Back to Journals » Drug Design, Development and Therapy » Volume 19
Preliminary Developmental Safety Assessment of Allylestrenol in Pregnant SD Rats: Evaluation of F0 and Early F1 Generational Endpoints
Authors Jiang L, Zhuan S, Jin W ![]() , Wen H, Li D
, Wen H, Li D
Received 20 March 2025
Accepted for publication 13 November 2025
Published 26 December 2025 Volume 2025:19 Pages 11659—11673
DOI https://doi.org/10.2147/DDDT.S529390
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Tin Wui Wong
Lai Jiang,1 Shengnan Zhuan,2 Wenjie Jin,3 He Wen,3 Dajin Li4
1Department of Obstetrics and Gynecology, The First Affiliated Hospital of USTC, Division of Life Sciences and Medicine, University of Science and Technology of China, Hefei, Anhui, 230001, People’s Republic of China; 2Department of Obstetrics and Gynecology, Bengbu Medical University, Bengbu, Anhui, 233030, People’s Republic of China; 3Tianjin Tiancheng New Drug Evaluation Co., Ltd, Tianjin, People’s Republic of China; 4Haboratory of Reproduction Immunology, Shanghai Key Laboratory of Female Reproductive Endocrine Related Diseases, Obstetrics and Gynecology Hospital, Fudan University Shanghai Medical College, Shanghai, 200001, People’s Republic of China
Correspondence: Dajin Li, Email [email protected] He Wen, Email [email protected]
Objective: The objective of this study was to evaluate the developmental and hormonal safety profile of different doses of allylestrenol administered during gestation in SD rats, focusing on selected physiological and reproductive indices in the Parental (F0) and First-generation offspring (F1) generations.
Methods: In this study, 138 successfully mated Sprague-Dawley (SD) female rats were randomly divided into solvent control (Sol-C), progesterone injection control (PgI-C), progesterone soft-gel capsule control (PgSC-C), low-dose allylestrenol (Alt-LD, 5 mg kg− 1), medium-dose allylestrenol (Alt-MD, 10 mg kg− 1), and high-dose allylestrenol (Alt-HD, 20 mg kg− 1) groups, with 16 successfully mated female rats in each group. Satellite groups were also established, each consisting of 7 successfully mated female rats.
Results: Estradiol levels on gestation day 19 (GD19) were reduced in the Alt-HD compared to the Sol-C (P < 0.05). Compared with the Sol-C, there was no significant change in the anogenital distance (AGD) on postnatal day 21 (PND21) and PND54 in both male and female littermates in each allylestrenol dose group (P > 0.05). Testosterone levels were significantly elevated in the Alt-MD and Alt-LD groups compared to the Sol-C (P < 0.05). On GD19, the estradiol level was significantly reduced only in the Alt-HD group (9.981 ± 0.514 ng/mL) compared with the Sol-C group (18.725 ± 4.770 ng/mL; P < 0.05). The low-dose (Alt-LD: 17.575 ± 3.017 ng/mL) and medium-dose (Alt-MD: 11.927 ± 3.663 ng/mL) groups showed numerically lower values than Sol-C, but the difference was not statistically significant after adjustment. 95% CIs for Sol-C, Alt-MD, and Alt-HD were [17.1, 20.3], [10.2, 13.7], and [9.4, 10.5] ng/mL, respectively.
Conclusion: Allylestrenol at doses of 5, 10, and 20 mg kg− 1 did not result in overt developmental abnormalities in maternal or offspring rats across selected early endpoints. However, further long-term and functional evaluations across generations are needed to fully establish its safety profile.
Keywords: allylestrenol, SD rats, normal pregnancy, progesterone, reproductive toxicity
Introduction
Progesterone is a key hormone that plays an essential role in the maintenance of pregnancy.1 In early gestation, it supports embryo implantation and uterine quiescence by reducing tubal contractility, modulating cervical mucus, promoting endometrial decidualization, and suppressing maternal immune responses to the fetus. Later in pregnancy, progesterone prevents preterm labor by maintaining relaxation of the myometrial musculature.2 Clinically, progesterone supplementation is widely used to prevent threatened miscarriage, recurrent pregnancy loss, and premature labor.3 However, oral administration of natural progesterone is often associated with low bioavailability due to first-pass metabolism, while injectable formulations can cause pain, inflammation, or compliance issues.4,5 These limitations have prompted interest in synthetic progestins, which offer improved stability, potency, and patient acceptability.
One such compound is allylestrenol, a synthetic progestin with high oral bioavailability and potent pregnancy-maintaining activity. Allylestrenol enhances placental endocrine function, increases endogenous progesterone and HCG secretion, and supports uterine quiescence by antagonizing prostaglandins and oxytocin.6–9 Clinically, it has been shown to prolong gestation and improve neonatal outcomes without androgenic, estrogenic, or glucocorticoid side effects.10–13 Allylestrenol has been reported to be approximately 5–10 times more potent than progesterone in maintaining uterine quiescence in vivo,14 with docking studies showing binding energies of −8.1 kcal/mol versus −5.7 kcal/mol for progesterone.14
Despite allylestrenol’s extensive clinical use in Asia for decades, concerns persist regarding the safety of progestin use during pregnancy. Some progestins have been associated with developmental toxicity, including urethral hypospadias, due to androgenic effects.15 Although allylestrenol is believed to lack androgenic activity, few studies have directly evaluated its developmental or transgenerational safety in preclinical models.
Therefore, the objective of this study was to assess the effects of allylestrenol exposure during gestation on maternal reproductive performance and offspring development in SD rats. Specifically, we evaluated selected indices in the (F0) and First-generation offspring (F1) generations, including maternal hormone levels, fetal viability, organ weights, and hormonal parameters, to generate foundational safety data for this widely used synthetic progestin.
Materials and Methods
Animals
Nine-week-old SD rats (SPF, n = 280; males, n = 100; females, n = 180) were purchased from Beijing Vital River Laboratory Animal Technology Company (Beijing, China; experimental animal quality certificate number: 110011231108836813). The animals were subjected to reception, quarantine, and adaptive breeding for 10 days in a barrier environment with a temperature of 20°C to 26°C and relative humidity of 40% to 70%. Feed was provided by Beijing Keao Xieli Feed Company (Beijing, China), and animals had free access to food and water.
The study of animals used in this study follows the Guide for the Care and Use of Laboratory Animals (2011, 8th edition). All experiments followed the ARRIVE (Animal Research: Reporting of In Vivo Experiments) guidelines.6 The institute is accredited by the International Association for Assessment and Accreditation of Laboratory Animal Care International (AAALAC). The method of animal use was approved by the Institutional Animal Care and Use Committee (IACUC), approval number: 2023071101.
Animals were housed in polysulfone cages with stainless steel wire lids (cage size: 440×280 × 200 mm) at a maximum density of three rats per cage. The facility maintained a controlled environment with a 12-hour light/12-hour dark cycle, temperature of 20–26°C, and relative humidity of 40–70%. Corn cob bedding was used, and environmental enrichment included nesting materials and PVC tunnels. All animals were housed under Specific Pathogen-Free (SPF) conditions. Routine health monitoring was conducted weekly, and animals showing signs of illness, injury, or abnormal behavior were excluded from the study.11
Randomization was performed using computer-generated random numbers based on body weight distribution to ensure comparable groups.15 Blinding was implemented for all personnel responsible for outcome assessment and hormone analysis; dosing personnel were separated from those involved in data collection.
Preparation of Allylestrenol and Solvent Control Solution
Allylestrenol was administered via oral gavage in a suspension using 0.5% carboxymethyl cellulose sodium (CMC-Na) as the vehicle based on solubility compatibility data with allylestrenol, and the final formulation pH was adjusted to 6.8–7.2.5 The test substance was a white crystalline solid formulation (Changzhou Siyao Pharmaceutical Company Co., Ltd, China; Batch No. A202303011).
Analytical testing confirmed the homogeneity and target concentration (±10%) of the allylestrenol suspensions prior to and during the dosing period using validated HPLC methods. Stability studies showed that the allylestrenol suspension remained stable for up to 4 hours at room temperature (25 ± 2°C). Therefore, freshly prepared suspensions were used within 4 hours of preparation to ensure dose accuracy.
The selected doses of 5, 10, and 20 mg kg−1 were based on reported clinical dosing equivalence, preliminary tolerability data in rodents, and existing literature indicating effectiveness and safety at these levels. The high dose (20 mg kg−1) was chosen to represent a limit test level in repeated-dose studies.
Preparation of Allylestrenol Solution
Allylestrenol (white crystalline solids, Changzhou Siyao Pharmaceutical Company Co., Ltd, Jiangsu, China; batch No. A202303011) was weighed a certain amount and transferred to a mortar. A small amount of solvent was added, ground, and stirred until there were no large particles, and then transferred to a measuring cylinder. The mortar was rinsed with solvent in small amounts several times until there was no residue of the drug, and the rinse solution was poured into a measuring cylinder. Finally, the solvent was added to finalize the volume, sonicated, and mixed for use. The test solution should be used immediately after preparation, and other dose groups were prepared in the same way, with concentrations of 5, 10, and 20 mg/mL for the low, medium, and high dose groups, respectively.
Experimental Methods
Mated Examination and Grouping of Laboratory Animals
A total of 100 males and 180 females were used for mating. Each male rat was paired with a single female during the 6-day cohabitation period; no males were reused across females. Vaginal plug detection was used to confirm successful mating, and females were randomly assigned to main or satellite groups post-confirmation. Successfully mated female rats that met the experimental conditions were randomly grouped into solvent control (Sol-C), progesterone injection control (PgI-C), progesterone soft-gel capsule control (PgSC-C), low-dose allylestrenol (Alt-LD, 5 mg kg−1), medium-dose allylestrenol (Alt-MD, 10 mg kg−1), and high-dose allylestrenol (Alt-HD, 20 mg kg−1) groups, with 16 successfully mated female rats in each group. Satellite groups for hormone detection were also set up in the same way with 7 successfully mated female rats in each group, including Sol-C (satellite), PgI-C (satellite), PgSC-C (satellite), Alt-LD (satellite), Alt-MD (satellite), and Alt-HD (satellite) groups. The satellite groups were established specifically for serial hormone analysis (testosterone, progesterone, and estradiol) at three timepoints (GD6, GD19, and LD21) to avoid repeated blood sampling from the main cohort and minimize stress-related hormonal alterations.
Female rats were randomly assigned to treatment groups using computer-generated random numbers. Investigators performing endpoint assessments and hormonal analyses were blinded to group allocations to minimize observer bias.
Drug Administration to Parental Generation Rats
Successfully mated female rats were administered the investigational and comparator drugs once daily from gestation day (GD) 6 to GD19, totaling 14 doses. Allylestrenol was given by oral gavage at dose levels of 5, 10, and 20 mg kg−1 (Sol-C, Alt-LD, Alt-MD, and Alt-HD groups) with an administration volume of 1 mL/kg. These doses were based on prior range-finding studies and represent approximately 1×, 2×, and 4× multiples of anticipated human clinical exposure, adjusted for interspecies scaling.
Comparator progesterone formulations were selected and administered to reflect their intended clinical use. Progesterone injection (PgI-C group; 1 mL: 20 mg, approval no. H44020229, Guangzhou Baiyunshan Mingxing Pharmaceutical Co., Ltd, China) was administered intramuscularly at 1.35 mL/kg to preserve bioavailability comparable to clinical practice. Progesterone soft-gel capsules (PgSC-C group; 100 mg/capsule, approval no. HJ20160265, Cyndea Pharma, S.L.) were administered by oral gavage at 0.8 mL/kg to simulate human oral intake while ensuring consistent dosing and overcoming voluntary ingestion variability in rodents.
Index Examination of the F0 Generation
General Status
During pregnancy, delivery, and lactation, the animals’ appearance, behavioral activities, and mortality were observed daily, as well as vaginal bleeding, abortion, premature delivery, dystocia, and abnormal signs were recorded.
Body Weight and Food Intake
Body weight was measured on GD0, GD3, GD6, GD9, GD12, GD15, GD18, and GD21. Food intake was recorded daily between GD0-1, and then every three days [GD3, GD7, GD9, GD12-13, GD19-20, and on lactation day (LD) 0–1], using pre-weighed chow and subtracting uneaten food to calculate intake per animal.
Reproductive Ability
Delivery and litter were examined from GD20 to GD24. After 21 days of lactation, the females were euthanized, and gross dissection was examined, as well as the number of corpora lutea and implantations. Conception rate (%) = (number of pregnant animals/number of successfully mated female rats) × 100, pre-implantation loss rate (%) = [(number of corpora lutea - number of implantations) / number of corpora lutea] × 100, post-implantation loss rate (%) = [(number of implantations - number of live fetuses) / number of implantations] × 100, stillbirth incidence rate (%) = (number of stillbirths/number of implantations) × 100, relative organ weight = (organ weight/body weight) × 1000.
Hormone Levels
Testosterone, progesterone and estradiol levels of pregnant rats in the satellite group were sampled at GD6 (before the first dose), GD19 (after the last dose) and LD21 (on the day of whelping), respectively. If no LD21 sample was collected, that pregnant rat was excluded.
Serum hormone levels (testosterone, progesterone, estradiol) were measured using ELISA kits (Manufacturer: [eg, Cusabio Biotech Co., Ltd]; Catalogue Nos.: CSB-E05110r, CSB-E05104r, CSB-E07401r). The lower limit of quantification (LLOQ) for each assay was: testosterone = 0.2 ng/mL, progesterone = 0.5 ng/mL, estradiol = 0.1 ng/mL. Intra- and inter-assay coefficients of variation (CVs) were <10% and <15%, respectively. Each sample was run in duplicate, and any discrepancies >15% were repeated.
Index Examination of the F1 Generation
General Status
The F1 generation offspring were weaned on postnatal day 21 (PND21). The whole nest with the dam was observed before weaning, and individual animals were observed after weaning. The animals were observed daily for signs of appearance, behavioral activities, and mortality, and abnormal signs were recorded.
Appearance and Sex
The sex and appearance of the newborns were checked. The ratio of male fetuses (%) = (number of male live fetuses / total number of live fetuses) × 100.
Anogenital Distance (AGD) and Male Hypospadias
All offspring were examined for AGD on PND21, and male offspring were examined for AGD on PND54. Male offspring were examined for hypospadias on PND21 and PND54.
Incidence rate of male hypospadias (%) = (number of male hypospadias fetuses/number of male fetuses) × 100.
AGD was measured using digital Vernier calipers with 0.01 mm precision. The anatomical landmarks for measurement were from the center of the anus to the base of the genital tubercle.7 Measurements were performed in duplicate by two trained, blinded observers. The mean of both measurements was used for analysis, and discrepancies >5% were resolved by consensus.
Male offspring were examined for hypospadias on PND21 and PND54. Diagnostic criteria included abnormally located urethral meatus (eg, along the shaft or perineal area) and presence of chordee.9
Gross Dissection, Organ Weights, and Hormone Levels
All offspring were euthanized and subjected to gross autopsy. Testes and epididymis were weighed in male offspring on PND54, and blood samples were collected from the smallest numbered male and female offspring in each litter for estradiol, progesterone, and testosterone level measurements.
Prior to weighing, all reproductive organs were dissected carefully and trimmed of attached adipose and mesenteric tissues. Weighing was conducted using a calibrated digital balance (Mettler Toledo; calibration certificate valid within 6 months of experiment). Relative organ weight was calculated as (organ weight/body weight) × 1000.
Statistical Analysis
All statistical analyses were conducted using GraphPad Prism (version 10.1.2), SPSS (version 29.0), and R (version 4.3.1) with relevant packages (lme4, emmeans, and car). Data normality and homogeneity of variance were assessed using the Shapiro–Wilk and Levene’s tests, respectively. Depending on these results, either one-way ANOVA followed by Dunnett’s multiple comparisons test or the non-parametric Kruskal–Wallis test followed by Dunn’s post hoc test was applied. Categorical variables were analyzed using the Chi-square or Fisher’s exact test, as appropriate. All statistical tests were two-tailed with a significance threshold of α = 0.05.
For all F1 endpoints—including anogenital distance (AGD), hormone levels, and organ weights—the litter was considered the experimental unit in accordance with OECD and ICH guidelines. When multiple pups were sampled from a litter, their values were averaged to generate a single representative value. For more complex analyses requiring individual-level data, linear mixed-effects models (LMMs) or nested ANOVA were applied, with litter included as a random factor to account for intra-litter correlation. This approach prevents pseudo-replication and ensures appropriate control of Type I error. For multiple comparisons, either Dunnett-adjusted P-values or Benjamini–Hochberg false discovery rate (FDR) corrections were applied as appropriate.
For each key outcome, group means ± standard deviations, percentage change from control, and effect sizes (Cohen’s d or η2) with 95% confidence intervals were reported to reflect the magnitude and precision of observed effects.
Sample sizes were determined using a priori power analysis with GPower 3.1, which indicated that 14–16 litters per group would be sufficient to detect medium effect sizes (Cohen’s d = 0.65) with 80% power at α = 0.05, assuming an intra-group standard deviation of 15–20%. Based on this, 16 pregnant rats per group were included in the main study, and 7 per group in the satellite study.
Results
Basic State and Index Changes of the F0 Generation
General Status of the F0 Female Rats
The general condition of the female rats in each allylestrenol dose group was similar to that of the Sol-C, mainly reflected in the good general condition of the animals during gestation without any abnormality. During lactation, the mothers in each group were lactating normally, and some of the mothers were found to eat their pups, with percentages of 4/23, 5/22, 2/23, and 4/21, respectively. Abnormal conditions were observed in the PgI-C and PgSC-C groups. In the PgI-C, edema and hair loss at the injection site occurred during pregnancy as the dosing cycle lengthened. After 21 days of lactation, all hair loss recovered, but most of the edema at the injection site did not resolve. During lactation, some females (4/21) exhibited pup-eating behavior, two females did not lactate or care for their pups, and one female did not swallow the placenta. In the PgSC-C, during the administration period of pregnancy, the rats showed decreased activity, depression, ataxia, and disappearance of the righting reflex from GD6 to GD19 but recovered after drug withdrawal. During the non-administration period of pregnancy, the animals were generally in good condition with no abnormalities. One female rat was found dead on GD8 (2 days after administration), and no obvious abnormalities were found in various organs by necropsy. Comprehensive clinical observation and gross anatomical examination showed that the cause of death could not be ruled out as being related to the PgSC-C. During lactation, the mothers were normal, but some of them (7/19) exhibited pup-eating behavior.
Growth and Feeding of the F0 Female Rats
There were no significant changes in body weight and weight gain in the PgI-C and allylestrenol dose groups compared with the Sol-C (P > 0.05) (Figure 1A and B). In the PgSC-C, only the body weight gain from GD6 to GD9 was significantly reduced (P < 0.01), and the body weight gain from GD0 to GD21 was slightly reduced (P < 0.05), with decreases of 73.8% and 15.4%, respectively (Figure 1C), showing a statistically significant difference. The other time points of the weight check showed no statistical differences (P > 0.05). There was no significant change (P > 0.05) in food intake in all groups when compared to the Sol-C (Figure 1D).
 |
Figure 1 Growth and food intake of pregnant rats. (A) Body weight of pregnant rats; (B) Weight gain in pregnant rats (3 days); (C) Weight gain in pregnant rats (before administration, during drug administration, net weight gain); (D) Food intake of pregnant rats. |
Gross Dissection of the F0 Female Rats
There were no obvious macroscopic abnormalities on the body surface, natural orifices, thoracic cavity, abdominal cavity, or pelvic cavity in all groups of animals.
Reproductive Capacity of the F0 Female Rats
Compared with the Sol-C, there were no significant differences in the pregnancy rate, gestational days, number of corpora lutea, number of implantations, number of live fetuses, pre-implantation/post-implantation loss rate, and stillbirth incidence (litter) in the PgSC-C and the allylestrenol dose groups (P > 0.05). However, most of the animals in the PgI-C had difficulty in delivery and/or prolonged labor. Compared with the Sol-C, the gestational days, implantation loss rate, and stillbirth incidence were significantly increased (P < 0.01), while the number of live fetuses was significantly decreased (P < 0.05) (Table 1). Detailed reproductive parameters including pregnancy rate, gestational length, corpora lutea count, and fetal outcomes are summarized in Table 1.
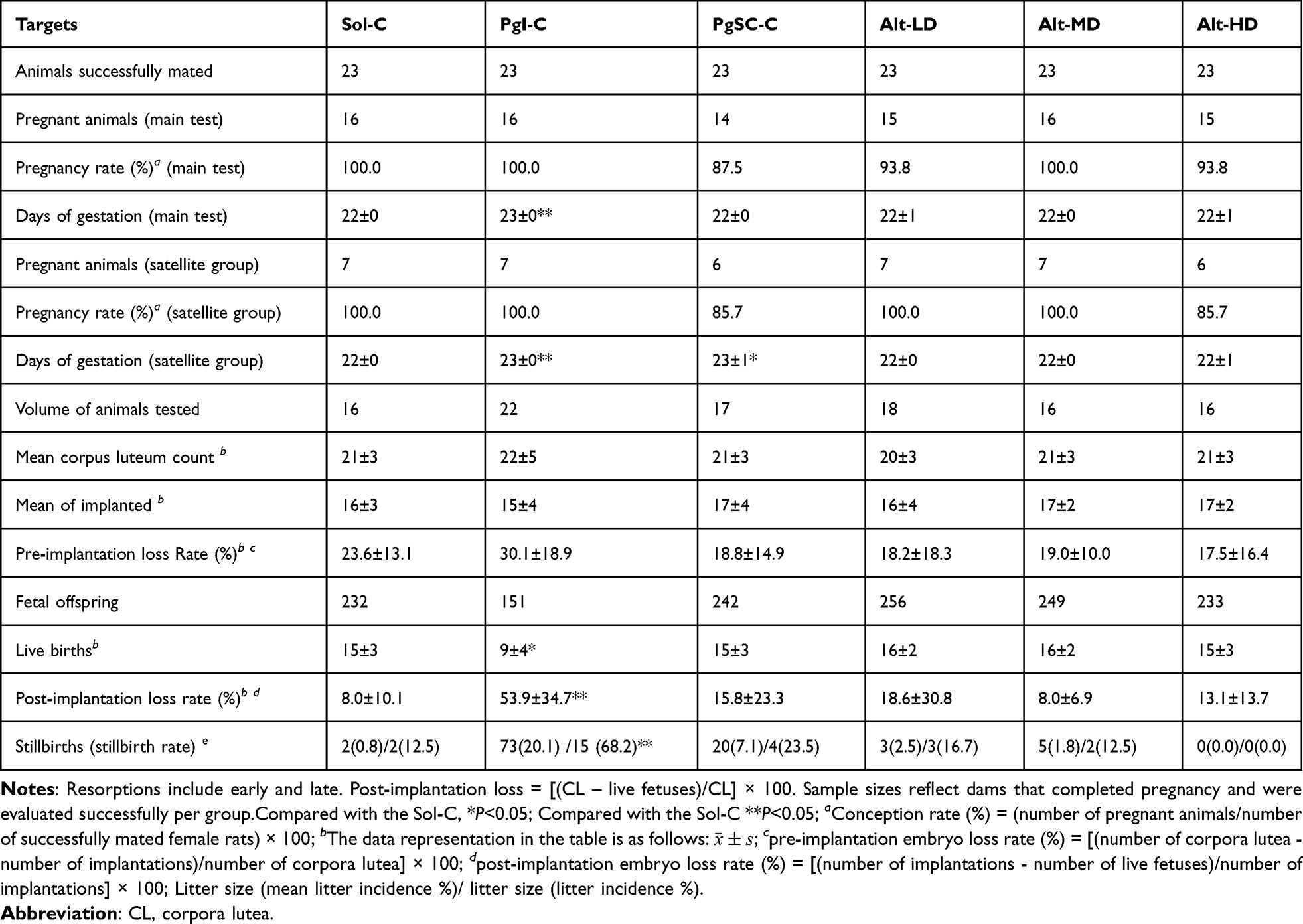 |
Table 1 Reproductive and Developmental Outcomes in F0 Females (Main-Test Animals Only). Values are Presented as Mean ± Standard Deviation |
Effects on Hormone Levels in the F0 Female Rats
The changes in hormone levels in each group compared to the Sol-C were as follows: There was no significant change in testosterone levels in the different dosage groups of allylestrenol and PgI-C, and there was no statistical difference (P > 0.05). The GD19 testosterone level in the PgSC-C increased by 64.7%, and the statistical difference was significant (P < 0.01). There was no significant effect on progesterone levels between all groups (P > 0.05). No significant difference in estradiol levels was observed in PgI-C, PgSC-C, Alt-LD and Alt-MD groups (P > 0.05).
On gestation day 19, estradiol levels in the Alt-HD group (9.98 ± 0.51 ng/mL) were significantly reduced compared to Sol-C controls (18.73 ± 4.77 ng/mL), representing a 46.7% decrease (Cohen’s d = 2.48; 95% CI: −11.02 to −6.79; P < 0.01, Dunnett-adjusted). The Alt-LD (17.58 ± 3.02 ng/mL) and Alt-MD (11.93 ± 3.66 ng/mL) groups showed non-significant trends toward reduction (−6.1% and −36.3%, respectively) (Table 2). Numerical values for testosterone, progesterone, and estradiol concentrations at GD6, GD19, and LD21 across all treatment groups are provided in Table 2. Only the PgSC-C group exhibited a statistically significant increase in testosterone levels on GD19 (12.94 ± 1.72 ng/mL) compared to Sol-C (9.86 ± 1.93 ng/mL; P = 0.021, Dunnett-adjusted; Cohen’s d = 1.71). Although testosterone levels were numerically elevated in the Alt-MD (11.45 ± 2.01 ng/mL) and Alt-HD (11.98 ± 2.13 ng/mL) groups, these differences were not statistically significant.
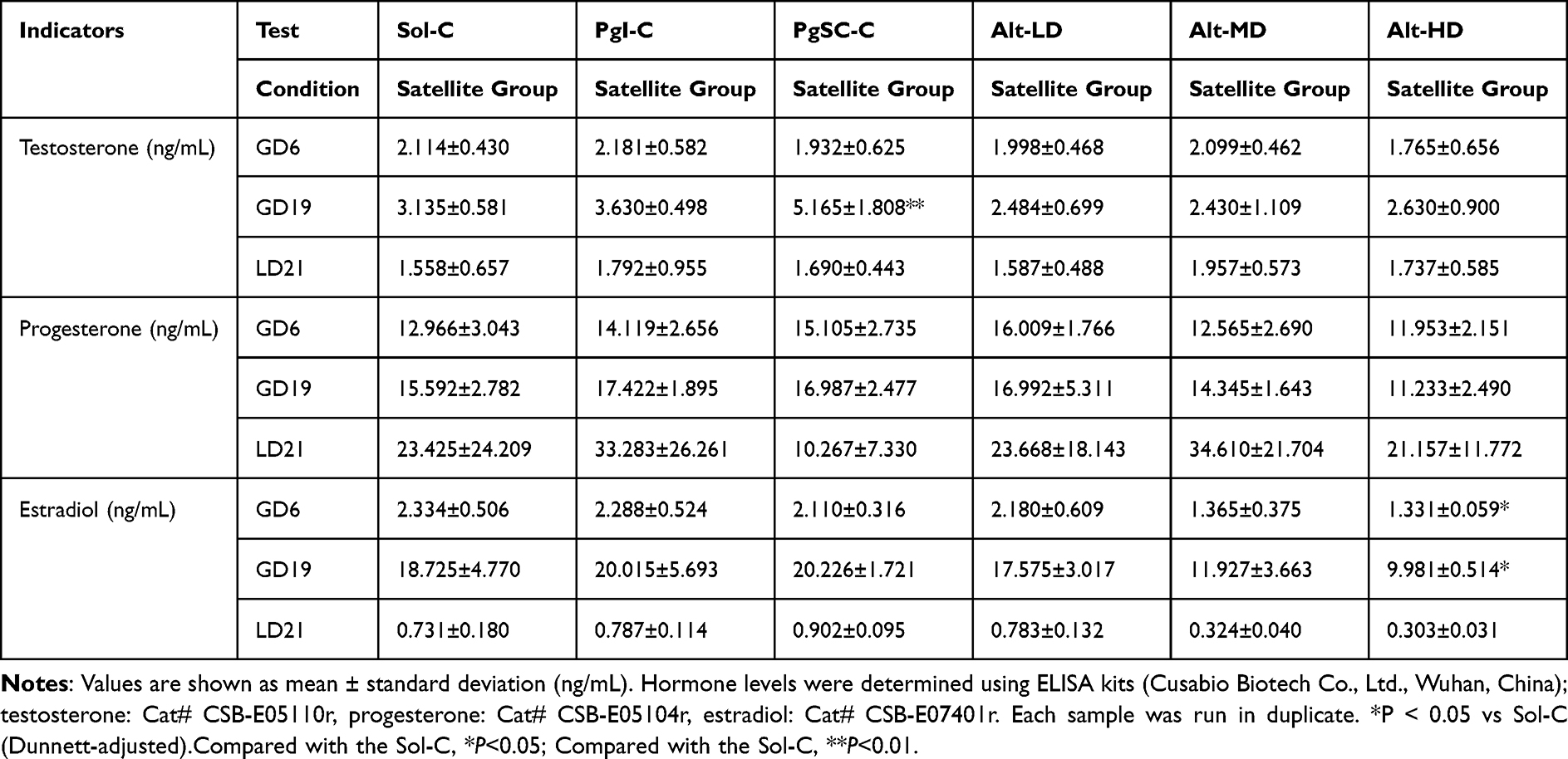 |
Table 2 Maternal Serum Concentrations of Testosterone, Progesterone, and Estradiol on Gestational Days 6 and 19 and Lactation Day 21 |
Basic State and Index Changes of the F1 Generation
General State of the F1 Generation
The surviving offspring in each group were generally in good condition before and after weaning, with good growth and development, normal activities, no abnormal secretions from the eyes, nose, and mouth, and normal urine and feces. Before weaning, some litter deaths occurred in the Alt-LD, Alt-MD, Alt-HD, PgI-C, PgSC-C, and Sol-C groups, with rates of 3/22, 6/22, 5/21, 4/21, 4/19 and 4/23, respectively. After weaning, 8(from 2 F0 females), 8(3), 2(2), and 1(1) male pups from the Sol-C, PgSC-C, Alt-LD, Alt-MD, and Alt-HD groups died during PND24~26.
Comparison of Appearance and Sex of the F1 Generation
In the Sol-C, PgSC-C, Alt-LD, Alt-MD, and Alt-HD groups, some of the offspring rats were small fetuses (numbers were 4, 3, 3, 1, and 2, respectively), while no small mouse fetuses appeared in the PgI-C. There was no significant difference in the average incidence of small fetuses, the litter incidence, or the sex ratio between each group and the Sol-C (P > 0.05) (Figure 2A–C). There were no obvious abnormalities in the appearance of offspring rats in each group.
 |
Figure 2 Comparison of the Appearance and Sex of Rats. Note: (A) Number of Fetal Rats and Litter Size (B) Incidence of Small Fetuses (C) Sex of Pups in Groups. |
AGD and Hypospadias in the F1 Generation
Male offspring rats in each group were examined on PND21 and PND54, and no hypospadias were found. At PND21, the mean AGD in male pups from the Alt-HD group was 5.21 ± 0.42 mm versus 5.35 ± 0.47 mm in the Sol-C group — a non-significant 2.6% decrease (Cohen’s d = 0.31; 95% CI: −0.39 to 0.13). Female AGD values also showed no significant difference (Alt-HD: 2.42 ± 0.28 mm vs Sol-C: 2.45 ± 0.30 mm; P > 0.05). The AGD of female and male offspring animals in the PgI-C on PND21 were slightly longer (P < 0.01), with increases of 17.0% and 17.3%, respectively. However, there was non-significant (P > 0.05) change in the AGD on PND54 in the PgI-C (Figure 3). Anogenital distance (AGD) results in male and female offspring at PND21 and PND54 are shown in Figure 3.
 |
Figure 3 Anogenital distance (AGD) in male and female offspring on PND21 and PND54. Bars represent mean ± SD of litter averages. Statistical analysis used litter as the unit (n = number of litters per group). AGD measured in millimeters using calibrated calipers. |
Gross Dissection and Organ Weight of the F1 Generation
Upon dissection, one male littermate in the Sol-C was found to have no thymus, while the other animals showed no obvious macroscopic abnormalities on the body surface, natural orifices, cranial cavity, thoracic cavity, abdominal cavity, pelvic cavity, or intraluminal tissues and organs. Testicular weight in F1 males from the Alt-MD group averaged 1.89 ± 0.22 g, compared to 1.92 ± 0.25 g in the Sol-C group, reflecting a 1.6% decrease (Cohen’s d = 0.12; 95% CI: −0.14 to 0.09; P = 0.64). Epididymal weights were also comparable across groups (Figure 4). Testicular and epididymal weights and relative organ weights across groups are illustrated in Figure 4.
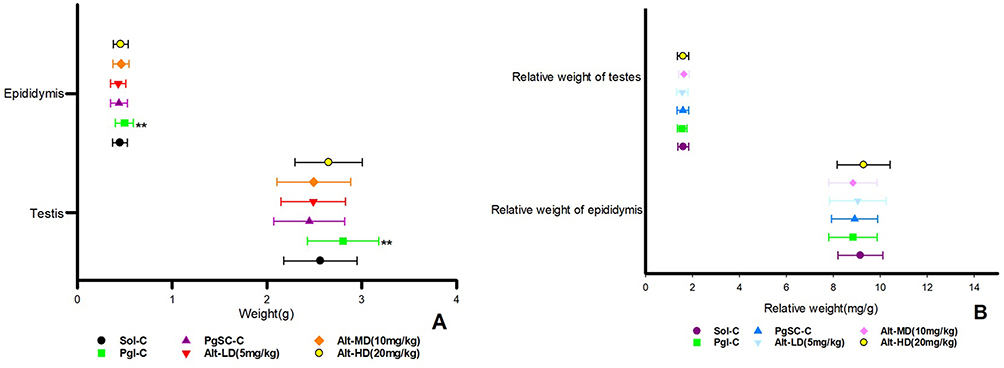 |
Figure 4 The weight and relative weight of epididymis and testicles of offspring rats in each group. (A) Weight of epididymis and testicles; (B) Relative weight of epididymis and testicles. Compared with the Sol-C, **P < 0.01. |
Hormone Levels of the F1 Generation
Hormone Levels of the F1 Males
There was no significant difference in the levels of testosterone, progesterone, and estradiol between the male offspring of each group compared with the Sol-C (P > 0.05).
Hormone Levels of the F1 Females
The testosterone levels in the PgI-C and Alt-HD had no significant changes compared with Sol-C (P > 0.05). However, there were significant (P < 0.01) increases in testosterone levels among PgSC-C, Alt-LD, and Alt-MD, which increased by 64.5%, 48.4%, and 35.7%, respectively (P > 0.05). Progesterone levels in the Alt groups and PgSC-C did not change significantly compared with the Sol-C (P > 0.05); while in the PgI-C the progesterone level increased significantly (P <0.05), with a growth of 34.9%. Compared with the Sol-C, there was no significant change in the estradiol level in PgI-C (P > 0.05). However, the estradiol level in the PgSC-C decreased slightly (P < 0.05), with a decrease of 13.9%; the estradiol levels in the Alt groups all decreased slightly (P < 0.05), with decreases of 15.8%, 17.4%, and 21.2%, respectively. When compared with PgI-C and PgSC-C, estradiol levels in the Alt group were the same in each group, and no significant differences were found. (P > 0.05) (Table 3). Full hormone profiles (testosterone, progesterone, estradiol) for male and female F1 offspring are provided in Table 3.
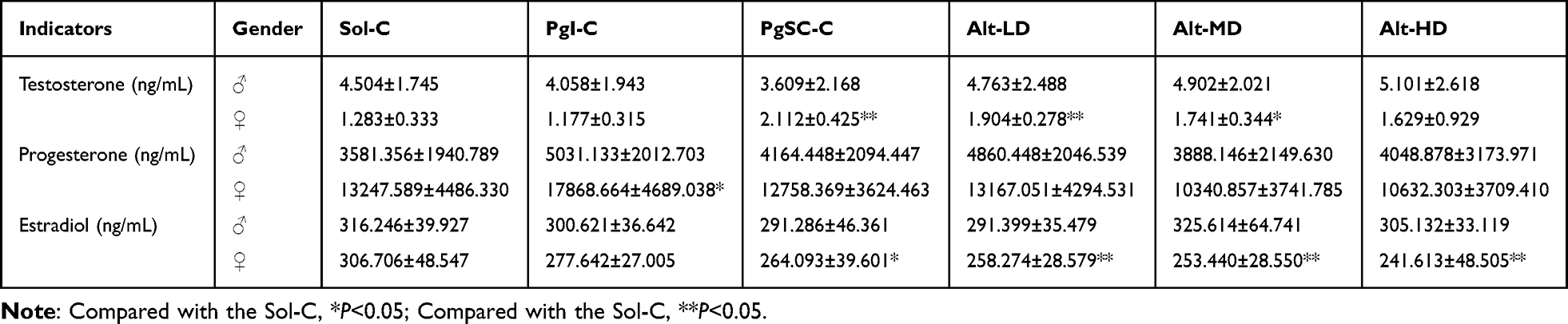 |
Table 3 Statistical Table of Hormone Content in the F1 Generation |
Discussion
Threatened abortion, recurrent miscarriage, and threatened preterm birth represent distinct but interrelated complications of pregnancy that often stem from overlapping pathophysiological mechanisms, including luteal insufficiency, hormonal imbalance, and immune dysregulation.16–18 Progesterone therapy remains a cornerstone in the management of these conditions, given its essential role in maintaining uterine quiescence, modulating the maternal immune response, and supporting early gestational development.1,2 However, natural progesterone is limited by poor oral bioavailability due to hepatic first-pass metabolism, while intramuscular formulations can cause discomfort, inflammation, and poor patient adherence.4,5,19
These clinical limitations have prompted growing interest in alternative synthetic progestogens with improved pharmacological profiles. Allylestrenol, with the chemical structure 17-alpha-allyl-4-estren-17-beta-ol, is a synthetic progestin similar to progesterone that has been used in several Asian countries for over five decades to manage threatened miscarriage and preterm labor. Despite its long-standing clinical use, concerns persist over the developmental safety of progestins during pregnancy, with some compounds linked to adverse fetal outcomes such as hypospadias, mediated by off-target androgenic effects.19–22 Although allylestrenol is generally regarded as non-androgenic, its developmental safety profile—especially in preclinical models—remains inadequately characterized.
In the current study, allylestrenol was administered orally to pregnant SD rats at doses of 5, 10, and 20 mg/kg from gestational day (GD) 6 to 19. Across all dose groups, no statistically significant changes were observed in maternal body weight gain (GD0–GD21) compared to the solvent control (Sol-C: 110.2 ± 9.4 g vs Alt-HD: 107.8 ± 10.6 g; P > 0.05). Anogenital distance (AGD) in male offspring at PND21 in the Alt-HD group was 5.21 ± 0.42 mm vs 5.35 ± 0.47 mm in Sol-C (absolute difference: −0.14 mm; 95% CI: −0.39 to 0.13; Cohen’s d = 0.31; P = 0.27). Similarly, the stillbirth rate in Alt-HD dams was 0.0% compared with 0.8% in Sol-C and 20.1% in the PgI-C group. Testicular weights in F1 males from the Alt-MD group (1.89 ± 0.22 g) were also comparable to Sol-C (1.92 ± 0.25 g; absolute difference: −0.03 g; P = 0.64). These findings suggest a lack of gross developmental toxicity or androgenic effects under the exposure conditions tested.
A notable exception was the statistically significant reduction in maternal estradiol levels on GD19 in the Alt-HD group (9.98 ± 0.51 ng/mL) compared to Sol-C (18.73 ± 4.77 ng/mL), corresponding to a 46.7% decrease (P < 0.01; Cohen’s d = 2.48; 95% CI: −11.02 to −6.79). Although this decline was substantial, it did not correlate with downstream effects on offspring viability, AGD, or reproductive organ weights. Reported reference ranges for serum estradiol in late gestation (GD18–20) in SD rats typically span 15–25 ng/mL,19–24 suggesting that the observed estradiol level in Alt-HD dams falls slightly below the lower physiological threshold. However, the absence of significant morphological or functional abnormalities in the F1 generation indicates that this transient hormonal suppression may not have disrupted key developmental windows, potentially due to compensatory endocrine mechanisms or limited exposure duration. Nevertheless, this estradiol reduction should be monitored in future multigenerational studies.
To achieve full exposure of allylestrenol in the parental body, the administration period for the F0 females was prolonged as far as possible (GD6~19). Results showed that there were no significant changes in body weight, body weight gain, and food intake in the allylestrenol groups, with no obvious abnormalities being found for animals in each group after dissection. By contrast, in the F1 offspring, no significant abnormalities of general appearance and anatomy of littermates were observed across all groups, no hypospadias were observed, and no significant differences in AGD worth noting Chen et al25 used progesterone combined with allylestrenol in elderly patients with threatened abortion, which showed that, compared with progesterone alone, combined treatment can improve pregnancy outcomes and increase β-human chorionic gonadotropin (β-hCG), and progesterone levels. Yu et al26 conducted an observational study on clinical pregnancy in patients treated with assisted reproductive technology, and the results showed that allylestrenol had similar clinical pregnancy rates and pregnancy outcomes compared to dydrogesterone. Allylestrenol reduced the incidence of multiple pregnancies while improving endometrial thickness, shape, and blood flow. This shows that allylestrenol has certain clinical efficacy in improving pregnancy outcomes, which supports the results of this study.
For the Alt group, only limited changes in hormone levels between the parental and female offspring were demonstrated in testosterone, progesterone, and estradiol sex hormone levels. Estradiol levels in F0 females on GD19 demonstrated a dose-related downward trend, with statistically significant reduction observed only in the high-dose group. While values in the low- and medium-dose groups were numerically lower than controls, they did not reach statistical significance after correction. The estradiol level of F1 females decreased by 15.8%-21.2%, which was similar to that of the PgI-C and PgSC-C. Considering that the estradiol level is easily affected by the different periods in physiological states of F0 females (pre-estrus, estrus, post-estrus, and inter-estrus), which resulted in large individual fluctuations and a wide normal range of values. Also, in combination with the growth and autopsy of offspring, it could be concluded that a slight decrease in estradiol does not lead to significant organic and functional changes in the animals. Therefore, allylestrenol doses of 5, 10, and 20 mg kg−1 are safe. Shen et al27 treated patients with threatened abortion with progesterone combined with allylestrenol, and the results showed that it could improve the levels of monocyte chemoattractant protein-1 (MCP-1), interleukin-6 (IL-6), transforming growth factor-β1 (TGF-β1), progesterone-induced blocking factor (PIBF), and sex hormones, while increasing the rate of fetal protection. Li Q et al28 used allylestrenol combined with ritodrine to treat patients with threatened abortion. As a result, the pregnancy time and delivery time were prolonged, IL-17, IL-10, and IL-6 levels were improved, and the neonatal weight and Apgar score were also significantly optimized. Moreover, the incidence of neonatal deaths, malformations and asphyxia is reduced. It is suggested that allylestrenol can improve pregnancy-related indicators.
No direct statistical correlation was observed between maternal estradiol reductions in Alt-HD dams and changes in offspring AGD, viability, or organ weight. Similarly, elevated fetal testosterone in PgSC-C was not associated with significant increases in AGD or reproductive organ weights.13 These findings may reflect a threshold effect, compensatory endocrine mechanisms, or insufficient duration of exposure to elicit phenotypic changes.
The ~46% reduction in estradiol levels observed in Alt-HD dams on gestation day 19 falls below typical physiological ranges reported during late rat pregnancy (15–25 ng/mL). This reduction was not accompanied by overt developmental abnormalities in offspring, suggesting that either the timing, duration, or magnitude of estradiol suppression was insufficient to disrupt key developmental windows. In this study, the litter was used as the primary experimental unit for analyzing offspring endpoints such as anogenital distance (AGD), organ weights, and serum hormone levels, consistent with OECD TG 443 and ICH S5 guidelines.9 When multiple pups were evaluated per litter, their values were averaged to yield one representative measurement per dam, thereby preventing pseudo-replication and minimizing Type I error risk. While this conservative approach strengthens the validity of statistical comparisons, it may limit the detection of subtle within-litter variation or sex-specific differences. For more granular insights, future analyses will apply linear mixed-effects models (LMMs) that incorporate individual-level data with litter as a random factor. This method will allow for partitioning of within-litter and between-litter variances, providing a more detailed understanding of potential exposure effects while maintaining appropriate error control.4 Conversely, PgSC-C males exhibited elevated fetal testosterone, which aligns with known androgenic programming during male reproductive development, yet did not manifest in structural abnormalities. These findings suggest a possible decoupling between transient hormonal shifts and phenotypic endpoints under the current exposure conditions.
The observed dose-dependent decrease in estradiol levels in Alt-LD to Alt-HD groups, particularly the 46.7% reduction at 20 mg/kg on GD19, may reflect a pharmacodynamic effect of allylestrenol rather than off-target toxicity. One plausible mechanism involves suppression of aromatase activity or hypothalamic-pituitary-gonadal (HPG) axis feedback. Specifically, allylestrenol may exert negative feedback on the hypothalamus or pituitary, reducing luteinizing hormone (LH) and follicle-stimulating hormone (FSH) secretion, thereby downregulating ovarian estrogen synthesis. Alternatively, it may modulate placental aromatase expression or activity, which converts androgens to estrogens during pregnancy. Although these mechanisms remain speculative, they are consistent with the known hormonal feedback loops of steroidal progestins.24,25 Future mechanistic studies involving gonadotropin profiling and aromatase expression assays in ovarian or placental tissues would help elucidate these pathways.
Although allylestrenol has demonstrated efficacy in improving pregnancy outcomes in women with threatened abortion or luteal insufficiency,25–28 the present study employed healthy, gestationally normal SD rats. This choice follows standard developmental toxicity testing guidelines designed to detect baseline reproductive hazards, not therapeutic efficacy under pathological conditions. The lack of adverse findings in this non-diseased model still offers important insight into the developmental safety of allylestrenol. However, it does not fully recapitulate human clinical indications. Thus, while this study provides foundational toxicological data, future evaluations in disease-relevant animal models—such as rats with pharmacologically induced luteal insufficiency or inflammation-driven preterm labor—would enhance translational relevance and allow better alignment between preclinical pharmacology and clinical use.
While some endpoints did not meet statistical significance thresholds, the observed biological patterns suggest potential toxicological effects. Notably, the >50% post-implantation loss in the PgI-C group—though not statistically compared here—raises concern regarding reproductive toxicity associated with this formulation. In contrast, small differences in stillbirth rates (eg, 0.8% vs 1.8%) across groups lacked both statistical and biological significance. These findings underscore the importance of interpreting effect size and clinical relevance, not just P-values, in reproductive toxicology.
One maternal death occurred in the PgSC-C group on gestational day 8; though no necropsy or histopathological assessment was performed, this may be related to anesthetic complications during blood collection.15 Transient depression and ataxia observed in PgSC-C animals could be due to vehicle-related CNS effects or excipient-induced stress; this warrants further exploration in dedicated safety studies. Localized edema at the PgI-C injection site likely reflects formulation intolerance, and future studies should incorporate histological examination or photographic documentation of injection sites to better characterize local tissue reactions. No other treatment-related mortality or severe clinical signs were observed.
This study has several limitations that should be acknowledged. First, the long-term effects of allylestrenol on the reproductive capacity and endocrine health of F1 offspring were not assessed, as mating trials and sexual maturation endpoints were not included. Second, evaluation of F2 generation outcomes was not performed, which limits conclusions on transgenerational safety. Third, only serum hormone levels were measured without corresponding tissue-level assessments, which may underestimate subtle local effects. Finally, the study was conducted in a single rodent species (SD rats); extrapolation to humans would benefit from confirmation in other animal models. Future studies will address these gaps by incorporating histological, molecular, and functional reproductive endpoints across generations and species.
Conclusion
Allylestrenol administration at doses of 5, 10, and 20 mg/kg in SD rats pregnant during the gestational duration did not exhibit any overt developmental issues in F1 offspring. No remarkable differences in the physical appearance of offspring, anogenital distance, and androgenic effect indication were found by the investigation. The estradiol, testosterone, and progesterone hormone levels changed only marginally during the course of the study, with estradiol levels decreasing dose dependently by up to 46% (P < 0.05). This study shows that allylestrenol is a safe treatment for the maintenance of pregnancy at the doses tested without any evidence of gross developmental toxicities. Additional studies are needed to evaluate the long-term consequences and functional reproductive health, as well as a transgenerational effect assessment, to confirm the extended safety profile of allylestrenol.
Data Sharing Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Ethics Approval and Consent to Participate
The study of animals used in this study follows the Guide for the Care and Use of Laboratory Animals (2011, 8th edition). The institute is accredited by the International Association for Assessment and Accreditation of Laboratory Animal Care International (AAALAC). The method of animal use was approved (approval number: 2023071101) by the Institutional Animal Care and Use Committee (IACUC) of Fudan University Shanghai Medical College. The consent to participate was not applicable.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work. Lai Jiang and Shengnan Zhuan are co-first authors of the article. Dajin Li and He Wen are Co-corresponding authors.
Funding
There is no funding to report.
Disclosure
All authors disclosed no relevant relationships.
References
1. Bulletti C, Bulletti FM, Sciorio R, et al. Progesterone: the key factor of the beginning of life. Int J Mol Sci. 2022;23(22):14138. doi:10.3390/ijms232214138
2. Nagy B, Szekeres-Barthó J, Kovács GL, et al. Key to life: physiological role and clinical implications of progesterone. Int J Mol Sci. 2021;22(20):11039. doi:10.3390/ijms222011039
3. Dhillon-Smith RK, Melo P, Kaur R, et al. Interventions to prevent miscarriage. Fertil Sterility. 2023;120(5):951–954. doi:10.1016/j.fertnstert.2023.08.955
4. Shynlova O, Nadeem L, Lye S. Progesterone control of myometrial contractility. J Steroid Biochem Mol Biol. 2023;234:106397. doi:10.1016/j.jsbmb.2023.106397
5. Di Renzo GC, Tosto V, Tsibizova V. Progesterone: history, facts, and artifacts. Best Pract Res Clin Obstet Gynaecol. 2020;69:2–12. doi:10.1016/j.bpobgyn.2020.07.012
6. Ge BB, Kong S. Clinical study on Yunkang Syrup combined with allylestrenol in treatment of threatened abortion. Drugs Clinic. 2022;37(7):1579–1583.
7. Deng T, Liao X, Zhu S, et al. Recent advances in treatment of recurrent spontaneous abortion. Obstet Gynecology Surv. 2022;77(6):355–366. doi:10.1097/OGX.0000000000001033
8. Zheng CC, Wang HP, Xu BH, et al. Toxicity of Jingfang granule extract on embryo-fetal developmental of rats. Chin J Pharmacovigil. 2022;19(5):493–497.
9. Lim S, McDougall ARA, Goldstein M, et al. Analysis of a maternal health medicines pipeline database 2000–2021: new candidates for the prevention and treatment of fetal growth restriction. BJOG. 2023;130(6):653–663. doi:10.1111/1471-0528.17392
10. Yu N, Kwak-Kim J, Bao S, et al. Unexplained recurrent pregnancy loss: novel causes and advanced treatment. J Reprod Immunol. 2023;155(1):103785. doi:10.1016/j.jri.2022.103785
11. Bilir A, Atay E, Firat F, et al. Investigation of developmental toxicity of favipiravir on fetal bone and embryonic development. Birth Defects Res. 2022;114(17):1092–1100. doi:10.1002/bdr2.2073
12. Yang X, Tian Y, Zheng L, et al. The update immune-regulatory role of pro- and anti-inflammatory cytokines in recurrent pregnancy losses. Int J Mol Sci. 2022;24(1):132. doi:10.3390/ijms24010132
13. Zhang CP, Liu JY, Liu FY, et al. Clinical study on gushen antai pills combined with allylestrenol in treatment of threatened abortion. Drugs Clinic. 2020;35(5):893–897.
14. Malhotra N. Oral allylestrenol: a pregnancy-supporting progestogen. J South Asian Feder Obs Gynae. 2017;9(4):297–303.
15. Akay MA, Yıldız GE. Impact of gestational and parental factors and maternal intake of progesterone on the development of hypospadias: a retrospective case-control study. Taiwan J Obstet Gynecol. 2021;60(5):894–898. doi:10.1016/j.tjog.2021.08.001
16. Cornish EF, McDonnell T, Williams DJ, et al. Chronic inflammatory placental disorders associated with recurrent adverse pregnancy outcome. Front Immunol. 2022;13(1):825075. doi:10.3389/fimmu.2022.825075
17. Giouleka S, Tsakiridis I, Arsenaki E, et al. Investigation and management of recurrent pregnancy loss: a comprehensive review of guidelines. Obstet Gynecol Surv. 2023;78(5):287–301. doi:10.1097/OGX.0000000000001133
18. Li XX, Ding JP. Clinical study on baotailing capsules combined with allylestrenol in treatment of habitual abortion. Drugs Clinic. 2020;35(5):975–979.
19. Sivaraman L, Sanderson T. Gamma secretase inhibition: effects on fertility and embryo-fetal development in rats. Toxicol Appl Pharmacol. 2023;469(1):116512. doi:10.1016/j.taap.2023.116512
20. Shen J, Lu BB, Lu F, et al. Embryo-fetal toxicity and concomitant toxicokinetic assessment of PDQ in SD rats. Chin J New Drugs. 2022;31(14):1426–1432.
21. Devall AJ, Papadopoulou A, Podesek M, et al. Progestogens for preventing miscarriage: a network meta-analysis. Cochrane Database Syst Rev. 2021;4(4):13792.
22. Lv D, Liu H, An Q, et al. Association of adverse fetal outcomes with placental inflammation after oral gestational exposure to hexafluoropropylene oxide dimer acid (GenX) in Sprague-Dawley rats. J Hazard Mater. 2024;461(1):132536. doi:10.1016/j.jhazmat.2023.132536
23. Azmi MB, Khan W, Azim MK, et al. Identification of potential therapeutic intervening targets by in-silico analysis of nsSNPs in preterm birth-related genes. PLoS One. 2023;18(3):e0280305. doi:10.1371/journal.pone.0280305
24. Pang YY, Ma CL. Real-world pharmacological treatment patterns of patients with threatened miscarriage in China from 2014 to 2020: a cross-sectional analysis. J Clin Pharm Ther. 2022;47(2):228–236. doi:10.1111/jcpt.13536
25. Chen Y, Ai MH. The effects of progesterone combined with allylestradiol on serum CA125, β-hCG and P in elderly patients with threatened abortion. Chin J Hum Sex. 2021;30(9):101–103.
26. Yu Y, Yu T, Ding W, et al. Evaluation of allylestrenol for clinical pregnancies in patients treated with assisted reproductive techniques: a retrospective, propensity score matched, observational study. BMC Pregnancy Childbirth. 2023;23(1):660. doi:10.1186/s12884-023-05970-2
27. Shen M, Du EQ, Wang P. Effects of progesterone combined with alllestradiol tablets on MCP-1, PIBF and sex hormone levels in patients with threatened abortion. Maternal Child Health Care China. 2022;37(10):1838–1842.
28. Li Q, Li C, Jin H. Efficacy of allylestrenol combined with ritodrine on threatened premature labor and its influence on inflammatory factors in peripheral blood. Exp Ther Med. 2020;19(2):907–912. doi:10.3892/etm.2019.8273
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.