Back to Journals » Drug Design, Development and Therapy » Volume 19
Pharmacological Potential and Molecular Targets of Tetrahydrofurofuranoid Lignans From Magnoliae Flos
Authors Yi X, Xiao Z, Chen J, Chen G, Ma P
Received 19 June 2025
Accepted for publication 9 September 2025
Published 3 October 2025 Volume 2025:19 Pages 9011—9027
DOI https://doi.org/10.2147/DDDT.S547924
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Anastasios Lymperopoulos
Xiaohan Yi, Zixuan Xiao, Jiaxue Chen, Guoren Chen, Ping Ma
School of Basic Medicine, Chengdu University of Traditional Chinese Medicine, Chengdu, 611137, People’s Republic of China
Correspondence: Ping Ma, School of Basic Medicine, Chengdu University of Traditional Chinese Medicine, No.1166, Liutai Avenue, Wenjiang District, Chengdu, 611137, People’s Republic of China, Email [email protected]
Abstract: Tetrahydrofurofuranoid lignans (TFLs) are lignans extracted from the dried flower buds of Magnoliae Flos (MF). The chemical structures of the seven major constituents, magnolin, fargesin, aschantin, lirirresinol B dimethyl ether, epimognolin, eudesmin, and kobusin, are strikingly similar. Despite their potential therapeutic effects against various pathological conditions, their underlying mechanisms have not been systematically analyzed. The primary objective of this review was to examine the mechanisms by which TFLs exert therapeutic effects on inflammation, allergies, cancer, cardiovascular diseases, metabolic disorders and osteoarticular diseases. Data were retrieved from scientific databases, including PubMed, Web of Science, and Google Scholar, up to November 2024. The search terms employed included “Magnoliae Flos” or “Magnolia” and “Lignans” “Tetrahydrofurofuranoid” “Magnolin” “Fargesin” “Aschantin” “Lirirresinol B dimethyl ether” “Epimagnolin” “Eudesmin” and “Kobusin.” TFLs have been demonstrated to treat various inflammatory diseases through inhibition of signaling pathways, including NF-κB and MAPK. Additionally, allergic reactions are suppressed via inhibition of mast cell degranulation-related receptors. Furthermore, anticancer activity is achieved by triggering signaling pathways such as PI3K/AKT/mTOR, ERK/MEK, and P53. Besides enhancing glucose metabolism, TFLs suppress adipogenesis through downregulation of pathways involving PI3K/Akt, Akt/AMPK, and P-selectin expression. Fargesin, a notable compound derived from MF, exerts cardioprotective effects by inhibiting the cAMP/PKA pathway and counteracts atherosclerosis through the downregulation of the TLR4/NF-κB signaling pathway. Notably, TFLs also demonstrate the ability to reduce melanin synthesis by suppressing tyrosinase, associated proteins, MITF, and enzymes involved in melanogenesis. Growing research highlights diverse pharmacological attributes of TFLs, providing critical insights for future studies aimed at developing novel therapeutics exhibiting enhanced efficacy and minimal toxicity for inflammation, cancer, cardiovascular disorders, diabetes, skin ultraviolet damage and osteoporosis.
Keywords: Xin-yi, Magnolia, Lignans, pharmacology, mechanisms, toxicity
Introduction
Magnoliae Flos (MF), comprising the dried flower buds of Magnolia species harvested in late winter or early spring before bloom, has been historically employed in traditional medicine for treating conditions such as allergic rhinitis, headaches, sinusitis, rheumatoid arthritis, and asthma, as shown in Figure 1. Magnolia denudata Desr.,Magnolia biondii Pamp.,Magnolia fargesii Cheng.,Magnolia sprengeri Pamp.,Magnolia liliiflora Desr.,and Magnolia kobus DC are the most frequently used medicinal materials in China, Japan, and South Korea. Currently, only a few species from the genus Magnolia are considered medicinal sources. MF is referred to as Xin-yi in China, Shin-Yi in South Korea, and Shen-i in Japan. The Pharmacopoeia of the People’s Republic of China includes M. denudata, M. biondii, and M. sprengeri.1–3
 |
Figure 1 Magnoliae Flos denotes fresh flower buds. After sun-drying, these buds yield the TCM material known as Xinyi, employed for various therapeutic indications. Created in BioRender. xiaohan, Y. (2025) https://BioRender.com/duizcpq. |
The primary constituents of MF species are categorized into two classes: lipid-soluble and water-soluble components. Lipid-soluble compounds, typically extracted using alcoholic solvents, are abundant in volatile oils and lignans.2,4 Lipid-soluble constituents are the primary active components of MF. Numerous recent studies have investigated the chemical composition of volatile components and lignans in this medicinal plant. TFLs, as non-volatile lignans, represent the most abundant and bioactive constituents in MF lignan studies. Due to their relatively high stability, crude extracts can be obtained through alcohol extraction (MFE) or aqueous extraction (MKE).5 These bioactive lignans have demonstrated efficacy in inhibiting inflammatory processes, exerting anti-allergic and anti-tumor effects, improving glucose metabolism, and providing cardiovascular protection. However, a systematic synthesis of available evidence remains lacking. This review comprehensively evaluates TFLs concerning chemical composition, clinical applications, pharmacological mechanisms, pharmacokinetic profiles, toxicological properties, and critical influencing factors. Furthermore, perspectives on future research directions are provided to facilitate the development of therapeutics derived from MF and optimize their clinical application.
TFLs represent optically active, naturally occurring dimeric compounds composed primarily of two phenylpropane (C6-C3) units interconnected by β’-β’ bonds. These structural units feature a characteristic 2,6-diaryl-3,7-dioxabicyclo [3.3.0] octane skeleton linked via positions 8–8’, 7-O-9′, and 9-O-7′, as in Figure 2a.6,7 Structures of these compounds vary based on stereochemical configurations of oxygen-containing substituents in the hydrocarbon chain and benzene ring. As mixtures of enantiomers, they exhibit significant variation in enantiomeric composition. Optical purity of TFLs and furan lignans has not been demonstrated through chiral high-performance liquid chromatography (HPLC) analysis.8,9 According to plasma pharmacochemistry theory, only components absorbed into the bloodstream exert pharmacological effects. Research has demonstrated that seven TFLs from MF, magnolin, fargesin, aschantin, lirirresinol B dimethyl ether, epimagnolin, eudesmin, and kobusin can be absorbed and metabolized in the bloodstream.10 As presented in Figure 2b, their chemical structures exhibit high similarity.
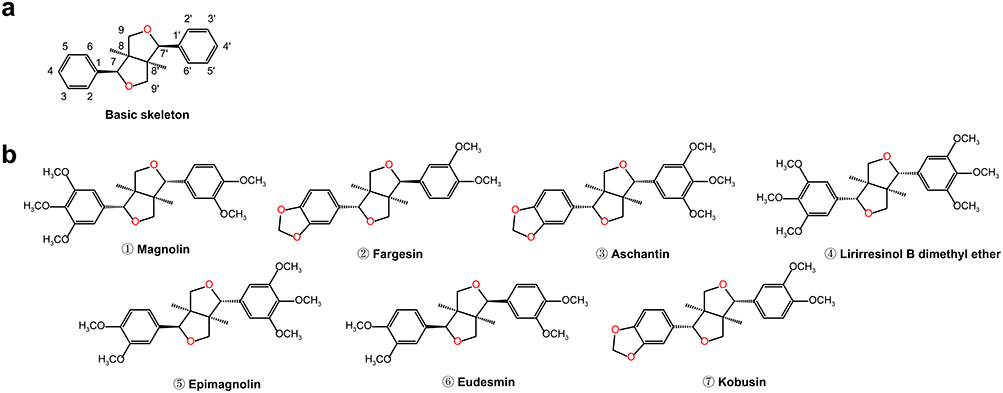 |
Figure 2 The chemical structures of bioactive tetrahydrofurofuranoid lignans isolated from Magnoliae Flos. (a) Skeletal framework of tetrahydrofurofuranoid lignans. (b) ①–⑦: Chemical structures of seven tetrahydrofurofuranoid lignans. |
The earliest written record of medicinal use of Xin-yi (MF) traces back to “Shen Nong’s Classic of Materia Medica” describing only its properties, flavors, and efficacy without details about plant characteristics or geographical distribution. During the Song Dynasty, Xinyi Powder, primarily composed of Xin-yi, became a classic prescription for nasal ailments, which remains effective and in use today.11 Xinyi Qingfei Decoction, originating from “The Canon of Surgery” is used to treat cough induced by lung heat and nasal congestion accompanied by thick discharge.12 Cangerzi San, a traditional Chinese medicine prescription from “Jisheng Fang” treats rhinitis. In this formula, Cangerzi and Xin-yi are warm, pungent herbs effective in treating allergic rhinitis and asthma by opening nasal passages.13 Recently, MF has been widely utilized in clinical treatments, and lignans have been identified as its primary active components. Notably, tetrahydrofurofuran-type lignans are the main type absorbed into the bloodstream, exerting pharmacological effects.14 Xinfangbiyan capsules, containing Xin-yi as a primary component, are utilized to treat chronic rhinitis, sinusitis, and related conditions in traditional Chinese medicine. Several studies have isolated magnolin from this preparation.15 Xinyi Biyan Pill is therapeutically employed for managing both acute and chronic rhinitis as well as allergic rhinitis. Magnolin, detected through ultra-high performance liquid chromatography coupled with a diode array detector fingerprinting and multi-component ultra-high performance liquid chromatography-tandem mass spectrometry (UPLC-MS/MS) analysis, was identified as a component of Xinyi Biyan Pill.16 Another traditional Korean medicine, NDC-052, containing magnolin and epimagnolin A, originates from M. fargesii. Clinical trials have demonstrated its effectiveness in alleviating symptoms and improving pulmonary function in patients with moderate asthma, leading to extensive clinical use.17,18
Pharmacological Properties of Tetrahydrofurofuranoid Lignans
Inflammatory Diseases
In 1987, Pan et al19 discovered that MF could antagonize platelet-activating factor. They extracted six active constituents: magnolin, fargesin, aschantin, lirirresinol B dimethyl ether, eudesmin, and kobusin, suggesting potential involvement in cardiovascular diseases, respiratory conditions, and inflammation. Numerous recent studies have demonstrated that TFLs possess significant anti-inflammatory properties, exhibiting inhibitory effects on diverse inflammatory disorders. Their actions effectively control the progression of conditions including hepatitis, osteoarthritis, pneumonia, periodontitis, neuritis, and inflammatory bowel disease. TFLs primarily exert their anti-inflammatory effects by regulating inflammatory factor production and inhibiting associated pathways.
Inhibition of TNF-α
Research has demonstrated that M. biondii extracts significantly suppress TNF-α production in LPS-induced RAW 264.7 macrophages. Eudesmin, magnolin, and lirirresinol B dimethyl ether were isolated from these extracts.20 Cho et al21 confirmed that eudesmin markedly inhibits TNF-α production and T lymphocyte proliferation. Ahn et al22 showed that six lignans (magnolin, fargesin, aschantin, lirirresinol B dimethyl ether, eudesmin, and kobusin) effectively suppress TNF-α-induced adhesion molecule expression in THP-1 cells. Furthermore, MFE significantly decreased inflammatory mediators such as TNF-α, IL-1β, and COX-2, effectively preventing tissue damage and alveolar bone loss in periodontitis rat models.22 Collectively, lignan constituents from MF markedly inhibit the activation of numerous pro-inflammatory mediators, especially TNF-α.
NF-κB and MAPK Pathways
Numerous in vitro investigations have demonstrated that constituents including MFE,23 MKE,24 magnolin,25 fargesin,26 and lirirresinol B dimethyl ether27 concurrently suppress NF-κB and MAPK signaling pathways in LPS-stimulated macrophages, thus reducing the production of pro-inflammatory cytokines. Epimagnolin28 exhibited comparable inhibitory activity in macrophages stimulated by Phorbol 12-myristate 13-acetate (PMA). Complementary in vivo studies confirmed that fargesin alleviates ulcerative colitis29 and cadmium-induced acute lung injury30 by inhibiting NF-κB signaling. Additionally, fargesin simultaneously suppressed both NF-κB and MAPK pathways, markedly reducing pro-inflammatory cytokines, cartilage breakdown, and synovial inflammation in mice with collagen-induced osteoarthritis.31 Similarly, magnolin attenuated osteoarthritis progression by targeting NF-κB signaling and preserving cartilage integrity in osteoarthritic rat models.32 Epimagnolin and fargesin decreased ERK phosphorylation, suppressed inducible nitric oxide synthase (iNOS) expression, and lowered NO generation, thereby alleviating inflammation in human respiratory epithelial cells.33 These observations have been substantiated through rigorous in vitro and in vivo experiments. Seven lignans, including eudesmin, magnolin, lirirresinol B dimethyl ether, epimagnolin, kobusin, aschantin, and fargesin, isolated from the CHCl3 fraction of MF, inhibited cigarette smoke condensate-induced IL-6 expression in human airway epithelial cells through suppression of ERK phosphorylation. In murine models, these compounds significantly reduced inflammatory cell infiltration and pro-inflammatory cytokine secretion. Notably, aschantin exerted similar effects by selectively inhibiting Akt phosphorylation.34 Lirirresinol B dimethyl ether demonstrated therapeutic effects in acute liver fibrosis mouse models and inhibited carbon tetrachloride-induced hepatocellular carcinoma via the NF-κB/COX-2 pathway.35
TFLs also exhibit anti-neuroinflammatory effects. In vitro, aschantin, kobusin, and fargesin blocked NF-κB pathways and reduced iNOS expression in activated BV-2 microglial cells, thereby suppressing NO production.36 In vivo, magnolin alleviated paclitaxelinduced cold allodynia via the MAPK/ERK pathway.37
Collectively, these studies illustrate that TFLs exert anti-inflammatory properties primarily by concurrently inhibiting the NF-κB and MAPK signaling cascades. Specifically, TFLs markedly decrease phosphorylation levels of P65/P50 in the NF-κB pathway and diminish phosphorylation of ERK, JNK, and p38 within the MAPK pathway. This simultaneous blockade curtails inflammation by significantly lowering the production of critical inflammatory mediators such as TNF-α, IL-6, iNOS, COX-2, and IL-1β. Given the frequent interplay and synergistic effects observed between the NF-κB and MAPK signaling pathways, these findings provide a comprehensive elucidation of the molecular basis for TFLs’ anti-inflammatory actions, as shown in Figure 3. Moreover, MKE38 was reported to suppress pro-inflammatory cytokines and ERK1/2 signaling in human keratinocytes stimulated by TNF-α and IFN-γ, while also inhibiting JAK/STAT signaling pathways, thus alleviating scratching behaviors in mice induced with 2,4-dinitrochlorobenzene to mimic atopic dermatitis. Such diverse regulatory mechanisms may be due to variations in the bioactive constituents within TFLs crude extract mixtures.
 |
Figure 3 Under LPS-induced inflammatory conditions, the inactive NF-κB complex (p50, p65, and IκB) is activated, leading to IκB phosphorylation and degradation and subsequent activation of the NF-κB pathway. Concurrently, AP-1 is regulated by MAPK family members (p38, ERK, and JNK), promoting the release of inflammatory mediators. Tetrahydrofurofuranoid lignans have been shown to suppress activation of both pathways. Created in BioRender. xiaohan, Y. (2025) https://BioRender.com/dl0f2wz. |
Others
Research has also highlighted magnolin’s protective effects on the intestinal barrier through inhibition of the PI3K/Akt pathway, thereby reducing pro-inflammatory cytokine production and alleviating colitis resembling Crohn’s disease both in vitro and in vivo.39 However, another study reported that low concentrations of magnolin suppressed 5-lipoxygenase (ALOX5) expression without influencing glutathione peroxidase 4 in dextran sulfate sodium-induced colitis in mice. Interestingly, higher magnolin doses failed to produce significant effects, possibly due to concurrent inhibition of ferroptosis and intestinal epithelial cell regeneration, a hypothesis that requires additional verification.40 Additionally, magnolin conferred protection to liver and kidney tissues by reducing oxidative stress, inflammation, and cytotoxicity induced by the chemotherapeutic agent cyclophosphamide via activation of the Nrf2/HO-1 signaling pathway.41
In conclusion, experimental evidence supports the anti-inflammatory effects of TFLs through the NF-κB and MAPK pathways, which represent the primary mechanisms regulating inflammation. Nevertheless, comprehensive investigations remain necessary for clinical validation.
Allergic Diseases
Mast cells (MCs) are pivotal mediators in allergic reactions, activated and triggered into degranulation by virtually all known allergenic stimuli.42 A principal activation receptor present on all mast cells is the high-affinity IgE/Fc receptor. Compound 48/80 is widely recognized as a strong inducer of MC degranulation, primarily via the MAS-related G protein-coupled receptor (MRGPRX2) on human MCs.43 Calcium influx and activation of protein kinase C are critical factors in MC degranulation. Following activation, increased intracellular Ca2+ levels significantly promote degranulation,44 releasing inflammatory mediators such as histamine, leukotrienes, prostaglandins, cytokines, proteases, kinases, and nitric oxide, thereby sensitizing cells.45 As demonstrated in Figure 4, TFLs have been identified as effective inhibitors of MC degranulation by suppressing calcium signaling and reducing allergenic mediator release, providing a promising therapeutic strategy for allergic reactions.
 |
Figure 4 Mast cell activation via two distinct pathways, IgE-dependent and MRGPRX2-mediated signaling, stimulates phospholipase C (PLCγ and PLCβ, respectively), generating IP3 and DAG. IP3 interacts with its receptor (IP3R) on the endoplasmic reticulum (ER), inducing Ca2+ release. ER calcium depletion induces STIM1 accumulation adjacent to the plasma membrane, activating ORAI1 channels and facilitating store-operated calcium entry. Increased intracellular Ca2+ subsequently activates the calcium-dependent chloride channel ANO1, resulting in chloride efflux, membrane depolarization, and amplified calcium influx, thereby establishing a positive feedback loop that enhances downstream responses. Magnolin, fargesin, aschantin, lirioresinol dimethyl ether B, epimagnolin, eudesmin, and kobusin inhibited IgE-induced histamine release from mast cells. Notably, magnolin did not suppress histamine release triggered by MRGPRX2 activation. Furthermore, magnolin, fargesin, and eudesmin effectively inhibited ORAI1 calcium channel activity, with eudesmin also exhibiting inhibitory effects on the ANO1 calcium-activated chloride channel. Created in BioRender. xiaohan, Y. (2025) https://BioRender.com/6xmgl84. |
Inhibition of Histamine
As early as 1999, MKE was reported to inhibit histamine release in rat peritoneal mast cells induced by either compound 48/80 or anti-DNP IgEin a dose-dependent manner, though specific lignan constituents were not identified at that time.46 Subsequent research isolated nine bisepoxylignans from MFE, TFLs of which showed inhibitory effects on bone marrow-derived mast cells and IgE-mediated passive cutaneous anaphylaxis in mouse models. Fargesin exhibited the most potent activity (IC50 = 93.03 μM). Increased methoxylation on the benzene ring correlated with diminished anti-allergic efficacy, and symmetrical bisepoxylignans showed weaker anti-allergic effects compared to asymmetrical compounds.47 Fargesin and eudesmin exhibited anti-allergic activities by reducing histamine and β-hexosaminidase release from MRGPRX2-stimulated MCs.48 A systematic comparative evaluation of six Magnolia species using a compound 48/80-induced histamine release assay in rat peritoneal mast cells revealed the inhibitory potency (IC50 values) in descending order as follows: M. biondii (0.37 mg/mL) being the most potent, kobus (0.68 mg/mL) > M. liliflora (7.80 mg/mL) > M. denudata (12.63 mg/mL) > M. sprengeri (24.75 mg/mL) > M. sargentiana (57.82 mg/mL). Notably, magnolin only marginally inhibited histamine release, whereas fargesin had no significant effect.49 These contradictory findings regarding fargesin’s inhibition of the MRGPRX2 pathway may reflect substantial variations in TFLs content or composition among different Magnolia species.
Suppression of Ca2+ Signaling
Further studies indicated magnolin’s ability to suppress IgE/Ag-triggered pseudo-allergic responses by attenuating Ca2+ influx, mast cell degranulation, and chemokine/cytokine secretion, demonstrating dose-dependent effects in vitro and in vivo. Nevertheless, magnolin did not significantly affect histamine release from MCs activated by compound 48/80 or substance P,50 indicating a lack of inhibitory effect on non-IgE-mediated activation pathways. In an in vitro study, Joo Hyun Nam’s team isolated magnolin, eudesmin, and fargesin from a 70% aqueous-30% ethanolic extract of FM (FMEtOH). Experiments demonstrated that both FMEtOH and eudesmin inhibited the classical pathway of TMEM16A Anoctamin 1 (ANO1) calcium-activated chloride channels in ANO1-overexpressing HEK293 cells and airway epithelial cells, whereas magnolin and fargesin showed negligible inhibitory effects.51 In another in vitro study, the same group found that all three compounds (magnolin, eudesmin, and fargesin) suppressed allergic responses by inhibiting Oral calcium release-activated calcium channel protein 1 (ORAI1) channel activity (a GPCR-coupled channel), CD4+ T-cell proliferation, and MC degranulation, with fargesin exhibiting the strongest inhibition.52 These results suggest ion channel selectivity differences between magnolin and fargesin, though precise mechanisms require further elucidation.
Additionally, fargesin, eudesmin, and lirirresinol B dimethyl ether suppress leukotriene synthesis through ALOX5 inhibition, thus exhibiting anti-allergic properties.53 Aschantin and kobusin effectively mitigate allergic responses and significantly downregulate IL-2 production in Jurkat T cells.54
Cancer Diseases
Cancer is a disease that can involve multiple organs and tissues. Its development involves complex cellular processes, including proliferation, survival, invasion, metastasis, and mutations.55 Commonly activated or mutated signaling pathways in cancer include PI3K/AKT/mTOR and RAS/MEK/ERK. Additionally, the P53, STAT3, Wnt/β-catenin, and MYC pathways significantly contribute to cancer development and represent targets for therapeutic strategies.56 Experimental evidence demonstrates that TFLs inhibit cancer cell proliferation and tumor growth (Table 1).
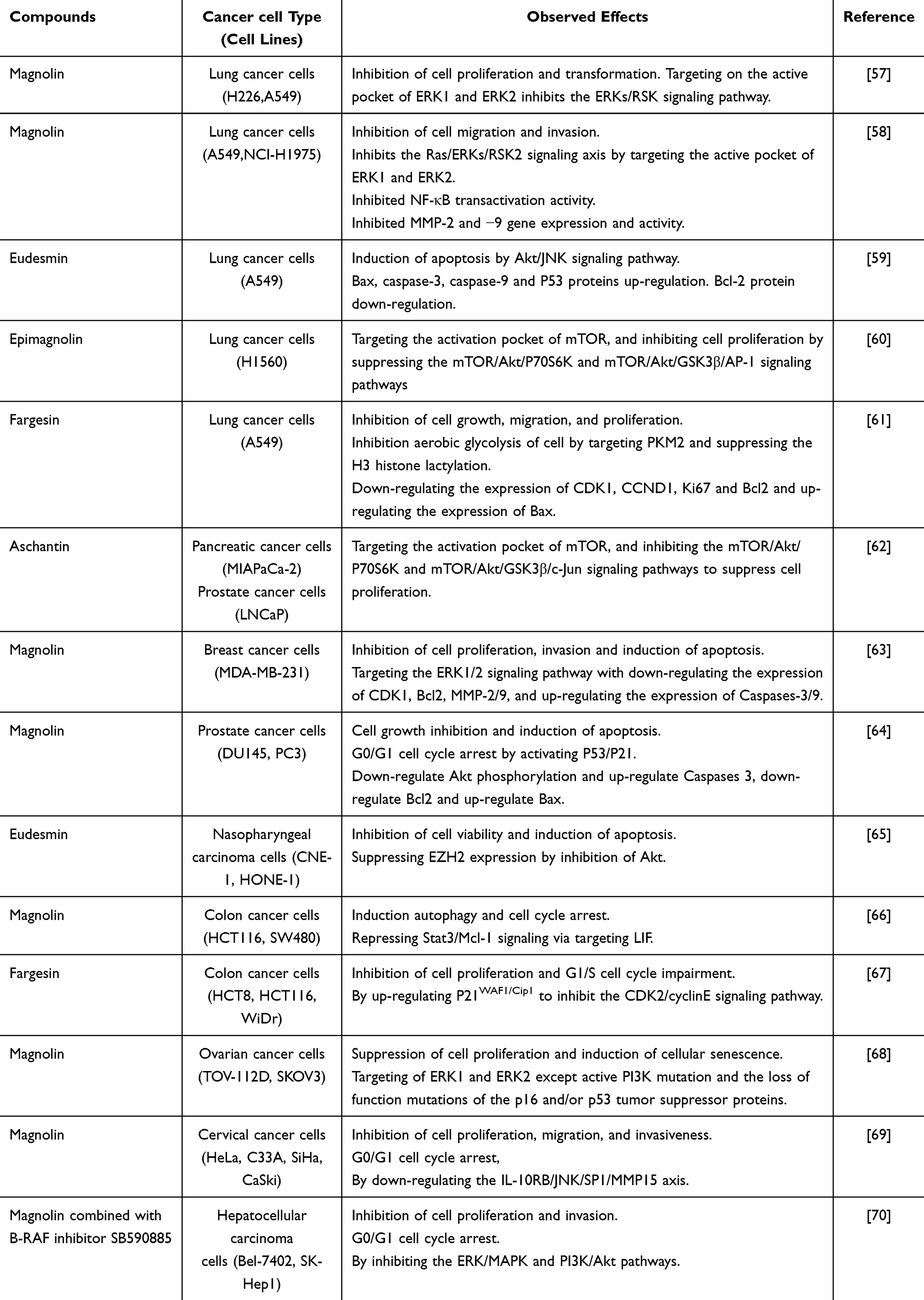 |
Table 1 Pharmacological Mechanisms of TFLs Involved in Its Anticancer Activities |
PI3K/AKT/mTOR
The phosphatidylinositide 3-kinase PI3K/AKT signaling cascade is pivotal in regulating glucose metabolism, protein synthesis, cellular proliferation, and growth, and abundant evidence highlights its significance as a target in oncology.71 The serine/threonine kinase mammalian target of rapamycin (mTOR), consisting of mTORC1 and mTORC2 complexes, orchestrates AKT-driven cell growth and transformation.72 mTORC2 directly phosphorylates AKT at Ser473, activating it fully and subsequently regulating mTORC1 downstream activities.71 Yong-Yeon Cho and colleagues identified aschantin as targeting the catalytic site of mTOR kinase, impeding complete AKT activation, thereby inhibiting the AKT/mTORC1/P70S6K axis and selectively obstructing downstream GSK3β signaling, effectively preventing EGF-mediated transformation and proliferation of MIAPaCa-2 and LNCaP cancer cell lines.62 Their further investigations indicated epimagnolin notably impeded H1650 lung cancer cell proliferation via similar pathways, whereas H460 cells remained unaffected.60 Moreover, eudesmin displayed robust antitumor efficacy against A549 lung cancer cells through inhibiting AKT phosphorylation and facilitating JNK activation, consequently inducing apoptosis both in vitro and in vivo.59 Eudesmin also inhibited EZH2 gene expression in nasopharyngeal carcinoma (NPC) by suppressing AKT, thereby reducing NPC cell proliferation, migration, and invasion.65
RAS/MEK/ERK
Extracellular signal-regulated kinases (ERKs), critical MAPK family constituents, function downstream of surface receptors and cytoplasmic signaling molecules. ERKs often exhibit dysregulated activity in cancers and various diseases.73 Tumor promoters, including EGF and TPA, initiate phosphorylation of ERK1/2 and RSKs, facilitating G1/S transition and promoting cellular proliferation.74 Magnolin, a primary bioactive component of TFLs, exhibits significant anticancer properties. Yong-Yeon Cho’s team demonstrated magnolin’s ability to inhibit the ERK1/2 and downstream ERKs/RSK2 pathways, mediating its anticancer effects through these critical signaling mechanisms.57 Another study showed magnolin suppresses NF-κB transactivation by blocking ERKs/RSK2 signaling, downregulating matrix metalloproteinases (MMP)2 and 9, and effectively inhibiting lung cancer cell proliferation and migration.58 Furthermore, systematic evaluation revealed magnolin significantly inhibited ovarian cancer cell growth, but minimally affected pancreatic, colon, and breast cancer cells. Interestingly, magnolin suppressed ERK1/2 phosphorylation in TOV-112D ovarian cancer cells, whereas it was ineffective in SKOV3 cells, possibly due to constitutive PI3K activation and p16/p53 mutations in the latter.68 The same team confirmed that neither epimagnolin nor aschantin exerted antiproliferative or antimigratory effects via ERKs/RSK2 signaling.60 Shengchu Zhang’s team found that magnolin effectively inhibited breast cancer progression by suppressing MEK1/2 and ERK1/2 phosphorylation, attenuating proliferation and invasion, and inducing apoptosis in triple-negative breast cancer (MDA-MB-231) cells.63 These differing outcomes may stem from cell-type-specific variations in signaling regulation. A combination therapy study revealed synergistic effects between magnolin and a BRAF inhibitor (SB590885), targeting both PI3K/AKT/mTOR and ERK/MEK pathways.70 Integration of chemotherapeutics with traditional Chinese medicine in future therapies may significantly enhance anticancer efficacy, warranting further investigation.
P53/P21
The tumor suppressor p53, functioning as a transcriptional regulator, modulates pivotal gene expressions such as CDKN1A (p21) for inducing cell cycle arrest and senescence, and pro-apoptotic genes like BCL2-associated X protein (BAX) and p53 upregulated modulator of apoptosis. Given its central regulatory role, targeting p53 has emerged as a promising cancer therapeutic strategy.75 The CDK inhibitor p21WAF1/CIP1 mediates p53-dependent cell cycle arrest, apoptosis, and senescence following DNA damage.76 Recent research indicated magnolin effectively induced cell cycle arrest in prostate cancer cells, initiating apoptosis through activation of the p53/p21 signaling pathway both in vitro and in vivo, significantly hindering tumor development and cell viability.64 Additionally, fargesin disrupted the cell cycle by suppressing CDK2/cyclin E, thereby inhibiting colon cancer cell proliferation via enhanced expression of p21WAF1/CIP1.67
Others
Additionally, fargesin disrupted the cell cycle by suppressing CDK2/cyclin E, thereby inhibiting colon cancer cell proliferation via enhanced expression of.66 Interestingly, magnolin also inhibits migration and invasion in cervical cancer cells by targeting IL-10 receptor B and suppressing JNK/SP1-mediated MMP15 expression.69 Recent studies indicate that fargesin suppresses lung cancer cell growth and proliferation by targeting pyruvate kinase muscle isozyme M2, a key rate-limiting enzyme in glycolysis, through the glycolytic metabolism pathway linked to histone Histone H3 lactylation.61
These studies highlight distinct signaling pathways through which TFLs affect various cancers. Despite their similar chemical structures, minor structural variations among TFLs significantly influence molecular targeting of cancer-related proteins.
Cardiovascular Diseases
Fargesin exhibits significant cardioprotective effects, potentially acting through β1-adrenergic receptor (β1-AR) antagonism. It safeguards cardiac tissue by suppressing the cAMP/PKA pathway and reducing ischemia/reperfusion-induced apoptosis through diminished Caspase-3 activation in experimental animal models.77 Additionally, fargesin effectively reduces blood pressure and induces vasodilation, potentially via antioxidative mechanisms and stimulation of NO synthesis, an effect independent of dosage.78 These findings indicate fargesin’s capability to enhance peripheral circulation and ameliorate myocardial ischemia. Moreover, fargesin significantly diminishes atherosclerotic lesions and vascular inflammation in apoE−/− mice by enhancing reverse cholesterol transport through elevated ATP-binding cassette transporter A1 and ATP-binding cassette transporter G1 expressions mediated by the CEBPαS21/LXRα pathway, and reducing inflammation via TLR4/NF-κB pathway inhibition.79 Consequently, fargesin holds substantial promise as a therapeutic candidate for cardiovascular pathologies.
Metabolic-Related Diseases
MKE was first reported as a potential preventive agent against type 1 diabetes. Pretreatment with MKE completely prevented low dose streptozotocin-induced diabetes in mice, accompanied by reduced NO production in RIN-m5F cells and pancreatic islets. This treatment protected β-cells from cytokine-induced toxicity through inhibition of the NF-κB pathway.80 Recent studies show that fargesin influences metabolism and adipogenesis. Magnolin, epimagnolin, and kobusin isolated from MFE significantly enhanced basal glucose uptake in L6 myotubes, with fargesin exhibiting the strongest effect. Mechanistic studies indicated fargesin specifically acts through the insulin-mediated PI3K/Akt pathway rather than the AMPK pathway, thereby promoting GLUT4 translocation.81 However, mechanisms underlying glucose uptake stimulation by epimagnolin and kobusin remain unclear. Moreover, fargesin upregulated GLUT4 protein expression and increased phosphorylation of Akt, AMPK, and acetyl-CoA carboxylase in 3T3-L1 adipocytes and white adipose tissue of high-fat diet-induced obese mice.82 An in vitro study utilizing methano and dichloromethane extracts of MF(MF-MD) demonstrated that fargesin, magnolin, eudesmin, and epimagnolin inhibited key transcription factors for lipid synthesis, including Sterol Regulatory Element-Binding Protein 1, Peroxisome Proliferator-Activated Receptor γ, and CCAAT/Enhancer-Binding Protein α.83 Additionally, eudesmin inhibited adipocyte differentiation by downregulating the S6K1-H2BS36p axis and suppressing Wnt gene expression in mouse mesenchymal stem cells.84 Overall, TFLs possess therapeutic potential against insulin resistance and obesity-related diseases by promoting glucose metabolism and inhibiting adipogenesis.
Skin Anti-Darkening Effect
Early studies demonstrated MKE’s dose-dependent inhibition of B16F10 melanoma cell viability and attenuation of dermal inflammation via dual mechanisms: reducing tyrosinase and tyrosinase-related protein 1 expression and scavenging oxidative radicals.85 Recent studies indicated MFE effectively suppressed high mobility group box 1 protein expression and migration in ultraviolet B(UVB)-exposed HaCaT cells and epidermal models, thereby diminishing inflammatory cytokine release and cutaneous damage. Bioactive constituents from MFE identified included magnolin, eudesmin, and fargesin.86 Furthermore, in vitro and in vivo experiments demonstrated that fargesin possesses anti-melanoma properties partly mediated by suppression of the PKA/CREB and p38/MAPK pathways.87 Conversely, another study reported MFE stimulated melanin biosynthesis in B16-F1 and HMV-II cells, isolating seven TFLs, among which magnolin promoted melanin production via activation of melanogenic enzymes (tyrosinase, tyrosinase-related protein 1, tyrosinase-related protein) through PKA and p38 MAPK-dependent signaling pathways.88 This finding notably contrasts with earlier observations.
Osteoarticular Diseases
Magnolin suppressed IL-1α stimulation in murine models of rheumatoid arthritis, inhibiting synovial cell proliferation and reducing joint capsule inflammation.89,90 Additionally, MFE has therapeutic potential for osteoporosis, as demonstrated by inhibition of MMPs and cathepsin K expression, thereby reducing nuclear factor-κB ligand-mediated osteoclast differentiation and bone resorption.91 Magnolin, fargesin, aschantin, and lirirresinol B dimethyl ether inhibited MMP-9 and cathepsin K activity associated with bone destruction caused by breast cancer metastasis. These lignans significantly reduced osteoclast bone resorption, possibly linked to decreased expression of the osteolytic factor parathyroid hormone-related protein mRNA.92
Other Pharmacological Effects
Epimagnolin A can function as a competitive substrate for ATP-binding cassette subfamily B member 1, enhancing the sensitivity of chemotherapy drugs.93 Additionally, MF-MD dose-dependently suppress intracellular reactive oxygen species (ROS) generation. Bioactive compounds isolated from these extracts include magnolin, fargesin, epimagnolin, and eudesmin, with fargesin demonstrating the greatest hydroxyl radical scavenging activity.94 MKE also protects ovarian tissues by activating the Akt pathway and inhibiting oxidative stress.95 Moreover, magnolin mitigates contrast agent-induced kidney injury by reducing oxidative stress, suppressing ROS production, calcium protease I, and caspase 3 activity, and increasing Bcl-2 activity.96 Recently, TFLs derivatives have attracted significant research attention, demonstrating anti-inflammatory,89 anti-angiogenic, anti-rheumatic,97 and neuroprotective effects.98 In summary, TFLs exhibit pronounced advantages in mitigating oxidative stress and scavenging free radicals.
Pharmacokinetics
Chinese herbal medicines exhibit unique advantages as effective therapeutic and preventive agents. Currently, studies demonstrate that the pharmacokinetic parameters of magnolin, fargesin, epimagnolin, eudesmin, lirirresinol B dimethyl ether, and kobusin are dose-independent after oral or intravenous administration.99 Following oral administration, magnolin is rapidly absorbed with bioavailability (F) ranging from 54.3% to 76.4%.100 Pharmacokinetic analyses indicate magnolin follows a two-compartment model in rats, characterized by rapid absorption (Tmax ≈ 1.0 h) and slow elimination (t1/2β = 7.96 ± 2.57 h). The volume of distribution (Vd/F) was 0.012 ± 0.008.101 A comparative study in Sprague-Dawley rats evaluated fargesin pharmacokinetics after oral (50 mg/kg) and intravenous (20 mg/kg) administration. Oral administration showed a Tmax of 290 ± 24.49 min, Cmax of 464.38 ± 152.11 ng/mL, and an elimination half-life of 106.39 ± 84.93 min. Intravenous administration achieved peak concentrations earlier (Tmax = 20 ± 1.28 min, Cmax = 2049.0 ± 723.43 ng/mL) with a shorter half-life (80.75 ± 36.19 min). Notably, fargesin was detectable in major organs, including heart, liver, lungs, and kidneys.102 However, combined use of herbal medicines and therapeutic drugs may lead to adverse interactions. Numerous studies have focused on the human liver cytochrome P450 (CYP) enzymes (including CYPs 1A1, 1A2, 2A6, 2B6, 2C8, 2C9, 2C19, 2D6, 2E1, 2J2, 3A4, and 3A5) and uridine 5′-diphospho-glucuronosyltransferase (UGT) (including UGT1A1, UGT1A3, UGT1A4, UGT1A6, UGT1A9, and UGT2B7) to investigate TFLs metabolism in hepatocytes. Hepatic metabolism represents the principal elimination route for fargesin in both experimental animal models and human subjects. Fargesin acts as a competitive inhibitor of CYP2C9 and demonstrates time-dependent inhibitory effects on the activities of CYP2C8, CYP2C19, and CYP3A4.103,104 Similarly, aschantin inhibits the activities of CYP2C8, CYP2C9, CYP2C19, and CYP3A4.105 Conversely, magnolin exhibits a reduced likelihood of drug interactions owing to the involvement of several CYP enzymes in its metabolic pathways.106 Furthermore, eudesmin, magnolin, fargesin, epimagnolin A, and lirirresinol B dimethyl ether non-competitively inhibit UGT1A1 and competitively inhibit UGT1A3. Consequently, co-administration with drugs metabolized by these enzymes may increase the risk of drug interactions.107 Therefore, pharmacokinetic drug interactions mediated by TFLs through UGT1A1 and UGT1A3 inhibition should be evaluated in vivo.
Toxicology
Current toxicological evidence regarding TFLs remains limited. In vitro, cell viability was assessed using the MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay revealed that eudesmin exhibits concentration-dependent (2.5–80 μM) antitumor activity against A549 cells, with an IC50 of 18.3 μM.64 Using the cell Counting Kit-8 assay measured cell proliferation, magnolin reduced chondrocyte viability at concentrations ≥20 μM, with an IC50 of 58.05 μM.49 Acute toxicity and mucosal irritation tests in guinea pigs administered MF extract at 60-fold the standard dosage found no chronic toxicity or nasal mucosal irritation. Thus, these studies indicate MF is safe for nasal administration.108 However, TFLs metabolism in the body remains unclear, with insufficient quantitative toxicological data available. Further experimental research is required to evaluate the safety, toxicity, and mechanisms of action for these compounds.
TFLs Content Differentiation
Our analysis revealed significant variability in TFLs content, primarily influenced by plant genetic diversity and extraction methods. Previous studies demonstrated distinct TFLs compositions in different Magnolia varieties. Wolfgang Schuhly et al109 obtained dichloromethane extracts from M. liliiflora, which contained only trace amounts of magnolin. In contrast, magnolin and fargesin were undetectable in M. denudata. Subsequently, methanolic extract liquid chromatography-mass spectrometry was employed to isolate seven lignans from M. biondii. A variety of extraction methods, including solvent extraction,110 accelerated solvent extraction,111 and microwave-assisted extraction,112 facilitate qualitative and quantitative determination of MF components. Analytical methods include ultraviolet spectrophotometry, thin-layer chromatography, HPLC, gas chromatography-mass spectrometry (GC-MS), and liquid chromatography-mass spectrometry.2 Variations in extraction and measurement methodologies result in differing quantification of MF constituents (Table 2). This underscores the necessity of optimizing extraction techniques for efficient and practical natural product research. Due to extensive species diversity within the Magnoliaceae family, medicinal magnolia quality standardization is essential. Research indicates mitochondrial DNA heterogeneity and dynamic structural evolution arising from repetitive recombination within Magnolia species mitochondria.113 Controlling genetic diversity among MF varieties could effectively address this challenge. While TFLs are primary bioactive components in MF and also present in magnolia officinalis leaves and bark,114–116 these sources were not addressed in the present study.
 |
Table 2 Chemical Compounds of Magnoliae Flos: a Comparison of Different Extraction and Analytical Techniques |
Discussion and Perspective
The review covers biomolecules, pharmacology, pharmacokinetics, and toxicology, providing valuable insights into TFLs; however, several issues require further investigation. First,the effects of fargesin isolated from different crude extracts on the inhibition of the MRGPRX2 pathway are inconsistent. Multi-omics approaches (metabolomics-transcriptomics) are recommended to investigate mechanisms underlying the actions of crude extracts and isolated compounds. Second, fargesin demonstrates cardiovascular protective effects and may function as a potential β1-AR antagonist. Furthermore, fargesin demonstrates considerable potential in modulating glucose and lipid metabolism, positioning it as a promising candidate for hypoglycemic therapeutic development. However, further robust experimental and clinical data are required to substantiate this potential and elucidate the underlying mechanisms.Third, the effects of TFLs on melanogenesis are contradictory. Although their potential antioxidant and free radical-scavenging activities have been confirmed, further comparative studies among the seven identified TFLs are needed. Fourth, TFLs exhibit broad anti-tumor activity, and studies suggest magnolin may enhance chemotherapeutic drug sensitivity, indicating potential synergistic effects between TFLs and chemotherapy agents. Fifth, toxicological evidence remains insufficient, relying predominantly on in vitro studies. Comprehensive in vivo toxicological and safety evaluations are thus necessary. Sixth, lignan content and quality in MF vary significantly based on cultivar, geographical origin, and processing methods. To ensure experimental reproducibility, fingerprint profiles should be updated regularly, and a one-test-multiple-evaluation approach should be employed to confirm cultivar identity and origin. Seventh, some studies utilized MKE. As lignans are lipophilic compounds with limited water solubility, the use of water extracts may result in the loss of TFLs components and inclusion of water-soluble active constituents, such as alkaloids and flavonoid glycosides, potentially causing experimental bias. Therefore, future studies should focus on evaluating the bioactivity of isolated components from crude extracts through in vitro and in vivo experiments. Eighth, this review has limitations, as it summarizes only the seven TFLs known to be absorbed into the bloodstream, rather than covering all TFLs.
Conclusion
TFLs, as primary bioactive lignans from MF, exhibit potent pharmacological activities, including anti-inflammatory, anti-allergic, anticancer, cardiovascular protective effects, regulation of glucose and lipid metabolism, inhibition of melanogenesis, and treatment of osteoarticular diseases. These effects primarily involve modulation of the NF-κB, MAPK, PI3K–AKT, P53/P21, cAMP/PKA, Nrf2/HO-1, and LIF/Stat3 pathways. Based on existing evidence, MF represents a promising traditional Chinese medicinal herb with multiple pharmacological properties; however, it remains underappreciated clinically, with use predominantly limited to rhinitis, headache, and asthma. This review provides scientific evidence supporting the further development and broader application of TFLs from MF.
Acknowledgments
The authors acknowledge the BioRender (www.biorender.com), as portions of Figure 1, along with Figures 3 and 4 in this review were created using the BioRender platform.
Funding
This research was funded by the National Natural Science Foundation of China (NSFC) [grant numbers, 81173365].
Disclosure
The authors report no conflicts of interest in this work.
References
1. Tang W, Eisenbrand GMS. In: Chinese Drugs of Plant Origin. Berlin Heidelberg: Springer; 1992. 639–646. doi:10.1007/978-3-642-73739-8_84
2. Shen Y, Li CG, Zhou SF, Pang ECK, Story DF, Xue CCL. Chemistry and bioactivity of flos Magnoliae, a Chinese herb for rhinitis and sinusitis. Current Medicinal Chemistry. 2008;15(16):1616–1627. doi:10.2174/092986708784911515
3. The Pharmacopoeia Commission of People’s Republic of China. Pharmacopoeia of the People’s Republic of China. Beijing, China: Chemical Industry Press; 2005.
4. Zhou X, Chen C, Ye X, Song F, Fan G, Wu F. Analysis of lignans in Magnoliae Flos by turbulent flow chromatography with online solid-phase extraction and high-performance liquid chromatography with tandem mass spectrometry. J Sep Sci. 2016;39(7):1266–1272. doi:10.1002/jssc.201501167
5. Wang T. Study on Chemical Characteristics of Magnoliae Flos andProduction Quality of Its Extract [Master’s Thesis]. Chengdu University of Traditional Chinese Medicine; 2018.
6. Shen Y. Study on a Chinese herbal Flos Magnoliae (Xin-Yi) identification and pharmacological actions [PhD diss]. RMIT University; 2007.
7. Xu W-H, Zhao P, Wang M, Liang Q. Naturally occurring furofuran lignans: structural diversity and biological activities. Nat Prod Res. 2019;33(9):1357–1373. doi:10.1080/14786419.2018.1474467
8. Umezawa T. Diversity in lignan biosynthesis. Phytochem Rev. 2003;2(3):371–390. doi:10.1023/B:PHYT.0000045487.02836.32
9. Wang YX, Zhou L, Wang J, et al. Enantiomeric lignans with anti-β-amyloid aggregation activity from the twigs and leaves of pithecellobium clypearia benth. Bioorg Chem. 2018;77:579–585. doi:10.1016/j.bioorg.2018.02.001
10. JS K, JY K, Lee HJ, et al. Suppression of inducible nitric oxide synthase expression by furfuran lignans from flower buds of Magnolia fargesii in BV-2 microglial cells. Phytother Res. 2010;24(5):748–753. doi:10.1002/ptr.3028
11. Wang LD, Lin YX, Wang Y, et al. Analysis and verification of the classical formula Xinyi powder based on ancient and modern literature. China Pharm. 2021;32(18):2300–2304.
12. Wang B, Jin R, Liu J, et al. Study on the common characteristics of the compatibility of medicinal tastes in allergic rhinitis of different syndromes. Herald of Medicine. 2023;42(04):488–495.
13. Liu ZY, Ren YQ, Wang XG, et al. Study on the therapeutic mechanism of cangerzi san combined with GUOOMINJIAN in allergic rhinitis rats based on JNK signal pathway. Pharmacol Clini Chinese Mater Med. 2020;36(03):198–203. doi:10.13412/j.cnki.zyyl
14. Yang G. Quality control and pharmacokinetic study of M. biondii based on chemical characterization and improved pattern recognition approach [PhD dissertation]. The Second Military Medical University; 2017.
15. Yang X, SP Y, Yuan L, Liu L. Quality standard of Xinfang Biyan capsule. Chinese Traditional Patent Med. 2005;9:1033–1036.
16. Lu Y, He Z, Wang Q, et al. An advanced strategy for quality evaluation of Xinyi Biyan Pill by UPLC-DAD fingerprinting combined with multi-components UPLC-MS/MS analysis. J Pharm Biomed Anal. 2024;239:115858. doi:10.1016/j.jpba.2023.115858
17. Park CS, Kim TB, Lee JY, et al. Effects of add-on therapy with NDC-052, an extract from magnoliae flos, in adult asthmatic patients receiving inhaled corticosteroids. Korean J Intern Med. 2012;27(1):84–90. doi:10.3904/kjim.2012.27.1.84
18. Kim NJ, Oh SR, Lee HK, Lee HS. Simultaneous determination of magnolin and epimagnolin A in rat plasma by liquid chromatography with tandem mass spectrometry: application to pharmacokinetic study of a purified extract of the dried flower buds of Magnolia fargesii, NDC-052 in rats. J Pharm Biomed Anal. 2009;50(1):53–57. doi:10.1016/j.jpba.2009.03.036
19. Pan JX, Hensens OD, Zink DL, Chang MN, Hwang SB. Lignans with platelet activating factor antagonist activity from magnolia biondii. Phytochemistry. 1987;26(5):1377–1379. doi:10.1016/S0031-9422(00)81816-6
20. Chae SH, Kim PS, Cho JY, et al. Isolation and identification of inhibitory compounds on TNF-α production fromMagnolia fargesii. Arch Pharm Res. 1998;21(1):67–69. doi:10.1007/BF03216755
21. Cho JY, Yoo ES, Baik KU, Park MH. Eudesmin inhibits tumor necrosis factor-α production and T cell proliferation. Arch Pharm Res. 1999;22(4):348–353. doi:10.1007/BF02979056
22. Ahn K-S, Jung KY, Kim J-H, Oh SR, Lee H-K. Inhibitory activity of lignan components from the flower buds of magnoliae fargesii on the expression of cell adhesion molecules. Biol Pharm Bull. 2001;24(9):1085–1087. doi:10.1248/bpb.24.1085
23. Ham JR, Yun KW, Lee M-K. Anti-inflammatory and antioxidant in vitro activities of magnoliae flos ethanol extract. Prev Nutr Food Sci. 2021;26(4):485–491. doi:10.3746/pnf.2021.26.4.485
24. Gil T-Y, Jin B-R, Cha -Y-Y, An H-J. Magnoliae flos downregulated lipopolysaccharide-induced inflammatory responses via NF-κB/ERK-JNK MAPK/STAT3 pathways. Mediators Inflamm. 2022;2022:6281892. doi:10.1155/2022/6281892
25. Lee H-J, Lee S-J, Lee S-K, Choi B-K, Lee D-R. Magnolia kobus extract inhibits periodontitis-inducing mediators in porphyromonas gingivalis lipopolysaccharide-activated RAW 264.7 cells. Curr Issues Mol Biol. 2023;45(1):538–554. doi:10.3390/cimb45010036
26. Pham T-H, Kim M-S, Le M-Q, et al. Fargesin exerts anti-inflammatory effects in THP-1 monocytes by suppressing PKC-dependent AP-1 and NF-ĸB signaling. Phytomedicine. 2017;24:96–103. doi:10.1016/j.phymed.2016.11.014
27. Su Y, Xiong S, Lan H, Xu L, Wei X. Molecular mechanism underlying anti-inflammatory activities of lirioresinol B dimethyl ether through suppression of NF-κB and MAPK signaling in in vitro and in vivo models. Int Immunopharmacol. 2019;73:321–332. doi:10.1016/j.intimp.2019.05.020
28. Chun H-W, Kim S-J, Pham T-H, et al. Epimagnolin A inhibits IL-6 production by inhibiting p38/NF-κB and AP-1 signaling pathways in PMA-stimulated THP-1 cells. Environ Toxicol Int J. 2019;34(7):796–803. doi:10.1002/tox.22746
29. Yue B, Ren Y-J, Zhang -J-J, et al. Anti-inflammatory effects of fargesin on chemically induced inflammatory bowel disease in mice. Molecules. 2018;23(6):1380. doi:10.3390/molecules23061380
30. Zhang Y, Ma R, Wang J. Protective effects of fargesin on cadmium-induced lung injury through regulating aryl hydrocarbon receptor. J Biochem Mol Toxicol. 2022;36(11):e23197. doi:10.1002/jbt.23197
31. Lu J, Zhang H, Pan J, et al. Fargesin ameliorates osteoarthritis via macrophage reprogramming by downregulating MAPK and NF-κB pathways. Arthritis Res Ther. 2021;23(1):142. doi:10.1186/s13075-021-02512-z
32. Xu K, Gao Y, Yang L, Liu Y, Wang C. Magnolin exhibits anti-inflammatory effects on chondrocytes via the NF-κB pathway for attenuating anterior cruciate ligament transection-induced osteoarthritis. Connective Tissue Research. 2021;62(4):475–484. doi:10.1080/03008207.2020.1778679
33. Baek JA, Lee YD, Lee CB, et al. Extracts of Magnoliae flos inhibit inducible nitric oxide synthase via ERK in human respiratory epithelial cells. Nitric Oxide. 2009;20(2):122–128. doi:10.1016/j.niox.2008.10.003
34. Lee S-U, Ryu HW, Lee S, et al. Lignans isolated from flower buds of Magnolia Fargesii attenuate airway inflammation induced by cigarette smoke in vitro and in vivo. Front Pharmacol. 2018;9:970. doi:10.3389/fphar.2018.00970
35. Shehzad A, Rehmat S, Ul-Islam S, et al. Lirioresinol B dimethyl ether inhibits NF-κB and COX-2 and activates IκBα expression in CCl4-induced hepatic fibrosis. BMC Complement Med Ther. 2020;20(1):49. doi:10.1186/s12906-020-2839-3
36. Kim JS, Kim JY, Lee HJ, et al. Suppression of inducible nitric oxide synthase expression by furfuran lignans from flower buds of Magnolia fargesii in BV-2 microglial cells. Phytother Res. 2010;24(5):748–753. doi:10.1002/ptr.3028
37. Kim N, Chung G, Son S-R, et al. Magnolin inhibits paclitaxel-induced cold allodynia and ERK1/2 activation in mice. Plants. 2023;12(12):2283. doi:10.3390/plants12122283
38. Wang F, Wen H, Liu L, Aisa HA, Xin X. A pair of epimers of lignan alleviate neuroinflammatory effects by modulating iNOS/COX-2 and MAPK/NF-κB signaling pathways. Inflammation. 2024;48(1):361–371. doi:10.1007/s10753-024-02080-9
39. Zhang M, Song X, Liu S, et al. Magnolin inhibits intestinal epithelial cell apoptosis alleviating Crohn’s disease-like colitis by suppressing the PI3K/AKT signalling pathway. Int Immunopharmacol. 2024;134:112181. doi:10.1016/j.intimp.2024.112181
40. Yao T, Yao YY, Wang JZ, Jiang SM, Li LJ. Magnolin alleviated DSS-induced colitis by inhibiting ALOX5-mediated ferroptosis. Kaohsiung J Med Sci. 2024;40(4):360–373. doi:10.1002/kjm2.12806
41. Ince S, Demirel HH, Demirkapi EN, et al. Magnolin alleviates cyclophosphamide-induced oxidative stress, inflammation, and apoptosis via Nrf2/HO-1 signaling pathway. Toxicol Res. 2024;13(4):tfae129. doi:10.1093/toxres/tfae129
42. Stevens WW, Kraft M, Eisenbarth SC. Recent insights into the mechanisms of anaphylaxis. Curr Opin Immunol. 2023;81:102288. doi:10.1016/j.coi.2023.102288
43. Castells M, Madden M, Oskeritzian CA. Mast cells and mas-related g protein-coupled receptor X2: itching for novel pathophysiological insights to clinical relevance. Curr Allergy Asthma Rep. 2025;25(1):5. doi:10.1007/s11882-024-01183-5
44. Draber.P P, Halova I, Polakovicova I, Kawakami T. Signal transduction and chemotaxis in mast cells. European Journal of Pharmacology. 2016;778:11–23. doi:10.1016/j.ejphar.2015.02.057
45. Baran J, Sobiepanek A, Mazurkiewicz-Pisarek A, et al. Mast cells as a target—a comprehensive review of recent therapeutic approaches. Cells. 2023;12(8):1187. doi:10.3390/cells12081187
46. Kim H-M, Yi J-M, Lim K-S. Magnoliae flos inhibits mast cell-dependent immediate-type allergic reactions. Pharmacol Res. 1999;39(2):107–111. doi:10.1006/phrs.1998.0414
47. Zhang X, Qian F, Tan -J-J, et al. Bioassay-guided isolation of bisepoxylignans from the flower buds of Magnolia biondii Pamp and their antiallergic effects. RSC Adv. 2017;7(54):34236–34243. doi:10.1039/C7RA01476G
48. Lin Y, Xu J, Jia Q, et al. Cell membrane chromatography coupled online with LC-MS to screen anti-anaphylactoid components from Magnolia biondii Pamp. targeting on Mas-related G protein-coupled receptor X2. J Sep Sci. 2020;43(13):2571–2578. doi:10.1002/jssc.202000014
49. Shen Y, Pang EC, Xue CC, Zhao ZZ, Lin JG, Li CG. Inhibitions of mast cell-derived histamine release by different Flos Magnoliae species in rat peritoneal mast cells. Phytomedicine. 2008;15(10):808–814. doi:10.1016/j.phymed.2008.04.012
50. Ma P, Che D, Zhao T, et al. Magnolin inhibits IgE/Ag-induced allergy in vivo and in vitro. Int Immunopharmacol. 2019;76:105867. doi:10.1016/j.intimp.2019.105867
51. Kim HJ, Nam YR, Nam JH. Flos Magnoliae inhibits chloride secretion via ANO1 inhibition in Calu-3 cells. Am J Chin Med. 2018;46(5):1079–1092. doi:10.1142/S0192415X18500568
52. Hong PTL, Kim HJ, Kim WK, Nam JH. Flos magnoliae constituent fargesin has an anti-allergic effect via ORAI1 channel inhibition. Korean J Physiol Pharmacol. 2021;25(3):251–258. doi:10.4196/kjpp.2021.25.3.251
53. Lim H, Son KH, Bae KH, Hung TM, Kim YS, Kim HP. 5-lipoxygenase-inhibitory constituents from Schizandra fructus and Magnolia flos. Phytother Res. 2009;23(10):1489–1492. doi:10.1002/ptr.2783
54. Nguyen TTM, Lee H-S, Nguyen TT, et al. Four new Lignans and IL-2 Inhibitors from Magnoliae Flos. Chem Pharm Bull. 2017;65(9):840–847. doi:10.1248/cpb.c17-00314
55. Farber E, Rubin H. Cellular adaptation in the origin and development of cancer. Cancer Res. 1991;51(11):2751–2761.
56. Bhuia MS, Wilairatana P, Chowdhury R, et al. Anticancer potentials of the lignan Magnolin: a systematic review. Molecules. 2023;28(9):3671. doi:10.3390/molecules28093671
57. Lee C-J, Lee HS, Ryu HW, et al. Targeting of magnolin on ERKs inhibits Ras/ERKs/RSK2-signaling-mediated neoplastic cell transformation. Carcinogenesis. 2014;35(2):432–441. doi:10.1093/carcin/bgt306
58. Lee C-J, Lee M-H, Yoo S-M, et al. Magnolin inhibits cell migration and invasion by targeting the ERKs/RSK2 signaling pathway. BMC Cancer. 2015;15(1):576. doi:10.1186/s12885-015-1580-7
59. Jiang -L-L, Sun B-R, Zheng C, Yang G-L. The antitumour effects of eudesmin on lung cancer by inducing apoptosis via mitochondria-mediated pathway in the tumour cells. Pharm Biol. 2017;55(1):2259–2263. doi:10.1080/13880209.2017.1401647
60. Yoo S-M, Lee C-J, Kang HC, et al. Epimagnolin targeting on an active pocket of mammalian target of rapamycin suppressed cell transformation and colony growth of lung cancer cells. Mol, Carcinog. 2019;58(7):1221–1233. doi:10.1002/mc.23005
61. Guo Z, Tang Y, Wang S, et al. Natural product fargesin interferes with H3 histone lactylation via targeting PKM2 to inhibit non-small cell lung cancer tumorigenesis. Biofactors. 2024;50(3):592–607. doi:10.1002/biof.2031
62. Lee C-J, Jang J-H, Lee J-Y, et al. Aschantin targeting on the kinase domain of mammalian target of rapamycin suppresses epidermal growth factor-induced neoplastic cell transformation. Carcinogenesis. 2015;36(10):1223–1234. doi:10.1093/carcin/bgv113
63. Wang J, Zhang S, Huang K, Shi L, Zhang Q. Magnolin Inhibits proliferation and invasion of breast cancer MDA-MB-231 cells by targeting the ERK1/2 signaling pathway. Chem Pharm Bull. 2020;68(5):421–427. doi:10.1248/cpb.c19-00820
64. Huang Y, Zou X, Zhang X, et al. Magnolin inhibits prostate cancer cell growth in vitro and in vivo. Biomed Pharmacother. 2017;87:714–720. doi:10.1016/j.biopha.2017.01.010
65. Yu M, Li Y, Li M, Lu D. Eudesmin exerts antitumor effects by down-regulating EZH2 expression in nasopharyngeal carcinoma cells. Chem Biol Interact. 2019;307:51–57. doi:10.1016/j.cbi.2019.04.028
66. Yu H, Yin S, Zhou S, et al. Magnolin promotes autophagy and cell cycle arrest via blocking LIF/Stat3/Mcl-1 axis in human colorectal cancers. Cell Death Dis. 2018;9(6):702. doi:10.1038/s41419-018-0660-4
67. Lee G-E, Lee C-J, An H-J, et al. Fargesin inhibits EGF-induced cell transformation and colon cancer cell growth by suppression of CDK2/Cyclin E signaling pathway. Int J Mol Sci. 2021;22(4):2073. doi:10.3390/ijms22042073
68. Song J-H, Lee C-J, An H-J, et al. Magnolin targeting of ERK1/2 inhibits cell proliferation and colony growth by induction of cellular senescence in ovarian cancer cells. Mol, Carcinog. 2019;58(1):88–101. doi:10.1002/mc.22909
69. Lin C-L, Ying T-H, Yang S-F, Lin C-L, Chiou H-L, Hsieh Y-H. Magnolin targeting of the JNK/Sp1/MMP15 signaling axis suppresses cervical cancer microenvironment and metastasis via microbiota modulation. Cancer Lett. 2024;583:216584. doi:10.1016/j.canlet.2023.216584
70. Wang W, Xiao Y, Li S, et al. Synergistic activity of magnolin combined with B-RAF inhibitor SB590885 in hepatocellular carcinoma cells via targeting PI3K-AKT/mTOR and ERK MAPK pathway. Am J Transl Res. 2019;11(6):3816–3824.
71. Yap TA, Garrett MD, Walton MI, Raynaud F, de Bono JS, Workman P. Targeting the PI3K–AKT–mTOR pathway: progress, pitfalls, and promises. Curr Opin Pharmacol. 2008;8(4):393–412. doi:10.1016/j.coph.2008.08.004
72. Guertin DA, Sabatini DM. Defining the role of mTOR in cancer. Cancer Cell. 2007;12(1):9–22. doi:10.1016/j.ccr.2007.05.008
73. Roberts PJ, Der CJ. Targeting the Raf-MEK-ERK mitogen-activated protein kinase cascade for the treatment of cancer. Oncogene. 2007;26(22):3291–3310. doi:10.1038/sj.onc.1210422
74. Yoo S-M, Cho SJ, Cho -Y-Y. Molecular targeting of ERKs/RSK2 signaling axis in cancer prevention. J Cancer Prev. 2015;20(3):165–171. doi:10.15430/JCP.2015.20.3.165
75. Yip HYK, Papa A. Signaling pathways in cancer: therapeutic targets, combinatorial treatments, and new developments. Cells. 2021;10(3):659. doi:10.3390/cells10030659
76. Shamloo B, Usluer S. P21 in cancer research. Cancers. 2019;11(8):1178. doi:10.3390/cancers11081178
77. Wang X, Cheng Y, Xue H, Yue Y, Zhang W, Li X. Fargesin as a potential β1 adrenergic receptor antagonist protects the hearts against ischemia/reperfusion injury in rats via attenuating oxidative stress and apoptosis. Fitoterapia. 2015;105:16–25. doi:10.1016/j.fitote.2015.05.016
78. Sha S, Xu D, Wang Y, Zhao W, Li X. Antihypertensive effects of fargesin in vitro and in vivo via attenuating oxidative stress and promoting nitric oxide release. Can J Physiol Pharmacol. 2016;94(8):900–906. doi:10.1139/cjpp-2015-0615
79. Wang G, Gao J-H, He L-H, et al. Fargesin alleviates atherosclerosis by promoting reverse cholesterol transport and reducing inflammatory response. Biochim Biophys Acta Mol Cell Biol Lipids. 2020;1865(5):158633. doi:10.1016/j.bbalip.2020.158633
80. Kim E-K, Song M-Y, Kim I-S, et al. Beneficial effect of Flos magnoliae extract on multiple low dose streptozotocin-induced type 1 diabetes development and cytokine-induced beta-cell damage. Int J Mol Med. 2008;22(4):481–488.
81. Choi -S-S, Cha B-Y, Choi B-K, et al. Fargesin, a component of Flos Magnoliae, stimulates glucose uptake in L6 myotubes. J Nat Med. 2013;67(2):320–326. doi:10.1007/s11418-012-0685-4
82. Lee Y-S, Cha B-Y, Choi -S-S, et al. Fargesin improves lipid and glucose metabolism in 3T3-L1 adipocytes and high-fat diet-induced obese mice. Biofactors. 2012;38(4):300–308. doi:10.1002/biof.1022
83. Kong C-S, Lee JI, Kim J-A, Seo Y. In vitro evaluation on the antiobesity effect of lignans from the flower buds of Magnolia denudata. J Agric Food Chem. 2011;59(10):5665–5670. doi:10.1021/jf200230s
84. Nam KH, Yi SA, Lee J, et al. Eudesmin impairs adipogenic differentiation via inhibition of S6K1 signaling pathway. Biochem Biophys Res Commun. 2018;505(4):1148–1153. doi:10.1016/j.bbrc.2018.09.188
85. Huang H-C, Hsieh W-Y, Niu Y-L, Chang T-M. Inhibition of melanogenesis and antioxidant properties of Magnolia grandiflora L. flower extract. BMC Complement Altern Med. 2012;12(1):72. doi:10.1186/1472-6882-12-72
86. Huang F, Liu Q, Lu Y. Magnolia biondii flower extract attenuates UVB -induced skin damage through high-mobility group box protein B1. Int J Cosmet Sci. 2024;46(5):775–785. doi:10.1111/ics.12959
87. Fu T, Chai B, Shi Y, Dang Y, Ye X. Fargesin inhibits melanin synthesis in murine malignant and immortalized melanocytes by regulating PKA/CREB and P38/MAPK signaling pathways. J Dermatol Sci. 2019;94(1):213–219. doi:10.1016/j.jdermsci.2019.03.004
88. Uto T, Tung NH, Ohta T, Shoyama Y. (+)-magnolin enhances melanogenesis in melanoma cells and three-dimensional human skin equivalent; involvement of PKA and p38 MAPK signaling pathways. Planta Med. 2022;88(13):1199–1208. doi:10.1055/a-1740-7325
89. Kimura M, Suzuki J, Yamada T, et al. Anti-inflammatory effect of neolignans newly isolated from the crude Drug “Shin-i” (Flos Magnoliae). Planta med. 1985;51(04):291–293. doi:10.1055/s-2007-969493
90. Kobayashi S, Kobayashi H, Matsuno H, Kimura I, Kimura M. Inhibitory effects of anti-rheumatic drugs containing magnosalin, a compound from `Shin-i’ (Flos magnoliae), on the proliferation of synovial cells in rheumatoid arthritis models. Immunopharmacology. 1998;39(2):139–147. doi:10.1016/s0162-3109(98)00004-6
91. Jun AY, Kim H-J, Park -K-K, et al. Extract of Magnoliae Flos inhibits ovariectomy-induced osteoporosis by blocking osteoclastogenesis and reducing osteoclast-mediated bone resorption. Fitoterapia. 2012;83(8):1523–1531. doi:10.1016/j.fitote.2012.08.020
92. Jun AY, Kim H-J, Park -K-K, et al. Tetrahydrofurofuran-type lignans inhibit breast cancer-mediated bone destruction by blocking the vicious cycle between cancer cells, osteoblasts and osteoclasts. Invest New Drugs. 2014;32(1):1–13. doi:10.1007/s10637-013-9969-0
93. Mitani Y, Satake K, Tsukamoto M, et al. Epimagnolin A, a tetrahydrofurofuranoid lignan from Magnolia fargesii, reverses ABCB1-mediated drug resistance. Phytomedicine. 2018;51:112–119. doi:10.1016/j.phymed.2018.06.030
94. Seo Y. Antioxidant activity of the chemical constituents from the flower buds of Magnolia denudata. Biotechnol Bioproc E. 2010;15(3):400–406. doi:10.1007/s12257-009-0219-4
95. Kim MR, Kim D-I, Park SY, Kang HJ, Park S-D, Lee J-H. The protective role of Magnoliae Flos in preventing ovotoxicity and managing ovarian function: an in vitro and in vivo study. Int J Mol Sci. 2024;25(12):6456. doi:10.3390/ijms25126456
96. Wang F, Zhang G, Zhou Y, et al. Magnolin protects against contrast-induced nephropathy in rats via antioxidation and antiapoptosis. Oxid Med Cell Longev. 2014;2014:203458. doi:10.1155/2014/203458
97. Tanaka K, Konno Y, Kuraishi Y, Kimura I, Suzuki T, Kiniwa M. Synthesis of a magnosalin derivative, 4-(3,4,5-trimethoxyphenyl)-6-(2,4,5-trimethoxyphenyl)-2-diethylaminopyrimidine, and the anti-angiogenic and anti-rheumatic effect on mice by oral administration. Bioorg Med Chem Lett. 2002;12(4):623–627. doi:10.1016/s0960-894x(01)00810-1
98. Ma Q, Wei R, Zhang X, Sang Z, Dong J. Structural elucidation and neuroprotective activities of lignans from the flower buds of Magnolia biondii Pamp. Naturforsch C J Biosci. 2020;76(3–4):147–152. doi:10.1515/znc-2020-0100
99. Jeong JH, Kim DK, Ji HY, Oh SR, Lee HK, Lee HS. Liquid chromatography-atmospheric pressure chemical ionization tandem mass spectrometry for the simultaneous determination of dimethoxyaschantin, dimethylliroresinol, dimethylpinoresinol, epimagnolin A, fargesin and magnolin in rat plasma. Biomed Chromatogr. 2011;25(8):879–889. doi:10.1002/bmc.1538
100. Kim NJ, Song WY, Yoo SD, Oh SR, Lee HK, Lee HS. Pharmacokinetics of magnolin in rats. Arch Pharm Res. 2010;33(6):933–938. doi:10.1007/s12272-010-0617-3
101. Yu Y, Yuan HY, Zeng JP. Pharmacokinetic study of Magnolin in rats. China Pharm. 2015;26(25):3522–3524.
102. Zhang ZH. The study of pharmacokinetics and tissue distributionof fargesin and its preparation [Master’s Thesis]. The Fourth Military Medical University; 2016.
103. Kim J-H, Kwon -S-S, Jeong H-U, Lee HS. Inhibitory effects of dimethyllirioresinol, epimagnolin a, eudesmin, fargesin, and magnolin on cytochrome P450 enzyme activities in human liver microsomes. Int J Mol Sci. 2017;18(5):952. doi:10.3390/ijms18050952
104. Lee MS, Park EJ, Cho -Y-Y, Lee JY, Kang HC, Lee HS. Comparative metabolism of fargesin in human, dog, monkey, mouse, and rat hepatocytes. Toxicol Res. 2024;40(1):125–137. doi:10.1007/s43188-023-00211-2
105. Kwon -S-S, Kim J-H, Jeong H-U, Cho YY, Oh S-R, Lee HS. Inhibitory effects of aschantin on cytochrome P450 and uridine 5′-diphospho-glucuronosyltransferase enzyme activities in human liver microsomes. Molecules. 2016;21(5):554. doi:10.3390/molecules21050554
106. Kim DK, Liu K-H, Jeong JH, et al. In vitro metabolism of magnolin and characterization of cytochrome P450 enzymes responsible for its metabolism in human liver microsomes. Xenobiotica. 2011;41(5):358–371. doi:10.3109/00498254.2010.549968
107. Park R, Park EJ, Cho -Y-Y, et al. Tetrahydrofurofuranoid Lignans, Eudesmin, Fargesin, Epimagnolin A, Magnolin, and Yangambin Inhibit UDP-Glucuronosyltransferase 1A1 and 1A3 Activities in Human Liver Microsomes. Pharmaceutics. 2021;13(2):187. doi:10.3390/pharmaceutics13020187
108. Lv LL, Qian XL, Sun R. Experimental study on safety in guinea pig about the different components of flos magnolia nose drop. Chin J Pharmacovig. 2013;10(02):74–76.
109. Chühly W, Skarbina J, Kunert O, Nandi OI, Bauer R. Chemical characterization of Magnolia biondii (Flos Magnoliae, Xin Yi). Nat Prod Commun. 2009;4(2):231–234.
110. Gao X, Shen Y, Yang L, Shu L, Li G, Hu Q-F. 8-O-4’-Neolignans from flower buds of Magnolia fargesii and their biological activities. J Braz Chem Soc. 2012;23(7):1274–1279. doi:10.1590/S0103-50532012000700011
111. Zhao W, Zhou T, Fan G, Chai Y, Wu Y. Isolation and purification of lignans from Magnolia biondii Pamp by isocratic reversed-phase two-dimensional liquid chromatography following microwave-assisted extraction. J Sep Sci. 2007;30(15):2370–2381. doi:10.1002/jssc.200700098
112. LU YH, Gao Y, Wang ZT, Liu JQ. A Benzofuranoid Neolignan from Magnolia biondii Pamp. J Chin Pharmace Sci. 2005;14(3):137.
113. Dong S, Chen L, Liu Y, et al. The draft mitochondrial genome of Magnolia biondii and mitochondrial phylogenomics of angiosperms. PLoS One. 2020;15(4):e0231020. doi:10.1371/journal.pone.0231020
114. Du J, Wang M-L, Chen R-Y, Yu D-Q. Chemical constituents from the leaves of Magnolia denudata. J Asian Nat Prod Res. 2001;3(4):313–319. doi:10.1080/10286020108040371
115. Seo S-M, Lee H-J, Lee O-K, et al. Furofuran lignans from the bark of Magnolia kobus. Chem Nat Compd. 2008;44(4):419–423. doi:10.1007/s10600-008-9093-0
116. Lee H-J, Seo S-M, Lee O-K, et al. Lignans from the bark of Magnolia kobus. Helvetica Chim Acta. 2008;91(12):2361–2366. doi:10.1002/hlca.200890256
117. Li J, Wen J, Tang G, et al. Development of a comprehensive quality control method for the quantitative analysis of volatiles and lignans in Magnolia biondii pamp. by near infrared spectroscopy. Spectrochim Acta A Mol Biomol Spectrosc. 2020;230:118080. doi:10.1016/j.saa.2020.118080
118. Lee DH, Kwon SY, Woo MH, Lee JH, Son KH. Phytochemical studies on Magnoliae Flos (I) isolation of lignans from the flower buds of Magnolia biondii. Nat Product Sci. 2013;19(2):160–165.
119. Feng W-S, He Y-H, Zheng X-K, et al. Lignans from flower buds of Magnolia biondii. Zhongguo Zhong Yao Za Zhi. 2018;43(5):970–976. doi:10.19540/j.cnki.cjcmm.2018.0028
120. Li J, Tanaka M, Kurasawa K, Ikeda T, Nohara T. Lignan and neolignan derivatives from Magnolia denudata. Chem Pharm Bull. 2005;53(2):235–237. doi:10.1248/cpb.53.235
121. Li J, Mei ZY, Luo HW, Huang XJ, Yang GZ. Study on the Aromatic ditetrahydrofuran constituents from magnolia denudata desr. J South-Central Minzu Univ. 2015;34(03):37–40.
122. Mei ZY, Luo HW, Zou DJ, et al. Study on the furofurans lignan from the buds of magnolia denudata desr. J Central China Normal Univ. 2014;48(4):525–527,543. doi:10.19603/j.cnki.1000-1190.2014.04.014
123. Kim JY, Lim HJ, Lee da Y, et al. In vitro anti-inflammatory activity of lignans isolated from Magnolia fargesii. Bioorg Med Chem Lett. 2009;19(3):937–940. doi:10.1016/j.bmcl.2008.11.103
124. Li J, Tanaka M, Kurasawa K, Ikeda T, Nohara T. Studies of the chemical constituents of the flower buds of Magnolia kobus and M. salicifolia. J Nat Med. 2007;61(2):222–223. doi:10.1007/s11418-006-0110-y
125. Khumploy P. Anti-Oxidant and Anti-Inflammatory Compounds From Magnolia Liliifera (L.) Baill. Flower and Dendrobium Signatum Rchb.f. Chulalongkorn University Theses and Dissertations (Chula ETD); 2020. doi:10.58837/CHULA.THE.2020.91
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.