Back to Journals » Journal of Experimental Pharmacology » Volume 17
Psidium guajava Leaves n-Hexane Fraction Antibacterial Activity and the Inhibition of Gene Expression of gtfB and gtfC in the Combination Streptococcus mutans ATCC 25175 and Veillonella parvula ATCC 10790T
Authors Setiawan AS ![]() , Subarnas A, Djais AA, Milanda T, Ichwan SJA
, Subarnas A, Djais AA, Milanda T, Ichwan SJA
Received 30 March 2025
Accepted for publication 13 August 2025
Published 8 September 2025 Volume 2025:17 Pages 639—650
DOI https://doi.org/10.2147/JEP.S531035
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Prof. Dr. Abdelwahab Omri
Ame Suciati Setiawan,1,2 Anas Subarnas,3 Ariadna Adisatya Djais,4 Tiana Milanda,5 Solachudin Jauhari Arief Ichwan6
1Doctoral Program Pharmacy, Faculty of Pharmacy, Padjadjaran University, Bandung, West Java, Indonesisa; 2Department of Oral Biology, Faculty of Dentistry, Padjadjaran University, Bandung, West Java, Indonesia; 3Department of Pharmacology and Clinical Pharmacy, Faculty of Pharmacy, Padjadjaran University, Bandung, West Java, Indonesia; 4Department of Oral Biology Faculty of Dentistry, University of Indonesia, Jakarta, Indonesia; 5Department of Biological Pharmacy, Faculty of Pharmacy, Padjadjaran University, Bandung, West Java, Indonesia; 6PAPRSB Institute of Health Sciences, University of Brunei Darussalam, Brunei, Darussalam
Correspondence: Ame Suciati Setiawan, Email [email protected]
Background: Oral biofilm is an extracellular polymeric matrix produced by Streptococcus mutans as the early colonizer in the formation of an oral biofilm. Glucosyltransferase (Gtfs) enzyme is a virulence factor for S. mutans in biofilm that is resistant to antibacterial agents. Veillonella parvula influences S. mutans to increased quantities of Gtfs, thereby accelerating the growth of oral biofilms. Decreasing the production of Gtfs by the use of additional materials is necessary to inhibit biofilm formation.
Objective: The study was determined the inhibitory activity of the Psidium guajava leaves n-hexane fraction on the production of Gtfs by a single and combination of S. mutans and V. parvula.
Methods: Antibacterial effect of the P. guajava leaves n-hexane fraction against S. mutans and V. parvula was examined by the inhibition zone and, and MBC. The antibiofilm was examined on the expression of gtfB and gtfC in S. mutans, V. parvula, and their combination using qRT-PCR. The results were analyzed with Kruskal–Wallis test and Mann–Whitney U-test.
Results: P. guajava leaves inhibited the growth of S. mutans and V. parvula at the lowest concentration (3.125%), with the inhibition zones are larger than chlorhexidine (CHX). The effect on the Gtfs activity shown the gene expression of gtfB remained unchanged in both single S. mutans and combined species, while it was decreased in the single V. parvula at a concentration of 0.78%, but itdoes not exhibit a significant difference between single and double species. Conversely, gtfC gene expression in the dual-species was reduced 90.43% by the n-hexane fraction at a concentration of 0.78%.
Conclusion: The P. guajava leaves n-hexane fraction exhibited antibacterial activity against S. mutans and V. parvula. It may exhibit antibiofilm formation, as it could reduce the expression of the gtfC in the double species bacteria.
Keywords: V. parvula, S. mutans, glucosyltransferase, P. guajava, antibiofilm
Introduction
Oral biofilm is characterized by a layer of gram-positive and gram-negative bacteria that generate a polysaccharide matrix, facilitating their adherence and proliferation on both biotic (oral mucous) and abiotic (teeth, dental restoration, implant) hard surfaces.1 Gram-positive bacteria are the early colonizers in oral biofilm production due to their capacity to adhere to the pellicle on the tooth surface. These bacteria generate acid, which can facilitate the adhesion of more bacteria, including gram-negative species.2 Gram-negative microbes are generally incapable of directly adhering to tooth surfaces. Rather, they establish a connection with the earliest colonizers through a series of specific interactions known as coaggregation.2
Oral biofilm forms due to the capacity of Streptococcus mutans to metabolize sucrose, resulting in the production of organic acids (lactic acid) and glucosyltransferase, which facilitates bacterial adherence to tooth surfaces and other microbes.3 Bacteria generate extracellular polymeric substances (EPS) that induce chronic infections, and potentially resulting in the demineralization of dental enamel by increasing the adhesion to the tooth surface or between other microorganism.1,4 The formation of biofilm begins with the adhesion of initial colonies, such as S. mutans species, to the proteins and glycoproteins in saliva on the tooth surface.5,6
S. mutans is an anaerobic gram-positive cocci bacterium that is commonly found as an early colonizer in the formation of oral biofilms that cause dental caries. The virulence factor of S. mutans is the sucrose-dependent adhesion mechanism, which is responsible for the colonization of tooth surfaces, and adhesion contributes to the formation of oral biofilm facilitated by the Gtfs.7,8 Gtfs enzyme catabolizes sucrose into glucan through glycosidic bonds and forms an extracellular polysaccharide (EPS) matrix.8 Recent studies have indicated that Gtfs can modulate the interaction between S. mutans and other pathogenic bacteria, consequently affecting the formation and virulence of oral biofilm.8
Veillonella parvula is a gram-negative anaerobic coccus and an early colonizer of dental plaque and S. mutans.7,9 This bacterium cannot ferment glucose or other sugars but can metabolize lactic acid produced by S. mutans for its growth.7,9 V. parvula requires S. mutans to have a symbiotic relationship for growth. The presence of V. parvula can promote S. mutans in biofilm formation. The biofilm resulting from single S. mutans exhibited a lower pH than single V. parvula, and the double species biofilm of S. mutans combined with V. parvula had the same pH as S. mutans. This means that the existence of V. parvula cannot increase the pH of the biofilm, even if the lactic acid from S. mutans has been consumed by V. parvula.10 The existing of V. parvula also increased thickness, and a greater EPS matrix compared to single-species biofilms.10 Gene expression of gtfB and gtfC produced by S. mutans is upregulated with the existence of V. parvula in biofilm formation.10 Consequently, the presence of V. parvula increases the risk of caries development and improves the resistance to external bactericidal agents.10,11 Previous studies have concluded that there is a relationship between the growth of V. parvula and S. mutans in biofilm formation; thus, preventive measures against caries are needed for both species.
Chlorhexidine (CHX) is the gold standard as an antibiofilm agent; however, it has numerous side effects and drawbacks, making a substitute for chlorhexidine as an antibiofilm agent necessary. One of the materials that can be used is an herbal plant, namely guava leaves. Psidium guajava is a plant that grows in tropical regions and has pharmacological effects on several disorders. P. guajava leaf extract has been reported to exert antibacterial effects against S. mutans.12 The compounds found in P. guajava can reduce the number of S. mutans bacterial colonies, inhibit the activity of Gtfs enzymes, and inhibit biofilm formation by glucan.13 Furthermore, P. guajava has antibacterial activity against both gram-positive and gram-negative bacteria because of its rich flavonoid, guajaverin, and quercetin contents.14 These studies show that the extract of P. guajava leaves has antibacterial effects on S. mutans; however, its effectiveness against V. parvula is not yet known. Therefore, it is of interest to investigate the antibacterial effect of P. guajava leaves on S. mutans and V. parvula in a single species and to examine the effect of the sample on biofilm formation by single and double species of S. mutans and V. parvula.
Materials and Methods
Plant Materials
P. guajava leaves used in this study were harvested from the Spices and Medicinal Plants Research Center (BALITTRO) in Lembang, West Java, Indonesia and were approximately 3–4 months old. Plant determinations were conducted at the School of Life Sciences and Technology, Institute of Technology, Bandung, West Java, Indonesia with the sample no. 3974/IT1. C11.2/TA.00/2022 https://herbarium.sith.itb.ac.id/koleksi/ and it authenticated by Arifin Surya Dwipa Irsyam (https://www.scopus.com/authid/detail.uri?authorId=57211286941;https://herbarium.sith.itb.ac.id/profil-kurator/) showed that the leaves were Psidium guajava L. Leaves were washed and dried in open air and protected from direct sunlight.
Fraction Preparation
Dried leaves were powdered and extracted with ethanol 70% (three times, each 24 h) at a room temperature using a maceration method to produce a diluted ethanol extract, the solvent was then evaporated under reduced pressure at 50°C to obtain a concentrated extract.15 The concentrated ethanol extract is added to an ethanol-water solvent to dissolve the extract, then add n-hexane solvent (1:1) to obtain hexane and water layers. Both layers are separated. The procedure is repeated three times, then placed into the evaporator to produce a thick fraction.16 The n-hexane fraction was employed in this study to assess its inhibitory activity against S. mutans and V. parvula bacteria, as well as its effects on GtfS enzyme activity and the expression of gtfB and gtfC genes. This choice was due to the low polarity of the n-hexane fraction, facilitating its binding to the hydrophobic regions of bacterial cells, in contrast to the higher polarity of the ethanol extract and ethyl acetate fraction.17
Phytochemical Test
Qualitative phytochemical analysis of the P.guajava n-hexane fractions was performed to identify the principal classes of the compounds (tannins, saponins, flavonoids, alkaloids, phenols, glycosides, steroids, and terpenoids) following recognized techniques.
Phenolic Tested
The fraction was combined with three drops of FeCl3 to determine the presence of phenols. A positive sample changed color to greenish-blue.18
Tannin Tested
A small quantity of the fraction was mixed with distilled water and boiled. Filtrate produced was mixed with ferric chloride 1%, and the appearance of a blackish-green color indicated the presence of tannin.16
Flavonoid Test
The guava leaf n-hexane fraction was combined with ethanol and then filtered. The filtrate was combined with HCl concentrate and a magnesium ribbon. The emergence of a pink or red color indicates the presence of flavonoids. Another flavonoid test was conducted by adding distilled water and NaOH to the fraction, and the emergence of a yellowish color indicated the presence of flavonoids.16
Saponin Test
A number of fraction samples were placed in a test tube and distilled water was added. The mixture was shaken until froth was produced. The sample was boiled if it did not produce froth. Saponins were present when the foam was formed for more than 3 min.16
Terpenoid and Steroid Test
The fractions were then combined with chloroform and sulfuric acid. The development of a double layer (with the upper layer being red and the lower layer greenish) indicated the presence of steroids, whereas the emergence of a reddish-brown color indicated the presence of terpenoids.16
Alkaloid Test
The fraction sample was combined with 2% H2SO4 and subjected to heating for 2 min. After boiling, the solution was filtered and several drops of Dragendroff’s reagent were added. The orange precipitate confirms the presence of alkaloids.18
Bacterial Culture and Inoculation
S. mutans ATCC 25175 was pre-cultured in tryptic soy agar (TSA) media and incubated in an anaerobic atmosphere at 37°C for 24 h. V. parvula ATCC 10790T was precultured using defibrinated sheep blood agar and 2% lactic acid in an anaerobic atmosphere (90% N2, 5% H2, 5% CO2) at 37°C for 48 h.10 The culture results were then identified and subjected to gram staining using the HiMedia Gram Stains-Kit (Country of origin INDIA, company certified ISO 9001–2015 and ISO 13485–2016, WHO GMP). The preculture results were made into a suspension with turbidity according to the 0.5 McFarland standard (concentration 1–2 × 108 CFU/mL).
Bacterial inoculation was performed by collecting a bacterial suspension using a sterile cotton swab and spreading it evenly on Brain Heart Infusion agar + 1% sucrose, incubated in an anaerobic atmosphere at 37°C for 48 h.
Antibacterial Test
The antibacterial test was conducted using the disc diffusion method with the n-hexane fractions at concentrations of 50, 25, 12.5, 6.25, and 3.13% against the S. mutans and V. parvula bacteria. The negative control is ethanol 50% as a solvent and positive control is CHX 0.2%. The paper disc used had a diameter of 6 mm, was given 20 μL of the fraction solution, and was then placed on top of the prepared bacterial inoculum. The agar was then incubated under anaerobic conditions at 37°C for 48 h for S. mutans and V. parvula, and the inhibition zones were measured using calipers. The inhibition test was conducted with two repetitions.
Biofilm Formation
Biofilm formation was carried out using Brain Heart Infusion and 1% sucrose (BHIS) broth, which was placed in a 96-well flat-bottom plate. The bacteria used were single S. mutans, V. parvula, and double species S. mutans and V. parvula. Determination of the minimum bactericidal concentration (MBC) of the extract and fractions was performed using the broth serial dilution method with a 96-well flat-bottom plate. In well 1 of the 96-well flat-bottom plate, 100 µL of media was added as a negative control. In well 2, 100 µL of media and 10 µL of the test bacterial suspension were added. In wells 3–12, 50 µL of medium, 10 µL of the test bacterial culture, and 50 µL of the n-hexane fraction were added at up to 10 concentrations. Sample concentration is highest in well no. 3 and decreases to well no. 12. The test materials used had concentrations from 25% decreased to 10 concentrations below it. Microwells were incubated for a minimum of 48 h in an anaerobic atmosphere at 37°C. Samples were taken using an ose and streaked on agar media and incubated for a minimum of 48 h in an anaerobic atmosphere at 37°C to determine the MBC so that further tests could be conducted. The medium in each well was discarded, washed, transferred to a 1.5 mL Eppendorf tube using phosphate buffered saline (PBS), and stored at –20°C.19
Glucosyltransferase Analysis
Quantitative reverse transcription PCR (qRT-PCR) was performed to determine the mRNA expression levels of gtfB and gtfC genes. The protocols used for the isolation, purification, and reverse transcription of total bacterial RNA into cDNAs were described using the SensiFAST cDNA Synthesis Kit with catalog number BIO-65053: and batch number BIO-65054 (http://www.bioline.com/), with the following protocol used for the isolation, purification, and reverse transcription of total bacterial RNA into cDNA with the following composition: RNA template (1 ng to 1 µg) of 5 µL, 5× transcription buffer of 4 µL, reverse transcriptase of 1 µL, nuclease-free water (NFW) of 10 µL (added until the total sample volume reached 20 µL), mixed, and then placed into the thermocycler. The samples were stored at –20°C and for long-term storage at –80°C. Quantitative tests of gtfB and gtfC gene expression were conducted as for the quantitative DNA test using SensiFAST SYBR ®Hi-ROX Kit with batch number BIO-92005: (http://www.bioline.com/sensifast) with the following composition: (1) SYBR reagent 10 μL, (2) primer F 0.8 μL, (3) primer R 0.8 μL, (4) test sample with a concentration of 25 ng/μL sebanyak 3 μL, dan (5) NFW 1,2 μL. The total mixture used is 20 µL. The samples are placed into the MicroAmpTM Fast Optical 48-well Reaction Plate and covered with an optical adhesive film. They are then inserted into the PCR machine using the three-step cycling method with 40 cycles. The results were recorded by observing the CT values of each test sample, followed by data analysis. All primers used are listed in Table 1.
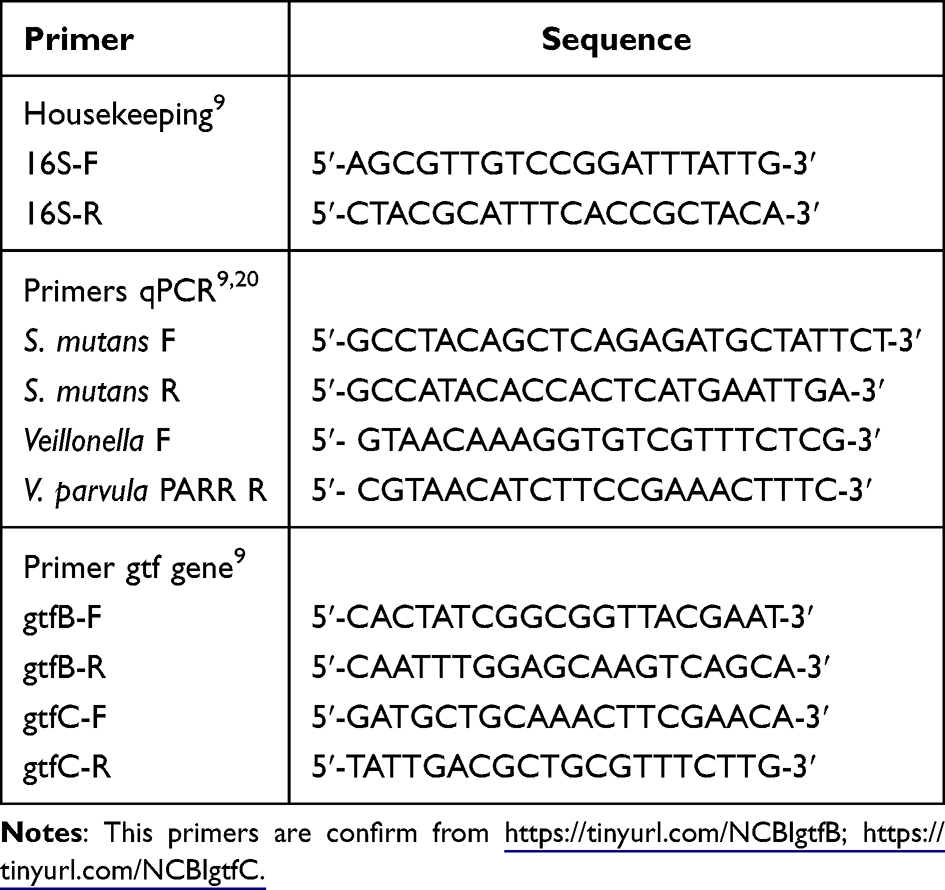 |
Table 1 List of Primers |
Statistical Analysis
Data were recorded and analyzed using a non-parametric test with the Kruskal–Wallis method, followed by the Mann–Whitney test to perform a difference test between two variables with a P- value of 0.05.
Research Time
The research conducted from January to July 2024.
Results
Phytochemical Screening
Phytochemical screening was used to assess the composition and predominance of constituents within the plant fraction. This study examined the secondary metabolites present in the n-hexane fraction of guava leaves (Table 2). This fraction contained phenols, tannins, flavonoids, terpenoids, steroids, and alkaloids. The antibacterial potential may be attributed to the presence of these phytochemical constituents.
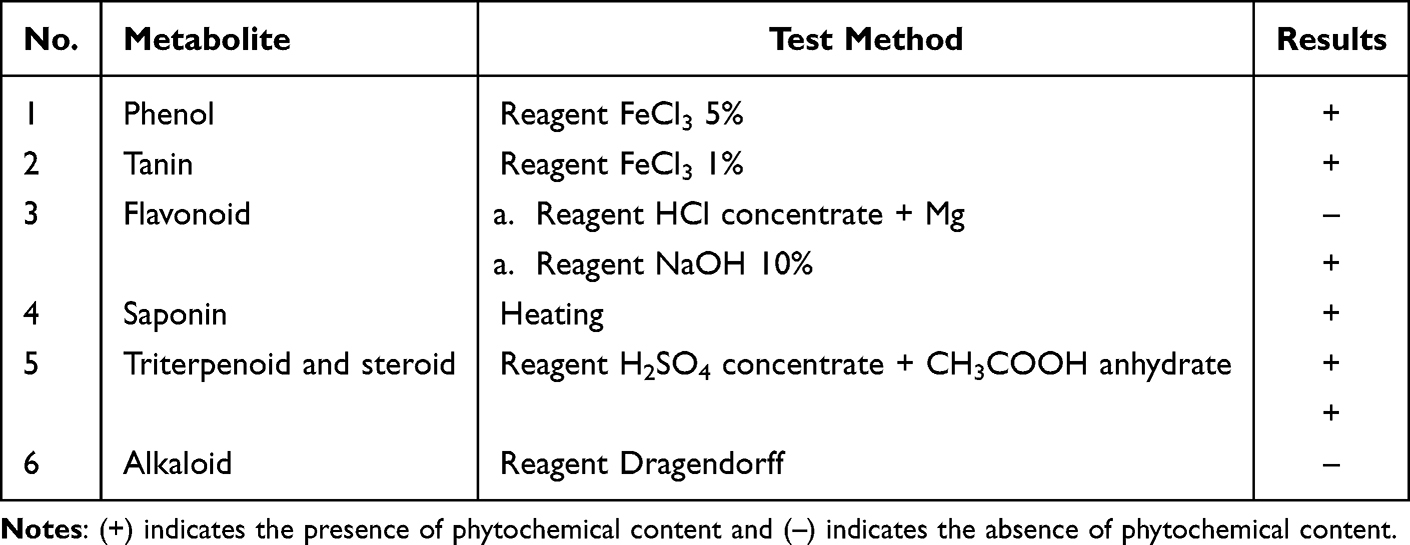 |
Table 2 Phytochemical Screening of n-Hexan Fraction |
Antibacterial Effects
The results of the antibacterial tests on the n-hexane fraction of P. guajava leaves against S. mutans and V. parvula are shown in Table 3. The n-hexane fraction with a concentration of 3.125% inhibits the growth of S. mutans and V. parvula with inhibition zones of 9.15±2.62 mm and 9.3±0.00 mm, respectively, but at higher concentrations, the inhibition zone produced is greater against S. mutans compared to V. parvula. The inhibitory effect of the n-hexane fraction of P. guajava leaves against V. parvula and S. mutans was lower than that of CHX (inhibitory zones: 17.1 ± 0.57 mm and 16.60 ± 0.57 mm). Figure 1 shows the zones of inhibition.
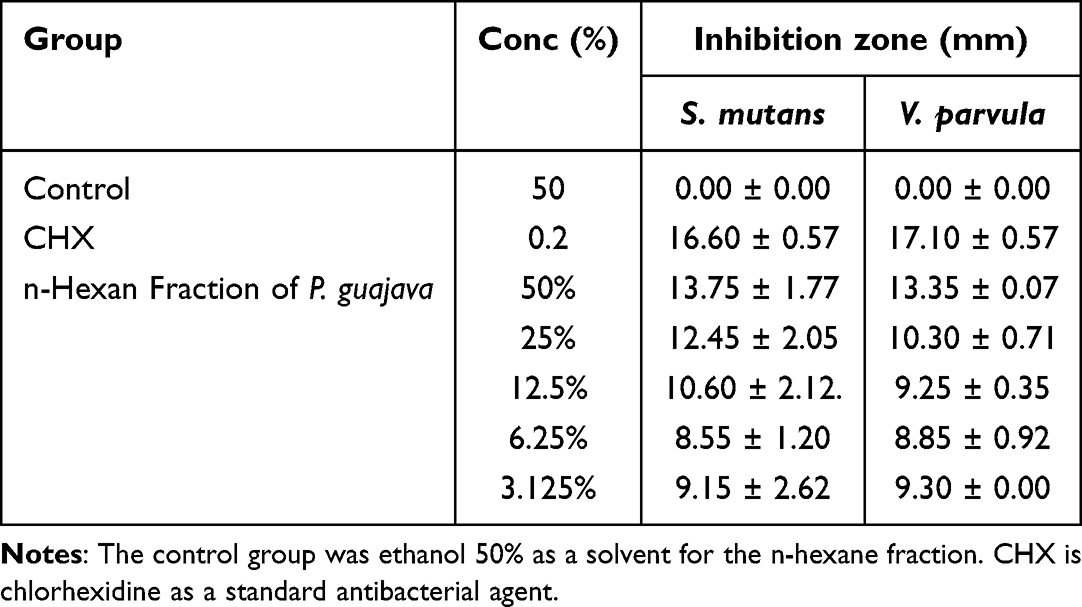 |
Table 3 Table of Inhibition Zone of P. guajava n-Hexan Fraction |
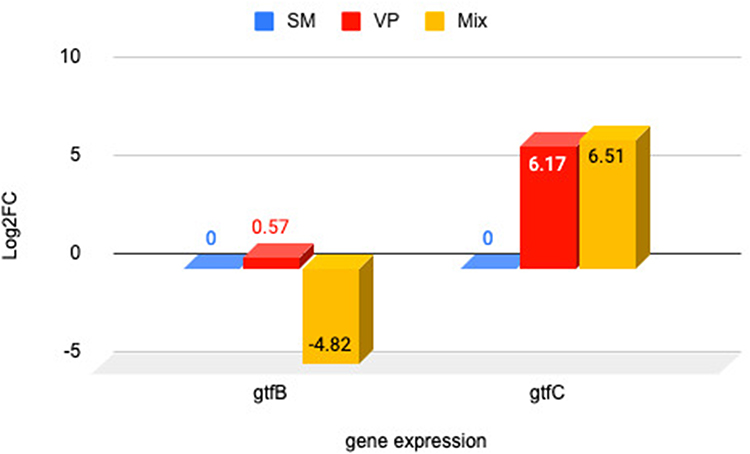 |
Figure 1 Diagram of changes in gtfB and gtfC gene expression in single species and double species V. parvula and S. mutans without n-hexane fraction treatment. Notes: Negative (–) value is indicated the downregulated of the gene expression fold change. Positive (+) value is indicated the upregulated of the gene expression fold change. |
The MBC of the n-hexane fraction of P. guajava leaves and CHX was determined by the lack of bacterial proliferation at the lowest concentration. The MBC of the n-hexane fraction on the single S. mutans and V. parvula bacteria was 1.56% and 0.78%, respectively, as no bacterial growth at these minimum concentrations. For both bacteria presented in the combination or dual-species, the MBC of the n-hexane fraction was 0.78%. Furthermore, administration of CHX as a standard antibacterial agent inhibited the growth of bacteria in single and dual species, whereas no antibacterial effect was observed in the control group. Table 4 shows the MBC of the n-hexane fraction of P. guajava leaves for single and dual species.
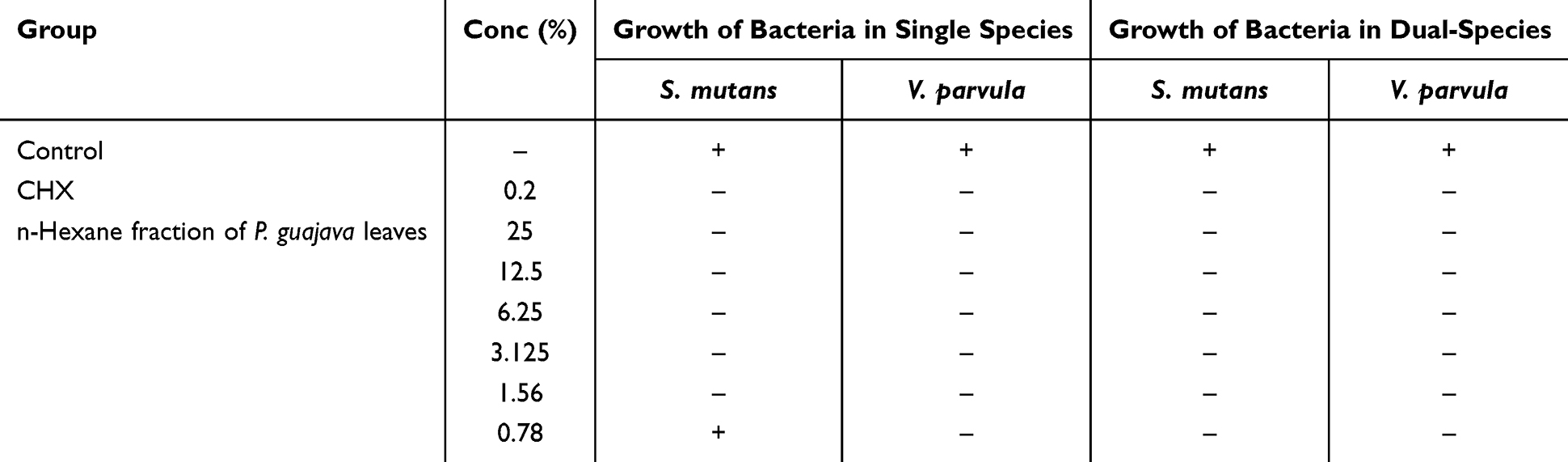 |
Table 4 Minimium Inhibitory Concentration of P. guajava n-Hexane Fraction of V. parvula and S. mutans |
Expression of gtfB and gtfC Genes
The gene expression of gtfB and gtfC was determined from the number of fold changes (log2FC) in the cDNA, with the control being untreated bacteria. An intuitive measure of the extent of the difference between groups is provided by fold change. The use of log2 enables the direct interpretation of the number of times the marker expression was greater or less in the treatment group in multiples of 2, as well as the number of folds by which the expression changes for both up- and downregulation.21 As seen in Figure 1, prior to the application of the n-hexane fraction, the gtfB gene expression level in the single V. parvula was observed to be 0.57 log2FC upregulated than that in the single S. mutans, whereas in the dual-species, the expression level of the gtfB gene was −4.82 log2FC downregulated. Theoretically, V. parvula cannot produce Gtfs because it does not metabolize carbohydrates directly. In this study, it was found that V. parvula can express the gtfB and gtfC genes, but not in large quantities. The theory remains unexplained due to the necessity for additional investigation.
In the administration of the n-hexane fraction, the expression of the gtfB gene remained unchanged in both single S. mutans and combined species, while it diminished by a factor −3.62 log2FC downregulated in the single V. parvula at a concentration of 0.78%. CHX showed increased on the levels of gtfB gene expression in either the single or combined species (Figure 2). The results were not significantly different between single S. mutans, V. parvula, and double species (P 0.387 >0.05), it is present at Table 5.
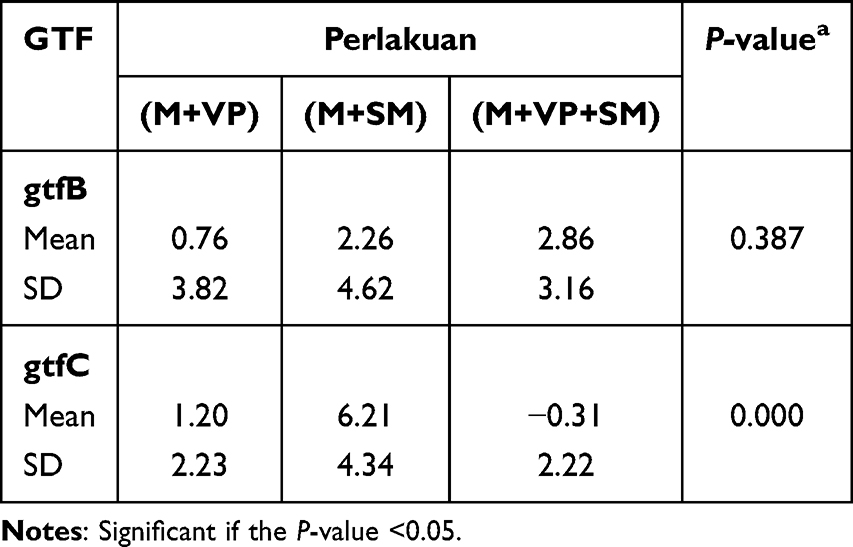 |
Table 5 Comparison of the Effect of n-Hexane Fraction on the Expression of gtfB and gtfC Genes in Single and Double Species |
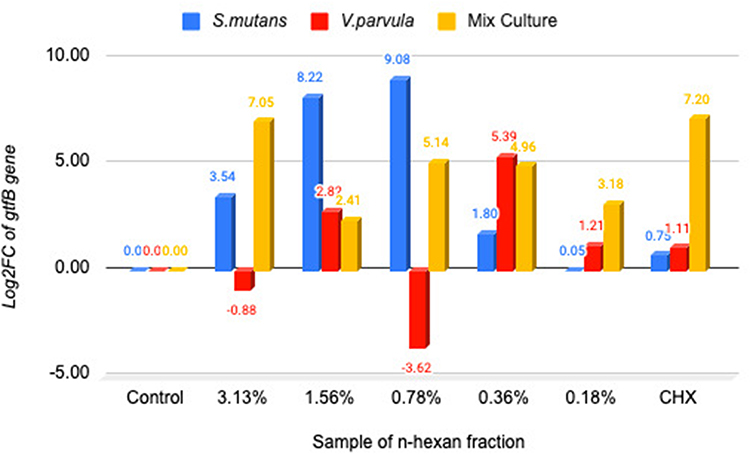 |
Figure 2 Diagram of changes in gtfB gene expression in single species and double species V. parvula and S. mutans by the n-hexane fraction. Notes: Negative (–) value is indicated the downregulated of the gene expression fold change. Positive (+) value is indicated the upregulated of the gene expression fold change. |
Before administration of the n-hexane fraction, gtfC gene expression showed a increase in the case of single V. parvula and double species relative to the single S. mutans. Application of the n-hexane fraction reduced the expression of the gtfC gene in the dual-species at a concentrations of 3.125, 0.78, and 0.18% by a factors of −2.49, −3.39, and −1.36 log2FC downregulated. However, CHX did not decrease gtfC expression in either the individual S. mutans or V. parvula (Figure 3). The results exhibited a significant differences among single S. mutans, V. parvula, and dual species to the expression of gtfC (P 0.00 <0.05), it is present at Table 5
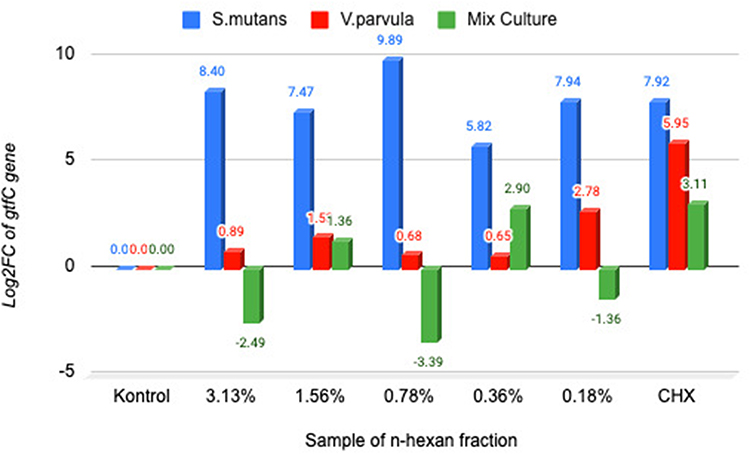 |
Figure 3 Diagram log2FC gtfC in single species V. parvula ATCC and S. mutans ATCC as well as double species. Notes: Negative (–) value is indicated the downregulated of the gene expression fold change. Positive (+) value is indicated the upregulated of the gene expression fold change. |
Discussion
S. mutans is a pathogenic microbe capable of forming oral biofilms, resulting in tooth caries, bacteremia, and bacterial endocarditis.22 V. parvula is an early colonizer of oral biofilm formation that may metabolize lactic acid produced by S. mutans and possesses the capability to establish hard tooth surfaces, potentially resulting in tooth demineralization.11 The development of biofilms and caries can be reduced using antiplaque and antiseptic agents; however, their use is now limited to lowering the risk of developing resistance to multiple drugs.22
P. guajava leaves have been reported to exhibit antioxidant, hepatoprotective, anti-inflammatory, antispasmodic, anticancer, antimicrobial, antihyperglycemic, analgesic, and antibacterial effects.12 In this study, we investigated the antibacterial effect of the n-hexane fraction of P. guajava leaves on single S. mutans and P. parvula and their combination or dual-species. The n-hexane fraction was examined for its inhibitory effect on Gtfs activity, which contributes to biofilm formation. Gtfs with their ability to catalyze the synthesis of various glucans, are categorized into several types based on their enzymatic activity, including GtfB and GtfC, which play important roles in the formation of dental biofilms.3,19
As shown in Table 2, the n-hexane fraction of P. guajava leaves inhibited the growth of S. mutans and V. parvula at all concentrations tested. A previous study reported that P. guajava leaf extract strongly inhibited S. mutans,22 which is in accordance with this result. However, the antibacterial activity of P. guajava leaf extract against V. parvula has not yet been reported. In this study, the n-hexane fraction had a greater inhibitory effect against V. parvula than against S. mutans, as observed in their inhibition zones (Figure 1). These findings suggest that the n-hexane fraction of P. guajava leaves may inhibit V. parvula rather than S. mutans.
The antibacterial activity of the n-hexane fraction might be due to the fact that the n-hexane fraction contains lipophilic active compounds that can easily penetrate the lipid-bilayer in the cell membrane. This leads to membrane impairment and efflux of intracellular substances,23 which cause bacterial growth inhibition or death. S. mutans is a gram-positive bacterium characterized by a peptidoglycan-rich cell wall that facilitates absorption of antibacterial drugs. Conversely, the outer layer of the cell wall of gram-negative bacteria contains a lipopolysaccharide layer, which may block the penetration of antimicrobial agents into the bacteria.24 V. parvula is a gram-negative bacterium characterized by lipopolysaccharide coating in its cell wall; therefore, in this study the inhibitory zone produced by the n-hexane fraction of P. guajava on V. parvula was larger than that produced by S. mutans. This condition can occur by the non-polar properties of n-hexane fraction, that it has hydrophobic bioactive compounds are insoluble in water. This fraction is effective against both gram-positive and gram-negative bacteria, but its antibacterial activity is more limited depending on the bacterial conditions. The target of its action is the lipid layer of the cell membrane, causing membrane damage and leakage into the intracellular space.23
Furthermore, the MBC of the n-hexane fraction determined using the broth microdilution method was 1.56% and 0.78% for S. mutans and V. parvula bacteria, respectively, and 0.78% for the dual species. The concentration used to examine Gtf inhibition was 3.125%, which was determined from the MBC value increasing three levels.
Multiple variables, including glucose availability, ambient pH, and growth rate or phase, affect the transcription, translation, and secretion of the gtf gene. A reduction in pH resulting from the presence of carbohydrates, an elevation in metabolizable carbohydrates identified by the phosphotransferase system, or the synthesis of glycolytic intermediates can modulate the expression of the gtf gene. Regulation of the expression of the gtfB and gtfC genes as virulence factors does not occur randomly, but is instead regulated by a two-component signaling system, one of which is VicRK. Previous studies have shown that VicR directly increases the expression of gtfB and gtfC, two genes involved in the synthesis of glucans necessary for biofilm formation and maturation. Therefore, the activation of VicRK is usually associated with increased virulence, especially in the context of biofilm formation and resistance to acidic environments.8
In assessing how the n-hexane fraction influenced gene expression of gtfB activity, it was found that this fraction did not decreased the gene expression of gtfB activity in either the single S. mutans or in the dual species. However, in the case of V. parvula, gtfB gene expression was decreased by −3.62 log2FC at the concentration of 0. 78%. In contrast, gene expression of gtfC activity in the dual-species was −2.49, −3.39, and −1.36 log2FC downregulated by the n-hexane fraction at the concentrations of 3.125, 0.78, and 0.18%. Moreover, CHX, a standard antibacterial agent, did not alter the expression levels of gtfB and gtfC in either single or combined species.
This fact shows that the n-hexane fraction has a specific effect in disrupting virulence gene expression in the context of interspecies interactions, not just against S. mutans individually. According to the established mechanism, VicRK directly governs the expression of gtfB and gtfC, suggesting that the reduction in the expression of these genes – especially gtfC in dual-species – may be associated with the suppression of the VicRK system’s activation.19 However, in this study, the expression of the VicRK gene itself has not been directly analyzed, so this correlation remains hypothetical.
The expression of the gtf gene will be related to the production of the gtf enzyme itself. It has been known that Gtfs, mainly GtfB and GtfC, produced by S. mutans contribute to the biofilm formation This enzyme enables the conversion of sucrose into adhesive glucan. GtfB and GtfC are responsible for the formation of water-insoluble glucans, while GtfD synthesizes water-soluble glucans.19 Gtfs produced by S. mutans exert a greater influence on the development and composition of biofilms than on the bacterial populations present within the biofilm. Gtfs can also be associated with various oral cavity bacteria that do not produce Gtfs, thereby transforming non-Gtf-producing bacteria into glucan-producing bacteria.3,19 The results of this study indicate that the formation of GtfB and GtfC in the biofilm generated by V. parvula may be due to the ability of Gtfs produced by S. mutans to be adsorbed onto V. parvula, thereby enabling the production of glucan.
The initial phase in inhibiting biofilm formation is the planktonic stage. In this phase, the bacteria begin to multiply and adhere to the tooth surface. This process leads to bacterial colonization, which promotes biofilm formation in the subsequent phases. Antibacterial agents are required to avoid hydrophobic bonding as they help prevent biofilm formation or dental caries. This is important because S. mutans can adhere to the surface of tooth enamel through hydrophobic interactions.22
Previous studies have reported that the n-hexane fraction of P. guajava contains hydrophobic or lipophilic compounds, such as triterpenoids (β-sitosterol, lupeol, and betulinic acid), lipophilic flavonoids (trace amounts of apigenin and quercetin derivatives), acids and esters (palmitic acid, oleic acid, and stearic acid), phenols (trace amounts of lipophilic phenols), essential oils (caryophyllene, humulene, and limonene), and long-chain hydrocarbons (n-hexane derivatives).20 Phytochemical screening of the n-hexane fraction in our study revealed the presence of phenols, tannins, flavonoids, triterpenoids, and alkaloid groups, which is in accordance with previous research on the phytochemical composition of P. guajava. The findings of this study suggest that P. guajava leaves contain bioactive components, including tannins, alkaloids, polyphenols, and flavonoids, which are recognized for their antibacterial properties in medicinal plants and can exhibit activity against gram-positive and gram-negative microorganisms.13,25
Tannins exhibit effective antibacterial activity owing to the presence of tannic acid. The antibacterial action is caused by the disruption of the bacterial membrane lipid layer, resulting in the release of cellular fluid and subsequent cell destruction. Polyphenols exhibit antibacterial properties against human oral cavity infections by directly targeting pathogens and suppressing virulence factors. Polyphenols synergistically enhance the effectiveness of drugs against drug-resistant bacteria.13
The compounds present in P. guajava can influence the Gtfs enzyme responsible for biofilm development. The results of this study indicated a reduction in the Gtfs enzyme following the application of P. guajava leaves.13 The antibacterial activity of the extract against planktonic oral microbes was the basis for selecting this plant. The purpose of the test was to observe the attachment of early colonization of plaque and pellicle formation. The adhesion of early plaque colonization to the pellicle was substantially inhibited by P. guajava. Furthermore, it interferes with the early phases of biofilm formation. According to recent studies, P. guajava exhibits remarkable biofilm inhibition and antibacterial activity against S. mutans.22
Primary analysis of the interaction between these bioactive compounds and the active site of Gtfs was conducted using a docking simulation technique. These results clearly demonstrate that the phytochemical compounds isolated from P. guajava also interact with the sucrose active site of Gtfs and interfere with sucrose binding. Consequently, these phytochemical compounds can inhibit the formation of glucan, a primary cariogenic compound. The therapeutic potential of P. guajava leaves is evident in the comprehensive results of this study, which demonstrate its efficacy against the cariogenic pathogen S. mutans.13
The hexane fraction of P. guajava also contains fatty acids that affect Gtfs; however, the magnitude of this effect is not higher than that of flavonoids.13 The activity of Gtfs enzymes is disrupted by long-chain unsaturated fatty acids (oleic acid). This is due to the capacity of fatty acids to interact with the hydrophobic regions of the enzyme, which in turn affects its structure and function. Saturated fatty acids, including palmitic acid, have less impact; however, they can still disrupt enzymatic activity under certain circumstances. A study demonstrated that glucan synthesis is inhibited by fatty acids derived from plant extracts, such as guava, as a result of reduced Gtfs activity.13
This study revealed that the n-hexane fraction of P. guajava affects the expression of gtfC. GtfC is a glucan that is both soluble and insoluble in water, and possesses hydrophobic properties. GtfC also develops during the initial phase of biofilm formation. Previous studies have demonstrated that components of the n-hexane fraction of P. guajava can prevent the attachment of bacteria to the tooth surface, attachment of bacteria to hydrophobic enzymes, and the development of biofilms at an early stage.22 This aligns with the study findings, indicating that the n-hexane fraction diminishes the expression level of the gtfC gene, which encodes the hydrophobic Gtfs enzyme and is involved in the initial phases of biofilm development.
The limitations of this study are the small number of samples tested and the inability to determine the minimum inhibitory concentration of the biofilm owing to the dark color of the hexane fraction, thus biasing the test results.
Conclusion
V. parvula affects the formation of glucosyltransferase enzymes in S. mutans, marked by changes in the expression of gtfB and gtfC genes in the double species biofilm. The expression of the gtfB gene decreases in the double species of S. mutans and V. parvula compared to the expression of the gtfB gene in single species S. mutans, while the expression of the gtfC gene increases with the presence of V. parvula.
The n-hexane fraction of guava leaves has the ability to decrease the expression level of the gtfC gene in double species when compared to single S. mutans. However, the n-hexane fraction is unable to reduce the expression level of the gtfB gene in either single or double species. It can be inferred that the n-hexane fraction has the capacity to impede the formation of biofilm adhesion on tooth surfaces, and this inhibition takes place during the planktonic phase.
Acknowledgments
We thank the Spices and Medicinal Plants Research Centre (BALITTRO) in Lembang, West Java, Indonesia, for granting us the opportunity to collect the samples. We also thank the School of Life Sciences and Technology at the Institute of Technology of Bandung for performing the sample analyses. We are grateful to the Faculty of Dentistry at the University of Indonesia for granting access to the Oral Biology Laboratory for conducting the experiments.
Funding
This research received an Internal Research Grant Padjadjaran University Doctoral Dissertation Research scheme by Directorate of Research and Community Engagement Padjadjaran University with grant number 1978/UN6.3.1/PT.00/2024.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Berger D, Rakhamimova A, Pollack A, Loewy Z. Oral biofilms: development, control, and analysis. High Throughput. 2018;7(3):1–8. doi:10.3390/ht7030024
2. Ruhal R, Kataria R. Biofilm patterns in gram-positive and gram-negative bacteria. Microbiol Res. 2021;251(March):126829. doi:10.1016/j.micres.2021.126829
3. Bowen WH, Koo H. Biology of streptococcus mutans-derived glucosyltransferases: role in extracellular matrix formation of cariogenic biofilms. Caries Res. 2011;45(1):69–86. doi:10.1159/000324598
4. Bowen WH, Burne RA, Wu H, Koo H. Oral biofilms: pathogens, matrix, and polymicrobial interactions in microenvironments. Trends Microbiol. 2018;26(3):229–242. doi:10.1016/j.tim.2017.09.008
5. Spormann AM, Thormann K, Saville R, Shukla S, Entcheva P. Microbial biofilms. Nanoscale Technol Bioll Syst. 2004;341–357.
6. Lin NJ. Biofilm over teeth and restorations: what do we need to know? Dent Mater. 2017;33(6):667–680. doi:10.1016/j.dental.2017.03.003
7. Samaranayake L. Essential Microbiology for Dentistry.
8. Zhang Q, Ma Q, Wang Y, Wu H, Zou J. Molecular mechanisms of inhibiting glucosyltransferases for biofilm formation in Streptococcus mutans. Int J Oral Sci. 2021;13(1):1–8. doi:10.1038/s41368-021-00137-1
9. Luppens SBI, Kara D, Bandounas L, et al. Effect of Veillonella parvula on the antimicrobial resistance and gene expression of Streptococcus mutans grown in a dual-species biofilm. Oral Microbiol Immunol. 2008;23(3):183–189. doi:10.1111/j.1399-302X.2007.00409.x
10. Liu S, Chen M, Wang Y, et al. Effect of Veillonella parvula on the physiological activity of Streptococcus mutans. Arch Oral Biol. 2020;109(September 2019):104578. doi:10.1016/j.archoralbio.2019.104578
11. Wei Y, Zhang Y, Zhuang Y, et al. Veillonella parvula acts as a pathobiont promoting the biofilm virulence and cariogenicity of Streptococcus mutans in adult severe caries. Microbiol Spectrum. 2024;12(11). doi:10.1128/spectrum.04318-23
12. Dintenfass L, Sharp A. Efficacy of Psidium guajava leaf extract on Streptococcus mutans and Enterococcus faecalis – an in vitro study. Ann Surg. 2019;07(05):984–995.
13. Bhagavathy S, Mahendiran C, Kanchana R. Identification of glucosyl transferase inhibitors from Psidium guajava against Streptococcus mutans in dental caries. J Tradit Complement Med. 2019;9(2):124–137. doi:10.1016/j.jtcme.2017.09.003
14. Razak FA, Othman RY, Rahim ZHA. The effect of Piper betle and Psidium guajava extracts on the cell-surface hydrophobicity of selected early settlers of dental plaque. J Oral Sci. 2006;48(2):71–75. doi:10.2334/josnusd.48.71
15. Nn A. A review on the extraction methods use in medicinal plants, principle, strength and limitation. Med Aromat Plants. 2015;04(03):3–8.
16. Abubakar AR, Haque M. Preparation of medicinal plants: basic extraction and fractionation procedures for experimental purposes. Asian J Pharm Clin Res. 2017;7(10):1–5.
17. Wiener MC, Horanyi PS. How hydrophobic molecules traverse the outer membranes of Gram-negative bacteria. Proc Natl Acad Sci U S A. 2011;108(27):10929–10930. doi:10.1073/pnas.1106927108
18. Dubale S, Kebebe D, Zeynudin A, Abdissa N, Suleman S. Phytochemical screening and antimicrobial activity evaluation of selected medicinal plants in Ethiopia. J Exp Pharmacol. 2023;15(February):51–62. doi:10.2147/JEP.S379805
19. Ren Z, Cui T, Zeng J, et al. Molecule targeting glucosyltransferase inhibits Streptococcus mutans biofilm formation and virulence. Antimicrob Agents Chemother. 2016;60(1):126–135. doi:10.1128/AAC.00919-15
20. Afzal M, Iqbal R, Mahmood Z, Zeshan B, Wattoo JI. Study of GC-MS and HPLC characterized metabolic compounds in Guava (Psidium guajavaL.) leaves. Pak J Agric Sci. 2019;56(3):709–713.
21. Lötsch J, Kringel D, Ultsch A. Revisiting fold-change calculation: preference for median or geometric mean over arithmetic mean-based methods. Biomedicines. 2024;12(8):1639. doi:10.3390/biomedicines12081639
22. Gomashe AV, and SAA, Kasulkar A. Original research article investigation of biofilm inhibition activity and antibacterial activity of Psidium guajava Plant extracts against Streptococcus mutans causing dental plaque. Int J Curr Microbiol Appl Sci. 2014;3(9):335–351.
23. Alamin MA, El Samia Badwi MA, Alqurashi AM, Elsheikh AS. Bactericidal activity of Psidium guajava leaves against some pathogenic microbes. IOSR J Dental Med Sci. 2016;15(3):61–70.
24. Biswas B, Rogers K, McLaughlin F, Daniels D, Yadav A. Antimicrobial activities of leaf extracts of Guava (Psidium guajava L.) on two gram-negative and gram-positive bacteria. Int J Microbiol. 2013;2013:1–7. doi:10.1155/2013/746165
25. Joseph B, Priya RM, Helen PAM, Sujatha S. Bio-active compounds in essential oil and its effects of antimicrobial, cytotoxic activity from the Psidium Guajava (L.) leaf. J Adv Biotechnol. 2010;9(2000):10–14.
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.