Back to Journals » International Journal of Nanomedicine » Volume 21
Potential Application of Plant-Derived Extracellular Vesicles in Treatment of Burn Wounds
Authors He X, Yang Y, Niu H, Hu C, Lin J ![]()
Received 28 August 2025
Accepted for publication 31 December 2025
Published 10 January 2026 Volume 2026:21 563793
DOI https://doi.org/10.2147/IJN.S563793
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Prof. Dr. RDK Misra
Xinyu He,1,2,* Ya Yang,1,2,* Hong Niu,3 Congxia Hu,4 Jihui Lin1,2
1Department of Nursing, The Affiliated Hospital, Southwest Medical University, Luzhou, Sichuan, 646000, People’s Republic of China; 2Wound Healing Basic Research and Clinical Application Key Laboratory of LuZhou, School of Nursing, Southwest Medical University, Luzhou, Sichuan, 646000, People’s Republic of China; 3School of Pharmacy, Southwest Medical University, Luzhou, Sichuan, 646000, People’s Republic of China; 4Department of Endocrinology and Metabolism, The Affiliated Hospital, Southwest Medical University, Luzhou, Sichuan, 646000, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Congxia Hu, Department of Endocrinology and Metabolism, The Affiliated Hospital, Southwest Medical University, No. 1, Section 1, Xianglin Road, Longmata District, Luzhou, Sichuan Province, 646000, People’s Republic of China, Email [email protected] Jihui Lin, Department of Nursing, The Affiliated Hospital, Southwest Medical University, No. 1, Section 1, Xianglin Road, Longmata District, Luzhou, Sichuan Province, 646000, People’s Republic of China, Email [email protected]
Abstract: Burns are triggered by exposure of skin to heat sources, chemicals, electric currents, or radiation. Due to the cross-interaction of multiple factors, the healing mechanism of burns is complex. In recent years, plant-derived extracellular vesicles (P-EVs) have been found to possess characteristics such as anti-inflammatory, antioxidant, antibacterial, and promotion of angiogenic, owing to their rich content of proteins, nucleic acids, and secondary metabolites, which make P-EVs have great application prospects in the field of tissue regeneration. This article, based on the specificities of healing mechanisms of burn wounds compared to general wounds, systematically reviews the extraction techniques of P-EVs, their bioactive components, and the molecular mechanisms in repair of burn wounds. Moreover, P-EVs have certain advantages in biocompatibility and promoting cell proliferation for repair, providing innovative strategies for the treatment of burn wounds directly or as a complement for the existing therapeutics. However, their clinical application still faces challenges such as the lack of standardized extraction processes, unclear targeting mechanisms, and long-term safety assessment. Future research can combine multidisciplinary technologies to further optimize the extraction and targeting of P-EVs, promoting their translational application in regenerative medicine. This article aims to provide theoretical references for the basic research and clinical practice of P-EVs in repair of burn wounds and to envision their development prospects as a new treatment method for burn wounds.
Keywords: Burns, burn wounds healing, exosomes, plant-derived extracellular vesicles
Introduction
The skin, the largest organ in the human body, performs a wide range of vital functions.1 It serves as a protective barrier, shielding the body from bacteria, viruses, and other harmful substances.1 Beyond protection, the skin plays a key role in regulating body temperature and facilitates the perception of touch, temperature changes, and pain.1 Burns, which are injuries to the tissue, can result from exposure to heat (flames, hot liquids, or heated objects), chemicals, electrical currents, or radiation. These injuries often lead to tissue necrosis and functional impairment, making burns a significant and challenging category of clinical trauma.2 Among the various types of burns, thermal burns, which are caused by flames or wet sources and categorized according to the depth of tissue damage, are the most prevalent.3 Burn wounds healing is a complex physiological process that involves several stages, including inflammation, cell proliferation, neovascularization, extracellular matrix remodeling, and re-epithelialization.4,5 For optimal recovery, these processes must occur in precise coordination to restore the skin’s functionality.6,7 Burns are a global public health problem, which causing approximately 180, 000 deaths annually, according to the estimation of WHO.8 Critically, more than two-thirds of these fatalities occur in low- and middle-income countries (LMICs), with the highest burden concentrated in the African and South-East Asian regions.8 Additionally, non-fatal burns result in significant morbidity, including prolonged hospitalization, permanent disfigurement, and disability, which often lead to stigma and rejection.8 While direct medical costs vary by region, they universally impose substantial economic burdens.9,10 In South Africa, it is estimated that 26 million US dollars is spent each year on treating burns caused by kerosene (paraffin) stove accidents.8 Burn wounds healing, however, poses unique challenges. Patients often encounter complications, including insufficient autologous skin for grafts and the risk of immune rejection when using allografts. These barriers complicate the treatment process and contribute to the broader public health burden.11,12 Consequently, it is necessary to develop innovative and effective strategies to enhance the treatment of burn wounds.
For skin wounds, a variety of promising intervention strategies have been developed, including peptides and hydrogel.13–15 While bandages and gauze are effective in stopping bleeding, they are non-degradable, susceptible to infection, and incapable of maintaining the moist environment crucial for optimal healing.16,17 Hydrogels are commonly used in burn wounds care to absorb exudate and retain moisture. However, hydrogels lack targeted anti-inflammatory and antioxidant properties, which significantly limits their therapeutic potential.18 Peptides have the ability to fight bacteria and act as antioxidants, but they have poor cell penetration ability and are difficult to target diseased tissues.19–22
These restrictions have prompted people to explore new treatment strategies. In this case, P-EVs have attracted attention. Exosomes are membranous vesicles composed of lipid membranes with a phospholipid bilayer structure, which play a vital role in both intercellular and interspecies communication.23 Exosomes exhibit favorable characteristics for potential applications, persisting stably in various body fluids such as ascites, plasma, serum, urine, breast milk, and tears.24 They also demonstrate high biocompatibility, low immunogenicity, low cytotoxicity, and a relatively long circulation half-life.24 Researchers have found that P-EVs have a similar structure to exosomes.25 P-EVs contain a variety of bioactive substances, including proteins, nucleic acids, and lipids.26 These bioactive components can regulate key cellular processes of burn wounds healing such as proliferation, migration, differentiation, and angiogenesis.27–29 Furthermore, P-EVs can effectively deliver DNA and proteins, functional siRNAs, and various other therapeutic drugs to target cells without altering their biological activity.30 Thus, P-EVs are expected to become complementary therapies for clinical treatment of burn wounds. For example, P-EVs derived from bitter melon were found to enhance vascular endothelial cell synthesis and promote the recovery of damaged cells, highlighting their considerable potential in burn wounds repair.31 Therefore, P-EVs have broad application prospects in management of burn wounds. This article reviews the potential application of P-EVs in healing of burn wounds and analyzes its mechanism of action. Additionally, the challenges and prospects of P-EVs in clinical applications were discussed, which aimed to bridge the gap between basic research and practical application of P-EVs.
Physiological Characteristics and Repair Mechanisms of Burn Wounds
Physiological Characteristics of Burn Wounds
Burns are severe traumatic injuries that can cause extensive bodily harm and often result in long-term consequences, such as functional impairments and changes in physical appearance.32 For patients, the prolonged treatment process is frequently accompanied by intense pain and significant psychological distress.32 For their families, the high cost of medical care and the demands of long-term recovery can pose substantial financial challenges.32 Additionally, the expenses associated with burn wounds treatment and rehabilitation place a considerable strain on the broader healthcare system.32 Therefore, effective management of initial tissue damage and timely interventions during the healing process are critical components of early burn wounds treatment.33 Thermal injuries to the skin lead to a marked increase in vascular permeability, which is the primary cause of post-burn edema and hypovolemic shock.34 This phenomenon is marked by a significant increase in vascular wall permeability, leading to the leakage of large volumes of plasma-like fluid, extensive exudation from the burn wounds, and fluid accumulation in both tissue interstitial spaces and potential third-spacing compartments.34 Such fluid shifts affect not only the burn site but also surrounding uninjured tissues, causing pronounced edema.34 In severe cases, generalized edema can give rise to life-threatening complications, including cerebral edema due to increased intracranial pressure, elevated abdominal pressure, and fascial compartment syndrome.34 If left untreated, these conditions can lead to irreversible organ damage or prove fatal.34
Clinical Heterogeneity of Burns
The clinical manifestations and pathophysiological responses of burns show significant heterogeneity, which is mainly attributed to the causes, the systemic reactions, and the long-term outcomes.2 Different injury causes usually lead to different patterns of tissue damage. For instance, alkaline chemical injuries may cause liquefactive necrosis, while acidic burns may result in coagulative necrosis.2,35 Frostbite may involve cellular crystallization and ischemia-reperfusion injury.2,35 Severe burns can trigger a unique and complex systemic response, including a prolonged state of high metabolism lasting for several year and immune dysfunction which increases the patient’s susceptibility to infections36,37 The intensity and duration of this response vary from patient to patient, and the effects of burns are long-lasting.2 The recovery of patients will not only be involved in scar issues, but their mental health, quality of life, and the ability of return to work will also be continuously affected.38,39
Based on the severity, burns can be classified into different degrees that are superficial thickness (first degree), superficial partial thickness (second degree), deep partial thickness (second degree), full thickness (third degree) and fourth degree.2 The clinical characteristics and treatment methods vary according to the different degrees of burn (Table 1). 2 Grasping the complexities of these burn wounds is essential for creating effective treatment strategies and enhancing the healing process.33 Targeted treatments should be carried out based on the actual clinical situation. The reason why first-degree and second-degree burn wounds can heal spontaneously lies in the fact that they retain the source of cells for epithelial regeneration.40 These remaining epithelial cells form the basis for re-epithelialization and drive the wound to be repaired through the natural healing process.40,41 However, the healing dynamics of third-degree and fourth-degree full-thickness skin burn wounds have undergone fundamental changes.40 Due to the thermal injury, the entire layer of dermis and all its epidermal appendages were damaged, and the wounds completely lost its endogenous epithelial regeneration ability.40 The current gold standard treatment is early escharotomy and skin grafting, aiming to replace the necrotic tissue with autologous skin through transplantation, thereby achieving wound coverage and fundamentally altering the healing trajectory.40,42 Therefore, accurate evaluation of the depth of burn wounds involves determination of their inherent potential for healing essentially. This is of crucial importance for making the right decisions regarding treatment ranging from conservative therapy to surgical intervention.
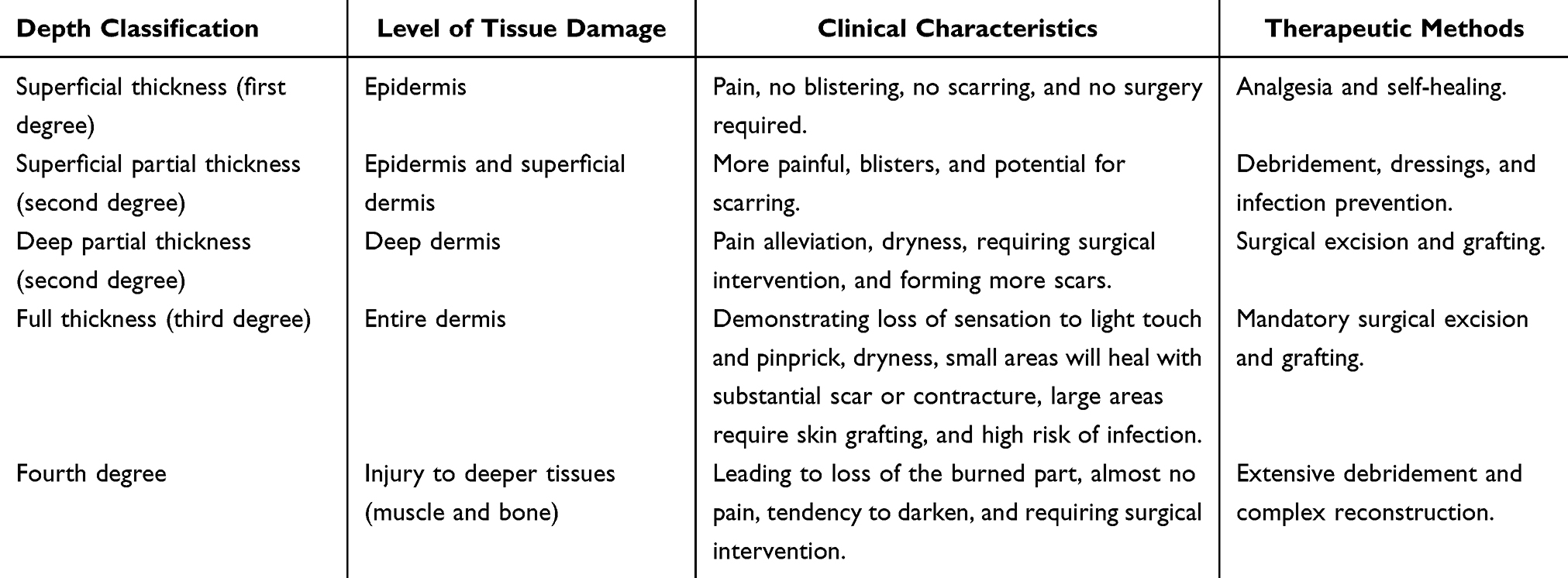 |
Table 1 The Clinical Characteristics and Therapeutic Methods of Different Degrees of Burns2 |
Mechanisms of Burn Wounds Repair
General Mechanisms of Natural Wound Healing
The natural wound healing stage is the foundation for understanding the healing process of burn wounds.2 Wound healing is a highly intricate repair process involving the precise coordination and integration of various biomolecules.43 This process encompasses the regulation of cellular activity, the reconstruction of the vascular system, and the reorganization of tissue structure.43 Together, these interdependent and closely interconnected mechanisms form a well-organized wound repair network, which is essential for ensuring complete and effective healing.43 Wound healing phases include hemostasis, inflammation, proliferation, and remodeling of the extracellular matrix (ECM) (Figure 1). 44 Far from being linear or independent, these stages are dynamic, continuous, and intricately linked, maintaining the integrity and progression of the wound repair process.45,46 Critical participants in this process include skin cells, immune cells, ECM components, cytokines, and chemokines, all of which play pivotal roles in modulating and orchestrating the intricate cascade of events.45,46
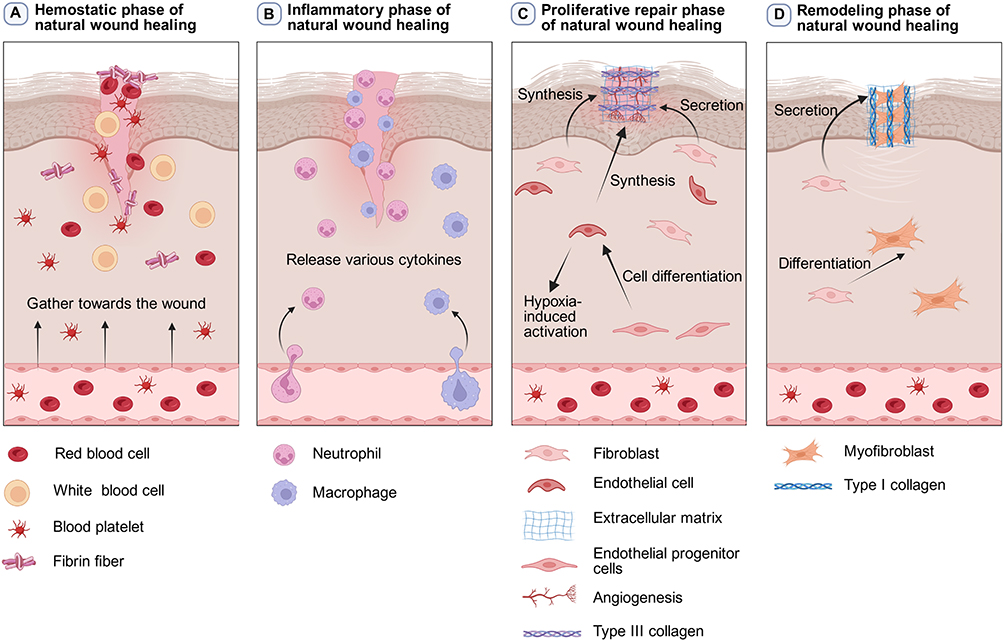 |
Figure 1 The general healing process of natural wound healing. Natural wound healing involves multiple stages, including hemostasis, inflammation, proliferation, and remodeling. (A) During the hemostasis period, platelets, red blood cells, white blood cells, and fibrin fibers among other blood components will gather at the injured area to form a blood clot, which will seal the wound and limit blood loss. (B) During the inflammatory period, neutrophils and macrophages gradually infiltrate the wound and release various cytokines. (C) During the proliferation and repair period, endothelial progenitor cells undergo cell differentiation to be-come vascular endothelial cells. These vascular endothelial cells will be activated in the hypoxic environment of burn wounds, thereby initiating angiogenesis. Activated fibroblasts and vascular synthetic cells secrete extracellular matrix and attach to the wound site. At this stage, the activated fibroblasts mainly secrete type III collagen, forming a temporary matrix. (D) During the remodeling period, fibroblasts differentiate into myofibroblasts which can promote contraction and closure of wounds, and the type III collagen will be remodeled into more the resilient type I collagen. |
The Special Pathological and Physiological Mechanisms of Burn Wounds Repair
Hemostatic Phase of Wound Healing
After a skin injury, the body swiftly activates hemostatic mechanisms to prevent further blood loss.47 In the early stages, blood components such as platelets, red blood cells, white blood cells, and fibrin fibers accumulate at the injury site, forming a blood clot that seals the wound and minimizes blood loss.47 Concurrently, platelets and damaged tissues release specific cytokines that guide fibrin to the wound surface, creating a cellular scaffold that supports immune cell activity and prepares them for the next phase of healing.47 Both normal wounds and burn wounds exhibit vasoconstriction responses.48 The early vasoconstriction in normal wounds is to achieve local hemostasis, but in severe burn wounds, the early vasoconstriction is part of the systemic stress response and shock, which can lead to reduced tissue perfusion.48,49
Inflammatory Phase of Burn Wounds Healing
The inflammatory phase is a critical stage in wound healing, serving to prevent infection and prepare the tissue for subsequent repair.50 Neutrophils act as the first responders during this phase, playing an essential role in eliminating debris and pathogens.50 Beyond their antimicrobial functions, neutrophils release various cytokines and chemokines that amplify the immune response by attracting additional immune cells to the wound site.50 Their population peaks within 1 to 2 days after injury.50 As neutrophil numbers diminish, macrophages begin to infiltrate the wound, with their activity peaking around day 5. By day 14, macrophage levels gradually decline and stabilize.50 Macrophages release various growth factors and cytokines, including platelet-derived growth factor (PDGF), transforming growth factor-β (TGF-β), fibroblast growth factor (FGF), and interleukin-6 (IL-6).51,52 They also produce pro-inflammatory mediators such as interleukin-1 beta (IL-1β) and tumor necrosis factor-alpha (TNF-α), which are crucial for maintaining the inflammatory phase.51,52 In addition to driving the immune response, macrophages facilitate the resolution of inflammation by clearing residual neutrophils and cellular debris.53,54 They also release cytokines and growth factors that stimulate the differentiation, proliferation, and migration of endothelial cells, keratinocytes, and fibroblasts.53,54 These processes are vital for neovascularization (the formation of new blood vessels), re-epithelialization (the regeneration of skin cells), and ECM deposition.53,54 Together, these activities not only contribute to wound closure but also lay the foundation for tissue remodeling during the proliferative phase.53,54
During the inflammatory phase, burn wounds share the basic biological process mediated by immune cells, which aims to eliminate damage and resist infection, with ordinary wounds.2,48 However, severe burn wounds amplify this localized, controllable response into a severe, prolonged and systemic pathological storm.2 Its characteristics include systemic capillary leakage and shock, an overactive unique mediator system (such as the kinin system), and contradictory systemic immunosuppression.2,48 This uncontrolled inflammatory response is not only the main cause of life-threatening issues during the acute phase of burn wounds, but also lays the groundwork for long-term complications such as delayed healing, extremely high metabolism and pathological scarring.2 Due to the excessive inflammation of the burn wounds that leads to systemic immunosuppression, and the damage caused by heat to the skin’s protective barrier, one of the main challenges faced by burn wounds is their susceptibility to infection.55 This damage compromises the barrier’s natural defense mechanisms, leading to a nutrient-rich exudate buildup.55 This microenvironment, in turn, becomes a fertile breeding ground for pathogenic microorganisms, which can quickly colonize and proliferate within the wound, dramatically increasing the risk of infection.55
Proliferative Repair Phase of Burn Wounds Healing
During the proliferation and repair phase, activated fibroblasts play a central role in the formation of granulation tissue and the synthesis of key ECM components.56,57 In this stage, fibroblasts mainly secrete type III collagen which transform into type I collagen during the ECM remodeling phase.58 This collagen serves as a structural scaffold, supporting neovascularization (the formation of new blood vessels) and the recruitment of inflammatory cells.56,57 Meanwhile, the hypoxic environment at the wound site stimulates the activation of vascular endothelial cells (EC), thereby triggering angiogenesis.59 Simultaneously, endothelial progenitor cells (EPCs) are mobilized, migrating from the bone marrow into the circulatory system.59 These progenitor cells differentiate into endothelial cells, which contribute to the formation of new blood vessels at the injury site.59 The activities of fibroblasts and capillarogenesis are intricately coordinated.60 Together, they promote the synthesis of ECM and granulation tissue, contributing to re-epithelialization.60 As the newly deposited ECM accumulates, it undergoes continuous degradation and remodeling by enzymes such as collagenase and elastase.61 This remodeling process persists until the ECM achieves equilibrium, establishing the structural foundation required for normal wound closure.61 During the proliferative phase, burn wounds and normal wounds share the basic biological processes mediated by fibroblasts and keratinocytes, which aim to close the wound through the formation of granulation tissue, angiogenesis, and re-epithelialization.2,48 However, the proliferative phase of severe burn wounds initiates in an extremely unfavorable internal environment: the continuous extremely high metabolic rate consumes a large amount of energy and substrates necessary for repair, while the uncontrolled inflammation disrupted the normal repair signals.62,63 Furthermore, deep burn wounds destroy the basal layer of the re-epithelializing epidermis, forcing the healing process to start from the deeper skin appendages with great difficulty.64 These factors collectively lead to the often-abnormal proliferation and repair of burn wounds, and the final outcome is highly dependent on the depth of the injury and is prone to developing into hypertrophic scars and contractures.65,66
Remodeling Phase of Burn Wound Healing
The remodeling phase represents the final stage of wound healing, following the proliferation phase.56,57,67 During this critical stage, fibroblasts transform into myofibroblasts, which possess contractile properties that facilitate wound contraction and closure.56,57,67 Simultaneously, the ECM undergoes structural alterations. Macrophages release various enzymes that further remodel the ECM, reshape the wound architecture, and support tissue maturation.56,57,67 Gradually, granulation tissue is replaced by connective tissue, allowing the wound to heal and ultimately close.57
While the human body demonstrates an innate capacity to repair epidermal burn wounds, severe burn wounds often lead to long-term complications, including impaired wound healing, metabolic imbalances, immune dysfunction, cardiovascular problems, and psychological trauma.2 These complications may result in chronic pain, diminished quality of life, reduced work productivity, and an elevated risk of mortality.39,40,68 Together, they impose lifelong physical and mental health challenges on patients.39,40,68 During the remodeling period, both burn wounds and ordinary wounds undergo collagen remodeling and scar maturation, including the conversion of collagen types and the remodeling of the matrix mediated by myofibroblasts.48 However, the remodeling process of burn wounds is severely pathologically deviated under the drive of continuous inflammation and extremely high metabolism.62,63 Its core feature is a significant imbalance in collagen metabolism, leading to the formation of hypertrophic scars.62,63 The clinical outcome of this kind of abnormal repair is devastating, often resulting in functional impairments such as scar contractures and long-term abnormal pigmentation.69,70 Therefore, the re-formation period of burn wounds is not merely a local repair endpoint, but rather a long and challenging stage that is closely linked to the patient’s overall pathophysiology and determines their long-term quality of life.
Other Factors Affecting Burn Wound Healing
The severity of a burn wound, the risk of infection, and individual patient characteristics are pivotal in determining the healing trajectory of burn wounds.71–74 Burn depth and severity are primary factors influencing the extent of tissue damage and the corresponding healing mechanisms.71 For example, superficial burns, such as first-degree burns, affect only the outermost layers of the skin, causing redness and mild discomfort.72 These burns generally require minimal treatment and heal relatively quickly.72 Conversely, deeper burn wounds, like third-degree burns, extend into the subcutaneous fat, muscle, and even bone in severe cases.72 Such injuries heal slowly, often necessitate surgical interventions like skin grafting, and frequently result in permanent scarring.72 Infection poses a critical threat to the healing of burn wounds, as pathogens can exacerbate the inflammatory response, prolong inflammation, and impede the healing process.73 Therefore, to mitigate infection and support optimal healing, maintaining a sterile environment and utilizing antimicrobial therapies are indispensable.
Reducing local inflammation and optimizing the wound microenvironment are vital strategies for promoting healing and minimizing scar formation.75,76 The normal restoration of skin barrier function depends on maintaining specific physiological conditions over a sustained period.77 Any disruption in this process can impair wound healing, resulting in chronic or poorly healing wounds.77 Chronic wounds are often characterized by dysfunctional and delayed cellular activity, triggering a pathological inflammatory response that culminates in persistent ulcers.77 Common underlying causes of chronic wounds include diabetes, ischemia, venous stasis, and pressure ulcers.77 Additionally, comorbidities and the overall health of the patient significantly influence the healing process. Conditions such as malnutrition and diabetes can obstruct the body’s natural healing mechanisms, leading to delays or incomplete recovery.74 For instance, deficiencies in essential micronutrients can hinder the healing process by impairing cellular repair, immune function, and tissue regeneration.74 In diabetic patients, wound healing is often impaired due to abnormal inflammatory responses, impaired granulation tissue formation, insufficient angiogenesis, and poor epithelialization.78 These challenges are partly from disruptions in epidermal intercellular junctions and disconnections between basal cells and basement membranes, which hinder the activation of normal skin repair.78
In short, due to the unique pathological and physiological characteristics of burn wounds, such as excessive inflammatory response, blocked cell proliferation, and abnormal tissue remodeling. Traditional gauze and bandages, although helpful in stopping bleeding, often fail to provide a moist environment and increase the risk of infection. Hydrogels can absorb exudate, but lack targeted anti-inflammatory or antioxidant functions. Although antimicrobial peptides have the potential for antibacterial and antioxidant properties, their poor cell permeability and limited targeting to tissues limit their efficacy. P-EVs carry a variety of bioactive molecules and can regulate key cellular processes, such as proliferation, migration, differentiation, and angiogenesis. P-EVs can also transport proteins and DNA to target cells without affecting their activity. Therefore, P-EVs are expected to become a supplementary strategy for burn treatment.
Biological Characterization of P-EVs
Composition and Origin of P-EVs
Exosomes are small membrane vesicles composed of lipid bilayers, usually between 30 ~150 nm in diameter.23,79 These vesicles are usually “saucer-like” or “cup-shaped” and are formed by the fusion of intracellular multivesicular bodies with the cell membrane, and then released into the extracellular space.23,79 Exosomes play an important role in intercellular and interspecies communication by delivering various bioactive molecules.23,79 Exosome membranes are rich in proteins, while vesicle contents typically include lipids, RNA (such as mRNA and microRNA), DNA, and other molecules capable of regulating a variety of physiological and pathological processes.23,79 Recently, researchers have found that P-EVs have a similar structure to exosomes.25,26 These vesicles are secreted by plant cells and are naturally non-toxic.80
P-EVs are mainly composed of lipid bilayers, and the biological active molecules such as proteins, nucleic acids, and secondary metabolites were encapsulated within or embedded in the lipid bilayers (Figure 2). 80 The lipid composition of membrane is mainly composed of phosphatidylcholine (PC), phosphatidylethanolamine (PE), phosphatidic acid (PA), and digalactosyldiacylglycerol (DGDG) et.al81,82 In terms of protein composition, P-EVs contain a variety of functional proteins. Annexins, heat shock proteins (HSPs), and aquaporins have been widely identified to play crucial roles in vesicle biogenesis, membrane fusion, and immune regulation processes.83,84 For instance, heat shock protein 70 (HSP70) can be successfully delivered to human cells as functional exogenous proteins.85 Tetraspanin 8 (TET8) and tetraspanin 9 (TET9), which are members of the tetraspanin family, are involved in the formation and secretion of P-EVs, the transmission of small RNAs, and the immune response of plant.86 Additionally, actions have been identified in various P-EVs, which suggests that P-EVs may play an important role in active processes related to the cell skeleton, such as organelle movement and vesicle transport.87 Among the components of nucleic acids in P-EVs, plant-specific microRNAs (miRNAs) play a crucial role in mediating their core functions.80 For instance, miR18a derived from grapefruit-derived extracellular vesicles (Gf-EVs) can inhibit the growth of colon tumors and liver metastasis by regulating the β-catenin and IFN-γ/IRF2 axis in macrophages.30 The gma-miR-159a of soybean-derived extracellular vesicles (Sb-EVs) can induce apoptosis in human colon cancer cells through the TCF7 inhibition mechanism.88,89 Ginger-derived extracellular vesicles (G-EVs) contain miRNAs such as csi-miR396e-5p and bdi-miR5179, which exhibit significant anti-inflammatory effects in human intestinal cells.90 In addition, messenger RNAs (mRNAs), long non-coding RNAs (lncRNAs), circular RNAs (circRNAs), siRNAs, and piRNAs were also identified in P-EVs.25,91–93 Moreover, P-EVs also contain a wide variety of plant-specific secondary metabolites, including flavonoids, phenolic acids, and alkaloids, etc.94,95 These compounds were reported to possess strong anti-inflammatory and antioxidant activities.94,95 They can work in synergy with nucleic acids and proteins to form the comprehensive pharmacological basis of PELNs for promoting wound healing and regulating the inflammatory microenvironment.94,95 The secondary metabolites are distributed within the membranes and cavities of P-EVs based on their chemical properties.96,97 Hydrophobic metabolites such as flavonoids and hydrophilic metabolites such as certain alkaloids tend to be embedded in the lipid bilayer, but hydrophilic metabolites such as phenolic acids and glycosides are dissolved in the aqueous phase within the cavity.96,97
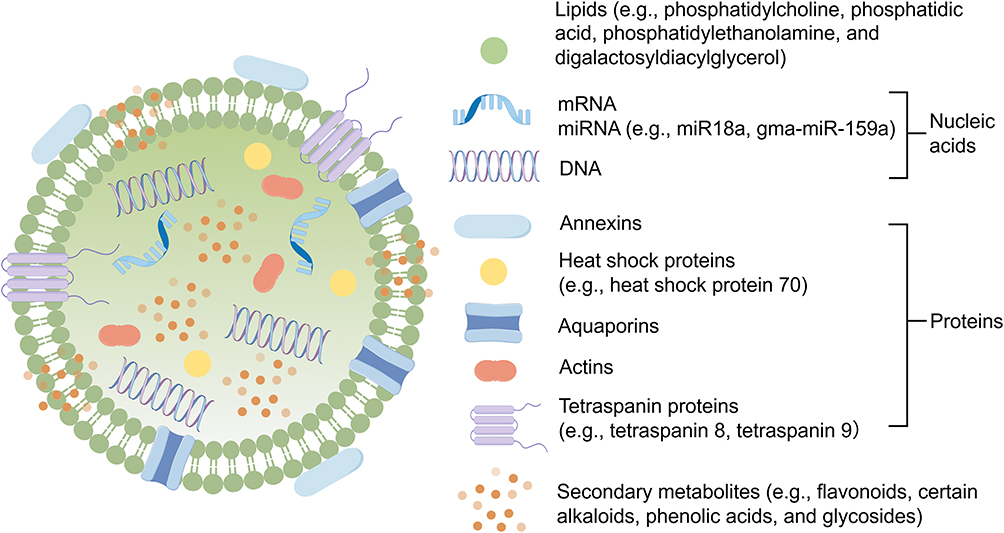 |
Figure 2 The generalized composition of P-EVs. P-EVs are mainly composed of lipid bilayers, and the bioactive substances such as proteins, nucleic acids, and secondary metabolites are encapsulated within or embedded in the lipid bilayers. Bioactive substances are present within P-EVs, embedded in the lipid bilayer, or adsorbed on their surface. |
Comparison of P-EVs with Mammalian Exosomes and Artificial Nanoparticles
Exosomes are a specific subtype of extracellular vesicles which are derived from endosomes of mammalian cells. They form in multivesicular bodies within the cell and are subsequently released into the extracellular space through the fusion of multivesicular bodies with the cell membrane.98 P-EVs is a broad term that encompasses all types of lipid-coated nanoparticles secreted from plant cells. Unlike the mammalian system, the exact biogenesis pathways of many P-EVs have not been fully elucidated.98,99 Therefore, in accordance with the principle of using operational terms for EVs with less studies as recommended by the MISEV guidelines, the general term “P-EVs” was used to refer to these vesicles. P-EVs are typically isolated from plant extracellular fluids or culture supernatants and have similar sizes (50 ~ 200 nanometers) and bilayer membrane structures to mammalian EVs.98,99 Artificial nanoparticles are synthetic nano-delivery vehicles, such as liposomes, polymer nanoparticles, and solid lipid nanoparticles, whose formation is driven by physical or chemical methods rather than biological processes.100
P-EVs are nano-sized vesicles in form, which are similar to mammalian exosomes, both functionally play significant roles in a non-cell-autonomous manner, and even participate in inter-species information exchange.101,102 P-EVs originate from plants, while exosomes were initially discovered and studied in mammalian cells.102 Furthermore, P-EVs can be isolated from a diverse array of plants, making them a readily available and potentially cost-effective resource.29 There are more clinical studies on exosomes, and some of them have entered the clinical trial stage, while the clinical research on P-EVs is relatively less.102 In addition, P-EVs pose no risk of being human pathogens and can be easily produced on a large scale.100 Mammalian exosomes may carry dangerous molecules, such as tumor substances and viruses.100 Large-scale production of exosomes relies on cell culture, which is costly and has limited output.100
Both P-EVs and artificial nanoparticles can be used as nano-scale carriers for drug delivery.100 P-EVs do not require chemical synthesis and possess natural lipid membrane structures with high stability. They are derived from food and exhibit low immunogenicity. However, their content is limited by plant types, thus requiring modification.100 However, artificial nanoparticles need to be synthesized artificially, which lead to their stability being affected by the preparation process, and lead to them having a risk of triggering immune reactions.100 Artificial nanoparticles can flexibly carry multiple drugs, but the preparation process is complex and the cost for large-scale production is high.100
Separation and Extraction Methods for P-EVs
Collectively, the isolation and extraction of P-EVs from plant material can be achieved using several techniques.
- Ultracentrifugation is a commonly method which gradually removes cellular debris and larger vesicles through stepwise centrifugation (Figure 3). 103,104 First, setting the centrifuge at 3000 × g for 20 minutes to remove dead cells and large cell fragments.105–107 Then, setting the centrifuge at 10000 × g for 40 minutes to remove cell debris and most of the fibers.105–107 Next, collecting the supernatant and filtrating through a 0.22 μm filter to remove pectin, small fibers, and other large molecular polymers.105–107 Finally, through an ultracentrifuge at 120, 000 × g for 90 minutes, collecting the precipitate as P-EVs.105–107 During this process, particles with different densities and sizes settle at distinctive rates under centrifugal force, ultimately yielding P-EVs.103,104 However, this method carries the risk of damaging P-EVs and may lead to reduced purity.103,104
- Sucrose density gradient centrifugation serves as an alternative method for the isolation and purification of P-EVs. Although this approach yields higher purity, it shares certain limitations with ultracentrifugation, including the potential for P-EVs damage and a time-intensive process.108
- Ultrafiltration and polymer precipitation: Ultrafiltration employs filter membranes with varying pore sizes to isolate P-EVs from particles of differing sizes. Polymer precipitation involves the use of polymers, such as polyethylene glycol, which interact with surface proteins on P-EVs to promote their precipitation.103 These techniques are widely utilized for P-EVs purification, but they tend to yield low recovery rates.103
- Immunoaffinity, volumetric exclusion chromatography, and microfluidics are emerging technologies that enable more precise separations and demonstrate relatively improved results.103,109 However, they continue to face limitations in efficiency and optimization.103,109
- Tangential flow filtration (TFF). The TFF system consists of two filters with different pore sizes, connected to a power pump that circulates the fluid continuously.110 This continuous circulation enhances the separation of specific size ranges through double filtration.110 Some researchers combined TFF with other exosome extraction methods and demonstrated this approach could increase the yield of exosomes effectively.111,112
 |
Figure 3 Isolation of P-EVs based on ultracentrifugation. First, various plants juice is obtained through extraction. Then, the juice is centrifuged in a low-speed to obtain the supernatant which contains small molecules such as proteins, DNA and extracellular vesicles, and the precipitate which contains cell debris, dead cells, most of the fibers, and cell nuclei will be discarded. Next, the supernatant is collected and filtered with a 0.22 μm filter membrane to remove pectin, small fibers, and other large molecular polymers. Finally, ultracentrifugation is performed on the supernatant, and the precipitate is collected and saved as P-EVs. |
In Summary, the choice of extraction and isolation methods for P-EVs largely depends on the unique characteristics of the sample and the intended application. Each method offers distinct advantages, but also comes with specific challenges, such as the risk of P-EVs damage, purity reduction, and time-consuming operation processes which can impede efficiency, particularly in large-scale applications. However, recent advancements in microfluidics, as well as electrical, acoustic, and force-based separation techniques, show great promise for enhancing both the efficiency and the delicacy of vesicle extraction processes.
Identification of P-EVs
The identification of P-EVs depends on the shape, size, and surface charge. Commonly, characterization is carried out using electron microscopes. Scanning electron microscopes (SEM) is suitable for observing the surface morphology and three-dimensional structure of P-EVs, but its resolution is lower than that of transmission electron microscopes (TEM)113,114 TEM has high resolution and can provide detailed morphological information of P-EVs.115,116 However, TEM assay needs to be operated in a vacuum environment and the samples need to undergo complex processes, which may damage the structure of P-EVs.115,116 Atomic force microscopes (AFM) can non-destructively capture and detect P-EVs, and display the morphology and structure of P-EVs clearly.117 However, the operation is rather complex, and strict requirements are imposed on the sample surface.117
The size and Zeta potential of P-EVs are important parameters that affect their biological functions such as intracellular uptake and drug delivery.118 The size and Zeta potential of P-EVs are typically characterized using Dynamic Light Scattering (DLS) and Nanoparticle Tracking Analysis (NTA). DLS has the advantages of simple operation and high efficiency but is prone to be affected by large particles during particle size analysis.119,120 Therefore, the measurement results in a single dispersion system are relatively accurate.119 NTA can capture and analyze the movement trajectories of individual particle, which can reduce the interference from large particles, improve measurement accuracy, and provide concentration data of P-EVs.119,120
Role and Advantages of P-EVs
P-EVs, which are similar to animal-derived exosomes, can be internalized by both plant and animal cells, thereby facilitating intercellular communication.85 Enriched with plant-specific active ingredients, P-EVs held significant promise as tools for disease treatment and drug delivery.29,121 Unlike vesicles derived from mammalian sources, P-EVs were inherently non-pathogenic and free from risks associated with infection or biological contamination due to their natural origin.122,123 This quality makes P-EVs a safer and more reliable alternative for applications in human therapeutic and drug delivery.122,123 P-EVs can deliver exogenous proteins to human cells efficiently, and the delivered proteins remain functionally active.85 The use of Citrus limon-derived EVs as carriers for doxorubicin (DOX) did not affect the pharmacotoxicity of DOX. Studies on HeLa and HEK293T cell lines showed that Citrus limon-derived EVs do not exhibit cytotoxic effects and can be considered as safe drug carriers.124 P-EVs can effectively transport therapeutic drugs across cellular barriers, which represents a significant advantage in drug delivery. Furthermore, pharmacological studies have underscored the anti-inflammatory, anti-tumor, and immunomodulatory properties of P-EVs. Gf-EVs could efficiently deliver biotinylated DNA and proteins, functional siRNAs, and various other therapeutic agents to target cells without altering their biological activity.29 G-EVs were mainly taken up by intestinal epithelial cells (IECs) and macrophages.107 Although G-EVs can be taken up by intestinal epithelial cells and macrophages, the precise molecular mechanism of their targeted delivery has not yet been clarified.107 The targeting ability of P-EVs may stem from their complex surface molecular composition. Taking G-EVs as an example, their membranes are rich in specific lipids such as phosphatidic acid and monogalactosyldiacylglycerol, as well as carry a small amount of membrane proteins such as aquaporins and chloride ion channels.107 A large amount of negatively charged phospholipids (such as phosphatidylcholine), which present on the surface of P-EVs, may interact through charges or act as ligands to bind to the receptors (such as integrin family members or other adhesion molecules) of target cells, thereby mediating specific recognition and uptake.125 In addition, G-EVs could alleviate acute colitis, enhance intestinal repair, and prevent chronic colitis and colitis-associated cancer (CAC).107 Thus, the medicinal properties of P-EVs, which are combined with high efficiency of cellular uptake, exceptional biocompatibility, and stability, highlights their immense potential in treatment of a wide range of diseases, such as inflammatory bowel disease, colitis-associated cancer, acute liver failure, and nonalcoholic fatty liver disease.29,107
P-EVs exhibit a range of distinctive biological functions, including immune modulation, cellular repair, and tissue regeneration.29,107 Their remarkable potential in wound healing and anti-inflammatory therapies stems from their capacity to enhance tissue repair and inflammation regulation, making them highly advantageous in such applications.29,107 Furthermore, P-EVs were notable for their high production yield, ease of extraction, cost-efficiency, eco-friendly manufacturing processes, and inherent safety.29,121 In conclusion, P-EVs represent an emerging and innovative class of nanomedicine. Their plant-based composition ensures safe, effective, and biocompatible therapeutic versatility, particularly in the domains of drug delivery, wound care, and anti-inflammatory treatments. The affordability, high yield, and natural origin of P-EVs make them unparalleled candidates for advancing nanomedicine and biopharmaceutical applications. Additionally, G-EVs could ameliorate dextran sulfate sodium (DSS)-induced murine colitis by upregulating expression of heme oxygenase-1 (HO-1) and inhibiting production of interleukin-1β (IL-1β) and tumor necrosis factor-α (TNF-α) in intestinal macrophages.126 And Brucea javanica-derived extracellular vesicles (BJ-EVs) could suppress tumor progression and metastasis by delivering 10 distinct miRNAs into 4T1 cells, which could significantly retard the growth and metastasis of 4T1 cells by regulating the PI3K/Akt/mTOR signaling pathway.127 Thus, P-EVs ability to facilitate cross-species cellular communication and deliver therapeutic outcomes suggests a promising potential for advancing phytotherapy.
Mechanisms of Burn Wound Repair by P-EVs
P-EVs exhibit a multimodal therapeutic profile in burn wound management, including enhanced cellular regeneration and proliferation, anti-inflammatory modulation, pro-angiogenic stimulation, regulation of redox homeostasis, and antimicrobial activity (Figure 4 and Table 2).
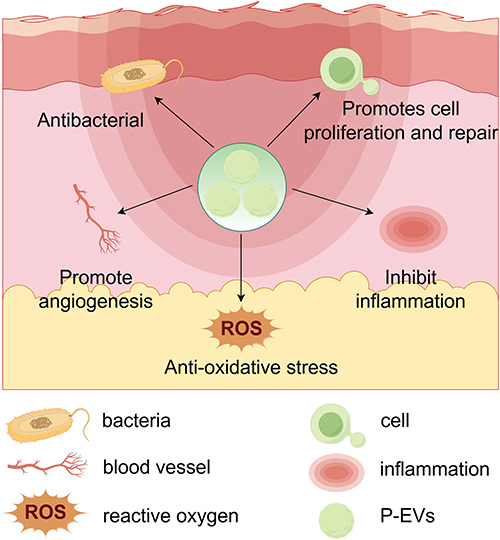 |
Figure 4 The regulatory mechanism of P-EVs in burn wounds healing process. P-EVs exhibit a multimodal therapeutic profile in burn wounds management, including enhanced cellular regeneration and proliferation, anti-inflammatory modulation, pro-angiogenic stimulation, regulation of redox homeostasis, and antimicrobial activity. |
 |
Table 2 Functions of Different Plant-Derived Extracellular Vesicles (P-EVs) |
Promoting Repair and Proliferation of Cells for Burn Wound Healing
Burn wounds typically progress through four stages: hemostasis, inflammatory response, tissue proliferation, and remodeling.44 The early inflammatory phase plays a critical role in determining the extent and development of scarring during later stages.50 As the largest organ in the body, the skin generally retains its function during the healing of minor wounds.1 However, severe trauma or an abnormal inflammatory response could result in excessive tissue repair, leading to the formation of scar tissue, which not only negatively impacts appearance but also may increase the risk of cancer in extreme cases.128,129 P-EVs have demonstrated promising potential in enhancing burn wound healing due to their diverse biological effects, including anti-inflammatory, pro-angiogenic, and antioxidant properties. Furthermore, P-EVs are rich in bioactive molecules such as proteins, lipids, and nucleic acids, which aid in cell repair, proliferation, and migration, making them highly effective for wound recovery.
For instance, P-EVs have been shown to enhance the proliferation of human dermal fibroblasts and keratinocytes, which are involved in tissue regeneration and repair.130,131 Their activation facilitates the formation and restoration of new tissue in burn wounds, significantly expediting the healing process.130,131 P-EVs of physalis peruviana fruit were demonstrated to have therapeutical bioactivities, including regeneration of dermis, induction of actin cytoskeletal rearrangement, stimulation of type I collagen biosynthesis, and downregulation of matrix metalloproteinase-1 (MMP-1) secretion.132 Although these functional studies provide experimental evidence for the role of P-EVs in promoting cell repair and proliferation, the underlying deeper signaling pathways still need to be further elucidated. P-EVs, which were isolated from cabbage (Cabex) and red cabbage (Rabex), showed potent cytoprotective properties, by inhibiting the activation of Caspase-3, they alleviate the apoptosis of human keratinocytes and dermal fibroblasts, which highlight their potential in burn wound healing.27
Additionally, P-EVs derived from Aloe vera peel showed excellent cytocompatibility with human skin cells, which enhanced the proliferation and migration of keratinocytes and fibroblasts.133 Wheat grass-derived extracellular vesicles (Wg-EVs) not only upregulated the gene expression of type I collagen (COL1A) significantly, which provided a crucial matrix environment for cell migration and tissue reconstruction, but also significantly promoted the proliferation and migration of human dermal fibroblasts, keratinocytes, and endothelial cells.134 Similarly, P-EVs derived from tomato (Solanum lycopersicum) were found to promote the migration of human keratinocytes and mouse fibroblasts, further highlighting their therapeutic potential for burn wound healing.28 Lemon-derived extracellular vesicles (L-EVs) could promote fibroblasts to migrate to the scratch area at a higher speed and a greater migration rate.135 Collectively, these findings suggest that P-EVs can accelerate the wound healing process by fostering cell repair, inhibiting cell apoptosis, enhancing migration, and promoting tissue regeneration.
Anti-Inflammatory Effects
Following a burn wound, the acute inflammatory response occurs locally at the wound site.50 Pro-inflammatory cytokines also have a positive effect on wound healing. For instance, pro-inflammatory cytokines initiate wound healing by recruiting immune cells and promoting proliferation, but excessive inflammation can impair healing and lead to scarring.136,137 Therefore, balanced regulation of these cytokines is crucial for optimal wound repair. However, an excessive inflammatory reaction not only delays the healing process but also may lead to secondary injuries.50 P-EVs exhibit remarkable anti-inflammatory properties by regulating immune cell activity and suppressing excessive inflammatory responses, which play a crucial role in the effective healing of burn wounds. Various P-EVs-based strategies have been proven to be effective in mitigating excessive inflammation, preventing infections, and accelerating tissue regeneration.
P-EVs derived from Aloe vera peel, through the key proteins they carry such as glutathione S-transferase P and phosphoenolpyruvate carboxykinase, effectively inhibited the secretion of TNF-α, IL-1β and IL-6 in macrophages and keratinocytes.138 This indicates that targeting these inflammatory regulatory proteins may be an effective strategy for suppressing excessive inflammatory responses at the wound site. P-EVs derived from cabbage exhibit dose-dependent inhibition of the expression of key pro-inflammatory mediators (such as IL-6, IL-1β and COX-2).27 Through comparing the differences in mRNA and protein levels, the study found that the inhibition of COX-2 by it might involve post-transcriptional regulatory mechanisms.27 In addition, garlic-derived extracellular vesicles (Ga-EVs) were shown to inhibit the NF-κB/NLRP3 signaling and block the CCR2/CCR5-mediated immune cell infiltration, which could form a powerful synergistic anti-inflammatory network.139 Although this result comes from a liver model, the NF-κB pathway has been regarded as a typical pro-inflammatory signaling pathway.140 These results suggest that P-EVs have the potential to regulate the excessive inflammatory response of burn wounds.
Similarly, another study assessed the anti-inflammatory effects of Allium tuberosum-derived extracellular vesicles (At-EVs) in RAW264.7 cells and mice with colitis. Results showed that At-EVs inhibit the activation of NF-κB, down-regulate the mRNA and protein levels of TNF-α, IL-1β, and IL-6, and up-regulate the protein level of the anti-inflammatory factor IL-10, thereby exerting an anti-inflammatory effect.141 G-EVs significantly reduced the mRNA and protein levels of TNF-α, IL-6, and IL-1β, and increased the expression of IL-10 and IL-22.107 Lemon-derived extracellular vesicles (L-EVs) possess powerful anti-inflammatory properties.142 The mechanism is not based on a single target, but rather involves the coordinated inhibition of the ERK and NF-κB two core inflammatory signaling pathways, exerting effects at multiple levels such as in macrophages and T lymphocytes.142 These results further highlight the potential of P-EVs in managing inflammation associated with burn wounds.
Promotion of Angiogenesis
Angiogenesis plays a crucial role in wound healing, particularly in burn injuries, where an enhanced blood supply is essential for tissue repair and regeneration.143 Research has demonstrated that P-EVs extracted from bitter melon contain key bioactive molecules, including vascular endothelial growth factor (VEGF) and CD31.31 These molecules can directly accelerate the healing of burn wounds by stimulating vascular endothelial synthesis and supporting the recovery of damaged cells.31 Notably, they had a significant effect in promoting the repair of deep second-degree burn wounds and held promise as a potential therapeutic agent for burn treatment.31 Gf-EVs were shown to enhance the tube-forming ability of endothelial cells, upregulate the expression of ECM and cytoskeletal components such as collagen, fibronectin, laminin and vimentin, and regulate the balance of migratory cytokines (such as RANTES, EGF, IGF-1) and inhibitory factors (such as TIMP-1).144 At the same time, they protect the function of endothelial cells through anti-oxidative stress, and jointly create a microenvironment conducive to angiogenesis.144 In addition, through in vitro tube formation experiments, it was confirmed that Wg-EVs have the potential to promote angiogenesis. However, the molecular mechanism by which wheat exosomes coordinate the proliferation, migration and lumen formation of endothelial cells remains a problem that needs to be addressed in future research.134 L-EVs significantly increased the expression of vascular biomarkers CD31 and VEGF, suggesting therapeutic potential for endothelial cell regeneration.135 These studies have demonstrated the ability of P-EVs to promote angiogenesis and have shown great potential in promoting wound healing in burn patients.
Antioxidant Effects
Burn wounds predominantly induce oxidative stress through excessive free radical generation, which can directly compromise tissue integrity and disrupt regenerative pathways.145 The natural antioxidant components in P-EVs, such as polyphenolic compounds and flavonoids, demonstrate significant antioxidant activity.146 These components help reduce the production of free radicals, mitigate oxidative damage, and support wound healing.146 P-EVs, which were isolated from the strawberry juice of Fragaria x ananassa (cv. Romina), were enriched with anthocyanins, folic acid, flavonols, and vitamin C, all of which exhibited strong antioxidant properties.146 Further experiments indicated that mesenchymal stem cells (MSCs) efficiently internalized strawberry-derived extracellular vesicles (S-EVs) without compromising cell viability.146 Moreover, pretreatment of MSCs with S-EVs effectively inhibited oxidative stress in a dose-dependent manner.146 Similarly, blueberry-derived extracellular vesicles (B-EVs) could activate the nuclear factor E2-related factor 2 (Nrf2) signaling pathway, thereby directly enhancing the antioxidant defense capacity of cells. This is manifested by a significant reduction in reactive oxygen species (ROS) levels, as well as the restoration of the activities of endogenous antioxidant enzymes such as superoxide dismutase (SOD) and glutathione (GSH).147 Furthermore, L-EVs are rich in various flavonoids, such as chuanxiong sinapine, vicarin-2, and stachyose glycoside, etc., which can activate the AhR/Nrf2 signaling pathway and upregulate the cell’s antioxidant defense system, thereby effectively eliminating ROS and promoting tissue repair.148 Through non-targeted metabolomics analysis, another study revealed that L-EVs were rich in natural antioxidants such as vitamin C, flavonoids, and citric acid, which can neutralize free radicals, alleviate oxidative stress, and protect cells from oxidative damage caused by high sugar environment.135 Aloe vera peel-derived extracellular vesicles (AV-EVs) can activate the core transcription factor Nrf2, thereby initiating the intrinsic antioxidant defense program of the cells.133 This is manifested by a dose-dependent increase manner in the activity of key antioxidant enzymes, such as superoxide dismutase (SOD) and catalase (CAT), and by significant upregulation of the expressions of Nrf2, HO-1, CAT, and SOD at the genetic level.133 Furthermore, AV-EVs are rich in superoxide dismutase (SOD), thioredoxin-dependent peroxidase oxidoreductase, and ascorbate peroxidase, etc.138 These enzymes can form a coordinated antioxidant defense network that can effectively remove excessive ROS and protect cells from oxidative damage.138 Notably, the source of P-EVs is important in the future treatment of burn wounds. Plants deriving from organic farming should be the first choice for P-EVs isolation, since those from intensive farming may deliver residual pesticides.149 The production of P-EVs from organic farming was 22% ~ 49% higher than that from intensive farming, and the total antioxidant capacity of P-EVs from organic fruits and vegetables was also 6% ~ 21% stronger than that from intensive fruits and vegetables.149 The P-EVs contain various antioxidant molecules, including enzymatic antioxidants such as superoxide dismutase (SOD) and catalase; non-enzymatic antioxidants such as glutathione (GSH), ascorbic acid (vitamin C), citric acid, and energy molecules such as ATP.149 These components can directly eliminate ROS and thereby reduce oxidative damage.149 The antioxidant components of P-EVs from the mixed fruits and vegetables were abundant and had a significant synergistic effect, which also demonstrated strong stability (resistant to decomposition) and high bioavailability.149,150 P-EVs exhibit antioxidant properties and are expected to create a favorable microenvironment for the repair of burned tissues.
Antimicrobial Effects
Burn wounds are highly susceptible to bacterial infection, often involving pathogens such as Escherichia coli, which can significantly delay the healing process. Therefore, it is of utmost importance to develop effective strategies to control infection of burn wounds73,151 Peppermint leaf-derived extracellular vesicles (Pl-EVs) could disrupt the integrity of bacterial cell membrane by carrying internal miRNAs, phospholipids, membrane proteins, and other bioactive molecules, which could increase the permeability of the cell membrane, cause the leakage of intracellular contents, and finally achieve antibacterial effects.152 Banana-derived extracellular vesicles (B-EVs) exhibit broad-spectrum antibacterial activity, with particularly significant inhibitory effects on Gram-positive bacteria (such as Staphylococcus aureus and Bacillus subtilis).153 B-EVs are rich in key components such as gallic acid, chlorogenic acid, and methyl gallate.153 These molecules can induce excessive accumulation of ROS within the bacteria, triggering oxidative stress, which subsequently disrupts the electron transfer chain, interferes with signal transduction, and causes DNA damage, ultimately leading to metabolic disorders and death of the bacteria.153 Moreover, these bioactive compounds may also inhibit key enzyme systems related to bacterial energy metabolism and cell wall synthesis.153
Prospects and Challenges of P-EVs in Burn Wounds
Prospects
P-EVs offer significant potential as an innovative approach for burn wound repair due to their remarkable biological properties, such as biocompatibility, anti-inflammatory effects, promotion of cell proliferation, and enhancement of angiogenesis. These characteristics make P-EVs promising as drugs that can accelerate the healing process of burn wounds. Composed of natural bioactive molecules such as proteins, lipids, and nucleic acids, P-EVs regulate critical aspects of wound healing, including cell migration, proliferation, and tissue regeneration. A significant advantage of P-EVs lies in their capacity to integrate with traditional therapies or biologics. For instance, using hydrogels enriched with P-EVs could extend the residence time of exosomes at the wound site, thereby prolonging their therapeutic efficacy.152,154,155 This synergistic capability enables P-EVs to enhance the effects of existing treatments, offering the potential to achieve better overall wound healing outcomes. Furthermore, the choice of plant sources is also very important. It is advisable to opt for organic crops. Some mixed plants also have synergistic effects.
Challenges
While P-EVs demonstrate significant potential for burn wound repair, several technical and regulatory challenges must be addressed before their widespread clinical application can be realized. (1) Lack of high-yield and standardized extraction methods. The current routine extraction and separation methods in the laboratory are time-consuming and have low yields, as well as may damage the integrity of the P-EVs due to mechanical damage. Therefore, it is necessary to develop a reliable, scalable, and standardized separation scheme for P-EVs. Integrating multiple separation technologies into a single platform may lead to a more efficient, scalable, and cost-effective separation system for P-EVs, while the purity, sterility, and retention of biological activity of the extracted substances should be taken into consideration. (2) Customization for characteristics of burn wounds. The multifunctional hydrogel, which is injectable, can detect wound conditions, and possesses antibacterial properties, has achieved excellent results in burn wounds healing.156–158 Regarding the clinical heterogeneity of burn wounds, P-EVs can also be loaded into multifunctional hydrogels, which can enable a synergistic effect, addresses the short half-life issue of P-EVs, and finally achieve a sustained-release effect.159 (3) Improved targeting and bioactivity. To maximize therapeutic efficacy, the targeting ability of P-EVs must be enhanced, ensuring their precise delivery to damaged cells within burn wounds. Exosomes interact with target cell receptors through their surface proteins such as integrins, transmembrane proteins (e.g., CD63, CD81), and lectins, thereby achieving organ-specific distribution.160 This provides a paradigm for analyzing the natural targeting mechanism of P-EVs. For instance, the lipids such as monogalactosyldiacylglycerol (MGDG) and phosphatidic acid, which are abundant in G-EVs, along with their specific membrane proteins (such as aquaporins), are likely to act as natural targeting sites. The initial targeting process is mediated through charge, hydrophobicity, or specific ligand-receptor recognition.107 Future research can draw on the methods of exosome proteomics to systematically identify the characteristic surface molecules of P-EVs from different sources, and verify their interaction with integrins that are highly expressed at the burn site. Combining the targeted logic of exosomes with the inherent advantages of P-EVs is expected to lead to the development of a new generation of “green” nanomedicines, which can not only be used for treating burn wounds, but also for precise treatment of various diseases such as inflammatory disorders, cancer, and tissue repair. Drawing on the strategies of nanomedicine, specific targeting molecules could be attached to the surface of P-EVs, thereby creating a drug delivery system that can precisely identify and act on specific wound cells.161 (4) Unclear Mechanism of Action. The precise mechanism through which P-EVs exert their therapeutic effects remains inadequately understood. Extensive and in-depth studies are necessary to elucidate the underlying processes, which will guide their optimized clinical application. Multi-omics methods, such as proteomics, lipidomic, and RNA sequencing, can be employed to comprehensively identify the substances which were carried by P-EVs from different plant sources. This will help identify the key active components that are responsible for their therapeutic effects. Subsequently, the specific roles of these components in processes such as angiogenesis and anti-inflammation should be verified. (5) Ethical and Regulatory Considerations. The therapeutic use of P-EVs faces significant ethical and regulatory challenges, particularly due to varied regulatory frameworks across different countries and regions. The US FDA regulates therapies based on extracellular vesicles as biological products under the Public Health Service Act, requiring an Investigational New Drug (IND) application and subsequent Biologics License Application (BLA).162 The classification depends on the intended use, mechanism of action, and composition of P-EVs product. If P-EVs are engineered primarily as delivery vehicles for a defined drug (e.g., siRNA), they would be reviewed as a “combination product”.162 In the European Union (EU), therapies based on extracellular vesicles are predominantly classified as Advanced Therapy Medicinal Products (ATMPs), if their contents are deemed to have a direct therapeutic effect on human physiology, the EVs would be subject to rigorous oversight by the Committee for Advanced Therapies (CAT).162 The lack of specialized guidelines for P-EVs in different regions pose difficulties in promoting the global clinical application. Developing standardized guidelines for P-EVs use in medical applications is indispensable to addressing these hurdles. (6) Evaluation of Long-Term Safety and immunogenicity. Comprehensive evaluations of the long-term safety and immunogenicity of P-EVs remain necessary to confirm their reliability for clinical use. Establishment of safe dose range and concentration is crucial for clinical application. Although short-term (7-day) oral administration of P-EVs in healthy mice did not cause pathological changes in the major organs of the mice, the long-term safety of this treatment still requires supplementary data from animal experiments.107 In addition, the immunogenicity of P-EVs should be paid due attention to. On one hand, their inherent immunomodulatory ability is one of the mechanisms that promote healing of burn wounds. On the other hand, as non-autologous substances, P-EVs may be recognized by the host immune system and trigger unnecessary immune responses.133 To comprehensively assess the long-term biological safety of P-EVs, future studies should include a systematic toxicological evaluation in accordance with Good Laboratory Practice for Nonclinical Laboratory Studies (GLP). (7) Selection of plant sources. Organic agriculture and non-genetically modified approaches attribute the therapeutic effects entirely to the inherent biological activity of P-EVs, rather than to the interfering variables introduced by genetic engineering. The P-EVs derived from non-genetically modified plants have their components mainly influenced by the variety and growth conditions, and are relatively easy to standardize. Moreover, large-scale cultivation will require a significant number of agricultural resources, which raises concerns about land use, water consumption, and potential impacts.163 It is necessary to conduct regular assessments to mitigate environmental impacts and develop sustainable and ethical procurement strategies. Moreover, when selecting the plant sources for P-EVs, plants that have been treated with pesticides should be avoided, preventing the occurrence of pesticide residues. Although P-EVs showed great potential in treatment of burn wounds, their clinical application still faces challenges. Currently, there are no clinical studies on the application of P-EVs in burn wounds. The clinical studies on exosomes derived from mesenchymal stem cells have achieved preliminary positive results for diabetic foot ulcers.164 This provides a reference clinical development model for the application of P-EVs in burn wounds treatment. Furthermore, P-EVs production that complies with the Good Manufacturing Practice (GMP) standards is the cornerstone of clinical translation. Thus, the current methods for preparing P-EVs should strictly ensure their sterility, purity, and biological activity.
Summary
Burn wounds pose a substantial medical challenge due to factors such as deep tissue damage, severe inflammation, and an elevated risk of infection. There are certain limitations to the single traditional treatment method. In this context, P-EVs show considerable promise in advancing burn wound healing. The P-EVs derived from different plant sources exhibit unique biological activities in key aspects such as anti-inflammatory, antioxidant, promotion of angiogenesis, antibacterial, and acceleration of re-epithelialization due to the different combinations of miRNAs, proteins, and metabolites. Although P-EVs have potential in the treatment of burn wounds, the therapeutic properties of P-EVs from different sources varies.27,28 For the complex pathology of burn wounds, it requires simultaneous control of inflammation, protection from oxidative stress, and promotion of tissue regeneration.2 In this context, lemons-derived extracellular vesicles (L-EVs) present certain advantages and become a particularly promising candidate. The significant advantage lies in that L-EVs can not only effectively inhibit pro-inflammatory cytokines and eliminate reactive oxygen species, but also significantly increase the expression of vascular biomarkers CD31 and VEGF, demonstrating the potential for promoting angiogenesis.135,142,148 Therefore, although the choice of plant sources may depend on the specific treatment goals, the current evidence establishes L-EVs as the leading candidate for burn treatment owing to their comprehensive and multi-mechanistic mode of action, which is highly compatible with the clinical needs for management of burn wound.
P-EVs inherent functional diversity makes P-EVs potentially a supplementary option for burn treatment. Nevertheless, while P-EVs hold significant promise, their clinical application remains in its infancy. The clinical application of P-EVs still faces numerous challenges, including unclear mechanisms, safety concerns, and regulatory issues. These problems must be addressed in order to fully realize its potential in burn treatment.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was supported by Sichuan Science and Technology Program (No. 2025ZNSFSC0676), Scientific Research Startup Fund of the Affiliated Hospital of Southwest Medical University (No. 25050), Science and Technology Strategic Cooperation Programs of Luzhou Municipal People’s Government and Southwest Medical University (No. 2023LZXNYDJ044), and Undergraduate Innovation Training Program of Southwest Medical University (No. 2025299).
Disclosure
The authors declare no conflict of interest.
References
1. Nourian Dehkordi A, Mirahmadi Babaheydari F, Chehelgerdi M, Raeisi Dehkordi S. Skin tissue engineering: wound healing based on stem-cell-based therapeutic strategies. Stem Cell Res Ther. 2019;10(1):111. doi:10.1186/s13287-019-1212-2
2. Jeschke MG, van Baar ME, Choudhry MA, Chung KK, Gibran NS, Logsetty S. Burn injury. Nat Rev Dis Primers. 2020;6(1):11. doi:10.1038/s41572-020-0145-5
3. Rodgers K, Jadhav SS. The application of mesenchymal stem cells to treat thermal and radiation burns. Adv Drug Deliv Rev. 2018;123:75–22. doi:10.1016/j.addr.2017.10.003
4. Valente DS. New horizons in regenerative medicine: photostimulation of adipose tissue. Regen Med Rep. 2025;2(2):72. doi:10.4103/REGENMED.REGENMED-D-25-00001
5. Yang A, Yang A, Shu J, Wu X, Zhu Z. Extracellular vesicles combined with hyperbaric oxygen therapy as new strategies for the treatment of traumatic brain injury: a narrative review. Adv Technol Neurosci. 2025;2(4):187. doi:10.4103/ATN.ATN-D-25-00012
6. Lee DE, Ayoub N, Agrawal DK. Mesenchymal stem cells and cutaneous wound healing: novel methods to increase cell delivery and therapeutic efficacy. Stem Cell Res Ther. 2016;7(1):37. doi:10.1186/s13287-016-0303-6
7. Chen JS, Wong VW, Gurtner GC. Therapeutic potential of bone marrow-derived mesenchymal stem cells for cutaneous wound healing. Front Immun. 2012;3. doi:10.3389/fimmu.2012.00192
8. World Health Organization. Search results; 2023. Available from: https://www.who.int/home/search-results.
9. Sengoelge M, El-Khatib Z, Laflamme L. The global burden of child burn injuries in light of country level economic development and income inequality. Prevent Med Rep. 2017;6:115–120. doi:10.1016/j.pmedr.2017.02.024
10. Chinweuba AU, Chinweuba IS, Diorgu FC, et al. Economic burden of moderate to severe burns and its association with health-related quality of life of Nigerian women. BMC Women’s Health. 2021;21(1). doi:10.1186/s12905-021-01232-5
11. Liu L, Yu Y, Hou Y, et al. Human umbilical cord mesenchymal stem cells transplantation promotes cutaneous wound healing of severe burned rats. PLoS One. 2014;9(2):e88348. doi:10.1371/journal.pone.0088348
12. watt SM, Pleat JM. Stem cells, niches and scaffolds: applications to burns and wound care. Adv Drug Deliv Rev. 2018;123:82–106. doi:10.1016/j.addr.2017.10.012
13. Jia Q, Fu Z, Li Y, et al. Hydrogel loaded with peptide-containing nanocomplexes: symphonic cooperation of photothermal antimicrobial nanoparticles and prohealing peptides for the treatment of infected wounds. ACS Appl Mater Interfaces. 2024;16(11):13422–13438. doi:10.1021/acsami.3c16061
14. Ru ZQ, Wu YT, Yang CY, et al. Ultra-short cyclic peptide Cy RL-QN15 acts as a TLR4 antagonist to expedite oral ulcer healing. Zool Res. 2025;46(5):1187–1202. doi:10.24272/j.issn.2095-8137.2025.211
15. Wang L, Fu Z, Su Y, et al. Cyclic heptapeptide FZ1 acts as an integrin αvβ3 agonist to facilitate diabetic skin wound healing by enhancing angiogenesis. J Med Chem. 2025;68(18):19503–19520. doi:10.1021/acs.jmedchem.5c01734
16. Boyce ST, Lalley AL. Tissue engineering of skin and regenerative medicine for wound care. Burns Trauma. 2018;6. doi:10.1186/s41038-017-0103-y/5680409
17. Schiavon M, Francescon M, Drigo D, et al. The use of integra dermal regeneration template versus flaps for reconstruction of full-thickness scalp defects involving the calvaria: a cost–benefit Analysis. Aesth Plast Surg. 2016;40(6):901–907. doi:10.1007/s00266-016-0703-0
18. Shi C, Wang C, Liu H, et al. Selection of appropriate wound dressing for various wounds. Front Bioeng Biotechnol. 2020;8. doi:10.3389/fbioe.2020.00182
19. Li C, Xiong Y, Fu Z, et al. The direct binding of bioactive peptide Andersonin-W1 to TLR4 expedites the healing of diabetic skin wounds. Cell Mol Biol Lett. 2024;29(1):24. doi:10.1186/s11658-024-00542-4
20. Al Musaimi O, Lombardi L, Williams DR, Albericio F. Strategies for Improving Peptide Stability and Delivery. Pharmaceuticals. 2022;15(10):1283. doi:10.3390/ph15101283
21. Wu YT, Ru ZQ, Peng Y, et al. Peptide Cy RL-QN15 accelerates hair regeneration in diabetic mice by binding to the Frizzled-7 receptor. Zool Res. 2024;45(6):1287–1299. doi:10.24272/j.issn.2095-8137.2024.134
22. Li Y, Jia Q, Liu N, et al. Peptide RL-QN15 regulates functions of epidermal stem cells to accelerate skin wound regeneration via the FZD8/β-catenin axis. Exploration. 2025:20240090. doi:10.1002/EXP.20240090
23. Zhou Y, Tian T, Zhu Y, et al. Exosomes transfer among different species cells and mediating miRNAs delivery. J Cell Biochem. 2017;118(12). doi:10.1002/jcb.26077
24. Hao J, Ye Y, Zhang G, Shen H, Li J, Chen G. Mechanisms of nitric oxide in spinal cord injury. Med Gas Res. 2024;14(4):192. doi:10.4103/mgr.MEDGASRES-D-23-00006
25. Xiao J, Feng S, Wang X, et al. Identification of exosome-like nanoparticle-derived microRNAs from 11 edible fruits and vegetables. PeerJ. 2018;6:e5186. doi:10.7717/peerj.5186
26. Wu H, Shi M, Meng L, et al. Plant-derived extracellular vesicles as a novel tumor-targeting delivery system for cancer treatment. Front Cell Dev Biol. 2025;13:1589550. doi:10.3389/fcell.2025.1589550
27. You JY, Kang SJ, Rhee WJ. Isolation of cabbage exosome-like nanovesicles and investigation of their biological activities in human cells. Bioact Mater. 2021;6(12):4321–4332. doi:10.1016/j.bioactmat.2021.04.023
28. Daniello V, De Leo V, Lasalvia M, et al. Solanum lycopersicum (Tomato)-derived nanovesicles accelerate wound healing by eliciting the migration of keratinocytes and fibroblasts. Int J Mol Sci. 2024;25(5):2452. doi:10.3390/ijms25052452
29. Wang Q, Zhuang X, Mu J, et al. Delivery of therapeutic agents by nanoparticles made of grapefruit-derived lipids. Nat Commun. 2013;4(1):1867. doi:10.1038/ncomms2886
30. Teng Y, Mu J, Hu X, et al. Grapefruit-derived nanovectors deliver miR-18a for treatment of liver metastasis of colon cancer by induction of M1 macrophages. Oncotarget. 2016;7(18):25683–25697. doi:10.18632/oncotarget.8361
31. Liu Y, Gao Q, Zhao Y, et al. Effect of Momordica charantia exosome combined with mupirocin ointment on the wound healing of deep second degree scald. J Xuzhou Med Univ. 2023;43(8):584–589. [Chinese]. doi:10.3969/j.issn.2096-3882.2023.08.007
32. Vigiola Cruz M, Carney BC, Luker JN, et al. Plasma ameliorates endothelial dysfunction in burn injury. J Surg Res. 2019;233:459–466. doi:10.1016/j.jss.2018.08.027
33. Cartotto R, Callum J. A review on the use of plasma during acute burn resuscitation. J Burn Care Res. 2020;41(2):433–440. doi:10.1093/jbcr/irz184
34. D’Abbondanza JA, Shahrokhi S. Burn infection and burn sepsis. Surg Infect. 2021;22(1):58–64. doi:10.1089/sur.2020.102
35. Lee RC. Injury by electrical forces: pathophysiology, manifestations, and therapy. Curr Probl Surg. 1997;34(9):677–764. doi:10.1016/s0011-3840(97)80007-x
36. Jeschke MG, Gauglitz GG, Kulp GA, et al. Long-term persistance of the pathophysiologic response to severe burn injury. PLoS One. 2011;6(7):e21245. doi:10.1371/journal.pone.0021245
37. Jeschke MG, Chinkes DL, Finnerty CC, et al. Pathophysiologic response to severe burn injury. Ann Surg. 2008;248(3):387–401. doi:10.1097/SLA.0b013e3181856241
38. Burnett E, Gawaziuk JP, Shek K, Logsetty S. Healthcare resource utilization associated with burns and necrotizing fasciitis: a single-center comparative analysis. J Burn Care Res. 2017;38(6):e886–e891. doi:10.1097/BCR.0000000000000513
39. Logsetty S, Shamlou A, Gawaziuk JP, et al. Mental health outcomes of burn: a longitudinal population-based study of adults hospitalized for burns. Burns. 2016;42(4):738–744. doi:10.1016/j.burns.2016.03.006
40. Mason SA, Nathens AB, Byrne JP, et al. Increased rate of long-term mortality among burn survivors: a population-based matched cohort study. Ann Surg. 2019;269(6):1192–1199. doi:10.1097/SLA.0000000000002722
41. Rowan MP, Cancio LC, Elster EA, et al. Burn wound healing and treatment: review and advancements. Crit Care. 2015;19:243. doi:10.1186/s13054-015-0961-2
42. Herndon DN, Barrow RE, Rutan RL, Rutan TC, Desai MH, Abston S. A comparison of conservative versus early excision. Therapies in severely burned patients. Ann Surg. 1989;209(5):547–552;discussion552–553. doi:10.1097/00000658-198905000-00006
43. Amini-Nik S, Yousuf Y, Jeschke MG. Scar management in burn injuries using drug delivery and molecular signaling: current treatments and future directions. Adv Drug Deliv Rev. 2018;123:135–154. doi:10.1016/j.addr.2017.07.017
44. Piperigkou Z, Götte M, Theocharis AD, Karamanos NK. Insights into the key roles of epigenetics in matrix macromolecules-associated wound healing. Adv Drug Deliv Rev. 2018;129:16–36. doi:10.1016/j.addr.2017.10.008
45. Eming SA, Hammerschmidt M, Krieg T, Roers A. Interrelation of immunity and tissue repair or regeneration. Semin Cell Dev Biol. 2009;20(5):517–527. doi:10.1016/j.semcdb.2009.04.009
46. Eming SA, Krieg T, Davidson JM. Inflammation in wound repair: molecular and cellular mechanisms. J Invest Dermatol. 2007;127(3):514–525. doi:10.1038/sj.jid.5700701
47. Rodero MP, Licata F, Poupel L, et al. In vivo imaging reveals a pioneer wave of monocyte recruitment into mouse skin wounds. PLoS One. 2014;9(10):e108212. doi:10.1371/journal.pone.0108212
48. Abazari M, Ghaffari A, Rashidzadeh H, Badeleh SM, Maleki Y. A systematic review on classification, identification, and healing process of burn wound healing. Int J Low Extrem Wounds. 2022;21(1):18–30. doi:10.1177/1534734620924857
49. Nielson CB, Duethman NC, Howard JM, Moncure M, Wood JG. Burns: pathophysiology of systemic complications and current management. J Burn Care Res. 2017;38(1):e469–e481. doi:10.1097/BCR.0000000000000355
50. Rodero MP, Hodgson SS, Hollier B, Combadiere C, Khosrotehrani K. Reduced Il17a expression distinguishes a Ly6c lo MHCII hi macrophage population promoting wound healing. J Invest Dermatol. 2013;133(3):783–792. doi:10.1038/jid.2012.368
51. Weinheimer-Haus EM, Mirza RE, Koh TJ. Nod-like receptor protein-3 inflammasome plays an important role during early stages of wound healing. PLoS One. 2015;10(3):e0119106. doi:10.1371/journal.pone.0119106
52. Moreira E, Burghi G, Manzanares W. Update on metabolism and nutrition therapy in critically ill burned patients. Medicina Intensiva. 2018;42(5):306–316.
53. Delavary BM, Van Der Veer WM, Van Egmond M, Niessen FB, Beelen RHJ. Macrophages in skin injury and repair. Immunobiology. 2011;216(7):753–762. doi:10.1016/j.imbio.2011.01.001
54. Rodero MP, Khosrotehrani K. Skin wound healing modulation by macrophages. Int J Clin Exp Pathol. 2010;3(7):643–653.
55. Singh NP, Rani M, Gupta K, Sagar T, Kaur IR. Changing trends in antimicrobial susceptibility pattern of bacterial isolates in a burn unit. Burns. 2017;43(5):1083–1087. doi:10.1016/j.burns.2017.01.016
56. Rodrigues M, Kosaric N, Bonham CA, Gurtner GC. Wound healing: a cellular perspective. Physiol Rev. 2019;99(1):665–706. doi:10.1152/physrev.00067.2017
57. Wang PH, Huang BS, Horng HC, Yeh CC, Chen YJ. Wound healing. J Chin Med Assoc. 2018;81(2):94–101. doi:10.1016/j.jcma.2017.11.002
58. Reinke JM, Sorg H. Wound repair and regeneration. Eur Surg Res. 2012;49(1):35–43. doi:10.1159/000339613
59. Balaji S, King A, Crombleholme TM, Keswani SG. The role of endothelial progenitor cells in postnatal vasculogenesis: implications for therapeutic neovascularization and wound healing. Adv Wound Care. 2013;2(6):283–295. doi:10.1089/wound.2012.0398
60. Greaves NS, Ashcroft KJ, Baguneid M, Bayat A. Current understanding of molecular and cellular mechanisms in fibroplasia and angiogenesis during acute wound healing. J Dermatological Sci. 2013;72(3):206–217.
61. O’Kane S, Ferguson MW. Transforming growth factor beta s and wound healing. Int J Biochem Cell Biol. 1997;29(1):63–78. doi:10.1016/s1357-2725(96)00120-3
62. Finnerty CC, Jeschke MG, Branski LK, Barret JP, Dziewulski P, Herndon DN. Hypertrophic scarring: the greatest unmet challenge after burn injury. Lancet. 2016;388(10052):1427–1436. doi:10.1016/S0140-6736(16)31406-4
63. Gore DC, Chinkes DL, Hart DW, Wolf SE, Herndon DN, Sanford AP. Hyperglycemia exacerbates muscle protein catabolism in burn-injured patients. Crit Care Med. 2002;30(11):2438–2442. doi:10.1097/00003246-200211000-00006
64. Hettiaratchy S, Dziewulski P. ABC of burns: pathophysiology and types of burns. BMJ. 2004;328(7453):1427–1429. doi:10.1136/bmj.328.7453.1427
65. Tiwari VK. Burn wound: how it differs from other wounds? Indian J Plast Surg. 2012;45(2):364–373. doi:10.4103/0970-0358.101319
66. Atiyeh BS, Gunn SW, Hayek SN. State of the Art in Burn Treatment. World J Surg. 2005;29(2):131–148. doi:10.1007/s00268-004-1082-2
67. Sorg H, Tilkorn DJ, Hager S, Hauser J, Mirastschijski U. Skin wound healing: an update on the current knowledge and concepts. Eur Surg Res. 2017;58(1–2):81–94. doi:10.1159/000454919
68. Stone J, Gawaziuk JP, Khan S, et al. Outcomes in adult survivors of childhood burn injuries as compared with matched controls. J Burn Care Res. 2016;37(2):e166–e173. doi:10.1097/BCR.0000000000000323
69. Jeschke MG. Postburn hypermetabolism: past, present, and future. J Burn Care Res. 2016;37(2):86–96. doi:10.1097/BCR.0000000000000265
70. Jeschke MG, Gauglitz GG, Song J, et al. Calcium and ER stress mediate hepatic apoptosis after burn injury. J Cell Mol Med. 2009;13(8B):1857–1865. doi:10.1111/j.1582-4934.2009.00644.x
71. Palackic A, Jay JW, Duggan RP, et al. Therapeutic strategies to reduce burn wound conversion. Medicina. 2022;58(7):922. doi:10.3390/medicina58070922
72. Broussard KC, Powers JG. Wound dressings: selecting the most appropriate type. Am J Clin Dermatol. 2013;14(6):449–459. doi:10.1007/s40257-013-0046-4
73. Luterman A, Dacso CC, Curreri PW. Infections in burn patients. Am J Med. 1986;81(1):45–52.
74. Kassym L, Zhetmekova Z, Kussainova A, et al. Pressure ulcers and nutrients: from established evidence to gaps in knowledge. CMC. 2024;32. doi:10.2174/0109298673322825241018174928
75. Jeschke MG, Wood FM, Middelkoop E, et al. Scars. Nat Rev Dis Primers. 2023;9(1). doi:10.1038/s41572-023-00474-x
76. Shi L. Study on the Mechanism of Metformin to Improve Deep Partial Thickness Burn Wound Healing by Regulating Microenvironmental Macrophages and Fibroblasts [dissertation]. Nanchang University; 2024. Chinese.
77. Guo S, DiPietro LA. Factors affecting wound healing. J Dent Res. 2010;89(3):219–229. doi:10.1177/0022034509359125
78. Yates CC, Hebda P, Wells A. Skin wound healing and scarring: fetal wounds and regenerative restitution. Birth Defects Res Pt C. 2012;96(4):325–333. doi:10.1002/bdrc.21024
79. Aa F, Nn D, Mz Q, et al. Exosome biogenesis, bioactivities and functions as new delivery systems of natural compounds. Biotechnol Adv. 2018;36(1). doi:10.1016/j.biotechadv.2017.12.010
80. Tembo KM, Wang X, Bolideei M, et al. Exploring the bioactivity of microRNAs originated from plant-derived exosome-like nanoparticles (PELNs): current perspectives. J Nanobiotechnol. 2025;23(1):563. doi:10.1186/s12951-025-03602-9
81. Zhu H, He W. Ginger: a representative material of herb-derived exosome-like nanoparticles. Front Nutr. 2023;10:1223349. doi:10.3389/fnut.2023.1223349
82. Subha D, Harshnii K, Madhikiruba KG, Nandhini M, Tamilselvi KS. Plant derived exosome- like nanovesicles: an updated overview. Plant Nano Biology. 2023;3:100022. doi:10.1016/j.plana.2022.100022
83. Rutter BD, Innes RW. Extracellular vesicles isolated from the leaf apoplast carry stress-response proteins. Plant Physiol. 2017;173(1):728–741. doi:10.1104/pp.16.01253
84. Prado N, de Alché JD, Casado-Vela J, et al. Nanovesicles are secreted during pollen germination and pollen tube growth: a possible role in fertilization. Mol Plant. 2014;7(3):573–577. doi:10.1093/mp/sst153
85. Garaeva L, Kamyshinsky R, Kil Y, et al. Delivery of functional exogenous proteins by plant-derived vesicles to human cells in vitro. Sci Rep. 2021;11(1):6489. doi:10.1038/s41598-021-85833-y
86. Cai Q, Qiao L, Wang M, et al. Plants send small RNAs in extracellular vesicles to fungal pathogen to silence virulence genes. Science. 2018;360(6393):1126–1129. doi:10.1126/science.aar4142
87. Woith E, Fuhrmann G, Melzig MF. Extracellular vesicles—connecting kingdoms. IJMS. 2019;20(22):5695. doi:10.3390/ijms20225695
88. Liu J, Wang F, Weng Z, et al. Soybean-derived miRNAs specifically inhibit proliferation and stimulate apoptosis of human colonic Caco-2 cancer cells but not normal mucosal cells in culture. Genomics. 2020;112(5):2949–2958. doi:10.1016/j.ygeno.2020.05.011
89. Liu J, Wang F, Song H, et al. Soybean-derived gma-miR159a alleviates colon tumorigenesis by suppressing TCF7/MYC in mice. J Nutr Biochem. 2021;92:108627. doi:10.1016/j.jnutbio.2021.108627
90. Yin L, Yan L, Yu Q, et al. Characterization of the microRNA profile of ginger exosome-like nanoparticles and their anti-inflammatory effects in intestinal Caco-2 cells. J Agric Food Chem. 2022;70(15):4725–4734. doi:10.1021/acs.jafc.1c07306
91. Wang Y, Wang J, Ma J, Zhou Y, Lu R. Focusing on future applications and current challenges of plant derived extracellular vesicles. Pharmaceuticals. 2022;15(6):708. doi:10.3390/ph15060708
92. Sundaram GM. Dietary non-coding RNAs from plants: fairy tale or treasure? Noncoding RNA Res. 2019;4(2):63–68. doi:10.1016/j.ncrna.2019.02.002
93. Z P, L S, C M. Characterization and function of circular RNAs in plants. Front Mol Biosci. 2020;7. doi:10.3389/fmolb.2020.00091
94. Whitford W, Guterstam P. Exosome manufacturing status. Future Med Chem. 2019;11(10):1225–1236. doi:10.4155/fmc-2018-0417
95. Liu DK, Xu CC, Zhang L, et al. Evaluation of bioactive components and antioxidant capacity of four celery (Apium graveolens L.) leaves and petioles. Int J Food Prop. 2020;23(1):1097–1109. doi:10.1080/10942912.2020.1778027
96. Urzì O, Raimondo S, Alessandro R. Extracellular vesicles from plants: current knowledge and open questions. IJMS. 2021;22(10):5366. doi:10.3390/ijms22105366
97. Alfieri M, Leone A, Ambrosone A. Plant-derived nano and microvesicles for human health and therapeutic potential in nanomedicine. Pharmaceutics. 2021;13(4):498. doi:10.3390/pharmaceutics13040498
98. Théry C, Witwer KW, Aikawa E, et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): a position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J of Extracellular Vesicle. 2018;7(1). doi:10.1080/20013078.2018.1535750
99. Dad HA, Gu TW, Zhu AQ, Huang LQ, Peng LH. Plant exosome-like nanovesicles: emerging therapeutics and drug delivery nanoplatforms. Mol Ther. 2021;29(1):13–31. doi:10.1016/j.ymthe.2020.11.030
100. Logozzi M, Di Raimo R, Mizzoni D, Fais S. The potentiality of plant-derived nanovesicles in human health—a comparison with human exosomes and artificial nanoparticles. Int J Mol Sci. 2022;23(9):4919. doi:10.3390/ijms23094919
101. Zhang M, Viennois E, Xu C, Merlin D. Plant derived edible nanoparticles as a new therapeutic approach against diseases. Tissue Barriers. 2016;4(2):e1134415. doi:10.1080/21688370.2015.1134415
102. Cong M, Tan S, Li S, et al. Technology insight: plant-derived vesicles—how far from the clinical biotherapeutics and therapeutic drug carriers? Adv Drug Delivery Rev. 2022;182:114108. doi:10.1016/j.addr.2021.114108
103. Yang D, Zhang W, Zhang H, et al. Progress, opportunity, and perspective on exosome isolation - efforts for efficient exosome-based theranostics. Theranostics. 2020;10(8):3684–3707. doi:10.7150/thno.41580
104. Xu XH, Yuan TJ, Dad HA, et al. Plant exosomes as novel nanoplatforms for microRNA transfer stimulate neural differentiation of stem cells in vitro and in vivo. Nano Lett. 2021;21(19):8151–8159. doi:10.1021/acs.nanolett.1c02530
105. Momen-Heravi F. Isolation of extracellular vesicles by ultracentrifugation. In: Kuo WP, Jia S, editors. Extracellular Vesicles: Methods and Protocols. Springer; 2017:25–32. doi:10.1007/978-1-4939-7253-1_3
106. Posch A, ed.. Proteomic Profiling: Methods and Protocols. Vol. 2261. Springer US; 2021. doi:10.1007/978-1-0716-1186-9
107. Zhang M, Viennois E, Prasad M, et al. Edible ginger-derived nanoparticles: a novel therapeutic approach for the prevention and treatment of inflammatory bowel disease and colitis-associated cancer. Biomaterials. 2016;101:321–340. doi:10.1016/j.biomaterials.2016.06.018
108. Kilasoniya A, Garaeva L, Shtam T, et al. Potential of plant exosome vesicles from Grapefruit (Citrus × paradisi) and Tomato (Solanum lycopersicum) juices as functional ingredients and targeted drug delivery vehicles. Antioxidants. 2023;12(4):943. doi:10.3390/antiox12040943
109. Chen J, Li P, Zhang T, et al. Review on strategies and technologies for exosome isolation and purification. Front Bioeng Biotechnol. 2022;9:811971. doi:10.3389/fbioe.2021.811971
110. Kim K, Park J, Jung JH, et al. Cyclic tangential flow filtration system for isolation of extracellular vesicles. APL Bioeng. 2021;5(1):016103. doi:10.1063/5.0037768
111. Taghdi MH, Al-Masawa ME, Muttiah B, et al. Three-dimensional bioprinted gelatin—genipin hydrogels enriched with hUCMSC-derived small extracellular vesicles for regenerative wound dressings. Polymers. 2025;17(9):1163. doi:10.3390/polym17091163
112. Preto RM, Dos Santos VCT, Lordelo MVS, et al. Optimization of methods for isolation and purification of outer membrane vesicles (OMVs) from Neisseria lactamica. Appl Microbiol Biotechnol. 2025;109(1):82. doi:10.1007/s00253-025-13460-y
113. Lyu Z, Yao L, Chen W, Kalutantirige FC, Chen Q. Electron microscopy studies of soft nanomaterials. Chem Rev. 2023;123(7):4051–4145. doi:10.1021/acs.chemrev.2c00461
114. Castelli G, Logozzi M, Mizzoni D, et al. Ex vivo anti-leukemic effect of exosome-like grapefruit-derived nanovesicles from organic farming-the potential role of ascorbic acid. Int J Mol Sci. 2023;24(21):15663. doi:10.3390/ijms242115663
115. Gupta P, Rai N, Verma A, Gautam V. Microscopy based methods for characterization, drug delivery, and understanding the dynamics of nanoparticles. Med Res Rev. 2024;44(1):138–168. doi:10.1002/med.21981
116. Pascucci L, Scattini G. Imaging extracellular vesicles by transmission electron microscopy: coping with technical hurdles and morphological interpretation. Biochim Biophys Acta Gen Sub. 2021;1865(4):129648. doi:10.1016/j.bbagen.2020.129648
117. Liu Q, Fu Y, Qin Z, Wang Y, Zhang S, Ran M. Progress in the applications of atomic force microscope (AFM) for mineralogical research. Micron. 2023;170:103460. doi:10.1016/j.micron.2023.103460
118. Caponnetto F, Manini I, Skrap M, et al. Size-dependent cellular uptake of exosomes. Nanomed Nanotechnol Biol Med. 2017;13(3):1011–1020. doi:10.1016/j.nano.2016.12.009
119. Szatanek R, Baj-Krzyworzeka M, Zimoch J, Lekka M, Siedlar M, Baran J. The methods of choice for extracellular vesicles (EVs) characterization. Int J Mol Sci. 2017;18(6):1153. doi:10.3390/ijms18061153
120. Chan MY, Dowling QM, Sivananthan SJ, Kramer RM. Particle sizing of nanoparticle adjuvant formulations by dynamic light scattering (DLS) and nanoparticle tracking analysis (NTA). Methods Mol Biol. 2017;1494:239–252. doi:10.1007/978-1-4939-6445-1_17
121. Alzahrani FA, Khan MI, Kameli N, Alsahafi E, Riza YM. Plant-derived extracellular vesicles and their exciting potential as the future of next-generation drug delivery. Biomolecules. 2023;13(5):839. doi:10.3390/biom13050839
122. Feng J, Xiu Q, Huang Y, Troyer Z, Li B, Zheng L. Plant‐derived vesicle‐like nanoparticles as promising biotherapeutic tools: present and future. Adv Mater. 2023;35(24):2207826. doi:10.1002/adma.202207826
123. Mu N, Li J, Zeng L, et al. Plant-derived exosome-like nanovesicles: current progress and prospects. IJN. 2023;18:4987–5009. doi:10.2147/IJN.S420748
124. Steć A, Targońska M, Jaikishan S, et al. Incorporation of doxorubicin into plant-derived nanovesicles: process monitoring and activity assessment. Drug Delivery. 2025;32(1):2439272. doi:10.1080/10717544.2024.2439272
125. Jo H, F S, Gd T, Y J, H J, Kk W. Regulation of cell-cell interactions by phosphatidic acid phosphatase 2b/VCIP. EMBO J. 2003;22(7). doi:10.1093/emboj/cdg165
126. Wang B, Zhuang X, Deng ZB, et al. Targeted drug delivery to intestinal macrophages by bioactive nanovesicles released from grapefruit. Mol Ther. 2014;22(3):522–534. doi:10.1038/mt.2013.190
127. Yan G, Xiao Q, Zhao J, et al. Brucea javanica derived exosome-like nanovesicles deliver miRNAs for cancer therapy. J Control Release. 2024;367:425–440. doi:10.1016/j.jconrel.2024.01.060
128. Bruno S, Grange C, Collino F, et al. Microvesicles derived from mesenchymal stem cells enhance survival in a lethal model of acute kidney injury. PLoS One. 2012;7(3):e33115. doi:10.1371/journal.pone.0033115
129. Gatti S, Bruno S, Deregibus MC, et al. Microvesicles derived from human adult mesenchymal stem cells protect against ischaemia-reperfusion-induced acute and chronic kidney injury. Nephrol Dial Transplant. 2011;26(5):1474–1483. doi:10.1093/ndt/gfr015
130. Plikus MV, Wang X, Sinha S, et al. Fibroblasts: origins, definitions, and functions in health and disease. Cell. 2021;184(15):3852–3872. doi:10.1016/j.cell.2021.06.024
131. Talbott HE, Mascharak S, Griffin M, Wan DC, Longaker MT. Wound healing, fibroblast heterogeneity, and fibrosis. Cell Stem Cell. 2022;29(8):1161–1180. doi:10.1016/j.stem.2022.07.006
132. Natania F, Iriawati I, Ayuningtyas FD, Barlian A. Potential of plant-derived exosome-like nanoparticles from Physalis peruviana fruit for human dermal fibroblast regeneration and remodeling. Pharm Nanotechnol. 2024. doi:10.2174/0122117385281838240105110106
133. Kim MK, Choi YC, Cho SH, Choi JS, Cho YW. The antioxidant effect of small extracellular vesicles derived from aloe vera peels for wound healing. Tissue Eng Regen Med. 2021;18(4):561–571. doi:10.1007/s13770-021-00367-8
134. Şahin F, Koçak P, Güneş MY, Özkan İ, Yıldırım E, Kala EY. In vitro wound healing activity of wheat-derived nanovesicles. Appl Biochem Biotechnol. 2019;188(2):381–394. doi:10.1007/s12010-018-2913-1
135. Jin E, Yang Y, Cong S, et al. Lemon-derived nanoparticle-functionalized hydrogels regulate macrophage reprogramming to promote diabetic wound healing. J Nanobiotechnol. 2025;23(1):68. doi:10.1186/s12951-025-03138-y
136. Porwal D, Das B. Medical implications of gas producing probiotics: anti-inflammation and immunity enhancement for ameliorating disease severity. Med Gas Res. 2024;14(3):140–141. doi:10.4103/mgr.MEDGASRES-D-23-00041
137. Lu X, Hu W, Wang X, Wang Z, Yang P, Wang W. Protective role of methane in traumatic nervous system diseases. Med Gas Res. 2024;14(3):159–162. doi:10.4103/mgr.mgr_23_23
138. Ramírez O, Pomareda F, Olivares B, et al. Aloe vera peel-derived nanovesicles display anti-inflammatory properties and prevent myofibroblast differentiation. Phytomedicine. 2024;122:155108. doi:10.1016/j.phymed.2023.155108
139. Zhao X, Yin F, Fu L, et al. Garlic-derived exosome-like nanovesicles as a hepatoprotective agent alleviating acute liver failure by inhibiting CCR2/CCR5 signaling and inflammation. Biomater Adv. 2023;154:213592. doi:10.1016/j.bioadv.2023.213592
140. Lawrence T. The nuclear factor NF-κB pathway in inflammation. Cold Spring Harb Perspect Biol. 2009;1(6):a001651. doi:10.1101/cshperspect.a001651
141. Kang M, Kang M, Lee J, Yoo J, Lee S, Oh S. Allium tuberosum -derived nanovesicles with anti-inflammatory properties prevent DSS-induced colitis and modify the gut microbiome. Food Funct. 2024;15(14):7641–7657. doi:10.1039/D4FO01366B
142. Raimondo S, Urzì O, Meraviglia S, et al. Anti‐inflammatory properties of lemon‐derived extracellular vesicles are achieved through the inhibition of ERK/NF‐κB signalling pathways. J Cell Mol Med. 2022;26(15):4195–4209. doi:10.1111/jcmm.17404
143. Grad S, Ertel W, Keel M, Infanger M, Vonderschmitt DJ, Maly FE. Strongly enhanced serum levels of vascular endothelial growth factor (VEGF) after poly-trauma and burn. cclm. 1998;36(6):379–383. doi:10.1515/cclm.1998.064
144. Savcı Y, Kırbaş OK, Bozkurt BT, et al. Grapefruit-derived extracellular vesicles as a promising cell-free therapeutic tool for wound healing. Food Funct. 2021;12(11):5144–5156. doi:10.1039/d0fo02953j
145. Sulaiman M, Alyileili SR, Raghavankutty M, Kurup GM. Sulfated polysaccharide ascophyllan from Padina tetrastromatica enhances healing of burn wounds by ameliorating inflammatory responses and oxidative damage. Mol Biol Rep. 2020;47(11):8701–8710. doi:10.1007/s11033-020-05914-5
146. Perut F, Roncuzzi L, Avnet S, et al. Strawberry-derived exosome-like nanoparticles prevent oxidative stress in human mesenchymal stromal cells. Biomolecules. 2021;11(1):87. doi:10.3390/biom11010087
147. Zhao W, Bian Y, Wang Q, et al. Blueberry-derived exosomes-like nanoparticles ameliorate nonalcoholic fatty liver disease by attenuating mitochondrial oxidative stress. Acta Pharmacol Sin. 2022;43(3):645–658. doi:10.1038/s41401-021-00681-w
148. Urzì O, Cafora M, Ganji NR, et al. Lemon-derived nanovesicles achieve antioxidant and anti-inflammatory effects activating the AhR/Nrf2 signaling pathway. Iscience. 2023;26(7):107041. doi:10.1016/j.isci.2023.107041
149. Di Raimo R, Mizzoni D, Aloi A, et al. Antioxidant effect of a plant-derived extracellular vesicles’ mix on human skin fibroblasts: induction of a reparative process. Antioxidants. 2024;13(11):1373. doi:10.3390/antiox13111373
150. Logozzi M, Di Raimo R, Mizzoni D, Fais S. Nanovesicles from organic agriculture-derived fruits and vegetables: characterization and functional antioxidant content. IJMS. 2021;22(15):8170. doi:10.3390/ijms22158170
151. Alharbi NS, Khaled JM, Kadaikunnan S, et al. Prevalence of Escherichia coli strains resistance to antibiotics in wound infections and raw milk. Saudi J Biol Sci. 2019;26(7):1557–1562. doi:10.1016/j.sjbs.2018.11.016
152. Saroj S, Saha S, Ali A, et al. Plant extracellular nanovesicle-loaded hydrogel for topical antibacterial wound healing in vivo. ACS Appl Bio Mater. 2024. doi:10.1021/acsabm.4c00992
153. Hassanin DS, Abdelhady SR, Ghazi AK, Badawy WZ. Nanoparticles of banana peels as a potential source of bioactive compounds and their activities on HepG2. Sci Rep. 2025;15(1):11721. doi:10.1038/s41598-025-93382-x
154. Yu H, Zhang J, Yang L, et al. MSC-derived exosomes injectable hyaluronic acid hydrogel for enhanced chronic wound healing. J Control Release. 2025;385:113985. doi:10.1016/j.jconrel.2025.113985
155. Wang Y, Zhao G, S A, et al. In situ forming hypoxia‐induced exosome‐loaded hydrogel for enhanced diabetic wound healing. Macro Mater Eng. 2025;310(5):2400402. doi:10.1002/mame.202400402
156. Xie H, Wang Z, Wang R, Chen Q, Yu A, Lu A. Self‐healing, injectable hydrogel dressing for monitoring and therapy of diabetic wound. Adv Funct Mater. 2024;34(36):2401209. doi:10.1002/adfm.202401209
157. Wang Z, Qi B, Wang R, et al. Smart eutectogel with antibacterial activity for efficiently treating multidrug resistant infection, real-time monitoring and diabetic wound repair. Chem Eng J. 2025;520:166108. doi:10.1016/j.cej.2025.166108
158. Du J, Chu Y, Hu Y, et al. A multifunctional self-reinforced injectable hydrogel for enhancing repair of infected bone defects by simultaneously targeting macrophages, bacteria, and bone marrow stromal cells. Acta Biomater. 2024;189:232–253. doi:10.1016/j.actbio.2024.10.014
159. Y W, X Z, W X, et al. Microenvironment-responsive collagen hydrogel with Houttuynia Cordata Thunb vesicles for diabetic wound repair. Int J Biol Macromol. 2025;320(Pt 2). doi:10.1016/j.ijbiomac.2025.145840
160. He J, Ren W, Wang W, et al. Exosomal targeting and its potential clinical application. Drug Deliv Transl Res. 2022;12(10):2385–2402. doi:10.1007/s13346-021-01087-1
161. Xin L, Shen J, Huang Z, et al. Ultrasound-launched targeted nanoparticle enhances antibacterial sonodynamic therapy for effective eradication of pseudomonas aeruginosa biofilm. BIOI. 2024;5(1). doi:10.15212/bioi-2024-0001
162. Wang C, Tsai T, Lee C. Regulation of exosomes as biologic medicines: regulatory challenges faced in exosome development and manufacturing processes. Clin Transl Sci. 2024;17(8):e13904. doi:10.1111/cts.13904
163. Foley JA, DeFries R, Asner GP, et al. Global consequences of land use. Science. 2005. doi:10.1126/science.1111772
164. Kishta MS, Hafez AM, Hydara T, et al. The transforming role of wharton’s jelly mesenchymal stem cell-derived exosomes for diabetic foot ulcer healing: a randomized controlled clinical trial. Stem Cell Res Ther. 2025;16(1):559. doi:10.1186/s13287-025-04690-y
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.