Back to Archived Journals » International Journal of Clinical Transfusion Medicine » Volume 7
Potential of next-generation sequencing to match blood group antigens for transfusion
Authors Orzinska A ![]() , Guz K
, Guz K ![]() , Brojer E
, Brojer E
Received 20 May 2019
Accepted for publication 30 July 2019
Published 3 September 2019 Volume 2019:7 Pages 11—22
DOI https://doi.org/10.2147/IJCTM.S175142
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Cees Th. Smit Sibinga
Agnieszka Orzinska, Katarzyna Guz, Ewa Brojer
Institute of Hematology and Transfusion Medicine, Department of Hematological and Transfusion Immunology, Warsaw, Poland
Correspondence: Agnieszka Orzinska
Department of Hematological and Transfusion Immunology, Institute of Hematology and Transfusion Medicine, 14 Gandhi Street, Warsaw 02-776, Poland
Email [email protected]
Abstract: A review of the advances in applying next-generation sequencing (NGS) to transfusion medicine for the purpose of genotyping alleles encoding clinically important red blood cell and platelet antigens. NGS data from published studies confirm the possibility of antigen prediction based on sequencing of the whole genome, exome or targeted regions. What remains a challenge, to provide highly accurate NGS genotyping, is the further improvement of bioinformatic solutions for automated interpretation based on publicly accessible and improved reference databases appropriate for NGS methods as well as validation of a method based on the examination of a large number of individuals. There is no doubt, however, as to the future of NGS as a supplementary test used to provide highly compatible blood as well as to reduce the risk of patient’s alloimmunization. This is part of personalized medicine.
Keywords: alloimmunization, transfusion medicine, whole genome, exome, allele
Prevention of alloimmunization against blood group antigens
Transfusion is the procedure of introducing donor material with unknown blood cell antigens into the recipient’s circulatory system. The recipient’s immune system recognizes foreign antigens, produces specific antibodies and sensitization (alloimmunization) occurs.1,2 To date, more than 300 red blood cell (RBC) and 33 human platelet antigens (HPA) have been described.3,4 Antibodies against these antigens may have varying clinical significance and cause acute hemolytic reactions, delayed hemolytic reactions, hemolytic disease of the fetus/newborn or ineffective platelet transfusion in the case of sensitization to platelet antigens.5,6 Some alloantibodies are clinically irrelevant. According to the Serious Hazard of Transfusion data, hemolytic transfusion reactions were responsible for 8% of deaths attributed to the presence of antibodies acting against donor RBC antigens.7
The percentage of patients immunized as a result of blood transfusion varies from 5% to 50%.8–10 The underlying disease and ethnic origin of the donor as well as the recipient are also important factors in the risk of alloimmunization.11 Transfusion-dependent patients (with thalassemia or hemoglobinopathies such as sickle cell disease [SCD]) are particularly prone to sensitization. The prevalence of alloantibody detection in this group is up to 50% as compared to patients with histories of a single transfusion.12 It seems that the number of the recipient’s exposures to donor antigens may be significant in triggering the immunological mechanisms. It is therefore recommended to match the donor blood units with the recipient’s phenotype.
Since the beginning of transfusion history, serological methods have been the gold standard of pre-transfusion testing to determine blood group antigens and prevent hemolytic reactions in the recipient.13–15 Current blood transfusion procedures include compatibility of ABO antigens and focus on avoiding transfusion of RhD positive units to RhD negative recipients to prevent alloimmunization against the most immunogenic D antigen. The procedures assume that patients are treated with blood from donors compatible only in ABO/RhD antigens. Prevention of alloimmunization against antigens other than D is rather limited.17–19 A donor compatible in other antigens is selected only when a patient produces antibodies to a specific antigen. The process of identification of antibody specificity and selection of compatible donor is often complicated and requires multistep laboratory testing by highly qualified staff.16
Extended antigen typing is time-consuming, serological methods are costly and depend on the availability of reagents for antigen detection. The procedure is usually performed in reference laboratories which complicates and delays the delivery of blood for transfusion. Access to extended antigen-typed blood components requires the creation of a large registry of extended antigen-typed donors.
Blood group genotyping
Molecular techniques are an alternative approach for extended phenotype prediction.20,21 In part, they make it possible to overcome the limitations of serological tests.22–25 In immunohematology, molecular methods have become more popular since the 1990s when the genetic background of antigen specificities was first described.26,27 A single nucleotide variant (SNV) is the most common variation resulting from a change of amino acid in the protein sequence which leads to either the presence or absence of the antigen.28 This change determines the HPA, human neutrophil antigen as well as a large number of RBC antigens. The variability of erythrocyte antigens is also determined by other mechanisms such as deletions of the entire gene, indels or structural variants such as the hybrid genes RHD/RHCE or GYPA/GYPB. In addition, blood group polymorphisms may result from mutations in any other gene necessary for epitope expression (eg, KLF1, RHAG).29,30 Our knowledge about alleles encoding antigens is still expanding and antigen databases such as that of the International Society of Blood Transfusion are continuously updated.31,32
According to the guidelines and recommendations in developed countries, molecular methods are used in applications such as extended antigen genotyping for patients prone to alloimmunization, those who have been recently transfused or confirmation of genotype in cases with antibodies, especially when antisera are unavailable.33–35 Since the subject of the currently used genetic methods extends beyond the subject matter of this paper, all those interested can find more details in the cited papers.33–41 In general, at present, molecular blood antigen genotyping is commonly used not only in reference laboratories but also as a routine serological procedure in hospitals as an additional tool for donor/recipient antigen phenotype prediction and profiling complex cases. It simplifies further serological and molecular analysis. Genotyping determines the absence of antigens, which facilitates antibody identification and the indication for transfusion. Various commercial as well as home-made tests developed in reference laboratories are widely used for profiling patients both in individual tests as well as mass screening.33–40 Data from mass donor screening with blood group genotyping platforms have confirmed the high accuracy based on comparisons with serological results.36–40 DNA assays verify serological identification in cases of weak antigen expression and null phenotypes.41 However, DNA platforms have limitations that lead to the false determination of blood groups. The main disadvantage of all these solutions is the use of defined oligos complementary to known allelic variants. Genotyping fails for mutations in the tested region and gives a false negative result. False results are also reported when SNV occurs next to the tested region and affects the expression.42–44 The software used for the interpretation of results requires regular updating to be compatible with the updated allelic databases, but even then it is impossible to test all the known alternations.
Next-generation sequencing (NGS) technology
Recent years have witnessed the development of new molecular technologies for testing the genome sequence in high-throughput mode that overcome the limitations of routinely used DNA methods (Box 1).45,46 Studies of the whole human genome commenced with a project (1000 Genome Project) in which a full catalog of DNA variants from different populations was obtained by classical sequencing.47 Following this project, there was a significant development of technical solutions adapted to the needs of sequencing with a high throughput, the development of tools for reading DNA sequences and for collecting and analyzing the data obtained, and as a result, the development of the high-throughput solution known as NGS. The currently available technologies allow various levels of sequencing; either the whole genome (WGS), coding regions – exons (WES) or only selected genes or regions of interest. NGS technology significantly reduces the cost of testing. It has been successfully implemented in transplantation medicine for testing donors’ genotypes of HLA antigens in high-throughput mode.48 Over 9000 HLA alleles for over 500 individuals can be identified per run. Such a huge capacity is a green light for blood group and platelet gene screening in blood donors as well as for the setting up of a database with compatible blood units dedicated for transfusion.
 |
Box 1 Advantages of NGS overcoming the limitations of routinely used BG antigen genotyping platforms |
NGS in revealing unknown blood group variations
NGS is particularly effective for finding unknown variations responsible for different phenotypes in patients with antibodies of unknown specificity because it enables screening of the whole genome, exome or particular genes and finding a “needle in a haystack” – an unknown or rare variant. Recent studies have confirmed NGS effectiveness in resolving the molecular background of orphan antigens with an as yet unknown genetic basis.49–59 The WES strategy based on genomic DNA from patients with detected antibodies and their relatives combined with appropriate variant filtering have revealed one SNV in the GYPA gene.49 This discovery allowed the SARA antigen to join the MNS blood group system. The WES solution was applied for identification of a 17-bp deletion in the SMIM1 gene in Vel negative individuals.50 The WES strategy also helped to define the molecular background of the AnWj antigen negativity through indication of homozygous SNV in SMYD1 gene, which encodes a transcription factor not directly associated with blood group genes.15 NGS technology was recently used for resolving specificity of unknown antibodies acting against the AUG1 antigen and revealed a novel allele, encoding the AUG3 antigen.52,53 The WES approach was also used for recent identification of the missense variation in the PRNP gene responsible for the KANNO antigen negativity in the Japanese population.54 Another successful example was the use of NGS for solving the Xg blood group molecular background as reported by Yeh et al.55 The study revealed a variation in a GATA motif located beyond the XG gene but with an impact on its transcription.
Schoeman et al described the molecular background of seven unexplained cases with complex serological results.56 The WES method with the TruSightOne panel and Illumina technology revealed rare and novel variants in the Rh, Duffy, Lan, Cromer blood group systems and KLF1 from the transcription factor family. Similarly, with the use of customized panel targeting exons encoding 15 blood group systems and Ion Torrent sequencing platform, Jakobsen et al detected rare single nucleotide variations that may correspond to a variant or weakly expressed antigens.57 Wen et al tested RHAG and ANK1 sequences for explaining weaker Rh expression.58 The NGS analysis with a custom-made primer panel for exons of genes involved in RBC membrane disorders identified a homozygous variant of the RHAG gene which changes interactions of the RhAG-RhD complex. In a group of RhD negative individuals, El Wafi et al revealed five novel intronic RHD variants that probably affected the splicing of D protein.59
Moller et al analyzed the genomic sequences of genes encoding 36 blood group systems from over 2500 individuals enrolled in the 1000 Genome Projects.32 The study revealed 1241 non-synonymous variants. Their clinical role is unknown and 80% of them were absent in reference databases. Analysis of these sequencing data also led to the discovery of a huge deletion of 5–7 exons in the ABO gene described as the third common variation in an African population with O blood group phenotype.60
Montemayor-Garcia et al presented a new geographical distribution of allelic blood group variants, the description of which was based on genomic sequences from the same project.61 The report presented a novel distribution of known variants and 12 novel variants, probably antigenic in 4 cases, from the Kell, Kidd, Scianna and Cromer systems due to the predicted amino acid substitution.
All the above findings highlight the usefulness of NGS for detecting novel, unsuspected genetic variants. Each new discovery expands our knowledge and if they are submitted and introduced into the appropriate reference databases they will contribute to the better identification of patient/donor genotypes and phenotypes and more effective donor–recipient antigen matching. However, currently, the problem hindering the possibilities of studies is the lack of an accurately developed and verified, publicly accessible database of reference sequences in the form of a program adapted to work with the data obtained by NGS.
NGS in blood group screening
NGS is also effective in reducing the risk of post-transfusion alloimmunization since the huge capacity of one experiment enables the immediate and cost-effective determination of all RBC and platelet antigen genotypes. Study results support extended profiling of donors and patients for the best prophylactic antigen matching to prevent alloimmunization.16,22–24
Literature on the use of NGS for studying blood group antigens shows a picture of rapidly developing technology which may prove highly reliable. The technology is now being validated and upgraded especially in terms of antigens with a complex genetic background or with highly homological regions.62–67 Once the bioinformatics and economic solutions have been worked out, NGS will undoubtedly supersede other genetic methods used for predicting blood group phenotypes based on genotyping with predefined oligos. To date, difficulties in the sequencing of alleles encoding blood cell antigens have only partially been solved. Targeted panels for sequencing the coding regions of blood cell antigens have been designed. Some difficulties related to the high homology of the RHD/RHCE and GYA/GYPB/GYPE genes are beginning to be overcome, the database of blood group and platelet reference allele sequences is currently being improved for correct alignment, and special algorithms are being designed and amended for correct phasing and haplotype reconstruction from short-read sequences. The first software for automatic blood group phenotype interpretation based on WSG data has been developed. Automation of the NGS process required for high-throughput donor testing has been performed. The main task now is to improve the accuracy of NGS tests by creating an expanded database of allelic sequences found in different populations with related serological data. It will then be possible to develop a standardized panel for diagnostic tests for proper analysis of donor or recipient data. Numerous NGS reports on blood grouping reveal the presence of SNVs of unknown serological characteristics and unknown clinical significance that are absent in reference databases. The crucial step now is to supplement the reference database with hundreds of thousands of genomic sequences, paired with individual serological and genomic backgrounds obtained with other molecular techniques. Once the task has been performed NGS can be implemented into routine diagnostics to replace serological antigen determinations.
Table 1 presents a list of original papers considered to be milestones in the development of NGS technology and applications in transfusion medicine. They indicate the difficulties related to genotyping blood group antigens and introducing NGS as a basic tool for the next-generation serologists.
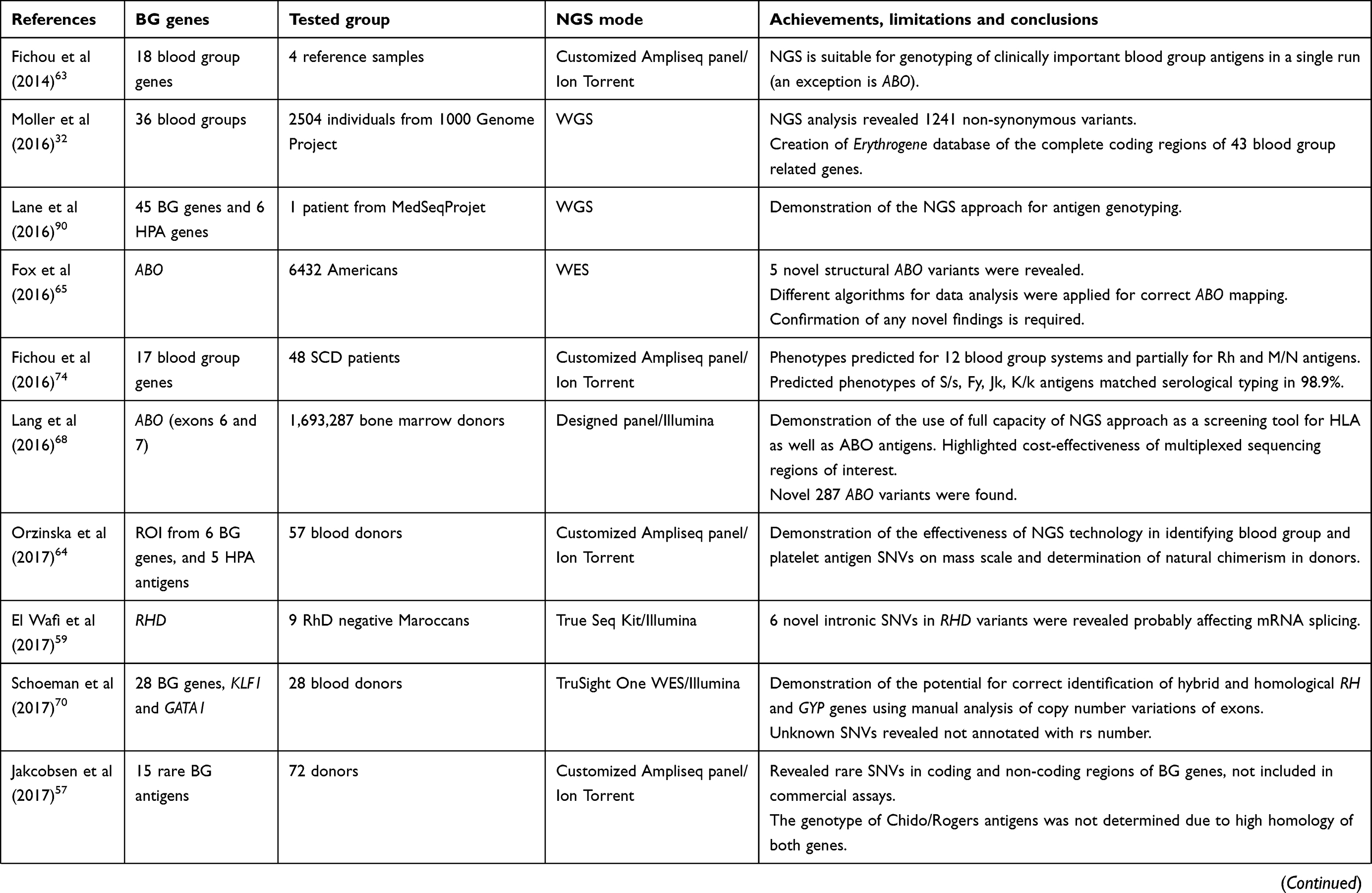 | 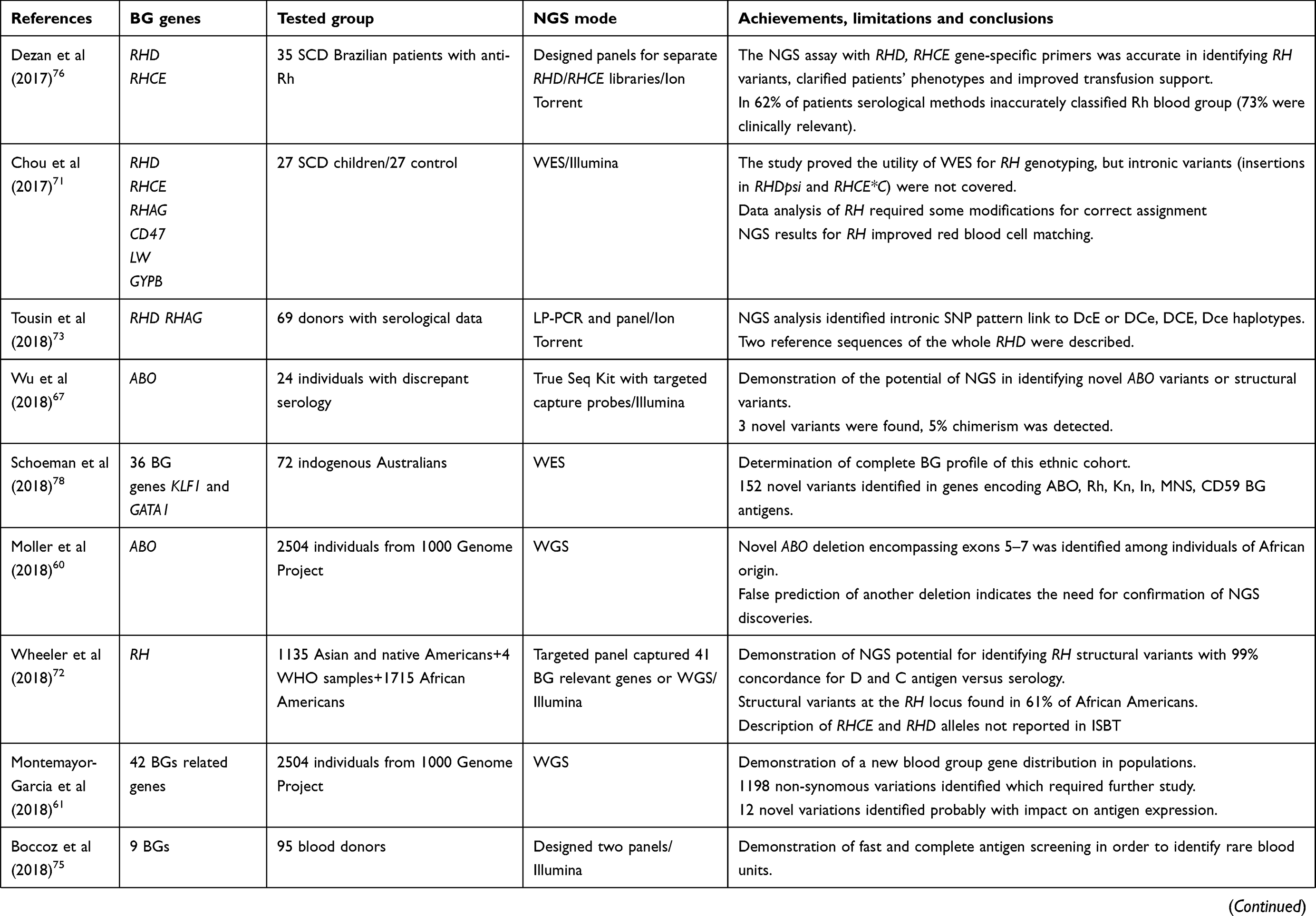 |  |
Table 1 Published reports on application of NGS to red blood cell and platelet antigen genotyping for donor/recipient antigen matching |
In their study, Fichou et al present NGS in comprehensive blood group typing and report the application of a targeted panel for NGS of 18 genes.63 In parallel, they explore the coding and intronic sequences encoding 15 blood group systems using four well-characterized DNA samples from the EQC program. To overcome the misalignment of sequenced regions from the homologous genes RH and GYP, the authors used specific primers for library preparation. In a single run, they obtained sequences that covered 86% of all coding regions, with the exception of the ABO antigen coding region. In our NGS study, we also focused on sequencing regions of interest flanking polymorphisms encoding blood group and platelet antigens in 57 blood donors and used primers designed by AmpliSeq software, complemented with RHD and RHCE*C/c specific primers incorporated into a customized mixture of primers.64 Our results were consistent with the serological and genotyping outcome, except for GYPA and intron 2 of RHCE*C with lack of or a poor number of amplicons. The above studies prove the utility of the NGS approach as a screening tool dedicated for testing donors and indicate the main difficulties concerning complex or homological genes.
Donor–recipient compatibility with regard to ABO is of crucial importance, so precise identification of ABO subtypes is a challenge. However, ABO gene genotyping is complicated by the complex genetic background of the A and B glycosyltransferases indirectly responsible for the presence of A and B antigens. NGS is the best method for their identification since the large numbers of variants are associated with SNVs in exons 6 and 7. A recent report on the testing of these regions was performed in parallel with HLA testing in bone marrow donors and proved the NGS capacity for ABO screening as well as its cost-effectiveness.48 Fox et al suggested another approach based on extraction of ABO data from WES of individuals of African origin revealing an unknown deletion.65 Their report demonstrates the importance of manual data inspection, despite applied algorithms, and the need to confirm with other molecular techniques in the case of novel variants. Lane et al report an accurate prediction of ABO from WGS data.66 Automated typing with a slightly modified algorithm called bloodTyper rendered 98% concordance with the serology. The work highlighted the necessity of further validation of the performance of software designed to interpret blood groups from data acquired from sequencing of known subtypes or hybrid ABO variants. Wu et al demonstrated the state of art and limitations in the NGS solution of the heterogeneity of the ABO molecular background in serologically discrepant samples.67 With the new technique of customized capture probes, the authors sequenced the entire ABO gene and successfully identified both known and novel alleles, microchimerism and cis/trans association. With this approach, however, it was impossible to phase the haplotypes encompassing exons 6 and 7 in all the samples. A longer sequencing platform (PacBio or Oxford Nanopore) seems to be the best solution for correct phasing of complex genes, as Lang et al have already demonstrated for confirmation of novel ABO allelic sequences identified in bone marrow donors.68
A current review of NGS technology applied to the prediction of Rh blood group antigens reveals a similar pattern of solutions and limitations as in the case of ABO described above. For Rh, the sequencing is even more complicated due to the presence of a high variety of alleles and mixed RHD/RHCE hybrid variants and a complex membrane structure. Interpretation of results from short NGS reads is extremely difficult because correct alignment to the reference common allele is often unclear. To overcome the similarity of RHD and RHCE, gene-specific primers were applied in targeted customized panels and separate libraries developed for both homological genes with different barcodes.69 The tools for data analysis and algorithms were modified for correct assignment. Schoeman et al demonstrated the need for manual interpretation of copy number variation for correct identification of RH rearrangements from the data of 28 individuals with a targeted exon panel.70 With the purpose of correct mapping and localization of RHD/RHCE genes, Chou et al calculated read depth proportion for each of the 54 patients with SCD tested in their study. Misalignment of fragments of RH genes was identified as the main source of WES errors which gave approximately 91.7% concordance with other genotyping methods.71 Lane et al proposed the bloodTyper tool for automated copy number analysis of RH from WGS data with a final 100% concordance for RhD typing and 99.5% accuracy at RhC typing.66 Recently, Wheeler et al have reported the application of a customized panel for specific RH detection.72 It makes use of information from flanking intronic regions for the correct mapping of short NGS reads of highly homological sequences including the problematic exons 1–2 and 8 misaligned to the opposite RH gene by other researchers. With this approach, the authors successfully identified RH structural variations in a large multi-ethnic group and correctly predicted RhC expression in 99.2% of hybrid cases. On the other hand, Tounsi et al used long-range PCR amplicons of the complete RHD and performed WGS of 69 individuals with known Rh serology in order to establish a reference RHD allele for NGS analyses.73 They found multiple intronic SNVs among them, 23 specific to DcE haplotype and 15 linked to DCe, DCE and Dce haplotypes, giving two references RHD sequences deposited to GenBank.
NGS prediction of other clinically relevant and rare blood group antigens is highly accurate with the exception of the highly homological genes GYP and CH/RG encoding the MNS and Chido–Rogers blood group systems.63,70,75 Study reports on the application of WGS, WES or a customized panel strategy demonstrate that for screening blood donor antigens and patient antigen profiling it is desirable to acquire an individual genotype of common and rare alleles in one step (see Table 1).
Applications of NGS in transfusion medicine
The application of NGS technology for blood typing contributes to the following aspects of patient care:
Regularly transfused patients are susceptible to sensitization against foreign donor antigens. The different ethnic origins of the donor/recipient, frequently of African origin and carrying Rh variants, are the main cause of alloimmunization. For such patients, precise NGS genotyping as described by Dezan et al has been proved extremely useful and cost-effective.76 NGS gene-specific assay for typing of 35 SCD patients with unexplained Rh antibodies revealed false serological Rh classification in 62% cases and misinterpretation of anti-Rhe alloantibodies as autoantibodies. According to NGS data, 10% of patients were reclassified as RhD positive which made it possible to save the already scarce/limited supplies of RhD negative blood products as well as to reduce the risk of alloimmunization against Rhc and Rhe antigens associated with RhD negative units from Caucasian donors.
Patients from specific populations or those with unexplained complex serological results will particularly benefit from NGS analysis used as a first-line test instead of relying on extended phenotyping, multistep antibody identification and numerous single or screening genetic tests.77–79 In 2014, Mazonson et al estimated the costs of pre-transfusion tests at about 195 USD per patient but for patients with autoimmune hemolytic anemia the cost increased to 1490 USD as compared to 65 USD for six blood genes when using the NGS approach.80
Patients with antibodies to high-frequency antigens and antibodies to numerous antigens will benefit when tested with the NGS approach which provides the recipient’s profile of antigens in one step. Moreover, if the donor is genotype-matched further alloantibody formation is limited.82 It is worth mentioning that there is a growing number of patients with WES or WGS performed for diagnostic reasons and their NGS blood group data is available for transfusion purposes. This also concerns immunized pregnant women who may take advantage of non-invasive prenatal testing to predict feto-maternal compatibility in fetal blood group or platelet antigens. The fetal and maternal blood group and platelet antigen genotypes are determined with a targeted panel for sequencing and in parallel the antigen profile is provided for potential transfusions in the fetus and the mother.83
The issue also concerns an important group of special donors relied upon for production of RBC panel reagents used for antibody screening and identification in all immunohematological laboratories worldwide. NGS typing of all blood group genes in these “products” will upgrade the interpretation of serological results and antigen matching.84
Summary
In the future, the NGS technique will contribute to an extension of the donor registry for immunized patients increasing transfusion safety and reducing the risk of delayed post-transfusion reactions. Well-characterized blood products will become more available. NGS provides a deeper insight into the sequence of genes and regions of interest, it determines SNVs and indels together with the quantitative measurement of copy number variations in one experiment with a large number of samples. An important advantage of this technology as compared to serological tests and other high-throughput genetic applications is the ability to identify variations of clinically significant antigens in a single sample. Other authors have also demonstrated NGS to be the correct technology for achieving significant progress in blood transfusion medicine. Implemented on a mass scale, this approach would facilitate access to donors with a known, extended phenotype, and donors with rare and atypical phenotypes. Establishing a digital database of results of NGS blood group testing is of the utmost importance. This sort of professional registry of both donors and recipients should be linked to open access serological data for national blood services. Before applying NGS technology to routine blood group diagnostics, there are some challenges to be faced.85,86 One of them is the analysis of NGS data which involves determination of the human reference sequences, validation and standardization of algorithms for interpretation and classification of genetic variants with their haplotype localization and for translation of final NGS results to the predicted phenotype. The current accuracy of NGS of blood group antigens is about 95% and should be validated in a large group after the development of a final study protocol together with an accessible program comparing with a reference sequence database. This step depends on data of sequenced individuals with paired serological data collected in a large-scale database. The economic aspect related to the implementation of NGS in routine diagnostics of blood donors as well as targeted groups of patients remains an open issue. The technology is undoubtedly cheaper but still too expensive. At the level of NGS laboratories, the preparation of CE-IVD marked products and external quality control programs are a challenge as well as the logistics of NGS testing in a centralized and cost-effective way.87 Another challenge are the huge amounts of NGS data that have to be collected, protected and used for other clinical purposes. Finally, the ethics related to NGS need to be adapted and supplemented with informed consent from both donors and patients. According to a round table discussion at the AABB Meeting, NGS typing of blood donors requires proper ethical guidelines concerning incidental genetic findings such as pathology or clinical phenotype known to be associated with the detected variations.88,89
Disclosure
The authors report no conflicts of interest in this work.
References
1. Klein HG, Anstee DJ. Haemolytic transfusion reactions. In: Mollison’s Blood Transfusion in Clinical Medicine.
2. Zimring JC, Welniak L, Semple JW, et al. Current problems and future directions of transfusion-induced alloimmunization: summary of an NHLBI working group. Transfusion. 2011;51(2):435–441. doi:10.1111/j.1537-2995.2010.03024.x
3. Storry JR, Clausen FB, Castilho L, et al. International society of blood transfusion working party on red cell immunogenetics and blood group terminology: report of the Dubai, Copenhagen and Toronto meetings. Vox Sang. 2019;114(1):95–102. doi:10.1111/vox.12717
4. Wikman A, Mörtberg A, Sachs UJ, Santoso S. Report on the 18th Platelet Immunology Workshop of the ISBT 2016. ISBT Sci Ser. 2017;12(1):214–222. doi:10.1111/voxs.12336
5. Garratty G. What is a clinically significant antibody? ISBT Sci Ser. 2012;7:54–57. doi:10.1111/j.1751-2824.2012.01594.x
6. Tormey CA, Hendrickson JE. Transfusion-related red blood cell alloantibodies: induction and consequences. Blood. 2019;133(17):1821–1830. doi:10.1182/blood-2018-08-833962
7. Available from: https://www.shotuk.org/wp-content/uploads/myimages/SHOT-Report-2017-WEB-Final-v4-25-9-18.pdf.
8. Yazdanbakhsh K, Ware RE, Noizat-Pirenne F. Red blood cell alloimmunization in sickle cell disease: pathophysiology, risk factors, and transfusion management. Blood. 2012;120(3):528–537. doi:10.1182/blood-2011-11-327361
9. Hendrickson JE, Tormey CA, Shaz BH. Red blood cell alloimmunization mitigation strategies. Transfus Med Rev. 2014;28(3):137–144. doi:10.1016/j.tmrv.2014.04.008
10. ST C, Jackson T, Vege S, Smith-Whitley K, Friedman DF, Westhoff CM. High prevalence of red blood cell alloimmunization in sickle cell disease despite transfusion from Rh-matched minority donors. Blood. 2013;122(6):1062–1071. doi:10.1182/blood-2013-03-490623
11. Fasano RM, Chou ST. Red blood cell antigen genotyping for sickle cell disease, thalassemia, and other transfusion complications. Transfus Med Rev. 2016;30(4):197–201. doi:10.1016/j.tmrv.2016.05.011
12. Zheng Y, Maitta RW. Alloimmunisation rates of sickle cell disease patients in the United States differ from those in other geographical regions. Transfus Med. 2016;26(3):225–230. doi:10.1111/tme.12314
13. MEM Y, Josephson CD, Winkler AM, et al. Red blood cell minor antigen mismatches during chronic transfusion therapy for sickle cell anemia. Transfusion. 2017;57(11):2738–2746. doi:10.1111/trf.14282
14. O’Suoji C, Liem RI, Mack AK, et al. Alloimmunization in sickle cell anemia in the era of extended red cell typing. Pediatr Blood Cancer. 2013;60:1487–1491. doi:10.1002/pbc.24530
15. Available from: https://www.transfusionguidelines.org/transfusion-handbook.
16. Belsito A, Costa D, Signoriello S, et al. Clinical outcome of transfusions with extended red blood cell matching in β-thalassemia patients: a single-center experience. Transfus Apher Sci. 2019;58(1):65–71. doi:10.1016/j.transci.2018.11.006
17. Ambruso DR. Transfusion complications in thalassemias. Transfusion. 2014;54(4):957–959. doi:10.1111/trf.12576
18. Matteocci A, Pierelli L. Red blood cell alloimmunization in sickle cell disease and in thalassaemia: current status, future perspectives and potential role of molecular typing. Vox Sang. 2014;106(3):197–208. doi:10.1111/vox.12086
19. Wahl SK, Garcia A, Hagar W, Gildengorin G, Quirolo K, Vichinsky E. Lower alloimmunization rates in pediatric sickle cell patients on chronic erythrocytapheresis compared to chronic simple transfusions. Transfusion. 2012;52(12):2671–2676. doi:10.1111/j.1537-2995.2012.03659.x
20. Westhoff CM. Blood group genotyping. Blood. 2019;133(17):1814–1820. doi:10.1182/blood-2018-11-833954
21. Wheeler MM, Johnsen JM. The role of genomics in transfusion medicine. Curr Opin Hematol. 2018;25(6):509–515. doi:10.1097/MOH.0000000000000469
22. Khan J, Delaney M. Transfusion support of minority patients: extended antigen donor typing and recruitment of minority blood donors. Transfus Med Hemother. 2018;45(4):271–276. doi:10.1159/000491883
23. Kulkarni S, Choudhary B, Gogri H, et al. Molecular genotyping of clinically important blood group antigens in patients with thalassaemia. Indian J Med Res. 2018;148(6):713–720. doi:10.4103/ijmr.IJMR_455_17
24. Belsito A, Magnussen K, Napoli C. Emerging strategies of blood group genotyping for patients with hemoglobinopathies. Transfus Apher Sci. 2017;56(2):206–213. doi:10.1016/j.transci.2016.11.007
25. Denomme GA, Anani WQ, Avent ND, et al. Red cell genotyping precision medicine: a conference summary. Ther Adv Hematol. 2017;8(10):277–291. doi:10.1177/2040620717729128
26. Reid ME, Lomas-Francis C, Olsson ML. The Blood Group Antigen Factbook.
27. Daniels G. Human Blood Groups.
28. Raud L, Férec C, Fichou Y. From genetic variability to phenotypic expression of blood group systems. Transfus Clin Biol. 2017;24(4):472–475. doi:10.1016/j.tracli.2017.06.011
29. Singleton BK, Frayne J, Anstee DJ. Blood group phenotypes resulting from mutations in erythroid transcription factors. Curr Opin Hematol. 2012;19(6):486–493. doi:10.1097/MOH.0b013e328358f92e
30. Polin H, Pelc-Klopotowska M, Danzer M, et al. Compound heterozygosity of two novel RHAG alleles leads to a considerable disruption of the Rh complex. Transfusion. 2016;56(4):950–955. doi:10.1111/trf.13476
31. Available from: http://www.isbtweb.org/working-parties/red-cell-immunogenetics-and-blood-group-terminology/.
32. Möller M, Jöud M, Storry JR, Olsson ML. Erythrogene: a database for in-depth analysis of the extensive variation in 36 blood group systems in the 1000 genomes project. Blood Adv. 2016;1(3):240–249. doi:10.1182/bloodadvances.2016001867
33. Belsito A, Costa D, Napoli C. Blood group genotyping for patients with autoimmune hemolytic anemia. Transl Res. 2014;164(2):177–178. doi:10.1016/j.trsl.2014.03.009
34. Svensson AM, Delaney M. Considerations of red blood cell molecular testing in transfusion medicine. Expert Rev Mol Diagn. 2015;15(11):1455–1464. doi:10.1586/14737159.2015.1086646
35. Shin KH, Lee HJ, Park KH, Hye BM, Chang CL, Kim HH. Extended red blood cell genotyping to investigate immunohematology problems. Ann Lab Med. 2018;38(4):387–388. doi:10.3343/alm.2018.38.4.387
36. McBean RS, Hyland CA, Flower RL. Blood group genotyping: the power and limitations of the Hemo ID Panel and MassARRAY platform. Immunohematology. 2015;31(2):75–80.
37. Molano A, Apraiz I, España P, et al. Performance evaluation study of ID RHD XT, a new genotyping assay for the detection of high-prevalence RhD negative and weak D types. Vox Sang. 2018;113(7):694–700. doi:10.1111/vox.12701
38. Jongruamklang P, Gassner C, Meyer S, et al. Matrix-assisted laser desorption/ionization time-of-flight mass spectrometry analysis of 36 blood group alleles among 396 Thai samples reveals region-specific variants. Transfusion. 2018;58(7):1752–1762. doi:10.1111/trf.14624
39. Finning K, Bhandari R, Sellers F, et al. Evaluation of red blood cell and platelet antigen genotyping platforms (ID CORE XT/ID HPA XT) in routine clinical practice. Blood Transfus. 2016;14(2):160–167. doi:10.2450/2015.0124-15
40. McBean RS, Hyland CA, Davis AC, et al. Blood group genotype analysis of Australian reagent red blood cell donors across three genotyping platforms: consistent detection of 70% phenotype genotype non concordance. ISBT Sci Ser. 2014;9(2):309–314. doi:10.1111/voxs.12166
41. Flegel WA, Gottschall JL, Denomme GA. Implementing mass-scale red cell genotyping at a blood center. Transfusion. 2015;55(11):2610–2615. doi:10.1111/trf.13168
42. Srivastava K, Stiles DA, Wagner FF, Flegel WA. Two large deletions extending beyond either end of the RHD gene and their red cell phenotypes. J Hum Genet. 2018;63(1):27–35. doi:10.1038/s10038-017-0345-3
43. Millard GM, Lopez GH, Turner EM, et al. Modified expression of the KEL2 (k) blood group antigen attributed to p.Leu196Val amino acid change three residues from the K/k antigen polymorphism site: implications for donor screening. Transfusion. 2019;59(3):1156–1158. doi:10.1111/trf.15106
44. Lopez GH, Turner RM, McGowan EC, et al. A DEL phenotype attributed to RHD Exon 9 sequence deletion: slipped-strand mispairing and blood group polymorphisms. Transfusion. 2018;58(3):685–691. doi:10.1111/trf.14439
45. Muzzey D, Evans EA, Lieber C. Understanding the basics of NGS: from mechanism to variant calling. Curr Genet Med Rep. 2015;3(4):158–165. doi:10.1007/s40142-015-0076-8
46. McBean RS, Hyland CA, Flower RL. Approaches to determination of a full profile of blood group genotypes: single nucleotide variant mapping and massively parallel sequencing. Comput Struct Biotechnol J. 2014;11(19):147–151. doi:10.1016/j.csbj.2014.09.009
47. 1000 Genomes Project Consortium; Auton A, Brooks LD, et al. A global reference for human genetic variation. Nature. 2015;526(7571):68–74. doi:10.1038/nature15393
48. Schöfl G, Lang K, Quenzel P, et al. 2.7 million samples genotyped for HLA by next generation sequencing: lessons learned. BMC Genomics. 2017;18(1):161. doi:10.1186/s12864-016-3396-5
49. McBean RS, Hyland CA, Hendry JL, Shabani-Rad MT, Flower RL. SARA: a “new” low-frequency MNS antigen (MNS47) provides further evidence of the extreme diversity of the MNS blood group system. Transfusion. 2015;55(6 Pt 2):1451–1456. doi:10.1111/trf.12973
50. Cvejic A, Haer-Wigman L, Stephens JC, et al. SMIM1 underlies the Vel blood group and influences red blood cell traits. Nat Genet. 2013;45(5):542–545. doi:10.1038/ng.2603
51. Yahalom V, Pillar N, Zhao Y, et al. SMYD1 is the underlying gene for the AnWj-negative blood group phenotype. Eur J Haematol. 2018;101(4):496–501. doi:10.1111/ejh.13133
52. McBean R, Liew YW, Wilson B, et al. Genotyping confirms inheritance of the rare At(a-) type in a case of haemolytic disease of the newborn. J Pathol Clin Res. 2015;2(1):53–55. doi:10.1002/cjp2.33
53. Millard GM, McGowan EC, Wilson B, et al. A proposed new low-frequency antigen in the Augustine blood group system associated with a severe case of hemolytic disease of the fetus and newborn. Transfusion. 2018;58(5):1320–1322. doi:10.1111/trf.14562
54. Omae Y, Ito S, Takeuchi M, et al. Integrative genome analysis identified the KANNO blood group antigen as prion protein. Transfusion. 2019. Epub ahead of print. doi:10.1111/trf.15319
55. Yeh CC, Chang CJ, Twu YC, et al. The molecular genetic background leading to the formation of the human erythroid-specific Xga/CD99 blood groups. Blood Adv. 2018;2(15):1854–1864. doi:10.1182/bloodadvances.2018018879
56. Schoeman EM, Roulis EV, Liew YW, et al. Targeted exome sequencing defines novel and rare variants in complex blood group serology cases for a red blood cell reference laboratory setting. Transfusion. 2018;58(2):284–293. doi:10.1111/trf.14393
57. Jakobsen MA, Dellgren C, Sheppard C, Yazer M, Sprogøe U. The use of next-generation sequencing for the determination of rare blood group genotypes. Transfus Med. 2017. doi:10.1111/tme.12496
58. Wen J, Verhagen OJHM, Jia S, et al. A variant RhAG protein encoded by the RHAG*572A allele causes serological weak D expression while maintaining normal RhCE phenotypes. Transfusion. 2019;59(1):405–411. doi:10.1111/trf.14969
59. El Wafi M, El Housse H, Zaid N, et al. Novel intronic RHD variants identified in serologically D-negative blood donors. Vox Sang. 2017;112(8):796–802. doi:10.1111/vox.12570
60. Möller M, Hellberg Å, Olsson ML. Thorough analysis of unorthodox ABO deletions called by the 1000 Genomes project. Vox Sang. 2018;113(2):185–197. doi:10.1111/vox.12613
61. Montemayor-Garcia C, Karagianni P, Stiles DA, et al. Genomic coordinates and continental distribution of 120 blood group variants reported by the 1000 Genomes Project. Transfusion. 2018;58(11):2693–2704. doi:10.1111/trf.14953
62. Leffler EM, Band G, Busby GBJ, et al. Resistance to malaria through structural variation of red blood cell invasion receptors. Science. 2017;356:6343. doi:10.1126/science.aam6393
63. Fichou Y, Audrézet MP, Guéguen P, Le Maréchal C, Férec C. Next-generation sequencing is a credible strategy for blood group genotyping. Br J Haematol. 2014;167(4):554–562. doi:10.1111/bjh.13084
64. Orzińska A, Guz K, Mikula M, et al. A preliminary evaluation of next-generation sequencing as a screening tool for targeted genotyping of erythrocyte and platelet antigens in blood donors. Blood Transfus. 2018;16(3):285–292. doi:10.2450/2017.0253-16
65. Fox K, Johnsen JM, Coe BP, et al. Analysis of exome sequencing data sets reveals structural variation in the coding region of ABO in individuals of African ancestry. Transfusion. 2016;56(11):2744–2749. doi:10.1111/trf.13797
66. Lane WJ, Westhoff CM, Gleadall NS, et al. Automated typing of red blood cell and platelet antigens: a whole-genome sequencing study. Lancet Haematol. 2018;5(6):e241–e251. doi:10.1016/S2352-3026(18)30053-X
67. Wu PC, Lin YH, Tsai LF, Chen MH, Chen PL, Pai SC. ABO genotyping with next-generation sequencing to resolve heterogeneity in donors with serology discrepancies. Transfusion. 2018;58(9):2232–2242. doi:10.1111/trf.14654
68. Lang K, Wagner I, Schöne B, et al. ABO allele-level frequency estimation based on population-scale genotyping by next generation sequencing. BMC Genomics. 2016;17:374. doi:10.1186/s12864-016-3328-4
69. Stabentheiner S, Danzer M, Niklas N, et al. Overcoming methodical limits of standard RHD genotyping by next-generation sequencing. Vox Sang. 2011;100(4):381–388. doi:10.1111/j.1423-0410.2010.01444.x
70. Schoeman EM, Lopez GH, McGowan EC, et al. Evaluation of targeted exome sequencing for 28 protein-based blood group systems, including the homologous gene systems, for blood group genotyping. Transfusion. 2017;57(4):1078–1088. doi:10.1111/trf.14054
71. Chou ST, Flanagan JM, Vege S, et al. Whole-exome sequencing for RH genotyping and alloimmunization risk in children with sickle cell anemia. Blood Adv. 2017;1(18):1414–1422. doi:10.1182/bloodadvances.2017007898
72. Wheeler MM, Lannert KW, Huston H, et al. Genomic characterization of the RH locus detects complex and novel structural variation in multi-ethnic cohorts. Genet Med. 2019;21(2):477–486. doi:10.1038/s41436-018-0074-9
73. Tounsi WA, Madgett TE, Avent ND. Complete RHD next-generation sequencing: establishment of reference RHD alleles. Blood Adv. 2018;2(20):2713–2723. doi:10.1182/bloodadvances.2018017871
74. Fichou Y, Mariez M, Le Maréchal C, Férec C. The experience of extended blood group genotyping by next-generation sequencing (NGS): investigation of patients with sickle-cell disease. Vox Sang. 2016;111(4):418–424. doi:10.1111/vox.12432
75. Boccoz SA, Fouret J, Roche M, et al. Massively parallel and multiplex blood group genotyping using next-generation-sequencing. Clin Biochem. 2018;60:71–76. doi:10.1016/j.clinbiochem.2018.07.010
76. Dezan MR, Ribeiro IH, Oliveira VB, et al. RHD and RHCE genotyping by next-generation sequencing is an effective strategy to identify molecular variants within sickle cell disease patients. Blood Cells Mol Dis. 2017;65:8–15. doi:10.1016/j.bcmd.2017.03.014
77. Quirino MG, Colli CM, Macedo LC, Sell AM, Visentainer JEL. Methods for blood group antigens detection: cost-effectiveness analysis of phenotyping and genotyping. Hematol Transfus Cell Ther. 2019;41(1):44–49. doi:10.1016/j.htct.2018.06.006
78. Schoeman EM, Roulis EV, Perry MA, Flower RL, Hyland CA. Comprehensive blood group antigen profile predictions for Western Desert Indigenous Australians from whole exome sequence data. Transfusion. 2019;59(2):768–778. doi:10.1111/trf.15047
79. Flores-Bello A, Mas-Ponte D, Rosu ME, Bosch E, Calafell F, Comas D. Sequence diversity of the Rh blood group system in Basques. Eur J Hum Genet. 2018;26(12):1859–1866. doi:10.1038/s41431-018-0232-1
80. Mazonson P, Efrusy M, Santas C, et al. The HI-STAR study: resource utilization and costs associated with serologic testing for antibody-positive patients at four United States medical centers. Transfusion. 2014;54(2):271–277. doi:10.1111/trf.12176
81. Portegys J, Rink G, Bloos P, Scharberg EA, Klüter H, Bugert P. Towards a regional registry of extended typed blood donors: molecular typing for blood group, platelet and granulocyte antigens. Transfus Med Hemother. 2018;45(5):331–340. doi:10.1159/000493555
82. Patel SR, Bennett A, Girard-Pierce K, et al. Recipient priming to one RBC alloantigen directly enhances subsequent alloimmunization in mice. Blood Adv. 2018;2(2):105–115. doi:10.1182/bloodadvances.2017010124
83. Orzińska A, Guz K, Mikula M, et al. Prediction of fetal blood group and platelet antigens from maternal plasma using next-generation sequencing. Transfusion. 2019;59(3):1102–1107. doi:10.1111/trf.15116
84. Scharberg E, Rink G, Portegys J, et al. The impact of using genotyped reagent red blood cells in antibody identification. Transfus Med Hemother. 2018;45(4):218–224. doi:10.1159/000491884
85. Hyland CA, Roulis EV, Schoeman EM. Developments beyond blood group serology in the genomics era. Br J Haematol. 2019;184(6):897–911. doi:10.1111/bjh.15747
86. Fichou Y, Férec C. NGS and blood group systems: state of the art and perspectives. Transfus Clin Biol. 2017;24(3):240–244. doi:10.1016/j.tracli.2017.06.002
87. Montemayor-Garcia C, Westhoff CM. The “next generation” reference laboratory? Transfusion. 2018;58(2):277–279. doi:10.1111/trf.14483
88. Flegel WA, Chen Q, Castilho L, et al. Molecular immunohaematology round table discussions at the AABB annual meeting, Orlando 2016. Blood Transfus. 2018;16(5):447–456. doi:10.2450/2018.0260-17
89. Anstee DJ. The relationship between blood groups and disease. Blood. 2010;115(23):4635–4643. doi:10.1182/blood-2010-01-261859
90. Lane WJ, Westhoff CM, Uy JM, et al. Comprehensive red blood cell and platelet antigen prediction from whole genome sequencing: proof of principle. Transfusion. 2016;56(3):743–754. doi:10.1111/trf.13416
91. Lane WJ, Aguad M, Smeland-Wagman R, et al. A whole genome approach for discovering the genetic basis of blood group antigens: independent confirmation for P1 and Xga. Transfusion. 2019;59(3):908–915. doi:10.1111/trf.15089
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.