Back to Journals » Drug Design, Development and Therapy » Volume 20
Proteome Microarray-Guided Global View: Multiple Pharmacological Targets of Icariin
Authors Gong G, Tang X, Yuan G, Yang X, Wang G, Zhang F ![]()
Received 14 November 2025
Accepted for publication 24 January 2026
Published 3 February 2026 Volume 2026:20 581728
DOI https://doi.org/10.2147/DDDT.S581728
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Anastasios Lymperopoulos
Guichun Gong,1,* Xianjin Tang,1,* Guanghun Yuan,1 Xinxing Yang,2 Guoqing Wang,1 Feng Zhang1,2
1Key Laboratory of Basic Pharmacology of Ministry of Education and Joint International Research Laboratory of Ethnomedicine of Ministry of Education and Key Laboratory of Basic Pharmacology of Guizhou Province and Laboratory Animal Center, Zunyi Medical University, Zunyi, Guizhou, 563000, People’s Republic of China; 2State Key Laboratory of Discovery and Utilization of Functional Components in Traditional Chinese Medicine, Guizhou Medical University, Guiyang, Guizhou, 561113, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Feng Zhang, Email [email protected]
Background: Icariin (ICA) is a major bioactive compound extracted from the traditional Chinese medicinal herb Epimedium, demonstrated a broad spectrum of pharmacological properties. However, a comprehensive and up-to-date study exclusively focusing on ICA direct binding proteins has not yet been conducted.
Methods: HuProt™ 20K human proteome microarray, the highest capacity human protein microarray, is suitable for the applications of small molecule targeting proteins. Here, we constructed an atlas of the ICA-binding proteins directly via proteome microarray.
Results: We showed that a total of 246 proteins which directly interacted with ICA. Subsequent PPI network analysis organized these proteins into 4 functionally distinct clusters, revealing that ICA interacts with proteins central to fundamental cellular processes, including protein folding chaperone complexes, the ubiquitin-proteasome system, apoptotic and PI3K-Akt signaling pathways, and nucleotide metabolism.
Conclusion: This study constructed an atlas of the ICA-binding proteins directly via microarray, which spans protein folding, ubiquitin-proteasome, apoptosis and nucleotide metabolism, providing a theoretical basis for ICA treatment of diseases caused by dysregulation of these core pathways.
Keywords: icariin, proteome microarray, pharmacological targets, protein networks, molecular mechanism
Introduction
Icariin (ICA) is a major bioactive compound extracted from the traditional Chinese medicinal herb Epimedium, commonly known as Horny Goat Weed or Yin Yang Huo. It is a prenylated flavonol glycoside with the chemical formula C33H40O15 and a molecular weight of 676.67 g/mol. Moreover, ICA can be metabolized by enzymes into glucuronide conjugates of flavonoid aglycones and isoflavonoids.1,2 Several researches have broadly explored the diverse pharmacological effects of ICA to articulate its traditional medicinal applications. ICA exerted a broad spectrum of pharmacological properties, including anti-oxidant properties, anti-cancer properties, anti-inflammatory properties, anti-depressant, and neuroprotective effects.3,4 However, a comprehensive and up-to-date study specifically focusing on ICA direct binding proteins has not yet been conducted.
The HuProt™ 20K Human Proteome Microarray contains over 21,000 unique human proteins, covering approximately 81% of the classical human proteome. These proteins are derived from 16,793 genes, including 15,889 classical human proteins described in the Human Protein Atlas. The proteins on the microarray exist in the correct three-dimensional structure for specific binding of small molecules, enabling highly sensitive detection of small molecule binding targets.5,6
Here, we used the HuProt 20K human proteome microarray to identify the binding proteins of ICA by biotinylating ICA, incubating it with the microarrays, and the binding capacity were detected with Cy5-conjugated streptavidin (Cy5-SA). In the end, we identified a total of 246 proteins that directly interacted with ICA. In addition, the above 246 candidate proteins were clustered using protein-protein interactions (PPI). Specifically, the top 4 ranked protein network clusters, relationship between diseases were presented. This article provides a valuable resource for ICA-binding proteins.
Materials and Methods
Reagents
AS (T2855, CAS 489–32-7) was purchased from TargetMol (USA). HuProt™ 20K human proteome microarray was obtained from the Johns Hopkins Medical Institutions Protein Microarray Core (CDI Laboratories, Inc).
HuProt™ 20K Human Proteome Microarray
HuProt™ 20K human proteome microarray assay was conducted by Wayen Biotechnologies (Shanghai, China). ICA was prepared as a 10 mM stock solution in DMSO and diluted in 1× PBS-T to a final working concentration of 10 µM for the assay. The HuProt array was equilibrated from −80°C and then blocked with 5% BSA in PBS-T on a shaker at room temperature, using an initial 5 min step followed by a 1.5 h incubation in the dark. After a brief wash, the array was incubated with the ICA solution for 1 h under the same conditions. Unbound compound was removed by 3 washes with PBS-T and 2 rinses with deionized water. For detection, the array was incubated with a 0.1% Cy5-Streptavidin solution for 20 min, followed by an identical wash series, and then dried by centrifugation. Fluorescent signals were captured by scanning at 635 nm using a GenePix 4000B scanner.5,6
Data Processing and Hit Identification
Raw fluorescence data were extracted from the scanned HuProt™ 20K microarray using GenePix™ Pro v6.0 software. To normalize data across the array and identify significant binders, a Z-score was computed for each individual protein spot. The Z-score for a given spot was defined as:
Z-score = (I - M) / SD. Where “I” represented the ratio of the median fluorescence signal value at the given point to the median background signal value at all points, “M” was the median of all I values, and SD was the standard deviation of all I values. A protein was considered a primary candidate hit only if both of its technical replicates surpassed a pre-defined Z-score significance threshold (Z-score ≥ 2.8). This refined list was defined as the set of high-confidence, specific ICA-binding proteins for downstream bioinformatic analysis.7
Network Analysis
To explore functional associations among the 246 high-confidence binding proteins, a PPI network was generated using the Search Tool for the Retrieval of Interacting Genes/Proteins (STRING, Version: 12.0) database, querying for Homo sapiens with a high-confidence interaction score threshold ≥ 0.70. The resulting network was imported into Cytoscape for visualization and analysis. Highly interconnected functional modules were identified using the MCODE (Molecular Complex Detection) plugin, and the top 4 ranked clusters by MCODE score were selected for subsequent pathway and enrichment analysis.
Functional Enrichment Analysis
To decipher the biological roles and pathway associations of the identified protein clusters, Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analyses were performed. The clusterProfiler R package was utilized for both analyses. Enrichment significance was assessed based on the hypergeometric distribution, and p-values were adjusted via the Benjamini-Hochberg (FDR) method. Terms and pathways with an FDR < 0.05 were deemed statistically significant.
Cellular Thermal Shift Assay (CETSA)
HEK293T cells were treated with ICA (10 μM) or vehicle control (DMSO) for 24 h. Cell lysates were divided into aliquots and heated at the indicated temperatures (45, 50, 55, 60, 65, 70, 75, 80, 85 °C) for 3 min. After 3 min at room temperature and cooling on ice, samples were centrifuged (12,000 rpm, 20 min, 4 °C). The resulting supernatants were prepared with loading buffer for Western Blot analysis.
Statistical Analysis
Results were reported as mean ± standard error of the mean (SEM). Differences among groups were compared by one-way analysis of variance (ANOVA) using GraphPad Prism software. Statistical significance was defined as P < 0.05.
Results
Global Profiling of ICA-Binding Proteins Using HuProt™ 20K Proteome Microarray
To identify ICA-binding proteins, we probed a human proteome microarray with a biotinylated ICA molecule. Briefly, the ICA-biotin conjugate (Biotin-ICA) was incubated with the human proteome microarray, and proteins with ICA-binding capacity were identified by adding Cy5-SA. To avoid false-positive detections, we compared this sample with another in which Biotin-ICA was omitted from the reaction and free biotin was included instead and identified as ICA-binding candidate proteins those exhibiting greater signals in the former reaction. In the end, we identified a total of 246 proteins that directly interacted with ICA. The protein name, ID and Z-score were listed in Table 1.
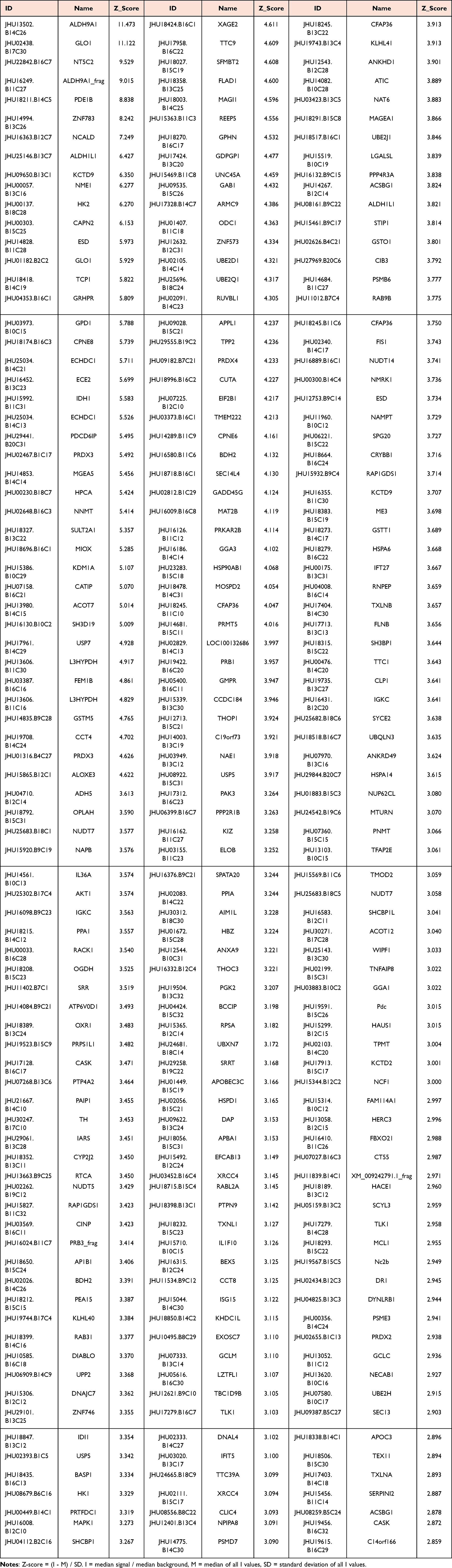 |
Table 1 The Pharmacological Targets of Icariin in HuProt™ 20K Human Proteome Microarray |
Interactions Network of ICA-Binding Proteins
To analysis the interaction network of ICA-binding proteins completely, the above 246 candidate proteins were clustered using PPI. The top 4 ranked protein network clusters were presented with confidence scores ≥ 0.7 and labeled by red, green, purple and yellow circle, respectively. Among them, sorting by descending order, the first-ranked network was protein folding chaperone (Score= 8.2), including PSMD7, TCP1, STIP1, RPSA, CCT8, HSPA6, CCT4, RUVBL1, DNAJC7, HSPA14, HSPD1. The second-ranked network was the cellular response to cadmium ion (Score= 4.667), including PSMB6, ELOB, UBE2D1, UBXN7, RACK1, HSP90AB1, PSME3. The third-ranked network was oxygen-dependent proline hydroxylation of hypoxia-inducible factor alpha (Score= 3.667), including PPP2R1B, TH, DIABLO, NCF1, MCL1, AKT1, MAPK1. The fourth-ranked network was ADP-ribose diphosphatase activity (Score=3), including NUDT7, NUDT14, NUDT5 (Figure 1).
 |
Figure 1 Interactions network of ICA-binding proteins. Protein-protein interaction (PPI) network of ICA-binding proteins. All proteins were divided into four significant protein clustering modules. Red dots represented cluster 1 proteins. Green dots represented cluster 2 proteins. Purple dots represented cluster 3 proteins. Yellow dots represented cluster 4 proteins. |
Relationship Between Cluster 1 Proteins and Diseases
Cluster 1 proteins included 11 nodes (PSMD7, TCP1, STIP1, RPSA, CCT8, HSPA6, CCT4, RUVBL1, DNAJC7, HSPA14, and HSPD1) and 41 interaction pairings with 8.2 score (Figure 2A). In addition, the enlarged-view, ID, name and Z-score for Cluster 1 proteins were also presented (Figure 2B). Furtherly, KEGG analysis showed that ICA-binding proteins were involved in legionellosis, prion disease and lipid and atherosclerosis (Figure 2C). Next, GO analysis indicated that in terms of the biological process, ICA-binding proteins were mainly enriched in chaperone-mediated protein folding and protein folding. In terms of cellular components, ICA-binding proteins were mainly enriched in protein folding chaperone complex. According to molecular function, ICA-binding proteins were mostly enriched in ATP-dependent protein folding chaperone, ATP hydrolysis activity, protein folding chaperone and unfolded protein binding (Figure 2D).
 |
Figure 2 Bioinformatics analysis of Cluster 1 proteins. (A) PPI network of Cluster 1 Proteins. (B) Enlarged-view and Z-score of Cluster 1 Proteins in proteome microarray. (C) KEGG analysis of Cluster 1 Proteins. (D) GO analysis of Cluster 1 Proteins. |
PSMD7 (26S proteasome non-ATPase regulatory subunit 7) is a core component of the 26S proteasome essential for ubiquitin-dependent protein degradation and promotes tumor progression in multiple cancers. In pancreatic cancer by deubiquitinating and stabilizing SOX2 to activate Notch1 signaling.8 In breast cancer, where its knockdown induces cell cycle arrest and apoptosis.9 In gastric cancer, where it is activated by FOXP3 and enhances proliferation and cisplatin resistance via RAD23B stabilization.10 Additionally, the PSMD7 polymorphism rs17336700 is associated with ankylosing spondylitis.11
TCP1 (T-complex protein 1), overexpressed in multiple cancers and linked to poor prognosis, promotes tumor progression via key pathways. In hepatocellular and pancreatic ductal adenocarcinoma, it drives proliferation by stabilizing c-Myc through AKT/GSK-3β/ERK and the Wnt7b/β-catenin pathway.12,13 Furthermore, it contributes to progression and drug resistance in other cancers partly via the PI3K/AKT/mTOR axis.14 In addition, in diffuse large B-cell lymphoma, it modulates ferroptosis sensitivity by stabilizing ACSL4.15
STIP1 (Stress-induced phosphoprotein 1) is a co-chaperone that regulates protein folding and quality control by interacting with HSPs. STIP1 plays roles in both oncogenesis and neurodegeneration. Its overexpression in ovarian, thyroid, liver, colorectal, and lung cancers correlates with poor prognosis, where it promotes tumor progression via mechanisms such as ALK2/SMAD pathway activation in ovarian cancer and likely JAK2/STAT3 signaling in others.16,17 Beyond cancer, STIP1 is linked to Parkinson’s disease (PD) through specific autoantibodies and shows neuroprotective effects in Alzheimer’s disease (AD) by mitigating β-amyloid toxicity.18
RPSA (Small ribosomal subunit protein uS2) is a ribosomal component essential for protein synthesis. It functions as a receptor for pathogens such as streptococcus pneumoniae and dengue virus, facilitating cellular adhesion and invasion with implications for infections like bacterial meningitis.19,20 In cancer, its overexpression is linked to increased tumor cell migration, invasion, and angiogenesis, partly through regulating telomerase and interacting with PrPC to promote oncogenic processes.21 Furthermore, altered RPSA expression is implicated in rheumatoid arthritis and may contribute to inflammatory responses during acute infections.22
CCT8 (Chaperonin containing TCP1 subunit 8), a subunit of the chaperonin complex, facilitates protein folding and is implicated in various diseases. CCT8 is involved in lung adenocarcinoma through AKT activation and in colorectal cancer by antagonizing p53.23 It correlates with poor prognosis and has also been associated with glioma and breast cancer. Beyond oncology, CCT8 facilitates tobamovirus spread24 and is essential for balanced T-cell immunity, with its deficiency skewing the immune response toward a Th1 phenotype and implicating it in immune pathology.25
HSPA6 (Heat shock protein family A member 6) is a heat shock protein that functions as a molecular chaperone. It plays a role in infectious diseases and cancer. Specifically, HSPA6 is induced upon Enterovirus 71 (EV71) infection, during which it colocalizes with the viral protein VP1 and may act as a host-specific factor.26 In glioblastoma, high HSPA6 expression is associated with poor prognosis, promoting tumor progression through enhanced proliferation, invasion, and resistance to apoptosis. Interestingly, HSPA6 may also function as a tumor suppressor in bladder cancer, lung cancer, and triple-negative breast cancer.27
CCT4 (Chaperonin containing TCP1 subunit 4) is a chaperonin involved in protein folding and has been implicated in several diseases. In hepatocellular carcinoma (HCC), CCT4 is overexpressed and interacts with Cdc20 to promote tumorigenesis.28 In hereditary sensory neuropathy (HSN), the CCT4 point mutation C450Y compromises protein stability.29 Meanwhile, Bardet-Biedl syndrome is associated with defects in the MKKS gene, which encodes a CCT-like protein.30 Beyond these specific conditions, CCT4 also regulates organ growth by modulating insulin/TOR signaling, a pathway whose dysregulation contributes to systemic disorders such as cancer and diabetes.31
RUVBL1 (RuvB-like 1) is overexpressed in numerous cancers, where it drives tumor progression and therapy resistance through diverse mechanisms. For instance, in breast cancer, RUVBL1 is ubiquitinated by DTL and cooperates with β-catenin to enhance non-homologous end joining (NHEJ) repair, thereby promoting radiation resistance.32 In tongue squamous cell carcinoma, it activates the CRAF/MEK/ERK pathway to increase cell viability, invasion, and chemotherapy resistance.33 In uveal melanoma, RUVBL1 enhances the transcriptional activity of CTNNB1 (β-catenin) via chromatin remodeling.34 Beyond these specific contexts, RUVBL1 is broadly implicated in oncogenesis through its fundamental roles in chromatin remodeling and DNA repair. Outside oncology, RUVBL1 is essential for ciliary function, its loss in renal tubules leads to a severe cystic kidney phenotype.35
DNAJC7 (DnaJ homolog subfamily C member 7) plays a critical role in proteostasis, with important functions in both neurodegeneration and cancer. Pathogenic variants of DNAJC7 are linked to amyotrophic lateral sclerosis (ALS) through impaired protein quality control.36 In tauopathies, it inhibits tau aggregation, a protective function that is lost in disease, associated mutations.37 Conversely, in HCC, DNAJC7 is overexpressed, where it promotes tumor progression and immune evasion, serves as a prognostic marker, and its knockdown induces cell cycle arrest.38
HSPA14 (Heat shock protein family A member 14) plays dual roles in cancer progression and viral infection. In breast cancer, its overexpression is associated with poor prognosis and may serve as a diagnostic biomarker, potentially through modulating tumorigenic processes and the tumor immune microenvironment.39 In leukemia, it contributes to resistance against endoplasmic reticulum stress‑induced drugs.40 Conversely, during HIV‑1 infection, HSPA14 expression is suppressed, its overexpression inhibits viral replication, likely by interacting with the viral long terminal repeat (LTR) or suppressing proviral transcription.41
HSPD1 (Heat shock protein family D member 1) plays roles in diverse pathological processes, including neurological disorders, cancer, and inflammation. Gene variants of HSPD1 are associated with fatal hypomyelinating leukodystrophy (HLD4) and spastic paraplegia type 13 (SPG13).42 In oncology, HSPD1 overexpression correlates with poor prognosis in non-small cell lung cancer and oral cancer, where it promotes tumor progression by reprogramming mitochondrial metabolism and activating pathways such as NF-κB.43 Inhibitors of HSPD1 can synergize with chemotherapy agents like oxaliplatin.44 Additionally, HSPD1 is implicated in arterial and inflammatory diseases, attributable to its established functions in maintaining protein homeostasis and modulating immune responses.
Relationships Between Cluster 2 Proteins and Diseases
Cluster 2 included 7 nodes (PSMB6, ELOB, UBE2D1, UBXN7, RACK1, HSP90AB1, PSME3) and 14 interaction pairings with 4.667 score (Figure 3A). In addition, enlarged-view, ID, name and Z-score for Cluster 2 proteins were also presented (Figure 3B). Furtherly, KEGG analysis showed that ICA-binding proteins were involved in proteasome, antigen processing and presentation, ubiquitin mediated proteolysis, protein processing in endoplasmic reticulum, pathways in cancer (Figure 3C). Next, GO analysis indicated that in terms of the biological process, ICA-binding proteins were mainly enriched in proteasome-mediated ubiquitin-dependent protein catabolic process. In terms of cellular components, ICA-binding proteins were mainly enriched in endopeptidase complex, peptidase complex, proteasome complex and ubiquitin ligase complex. According to molecular function, ICA-binding proteins were mostly enriched in ubiquitin-like protein ligase binding and ubiquitin protein ligase binding (Figure 3D).
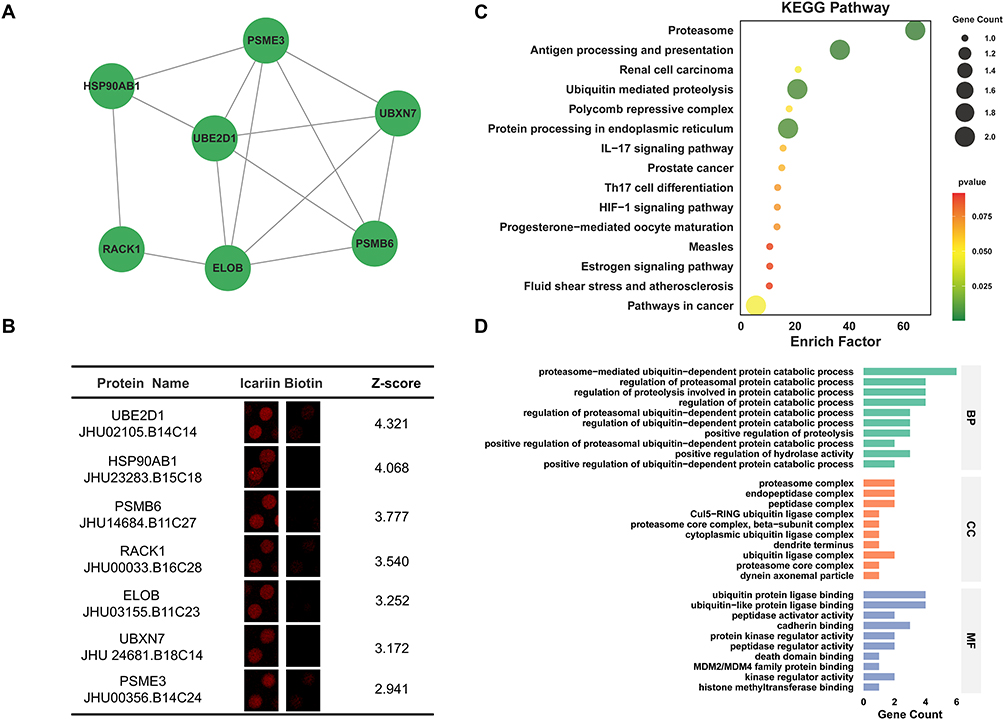 |
Figure 3 Bioinformatics analysis of Cluster 2 Proteins. (A) PPI network of Cluster 2 Proteins. (B) Enlarged-view and Z-score of Cluster 2 Proteins in proteome microarray. (C) KEGG analysis of Cluster 2 Proteins. (D) GO analysis of Cluster 2 Proteins. |
PSMB6 (Proteasome subunit beta type-6) is a catalytic subunit of the proteasome complex, widely expressed in the cytoplasm and nucleus.45 It is implicated in multiple diseases. In cancer, especially lung adenocarcinoma, its overexpression promotes tumor proliferation, invasion, and metastasis, correlating with poor prognosis.46 PSMB6 deficiency impairs proteasome activity and induces apoptosis, as observed in neurodegenerative and developmental zebrafish models.47 Furthermore, as an immunoproteasome component, PSMB6 participates in antigen presentation, and its mutations are linked to autoinflammatory disorders.48
ELOB (Elongin-B) is a core subunit of the elongin complex, involved in transcriptional regulation and ubiquitination.49 It is critically implicated in cancer progression, primarily as a component of the Cullin2-RBX1-ELOB E3 ubiquitin ligase complex that targets the tumor suppressor p14/ARF for degradation. High ELOB expression correlates with poor prognosis in breast cancer. Furthermore, ELOB/C are identified as “essential” genes for proliferation in several cancers (eg, prostate, pancreatic, liver), highlighting their therapeutic potential. Through its role in protein homeostasis, ELOB may also have indirect implications in neurodegenerative (eg, ALS, AD) and autoimmune disorders (eg, SLE, IBD).50,51
UBE2D1 (Ubiquitin conjugating enzyme E2 D1) is a widely expressed ubiquitin-conjugating enzyme involved in cancer and neurodegeneration. In breast cancer, its m6A-driven upregulation promotes proliferation and migration by modulating the TGF-β/Smad2/3 pathway.52 In hepatocellular carcinoma, UBE2D1 contributes to IL-6-induced DNA damage, chemoresistance, and poor prognosis.53 It also facilitates tumorigenesis by ubiquitinating regulators such as p53, KRAS, and TNFα/NF-κB pathway components.54 In neurodegeneration, impaired UBE2D1 function may hinder the clearance of misfolded proteins, promoting toxic aggregation.55
UBXN7 (UBX domain-containing protein 7) is a UBX domain-containing protein involved in ubiquitin-dependent processes. It is implicated in cancer and metabolic disease, showing moderate to strong nuclear expression in tumors. In clear cell renal carcinoma, its levels correlate with prognosis, and it acts as an adaptor for the CRL2/VHL complex to regulate HIF-1α.56 In hepatocellular carcinoma, the hepatitis B virus X protein (HBx) induces UBXN7 degradation, activating NF-κB and autophagy to promote viral replication.57 In diabetic kidney disease, the circular RNA circUBXN7 is upregulated, binds to IGF2BP2 to stabilize SP1 mRNA, and drives a pathogenic feedback loop promoting renal fibrosis.58
RACK1 (Receptor of activated C kinase 1) is a scaffolding protein that integrates cellular signaling pathways. Its aberrant overexpression in cancers, such as lung and liver cancer, correlates with poor prognosis and metastasis. In breast cancer, it promotes progression by stabilizing β-catenin to activate Wnt signaling and enhancing chemoresistance.59 In the nervous system, it has a neuroprotective role by promoting non-amyloidogenic APP processing, but its levels are reduced in AD.60 RACK1 also regulates proteostasis and stress granules, implicating it in other neurodegenerative diseases like frontotemporal dementia and ALS.61
HSP90AB1 (Heat shock protein HSP 90-beta) is a molecular chaperone that stabilizes and regulates diverse client proteins, including kinases and transcription factors. It contributes to disease pathogenesis through this stabilizing function. In cancer, it supports oncoprotein activity, and its methylation enhances tumor growth, making it a therapeutic target.62 In neurodegenerative diseases like Alzheimer’s and Huntington’s, it aids the accumulation of pathological proteins (eg, tau, mutant Huntingtin), and its inhibition promotes clearance.63 In inflammatory conditions, it amplifies responses by stabilizing pro-inflammatory signaling molecules.64
PSME3 (Proteasome activator complex subunit 3) is a proteasome activator that enhances protein degradation. It promotes cancer progression by increasing proteasome activity, which facilitates the degradation of tumor suppressor proteins and supports tumor cell proliferation. This activity may also disrupt antigen presentation, potentially aiding tumor immune evasion.65
Relationships Between Cluster 3 Proteins and Diseases
Cluster 3 included 7 nodes (PPP2R1B, TH, DIABLO, NCF1, MCL1, AKT1, MAPK1) and 11 interaction pairings with 3.667 score (Figure 4A). In addition, the enlarged-view, enlarged-view, ID, name and Z-score for Cluster 3 proteins were also presented (Figure 4B). Furtherly, KEGG analysis showed that ICA-binding proteins were involved in apoptosis and PI3K-Akt signaling pathway (Figure 4C). Next, GO analysis indicated that in terms of the biological process, ICA-binding proteins were mainly enriched in cellular response to inorganic substance, cellular response to metal ion, extrinsic apoptotic signaling pathway, response to metal ion. In terms of cellular components, ICA-binding proteins were mainly enriched in cytoplasmic side of membrane and cytoplasmic side of plasma membrane. According to molecular function, ICA-binding proteins were mostly enriched in phosphatidylinositol binding and phosphatidylinositol-3,4-bisphosphate binding (Figure 4D).
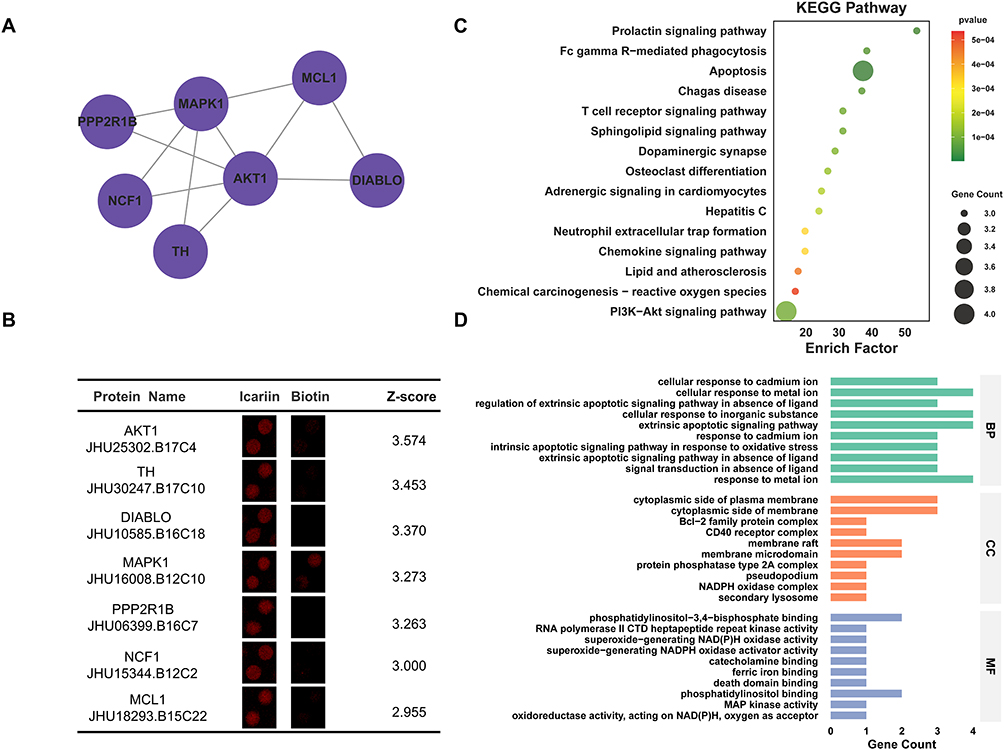 |
Figure 4 Bioinformatics analysis of Cluster 3 Proteins. (A) PPI network of Cluster 3 Proteins. (B) Enlarged-view and Z-score of Cluster 3 Proteins in proteome microarray. (C) KEGG analysis of Cluster 3 Proteins. (D) GO analysis of Cluster 3 Proteins. |
PPP2R1B encodes a regulatory subunit of protein phosphatase 2A (PP2A), which directs PP2A’s substrate specificity and modulates key cellular processes including the cell cycle and apoptosis.66 It functions as a tumor suppressor in several cancers. In colorectal cancer (CRC), low expression of PPP2R1B is associated with poor prognosis, metastasis, and reduced chemosensitivity, potentially through its inhibitory effect on the MAPK/ERK pathway.66 Beyond oncology, mutations in PPP2R1B are strongly linked to male infertility, specifically causing meiotic arrest and non-obstructive azoospermia, which underscores its essential role in spermatogenesis.67
TH (Tyrosine 3-hydroxylase) is the rate-limiting enzyme in catecholamine synthesis, and is essential for neurotransmission and stress responses.68 Its dysfunction is implicated in neurological disorders. In PD, impaired TH activity contributes to dopamine depletion and motor symptoms such as tremor and rigidity.69 Additionally, TH mutations are associated with dopa-responsive dystonia, a movement disorder characterized by muscle rigidity and impaired motor function.
DIABLO (Diablo IAP-binding mitochondrial protein), also known as SMAC, is a pro‑apoptotic mitochondrial protein that promotes caspase-mediated apoptosis by antagonizing inhibitor of apoptosis proteins (IAPs). In cancers such as renal, lung, and hepatocellular carcinoma, DIABLO is frequently downregulated. Its reduced expression correlates with advanced tumor stage, metastasis, and poor prognosis, primarily due to impaired apoptosis and enhanced chemoresistance, underscoring its potential as a therapeutic target.70,71
NCF1 (Neutrophil cytosol factor 1) is a core component of the NADPH oxidase 2 complex essential for generating antimicrobial reactive oxygen species (ROS) in phagocytes. Mutations in NCF1 are a primary cause of chronic granulomatous disease characterized by impaired ROS production and recurrent infections.72 Beyond immunodeficiency, dysregulation of NCF1 is implicated in chronic inflammatory and autoimmune disorders such as rheumatoid arthritis, where aberrant ROS activity contributes to tissue damage.73
MCL1 (Induced myeloid leukemia cell differentiation protein Mcl-1) is an anti‑apoptotic member of the BCL‑2 protein family that promotes cell survival by directly binding and inhibiting the pro‑apoptotic proteins BAK and BAX, thereby preventing mitochondrial outer membrane permeabilization and subsequent caspase activation. In cancer, MCL1 is frequently overexpressed in leukemia, lymphoma, and various solid tumors, where it enhances tumor cell survival and confers resistance to chemotherapy. Therefore, pharmacologically inhibiting MCL1 represents a promising therapeutic strategy to overcome treatment resistance in both hematological malignancies and solid tumors.74,75
AKT1 (RAC-alpha serine/threonine-protein kinase) is a central kinase in the PI3K/AKT pathway that, upon activation, phosphorylates downstream targets such as mTOR and FOXO to regulate cell growth, survival, and metabolism. Its dysregulation is a key driver in multiple cancers, including breast, colorectal, and lung cancer. In addition, it promotes tumor progression, inhibits apoptosis, and contributes to therapy resistance.76,77 Additionally, in AD, dysregulated AKT1 activity is implicated in the abnormal phosphorylation of tau protein, a process central to neurofibrillary tangle formation and neuronal dysfunction.78
MAPK1 (Mitogen-activated protein kinase 1) is a MAP kinase that transduces extracellular signals by phosphorylating transcription factors and cytoskeletal proteins, thereby regulating gene expression, cell cycle progression, and apoptosis. Its constitutive activation promotes cancer cell proliferation, survival, and tumor progression,79 while in neurological disorders it contributes to neuronal damage in stroke, synaptic dysfunction in AD, pathogenesis in PD, and dysregulation in spinocerebellar ataxias.80 Additionally, gain-of-function mutations in MAPK1 are associated with a neurodevelopmental RASopathy exhibiting Noonan syndrome-like features.81
Relationships Between Cluster 4 Proteins and Diseases
Cluster 4 included 3 nodes (NUDT7, NUDT14, NUDT5) and 3 interaction pairings with 3 score (Figure 5A). In addition, the enlarged-view, ID, name and Z-score for Cluster 4 proteins were also presented (Figure 5B). Furtherly, KEGG analysis showed that ICA-binding proteins were involved in peroxisome and purine metabolism (Figure 5C). Next, GO analysis indicated that in terms of the biological process, ICA-binding proteins were mainly enriched in ribose phosphate metabolic process. In terms of cellular components, ICA-binding proteins were mainly enriched in microbody, microbody lumen, peroxisomal matrix and peroxisome. According to molecular function, ICA-binding proteins were mostly enriched in magnesium ion binding and snoRNA binding (Figure 5D).
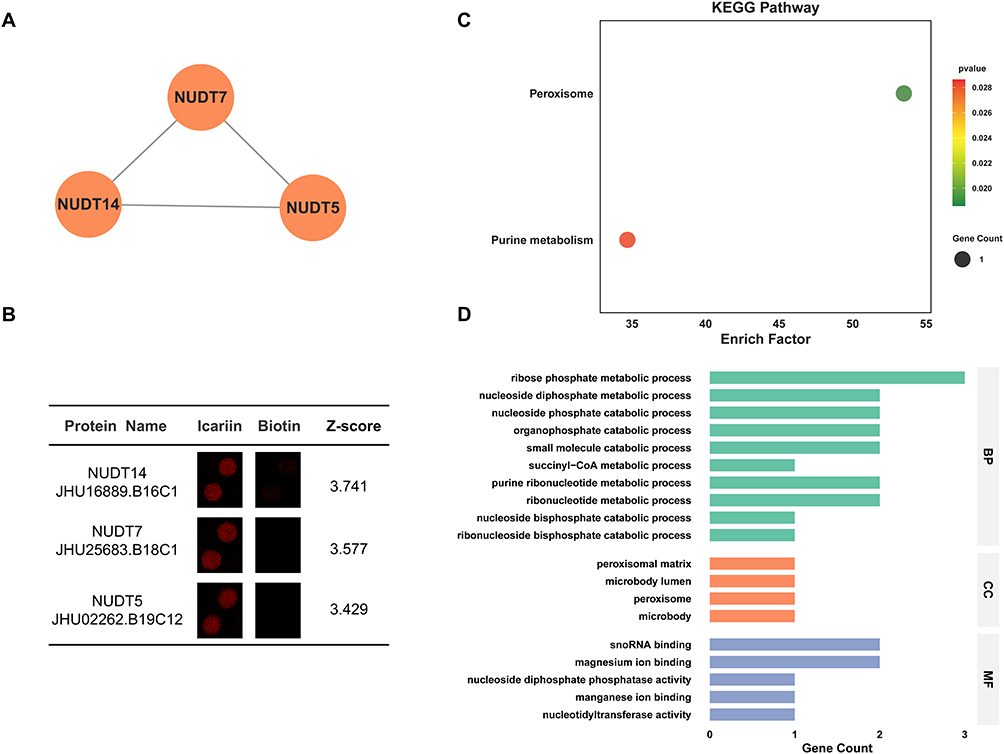 |
Figure 5 Bioinformatics analysis of Cluster 4 Proteins. (A) PPI network of Cluster 4 Proteins. (B) Enlarged-view and Z-score of Cluster 4 Proteins in proteome microarray. (C) KEGG analysis of Cluster 4 Proteins. (D) GO analysis of Cluster 4 Proteins. |
NUDT7 (Nucleoside diphosphate-linked moiety X motif 7) is a nucleoside diphosphate hydrolase that maintains nucleotide homeostasis. It critically regulates key metabolites including UDP‑glucose and UDP‑N‑acetylglucosamine, which are essential for glycosylation and lipid metabolism, and also modulates cyclic nucleotide and phosphoinositide signaling pathways.82 Dysregulation of NUDT7 has been implicated in cancer, where it appears to act as a tumor suppressor, as reduced expression correlates with enhanced cell proliferation and metastasis.83
NUDT14 (Nucleoside diphosphate-linked moiety X motif 14) is a uridine diphosphate hydrolase that specifically converts UDP to UMP, playing a key role in regulating uridine nucleotide levels for RNA metabolism. By promoting nucleotide recycling, it helps maintain cellular energy balance and metabolic homeostasis. NUDT14 has been linked to neurodevelopmental disorders and viral infection.84
NUDT5 (Nucleoside diphosphate-linked moiety X motif 5) is a multifunctional enzyme that plays dual roles in nucleotide metabolism and RNA processing.85 NUDT5 is implicated in immune-related diseases and viral infections, where its RNA processing activity may regulate viral replication or host immune responses. Its dysregulation is also linked to cancer progression, likely mediated through alterations in gene expression and stress signaling pathways.86 The interplay between its distinct enzymatic and RNA-binding activities remains an active area of research.
CETSA Validation of Top 3 ICA-Binding Proteins
HEK293T cells were treated with ICA (10 μM) or vehicle (DMSO) for 24 h. Cell lysates were heated at the indicated temperatures for 3 min. Soluble fractions were analyzed by Western blotting assay for protein expression of the cluster’s Top 3 ICA-binding proteins including TCP1 (Z-score = 5.822), CCT4 (Z-score = 4.702), UBE2D1 (Z-score = 4.321). As showed in Figure 6, ICA significantly enhanced the thermal stability of TCP1, CCT4, and UBE2D1 proteins, confirming its direct interactions with these three proteins. Quantitative analysis revealed that treatment with ICA (10 μM) increased the melting temperature of TCP1 (65°C), CCT4 (70°C), and UBE2D1 (55, 60 and 70°C), respectively, with statistical significance compared to the Control group (Figure 6).
 |
Figure 6 CETSA validation of Top 3 ICA-binding proteins. HEK293T cells were treated with ICA (10 μM) or vehicle (DMSO) for 24 h. Cell lysates were heated at the indicated temperatures for 3 min. Soluble fractions were analyzed by Western blotting assay for protein expression of TCP1 (A), CCT4 (B) and UBE2D1 (C). Notes: Data were expressed as mean ± SEM from three groups of cells. *P < 0.05 compared with Control group. |
Discussion
In this study, we employed a HuProt™ human proteome microarray to comprehensively identify the binding proteins of ICA, leading to the discovery of 246 candidate binding proteins. Subsequent PPI network analysis organized these proteins into 4 functionally distinct clusters, revealing that ICA interacts with proteins involved in central processes, including protein folding chaperone complexes, the ubiquitin-proteasome system, apoptotic and PI3K-Akt signaling pathways, and nucleotide metabolism. This multi-target profile suggests that the pharmacological effects of ICA are likely mediated through the modulation of interconnected biological networks rather than a single target.
The most significantly enriched cluster (Cluster 1) comprised key components of chaperone complexes. ICA-binding proteins were enriched in chaperone-mediated protein folding and protein folding. Given the well-established role of proteostasis failure in neurodegenerative diseases like AD and PD,87 the interaction of ICA with core chaperones (eg, HSPA6, HSPD1, DNAJC7, STIP1) provided a compelling molecular basis for its documented neuroprotective effects. For instance, the binding to DNAJC7 and STIP1 proteins directly implicated in tau aggregation and amyloid-β toxicity, respectively, suggesting a potential mechanism by which ICA could bolster cellular protein quality control to counteract pathological aggregation.37,88 Equally important, the same chaperones are overexpressed in pancreatic, breast, gastric and hepatocellular carcinomas where they stabilize oncogenic clients (c-Myc, SOX2, β-catenin, AKT1), promoting tumor progression and therapy resistance.89,90 Consequently, the poly-chaperone inhibition of ICA offers a mechanistic rationale for its broad anti-tumor activity.
Furthermore, the identification of proteasome subunits (PSMB6, PSME3) and ubiquitin-system enzymes (ELOB, UBE2D1) in Cluster 2 highlights the potential influence of ICA on protein degradation pathways. The proteasome is crucial for degrading misfolded proteins and regulating cell cycle proteins. Its dysfunction is linked to both cancer and neurodegeneration.87,89 ICA’s interaction with these components may indicate an ability to modulate proteolytic activity, which could impact tumor cell survival (eg, via MCL1 or cyclin regulation) and the clearance of toxic protein aggregates.
Clusters 3 and 4 connect ICA to critical signaling and metabolic hubs. The association with the apoptosis and PI3K-Akt pathway (via AKT1, MAPK1, DIABLO, MCL1) suggested ICA could influence cell survival decisions, relevant in both cancer (inhibiting proliferation) and neuroprotection (preventing neuronal apoptosis).91,92 Additionally, the interaction between ICA and NUDT family enzymes in Cluster 4 is involved in nucleotide and peroxisomal metabolism, indicating a potential role in regulating cellular energy homeostasis and redox balance. These processes are frequently disrupted in metabolic disorders and age-related diseases.
Limitations and Perspectives
The interactions were identified in a cell-free system, functional cellular assays and relevant animal models are needed to confirm whether ICA’s binding to these targets modulates the implicated pathways (eg, proteasome activity, chaperone function, or apoptosis) and to establish causal relationships.
Drug target is the point of direct action where a drug exerts its pharmacological effects. By clearly describing the structural characteristics and physiological functions of a target, it helps to clarify the direction of development and accelerate the discovery of new drugs. At the same time, the drug target is a key link in the disease mechanism. By describing the target in detail, we can deeply understand the molecular mechanism of disease development and provide the basis for precise treatment.
Conclusion
This study constructed an atlas of the ICA-binding proteins via microarray, which spans protein folding, ubiquitin-proteasome, apoptosis and nucleotide metabolism, providing a theoretical basis for ICA treatment of diseases caused by dysregulation of these core pathways.
Highlights
- Comprehensive profiling of 246 direct Icariin binding protein using a proteome microarray.
- Functional clustering of Icariin-binding proteins reveals key regulatory networks and pathways.
- The binding atlas provides a foundational resource for elucidating Icariin’s mechanisms and therapeutic potential.
Data Sharing Statement
The datasets generated and analyzed in this study are available in the Protein Microarray Database repository http://www.proteinmicroarray.cn/index.php/array/detail?chip_id=200.
Funding
This study was supported by the National Natural Science Foundation of China (Nos. 32460187, and 82404641).
Disclosure
The authors declare that they have no conflict of interests.
References
1. Wang S, Ma J, Zeng Y, et al. Icariin, an up-and-coming bioactive compound against neurological diseases: network pharmacology-based study and literature review. Drug Des Devel Ther. 2021;15:3619–19. doi:10.2147/dddt.S310686
2. Jin J, Wang H, Hua X, et al. An outline for the pharmacological effect of icariin in the nervous system. Eur J Pharmacol. 2019;842:20–32. doi:10.1016/j.ejphar.2018.10.006
3. Wang M, Gao H, Li W, et al. Icariin and its metabolites regulate lipid metabolism: from effects to molecular mechanisms. Biomed Pharmacother. 2020;131:110675. doi:10.1016/j.biopha.2020.110675
4. Zhu X, Wen S, Gul H, et al. Exploring regulatory network of icariin synthesis in Herba Epimedii through integrated omics analysis. Front Plant Sci. 2024;15:1409601. doi:10.3389/fpls.2024.1409601
5. Zhang HN, Yang L, Ling JY, et al. Systematic identification of arsenic-binding proteins reveals that hexokinase-2 is inhibited by arsenic. Proc Natl Acad Sci U S A. 2015;112(49):15084–15089. doi:10.1073/pnas.1521316112
6. Wang G, Li X, Li N, et al. Icariin alleviates uveitis by targeting peroxiredoxin 3 to modulate retinal microglia M1/M2 phenotypic polarization. Redox Biol. 2022;52:102297. doi:10.1016/j.redox.2022.102297
7. Jeong JS, Jiang L, Albino E, et al. Rapid identification of monospecific monoclonal antibodies using a human proteome microarray. Mol Cell Proteomics. 2012;11(6):
8. Luo C, Yu Y, Zhu J, et al. Deubiquitinase PSMD7 facilitates pancreatic cancer progression through activating Nocth1 pathway via modifying SOX2 degradation. Cell Biosci. 2024;14(1):35. doi:10.1186/s13578-024-01213-9
9. Zhao Y, Yang X, Xu X, et al. Deubiquitinase PSMD7 regulates cell fate and is associated with disease progression in breast cancer. Am J Transl Res. 2020;12(9):5433–5448.
10. Wang J, Liu R, Mo H, et al. Deubiquitinase PSMD7 promotes the proliferation, invasion, and cisplatin resistance of gastric cancer cells by stabilizing RAD23B. Int J Biol Sci. 2021;17(13):3331–3342. doi:10.7150/ijbs.61128
11. Niu Z, Lei R, Shi J, et al. A polymorphism rs17336700 in the PSMD7 gene is associated with ankylosing spondylitis in Chinese subjects. Ann Rheum Dis. 2011;70(4):706–707. doi:10.1136/ard.2010.130039
12. Tang N, Cai X, Peng L, et al. TCP1 regulates Wnt7b/β-catenin pathway through P53 to influence the proliferation and migration of hepatocellular carcinoma cells. Signal Transduct Target Ther. 2020;5(1):169. doi:10.1038/s41392-020-00278-5
13. Liu H, Chen L, Chen Y, et al. TCP1 promotes the progression of malignant tumours by stabilizing c-Myc through the AKT/GSK-3β and ERK signalling pathways. Commun Biol. 2025;8(1):563. doi:10.1038/s42003-025-07867-6
14. Zhang X, Shen L, Zhu Y, et al. Crosstalk of RNA methylation writers defines tumor microenvironment and alisertib resistance in breast cancer. Front Endocrinol. 2023;14:1166939. doi:10.3389/fendo.2023.1166939
15. Zhang S, Wang J, Huang G, et al. TCP1 expression alters the ferroptosis sensitivity of diffuse large B-cell lymphoma subtypes by stabilising ACSL4 and influences patient prognosis. Cell Death Dis. 2024;15(8):611. doi:10.1038/s41419-024-07001-0
16. Chao A, Lai CH, Tsai CL, et al. Tumor stress-induced phosphoprotein1 (STIP1) as a prognostic biomarker in ovarian cancer. PLoS One. 2013;8(2):e57084. doi:10.1371/journal.pone.0057084
17. Guo X, Yan Z, Zhang G, et al. STIP1 regulates proliferation and migration of lung adenocarcinoma through JAK2/STAT3 signaling pathway. Cancer Manag Res. 2019;11:10061–10072. doi:10.2147/cmar.S233758
18. Tan JSY, Lee B, Lim J, et al. Parkinson’s disease-specific autoantibodies against the neuroprotective co-chaperone STIP1. Cells. 2022;11(10). doi:10.3390/cells11101649
19. Sun Q, Li N, Jia L, et al. Ribosomal protein SA-positive neutrophil elicits stronger phagocytosis and neutrophil extracellular trap formation and subdues pro-inflammatory cytokine secretion against streptococcus suis serotype 2 infection. Front Immunol. 2020;11:585399. doi:10.3389/fimmu.2020.585399
20. Liu M, Li N, Guo W, et al. RPSA distribution and expression in tissues and immune cells of pathogen-infected mice. Microb Pathog. 2021;152:104609. doi:10.1016/j.micpath.2020.104609
21. Limone A, Maggisano V, Sarnataro D, et al. Emerging roles of the cellular prion protein (PrP(C)) and 37/67 kDa laminin receptor (RPSA) interaction in cancer biology. Cell Mol Life Sci. 2023;80(8):207. doi:10.1007/s00018-023-04844-2
22. Zhao Y, Guo G, Sun Y, et al. Membrane protein CRISPR screen identifies RPSA as an essential host factor for porcine epidemic diarrhea virus replication. J Virol. 2025;99(8):e0064925. doi:10.1128/jvi.00649-25
23. Liao Q, Ren Y, Yang Y, et al. CCT8 recovers WTp53-suppressed cell cycle evolution and EMT to promote colorectal cancer progression. Oncogenesis. 2021;10(12):84. doi:10.1038/s41389-021-00374-3
24. Fichtenbauer D, Xu XM, Jackson D, et al. The chaperonin CCT8 facilitates spread of tobamovirus infection. Plant Signal Behav. 2012;7(3):318–321. doi:10.4161/psb.19152
25. Oftedal BE, Maio S, Handel AE, et al. The chaperonin CCT8 controls proteostasis essential for T cell maturation, selection, and function. Commun Biol. 2021;4(1):681. doi:10.1038/s42003-021-02203-0
26. Jia J, Liu G, Zhong J, et al. Heat shock protein A6 is especially involved in enterovirus 71 infection. Front Microbiol. 2022;13:865644. doi:10.3389/fmicb.2022.865644
27. Zhou X, Ji Q, Li Q, et al. HSPA6 is correlated with the malignant progression and immune microenvironment of gliomas. Front Cell Dev Biol. 2022;10:833938. doi:10.3389/fcell.2022.833938
28. Li F, Liu CS, Wu P, et al. CCT4 suppression inhibits tumor growth in hepatocellular carcinoma by interacting with Cdc20. Chin Med J. 2021;134(22):2721–2729. doi:10.1097/cm9.0000000000001851
29. Sergeeva OA, Tran MT, Haase-Pettingell C, et al. Biochemical characterization of mutants in chaperonin proteins CCT4 and CCT5 associated with hereditary sensory neuropathy. J Biol Chem. 2014;289(40):27470–27480. doi:10.1074/jbc.M114.576033
30. Macario AJ, Grippo TM, Conway de Macario E. Genetic disorders involving molecular-chaperone genes: a perspective. Genet Med. 2005;7(1):3–12. doi:10.1097/01.gim.0000151351.11876.c3
31. Kim AR, Choi KW. TRiC/CCT chaperonins are essential for organ growth by interacting with insulin/TOR signaling in Drosophila. Oncogene. 2019;38(24):4739–4754. doi:10.1038/s41388-019-0754-1
32. Tian J, Wen M, Gao P, et al. RUVBL1 ubiquitination by DTL promotes RUVBL1/2-β-catenin-mediated transcriptional regulation of NHEJ pathway and enhances radiation resistance in breast cancer. Cell Death Dis. 2024;15(4):259. doi:10.1038/s41419-024-06651-4
33. Zhang XY, Liu Y, Rong Q, et al. RUVBL1 accelerates tongue squamous cell carcinoma by mediating CRaf/MEK/ERK pathway. iScience. 2024;27(4):109434. doi:10.1016/j.isci.2024.109434
34. Zhang C, Wu S. RUVBL1-modulated chromatin remodeling alters the transcriptional activity of oncogenic CTNNB1 in uveal melanoma. Cell Death Discov. 2023;9(1):132. doi:10.1038/s41420-023-01429-7
35. Kim CY, Lee HK, Lee H, et al. Ruvbl1 is essential for ciliary beating during xenopus laevis embryogenesis. Dev Reprod. 2023;27(3):159–165. doi:10.12717/dr.2023.27.3.159
36. Dilliott AA, Andary CM, Stoltz M, et al. DnaJC7 in amyotrophic lateral sclerosis. Int J Mol Sci. 2022;23(8). doi:10.3390/ijms23084076
37. Hou Z, Wydorski PM, Perez VA, et al. DnaJC7 binds natively folded structural elements in tau to inhibit amyloid formation. Nat Commun. 2021;12(1):5338. doi:10.1038/s41467-021-25635-y
38. Chen J, Yang Z, Cui Y, et al. Increased expression of DNAJC7 promotes the progression of hepatocellular carcinoma by influencing the cell cycle and immune microenvironment. J Cancer Res Clin Oncol. 2025;151(5):154. doi:10.1007/s00432-025-06202-0
39. Zhao R, Zhu X, Wei W, et al. The role of HSPA14 in breast cancer: implications for tumorigenesis, immune response modulation, and personalized therapies. Int J Hyperthermia. 2025;42(1):2452922. doi:10.1080/02656736.2025.2452922
40. Cabaud-Gibouin V, Durand M, Quéré R, et al. Heat-shock proteins in leukemia and lymphoma: multitargets for innovative therapeutic approaches. Cancers. 2023;15(3). doi:10.3390/cancers15030984
41. Bi M, Kang W, Sun Y. Expression of HSPA14 in patients with acute HIV-1 infection and its effect on HIV-1 replication. Front Immunol. 2023;14:1123600. doi:10.3389/fimmu.2023.1123600
42. Cömert C, Brick L, Ang D, et al. A recurrent de novo HSPD1 variant is associated with hypomyelinating leukodystrophy. Cold Spring Harb Mol Case Stud. 2020;6(3). doi:10.1101/mcs.a004879
43. Aluksanasuwan S, Somsuan K, Ngoenkam J, et al. Potential association of HSPD1 with dysregulations in ribosome biogenesis and immune cell infiltration in lung adenocarcinoma: an integrated bioinformatic approach. Cancer Biomark. 2024;39(3):155–170. doi:10.3233/cbm-220442
44. Wu T, Yu Y, Tu X, et al. Tubeimoside-I, an inhibitor of HSPD1, enhances cytotoxicity of oxaliplatin by activating ER stress and MAPK signaling pathways in colorectal cancer. J Ethnopharmacol. 2025;336:118754. doi:10.1016/j.jep.2024.118754
45. Huang WR, Chi PI, Chiu HC, et al. Avian reovirus p17 and σA act cooperatively to downregulate Akt by suppressing mTORC2 and CDK2/cyclin A2 and upregulating proteasome PSMB6. Sci Rep. 2017;7(1):5226. doi:10.1038/s41598-017-05510-x
46. Bian Y, Sui Q, Bi G, et al. Identification and validation of a proliferation-associated score model predicting survival in lung adenocarcinomas. Dis Markers. 2021;2021:3219594. doi:10.1155/2021/3219594
47. Wang J, Xu L, Yun X, et al. Proteomic analysis reveals that proteasome subunit beta 6 is involved in hypoxia-induced pulmonary vascular remodeling in rats. PLoS One. 2013;8(7):e67942. doi:10.1371/journal.pone.0067942
48. Yang C, Zheng X, Chen J, et al. Protein biomarker screening and functional analysis of salivary exosomes in patients with ulcerative colitis. Beijing Da Xue Xue Bao Yi Xue Ban. 2025;57(5):895–902. doi:10.19723/j.issn.1671-167X.2025.05.013
49. Lv J, Wu T, Xue J, et al. ASB1 engages with ELOB to facilitate SQOR ubiquitination and H(2)S homeostasis during spermiogenesis. Redox Biol. 2025;79:103484. doi:10.1016/j.redox.2024.103484
50. Sui XY, Ma XY, Hou Y, et al. Elongin B promotes breast cancer progression by ubiquitinating tumor suppressor p14/ARF. Cell Biol Toxicol. 2024;40(1):24. doi:10.1007/s10565-024-09864-7
51. Fischer S, Trinh VT, Simon C, et al. Peptide-mediated inhibition of the transcriptional regulator Elongin BC induces apoptosis in cancer cells. Cell Chem Biol. 2023;30(7):766–79.e11. doi:10.1016/j.chembiol.2023.05.012
52. Guan XQ, Yuan XN, Feng KX, et al. IGF2BP2-modified UBE2D1 interacts with Smad2/3 to promote the progression of breast cancer. Am J Cancer Res. 2023;13(7):2948–2968.
53. Zhou C, Bi F, Yuan J, et al. Gain of UBE2D1 facilitates hepatocellular carcinoma progression and is associated with DNA damage caused by continuous IL-6. J Exp Clin Cancer Res. 2018;37(1):290. doi:10.1186/s13046-018-0951-8
54. Mu X, Wei Y, Fan X, et al. Aberrant activation of a miR-101-UBE2D1 axis contributes to the advanced progression and chemotherapy sensitivity in human hepatocellular carcinoma. Cell Death Discov. 2024;10(1):422. doi:10.1038/s41420-024-02193-y
55. Andrews JL, Goodfellow FJ, Matosin N, et al. Alterations of ubiquitin related proteins in the pathology and development of schizophrenia: Evidence from human and animal studies. J Psychiatr Res. 2017;90:31–39. doi:10.1016/j.jpsychires.2017.01.009
56. Yang E, Fan X, Ye H, et al. Exploring the role of ubiquitin regulatory X domain family proteins in cancers: bioinformatics insights, mechanisms, and implications for therapy. J Transl Med. 2024;22(1):157. doi:10.1186/s12967-024-04890-9
57. Yuan S, Xu J, Wang M, et al. HBV X protein induces degradation of UBXN7, a novel negative regulator of NF-κB signaling, to promote HBV replication. Cell Mol Gastroenterol Hepatol. 2023;15(1):179–195. doi:10.1016/j.jcmgh.2022.09.003
58. Lin Z, Lv D, Liao X, et al. CircUBXN7 promotes macrophage infiltration and renal fibrosis associated with the IGF2BP2-dependent SP1 mRNA stability in diabetic kidney disease. Front Immunol. 2023;14:1226962. doi:10.3389/fimmu.2023.1226962
59. Jia Y, Zhang L, Zhou W, et al. Rack1 promotes breast cancer stemness properties and tumorigenesis through the E2F1-SOX2 axis. Cancer Cell Int. 2025;25(1):40. doi:10.1186/s12935-025-03678-1
60. He W, Shi X, Dong Z. The roles of RACK1 in the pathogenesis of Alzheimer’s disease. J Biomed Res. 2024;38(2):137–148. doi:10.7555/jbr.37.20220259
61. Masi M, Attanzio A, Racchi M, et al. Proteostasis deregulation in neurodegeneration and its link with stress granules: focus on the scaffold and ribosomal protein RACK1. Cells. 2022;11(16). doi:10.3390/cells11162590
62. Haase M, Fitze G. HSP90AB1: helping the good and the bad. Gene. 2016;575(2 Pt 1):171–186. doi:10.1016/j.gene.2015.08.063
63. Bao J, Chang W, Zhao Y. Diagnosis and drug prediction of parkinson’s disease based on immune-related genes. J Mol Neurosci. 2022;72(9):1809–1819. doi:10.1007/s12031-022-02043-5
64. Jin L, Yang G, Liu Y, et al. Bioinformatics data combined with single-cell analysis reveals patterns of immunoinflammatory infiltration and cell death in melanoma. Int Immunopharmacol. 2024;143(Pt 2):113347. doi:10.1016/j.intimp.2024.113347
65. Yi Z, Yang D, Liao X, et al. PSME3 induces epithelial-mesenchymal transition with inducing the expression of CSC markers and immunosuppression in breast cancer. Exp Cell Res. 2017;358(2):87–93. doi:10.1016/j.yexcr.2017.05.017
66. Liu W, Tang J, Gao W, et al. PPP2R1B abolishes colorectal cancer liver metastasis and sensitizes Oxaliplatin by inhibiting MAPK/ERK signaling pathway. Cancer Cell Int. 2024;24(1):90. doi:10.1186/s12935-024-03273-w
67. Du M, Yuan L, Zhang Z, et al. PPP2R1B is modulated by ubiquitination and is essential for spermatogenesis. FASEB J. 2021;35(5):e21564. doi:10.1096/fj.202002810R
68. Wang Y, Wang C, Liu M, et al. Segawa syndrome caused by TH gene mutation and its mechanism. Front Genet. 2022;13:1004307. doi:10.3389/fgene.2022.1004307
69. Gopinath A, Mackie P, Hashimi B, et al. DAT and TH expression marks human Parkinson’s disease in peripheral immune cells. NPJ Parkinsons Dis. 2022;8(1):72. doi:10.1038/s41531-022-00333-8
70. Paul A, Krelin Y, Arif T, et al. A new role for the mitochondrial pro-apoptotic protein SMAC/Diablo in phospholipid synthesis associated with tumorigenesis. Mol Ther. 2018;26(3):680–694. doi:10.1016/j.ymthe.2017.12.020
71. Espinosa M, Lizárraga F, Vázquez-Santillán K, et al. Coexpression of Smac/DIABLO and estrogen receptor in breast cancer. Cancer Biomark. 2021;30(4):429–446. doi:10.3233/cbm-200535
72. Kong M, Chen X, Lv F, et al. Serum response factor (SRF) promotes ROS generation and hepatic stellate cell activation by epigenetically stimulating NCF1/2 transcription. Redox Biol. 2019;26:101302. doi:10.1016/j.redox.2019.101302
73. Li Y, Li Z, Nandakumar KS, et al. Human NCF1(90H) variant promotes IL-23/IL-17-dependent mannan-induced psoriasis and psoriatic arthritis. Antioxidants. 2023;12(7). doi:10.3390/antiox12071348
74. Sancho M, Leiva D, Lucendo E, et al. Understanding MCL1: from cellular function and regulation to pharmacological inhibition. Febs J. 2022;289(20):6209–6234. doi:10.1111/febs.16136
75. Wu X, Luo Q, Liu Z. Ubiquitination and deubiquitination of MCL1 in cancer: deciphering chemoresistance mechanisms and providing potential therapeutic options. Cell Death Dis. 2020;11(7):556. doi:10.1038/s41419-020-02760-y
76. Alwhaibi A, Verma A, Adil MS, et al. The unconventional role of Akt1 in the advanced cancers and in diabetes-promoted carcinogenesis. Pharmacol Res. 2019;145:104270. doi:10.1016/j.phrs.2019.104270
77. Johnson SM, Gulhati P, Rampy BA, et al. Novel expression patterns of PI3K/Akt/mTOR signaling pathway components in colorectal cancer. J Am Coll Surg. 2010;210(5):767–76, 76–8. doi:10.1016/j.jamcollsurg.2009.12.008
78. Wang Y, Yang R, Gu J, et al. Cross talk between PI3K-AKT-GSK-3β and PP2A pathways determines tau hyperphosphorylation. Neurobiol Aging. 2015;36(1):188–200. doi:10.1016/j.neurobiolaging.2014.07.035
79. Deng R, Zhang HL, Huang JH, et al. MAPK1/3 kinase-dependent ULK1 degradation attenuates mitophagy and promotes breast cancer bone metastasis. Autophagy. 2021;17(10):3011–3029. doi:10.1080/15548627.2020.1850609
80. Sun J, Nan G. The extracellular signal-regulated kinase 1/2 pathway in neurological diseases: a potential therapeutic target (Review). Int J Mol Med. 2017;39(6):1338–1346. doi:10.3892/ijmm.2017.2962
81. Motta M, Pannone L, Pantaleoni F, et al. Enhanced MAPK1 function causes a neurodevelopmental disorder within the rasopathy clinical spectrum. Am J Hum Genet. 2020;107(3):499–513. doi:10.1016/j.ajhg.2020.06.018
82. Song J, Kim EH, Yang JH, et al. Upregulated FOXM1 stimulates chondrocyte senescence in Acot12(-/-)Nudt7(-/-) double knockout mice. Theranostics. 2023;13(15):5207–5222. doi:10.7150/thno.89033
83. Song J, Park S, Oh J, et al. NUDT7 loss promotes Kras(G12D) CRC development. Cancers. 2020;12(3). doi:10.3390/cancers12030576
84. Wang G, Ren G, Cui X, et al. Human cytomegalovirus RL13 protein interacts with host NUDT14 protein affecting viral DNA replication. Mol Med Rep. 2016;13(3):2167–2174. doi:10.3892/mmr.2016.4778
85. Qi H, Grace Wright RH, Beato M, et al. The ADP-ribose hydrolase NUDT5 is important for DNA repair. Cell Rep. 2022;41(12):111866. doi:10.1016/j.celrep.2022.111866
86. Tong XY, Quan Y, Zhang HY. NUDT5 as a novel drug target and prognostic biomarker for ER-positive breast cancer. Drug Discov Today. 2021;26(3):620–625. doi:10.1016/j.drudis.2020.11.031
87. Kinger S, Jagtap YA, Kumar P, et al. Proteostasis in neurodegenerative diseases. Adv Clin Chem. 2024;121:270–333. doi:10.1016/bs.acc.2024.04.002
88. Batko J, Antosz K, Miśków W, et al. Chaperones-A new class of potential therapeutic targets in alzheimer’s disease. Int J Mol Sci. 2024;25(6). doi:10.3390/ijms25063401
89. Paladino L, Vitale AM, Santonocito R, et al. Molecular Chaperones and Thyroid Cancer. Int J Mol Sci. 2021;22(8). doi:10.3390/ijms22084196
90. Heritz JA, Backe SJ, Mollapour M. Molecular chaperones: guardians of tumor suppressor stability and function. Oncotarget. 2024;15:679–696. doi:10.18632/oncotarget.28653
91. Khan MN, Choudhary D, Mehan S, et al. Molecular mechanisms of GDNF/GFRA1/RET and PI3K/AKT/ERK signaling interplay in neuroprotection: therapeutic strategies for treating neurological disorders. Neuropeptides. 2025;111:102516. doi:10.1016/j.npep.2025.102516
92. Hedayati N, Safari MH, Milasi YE, et al. Modulation of the PI3K/Akt signaling pathway by resveratrol in cancer: molecular mechanisms and therapeutic opportunity. Discov Oncol. 2025;16(1):669. doi:10.1007/s12672-025-02471-w
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.