Back to Archived Journals » Oncolytic Virotherapy » Volume 6
Oncolytic virus delivery: from nano-pharmacodynamics to enhanced oncolytic effect
Authors Yokoda R ![]() , Nagalo BM, Vernon B
, Nagalo BM, Vernon B ![]() , Oklu R
, Oklu R ![]() , Albadawi H
, Albadawi H ![]() , DeLeon TT, Zhou Y
, DeLeon TT, Zhou Y ![]() , Egan JB, Duda DG, Borad MJ
, Egan JB, Duda DG, Borad MJ
Received 30 June 2017
Accepted for publication 9 October 2017
Published 8 November 2017 Volume 2017:6 Pages 39—49
DOI https://doi.org/10.2147/OV.S145262
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Prof. Dr. Athanasios Kyritsis
Raquel Yokoda,1 Bolni M Nagalo,1 Brent Vernon,2 Rahmi Oklu,3 Hassan Albadawi,3 Thomas T DeLeon,1 Yumei Zhou,1 Jan B Egan,1 Dan G Duda,4 Mitesh J Borad1
1Division of Hematology Oncology, Department of Medicine, Mayo Clinic, Scottsdale, 2Department of Biomedical Engineering, Arizona State University, Tempe, 3Division of Vascular and Interventional Radiology, Department of Radiology, Mayo Clinic, Scottsdale, AZ, 4Department of Radiation Oncology, Massachusetts General Hospital, Boston, MA, USA
Abstract: With the advancement of a growing number of oncolytic viruses (OVs) to clinical development, drug delivery is becoming an important barrier to overcome for optimal therapeutic benefits. Host immunity, tumor microenvironment and abnormal vascularity contribute to inefficient vector delivery. A number of novel approaches for enhanced OV delivery are under evaluation, including use of nanoparticles, immunomodulatory agents and complex viral–particle ligands along with manipulations of the tumor microenvironment. This field of OV delivery has quickly evolved to bioengineering of complex nanoparticles that could be deposited within the tumor using minimal invasive image-guided delivery. Some of the strategies include ultrasound (US)-mediated cavitation-enhanced extravasation, magnetic viral complexes delivery, image-guided infusions with focused US and targeting photodynamic virotherapy. In addition, strategies that modulate tumor microenvironment to decrease extracellular matrix deposition and increase viral propagation are being used to improve tumor penetration by OVs. Some involve modification of the viral genome to enhance their tumoral penetration potential. Here, we highlight the barriers to oncolytic viral delivery, and discuss the challenges to improving it and the perspectives of establishing new modes of active delivery to achieve enhanced oncolytic effects.
Keywords: oncolytic viruses, oncolytic virotherapy, drug delivery systems, tumor microenvironment
Introduction
Efficient delivery of oncolytic viruses (OVs) remains a major challenge in the field of oncology limiting their therapeutic effect. This may account for the disparity between in vitro and in vivo preclinical studies1,2 and the relatively modest antitumor effects observed thus far in clinical trials.3
Three major limitations need to be addressed to enhance delivery: first, virus bioavailability determined by the host vascular dynamics, perfusion parameters and innate immune responses;4–6 second, OV biodistribution and propagation,7–9 usually impaired by the intra-tumoral microenvironment heterogeneity; and third, the amplification of the virus bystander killing effect by cell-to-cell contact or by intrinsic vector enhancement.5 In this review, we discuss these three aspects and provide alternatives to improve them.
A number of prior studies have been done to optimize systemic delivery of OVs.10–12 Although a systemic approach is still a major goal of therapy given its simplicity, it has been difficult, and clinical trials employing systemic delivery have had limited success thus far.3,4,13 Even locoregional approaches of administration – such as intraperitoneal delivery for ovarian cancer,14 intrapleural delivery for mesotheliomas,15 intracavitary delivery for gliomas16 and intradermal delivery for melanomas17 – have yielded inconsistent results in both preclinical and clinical studies. Despite the difficulties for a viral vector to reach and eventually infect extravascular tumor cells, the main mode of administration has remained direct intravascular infusion through a major vessel supplying the tumor or by local intra-lesional injection in a solid tumor, with some studies employing both modes of administration.14,18 As a result, the intra-lesional approach has been preferred due to limited viral inactivation by the innate immune system, lower probability of systemic toxicity and optimal delivery of viral load in a single dose. Nonetheless, intra-tumoral delivery needs to be enhanced given the presence of a heterogeneous extracellular matrix (ECM) and the fact that transvascular perfusion may not be achieved to a therapeutic level. Inasmuch as tumor neovasculature is often abnormal, it severely impacts the intra-tumoral spread of OVs and maintenance of viral propagation. Tumor angiogenesis results in heterogeneous pericyte coverage promoting transient perfusion, leading to a hypoxic and acidic microenvironment, with tendency to microvasculature coagulation. This leads to suboptimal infection, impacts treatment resistance and may result in tumor recurrence.19–21 In view of this, vector enhancement has been equally important to improve the potential of the virus platform.
The host complex and viral vector bioavailability
Strategies for improving vector delivery include shielding of the virus from host immune defenses, and the use of nanoparticles for active targeting and nanofilaments to improve vector propagation.
Shielding and surface modifications
Shielding is done by cell-based delivery approaches22 listed in Table 1, or through an interface with nanoparticle carriers.23 The first relies on passive delivery, given that direction and flow of viral propagation is dependent solely on the cell-based properties towards the target organ or tissue. The latter relies on active delivery, where nanoparticle physical properties can be used to promote monitoring and targeting of a specific organ. In this case, shielding may be done by physical interface with biomaterials such as encapsulation and coating with polymers, or biodegradable nanoparticles, liposomes or copolymers.24 It can also be achieved by chemical modification with biomaterials such as polyamidoamine, polyethylene glycol (PEG), poly-N-(2-hydroxypropyl) methacrylamide, polysaccharides, bioreducible polymers, arginine-grafted bioreducible polymers, cationic polymers, poly-ethylenimine, poly-L-lysine and cationic lipids.24–26 Another way of shielding with biomaterials is through immobilization of the vector to a material’s surface through a process termed reverse transfection, solid-phase delivery or substrate-mediated delivery. This may also provide controlled viral release rate, localizing the gene expression to the surroundings, diminishing systemic infectivity, maintaining an elevated local concentration and as such helping to overcome transport limitations. Other similar biomaterials include microporous scaffolds, hydrogels, silk-elastin-like polymers, recombinant polymers, alginate and poly (lactic-co-glycolic acid), chitosan, fibrin and collagen micelles.23,25 Both cell-based and biomaterial interface-based delivery approaches are summarized in Table 1.
 | Table 1 Selected examples of shielding the OV from host barriers Note: *Cationic polymers and cationic lipids may be classified as a way to stablish physical instead of chemical interface because they are formed by electrostatic interactions with anionic adenoviruses rather than through chemical conjugation. Abbreviations: OV, oncolytic virus; MV, measles virus; Ad, adenovirus; GBM, glioblastoma multiforme; NSCs, neural stem cells; VSV, vesicular stomatitis virus; VSV-G, vesicular stomatitis virus glycoprotein G; PLGA, poly(lactic-co-glycolic acid); PNLG, poly[2-(dibutylamino)ethylamine-L-glutamate]; PAMAM, polyamidoamine; EGFR+, epidermal growth factor receptor positive; CAR, coxsackie adenovirus receptor; PLL, poly(L-lysine); PEG, polyethylene glycol; poly-HPMA, poly-N-(2-hydroxypropyl) methacrylamide. |
Using physical properties of nanoparticles to enhance active delivery and vector propagation
Active delivery is desirable over passive delivery as transvascular extravasation leads to the same constraints for nano-therapeutic viral delivery that have been described for other nanotherapy approaches.19,51 In this sense, the modulation of the tumor microenvironment is fundamental to enhance tumor spread.
Specific biomaterials possess properties required to allow delivery of viral vectors through ultrasound (US) using microbubbles (MBs) and focused sonoporation.52–54 While using MBs, contrast is employed for US using inert gas to pre-produce MBs that are injected at the tumor followed by percutaneous US, with shocking waves propelling the MBs against the tumor matrix causing temporary cavitation which may considerably enhance OV delivery through increased extravasation.
Another strategy for improved real-time monitoring of delivery includes magnetic-viral complexes, detected by magnetic resonance55,56 which enables noninvasive therapy monitoring. From the active propagation perspective, nanofilaments57 can be used to enhance viral propagation, in a way comparable to the spontaneously formed tunneling nanotubes in mesothelioma cells. The goal is to use ultrafine actin-based cytoplasmic extensions for increased bystander killing due to amplification of cell-to-cell contact.
Appropriate control of viral delivery can also be achieved by nanoparticles. Some techniques provide the particles with stimuli-responsive properties for enabling antitumoral effect. An example is photodynamic virotherapy, where the particle–viral ligand is armed with a genetically encoded photosensitizer, such as photofrin or talaporfin,58 which provides light-induced antitumor effect when activated at a specific wavelength. Another approach is deployment of a synthesized pH-sensitive polymer with a bioreducible disulfide bond (methoxy-pegylated cystaminebisacrylamide) armed to release the viral particle load in an acidic environment.59 Particle ligand approaches have also been used to target the tumor. An example of such an approach is the use of penetrating peptides or homing peptides60 such as arginylglycylaspartic acid (Arg-Gly-Asp) which are frequently used as a targeting moiety in adenovirus/polymer complexes with high affinity for αv integrins,61 E-selectins62 and vascular endothelial growth factor (VEGF) receptors.63 Target ligands can also be growth factors or antibodies such as cetuximab, and even natural ligands such as folate and chitosan PEG-folic acid.64 Active delivery approaches using virus–particle ligand complexes are summarized in Table 2. Some of those strategies may facilitate the induction of virus persistence by evasion of the DNA and RNA sensing systems and thereby help to improve oncolytic effect. On the top of that, further effects on viral replication and necessary viral load for each type of nanoparticle used and specific target organ still need to be evaluated in a case-by-case fashion.
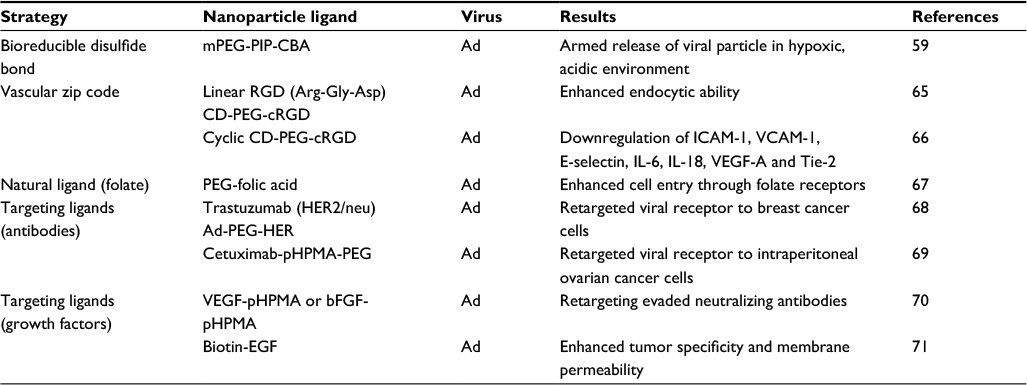 | Table 2 Selected examples of active delivery using complexes of virus–viral particle ligands Abbreviations: mPEG-PIP-CBA, methoxy-pegylated pH-sensitive polymer cystaminebisacrylamide; Ad, adenovirus; RGD, arginylglycylaspartic acid; CD-PEG-cRGD, cyclodextrin pegylated arginylglycylaspartic acid; ICAM-1, intercellular adhesion molecule 1; VCAM-1, vascular cell adhesion molecule 1; IL, interleukin; VEGF A, vascular endothelial growth factor A; PEG-folic acid, pegylated-folic acid; HER2/neu, human epidermal growth factor receptor 2/proto-oncogene neu; Ad-PEG-HER, pegylated adenovirus conjugated to herceptin; Cetuximab-pHPMA-PEG, cetuximab-pegylated conjugated with poly-N-(2-hydroxypropyl) methacrylamide; VEGF-pHPMA, vascular endothelial growth factor conjugated with poly-N-(2-hydroxypropyl)methacrylamide; bFGF-pHPMA, basic fibroblast growth factor conjugated with poly-N-(2-hydroxypropyl)methacrylamide; biotin-EGF, biotin conjugated with epidermal growth factor. |
Biomaterials shielding for improving monitoring and control of vector release
Biomaterial shielding has been important to diminish peripheral sequestration and to improve targeting by using specific ligands, while ensuring that size and molecular shape remain fairly uniform. This allows to predict hemodynamic interactions at a certain body temperature and blood viscosity depending on the polymer type used such as molecule, protein72 or hydrogels. This strategy also allows for prediction of the potential rate of viral release at the tumor level.73 In the long run, biomaterials may help track and predict viral delivery within reliable ranges. From this perspective, co-polymerization can result in enhanced tumor penetration. Similarly, optimal water solubility has been achieved by introducing a pH-sensitive cleavable linker and target moiety to a multi-arm copolymer and ultimately complexing it with viral particles.74,75
The tumor microenvironment: perfusion, permeability and retention
On one level, the more bioavailability a vector achieves inside the tumor, the more optimal the killing effect. On the other level, bioavailability can be increased by manipulation of host hemodynamics. Improving perfusion pressure may have an impact in a short window of time for locally administered compounds, as listed in Table 3. Animal studies have shown that promoting a hypertensive state through exercise also promotes extravasation for OV administration.76 Mayo Clinic studies8,76 have proposed a mathematical model of radial expansion and conflation of intra-tumoral infectious centers. This model also predicts the probability of tumor cell survival after the oncolytic phase and establishes perfusion pressure as a major determinant of intra-tumoral extravasation of OVs. The major impact was seen upon density of viral infection within the tumor achieved by increase and decrease in the mean arterial pressure.8,76
 | Table 3 Pharmacodynamic manipulation to enhance oncolytic virus bioavailability |
The focus on increasing permeability of tumor blood vessels has been critical since the enhanced perfusion and retention (EPR) effect was first described in 1986,77 exploiting the leaky nature of tumor vasculature.9,78 Convection is compromised on account of the high intra-tumoral interstitial pressure, dense heterogeneous stroma, lack of fenestration in the tumor endothelium and heterogeneous basement membranes hindering lymphatic drainage and impeding viral extravasation, all diminishing the EPR effect.19 The extent to which the EPR effect observed in murine models may translate to the human disease setting remains unclear.25
The tumor microenvironment as a barrier to OV delivery
Unlike normal vasculature, intra-tumoral vessels are immature, chaotic and mostly saccular, with a tortuosity that highly impacts effective blood perfusion.79 Nonuniform endothelial structure that promotes leakage in nonspecific areas plus constant changes in the tumor environment due to tumor growth and treatment may partially explain why a primary tumor may respond to treatment whereas its metastases may be unresponsive. The leakier a tumor becomes, the higher the interstitial fluid pressure, as the endothelial cells may not maintain pressure gradients across the endothelial wall and the drainage by lymphatics is dysfunctional. Stasis increases local hypoxia triggering upregulation of HIF-1 alpha and VEGF pathway activity, leading to a vicious cycle.80 Blocking VEGF has been shown to transiently “normalize” vascular structure and function, perhaps explaining why anti-VEGF antibodies have shown clinical efficacy (increased overall survival) when combined with chemotherapy in patients with mesothelioma, colon, lung, ovarian or cervical cancers. It was hypothesized that anti-VEGF approaches improve tumor perfusion by normalizing functionality of the tumor vasculature. The use of anti-VEGF and VEGF receptor therapies could enhance viral delivery in selected patients as well, akin to how these therapies have been enhancing immunotherapy.81 Similarly, another approach to normalize nitric oxide gradients to recover vessel function has been described.82
Another obstacle to viral penetration is the ECM, particularly in highly desmoplastic tumors, where nanoparticles injected directly inside the tumor are unable to move far from the injection site.84,85 One study co-injected local intra-tumoral bacterial collagenase with oncolytic herpes virus for melanoma treatment, and the distribution area of the OVs was found to increase threefold.86 Given that collagen is an integral structural component of the vascular wall, collagenase may never be used systemically. An alternate approach is to use an anti-fibrotic agent that can diminish collagen such as relaxin, which is known for reorganizing collagen during pregnancy, and has been shown to increase tumor penetration by OVs after 2 weeks.87 Another option is to use the metalloproteinase-1 and -8, which have been shown to increase OV delivery, improving distribution and yielding improved efficacy.88 A novel use for antihypertensive drugs from the angiotensin II receptor blocker class has been to modulate transforming growth factor beta activation and decrease collagen deposition.89 This mode of action has been employed in the prevention of esophageal sclerosis due to eosinophilic esophagitis. Losartan was the first candidate drug to be evaluated for decreasing collagen deposition in the tumor microenvironment. Dramatic decreases in collagen deposits and increase in tumor penetration were observed after 2 weeks.83 Another study in non-hypertensive patients with pancreatic adenocarcinoma demonstrated that use of candesartan was associated with a 6-month longer survival compared with the control group.90,91 In patients with hepatocellular carcinoma, candesartan downregulated the expression of VEGF-A, via the angiotensin II type I receptor, suggesting that it might be useful to inhibit angiogenesis in liver cancer.92 Studies with prostaglandin-I2 analogs have also demonstrated their efficiency in promoting vascular blood flow enhancement and extravasation at the tumor microenvironment, but safety studies must be performed prior to evaluating their potential for drug development.93
Intrinsic vector enhancement, selective replication and retargeting viral tropism
Therapeutic safety of vectors is proportional to their tumor target selectiveness. Selective targeting using tumor-specific promoters ensures that viral replication will be restricted to cancerous tissues while healthy tissues will remain unharmed. Examples of tumor-specific promoters are provided in Table 4. Another approach is to restrict tropism, and thereby enhance selectivity, by retargeting infection by a virus while ablating its ability to infect cells through its natural receptors.94 Instead, viruses are hexon swapped95 and pseudotyped with a more potent entry gene, or by fusing the entry gene to a single-chain antibody against upregulated tumor-specific receptors.96–98 To sum up, arming recombinant viruses with prodrug convertases, cytokines, and pH-releasing arms using a variety of envelopes, capsids and fibers may onset viral proteases only in a cancer-specific environment.99 Arming the viruses with a prodrug convertase will enable them to transform a nontoxic substrate or metabolite into a lethal drug within the tumor environment. Using this approach, the inclusion of cytotoxic genes and suicide genes was instituted in a herpes simplex virus (HSV) encoding thymidine kinase (TK) to sensitize dividing cells to ganciclovir.100 Still another strategy involves introduction of single-stranded RNA tumor suppressor or lethal genes with enhanced cytopathic and apoptotic effect (ADP-overexpressed adenovirus).101 Likewise, induction of autophagy can be accomplished by telomere-specific replication.102 Moreover, approaches where the virus may encode sequences for relaxin,103 decorin,104 hyaluronidase,105 heparanase106 and elastase107 from macrophages metalloelastase have tested the concept of tumor microenvironment targeting viruses with intrinsic capabilities.
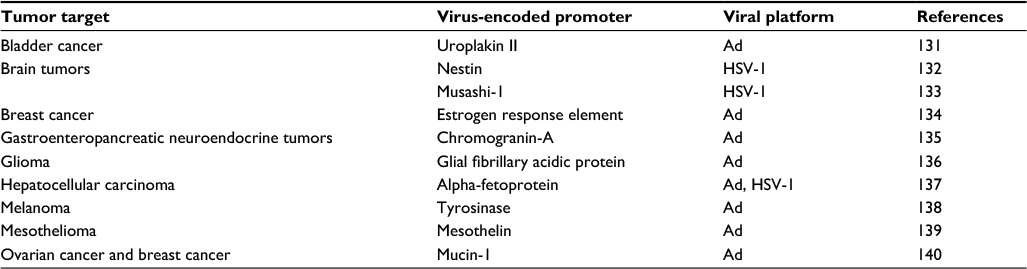 | Table 4 Selected examples of tumor-specific promoters Abbreviations: Ad, adenovirus; HSV-1, herpes simplex virus-1. |
Immunomodulation at the tumor microenvironment
Oncolysis in cancer cells may be limited by immune response. Although most tumor cells have defective immunomodulation and limited response to interferon (IFN) stimulation, normal cells can still sense the virus even if the progenies are replication deficient. Some viruses have developed mechanisms to evade or block the type-I IFN pathway at different levels.108 The most potent OVs are often wild types. First generations of OVs have been attenuated in an effort to achieve therapeutic safety, which led to reduced oncolytic potency. Hence, next generations are being engineered by modifying molecular patterns in order to boost oncolytic effect without compromising safety. Therefore, particular attention has been given to viral evasion of the DNA or RNA cytoplasmic sensing mechanisms, antiviral IFN blockages and molecular or sub-particle interactions that may improve oncolytic efficacy.99,109,110 On one hand, the virus should escape recognition by the host. On the other hand, the virus genome can be made to enhance tumor cell killing by using stimulatory cytokines.111 These include tumor necrosis factor-related apoptosis-inducing ligand,112 cytosine deaminase (CD),113 and immune-stimulatory cytokines genes such as interleukin (IL)-2,111 IL-12114 and IL-18.115 Some studies have employed a combination strategy with a TK-CD hybrid protein116 to enhance killing and cancer specificity.
At the transcriptional level, inhibition of angiogenesis by viral encoding genes has been achieved using short hairpin RNA-expressing oncolytic adenovirus-mediated inhibition of IL-8 resulting in antiangiogenesis and tumor growth inhibition.117 Similarly, a VEGF-specific short hairpin RNA-expressing adenovirus has been developed118 to block tumor growth and achieve potent inhibition of angiogenesis. These approaches exhibit ability of vector-encoded RNA knockdown for OV delivery. In addition, a vaccinia virus armed with the soluble VEGF receptor 1 protein developed antiangiogenic effect in a renal cancer cell model.119
Micro-RNA (miRNA) is evolving as a regulator of vector tropism,120,121 which is in contrast to the initial descriptions of RNA viruses where vector tropism could not be controlled through transcriptional targeting. This was evident in the case of coxsackievirus type A21, which causes off-target severe, often fatal myositis.122 miRNA techniques can promote detargeting of OVs during systemic administration, reshaping tumor tropism. Several insertions of combined miRNA target sites can be adapted to a single vector to detarget pivotal organs at risk for off-target side effects.123
Short-interfering RNA (si-RNA)124 delivery systems have been designed to increase tumor specificity. Small double-stranded RNAs impact posttranscriptional gene silencing as they target mRNAs that are then taken up by the RNA-induced silencing complex.125 They can be used to bind and guide cleavage of mRNA in a sequence homology-dependent manner.126 These may limit side effects, and toxicity, and take oncolytic virotherapy to a new safety level.
At the translational level, OV replication can be targeted by internal regulation of viral protein translation. This control is made through the internal ribosome entry site (IRES) element and is becoming a powerful tool to co-express genes of interest from a single mRNA, as IRES appears to play the role of a translational enhancer and may soon expand perspectives for better vectors.127 As such, multiple genes in viral payloads can be delivered using intrinsic genomic attributes, to impact viral delivery.
Local spread of the virus can be boosted by immunosuppressive drugs, such as cyclophosphamide.128 Pulsed application of immunosuppressive drugs is preferred, as seen with the prodrug 5-fluorocytosine and measles vaccine virus.129 Other immunomodulators such as cobra venom factor may also facilitate infection with HSV.130
Future directions
There is a need in the field of OV delivery to explore natural tropism of therapeutically modified viral platforms, such as hepatitis viruses for hepatocellular cancer and encephalitic viruses for brain tumors. Thus far, natural tropism of viruses has not been vastly explored from an oncolytic viral perspective.
Delivery of multiple distinct therapeutic viral vectors at the same time using biomaterials to bypass the viral load sequestration and neutralization by the immune system has not been attempted yet. This has the potential for simultaneous delivery of complementary viruses in an effort to achieve maximal synergy. Use of different viruses has been reported with better tumor penetration.141
Most current delivery approaches are passive in nature. Enhanced delivery could potentially be achieved using active delivery methods such as nanomachine-enabled propulsion of viral vectors. Simultaneously, the expectation is that active delivery could also contribute to the enoninvasive monitoring process of the OV targeting and propagation.
Improved monitoring methods for real-time viral injections and tagging viral particles for better in vivo visualization is also a growing necessity to assess viral delivery, especially for evaluating propagation after the first viral replication. To date, the gold standard for monitoring is biopsy. Bioluminescence and fluorescence optical methods are being developed along with noninvasive monitoring that allows deep tissue imaging. Examples of agents used for monitoring are radiotracer-coupled surface transporters used as the sodium iodide symporter,142 human norepinephrine transporter meta-iodobenzylguanidine which can be imagined by positron emission tomography or single photon emission computed tomography143 and human somatostatin receptor 2 radiolabeled with indium-111 along with vaccinia virus.144
Conclusion
Delivery of OVs remains a major challenge in the field of oncology. Rapidly evolving, innovative bioengineering and molecular approaches, at the host, tumor and viral level, are currently being tested. Overcoming the barriers to OV delivery will be paramount to clinical translation. A multimodal strategy based on novel viral engineering, host defense manipulation and novel active delivery techniques is necessary for more successful cancer therapy.
Disclosure
The authors report no conflict of interest in this work.
References
Saito Y, Sunamura M, Motoi F, et al. Oncolytic replication-competent adenovirus suppresses tumor angiogenesis through preserved E1A region. Cancer Gene Ther. 2006;13(3):242–252. | ||
Motoi F, Sunamura M, Ding L, et al. Effective gene therapy for pancreatic cancer by cytokines mediated by restricted replication-competent adenovirus. Hum Gene Ther. 2000;11(2):223–235. | ||
Seymour LW, Fisher KD. Oncolytic viruses: finally delivering. Br J Cancer. 2016;114(4):357–361. | ||
Russell SJ, Peng KW, Bell JC. Oncolytic virotherapy. Nat Biotechnol. 2012;30(7):658–670. | ||
Kaufman HL, Kohlhapp FJ, Zloza A. Oncolytic viruses: a new class of immunotherapy drugs. Nat Rev Drug Discov. 2015;14(9):642–662. | ||
Russell S, Miller A. Heterogeneous delivery is a barrier to the translational advancement of oncolytic virotherapy for treating solid tumors. Virus Adapt Treat. 2014;6(1):11–31. | ||
Allan KJ, Stojdl DF, Swift SL. High-throughput screening to enhance oncolytic virus immunotherapy. Oncolytic Virother. 2016;5:15–25. | ||
Bailey K, Kirk A, Naik S, et al. Mathematical model for radial expansion and conflation of intratumoral infectious centers predicts curative oncolytic virotherapy parameters. PLoS One. 2013;8(9):e73759. | ||
Nichols JW, Bae YH. EPR: evidence and fallacy. J Control Release. 2014;190:451–464. | ||
Grundy M, Coussios C, Carlisle R. Advances in systemic delivery of anti-cancer agents for the treatment of metastatic cancer. Expert Opin Drug Deliv. 2016;13(7):999–1013. | ||
Kim J, Li Y, Kim SW, Lee DS, Yun CO. Therapeutic efficacy of a systemically delivered oncolytic adenovirus – biodegradable polymer complex. Biomaterials. 2013;34(19):4622–4631. | ||
Ferguson MS, Lemoine NR, Wang Y. Systemic delivery of oncolytic viruses: hopes and hurdles. Adv Virol. 2012;2012:805629. | ||
Fisher K. Striking out at disseminated metastases: the systemic delivery of oncolytic viruses. Curr Opin Mol Ther. 2006;8(4):301–313. | ||
Matveeva OV, Guo ZS, Senin VM, Senina AV, Shabalina SA, Chumakov PM. Oncolysis by paramyxoviruses: preclinical and clinical studies. Mol Ther Oncolytics. 2015;2:15017. | ||
Belin LJ, Ady JW, Lewis C, et al. An oncolytic vaccinia virus expressing the human sodium iodine symporter prolongs survival and facilitates SPECT/CT imaging in an orthotopic model of malignant pleural mesothelioma. Surgery. 2013;154(3):486–495. | ||
Kaneda Y. A non-replicating oncolytic vector as a novel therapeutic tool against cancer. BMB Rep. 2010;43(12):773–780. | ||
Voit C, Kron M, Schwurzer-Voit M, Sterry W. Intradermal injection of Newcastle disease virus-modified autologous melanoma cell lysate and interleukin-2 for adjuvant treatment of melanoma patients with resectable stage III disease. J Dtsch Dermatol Ges. 2003;1(2):120–125. | ||
Apostolidis L, Schirrmacher V, Fournier P. Host mediated anti-tumor effect of oncolytic Newcastle disease virus after locoregional application. Int J Oncol. 2007;31(5):1009–1019. | ||
Danhier F. To exploit the tumor microenvironment: since the EPR effect fails in the clinic, what is the future of nanomedicine? J Control Release. 2016;244(Pt A):108–121. | ||
Jebar AH, Errington-Mais F, Vile RG, Selby PJ, Melcher AA, Griffin S. Progress in clinical oncolytic virus-based therapy for hepatocellular carcinoma. J Gen Virol. 2015;96(Pt 7):1533–1550. | ||
Busk M, Horsman MR. Relevance of hypoxia in radiation oncology: pathophysiology, tumor biology and implications for treatment. Q J Nucl Med Mol Imaging. 2013;57(3):219–234. | ||
Power AT, Bell JC. Cell-based delivery of oncolytic viruses: a new strategic alliance for a biological strike against cancer. Mol Ther. 2007;15(4):660–665. | ||
Jang JH, Schaffer DV, Shea LD. Engineering biomaterial systems to enhance viral vector gene delivery. Mol Ther. 2011;19(8):1407–1415. | ||
Lammers T. Improving the efficacy of combined modality anticancer therapy using HPMA copolymer-based nanomedicine formulations. Adv Drug Deliv Rev. 2010;62(2):203–230. | ||
Kim J, Kim PH, Kim SW, Yun CO. Enhancing the therapeutic efficacy of adenovirus in combination with biomaterials. Biomaterials. 2012;33(6):1838–1850. | ||
Duncan R, Vicent MJ. Do HPMA copolymer conjugates have a future as clinically useful nanomedicines? A critical overview of current status and future opportunities. Adv Drug Deliv Rev. 2010;62(2):272–282. | ||
Ong HT, Federspiel MJ, Guo CM, et al. Systemically delivered measles virus-infected mesenchymal stem cells can evade host immunity to inhibit liver cancer growth. J Hepatol. 2013;59(5):999–1006. | ||
Kaczorowski A, Hammer K, Liu L, et al. Delivery of improved oncolytic adenoviruses by mesenchymal stromal cells for elimination of tumorigenic pancreatic cancer cells. Oncotarget. 2016;7(8):9046–9059. | ||
Morshed RA, Gutova M, Juliano J, et al. Analysis of glioblastoma tumor coverage by oncolytic virus-loaded neural stem cells using MRI-based tracking and histological reconstruction. Cancer Gene Ther. 2015;22(1):55–61. | ||
Iankov ID, Blechacz B, Liu C, et al. Infected cell carriers: a new strategy for systemic delivery of oncolytic measles viruses in cancer virotherapy. Mol Ther. 2007;15(1):114–122. | ||
Power AT, Wang J, Falls TJ, et al. Carrier cell-based delivery of an oncolytic virus circumvents antiviral immunity. Mol Ther. 2007;15(1):123–130. | ||
Iankov ID, Msaouel P, Allen C, et al. Demonstration of anti-tumor activity of oncolytic measles virus strains in a malignant pleural effusion breast cancer model. Breast Cancer Res Treat. 2010;122(3):745–754. | ||
Sun CW, Willmon C, Wu LC, et al. Sickle cells abolish melanoma tumorigenesis in hemoglobin SS knockin mice and augment the tumoricidal effect of oncolytic virus in vivo. Front Oncol. 2016;6:166. | ||
Muthana M, Rodrigues S, Chen YY, et al. Macrophage delivery of an oncolytic virus abolishes tumor regrowth and metastasis after chemotherapy or irradiation. Cancer Res. 2013;73(2):490–495. | ||
Eisenstein S, Coakley BA, Briley-Saebo K, et al. Myeloid-derived suppressor cells as a vehicle for tumor-specific oncolytic viral therapy. Cancer Res. 2013;73(16):5003–5015. | ||
Bunuales M, Garcia-Aragoncillo E, Casado R, et al. Evaluation of monocytes as carriers for armed oncolytic adenoviruses in murine and Syrian hamster models of cancer. Hum Gene Ther. 2012;23(12):1258–1268. | ||
Liu X, Li YP, Zhong ZM, et al. Incorporation of viral glycoprotein VSV-G improves the delivery of DNA by erythrocyte ghost into cells refractory to conventional transfection. Appl Biochem Biotechnol. 2017;181(2):748–761. | ||
Mok H, Park JW, Park TG. Microencapsulation of PEGylated adenovirus within PLGA microspheres for enhanced stability and gene transfection efficiency. Pharm Res. 2007;24(12):2263–2269. | ||
Choi JW, Kim J, Bui QN, et al. Tuning surface charge and PEGylation of biocompatible polymers for efficient delivery of nucleic acid or adenoviral vector. Bioconjug Chem. 2015;26(8):1818–1829. | ||
Vetter A, Virdi KS, Espenlaub S, et al. Adenoviral vectors coated with PAMAM dendrimer conjugates allow CAR independent virus uptake and targeting to the EGF receptor. Mol Pharm. 2013;10(2):606–618. | ||
Park JW, Mok H, Park TG. Physical adsorption of PEG grafted and blocked poly-L-lysine copolymers on adenovirus surface for enhanced gene transduction. J Control Release. 2010;142(2):238–244. | ||
Chillon M, Lee JH, Fasbender A, Welsh MJ. Adenovirus complexed with polyethylene glycol and cationic lipid is shielded from neutralizing antibodies in vitro. Gene Ther. 1998;5(7):995–1002. | ||
Alemany R, Suzuki K, Curiel DT. Blood clearance rates of adenovirus type 5 in mice. J Gen Virol. 2000;81(Pt 11):2605–2609. | ||
Green NK, Herbert CW, Hale SJ, et al. Extended plasma circulation time and decreased toxicity of polymer-coated adenovirus. Gene Ther. 2004;11(16):1256–1263. | ||
Espenlaub S, Wortmann A, Engler T, Corjon S, Kochanek S, Kreppel F. Reductive amination as a strategy to reduce adenovirus vector promiscuity by chemical capsid modification with large polysaccharides. J Gene Med. 2008;10(12):1303–1314. | ||
Schek RM, Hollister SJ, Krebsbach PH. Delivery and protection of adenoviruses using biocompatible hydrogels for localized gene therapy. Mol Ther. 2004;9(1):130–138. | ||
Gustafson JA, Price RA, Greish K, Cappello J, Ghandehari H. Silk-elastin-like hydrogel improves the safety of adenovirus-mediated gene-directed enzyme-prodrug therapy. Mol Pharm. 2010;7(4):1050–1056. | ||
Kawamata Y, Nagayama Y, Nakao K, et al. Receptor-independent augmentation of adenovirus-mediated gene transfer with chitosan in vitro. Biomaterials. 2002;23(23):4573–4579. | ||
Breen A, Strappe P, Kumar A, O’Brien T, Pandit A. Optimization of a fibrin scaffold for sustained release of an adenoviral gene vector. J Biomed Mater Res A. 2006;78(4):702–708. | ||
Liao IC, Chen S, Liu JB, Leong KW. Sustained viral gene delivery through core-shell fibers. J Control Release. 2009;139(1):48–55. | ||
Torchilin V. Tumor delivery of macromolecular drugs based on the EPR effect. Adv Drug Deliv Rev. 2011;63(3):131–135. | ||
Bazan-Peregrino M, Arvanitis CD, Rifai B, Seymour LW, Coussios CC. Ultrasound-induced cavitation enhances the delivery and therapeutic efficacy of an oncolytic virus in an in vitro model. J Control Release. 2012;157(2):235–242. | ||
Bazan-Peregrino M, Rifai B, Carlisle RC, et al. Cavitation-enhanced delivery of a replicating oncolytic adenovirus to tumors using focused ultrasound. J Control Release. 2013;169(1–2):40–47. | ||
Carlisle R, Choi J, Bazan-Peregrino M, et al. Enhanced tumor uptake and penetration of virotherapy using polymer stealthing and focused ultrasound. J Natl Cancer Inst. 2013;105(22):1701–1710. | ||
Barnett BP, Arepally A, Karmarkar PV, et al. Magnetic resonance-guided, real-time targeted delivery and imaging of magnetocapsules immunoprotecting pancreatic islet cells. Nat Med. 2007;13(8):986–991. | ||
Almstatter I, Mykhaylyk O, Settles M, et al. Characterization of magnetic viral complexes for targeted delivery in oncology. Theranostics. 2015;5(7):667–685. | ||
Ady J, Thayanithy V, Mojica K, et al. Tunneling nanotubes: an alternate route for propagation of the bystander effect following oncolytic viral infection. Mol Ther Oncolytics. 2016;3:16029. | ||
Takehara K, Tazawa H, Okada N, et al. Targeted photodynamic virotherapy armed with a genetically encoded photosensitizer. Mol Cancer Ther. 2016;15(1):199–208. | ||
Choi JW, Jung SJ, Kasala D, et al. pH-sensitive oncolytic adenovirus hybrid targeting acidic tumor microenvironment and angiogenesis. J Control Release. 2015;205:134–143. | ||
Pike DB, Ghandehari H. HPMA copolymer-cyclic RGD conjugates for tumor targeting. Adv Drug Deliv Rev. 2010;62(2):167–183. | ||
Brooks PC, Clark RA, Cheresh DA. Requirement of vascular integrin alpha v beta 3 for angiogenesis. Science. 1994;264(5158):569–571. | ||
Luo J, Paranya G, Bischoff J. Noninflammatory expression of E-selectin is regulated by cell growth. Blood. 1999;93(11):3785–3791. | ||
Veikkola T, Karkkainen M, Claesson-Welsh L, Alitalo K. Regulation of angiogenesis via vascular endothelial growth factor receptors. Cancer Res. 2000;60(2):203–212. | ||
Choi JW, Lee YS, Yun CO, Kim SW. Polymeric oncolytic adenovirus for cancer gene therapy. J Control Release. 2015;219:181–191. | ||
Suh W, Han SO, Yu L, Kim SW. An angiogenic, endothelial-cell-targeted polymeric gene carrier. Mol Ther. 2002;6(5):664–672. | ||
Ogawara K, Kuldo JM, Oosterhuis K, et al. Functional inhibition of NF-kappaB signal transduction in alphavbeta3 integrin expressing endothelial cells by using RGD-PEG-modified adenovirus with a mutant IkappaB gene. Arthritis Res Ther. 2006;8(1):R32. | ||
Oh IK, Mok H, Park TG. Folate immobilized and PEGylated adenovirus for retargeting to tumor cells. Bioconjug Chem. 2006;17(3):721–727. | ||
Jung Y, Park HJ, Kim PH, et al. Retargeting of adenoviral gene delivery via Herceptin-PEG-adenovirus conjugates to breast cancer cells. J Control Release. 2007;123(2):164–171. | ||
Morrison J, Briggs SS, Green NK, et al. Cetuximab retargeting of adenovirus via the epidermal growth factor receptor for treatment of intraperitoneal ovarian cancer. Hum Gene Ther. 2009;20(3):239–251. | ||
Fisher KD, Stallwood Y, Green NK, Ulbrich K, Mautner V, Seymour LW. Polymer-coated adenovirus permits efficient retargeting and evades neutralising antibodies. Gene Ther. 2001;8(5):341–348. | ||
Park JW, Mok H, Park TG. Epidermal growth factor (EGF) receptor targeted delivery of PEGylated adenovirus. Biochem Biophys Res Commun. 2008;366(3):769–774. | ||
Erickson HP. Size and shape of protein molecules at the nanometer level determined by sedimentation, gel filtration, and electron microscopy. Biol Proced Online. 2009;11:32–51. | ||
Ramos J, Forcada J, Hidalgo-Alvarez R. Cationic polymer nanoparticles and nanogels: from synthesis to biotechnological applications. Chem Rev. 2014;114(1):367–428. | ||
Sun P, Chen D, Deng H, et al. “Bottom-up” construction of multi-polyprodrug-arm hyperbranched amphiphiles for cancer therapy. Bioconjug Chem. 2017;28(5):1470–1480. | ||
Zhang Y, Chen W, Yang C, Fan Q, Wu W, Jiang X. Enhancing tumor penetration and targeting using size-minimized and zwitterionic nanomedicines. J Control Release. 2016;237:115–124. | ||
Miller A, Nace R, Ayala-Breton CC, et al. Perfusion pressure is a critical determinant of the intratumoral extravasation of oncolytic viruses. Mol Ther. 2016;24(2):306–317. | ||
Matsumura Y, Maeda H. A new concept for macromolecular therapeutics in cancer chemotherapy: mechanism of tumoritropic accumulation of proteins and the antitumor agent smancs. Cancer Res. 1986;46(12 Pt 1):6387–6392. | ||
Fang J, Nakamura H, Maeda H. The EPR effect: unique features of tumor blood vessels for drug delivery, factors involved, and limitations and augmentation of the effect. Adv Drug Deliv Rev. 2011;63(3):136–151. | ||
Brown EB, Campbell RB, Tsuzuki Y, et al. In vivo measurement of gene expression, angiogenesis and physiological function in tumors using multiphoton laser scanning microscopy. Nat Med. 2001;7(7):864–868. | ||
Jain RK. Normalizing tumor microenvironment to treat cancer: bench to bedside to biomarkers. J Clin Oncol. 2013;31(17):2205–2218. | ||
Huang Y, Goel S, Duda DG, Fukumura D, Jain RK. Vascular normalization as an emerging strategy to enhance cancer immunotherapy. Cancer Res. 2013;73(10):2943–2948. | ||
Fukumura D, Kashiwagi S, Jain RK. The role of nitric oxide in tumour progression. Nat Rev Cancer. 2006;6(7):521–534. | ||
Diop-Frimpong B, Chauhan VP, Krane S, Boucher Y, Jain RK. Losartan inhibits collagen I synthesis and improves the distribution and efficacy of nanotherapeutics in tumors. Proc Natl Acad Sci U S A. 2011;108(7):2909–2914. | ||
Yuan F, Leunig M, Huang SK, Berk DA, Papahadjopoulos D, Jain RK. Microvascular permeability and interstitial penetration of sterically stabilized (stealth) liposomes in a human tumor xenograft. Cancer Res. 1994;54(13):3352–3356. | ||
Netti PA, Berk DA, Swartz MA, Grodzinsky AJ, Jain RK. Role of extracellular matrix assembly in interstitial transport in solid tumors. Cancer Res. 2000;60(9):2497–2503. | ||
McKee TD, Grandi P, Mok W, et al. Degradation of fibrillar collagen in a human melanoma xenograft improves the efficacy of an oncolytic herpes simplex virus vector. Cancer Res. 2006;66(5):2509–2513. | ||
Perentes JY, McKee TD, Ley CD, et al. In vivo imaging of extracellular matrix remodeling by tumor-associated fibroblasts. Nat Methods. 2009;6(2):143–145. | ||
Mok W, Boucher Y, Jain RK. Matrix metalloproteinases-1 and -8 improve the distribution and efficacy of an oncolytic virus. Cancer Res. 2007;67(22):10664–10668. | ||
Kumar V, Boucher Y, Liu H, et al. Noninvasive assessment of Losartan-induced increase in functional microvasculature and drug delivery in pancreatic ductal adenocarcinoma. Transl Oncol. 2016;9(5):431–437. | ||
Nakai Y, Isayama H, Ijichi H, et al. Phase I trial of gemcitabine and candesartan combination therapy in normotensive patients with advanced pancreatic cancer: GECA1. Cancer Sci. 2012;103(8):1489–1492. | ||
Cerullo M, Gani F, Chen SY, Canner JK, Pawlik TM. Impact of angiotensin receptor blocker use on overall survival among patients undergoing resection for pancreatic cancer. World J Surg. 2017;41(9):2361–2370. | ||
Fan F, Tian C, Tao L, et al. Candesartan attenuates angiogenesis in hepatocellular carcinoma via downregulating AT1R/VEGF pathway. Biomed Pharmacother. 2016;83:704–711. | ||
Tanaka S, Akaike T, Wu J, et al. Modulation of tumor-selective vascular blood flow and extravasation by the stable prostaglandin 12 analogue beraprost sodium. J Drug Target. 2003;11(1):45–52. | ||
Schneider U, Bullough F, Vongpunsawad S, Russell SJ, Cattaneo R. Recombinant measles viruses efficiently entering cells through targeted receptors. J Virol. 2000;74(21):9928–9936. | ||
Rivera AA, Davydova J, Schierer S, et al. Combining high selectivity of replication with fiber chimerism for effective adenoviral oncolysis of CAR-negative melanoma cells. Gene Ther. 2004;11(23):1694–1702. | ||
Bucheit AD, Kumar S, Grote DM, et al. An oncolytic measles virus engineered to enter cells through the CD20 antigen. Mol Ther. 2003;7(1):62–72. | ||
Nakamura T, Peng KW, Harvey M, et al. Rescue and propagation of fully retargeted oncolytic measles viruses. Nat Biotechnol. 2005;23(2):209–214. | ||
Hammond AL, Plemper RK, Zhang J, Schneider U, Russell SJ, Cattaneo R. Single-chain antibody displayed on a recombinant measles virus confers entry through the tumor-associated carcinoembryonic antigen. J Virol. 2001;75(5):2087–2096. | ||
Cattaneo R, Miest T, Shashkova EV, Barry MA. Reprogrammed viruses as cancer therapeutics: targeted, armed and shielded. Nat Rev Microbiol. 2008;6(7):529–540. | ||
Boviatsis EJ, Park JS, Sena-Esteves M, et al. Long-term survival of rats harboring brain neoplasms treated with ganciclovir and a herpes simplex virus vector that retains an intact thymidine kinase gene. Cancer Res. 1994;54(22):5745–5751. | ||
Yun CO, Kim E, Koo T, Kim H, Lee YS, Kim JH. ADP-overexpressing adenovirus elicits enhanced cytopathic effect by induction of apoptosis. Cancer Gene Ther. 2005;12(1):61–71. | ||
Tazawa H, Kagawa S, Fujiwara T. Oncolytic adenovirus-induced autophagy: tumor-suppressive effect and molecular basis. Acta Med Okayama. 2013;67(6):333–342. | ||
Kim JH, Lee YS, Kim H, Huang JH, Yoon AR, Yun CO. Relaxin expression from tumor-targeting adenoviruses and its intratumoral spread, apoptosis induction, and efficacy. J Natl Cancer Inst. 2006;98(20):1482–1493. | ||
Choi IK, Lee YS, Yoo JY, et al. Effect of decorin on overcoming the extracellular matrix barrier for oncolytic virotherapy. Gene Ther. 2010;17(2):190–201. | ||
Guedan S, Rojas JJ, Gros A, Mercade E, Cascallo M, Alemany R. Hyaluronidase expression by an oncolytic adenovirus enhances its intratumoral spread and suppresses tumor growth. Mol Ther. 2010;18(7):1275–1283. | ||
Watanabe Y, Kojima T, Kagawa S, et al. A novel translational approach for human malignant pleural mesothelioma: heparanase-assisted dual virotherapy. Oncogene. 2010;29(8):1145–1154. | ||
Lavilla-Alonso S, Bauer MM, Abo-Ramadan U, et al. Macrophage metalloelastase (MME) as adjuvant for intra-tumoral injection of oncolytic adenovirus and its influence on metastases development. Cancer Gene Ther. 2012;19(2):126–134. | ||
Christensen MH, Paludan SR. Viral evasion of DNA-stimulated innate immune responses. Cell Mol Immunol. 2017;14(1):4–13. | ||
Lam E, Stein S, Falck-Pedersen E. Adenovirus detection by the cGAS/STING/TBK1 DNA sensing cascade. J Virol. 2014;88(2):974–981. | ||
Champion BR, Fisher K, Seymour L. A PTENtial cause for the selectivity of oncolytic viruses? Nat Immunol. 2016;17(3):225–226. | ||
Choi IK, Li Y, Oh E, Kim J, Yun CO. Oncolytic adenovirus expressing IL-23 and p35 elicits IFN-gamma- and TNF-alpha-co-producing T cell-mediated antitumor immunity. PLoS One. 2013;8(7):e67512. | ||
Sova P, Ren XW, Ni S, et al. A tumor-targeted and conditionally replicating oncolytic adenovirus vector expressing TRAIL for treatment of liver metastases. Mol Ther. 2004;9(4):496–509. | ||
Leveille S, Samuel S, Goulet ML, Hiscott J. Enhancing VSV oncolytic activity with an improved cytosine deaminase suicide gene strategy. Cancer Gene Ther. 2011;18(6):435–443. | ||
Lee YS, Kim JH, Choi KJ, et al. Enhanced antitumor effect of oncolytic adenovirus expressing interleukin-12 and B7-1 in an immunocompetent murine model. Clin Cancer Res. 2006;12(19):5859–5868. | ||
Choi IK, Lee JS, Zhang SN, et al. Oncolytic adenovirus co-expressing IL-12 and IL-18 improves tumor-specific immunity via differentiation of T cells expressing IL-12Rbeta2 or IL-18Ralpha. Gene Ther. 2011;18(9):898–909. | ||
Freytag SO, Rogulski KR, Paielli DL, Gilbert JD, Kim JH. A novel three-pronged approach to kill cancer cells selectively: concomitant viral, double suicide gene, and radiotherapy. Hum Gene Ther. 1998;9(9):1323–1333. | ||
Yoo JY, Kim JH, Kim J, et al. Short hairpin RNA-expressing oncolytic adenovirus-mediated inhibition of IL-8: effects on antiangiogenesis and tumor growth inhibition. Gene Ther. 2008;15(9):635–651. | ||
Yoo JY, Kim JH, Kwon YG, et al. VEGF-specific short hairpin RNA-expressing oncolytic adenovirus elicits potent inhibition of angiogenesis and tumor growth. Mol Ther. 2007;15(2):295–302. | ||
Grunwald GK, Vetter A, Klutz K, et al. EGFR-targeted adenovirus dendrimer coating for improved systemic delivery of the theranostic NIS gene. Mol Ther Nucleic Acids. 2013;2:e131. | ||
Kelly EJ, Russell SJ. MicroRNAs and the regulation of vector tropism. Mol Ther. 2009;17(3):409–416. | ||
Ruiz AJ, Russell SJ. MicroRNA-based regulation of picornavirus tropism. J Vis Exp. 2017;120(2):e55033. | ||
Kelly EJ, Hadac EM, Greiner S, Russell SJ. Engineering microRNA responsiveness to decrease virus pathogenicity. Nat Med. 2008;14(11):1278–1283. | ||
Baertsch MA, Leber MF, Bossow S, et al. MicroRNA-mediated multi-tissue detargeting of oncolytic measles virus. Cancer Gene Ther. 2014;21(9):373–380. | ||
Fire A, Xu S, Montgomery MK, Kostas SA, Driver SE, Mello CC. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature. 1998;391(6669):806–811. | ||
Resnier P, Montier T, Mathieu V, Benoit JP, Passirani C. A review of the current status of siRNA nanomedicines in the treatment of cancer. Biomaterials. 2013;34(27):6429–6443. | ||
Kullberg M, McCarthy R, Anchordoquy TJ. Systemic tumor-specific gene delivery. J Control Release. 2013;172(3):730–736. | ||
Renaud-Gabardos E, Hantelys F, Morfoisse F, Chaufour X, Garmy-Susini B, Prats AC. Internal ribosome entry site-based vectors for combined gene therapy. World J Exp Med. 2015;5(1):11–20. | ||
Wakimoto H, Fulci G, Tyminski E, Chiocca EA. Altered expression of antiviral cytokine mRNAs associated with cyclophosphamide’s enhancement of viral oncolysis. Gene Ther. 2004;11(2):214–223. | ||
Yurttas C, Berchtold S, Malek NP, Bitzer M, Lauer UM. Pulsed versus continuous application of the prodrug 5-fluorocytosine to enhance the oncolytic effectiveness of a measles vaccine virus armed with a suicide gene. Hum Gene Ther Clin Dev. 2014;25(2):85–96. | ||
Ikeda K, Wakimoto H, Ichikawa T, et al. Complement depletion facilitates the infection of multiple brain tumors by an intravascular, replication-conditional herpes simplex virus mutant. J Virol. 2000;74(10):4765–4775. | ||
Zhang J, Ramesh N, Chen Y, et al. Identification of human uroplakin II promoter and its use in the construction of CG8840, a urothelium-specific adenovirus variant that eliminates established bladder tumors in combination with docetaxel. Cancer Res. 2002;62(13):3743–3750. | ||
Kambara H, Okano H, Chiocca EA, Saeki Y. An oncolytic HSV-1 mutant expressing ICP34.5 under control of a nestin promoter increases survival of animals even when symptomatic from a brain tumor. Cancer Res. 2005;65(7):2832–2839. | ||
Kanai R, Tomita H, Shinoda A, et al. Enhanced therapeutic efficacy of G207 for the treatment of glioma through Musashi1 promoter retargeting of gamma34.5-mediated virulence. Gene Ther. 2006;13(2):106–116. | ||
Hernandez-Alcoceba R, Pihalja M, Wicha MS, Clarke MF. A novel, conditionally replicative adenovirus for the treatment of breast cancer that allows controlled replication of E1a-deleted adenoviral vectors. Hum Gene Ther. 2000;11(14):2009–2024. | ||
Leja J, Dzojic H, Gustafson E, Oberg K, Giandomenico V, Essand M. A novel chromogranin-A promoter-driven oncolytic adenovirus for midgut carcinoid therapy. Clin Cancer Res. 2007;13(8):2455–2462. | ||
Hoffmann D, Meyer B, Wildner O. Improved glioblastoma treatment with Ad5/35 fiber chimeric conditionally replicating adenoviruses. J Gene Med. 2007;9(9):764–778. | ||
Hallenbeck PL, Chang YN, Hay C, et al. A novel tumor-specific replication-restricted adenoviral vector for gene therapy of hepatocellular carcinoma. Hum Gene Ther. 1999;10(10):1721–1733. | ||
Nettelbeck DM, Rivera AA, Balague C, Alemany R, Curiel DT. Novel oncolytic adenoviruses targeted to melanoma: specific viral replication and cytolysis by expression of E1A mutants from the tyrosinase enhancer/promoter. Cancer Res. 2002;62(16):4663–4670. | ||
Tsuruta Y, Pereboeva L, Glasgow JN, et al. A mosaic fiber adenovirus serotype 5 vector containing reovirus sigma 1 and adenovirus serotype 3 knob fibers increases transduction in an ovarian cancer ex vivo system via a coxsackie and adenovirus receptor-independent pathway. Clin Cancer Res. 2007;13(9):2777–2783. | ||
Kurihara T, Brough DE, Kovesdi I, Kufe DW. Selectivity of a replication-competent adenovirus for human breast carcinoma cells expressing the MUC1 antigen. J Clin Invest. 2000;106(6):763–771. | ||
Le Boeuf F, Diallo JS, McCart JA, et al. Synergistic interaction between oncolytic viruses augments tumor killing. Mol Ther. 2010;18(5):888–895. | ||
Wang J, Arulanandam R, Wassenaar R, et al. Enhancing expression of functional human sodium iodide symporter and somatostatin receptor in recombinant oncolytic vaccinia virus for in vivo imaging of tumors. J Nucl Med. 2017;58(2):221–227. | ||
Chen N, Zhang Q, Yu YA, et al. A novel recombinant vaccinia virus expressing the human norepinephrine transporter retains oncolytic potential and facilitates deep-tissue imaging. Mol Med. 2009;15(5–6):144–151. | ||
McCart JA, Mehta N, Scollard D, et al. Oncolytic vaccinia virus expressing the human somatostatin receptor SSTR2: molecular imaging after systemic delivery using 111In-pentetreotide. Mol Ther. 2004;10(3):553–561. |
 © 2017 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2017 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.