Back to Journals » Drug Design, Development and Therapy » Volume 9
Noncoding RNAs as potential biomarkers to predict the outcome in pancreatic cancer
Authors Jin K, Luo G, Xiao Z, Liu Z, Liu C, Ji S, Xu J, Liu L, Long J, Ni Q, Yu X
Received 16 November 2014
Accepted for publication 30 December 2014
Published 26 February 2015 Volume 2015:9 Pages 1247—1255
DOI https://doi.org/10.2147/DDDT.S77597
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Shu-Feng Zhou
Kaizhou Jin,1–3,* Guopei Luo,1–3,* Zhiwen Xiao,1–3 Zuqiang Liu,1–3 Chen Liu,1–3 Shunrong Ji,1–3 Jin Xu,1–3 Liang Liu,1–3 Jiang Long,1–3 Quanxing Ni,1–3 Xianjun Yu1–3
1Department of Pancreatic and Hepatobiliary Surgery, Fudan University Shanghai Cancer Center, 2Department of Oncology, Shanghai Medical College, Fudan University, 3Pancreatic Cancer Institute, Fudan University, Shanghai, People’s Republic of China
*These authors contributed equally to this work
Abstract: Pancreatic ductal adenocarcinoma (PDAC), a common digestive system cancer, is highly malignant and has a poor disease outcome. Currently, all available examination and detection methods cannot accurately predict the clinical outcome. Therefore, it is extremely important to identify novel molecular biomarkers for personalized medication and to significantly improve the overall outcome. The “noncoding RNAs” (ncRNAs) are a group of RNAs that do not code for proteins, and they are categorized as structural RNAs and regulatory RNAs. It has been shown that microRNAs and long ncRNAs function as regulatory RNAs to affect the progression of various diseases. Many studies have confirmed a role for ncRNAs in the progression of PDAC during the last few years. Because of the significant role of ncRNAs in PDAC, ncRNA profiling may be used to predict PDAC outcome with high accuracy. This review comprehensively analyzes the value of ncRNAs as potential biomarkers to predict the outcome in PDAC and the possible mechanisms thereof.
Keywords: pancreatic ductal adenocarcinoma, microRNA, long noncoding RNA, outcome prediction
Introduction
Pancreatic ductal adenocarcinoma
Pancreatic ductal adenocarcinoma (PDAC), a common digestive tumor, is highly malignant and has a poor patient outcome. The majority of patients have metastasis at diagnosis, and only approximately 15% of the tumors can be surgically removed.1 Despite the refinement of surgical techniques for treatment and the constant modification of surgical approaches for improving the resection rate, the median survival time of PDAC is only 13.4 months after resection combined with gemcitabine.2 For patients who are not candidates for curative surgery, the median survival time is only 7 months.3
Outcome prediction in pancreatic ductal adenocarcinoma
Pathological stage is the most important tool used to evaluate the outcome and predict the necessity and validity for adjuvant treatments in PDAC. Additionally, in the metastatic setting, only clinical prognostic factors, such as imaging examinations and serum tumor markers, are available to guide treatment decision making.4–6
Carbohydrate antigen 19-9 (CA19-9) has been extensively used to screen for PDAC in blood and predict the clinical cancer outcome.7,8 However, several benign diseases and multiple types of digestive adenocarcinoma, particularly advanced gastrointestinal cancers, may cause elevated serum CA19-9 levels, which suggests that the specificity of CA19-9 requires improvement for PDAC outcome predication.9 In the majority of patients with benign biliary obstruction, CA-19-9 levels are significantly elevated (>1,000 U/mL).10 Therefore, it is difficult to distinguish whether high preoperative levels of CA19-9 are related to tumor recurrence or other diseases.
Furthermore, PDAC is heterogeneous both in biology and genetics. Promising tools for stratifying patients according to tumor biological behaviors and aberrant genetic alterations are required. Moreover, the majority of patients with PDAC remain asymptomatic until they present with locally advanced or distally metastatic and surgically inoperable disease.11 Therefore, rather than a PDAC tissue biomarker, blood-based biomarkers are more critical to guide PDAC patient treatment.
Noncoding RNAs
“Noncoding RNAs” (ncRNAs) are a group of RNAs that do not code for proteins. They are categorized into two classes according to transcript size: “long noncoding RNAs” (lncRNAs) are greater than 200 nucleotides in length, and “small ncRNAs” are less than 200 nucleotides in length.12–14 Among the small ncRNAs, microRNAs (miRNAs) consist of approximately 22 nucleotides and are the product of precursor miRNAs consisting of 60–110 nucleotides digested by the RNase III, Dicer.15
MicroRNAs
“miRNAs” are widely distributed small ncRNAs, primarily responsible for the negative regulation of gene expression.16 Thus far, more than 2,500 miRNAs have been reported (miRBase, v 20.0).17 The majority of miRNAs function via base-pairing with complementary sequences of the messenger RNA (mRNA) three prime untranslated region (3′UTR), degrading target gene mRNA, and inhibiting the protein translation of target genes.18 miRNAs are involved in regulating various biological activities, including cell growth, differentiation, apoptosis, and proliferation,15,19 indicating that miRNAs play an important role in the development and progression of various diseases.20
In 2002, Calin et al identified a small genomic region in chromosome 13q14 that was commonly deleted in chronic lymphocytic leukemia and contained the miR-15a and miR-16-1 genes, suggesting a link between these miRNAs and chronic lymphocytic leukemia.21 That was the first time the significance of miRNA in human cancer was demonstrated. After this initial observation, additional miRNAs were found to be aberrantly expressed in various types of cancer,22–25 which indicates that the expression of miRNAs may be a better indicator of tumor prognosis than conventional protein-coding gene arrays.23
Long noncoding RNAs
Unlike miRNAs, lncRNAs are longer in length and more diverse. They are not highly conserved, and have been considered the transcriptional “noise”, by-product of RNA polymerase II and are not biologically functional for a long period of time.26,27 However, recently, studies have revealed the significance of lncRNAs in the regulation of multiple biological processes at different levels, including chromosome modifications, transcription, and post-transcription processing.28–31
The increased knowledge of the noncoding transcriptome in humans has revealed that lncRNAs are also involved in the regulation of cancer progression. In multiple solid adenocarcinomas, evidence suggests the value of lncRNAs for clinical outcome evaluation, indicating that the lncRNA signature can also improve the outcome prediction of various cancers, including pancreatic cancer.32–36
Noncoding RNAs as circulating biomarkers in cancer
In 2008, Mitchell et al37 reported remarkable insights into circulating miRNAs as biomarkers for cancer classification and prognostication. One of the most important advantages of using circulating miRNAs as biomarkers is their stability in plasma and serum, where they are most likely protected from RNase degradation by binding to Argonaute proteins.37,38 Recent studies corroborated the potential of circulating miRNAs, which appears to extend well beyond the oncology field.39,40 The specificity and simplicity of circulating miRNAs are particularly striking, suggesting that serum-based miRNAs could emerge as promising sources of biomarker information.
miRNAs are highly stable in plasma and have outcome-predictive potential in several malignancies thus far.41,42 Therefore, it is also possible that other types of ncRNAs, such as lncRNAs, are also stable in plasma and serum. Recent studies have confirmed that lncRNAs are relatively stable in plasma samples.43,44 However, the protective mechanism remains unclear.
Noncoding RNAs in pancreatic ductal adenocarcinoma outcome prediction
MicroRNAs as biomarkers for outcome prediction in pancreatic ductal adenocarcinoma
In 2005, the expression profiling of a large number of miRNAs demonstrated that 334 miRNAs are aberrantly expressed in PDAC.23 For circulating miRNAs in the serum of PDAC patients, several groups of researchers have demonstrated the presence and potential utility of circulating miRNAs (Table 1). Furthermore, the role of circulating miRNAs for PDAC outcome prediction has also been gradually revealed (Figure 1). Nevertheless, there are no direct correlations between circulating miRNA levels and CA19, which is not unexpected because these two factors are presumably regulated by different mechanisms.
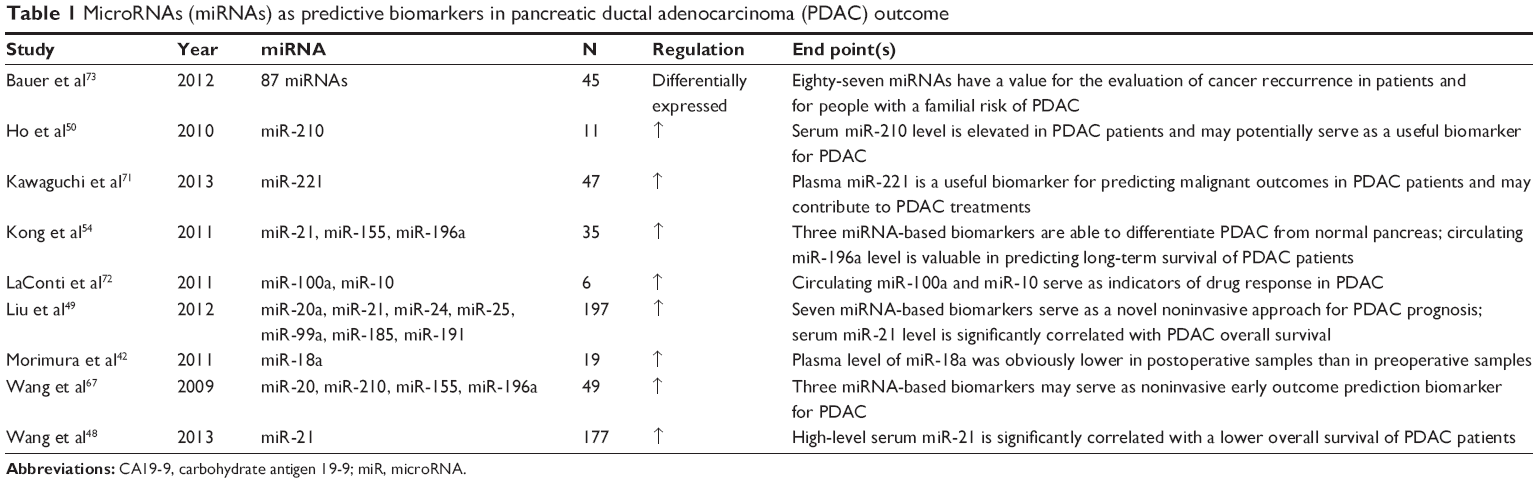 | Table 1 MicroRNAs (miRNAs) as predictive biomarkers in pancreatic ductal adenocarcinoma (PDAC) outcome |
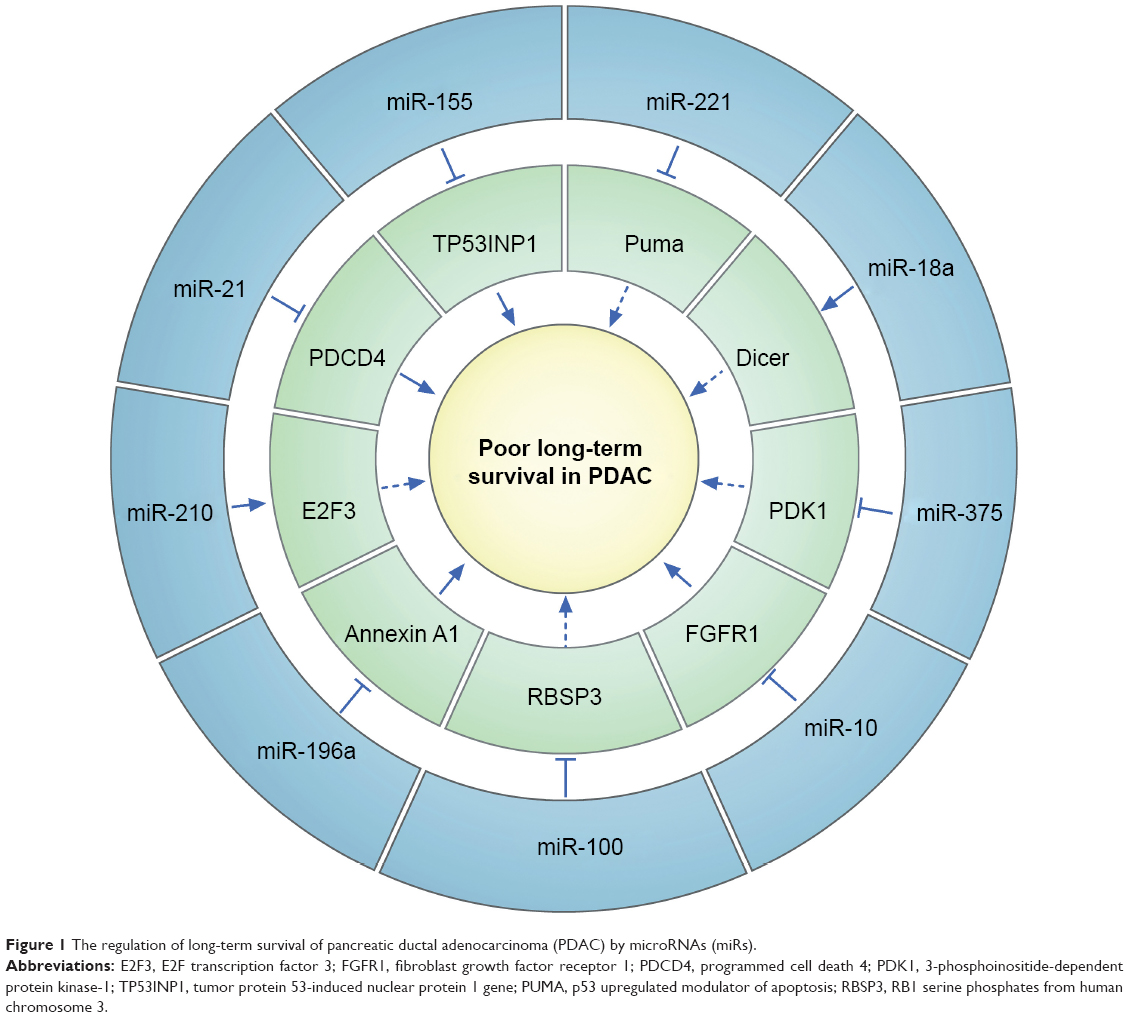 | Figure 1 The regulation of long-term survival of pancreatic ductal adenocarcinoma (PDAC) by microRNAs (miRs). |
MicroRNA-21
MicroRNA (miR)-21 is the most studied miRNA possibly related to the progression of pancreatic cancer, and this miRNA has several pancreatic cancer-related target genes, such as PDCD4, PTEN, and TIMP3.45 Of these, PDCD4, a tumor-suppressor gene, is the direct target of miR-21.46 Inhibiting the expression of miR-21 can upregulate the expression of PDCD4, reduce cell proliferation, and increase cell apoptosis simultaneously in the pancreatic cancer cell line MIA-Pa-Ca-2, indicating that miR-21 is a potential biomarker for oncological outcome and a target for treatment.47 It has also been demonstrated that gemcitabine downregulates miR-21 expression and promotes the expression of its target gene, FasL. The combination of FasL and gemcitabine promotes the apoptosis of PDAC cells.48
The abnormal expression of seven miRNAs – miR-21, miR-20a, miR-24, miR-25, miR-99a, miR-185, and miR-191 – in plasma was detected through the examination of 197 cases of PDAC.49 The positive rate of PDAC detection by the seven miRNA-based biomarkers was 96.2%, significantly higher than that of CA19-9 (46.2%) and carcinoembryonic antigen (30.8%) in the same sample set.49 These miRNAs can differentiate PDAC from pancreatitis and act as markers for predicting outcome, particularly plasma miR-21, which can function as an independent indicator for patient outcome.48,49
MicroRNA-210
As a hypoxia marker in PDAC, miR-210 was detected with fourfold increased expression in pancreatic cancer patients compared with in normal tissues (P<0.00004).50 Cancers with increased hypoxia have a poor outcome and greater resistance to chemotherapy and radiotherapy, and miR-210 likely mediates diverse epigenetic mechanisms, resulting in hypoxia.51,52 Camps et al53 showed that miR-210 in human breast cancer was not only a marker of tumor hypoxia in vivo but also an indicator of adverse outcomes clinically. Similarly, in PDAC patients, miR-210 may be a promising biomarker to predict disease outcome and sensitivity to chemotherapy.
MicroRNA-196a
The serum miR-196a level was found to have a potential value in predicting the median survival time of PDAC patients.54 Although the mechanism by which miR-196a promotes PDAC progression has not been completely elucidated, high expression of miR-196a is also associated with a poor outcome in pancreatic adenocarcinoma patients.55
Further functional analysis showed that the expression level of miR-196a in solid tumors promotes cell proliferation and suppresses cell apoptosis through downregulation of annexin 1.56 Overexpressed annexin 1 is a frequent biological marker and correlates with the differentiation of PDAC during tumorigenesis.57 These results suggest that miR-196a contributes to cancer progression and may be a prospective biomarker in PDAC.
MicroRNA-155
miR-155 is a typical multifunctional miRNA and was one of the first miRNAs found to be elevated in human cancer.58 An approximately twelvefold upregulation of the expression of miR-155 has been verified in PDAC tissues compared with in normal pancreatic tissues. miR-155 is overexpressed in PDAC cells and represses the expression of the tumor protein 53-induced nuclear protein 1 gene (TP53INP1), which is a proapoptotic stress-induced p53 target gene.59 TP53INP1 induces autophagy-dependent cell death and plays a tumor-suppressor role in multiple cancers, such as gastric cancer and pancreatic cancer.60,61 In serum, miR-155 can be used to differentiate the sera from PDAC patients from that of healthy controls.54 The ability of miR-155 to differentiate PDAC sufferers from healthy people demonstrates its utility as a first-line serum biomarker for PDAC outcome prediction.
MicroRNA-18a
MiR-18a is located in the miR-17-92 cluster and is highly expressed not only in pancreatic cancer tissues but also in patient serum.42,62 This miRNA was reported as a potential oncogene in various types of cancer, with its elevation caused by genome amplification or transcriptional activation by c-myc.63 c-myc alters the sensitivity of chemotherapeutic drugs by affecting the expression of the cell-cyclin protein cyclin D1.64 Transcriptional activation of cyclin D1 and c-myc induces the Gap 1- to Synthesis-phase transition in the cell cycle and results in DNA replication and ultimately mitosis, which are responsible for cell proliferation.65
A recent study showed the prognostic value of miR-18a in PDAC. MiR-18a was significantly lower in postoperative samples than preoperative samples (P=0.0077).42 Therefore, the relationships between miR-18a, c-myc, and cyclin D1 suggest that miR-18a is a potential outcome-predictive miRNA in PDAC.
Combination of microRNAs
miRNAs as a whole can significantly increase the reliability of outcome prediction. The expression of miR-21, miR-210, miR-155, and miR-196a in plasma has been shown to be increased in PDAC patients.66 Receiver operating characteristic curve analysis has shown that the specificity and sensitivity were significantly higher through miRNA combination, with values of 89% and 64%, respectively.67
A single gene or protein may not accurately predict patient outcome and guide treatment. The reliability is significantly increased if several miRNAs can predict the outcome. Other studies have also demonstrated that the putative target genes for these predictive miRNAs are involved in complex signaling networks that can affect pancreatic cancer tumor progression and predict disease outcome.68–70
Other microRNAs
Other circulating miRNAs, such as miR-10, miR-34b, miR-100a, miR-221, and miR-375, have also been confirmed as valuable biomarkers for predicting the clinical outcome in PDAC patients via the regulation of their target genes.71–73
Long noncoding RNAs as biomarkers for outcome prediction in pancreatic ductal adenocarcinoma
As yet, there is no direct evidence that circulating lncRNAs are molecular markers for PDAC outcome prediction; however, this has been reported in other digestive tumors, such as hepatocellular carcinoma and gastric cancer.43,74 Moreover, several lncRNAs have significant correlations with PDAC outcome and are involved in cancer progression. Therefore, these types of lncRNAs should also be investigated (Table 2).
 | Table 2 Long noncoding RNAs (lncRNAs) as predictive biomarkers in pancreatic ductal adenocarcinoma (PDAC) outcome |
Metastasis-associated lung adenocarcinoma transcript 1
Metastasis-associated lung adenocarcinoma transcript 1 (MALAT1) is the first lncRNA discovered to be related to the prognosis of PDAC. MALAT1 is located on chromosome 11q13 and was initially discovered by Ji et al75 when observing the metastatic process of non-small-cell lung cancer in 2003. Then Lin et al35 identified the negative expression of MALAT1 in the majority of adjacent normal tissues and more positive expression in PDAC tissues using in situ hybridization. In the next study, the expression of MALAT1 was also shown to be elevated in PDAC tissues, and its expression level was significantly related to the long-term survival of PDAC patients, which was considered an independent predictor of outcome.76
HOX transcript antisense RNA
HOX transcript antisense RNA (HOTAIR) is the first lncRNA found to have regulatory functions of reverse transcription.77 HOTAIR was also confirmed as related to the invasion and metastasis of solid tumors and is considered an important biomarker for disease outcome prediction.78–81 In PDAC, the higher expression of HOTAIR is related to cell invasion and tumor prognosis. Furthermore, in pancreatic cancer cell lines, HOTAIR knockout can reduce the proliferation and invasion of the tumor, and induce apoptosis, which further suggests that HOTAIR functions as both an oncogene and a negative prognostic factor.81
PVT1
Among lncRNAs, PVT1 functions as a MYC protein and oncogene, and these roles have been confirmed in various solid tumors, including Burkitt’s lymphoma, breast cancer, and lung cancer.82 In studies of PDAC cells, You et al83 used whole genome and transposon genetic screening to demonstrate that the functional inactivation of PVT1 enhanced the sensitivity of gemcitabine to the human pancreatic tumor cell line AsPDAC-1. In addition to drug sensitivity, MYC and PVT1 co-amplification is related to the rapid progression of and poor outcome in breast cancer.84 Therefore, the predictive role of PVT1 is quite promising in PDAC patients.
Other long noncoding RNAs
The mitogen-activated protein kinase (MAPK) signaling pathway is closely related to PDAC metastasis.85,86 Tahira et al87 evaluated 38 PDAC samples to examine the expression of intergenic and intronic lncRNAs in PDAC and metastatic tissues using gene microarray analysis. Their results revealed that the expression levels of nine MAPK signaling pathway-related intronic lncRNAs were significantly increased, which indicates that lncRNAs are involved in the regulation of PDAC.
Perspectives
ncRNAs are becoming potential prognostic markers in PDAC, with support from several studies (Tables 1 and 2). Several serum miRNAs, such as miR-21 and miR-196a, are negatively correlated between expression and overall survival. Additionally, several lncRNAs have also been found to be elevated in PDAC tissues, and their expression levels were significantly related to the outcome of PDAC patients. These findings suggest the potential value of ncRNAs in predicting the prognostic outcome of PDAC.
Moreover, increasing evidence suggests a role for ncRNAs in PDAC biology and progression.88 Recent studies have demonstrated that a variety of ncRNAs are frequently deregulated in PDAC and crosstalk with various biological processes, including epithelial-mesenchymal transition, cancer apoptosis regulation, and cancer-related signaling pathways, which may be crucial in tumor progression and affect patient outcome.89–93
However, as far as we are aware, no clinical trials are currently ongoing with ncRNA profiles as biomarkers in PDAC. The several studies described herein examined the predictive potential of ncRNAs in PDAC. Although they highlighted the promising roles of these molecules, four challenges remain to their clinical application: (1) The use of ncRNA for prognostic purposes requires assurance that the measured concentration represents the actual amount in the samples – however, commonly used approaches lack this assurance;94 (2) lncRNA is not conserved evolutionarily, and it is difficult to study these molecules using animal models prior to clinical applications; (3) some ncRNAs are abnormally expressed during the development and progression of various diseases, including tumors, cardiovascular diseases, and immune-system diseases, and the specificity of detection in a particular tumor requires further study;95–97 (4) PDAC is less common than other tumor types, such as lung cancer and colon cancer98 and, further, a relatively smaller proportion of patients with PDAC are candidates for resection, further diminishing the potential pool of patients with easily accessible specimens.
Conclusion
The increased understanding of the roles of ncRNAs as tumor suppressors or oncogenes will help design novel ncRNA-based approaches for outcome prediction and targeted therapy that counteract the aberrant expression of ncRNAs responsible for malignant progression. ncRNAs combined with other prognostic methods are a new direction for tumor biomarkers in the future that will allow clinicians to identify patients with the worst outcomes and then prolong survival.
Author contributions
Kaizhou Jin, Guopei Luo and Xianjun Yu designed study. Kaizhou Jin and Guopei Luo collected data. Zhiwen Xiao and Zuqiang Liu analyzed data. Chen Liu, Shunrong Ji, Jin Xu, Liang Liu, Jiang Long, Quanxing Ni and Xianjun Yu interpreted data. Kaizhou Jin, Zhiwen Xiao and Zuqiang Liu wrote the draft. Guopei Luo, Chen Liu, Shunrong Ji, Jin Xu, Liang Liu, Jiang Long, Quanxing Ni and Xianjun Yu revised it critically. All the authors approved the accuracy and integrity of the work and approved the version to be published.
Disclosure
Dr Yu has received research funding from the Sino-German Center (GZ857), the Science Foundation of Shanghai (13ZR1407500), the Shanghai Rising Star Program (12QH1400600, 14QA1400900), the Fudan University Young Investigator promoting program (20520133403), and the National Science Foundation of China (grant nos 81101807, 81001058, 81372649, 81372653, and 81172276). The authors report no other conflicts of interest in this work.
References
Stathis A, Moore MJ. Advanced pancreatic carcinoma: current treatment and future challenges. Nat Rev Clin Oncol. 2010;7(3):163–172. | ||
Zhou HY, Wang Y, Zhang J, et al. Retrograde vs Conventional Dissection Technique in Pancreatoduodenectomy: A Pilot Study. JAMA Surg. 2014. Epub April 23. | ||
Nieto J, Grossbard ML, Kozuch P. Metastatic pancreatic cancer 2008: is the glass less empty? Oncologist. 2008;13(5):562–576. | ||
Appel BL, Tolat P, Evans DB, Tsai S. Current staging systems for pancreatic cancer. Cancer J. 2012;18(6):539–549. | ||
Wong D, Ko AH, Hwang J, Venook AP, Bergsland EK, Tempero MA. Serum CA19-9 decline compared to radiographic response as a surrogate for clinical outcomes in patients with metastatic pancreatic cancer receiving chemotherapy. Pancreas. 2008;37(3):269–274. | ||
Liu L, Xu H, Wang W, et al. A preoperative serum signature of CEA+/CA125+/CA19-9≥1,000 U/mL indicates poor outcome to pancreatectomy for pancreatic cancer. Int J Cancer. 2014. Epub October 1. | ||
Locker GY, Hamilton S, Harris J, et al. ASCO 2006 update of recommendations for the use of tumor markers in gastrointestinal cancer. J Clin Oncol. 2006;24(33):5313–5327. | ||
Poruk KE, Gay DZ, Brown K, et al. The clinical utility of CA 19-9 in pancreatic adenocarcinoma: diagnostic and prognostic updates. Curr Mol Med. 2013;13(3):340–351. | ||
Duffy MJ, Sturgeon C, Lamerz R, et al. Tumor markers in pancreatic cancer: a European Group on Tumor Markers (EGTM) status report. Ann Oncol. 2010;21(3):441–447. | ||
Marrelli D, Caruso S, Pedrazzani C, et al. CA19-9 serum levels in obstructive jaundice: clinical value in benign and malignant conditions. Am J Surg. 2009;198(3):333–339. | ||
Bond-Smith G, Banga N, Hammond TM, et al. Pancreatic adenocarcinoma. BMJ. 2012;344(e2476). | ||
Mattick JS. Non-coding RNAs: the architects of eukaryotic complexity. EMBO Rep. 2001;2(11):986–991. | ||
Brosnan CA, Voinnet O. The long and the short of noncoding RNAs. Curr Opin Cell Biol. 2009;21(3):416–425. | ||
Costa FF. Non-coding RNAs: new players in eukaryotic biology. Gene. 2005;357(2):83–94. | ||
Slack FJ, Weidhaas JB. MicroRNA in cancer prognosis. N Engl J Med. 2008;359(25):2720–2722. | ||
Lim LP, Lau NC, Garrett-Engele P, et al. Microarray analysis shows that some microRNAs downregulate large numbers of target mRNAs. Nature. 2005;433(7027):769–773. | ||
miRBase: the microRNA database [database on the Internet]. Version 20. Manchester: University of Manchester Faculty of Life Sciences Griffiths-Jones Lab; nd. Available from: http://www.mirbase.org/. Accessed June 11, 2014. | ||
Lewis BP, Burge CB, Bartel DP. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell. 2005;120(1):15–20. | ||
Xiong J. Emerging roles of microRNA-22 in human disease and normal physiology. Curr Mol Med. 2012;12(3):247–258. | ||
Mo YY. MicroRNA regulatory networks and human disease. Cell Mol Life Sci. 2012;69(21):3529–3531. | ||
Calin GA, Dumitru CD, Shimizu M, et al. Frequent deletions and down-regulation of micro-RNA genes miR15 and miR16 at 13q14 in chronic lymphocytic leukemia. Proc Natl Acad Sci U S A. 2002;99(24):15524–15529. | ||
Volinia S, Calin GA, Liu CG, et al. A microRNA expression signature of human solid tumors defines cancer gene targets. Proc Natl Acad Sci U S A. 2006;103(7):2257–2261. | ||
Lu J, Getz G, Miska EA, et al. MicroRNA expression profiles classify human cancers. Nature. 2005;435(7043):834–838. | ||
Liu C, Cheng H, Shi S, et al. MicroRNA-34b inhibits pancreatic cancer metastasis through repressing Smad3. Curr Mol Med. 2013;13(4):467–478. | ||
Chen Z, Li J, Tian L, et al. MiRNA expression profile reveals a prognostic signature for esophageal squamous cell carcinoma. Cancer Lett. 2014;350(1–2):34–42. | ||
Ponting CP, Oliver PL, Reik W. Evolution and functions of long noncoding RNAs. Cell. 2009;136(4):629–641. | ||
Jia H, Osak M, Bogu GK, Stanton LW, Johnson R, Lipovich L. Genome-wide computational identification and manual annotation of human long noncoding RNA genes. RNA. 2010;16(8):1478–1487. | ||
Flynn RA, Chang HY. Active chromatin and noncoding RNAs: an intimate relationship. Curr Opin Genet Dev. 2012;22(2):172–178. | ||
Kung JT, Colognori D, Lee JT. Long noncoding RNAs: past, present, and future. Genetics. 2013;193(3):651–669. | ||
Hung T, Chang HY. Long noncoding RNA in genome regulation: prospects and mechanisms. RNA Biol. 2010;7(5):582–585. | ||
Mercer TR, Dinger ME, Mattick JS. Long non-coding RNAs: insights into functions. Nat Rev Genet. 2009;10(3):155–159. | ||
Hu Y, Chen HY, Yu CY, et al. A long non-coding RNA signature to improve prognosis prediction of colorectal cancer. Oncotarget. 2014;5(8):2230–2242. | ||
Sioss JA, Bhiladvala RB, Pan W, et al. Nanoresonator chip-based RNA sensor strategy for detection of circulating tumor cells: response using PCA3 as a prostate cancer marker. Nanomedicine. 2012;8(6):1017–1025. | ||
Quagliata L, Matter MS, Piscuoglio S, et al. Long noncoding RNA HOTTIP/HOXA13 expression is associated with disease progression and predicts outcome in hepatocellular carcinoma patients. Hepatology. 2014;59(3):911–923. | ||
Lin R, Maeda S, Liu C, Karin M, Edgington TS. A large noncoding RNA is a marker for murine hepatocellular carcinomas and a spectrum of human carcinomas. Oncogene. 2007;26(6):851–858. | ||
Cheng W, Zhang Z, Wang J. Long noncoding RNAs: new players in prostate cancer. Cancer Lett. 2013;339(1):8–14. | ||
Mitchell PS, Parkin RK, Kroh EM, et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc Natl Acad Sci U S A. 2008;105(30):10513–10518. | ||
Arroyo JD, Chevillet JR, Kroh EM, et al. Argonaute2 complexes carry a population of circulating microRNAs independent of vesicles in human plasma. Proc Natl Acad Sci U S A. 2011;108(12):5003–5008. | ||
Kodahl AR, Lyng MB, Binder H, et al. Novel circulating microRNA signature as a potential non-invasive multi-marker test in ER-positive early-stage breast cancer: a case control study. Mol Oncol. 2014;8(5):874–883. | ||
Sun Y, Zhang K, Fan G, Li J. Identification of circulating microRNAs as biomarkers in cancers: what have we got. Clin Chem Lab Med. 2012;50(12):2121–2126. | ||
Ng EK, Chong WW, Jin H, et al. Differential expression of microRNAs in plasma of patients with colorectal cancer: a potential marker for colorectal cancer screening. Gut. 2009;58(10):1375–1381. | ||
Morimura R, Komatsu S, Ichikawa D, et al. Novel diagnostic value of circulating miR-18a in plasma of patients with pancreatic cancer. Br J Cancer. 2011;105(11):1733–1740. | ||
Arita T, Ichikawa D, Konishi H, et al. Circulating long non-coding RNAs in plasma of patients with gastric cancer. Anticancer Res. 2013;33(8):3185–3193. | ||
Panzitt K, Tschernatsch MM, Guelly C, et al. Characterization of HULC, a novel gene with striking up-regulation in hepatocellular carcinoma, as noncoding RNA. Gastroenterology. 2007;132(1):330–342. | ||
Krichevsky AM, Gabriely G. miR-21: a small multi-faceted RNA. J Cell Mol Med. 2009;13(1):39–53. | ||
Frankel LB, Christoffersen NR, Jacobsen A, Lindow M, Krogh A, Lund AH. Programmed cell death 4 (PDCD4) is an important functional target of the microRNA miR-21 in breast cancer cells. J Biol Chem. 2008;283(2):1026–1033. | ||
Bhatti I, Lee A, James V, et al. Knockdown of microRNA-21 inhibits proliferation and increases cell death by targeting programmed cell death 4 (PDCD4) in pancreatic ductal adenocarcinoma. J Gastrointest Surg. 2011;15(1):199–208. | ||
Wang P, Zhuang L, Zhang J, et al. The serum miR-21 level serves as a predictor for the chemosensitivity of advanced pancreatic cancer, and miR-21 expression confers chemoresistance by targeting FasL. Mol Oncol. 2013;7(3):334–345. | ||
Liu R, Chen X, Du Y, et al. Serum microRNA expression profile as a biomarker in the diagnosis and prognosis of pancreatic cancer. Clin Chem. 2012;58(3):610–618. | ||
Ho AS, Huang X, Cao H, et al. Circulating miR-210 as a Novel Hypoxia Marker in Pancreatic Cancer. Transl Oncol. 2010;3(2):109–113. | ||
Huang X, Ding L, Bennewith KL, et al. Hypoxia-inducible mir-210 regulates normoxic gene expression involved in tumor initiation. Mol Cell. 2009;35(6):856–867. | ||
Kulshreshtha R, Ferracin M, Wojcik SE, et al. A microRNA signature of hypoxia. Mol Cell Biol. 2007;27(5):1859–1867. | ||
Camps C, Buffa FM, Colella S, et al. hsa-miR-210 Is induced by hypoxia and is an independent prognostic factor in breast cancer. Clin Cancer Res. 2008;14(5):1340–1348. | ||
Kong X, Du Y, Wang G, et al. Detection of differentially expressed microRNAs in serum of pancreatic ductal adenocarcinoma patients: miR-196a could be a potential marker for poor prognosis. Dig Dis Sci. 2011;56(2):602–609. | ||
Bloomston M, Frankel WL, Petrocca F, et al. MicroRNA expression patterns to differentiate pancreatic adenocarcinoma from normal pancreas and chronic pancreatitis. JAMA. 2007;297(17):1901–1908. | ||
Luthra R, Singh RR, Luthra MG, et al. MicroRNA-196a targets annexin A1: a microRNA-mediated mechanism of annexin A1 downregulation in cancers. Oncogene. 2008;27(52):6667–6678. | ||
Bai XF, Ni XG, Zhao P, et al. Overexpression of annexin 1 in pancreatic cancer and its clinical significance. World J Gastroenterol. 2004;10(10):1466–1470. | ||
Kluiver J, Poppema S, de Jong D, et al. BIC and miR-155 are highly expressed in Hodgkin, primary mediastinal and diffuse large B cell lymphomas. J Pathol. 2005;207(2):243–249. | ||
Gironella M, Seux M, Xie MJ, et al. Tumor protein 53-induced nuclear protein 1 expression is repressed by miR-155, and its restoration inhibits pancreatic tumor development. Proc Natl Acad Sci U S A. 2007;104(41):16170–16175. | ||
Seillier M, Peuget S, Gayet O, et al. TP53INP1, a tumor suppressor, interacts with LC3 and ATG8-family proteins through the LC3-interacting region (LIR) and promotes autophagy-dependent cell death. Cell Death Differ. 2012;19(9):1525–1535. | ||
Jiang PH, Motoo Y, Sawabu N, Minamoto T. Effect of gemcitabine on the expression of apoptosis-related genes in human pancreatic cancer cells. World J Gastroenterol. 2006;12(10):1597–1602. | ||
Szafranska AE, Davison TS, John J, et al. MicroRNA expression alterations are linked to tumorigenesis and non-neoplastic processes in pancreatic ductal adenocarcinoma. Oncogene. 2007;26(30):4442–4452. | ||
Woods K, Thomson JM, Hammond SM. Direct regulation of an oncogenic micro-RNA cluster by E2F transcription factors. J Biol Chem. 2007;282(4):2130–2134. | ||
Biliran H Jr, Banerjee S, Thakur A, et al. c-Myc-induced chemosensitization is mediated by suppression of cyclin D1 expression and nuclear factor-kappa B activity in pancreatic cancer cells. Clin Cancer Res. 2007;13(9):2811–2821. | ||
Kaldis P, Pagano M. Wnt signaling in mitosis. Dev Cell. 2009;17(6):749–750. | ||
Wang J, Chen J, Chang P, et al. MicroRNAs in plasma of pancreatic ductal adenocarcinoma patients as novel blood-based biomarkers of disease. Cancer Prev Res (Phila). 2009;2(9):807–813. | ||
Wang J, Chen J, Chang P, et al. MicroRNAs in plasma of pancreatic ductal adenocarcinoma patients as novel blood-based biomarkers of disease. Cancer Prev Res (Phila). 2009;2(9):807–813. | ||
Frampton AE, Castellano L, Colombo T, et al. MicroRNAs cooperatively inhibit a network of tumor suppressor genes to promote pancreatic tumor growth and progression. Gastroenterology. 2014;146(1):268–277.e18. | ||
Schultz NA, Andersen KK, Roslind A, Willenbrock H, Wojdemann M, Johansen JS. Prognostic microRNAs in cancer tissue from patients operated for pancreatic cancer – five microRNAs in a prognostic index. World J Surg. 2012;36(11):2699–2707. | ||
Greither T, Grochola LF, Udelnow A, Lautenschlager C, Wurl P, Taubert H. Elevated expression of microRNAs 155, 203, 210 and 222 in pancreatic tumors is associated with poorer survival. Int J Cancer. 2010;126(1):73–80. | ||
Kawaguchi T, Komatsu S, Ichikawa D, et al. Clinical impact of circulating miR-221 in plasma of patients with pancreatic cancer. Br J Cancer. 2013;108(2):361–369. | ||
LaConti JJ, Shivapurkar N, Preet A, et al. Tissue and serum microRNAs in the Kras(G12D) transgenic animal model and in patients with pancreatic cancer. PLoS One. 2011;6(6):e20687. | ||
Bauer AS, Keller A, Costello E, et al. Diagnosis of pancreatic ductal adenocarcinoma and chronic pancreatitis by measurement of microRNA abundance in blood and tissue. PLoS One. 2012;7(4):e34151. | ||
Xie H, Ma H, Zhou D. Plasma HULC as a promising novel biomarker for the detection of hepatocellular carcinoma. Biomed Res Int. 2013;2013:136106. | ||
Ji P, Diederichs S, Wang W, et al. MALAT-1, a novel noncoding RNA, and thymosin beta4 predict metastasis and survival in early-stage non-small cell lung cancer. Oncogene. 2003;22(39):8031–8041. | ||
Liu JH, Chen G, Dang YW, Li CJ, Luo DZ. Expression and prognostic significance of lncRNA MALAT1 in pancreatic cancer tissues. Asian Pac J Cancer Prev. 2014;15(7):2971–2977. | ||
Rinn JL, Kertesz M, Wang JK, et al. Functional demarcation of active and silent chromatin domains in human HOX loci by noncoding RNAs. Cell. 2007;129(7):1311–1323. | ||
Kogo R, Shimamura T, Mimori K, et al. Long noncoding RNA HOTAIR regulates polycomb-dependent chromatin modification and is associated with poor prognosis in colorectal cancers. Cancer Res. 2011;71(20):6320–6326. | ||
Gupta RA, Shah N, Wang KC, et al. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature. 2010;464(7291):1071–1076. | ||
Yang Z, Zhou L, Wu LM, et al. Overexpression of long non-coding RNA HOTAIR predicts tumor recurrence in hepatocellular carcinoma patients following liver transplantation. Ann Surg Oncol. 2011;18(5):1243–1250. | ||
Kim K, Jutooru I, Chadalapaka G, et al. HOTAIR is a negative prognostic factor and exhibits pro-oncogenic activity in pancreatic cancer. Oncogene. 2013;32(13):1616–1625. | ||
Pomerantz MM, Beckwith CA, Regan MM, et al. Evaluation of the 8q24 prostate cancer risk locus and MYC expression. Cancer Res. 2009;69(13):5568–5574. | ||
You L, Chang D, Du HZ, Zhao YP. Genome-wide screen identifies PVT1 as a regulator of Gemcitabine sensitivity in human pancreatic cancer cells. Biochem Biophys Res Commun. 2011;407(1):1–6. | ||
Borg A, Baldetorp B, Ferno M, Olsson H, Sigurdsson H. c-myc amplification is an independent prognostic factor in postmenopausal breast cancer. Int J Cancer. 1992;51(5):687–691. | ||
Campagna D, Cope L, Lakkur SS, Henderson C, Laheru D, Iacobuzio-Donahue CA. Gene expression profiles associated with advanced pancreatic cancer. Int J Clin Exp Pathol. 2008;1(1):32–43. | ||
Zhao Y, Shen S, Guo J, et al. Mitogen-activated protein kinases and chemoresistance in pancreatic cancer cells. J Surg Res. 2006;136(2):325–335. | ||
Tahira AC, Kubrusly MS, Faria MF, et al. Long noncoding intronic RNAs are differentially expressed in primary and metastatic pancreatic cancer. Mol Cancer. 2011;10:141. | ||
Shi S, Lu Y, Qin Y, et al. miR-1247 is correlated with prognosis of pancreatic cancer and inhibits cell proliferation by targeting neuropilins. Curr Mol Med. 2014;14(3):316–327. | ||
Arlt A, Muerkoster SS, Schafer H. Targeting apoptosis pathways in pancreatic cancer. Cancer Lett. 2013;332(2):346–358. | ||
Guo S, Xu X, Tang Y, et al. miR-15a inhibits cell proliferation and epithelial to mesenchymal transition in pancreatic ductal adenocarcinoma by down-regulating Bmi-1 expression. Cancer Lett. 2014;344(1):40–46. | ||
Deng S, Zhu S, Wang B, et al. Chronic pancreatitis and pancreatic cancer demonstrate active epithelial-mesenchymal transition profile, regulated by miR-217-SIRT1 pathway. Cancer Lett. 2014;355(2):184–191. | ||
Patel K, Kollory A, Takashima A, Sarkar S, Faller DV, Ghosh SK. MicroRNA let-7 downregulates STAT3 phosphorylation in pancreatic cancer cells by increasing SOCS3 expression. Cancer Lett. 2014;347(1):54–64. | ||
Zhang Y, Yang J, Cui X, et al. A novel epigenetic CREB-miR-373 axis mediates ZIP4-induced pancreatic cancer growth. EMBO Mol Med. 2013;5(9):1322–1334. | ||
Moltzahn F, Olshen AB, Baehner L, et al. Microfluidic-based multiplex qRT-PCR identifies diagnostic and prognostic microRNA signatures in the sera of prostate cancer patients. Cancer Res. 2011;71(2):550–560. | ||
Liang D, Xu X, Deng F, et al. miRNA-940 reduction contributes to human Tetralogy of Fallot development. J Cell Mol Med. 2014;18(9):1830–1839. | ||
Li F, Ma N, Zhao R, et al. Overexpression of miR-483-5p/3p cooperate to inhibit mouse liver fibrosis by suppressing the TGF-beta stimulated HSCs in transgenic mice. J Cell Mol Med. 2014;18(6):966–974. | ||
Liu Z, Chen L, Zhang X, et al. RUNX3 regulates vimentin expression via miR-30a during epithelial-mesenchymal transition in gastric cancer cells. J Cell Mol Med. 2014;18(4):610–623. | ||
Siegel R, Ma J, Zou Z, Jemal A. Cancer statistics, 2014. CA Cancer J Clin. 2014;64(1):9–29. |
 © 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.