Back to Journals » Pharmacogenomics and Personalized Medicine » Volume 14
PFN1 Gene Polymorphisms and the Bone Mineral Density Response to Alendronate Therapy in Postmenopausal Chinese Women with Low Bone Mass
Authors Zhao J, Liu L, Lv S, Wang C, Yue H, Zhang Z
Received 21 October 2021
Accepted for publication 15 December 2021
Published 23 December 2021 Volume 2021:14 Pages 1669—1678
DOI https://doi.org/10.2147/PGPM.S344818
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Martin H Bluth
Jiao Zhao,* Li Liu,* Shanshan Lv, Chun Wang, Hua Yue, Zhenlin Zhang
Shanghai Clinical Research Center of Bone Disease, Department of Osteoporosis and Bone Disease, Shanghai Jiaotong University Affiliated Sixth People’s Hospital, Shanghai, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Zhenlin Zhang; Hua Yue
Shanghai Clinical Research Center of Bone Disease, Department of Osteoporosis and Bone Disease, Shanghai Jiaotong University Affiliated Sixth People’s Hospital, Yishan Road 600, Shanghai, 200233, People’s Republic of China
Tel +86 21 64369181
Fax +86 21 64701361
Email [email protected]; [email protected]
Purpose: Alendronate is a widely used anti-osteoporotic drug. PFN1 gene is a newly identified early-onset Paget’s disease pathogenic gene. The purpose of this study is to study whether the genetic variations in this gene affect the clinical efficacy of alendronate in postmenopausal Chinese women with low bone mass.
Patients and Methods: Seven single nucleotide polymorphisms in PFN1 gene were genotyped. A total of 500 postmenopausal women with osteoporosis or osteopenia were included. All participants were treated with weekly alendronate 70 mg for 12 months. A total of 466 subjects completed the follow-up. Bone mineral density (BMD) of lumbar spine, femoral neck and total hip were measured at baseline and after treatment.
Results: After 12 months of treatment, the BMD of lumbar spine, femoral neck and total hip all increased significantly (all P < 0.001), with an average increase of 4.72 ± 5.31%, 2.08 ± 4.45%, and 2.42 ± 3.46%, respectively. At baseline, there were no significant differences in BMD at lumbar spine, femoral neck and total hip between different genotype groups (P > 0.05). We failed to identify any significant association between the genotypes or haplotypes of PFN1 and the BMD response to alendronate therapy.
Conclusion: Genetic polymorphisms of PFN1 may not be a major contributor to the therapeutic response to alendronate treatment in Chinese women with low bone mass.
Keywords: alendronate, bone mineral density, osteoporosis, PFN1 gene, single-nucleotide polymorphism
Introduction
Osteoporosis is a common skeletal disease characterized by low bone mass, reduced bone strength, deteriorated bone microarchitecture. Genetic factors contribute 60–80% of the variance in bone mineral density (BMD).1–3 With the development of the aging population, the incidence of osteoporosis and osteoporotic fracture has been increasing. An analysis of National Health and Nutrition Examination Survey 2005–2008 data revealed that 19% of older men and 30% of older women in the United States require antiosteoporosis therapy.4 With regard to osteoporosis in China, the rate of hip fractures in China is progressively increased approximately 10% per year from 2002 to 2006.5 Our study in 2019 indicated that the prevalence of vertebral fractures in the elderly in Shanghai was 17.2%, that of men was 17.0%, and that of women was 17.3%.6
Alendronate is a potent and specific inhibitor of osteoclast-mediated bone resorption. Because of its beneficial effects in increasing bone mass and reducing the risk of fracture, it is widely used as a first-line drug in postmenopausal women with osteoporosis in Europe, the Americas and Asia.7–12 Studies have demonstrated that treatment with 70 mg of alendronate once weekly decreases the risk of vertebral fractures by 50% and the risk of peripheral fractures by 20–30%.13,14 Nevertheless, the efficacy and safety of alendronate vary among different patients, with approximately 5–10% of cases having poor or no clinical response.15 The identification of the factors that influence the response to alendronate therapy can help optimize drug therapy. Variations in the human genome are considered the key cause of variable drug responses and adverse reaction development.16,17 The COL1A1 gene18 and VDR gene19–21 have been shown to be involved in the response to bisphosphonate therapy. Our previous research proved that MVK and FDFT1 polymorphisms are genetic determinants for response to alendronate therapy in postmenopausal Chinese women.22 We also clarified that there is no association between polymorphisms in OPG, RANKL and RANK gene and BMD response to alendronate.23 Recently, we demonstrated that the common variation of SOST gene is associated with the response to alendronate treatment in postmenopausal women with low BMD.24 However, to date, pharmacogenomics reports on alendronate are still scattered.
The PFN1 gene, located at 17p13.3, encodes the highly evolutionarily conserved protein Profilin 1. Profilin 1 is a small actin monomer-binding protein which is considered to be an essential control element for actin polymerization and cell migration.25,26 Profilin 1 is ubiquitously expressed in all cell types and in organs and tissues throughout the body, and is involved in the control of a variety of cell functions.25,27–31 We and two other research teams have reported that mutations in the PFN1 gene could lead to early-onset Paget’s disease with or without giant cell tumors.32–34 This finding indicates that PFN1 mutations can cause the imbalance of skeletal homeostasis and changes in bone mass and bone turnover status. In addition, Shirakawa et al have demonstrated in animal experiments that Pfn1 plays critical role in inhibiting osteoclast motility and bone resorption, thereby affecting the skeletal homeostasis of mice.35 So, we believe that PFN1 gene may be a key gene regulating the proliferation, differentiation and activity of osteoclasts.
We hypothesized that genetic polymorphisms in PFN1 gene is good candidate to explain the variable responses to alendronate treatment. Therefore, we conducted this study to explore the association between PFN1 gene variations and the BMD response to alendronate therapy in postmenopausal Chinese women with low mass.
Materials and Methods
Subjects
All participants were recruited by the department of Osteoporosis and Bone Disease in Shanghai Jiaotong University Affiliated Sixth People’s Hospital. Five hundred women were enrolled in this study. All study subjects were postmenopausal Han Chinese women with primary osteoporosis or osteopenia, without spontaneous menses for at least 1 year. All participants were treated with 70 mg of alendronate (Merck, Fosamax, Hangzhou, China) once weekly, 600 mg of calcium and 125 IU of vitamin D daily for 12 months. The study protocol was approved by the Committee of the Ethics of Human Research in the Shanghai Jiaotong University Affiliated Sixth People’s Hospital and was conducted in accordance with the Declaration of Helsinki. All study participants provided written informed consent.
The inclusion criteria were as follows: natural menopause after 40 years of age and a BMD of at least 1.0 SD below the peak mean bone density of healthy young women (−1.0 T-score) at the posterior–anterior L1–4, femoral neck, or total hip. The diagnosis of osteoporosis was based on L1-4, femoral neck or total hip with a T-score less than −2.5, or a history of fragility fractures of the spine or hip. Among women with osteopenia, the decisions of treatment were made depended on the high risk of osteoporotic fractures. We evaluated the high risk of fracture according to the history of low trauma fracture at humerus or radius, the 10-year probability of hip fracture ≥3% or of a major osteoporotic fracture (clinical spine, hip, forearm and humerus fracture) ≥20% by FRAX®.36
The exclusion criteria24 were as follows: a history of (1) chronic renal disease manifested by an endogenous creatinine clearance of <35 mL/min; (2) acute inflammation of the gastrointestinal tract (eg, gastritis and ulcerations); (3) esophagitis or certain malformations and malfunctions of the esophagus (eg, strictures and achalasia); (4) proton-pump inhibitor usage along with alendronate treatment; (5) inability to stand, walk or sit for 30 min after oral administration of alendronate; (6) hypersensitivity to alendronate or another ingredient in the therapeutic compound; (7) hypocalcemia (serum calcium (Ca) < 2.08 mmol/l) or hypophosphatemia (serum phosphorus (P) < 0.80 mmol/l); (8) increased serum parathyroid hormone (PTH) levels (normal values: 15–65 pg/mL); (9) serious residual effects of cerebral vascular disease; (10) diabetes mellitus, except for adult asymptomatic hyperglycemia controlled by diet; (11) chronic liver disease or alcoholism; (12) 12 weeks of corticosteroid therapy at pharmacologic levels; (13) 6 months of treatment with anticonvulsant therapy; (14) evidence of other metabolic or inherited bone diseases (eg, hyperparathyroidism or hypoparathyroidism, Paget’s disease, osteomalacia or osteogenesis imperfecta); (15) rheumatoid arthritis or collagen disease; (16) significant disease of any endocrine organ that would affect bone mass (eg, Cushing’s syndrome or hyperthyroidism); (17) any neurological or musculoskeletal condition that would be a nongenetic cause of low bone mass; (18) a body mass index (BMI) of <18 kg/m2 or >30 kg/m2; and (19) any previous treatment with bisphosphonate, sodium fluoride, calcitonin, a selective estrogen receptor modulator, strontium ranelate, or the recombinant form of PTH or current use of hormone replacement therapy.
BMD Measurements
Lunar prodigy dual-energy X-ray absorptiometry densitometer (DXA; GE Healthcare, Madison, WI, USA) was used to measure the BMD of lumbar spine (L1–4), left femoral neck and total hip at baseline and after 12 months of treatment, respectively. The BMD of the right hip was measured only in patients with a history of left hip fracture or surgery. The Prodigy encore software (ver. 6.70, standard-array mode; GE Healthcare) were used for data analysis. The machines were calibrated daily and coefficient of variance (CV) values for BMD of the L1–4, femoral neck and total hip were 1.39%, 2.22% and 0.70%, respectively.37 The long-term reproducibility of our DXA instrument during the study based on weekly repeated phantom measurements was 0.45%. Besides, we chose the least significant change (LSC) to estimate the effectiveness of treatment. And the LSC in BMD at L1–4, femoral neck and the total hip were 3.85%, 6.15% and 1.94%.37 Weight and height were measured using a calibrated balance beam scale and a calibrated stadiometer. The BMI was defined as weight/height2 in kg/m2.
PFN1 Genotyping
Seven tagSNPs in PFN1 gene (rs13204, rs78224458, rs238243, rs238242, rs113460962, rs4790714 and rs117337116) were selected from dbSNP (http://www.ncbi.nlm.nih.gov/snp/) and HapMap (http://hapmap.ncbi.nlm.nih.gov/) based on the following criteria: (1) minor allele frequency (MAF) higher than 5%; (2) pairwise linkage disequilibrium (LD) exceeding the threshold of 0.8 (r2 > 0.8) (Table 1). Genomic DNA was isolated from peripheral blood leukocytes via isopropanol-precipitating method. Genotyping was performed using the ABI PRISM SNaPshot multiplex kit (Applied Biosystems), an Mx3000p real-time PCR system (Stratagene), and GeneMapper 4.1 (Applied Biosystems).
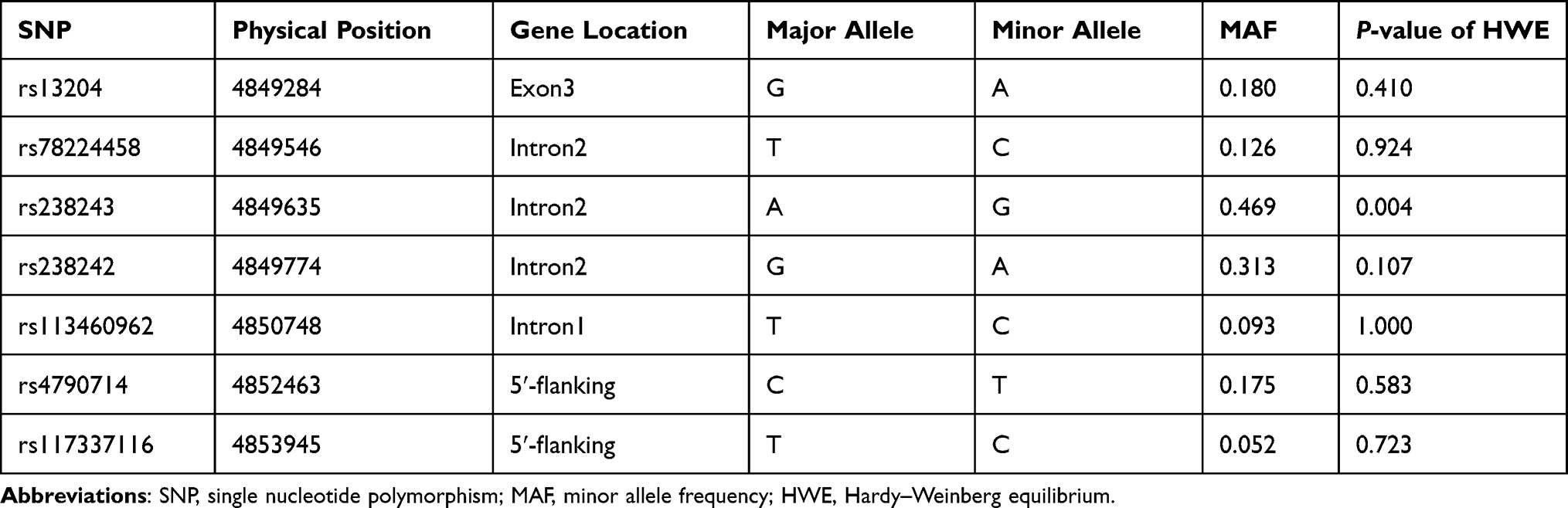 |
Table 1 Information on the 7 SNPs in This Study |
Statistical Analyses
Hardy–Weinberg equilibrium (HWE) was tested for each SNP using the χ2 test. The linkage disequilibrium block structure was assessed using the Haploview 4.2. PLINK (http://pngu.mgh.harvard.edu/purcell/plink/) was utilized in quality control filtering and haplotype association test. SNPs with call rates <75% and HWE tests with P-values <0.01 were excluded from further analysis. Descriptive statistics were given as the mean ± SD for normally distributed data and as the median and interquartile range for the non-normally distributed data. The BMD value at L1–4, femoral neck and total hip before and after alendronate treatment were compared by using the paired t-test. SPSS 22.0 (SPSS, Chicago, IL, USA) was used to analyzed these data. The response to alendronate treatment is estimated by the % change in BMD, defined as (12-months BMD – baseline BMD)/baseline BMD × 100%. The baseline and % change after 12 months treatment in BMD among different genotype groups were tested using PLINK in the ADD linear model. The differences in BMD at baseline and % change after 12 months treatment among haplotype groups were analyzed using PLINK in the GLMs linear model. According to the LSC in BMD, the participants were divided into two groups: responder(s) and non-responder(s). The association between genotype groups and the opportunity of being a responder after treatment was analyzed using PLINK in the ADD logistic regression model. The association between the haplotype groups and the opportunity of being a responder was analyzed using PLINK in the GLMs logistic regression model. All data were adjusted by the age and BMI covariates. P-values <0.05 were defined as statistically significant. The Bonferroni correction was used to adjust for multiple testing. For SNP analysis, P-value thresholds for statistical significance were 0.008; for haplotype analysis, P-value thresholds for statistical significance were 0.025.
Results
Basic Characteristics of Study Subjects
Among the 500 subjects participating in the study, 466 subjects completed 12 months of alendronate treatment and had BMD measurement at baseline and 12-months treatment, respectively. The average baseline age, height, weight and BMI were 64.88 ± 8.11 years, 153.76 ± 6.38 cm, 54.29 ± 8.26 kg and 22.96 ± 3.08 kg/m2, respectively. The mean BMD of the L1–4, femoral neck and total hip was 0.814 ± 0.149 g/cm2, 0.673 ± 0.104 g/cm2, 0.717 ± 0.107 g/cm2, respectively. After 12 months of treatment, there were no significant difference between baseline and follow-up height, weight or BMI. As expected with alendronate treatment, the BMD of L1–4, femoral neck and total hip all increased significantly (all P < 0.001), with average increases of 4.72 ± 5.31%, 2.08 ± 4.45%, and 2.42 ± 3.46%, respectively (Table 2).
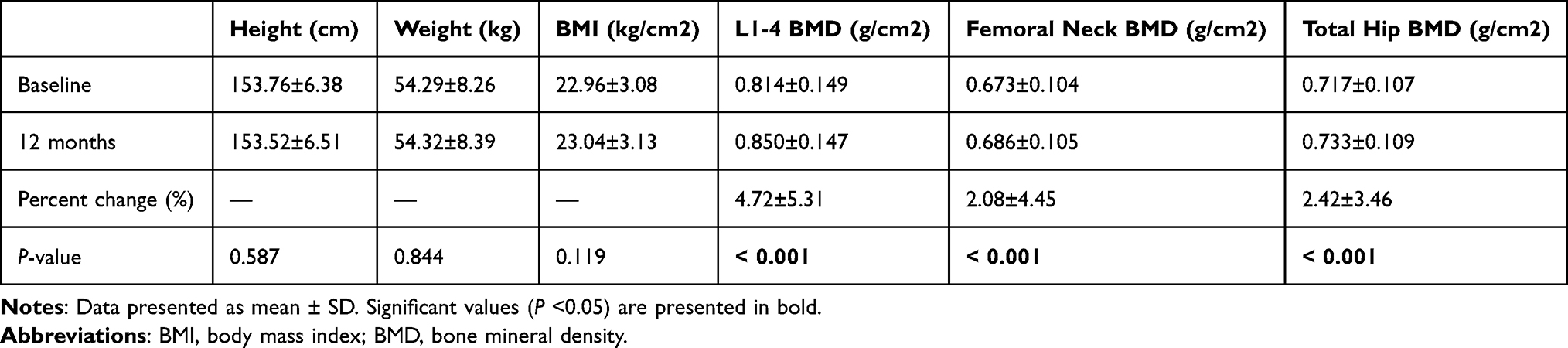 |
Table 2 Characteristic Changes of 466 Postmenopausal Women |
Allele Frequencies and Haplotype Structures
All 466 participants were genotyped. All tagSNPs were successfully genotyped and none of them failed the frequency test (MAF < 0.01). In the study, the population distribution of rs238243 deviated from HWE (P < 0.01) and were excluded from further analysis. The other 6 SNPs (rs13204, rs78224458, rs238242, rs113460962, rs4790714 and rs117337116) were compatible with HWE. Two haplotype blocks have been constructed from 6 SNPs (Figure 1).
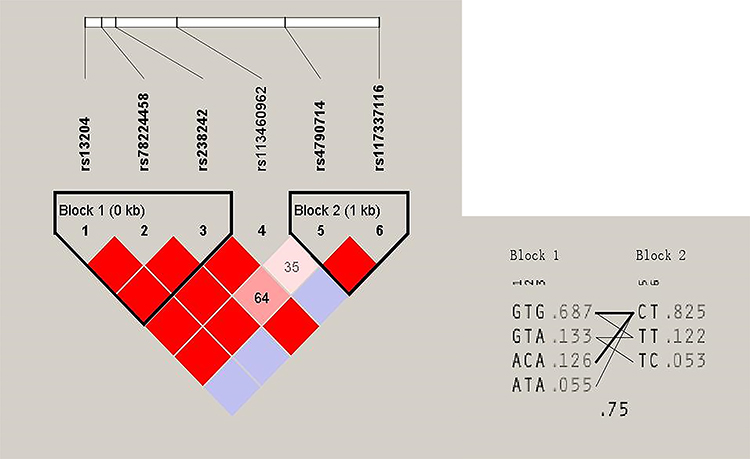 |
Figure 1 Haplotype linkage disequilibrium (LD) blocks in 466 postmenopausal women. Two LD blocks connecting SNP pairs are shaded based on the LD strength between SNPs by using the disequilibrium coefficient r2. The increasing degree of red of the cells represents the increasing strength of LD. LD plots with r2 values were generated using Haploview. Figure 1 shows the LD pattern in the PFN1 gene based on the 466 postmenopausal women with low bone mineral density. |
Association Between Genetic Polymorphisms and the Therapeutic Response to Alendronate Treatment
At baseline, there were no significant differences in BMD at L1–4, femoral neck and total hip between different genotype groups (P > 0.05). The associations between the 6 SNPs and the % change in BMD at L1–4, femoral neck or total hip were analyzed as the first step. Two haplotype blocks have been constructed from 6 SNPs (Figure 1). The rs13204, rs78224458 and rs238242 constituted block 1, and rs4790714, rs117337116 constituted block 2. After Bonferroni correction, no association was observed between any SNP or haplotype and BMD % change after 12 months of alendronate therapy in postmenopausal Chinese women with low BMD (Tables 3 and 4). In order to estimate the effectiveness of treatment, subjects were divided into responder(s) group and non-responder(s) group according to the LSC in BMD. The associations between 6 SNPs and being a responder to alendronate treatment at the L1–4, femoral neck or total hip were analyzed in the second step. In response to 70 mg weekly of alendronate, 55.0% subjects in spine BMD, 12.3% subjects in femoral neck, 57.8% subjects in total hip arrived an effective therapeutic range. Unfortunately, no association was found between any SNP or haplotype and treatment response in postmenopausal Chinese women with low BMD (Tables 5 and 6).
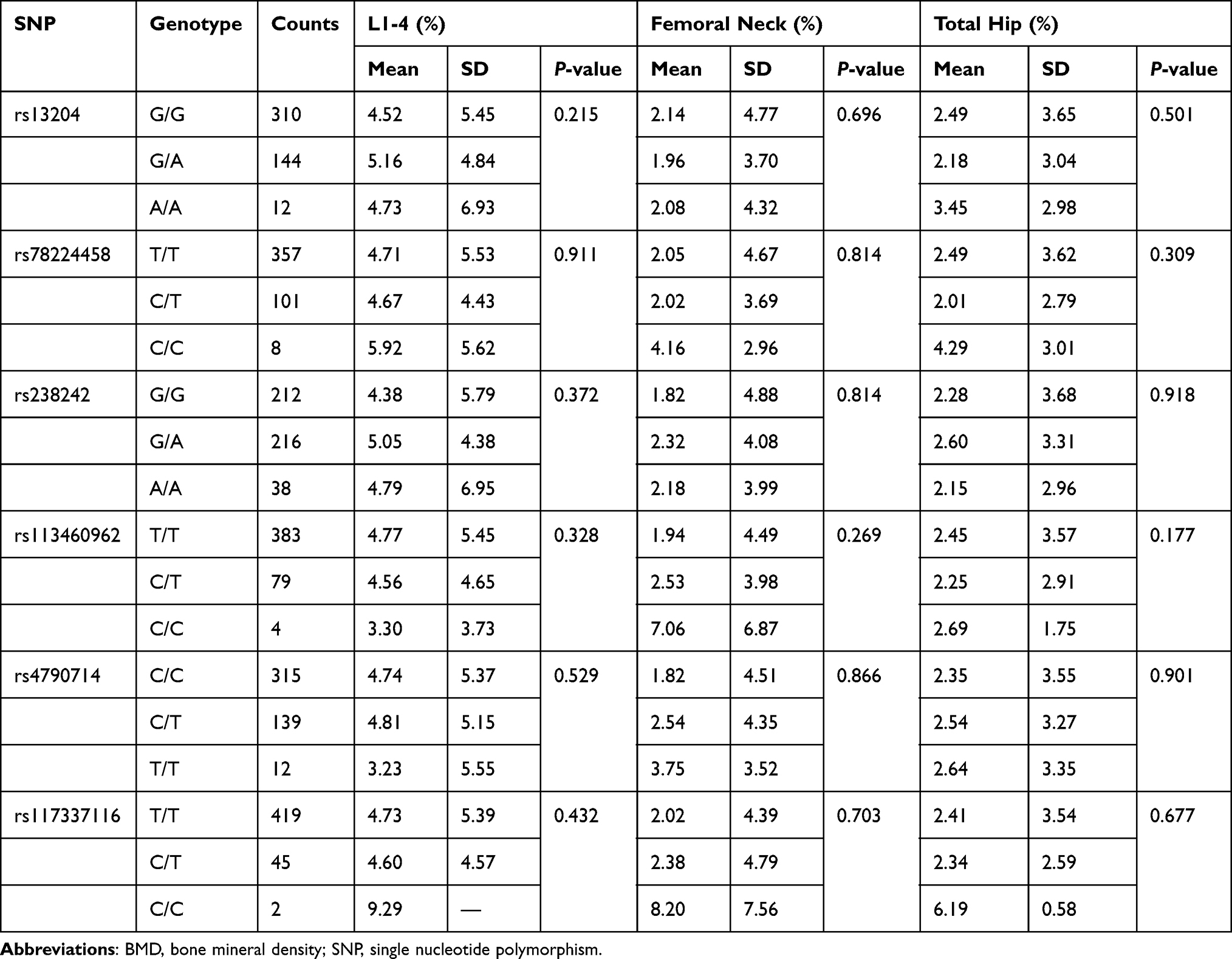 |
Table 3 The Association Between 6 Single Nucleotide Polymorphisms and the % Change in BMD |
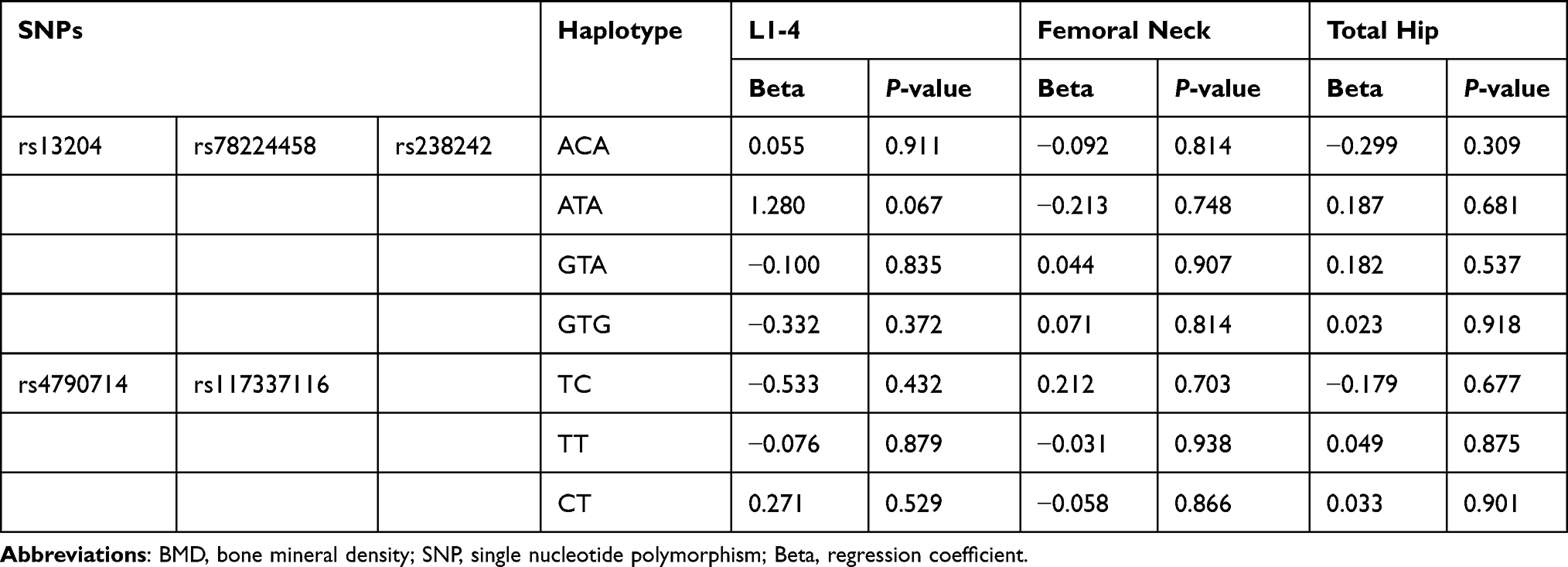 |
Table 4 The Association Between 2 Haplotypes and the % Change in BMD |
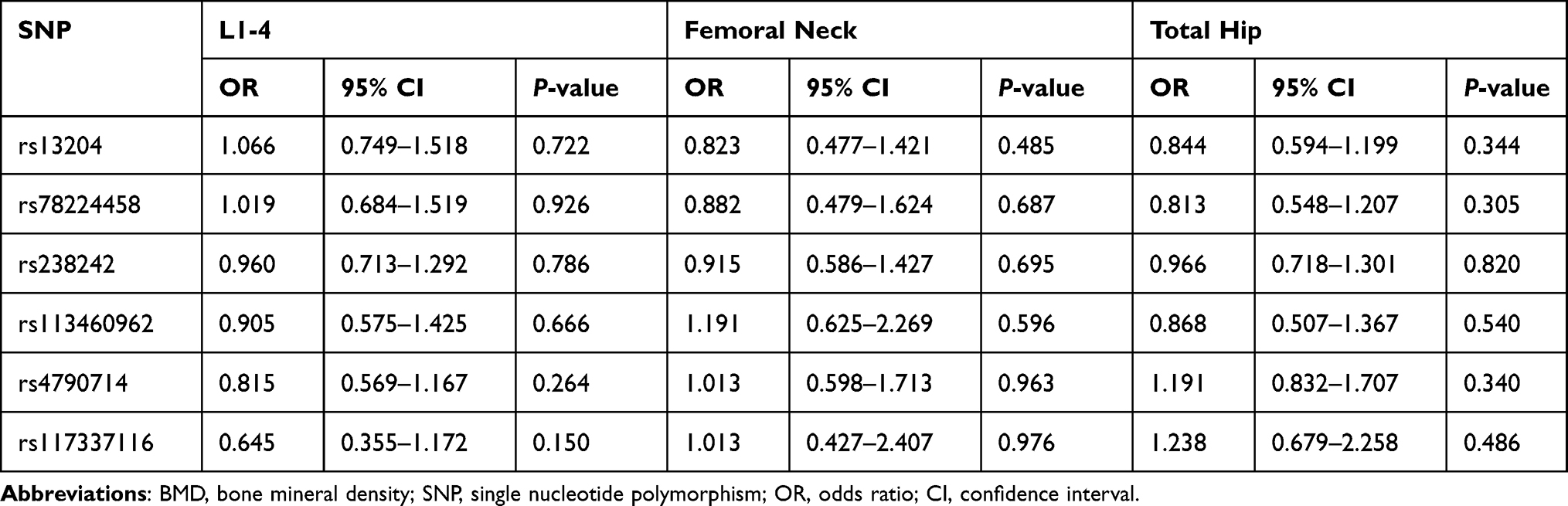 |
Table 5 The Association Between 6 Single Nucleotide Polymorphisms and BMD Response |
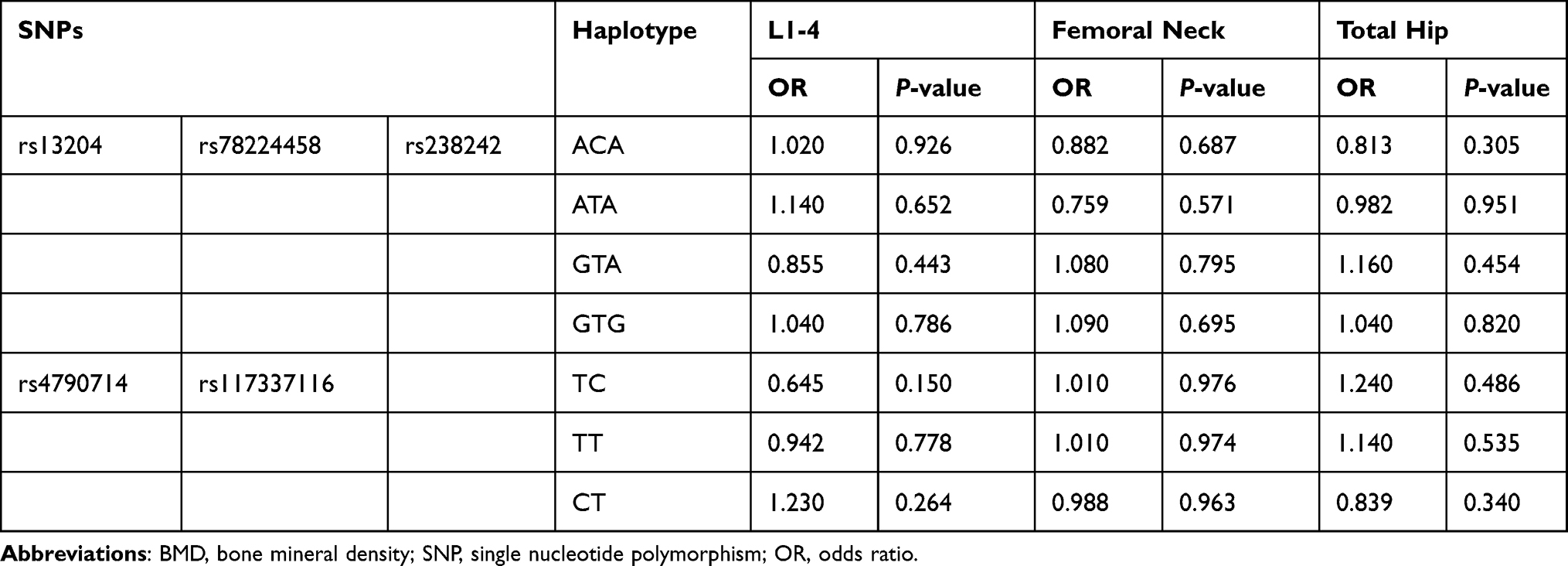 |
Table 6 The Association Between 2 Blocks and BMD Response |
Discussion
Osteoporosis is an important health problem worldwide. The effect of anti-osteoporosis treatment is determined by the bone-remodeling process, including the resorption of osteoclasts and the formation of osteoblasts. Alendronate is an important inhibitor of bone resorption and is the most commonly used bisphosphonate worldwide. It can increase BMD and decrease the risk of fragility fracture.38 It is well known that the treatment response to this drug is highly variable.22 Pharmacogenetics studies the genetic bases of individual drug response, determines the most effective drugs and the most beneficial doses for each patient. It has an important pharmacoeconomic impact on chronic diseases that require long-term treatment.39 As we all know, the response to alendronate treatment has a strong genetic component. Therefore, it is very important to clarify the genetic factors involved in anti-osteoporosis treatment, which will help to significantly improve the treatment effect. In previous studies, studies involving bone loss control, mevalonate pathway, farnesyl diphosphate synthase gene, geranylgeranyl diphosphate and other genes were analyzed.18,22,24,40–45 Despite all the advances in the field of pharmacogenetics in recent years, so far, these studies have found only a few SNPs that may be related to the response to anti-osteoporosis treatments. The pharmacogenetics of osteoporosis is still in its infancy, and pharmacogenomics research on anti-osteoporosis treatment is still insufficient.16
The PFN1 gene is a recently discovered pathogenic gene for early-onset Paget’s disease.32–34 It is widely expressed and highly expressed in blood and bone marrow cells.46 The PFN1 gene plays a central role in regulating de novo actin polymerization by enhancing ADP-ATP exchange on G-actin.47 Cell studies have confirmed that the PFN1 gene at least partially regulates the function of stem cells in bone marrow through the PFN1/Gα13/EGRL axis.48 Many studies have shown that PFN1 can stimulate the migration of endothelial cells, chondrocytes, human mesenchymal stem cells and neuronal cells.49,50 Zoidakis et al found that PFN1 plays an important role in skeletal development, and is a key molecule in sternal development and trabecular bone formation.51 Cell research found that in osteoblasts MC3T3-E1 (MC), the expression of PFN1 mRNA was down-regulated with osteogenic differentiation.52 And in MC cells, siRNA knocking out PFN1 can enhance the alkaline phosphatase activity induced by bone morphogenetic protein (BMP). Therefore, it can be considered that PFN1 is a new target of BMP, which can inhibit the differentiation of osteoblasts induced by BMP through transcription.52 Wanting et al conditionally knocked out profilin1 by using DMP1-CRE and PFN1-floxed mice. This conditional deletion of profilin1 specifically in osteocytes led to reduction in the levels of bone volume and BMD.53 This study indicated that PFN1 was expressed in osteocytes and regulates cell shape, migration and bone mass. In addition, Shirakawa et al investigated the osteoclast-specific functions of Pfn1 in mice and in cultured osteoclasts, and revealed its negative function in osteoclast movement and bone-resorbing activity.35 These studies have shown that PFN1 is closely related to the function of osteoblasts and osteoclasts as well as the regulation of bone mass. Therefore, we hypothesized PFN1 gene polymorphism as a candidate factor for the efficacy of alendronate.
To our knowledge, this is the first study reported a population-based association analysis of PFN1 gene polymorphism and response to alendronate treatment in postmenopausal Chinese women. In the present study, 500 postmenopausal Chinese women with osteoporosis or osteopenia were given alendronate at a dose of 70 mg weekly for 12 months, and 466 completed the therapy. In total, 7 SNPs in the PFN1 gene were genotyped in all patients. Initially, the association between candidate polymorphisms and baseline BMD, % change in BMD at L1–4, femoral neck and total hip were analyzed. No association was observed between any SNP or haplotype and baseline BMD or % change in BMD to 12 months of alendronate therapy in postmenopausal Chinese women with low BMD. Next, we considered that the % change of BMD may not represent the pharmacologic treatment response. According to the standards and guidelines established by ISCD 2007 Adult and Pediatric Official Positions, LSC should be considered to eliminate the error effect.54 Therefore, we divided the patients into responder(s) group and non-responder(s) group depending on our LSC at L1–4, femoral neck and total hip. Unfortunately, no SNP or haplotype was associated with the opportunity of being a responder at L–4, femoral neck or total hip. Although PFN1 has been proved to be the pathogenic gene of early-onset Paget’s disease, and it has been confirmed in cell and animal experiments that it is closely related to the skeletal homeostasis, we were unable to find a clear association between the PFN1 gene polymorphism and the response of Chinese postmenopausal women to alendronate treatment.
The results of this study may be due to the fact that although the PFN1 gene is consider to be closely related to the function of osteoblasts and osteoclasts, these studies have mainly focused on the cellular and animal levels. At present, the relationship between PFN1 and human bone mass is not clear. Another possible reason for failure to observe association of PFN1 polymorphism with the efficacy of alendronate is that unknown environmental factors (including other phenotypes or lifestyle factors such as vitamin D status, dietary characteristics, or medication compliance) may be confounding correlation between PFN1 polymorphism and the efficacy of alendronate, thereby masking the presence of a genetic association. And the number of subjects in our study was moderate. The rs238243 was detected deviated from HWE and excluded from further analysis. It was likely due to the relatively insufficient sample size. Therefore, it is necessary to testify the relationship between the excluded SNP and efficacy of alendronate treatment by enlarging samples size.
This is the first study to explore the genotype–treatment interaction between genetic variations in the PFN1 gene and BMD response to alendronate therapy in postmenopausal Chinese women with low BMD. However, we also acknowledge that our study has limitations. First, bone turnover markers were not evaluated. Bone turnover markers are also an important parameter to evaluate the effects of alendronate treatment. Second, 12 months follow-up time might not enough to detect the significant change of BMD. We believe that further studies with longer follow-up may be necessary.
Conclusion
In conclusion, this study demonstrated that polymorphisms of PFN1 gene may not be a major contributor to therapeutic response to alendronate in Chinese postmenopausal women with low BMD. Further studies that consider additional confounding factors or other types of polymorphisms such as insertion-deletions, copy number variants, and less common SNPs are needed for testing in a larger sample.
Acknowledgments
This study was supported by the National Key Research and Development Program of China (2018YFA0800801), National Basic Research Program of China (2014CB942903), National Natural Science Foundation of China (NSFC) (81770872, 81770874, 81974126 and 81770871), Clinical Science and Technology Innovation Project of Shanghai Shenkang Hospital Development Center (SHDC12018120), Shanghai Key Clinical Center for Metabolic Disease, Shanghai Health Commission Grant (2017ZZ01013), and Shanghai Municipal Key Clinical Specialty.
Funding
This study was supported by the National Key Research and Development Program of China (2018YFA0800801), National Basic Research Program of China (2014CB942903), National Natural Science Foundation of China (NSFC) (81770872, 81770874, 81974126 and 81770871), Clinical Science and Technology Innovation Project of Shanghai Shenkang Hospital Development Center (SHDC12018120), Shanghai Key Clinical Center for Metabolic Disease, Shanghai Health Commission Grant (2017ZZ01013), and Shanghai Municipal Key Clinical Specialty.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Seeman E, Bianchi G, Khosla S, Kanis JA, Orwoll E. Bone fragility in men–where are we? Osteoporos Int. 2006;17(11):1577–1583. doi:10.1007/s00198-006-0160-8
2. Pocock NA, Eisman JA, Hopper JL, et al. Genetic determinants of bone mass in adults. A twin study. J Clin Invest. 1987;80(3):706–710. doi:10.1172/JCI113125
3. Nguyen TV, Blangero J, Eisman JA. Genetic epidemiological approaches to the search for osteoporosis genes. J Bone Miner Res. 2000;15(3):392–401. doi:10.1359/jbmr.2000.15.3.392
4. Dawson-Hughes B, Looker AC, Tosteson AN, et al. The potential impact of the national osteoporosis foundation guidance on treatment eligibility in the USA: an update in NHANES 2005–2008. Osteoporos Int. 2012;23(3):811–820. doi:10.1007/s00198-011-1694-y
5. Xia WB, He SL, Xu L, et al. Rapidly increasing rates of hip fracture in Beijing, China. J Bone Miner Res. 2012;27(1):125–129. doi:10.1002/jbmr.519
6. Gao C, Xu Y, Li L, et al. Prevalence of osteoporotic vertebral fracture among community-dwelling elderly in Shanghai. Chin Med J. 2019;132(14):1749–1751. doi:10.1097/CM9.0000000000000332
7. Kanis JA, Burlet N, Cooper C, et al. European guidance for the diagnosis and management of osteoporosis in postmenopausal women. Osteoporos Int. 2008;19(4):399–428. doi:10.1007/s00198-008-0560-z
8. Watts NB, Adler RA, Bilezikian JP, et al. Osteoporosis in men: an Endocrine Society clinical practice guideline. J Clin Endocrinol Metab. 2012;97(6):1802–1822. doi:10.1210/jc.2011-3045
9. Ogawa S, Ouchi Y. [Therapeutic purpose and treatment guideline of osteoporosis]. Clin Calcium. 2012;22(6):885–889. Japanese.
10. Orimo H. [Bone and calcium update; diagnosis and therapy of metabolic bone disease update. Guideline for prevention and treatment of osteoporosis update]. Clin Calcium. 2011;21(12):123–143. Japanese.
11. Dawson-Hughes B. A revised clinician’s guide to the prevention and treatment of osteoporosis. J Clin Endocrinol Metab. 2008;93(7):2463–2465. doi:10.1210/jc.2008-0926
12. Delmas PD. The use of bisphosphonates in the treatment of osteoporosis. Curr Opin Rheumatol. 2005;17(4):462–466. doi:10.1097/01.bor.0000163448.51661.87
13. Cummings SR, Black DM, Thompson DE, et al. Effect of alendronate on risk of fracture in women with low bone density but without vertebral fractures: results from the Fracture Intervention Trial. JAMA. 1998;280(24):2077–2082. doi:10.1001/jama.280.24.2077
14. Quandt SA, Thompson DE, Schneider DL, Nevitt MC, Black DM. Effect of alendronate on vertebral fracture risk in women with bone mineral density T scores of-1.6 to −2.5 at the femoral neck: the Fracture Intervention Trial. Mayo Clin Proc. 2005;80(3):343–349. doi:10.4065/80.3.343
15. Eastell R, Barton I, Hannon RA, et al. Relationship of early changes in bone resorption to the reduction in fracture risk with risedronate. J Bone Miner Res. 2003;18(6):1051–1056. doi:10.1359/jbmr.2003.18.6.1051
16. Marini F, Brandi ML. Pharmacogenetics of osteoporosis: what is the evidence? Curr Osteoporos Rep. 2012;10(3):221–227. doi:10.1007/s11914-012-0110-7
17. Eisman JA. Genetics of osteoporosis. Endocr Rev. 1999;20(6):788–804. doi:10.1210/edrv.20.6.0384
18. Qureshi AM, Herd RJ, Blake GM, Fogelman I, Ralston SH. COLIA1 Sp1 polymorphism predicts response of femoral neck bone density to cyclical etidronate therapy. Calcif Tissue Int. 2002;70(3):158–163. doi:10.1007/s00223-001-1035-9
19. Palomba S, Numis FG, Mossetti G, et al. Effectiveness of alendronate treatment in postmenopausal women with osteoporosis: relationship with BsmI vitamin D receptor genotypes. Clin Endocrinol (Oxf). 2003;58(3):365–371. doi:10.1046/j.1365-2265.2003.01724.x
20. Palomba S, Orio F
21. Eisman JA. Pharmacogenetics of the vitamin D receptor and osteoporosis. Drug Metab Dispos. 2001;29(4 Pt 2):505–512.
22. Wang C, Zheng H, He JW, et al. Genetic polymorphisms in the mevalonate pathway affect the therapeutic response to alendronate treatment in postmenopausal Chinese women with low bone mineral density. Pharmacogenomics J. 2015;15(2):158–164. doi:10.1038/tpj.2014.52
23. Zheng H, Wang C, He JW, Fu WZ, Zhang ZL. OPG, RANKL, and RANK gene polymorphisms and the bone mineral density response to alendronate therapy in postmenopausal Chinese women with osteoporosis or osteopenia. Pharmacogenet Genomics. 2016;26(1):12–19. doi:10.1097/FPC.0000000000000181
24. Wang WJ, Fu WZ, He JW, Wang C, Zhang ZL. Association between SOST gene polymorphisms and response to alendronate treatment in postmenopausal Chinese women with low bone mineral density. Pharmacogenomics J. 2019;19(5):490–498. doi:10.1038/s41397-018-0059-8
25. Witke W. The role of profilin complexes in cell motility and other cellular processes. Trends Cell Biol. 2004;14(8):461–469. doi:10.1016/j.tcb.2004.07.003
26. Ding Z, Bae YH, Roy P. Molecular insights on context-specific role of profilin-1 in cell migration. Cell Adh Migr. 2012;6(5):442–449. doi:10.4161/cam.21832
27. Witke W, Podtelejnikov AV, Di Nardo A, et al. In mouse brain profilin I and profilin II associate with regulators of the endocytic pathway and actin assembly. EMBO J. 1998;17(4):967–976. doi:10.1093/emboj/17.4.967
28. Hein MY, Hubner NC, Poser I, et al. A human interactome in three quantitative dimensions organized by stoichiometries and abundances. Cell. 2015;163(3):712–723. doi:10.1016/j.cell.2015.09.053
29. Ertych N, Stolz A, Valerius O, Braus GH, Bastians H. CHK2-BRCA1 tumor-suppressor axis restrains oncogenic Aurora-A kinase to ensure proper mitotic microtubule assembly. Proc Natl Acad Sci U S A. 2016;113(7):1817–1822. doi:10.1073/pnas.1525129113
30. Zaidi AH, Manna SK. Profilin-PTEN interaction suppresses NF-κB activation via inhibition of IKK phosphorylation. Biochem J. 2016;473(7):859–872. doi:10.1042/BJ20150624
31. Lu H, Shamanna RA, de Freitas JK, et al. Cell cycle-dependent phosphorylation regulates RECQL4 pathway choice and ubiquitination in DNA double-strand break repair. Nat Commun. 2017;8(1):2039. doi:10.1038/s41467-017-02146-3
32. Wei Z, Li S, Tao X, et al. Mutations in profilin 1 cause early-onset Paget’s disease of bone with giant cell tumors. J Bone Miner Res. 2021;36(6):1088–1103. doi:10.1002/jbmr.4275
33. Scotto Di Carlo F, Pazzaglia L, Esposito T, Gianfrancesco F. The loss of profilin 1 causes early onset Paget’s disease of bone. Journal of Bone and Mineral Research: The Official Journal of the American Society for Bone and Mineral Research. 2020;35(8):1387–1398. doi:10.1002/jbmr.3964
34. Merlotti D, Materozzi M, Bianciardi S, et al. Mutation of PFN1 gene in an early onset, polyostotic Paget-like disease. J Clin Endocrinol Metab. 2020;105(8):2553–2565. doi:10.1210/clinem/dgaa252
35. Shirakawa J, Kajikawa S, Böttcher RT, et al. Profilin 1 negatively regulates osteoclast migration in postnatal skeletal growth, remodeling, and homeostasis in mice. JBMR Plus. 2019;3(6):e10130. doi:10.1002/jbm4.10130
36. Camacho PM, Petak SM, Binkley N, et al. American Association of Clinical Endocrinologists/American College of Endocrinology Clinical Practice Guidelines for the diagnosis and treatment of postmenopausal osteoporosis—2020 update executive summary. Endocr Pract. 2020;26(5):564–570. doi:10.4158/GL-2020-0524
37. Gao G, Zhang ZL, Zhang H, et al. Hip axis length changes in 10,554 males and females and the association with femoral neck fracture. J Clin Densitom. 2008;11(3):360–366. doi:10.1016/j.jocd.2008.04.005
38. Fisher JE, Rogers MJ, Halasy JM, et al. Alendronate mechanism of action: geranylgeraniol, an intermediate in the mevalonate pathway, prevents inhibition of osteoclast formation, bone resorption, and kinase activation in vitro. Proc Natl Acad Sci U S A. 1999;96(1):133–138. doi:10.1073/pnas.96.1.133
39. Marini F, Falchetti A, Silvestri S, et al. Modulatory effect of farnesyl pyrophosphate synthase (FDPS) rs2297480 polymorphism on the response to long-term amino-bisphosphonate treatment in postmenopausal osteoporosis. Curr Med Res Opin. 2008;24(9):2609–2615. doi:10.1185/03007990802352894
40. Chubachi S, Nakamura H, Sasaki M, et al. Polymorphism of LRP5 gene and emphysema severity are associated with osteoporosis in Japanese patients with or at risk for COPD. Respirology. 2015;20(2):286–295. doi:10.1111/resp.12429
41. Liu Y, Liu H, Li M, et al. Association of farnesyl diphosphate synthase polymorphisms and response to alendronate treatment in Chinese postmenopausal women with osteoporosis. Chin Med J. 2014;127(4):662–668.
42. Han LW, Ma DD, Xu XJ, et al. Association between geranylgeranyl pyrophosphate synthase gene polymorphisms and bone phenotypes and response to alendronate treatment in Chinese osteoporotic women. Chin Med Sci J. 2016;31(1):8–16. doi:10.1016/S1001-9294(16)30016-5
43. Olmos JM, Zarrabeitia MT, Hernández JL, et al. Common allelic variants of the farnesyl diphosphate synthase gene influence the response of osteoporotic women to bisphosphonates. Pharmacogenomics J. 2012;12(3):227–232. doi:10.1038/tpj.2010.88
44. Lima CA, Javorski NR, Souza AP, et al. Polymorphisms in key bone modulator cytokines genes influence bisphosphonates therapy in postmenopausal women. Inflammopharmacology. 2017;25(2):191–201. doi:10.1007/s10787-017-0322-7
45. Coffin JD, Homer-Bouthiette C, Hurley MM. Fibroblast growth factor 2 and its receptors in bone biology and disease. J Endocr Soc. 2018;2(7):657–671. doi:10.1210/js.2018-00105
46. Boopathy S, Silvas TV, Tischbein M, et al. Structural basis for mutation-induced destabilization of profilin 1 in ALS. Proc Natl Acad Sci U S A. 2015;112(26):7984–7989. doi:10.1073/pnas.1424108112
47. Witke W, Sutherland JD, Sharpe A, Arai M, Kwiatkowski DJ. Profilin I is essential for cell survival and cell division in early mouse development. Proc Natl Acad Sci U S A. 2001;98(7):3832–3836. doi:10.1073/pnas.051515498
48. Zheng J, Lu Z, Kocabas F, et al. Profilin 1 is essential for retention and metabolism of mouse hematopoietic stem cells in bone marrow. Blood. 2014;123(7):992–1001. doi:10.1182/blood-2013-04-498469
49. Ding Z, Lambrechts A, Parepally M, Roy P. Silencing profilin-1 inhibits endothelial cell proliferation, migration and cord morphogenesis. J Cell Sci. 2006;119(Pt 19):4127–4137. doi:10.1242/jcs.03178
50. Rust MB, Kullmann JA, Witke W. Role of the actin-binding protein profilin1 in radial migration and glial cell adhesion of granule neurons in the cerebellum. Cell Adh Migr. 2012;6(1):13–17. doi:10.4161/cam.19845
51. Zoidakis J, Makridakis M, Zerefos PG, et al. Profilin 1 is a potential biomarker for bladder cancer aggressiveness. Mol Cell Proteomics. 2012;11(4):
52. Lin W, Ezura Y, Izu Y, et al. Profilin expression is regulated by Bone Morphogenetic Protein (BMP) in osteoblastic cells. J Cell Biochem. 2016;117(3):621–628. doi:10.1002/jcb.25310
53. Lin W, Izu Y, Smriti A, et al. Profilin1 is expressed in osteocytes and regulates cell shape and migration. J Cell Physiol. 2018;233(1):259–268. doi:10.1002/jcp.25872
54. Lewiecki EM, Gordon CM, Baim S, et al. International society for clinical densitometry 2007 adult and pediatric official positions. Bone. 2008;43(6):1115–1121. doi:10.1016/j.bone.2008.08.106
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.