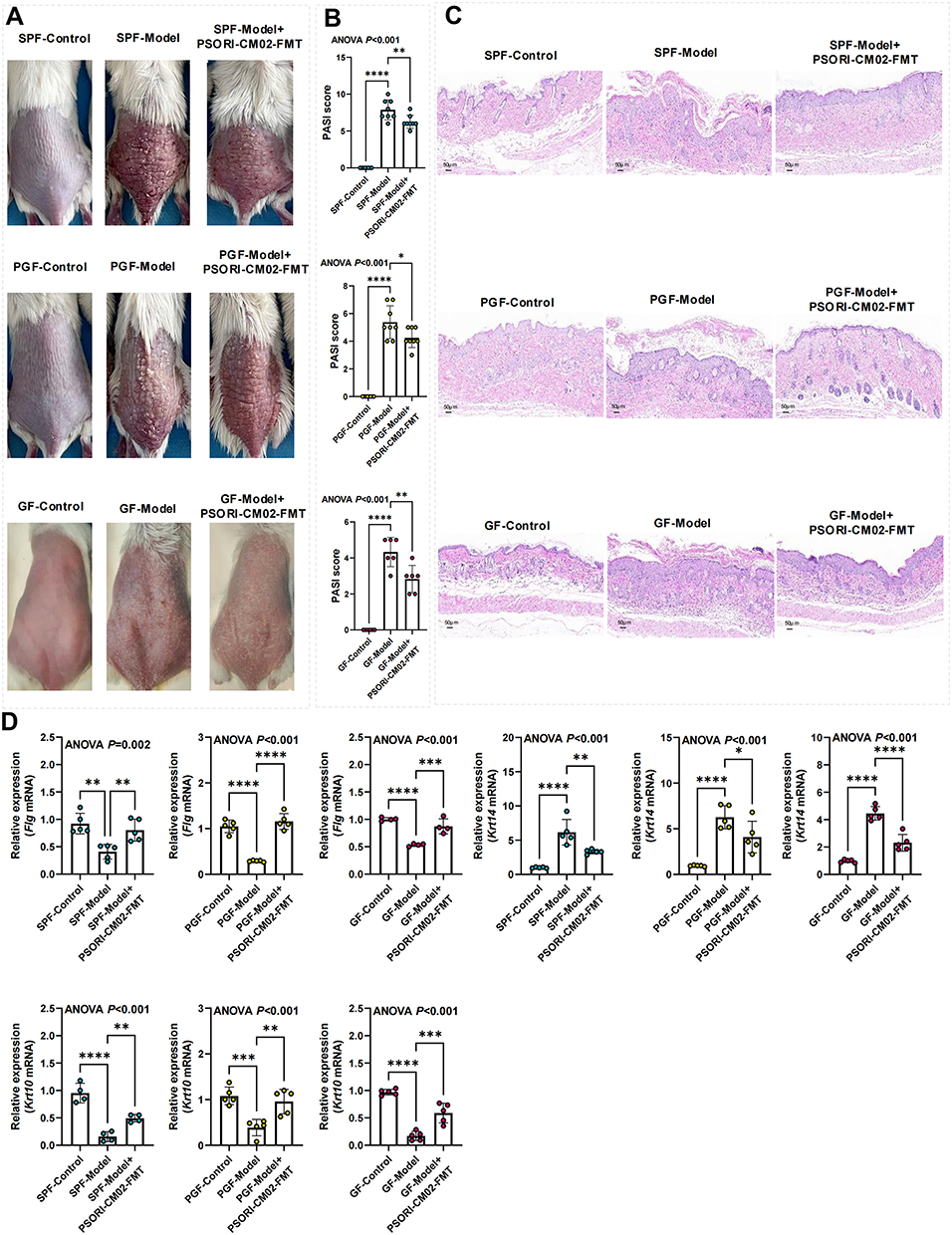 |
Figure 7 PSORI-CM02-FMT inhibits epidermal proliferation. (A) Skin images (n = 8 per group). (B) PASI scores (n = 6–8 per group). (C) HE staining (magnification ×200, scale bar = 50 μm). (D) Relative expression of Flg, Krt 10, and Krt 14 in skin tissue detected by RT-PCR (biological replicates n = 5 per group). Data are presented as mean ± standard deviation. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, compared with the model group. Abbreviations: SPF, Specific pathogen-free; PGF, pseudo germ-free; GF, germ-free; FMT, fecal microbiota transplantation; RT-PCR, Real-time quantitative reverse transcription-polymerase chain reaction; ANOVA, analysis of variance; PASI, psoriatic area and severity index. |
Skin and fecal samples from mice were analyzed for sphingolipid concentrations using ELISA. The results indicated that PSORI-CM02-FMT reduced the levels of ceramide, S1P, C1P, and SM in both skin and feces of psoriasis mice (Figure 8), suggesting that PSORI-CM02-FMT can inhibit sphingolipid synthesis in psoriasis mice.
 |
Figure 8 PSORI-CM02-FMT suppresses sphingolipid biosynthesis in skin and feces. Data are presented as mean ± standard deviation. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, ns, not significant, compared with the model group. Abbreviations: ANOVA, analysis of variance; S1P, sphingosine-1-phosphate; C1P, ceramide-1-phosphate; SM, sphingomyelin; ELISA, enzyme-linked immunosorbent assay; SPF, Specific pathogen-free; PGF, pseudo germ-free; GF, germ-free; FMT, fecal microbiota transplantation. |
The influence of PSORI-CM02-FMT on the gut microbiota composition in mice was additionally examined through 16S rDNA amplicon sequencing. The α-diversity, as indicated by the Shannon index, remained unaffected by the FMT (Figure 9A). Nevertheless, PCoA revealed distinct clustering in the gut microbiota configuration among the control, model, and PSORI-CM02-FMT groups (Figure 9B), suggesting that the intestinal microbial architecture in the psoriasis mice was influenced by PSORI-CM02-FMT. At the genus level, PSORI-CM02-FMT was observed to rehabilitate the makeup of the gut microbiota, aligning with the alterations observed subsequent to the administration of PSORI-CM02 (Figure 9C). A cladogram, derived from the LEfSe analysis of the microbiome data, is depicted in Figure 9D. Furthermore, modifications in the microbiota of both PGF and GF mice following PSORI-CM02-FMT were noted. The α and β diversities of the gut microbiota substantiated the successful colonization of PGF mice (Figure 10A and B). Given that PGF and GF mice are devoid of any indigenous microbes in their intestinal tracts, the observed microbiota shifts were more pronounced compared to those in SPF mice. At the genus level,contrary to our findings in SPF mice, PSORI-CM02-FMT exhibited an effect on reshaping the intestinal microbial community structure (Figure 10C–E, Supplementary Figure 2). However, in GF and PGF recipients, three genera that were associated with sphingolipid biosynthesis, including Bacteroides, Bifidobacterium, and Turicibacter, was detected and observed that PSORI-CM02-FMT tended to lower their combined abundance (Supplementary Figure 3), which was consist with the SPF mice.
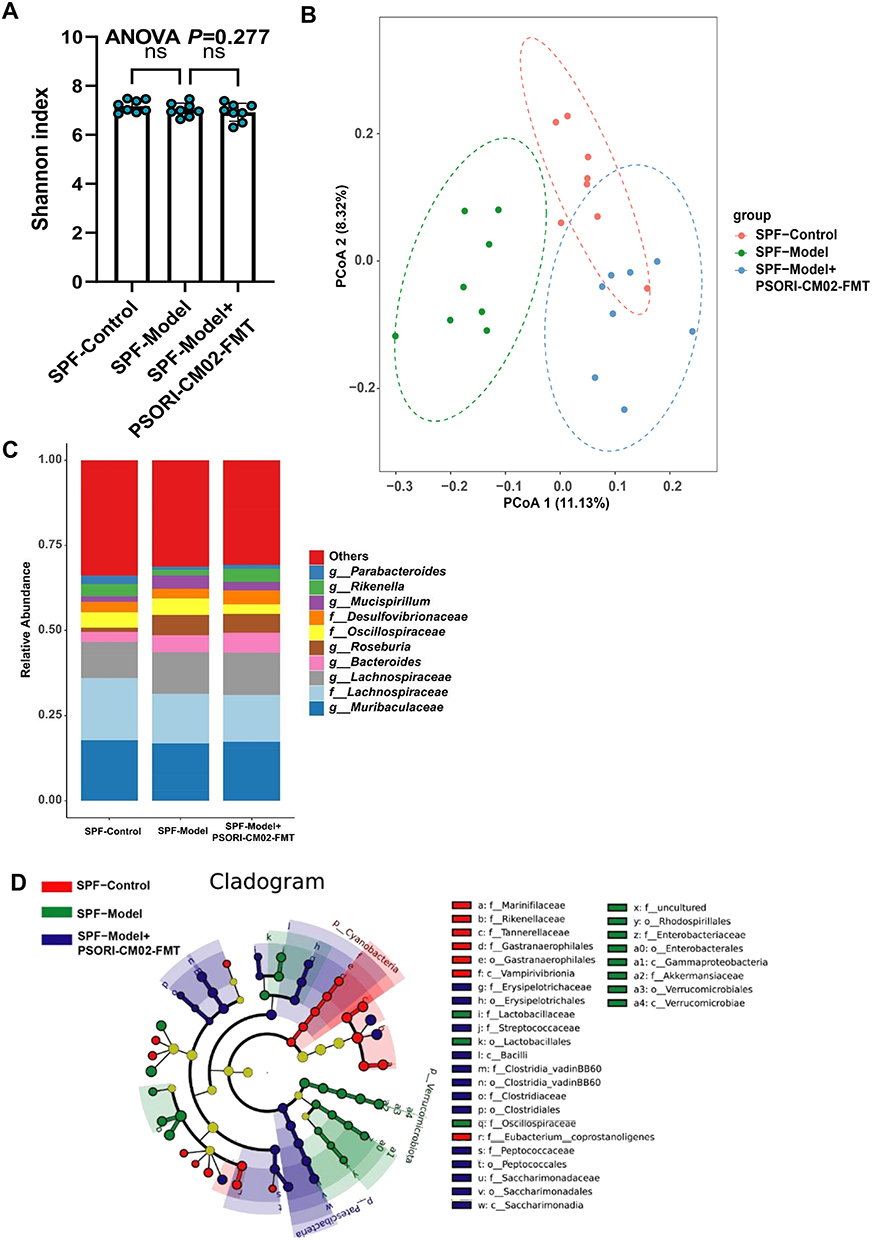 |
Figure 9 PSORI-CM02-FMT restores the intestinal balance in SPF mice. (A) α-diversity of SPF mice. (B) PCoA analysis of SPF mice. (C) Relative abundance of the top 10 microbiota at the genus level of SPF mice. (D) Cladogram of the LEfSe analysis results from SPF mice. Data are presented as mean ± standard deviation. ns, not significant, compared with the model group. Abbreviations: SPF, Specific pathogen-free; FMT, fecal microbiota transplantation; ANOVA, analysis of variance; LEfSe, linear discriminant analysis effect size; PCoA, principal coordinates analysis. |
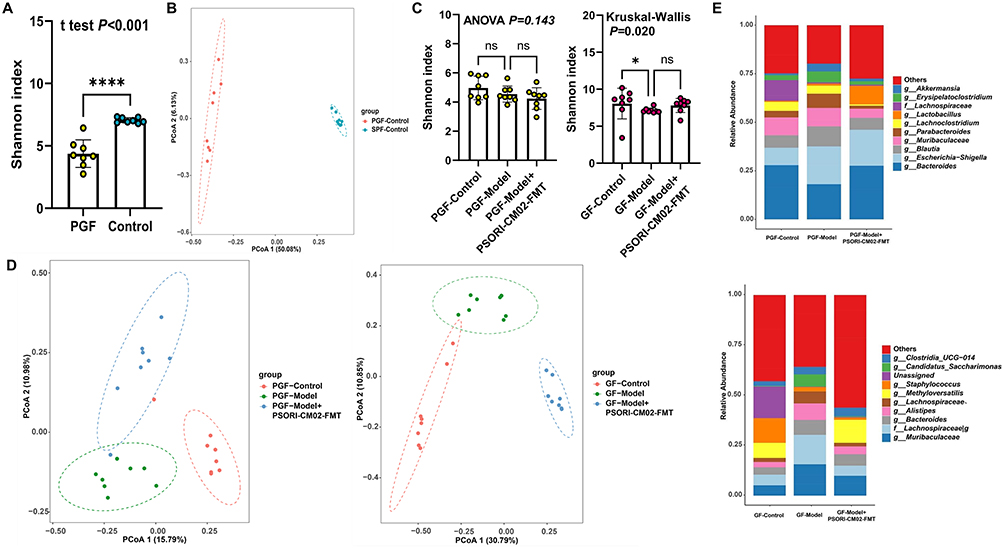 |
Figure 10 PSORI-CM02-FMT restores the intestinal balance in PGF and GF mice. (A) α-diversity for the comparison between the PGF and SPF mice. (B) PCoA analysis for the comparison between the PGF and SPF mice. (C) α-diversity of PGF and GF mice. (D) PCoA analysis of PGF and GF mice. (E) Relative abundance of the top 10 microbiota at the genus level of PGF and GF mice. Data are presented as mean ± standard deviation. ****p < 0.001, *p < 0.05, ns, not significant, compared with the model group. Abbreviations: SPF, Specific pathogen-free; PGF, pseudo germ-free; GF, germ-free; FMT, fecal microbiota transplantation; ANOVA, analysis of variance; LEfSe, linear discriminant analysis effect size; PCoA, principal coordinates analysis. |
Discussion
Psoriatic lesions arise from disrupted keratinocyte differentiation, in which sphingolipids act as both structural lipids and signaling molecules governing barrier integrity and cell fate.12,23 Transcriptomic profiling of patient keratinocytes revealed enrichment of sphingolipid biosynthetic genes, and selective inhibition of this pathway with carmofur promoted differentiation marker expression—establishing sphingolipid metabolism as a mechanistic driver of epidermal dysregulation. Despite documented anti-inflammatory, anti-angiogenic24 and pro-autophagic25 effects of PSORI-CM02,16,17 its impact on the gut–skin axis remained unexplored.18,19,26–29 We demonstrate that PSORI-CM02 attenuates IMQ-induced psoriasis while concurrently reducing the relative abundance of sphingolipid-producing taxa (Bacteroides, Bifidobacterium, Turicibacter, Acetobacter) and lowering ceramide, S1P, C1P and SM levels in skin, lymph nodes and gut. Multi-omics integration confirmed strong inverse correlations between these microbial taxa, sphingolipid content, and epidermal differentiation markers. Fecal microbiota transplantation from PSORI-CM02-treated donors reproduced the therapeutic phenotype in SPF, PGF and GF mice, confirming that the drug’s efficacy is transmissible via the microbiome and operates through suppression of microbial sphingolipid synthesis to restore normal keratinocyte differentiation.
The imbalance of the gut microbiome in individuals with psoriasis correlates with the intensity and phase of their condition.5 Diverse research exhibits inconsistent findings concerning the makeup of the gut microbiome among individuals with psoriasis.30–32 FMT is currently regarded as the most efficacious approach to rehabilitate a balanced diversity within the gastrointestinal microbiome.33 The therapeutic potential of FMT for psoriasis patients remains an area of investigation; however, preliminary outcomes from clinical studies are generating enthusiasm. Within the framework of a five-week clinical trial involving an intervention, a participant suffering from plaque psoriasis and irritable bowel syndrome underwent FMT twice, administered through endoscopic and colonoscopic procedures. Following the treatment, improvements were noted in the body surface area affected by psoriasis, PASI score, dermatology life quality index, gastrointestinal symptoms, and serum levels of tumor necrosis factor-alpha, with no adverse effects detected.8 However, no assessments of shifts in the gut microbiota were conducted, and the limited participant pool is a recognized constraint of the study. In our investigation, we performed FMT experiments on SPF, PGF, and GF psoriasis mice models. Given that PGF and GF mice lack any indigenous microorganisms in their gastrointestinal tracts, the observed microbiota alterations were more significant compared to those observed in SPF mice. Moreover, the data from SPF, PGF, and GF mice collectively indicated that FMT with PSORI-CM02 demonstrates analogous effects to those of PSORI-CM02 itself, thereby reinforcing the notion that PSORI-CM02 is capable of mediating its therapeutic benefits through the modulation of the intestinal microbiome.
Recently, the commensal gut microbes (Bacteroides, Bifidobacterium, Turicibacter, and Acetobacter) have been reported to produce sphingolipids, including ceramide, phosphoinositol, and deoxy-sphingolipids.34 Interestingly, these sphingolipids have been documented to exacerbate gastrointestinal inflammation and modulate host ceramide levels in animal models, and a deficiency in microbial-derived sphingolipids has been linked to inflammatory bowel diseases. Additionally, such a deficiency influences the quantities of host-produced sphingolipids in humans.34 In our research, PSORI-CM02-FMT was conducted on PGF and GF psoriasis mice, which led to the regulation of sphingolipid metabolism and consequently enhanced epidermal differentiation. Since SPF mice typically receive no ABX pre-treatment, the transplanted microbiota must coexist with the resident flora, yielding higher overall diversity; therefore, divergence in community structure between SPF and GF/PGF mice after FMT is inevitable In consist with the SPF mice, three genera that were associated with sphingolipid biosynthesis in GF and PGF recipients, was observed that PSORI-CM02-FMT tended to lower their combined abundance. This functional convergence despite compositional differences explains the consistent therapeutic outcome across models. The outcomes were in alignment with those observed in SPF mice, but they more robustly established the connection between the gut microbiota and sphingolipid metabolism.
This study acknowledges limitations that warrant consideration. The lack of a negative control group that received PSORI-CM02 without IMQ induction means that the isolated impact of PSORI-CM02 on the gut microbiota cannot be discerned. To resolve this ambiguity, future work will include SPF, GF and PGF cohorts that receive PSORI-CM02 or vehicle for 7 consecutive days without IMQ challenge. We also plan to employ psoriasis keratinocyte cell lines or primary cells to decipher the influence of PSORI-CM02 on sphingolipid metabolism and the process of epidermal stratification. Moreover, we aim to delineate the therapeutic spectrum of PSORI-CM02, with the goal of establishing the optimal and secure dosing parameters.
Conclusion
PSORI-CM02 exerts its therapeutic benefits by mediating the composition of the gut microbiota, subsequently regulating sphingolipid metabolism, and thereby enhancing epidermal differentiation. This innovative discovery not only broadens our comprehension of the pathogenic mechanisms involved in psoriasis but also presents novel perspectives on potential therapeutic targets and treatment modalities.
Abbreviations
PASI, psoriasis area, and severity index; IMQ, imiquimod; MTX, Methotrexate; HE, hematoxylin, and eosin; ELISA, enzyme-linked immunosorbent assay; PCA, Principal component analysis; PLS-DA, Partial least squares discriminant analysis; OPLS-DA, Orthogonal partial least-squares discrimination analysis; VIP, Variable Importance in Projection; KEGG, Kyoto Encyclopedia of Genes and Genomes; LEfSe, linear discriminant analysis effect size; SPF, Specific pathogen-free; PGF, pseudo germ-free; GF, germ-free; FMT, fecal microbiota transplantation; ABX, antibiotics; ANOVA, analysis of variance; S1P, sphingosine-1-phosphate; C1P, ceramide-1-phosphate; SM, sphingomyelin, PCoA, principal coordinates analysis.
Data Sharing Statement
The datasets during and/or analyzed during the current study available from the corresponding author on reasonable request.
Ethics Statement
This study was approved by the Ethics Committee of the Second Affiliated Hospital of Guangzhou University of Chinese Medicine (B2013-132-01). All participants provided informed consent, in accordance with the Declaration of Helsinki. All animal procedures were approved by the Animal Ethics Committee of the Second Affiliated Hospital of Guangzhou University of Chinese Medicine (Number 2021054).
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This research was financially supported by the National Natural Science Foundation of China (82004363, 82374313, U23A6012, U20A20397), Innovation Team and Talents Cultivation Program of National Administration of Traditional Chinese Medicine (ZYYCXTD-C-202204), Science and Technology Planning Project of Guangdong Province (2023B1212060063, 2020B1111100006, 2020B1212030006), State Key Laboratory of Dampness Syndrome of Chinese Medicine Special Fund (SZ2021ZZ29, SZ2021ZZ45), Youth Talent Promotion Project of China Association of Traditional Chinese Medicine (2021-QNRC2-B07), and Guangdong Provincial Hospital of Chinese Medicine Special Fund (YN2023ZH16, YN2024GZRPY028, YN2021DB03).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Leisner MZ, Riis JL, Schwartz S, Iversen L, Østergaard SD, Olsen MS. Psoriasis and risk of mental disorders in Denmark. JAMA Dermatol. 2019;155(6):745–747. doi:10.1001/jamadermatol.2019.0039
2. Nestle FO, Kaplan DH, Barker J. Psoriasis. N Engl J Med. 2009;361(5):496–509. doi:10.1056/NEJMra0804595
3. Lee EB, Amin M, Egeberg A, Wu JJ. Adverse events associated with apremilast use and withdrawal for psoriasis in a real-world setting. J Eur Acad Dermatol Venereol. 2018;32(10):e393–e394. doi:10.1111/jdv.15061
4. Wu M, Deng Y, Li S, et al. The immunoregulatory effects of Traditional Chinese medicine on psoriasis via its action on interleukin: advances and considerations. Am J Chin Med. 2018;46(4):739–750. doi:10.1142/S0192415X18500386
5. Buhaș MC, Gavrilaș LI, Candrea R, et al. Gut microbiota in psoriasis. Nutrients. 2022;14(14):2970. doi:10.3390/nu14142970
6. Wu CY, Chang YT, Juan CK, et al. Risk of inflammatory bowel disease in patients with rosacea: results from a nationwide cohort study in Taiwan. J Am Acad Dermatol. 2017;76(5):911–917. doi:10.1016/j.jaad.2016.11.065
7. Kim M, Choi KH, Hwang SW, Lee YB, Park HJ, Bae JM. Inflammatory bowel disease is associated with an increased risk of inflammatory skin diseases: a population-based cross-sectional study. J Am Acad Dermatol. 2017;76(1):40–48. doi:10.1016/j.jaad.2016.08.022
8. Yin G, Li JF, Sun YF, et al. Fecal microbiota transplantation as a novel therapy for severe psoriasis. Zhonghua Nei Ke Za Zhi. 2019;58:9–19.
9. Masuda-Kuroki K, Alimohammadi S, Di Nardo A. The role of sphingolipids and sphingosine-1-phosphate-sphingosine-1-phosphate-receptor signaling in psoriasis. Cells. 2023;12(19):2352. doi:10.3390/cells12192352
10. Matwiejuk M, Mysliwiec H, Chabowski A, Flisiak I. The role of sphingolipids in the pathogenesis of psoriasis. Metabolites. 2022;12(12):1171. doi:10.3390/metabo12121171
11. Matwiejuk M, Myśliwiec H, Lukaszuk B, et al. Crosstalk between serum and skin sphingolipids in psoriasis. Int J Mol Sci. 2023;24(19):14872. doi:10.3390/ijms241914872
12. Bocheńska K, Gabig-Cimińska M. Unbalanced sphingolipid metabolism and its implications for the pathogenesis of psoriasis. Molecules. 2020;25(5):1130. doi:10.3390/molecules25051130
13. Lamichhane S, Sen P, Alves MA, et al. Linking gut microbiome and lipid metabolism: moving beyond associations. Metabolites. 2021;11(1):55. doi:10.3390/metabo11010055
14. Bai X, Ya R, Tang X, Cai M. Role and interaction of bacterial sphingolipids in human health. Front Microbiol. 2023;14:1289819. doi:10.3389/fmicb.2023.1289819
15. Rohrhofer J, Zwirzitz B, Selberherr E, Untersmayr E. The impact of dietary sphingolipids on intestinal microbiota and gastrointestinal immune homeostasis. Front Immunol. 2021;12:635704. doi:10.3389/fimmu.2021.635704
16. Wang JY, hE ZH, Deng H, et al. The efficacy and safety of Shaoling decoction in the treatment of psoriasis vulgaris based on real-world data. Tradit Chin Drug Res Pharmacol. 2021;32(05):739–743.
17. Yao DN, Ye SY, Yu MY, Deng H, Yan YH, Lu CJ. The efficacy and safety of PSORI-CM02 in the treatment of psoriasis vulgaris. World Sci Technol Mod Tradit Chin Med Mater Med. 2023;25(02):716–723.
18. Chen H, Liu H, Lu C, et al. PSORI-CM02 formula increases CD4+ Foxp3+ regulatory T cell frequency and ameliorates imiquimod-induced psoriasis in mice. Front Immunol. 2017;8:1767. doi:10.3389/fimmu.2017.01767
19. Wu DH, Zhang MM, Li N, et al. PSORI-CM02 alleviates IMQ-induced mouse dermatitis via differentially regulating pro- and anti-inflammatory cytokines targeting of Th2 specific transcript factor GATA3. Biomed Pharmacother. 2019;110:265–274. doi:10.1016/j.biopha.2018.11.092
20. Scheiblich H, Schlütter A, Golenbock DT, Latz E, Martinez-Martinez P, Heneka MT. Activation of the NLRP3 inflammasome in microglia: the role of ceramide. J Neurochem. 2017;143(5):534–550. doi:10.1111/jnc.14225
21. Chen Y, Peng M, Li W, et al. Inhibition of inflammasome activation via sphingolipid pathway in acute lung injury by Huanglian Jiedu decoction: an integrative pharmacology approach. Phytomedicine. 2022;107:154469. doi:10.1016/j.phymed.2022.154469
22. Lkhagva E, Chung HJ, Ahn JS, Hong ST. Host factors affect the gut microbiome more significantly than diet shift. Microorganisms. 2021;9(12):2520. doi:10.3390/microorganisms9122520
23. Kim J, Krueger JG. The immunopathogenesis of psoriasis. Dermatol Clin. 2015;33(1):13–23. doi:10.1016/j.det.2014.09.002
24. Lu Y, Yang Y, Zhang J, et al. Anti-angiogenic efficacy of PSORI-CM02 and the associated mechanism in psoriasis in vitro and in vivo. Front Immunol. 2021;12:649591. doi:10.3389/fimmu.2021.649591
25. Yue L, Ailin W, Jinwei Z, et al. PSORI-CM02 ameliorates psoriasis in vivo and in vitro by inducing autophagy via inhibition of the PI3K/Akt/mTOR pathway. Phytomedicine. 2019;64:153054. doi:10.1016/j.phymed.2019.153054
26. Deng J, Tan S, Liu R, et al. Chinese medicine formula PSORI-CM02 alleviates psoriatic dermatitis via M-MDSCs and Th17 crosstalk. Front Pharmacol. 2020;11:563433. doi:10.3389/fphar.2020.563433
27. Lu C, Liu H, Jin X, et al. Herbal components of a novel formula PSORI-CM02 interdependently suppress allograft rejection and induce CD8+CD122+PD-1+ regulatory T cells. Front Pharmacol. 2018;9:88. doi:10.3389/fphar.2018.00088
28. Li L, Zhang HY, Zhong XQ, et al. PSORI-CM02 formula alleviates imiquimod-induced psoriasis via affecting macrophage infiltration and polarization. Life Sci. 2020;243:117231. doi:10.1016/j.lfs.2019.117231
29. Yuan X, Huang L, Lei J, Long Y, Li C. Study on anti-inflammatory effect and major anti-inflammatory components of PSORI-CM02 by zebrafish model. Evid Based Complement Alternat Med. 2020;2020:5604654. doi:10.1155/2020/5604654
30. Hidalgo-Cantabrana C, Gómez J, Delgado S, et al. Gut microbiota dysbiosis in a cohort of patients with psoriasis. Br J Dermatol. 2019;181(6):1287–1295. doi:10.1111/bjd.17931
31. Shapiro J, Cohen NA, Shalev V, Uzan A, Koren O, Maharshak N. Psoriatic patients have a distinct structural and functional fecal microbiota compared with controls. J Dermatol. 2019;46(7):595–603. doi:10.1111/1346-8138.14933
32. Zhang X, Shi L, Sun T, Guo K, Geng S. Dysbiosis of gut microbiota and its correlation with dysregulation of cytokines in psoriasis patients. BMC Microbiol. 2021;21(1):78. doi:10.1186/s12866-021-02125-1
33. Ramai D, Zakhia K, Ofosu A, Ofori E, Reddy M. Fecal microbiota transplantation: donor relation, fresh or frozen, delivery methods, cost-effectiveness. Ann Gastroenterol. 2019;32(1):30–38. doi:10.20524/aog.2018.0328
34. Brown EM, Ke X, Hitchcock D, et al. Bacteroides-derived sphingolipids are critical for maintaining intestinal homeostasis and symbiosis. Cell Host Microbe. 2019;25(5):668–680.e7. doi:10.1016/j.chom.2019.04.002
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.