Back to Journals » Pharmacogenomics and Personalized Medicine » Volume 13
Negative Association Between lncRNA HOTTIP rs3807598 C>G and Hirschsprung Disease
Authors Zheng Y, Zhuo Z, Xie X, Lu L, He Q, Zhong W
Received 13 February 2020
Accepted for publication 22 April 2020
Published 6 May 2020 Volume 2020:13 Pages 151—156
DOI https://doi.org/10.2147/PGPM.S249649
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Martin H Bluth
Yi Zheng,* Zhenjian Zhuo,* Xiaoli Xie, Lifeng Lu, Qiuming He, Wei Zhong
Department of Pediatric Surgery, Guangzhou Institute of Pediatrics, Guangdong Provincial Key Laboratory of Research in Structural Birth Defect Disease, Guangzhou Women and Children’s Medical Center, Guangzhou Medical University, Guangzhou 510623, Guangdong, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Wei Zhong
Department of Pediatric Surgery, Guangzhou Institute of Pediatrics, Guangdong Provincial Key Laboratory of Research in Structural Birth Defect Disease, Guangzhou Women and Children’s Medical Center, Guangzhou Medical University, 9 Jinsui Road, Guangzhou 510623, Guangdong, People’s Republic of China
Tel/Fax +86-18902268667
Email [email protected]
Background: Hirschsprung disease (HSCR) is a congenital disease that arises from defective intestinal neural system. LncRNA HOTTIP is a critical gene in various diseases, including HSCR. No epidemiological studies have explored the correlation between lncRNA HOTTIP single nucleotide polymorphisms (SNPs) and HSCR risk. We here lead as a pioneer to explore whether SNPs in lncRNA HOTTIP impact the risk of HSCR and HSCR subtypes in an unrelated Chinese population.
Methods: We used the TaqMan method to genotype rs3807598 C>G of the lncRNA HOTTIP gene using 1470 HSCR cases and 1473 healthy controls. Of them, 1441 cases and 1434 controls were successfully genotyped. We adopted odds ratios (ORs) and 95% confidence intervals (CIs) to quantify the relationship.
Results: We got an unexpected outcome that lncRNA HOTTIP SNP rs3807598 C>G could not modify the risk of HSCR (CG vs. CC: adjusted OR=0.89, 95% CI=0.74– 1.07; GG vs. CC: adjusted OR=1.10, 95% CI=0.89– 1.37; GG/CG vs CC: adjusted OR=0.95, 95% CI=0.80– 1.13; and GG vs. CC/CG: adjusted OR=1.19, 95% CI=0.99– 1.43). What’s more, risk effect of lncRNA HOTTIP rs3807598 C>G is still not obvious in stratification analysis by HSCR subtype.
Conclusion: Our studies did not provide statistical evidence of a correlation between lncRNA HOTTIP SNP rs3807598 C>G and susceptibility of HSCR in the Chinese population that is being studied. Further validation study with a larger sample size covering multi-ethnic groups is warranted.
Keywords: Hirschsprung disease, HOTTIP, polymorphism, susceptibility, Chinese
Introduction
Hirschsprung disease (HSCR) is the most frequent disorder of the enteric nervous system (ENS) in children.1 The prevalence of HSCR is about 1:2000–1:5000 in live births, with a 4:1 male predominance.2 Most of the HSCR are originated from defective migration and maturation of intestinal vagal neural cells during embryonal development.3 In over 80% of cases, aganglionosis is confined to the rectosigmoid colon (short-segment HSCR, S-HSCR).4 In another 20% of cases, aganglionosis can also affect significant lengths of the colon or even extend proximal to the sigmoid (long segment HSCR, L-HSCR). Extremely rarely (<5% of cases), aganglionosis affects longer tracts of the colon up to a total colonic aganglionosis (TCA) or even total intestinal aganglionosis (TIA).5 About 80% of patients present a phenotype as isolated or non-syndromic. The rest of them present a phenotype as syndromic which are accompanied by syndromes, such as Mowat-Wilson syndrome, or chromosomal abnormalities, such as trisomy 21, or other congenital abnormalities.6
Genetic variations are reported to be closely associated with predisposition to HSCR.7–9 Most of the genetic abnormalities in HSCR are found in genes that are known to be a necessity for the proliferation and maintenance of enteric neural crest cell (ENCC) and enteric nervous system (ENS) progenitors.10 RET gene mutations are the most commonly known genetic causes of HSCR, and they take up approximately 50% of familial HSCR and 15–20% of sporadic HSCR.11–13 More than 100 mutations in the RET gene have been identified in HSCR.14 Simple genetic mutations of the RET gene can cause HSCR by haploinsufficiency. Other major genes implicated in isolated HSCR include EDNRB,15 EDN3,16 NRG1,17,18 PHOX2B,19 and SOX10.20 In addition to the mentioned above, genetic variants in a dozen or more genes were found to predispose to HSCR.21,22 Despite the unequivocal implication of these important gene single-nucleotide polymorphisms (SNPs) in HSCR risk, additional SNPs with a modest effect await to be found to fully reveal the etiology of the disease.
Long non-coding RNAs (lncRNAs) are non-protein-coding RNAs that consist of over 200 nucleotides.23 More than 28,000 lncRNAs are encoded by the mammalian genome, according to the Encyclopedia of DNA Elements Project Consortium (GENCODE release 23). LncRNAs are presumed to have been implicated in various diseases by regulating diverse functions, including epigenetic, transcriptional and posttranscriptional regulation.24 Plenty of lncRNAs have been found to associate with the onset of the HSCR.25–27 LncRNA HOTTIP, short for HOXA transcript at the distal tip, is an enhancer-like lncRNA of the human HOXA locus.28 HOTTIP possesses great potentiality as a critical regulator in various diseases, including HSCR. SNPs, the most common type of genetic variation, affect the functions of lncRNAs by altering the secondary structure of lncRNAs or the expression of their coding genes.29 Indeed, recent efforts for the lncRNA HOTTIP gene identified several SNPs with strong effects on disease risk. However, the relevance of the vital lncRNA HOTTIP gene SNPs to HSCR risk has not been reported so far. Considering the important role of lncRNA HOTTIP in various diseases, it is reasonable to investigate lncRNA HOTTIP gene SNPs to add a bit evidence to fully unveil the genetic spectrum of HSCR. HSCR is a heterogenous disease, and different patients may be diagnosed with varied clinical manifestations. Each specific type of HSCR may have a specific genetic background. Therefore, it is of necessity to further perform subgroup detections in HSCR.
To shed light on this topic, we explored the correlation between lncRNA HOTTIP SNPs and the risk of HSCR using a set of 1470 HSCR cases and 1473 controls. Our findings aid in understanding genetic architectures and the etiology of HSCR.
Materials and Methods
Study Subjects
This research project was approved by the institutional review board of Guangzhou Women and Children’s medical center. Written informed consent was obtained from each subject’s guardians prior to recruitment. 1470 cases diagnosed with HSCR by pathological analysis after surgery were enrolled. According to the segment lengths of aganglionosis, recruited cases were divided into three subgroups: S-HSCR (1033), L-HSCR (294) and TCA (82). 1473 healthy controls without HSCR and neurological related diseases were recruited from the same region as cases. Relevant subjects’ descriptions with details could be found in the previously published studies.30,31 This study was conducted in accordance with the Declaration of Helsinki.
Genotyping
The potentially functional SNPs were selected by using the NCBI dbSNP database and SNPinfo (http://snpinfo.niehs.nih.gov/snpinfo/snpfunc.htm). The following criteria were applied: (1) the minor allele frequency (MAF) reported in HapMap was > 5% for Chinese Han subjects; (2) putative functional potentials SNPs which might affect transcription activity or binding capacity of the microRNA binding site; (3) SNPs covering the exons, introns, 3ʹ-, and 5ʹ- flanking sequences. Based on the criteria, two SNPs rs3807598 C>G and rs1859168 A>C of HOTTIP were selected. rs1859168 A>C was not included in the final genotyping due to its linkage disequilibrium (R2=0.647). Genomic DNA was extracted from participants’ blood using a TIANamp Blood DNA Kit (TianGen Biotech Co. Ltd., Beijing, China). SNP genotyping of rs3807598 C>G was conducted with TaqMan assay following the standard procedures.32–35 We additionally re-genotyped 10% of the samples. A 100% concordant rate indicates the reliability of the genotyping data.
Statistical Analysis
Hardy–Weinberg equilibrium (HWE) test was performed for rs3807598 C>G in controls using a χ2 test. First, we tested the differences of clinical characteristics in cases and controls using a χ2 test. We used multivariable logistic regression analysis to test for an association between SNP rs3807598 C>G and the risk of HSCR. Such association was quantified by odds ratios (ORs) and 95% confidence intervals (CIs). Two-sided P < 0.05 indicates differences were statistically significant. We did the statistical analyses using the SAS release 9.1 (SAS Institute, Cary, NC).
Results
Association of SNP rs3807598 C>G with HSCR Risk
Clinical characteristics of the study subjects are depicted in our previous reports. Among 1470 cases and 1473 healthy controls, 1441 cases and 1434 controls were successfully genotyped for HOTTIP rs3807598 C>G. The association results for HOTTIP rs3807598 C>G and HSCR risk are provided in Table 1. The genotype frequencies for rs3807598 C>G were 27.06% (CC), 48.79% (CG) and 24.15% (GG) in cases and 25.59% (CC), 53.21% (CG) and 21.20% (GG) in controls. We found no evidence of association signals between rs3807598 C>G and HSCR risk, after adjustments for age and gender (CG vs CC: adjusted OR=0.89, 95% CI=0.74–1.07; GG vs CC: adjusted OR=1.10, 95% CI=0.89–1.37; GG/CG vs CC: adjusted OR=0.95, 95% CI=0.80–1.13; and GG vs CC/CG: adjusted OR=1.19, 95% CI=0.99–1.43).
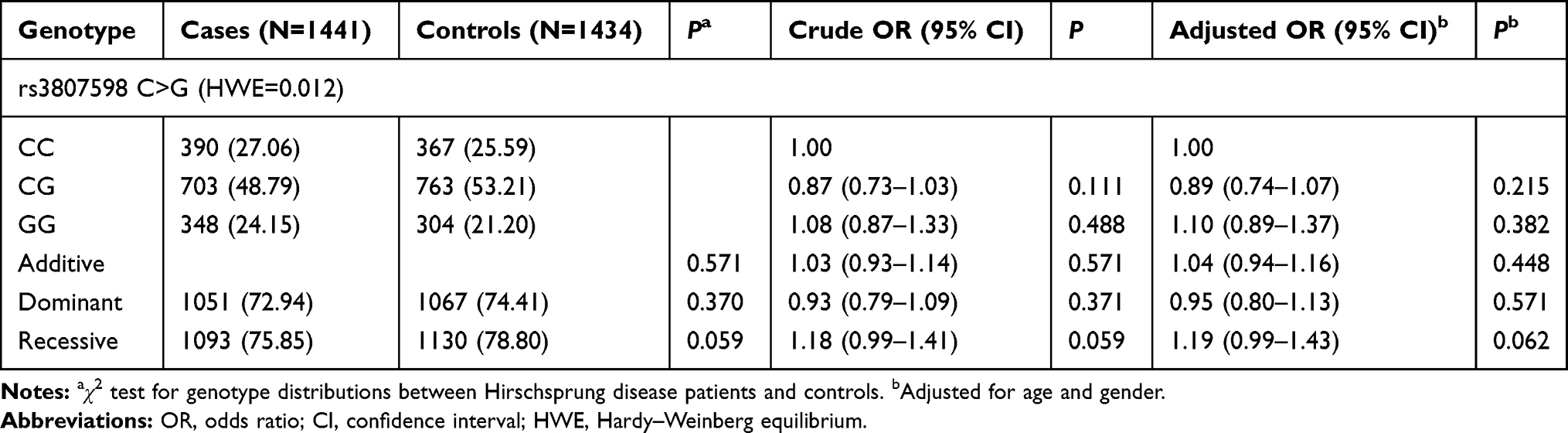 |
Table 1 Association Between HOTTIP rs3807598 C>G Polymorphism and Hirschsprung Disease Susceptibility |
Stratification Analysis
HSCR is a heterogenous disease characterized by widely clinical features, such as the aganglionosis lengths of the colon. SNPs might have played a specific role in a specific subtype of HSCR. Therefore, we assessed whether the association of rs3807598 C>G might be correlated to a specific HSCR subtype or not, ie S-HCSR, L-HCSR, or TCA (Table 2). In all subtype-stratified analyses, no significant correlations were found between rs3807598 C>G and all specific subtypes.
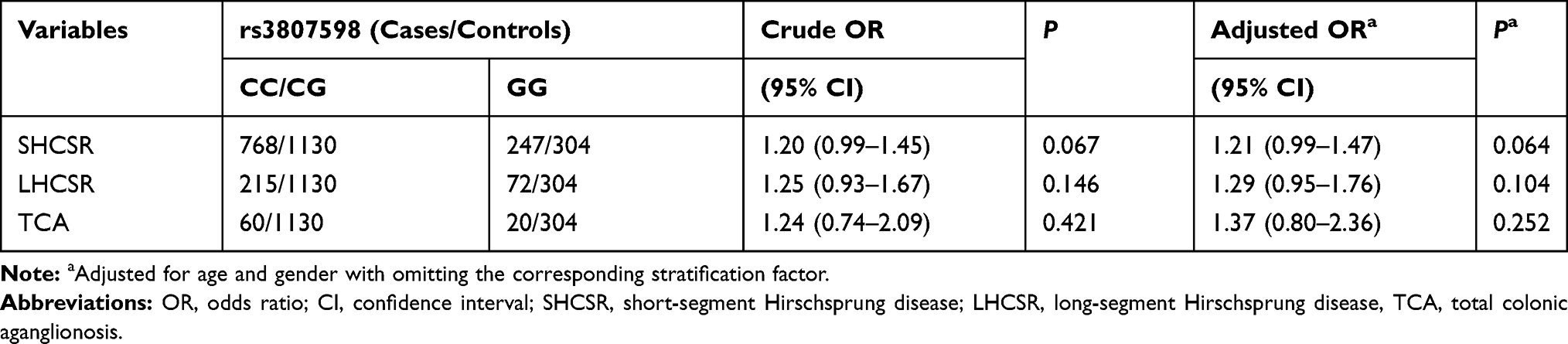 |
Table 2 Stratification Analysis for the Association Between HOTTIP rs3807598 C>G and Hirschsprung Disease Susceptibility (by Subtype) |
Discussion
With the list of newly identified lncRNAs with unknown function emerging, there is an urgent need to comprehensively dissect the role of individual lncRNA. In addition, most research paid close attention to the downstream functions of lncRNAs but neglected the relationship between clinical outcome and lncRNAs themselves. SNPs in the lncRNAs represent the original cause for the changing of the downstream functions. In this study, we explored the impact of potential functional SNPs of the HOTTIP gene on the HSCR risk using a large-scale sample from the Chinese population.
LncRNA HOTTIP gene is located at the homeobox A (HOXA) locus (7p15.2). It was originally identified in anatomically distal human fibroblasts and thus termed as “HOXA transcript at the distal tip”. HOTTIP could directly bind to WDR5/MLL complexes, and then results in the trimethylation of histone H3 lysine 4 and transcription activation 5ʹ HOXA genes.36 The lncRNA HOTTIP was found to be implicated in epigenetic alterations and tumorigenesis. Quagliata et al37 detected a significant up regulation of lncRNA HOTTIP in hepatocellular carcinoma tissues. High expression of HOTTIP is correlated to metastasis formation and serves as a poor prognosis of hepatocellular carcinoma. Xu et al38 determined that HOTTIP may exert a tumor suppressor by directly binding and inhibiting the expression of BRE in glioma cells. Interestingly, Xie et al39 found that both HOXA13 and HOTTIP were down-regulated in HSCR tissue compared to the control tissues, respectively. Further functional experiments indicated that knockdown of HOXA13 and HOTTIP could inhibit the proliferation and migration of HSCR cells. Wang et al40 carried out a case-control study focusing on onco-lncRNAs SNPs and hepatocellular cancer with 521 patients and 817 healthy control in the Chinese population. Among five SNPs in the HOTTIP gene (rs3807598, rs17501292, rs2067087, rs17427960, rs78248039) being investigated, rs17501292, rs2067087, and rs17427960 predisposes to an increased risk of hepatocellular cancer. Hu et al41 revealed that HOTTIP rs1859168 C allele was greatly linked to a decreased risk of pancreatic cancer, in both discovery and validation set. Further evidence indicated that decreased HOTTIP expression caused by C allele be causative of reduced risk of pancreatic cancer. Richards et al42 comprehensively investigated the role of 21 SNPs in HOXA lncRNA genes (HOXA10-AS, HOXA11-AS, and HOTTIP) on the risk of epithelial ovarian cancer in 1201 patients and 2009 controls. However, none of the 21 SNPs were correlated with epithelial ovarian cancer risk at a significance threshold, individually. HOTTIP SNPs exhibited disease-specific impact patterns. Till now, the role of lncRNA HOTTIP SNPs in HSCR risk remains unknown. Considering the vital role of individual HOTTIP gene SNPs in cancers and the important role of HOTTIP in HSCR, it is of great necessity to further explore the role of HOTTIP SNPs in HSCR risk.
Herein, we for the first time explored the correlation of HOTTIP SNPs with the HSCR susceptibility using a large sample size population. Unexpectedly, we failed to find a significant correlation between HOTTIP SNP rs3807598 C>G with the HSCR susceptibility. HSCR is a complex disease characterized by a combination of genetic and environmental factors. The impact of HOTTIP SNP rs3807598 C>G on HSCR risk may be masked by other factors. The joint gene-gene effects may also have a substantial impact on the risk of Hirschsprung disease. An integrative investigation by Gui et al43 found the interaction of variants in RET and NRG1 increases the risk to HSCR development. A single variant may only have a marginal influence on the risk of HSCR. Thus, it is tempting to speculate that this could in part explain such null impact of HOTTIP SNP rs3807598 C>G. Each specific type of HSCR may have a specific genetic background. Therefore, we further perform subgroup detections in HSCR. In all subtype-stratified analyses, no significant correlations were detected between rs3807598 C>G and all specific subtypes. Further study is required to validate the association in an independent cohort.
This study represents the first attempt to determine the relevance of HOTTIP gene SNPs to HSCR risk. The relatively large-scale sample size is the major advantage of our study. The study also has some limitations. Firstly, although the largest study conducted to-date with sample size exceeds 1,000, the statistical power in subgroup analysis was still limited. In addition, the studied population was restricted to the Han Chinese. The findings from the Asian based studies may not be applied to other ethnicities. Besides, we only analyzed one SNP here, more should be included in the future. Last, this study only estimated the significantly associated variants in the HOTTIP gene. Incorporation of environmental factors, gene-environment interactions, and functional evaluations subject to the association of HOTTIP SNPs for HSCR is required.
Conclusion
In summary, we failed to detect a correlation between HSCR risk and SNP rs3807598 C>G in the HOTTIP gene in the Chinese population. Beyond doubt, this first description of HOTTIP gene SNP and HSCR risk sheds light on the etiology of this intestinal disorder and emphasizes the need to expand our knowledge about lncRNAs SNPs in HSCR. Future investigation focus on this point in a larger sample size should be warranted.
Acknowledgments
This study was funded by grants from the Natural Science Foundation of Guangdong Province, China (No: 2019A1515010971), and Guangdong Provincial Key Laboratory of Research in Structural Birth Defect Disease (No: 2019B030301004).
Disclosure
The authors have no conflicts of interest to declare.
References
1. Langer JC. Hirschsprung disease. Curr Opin Pediatr. 2013;25(3):368–374. doi:10.1097/MOP.0b013e328360c2a0
2. Das K, Mohanty S. Hirschsprung disease - current diagnosis and management. Indian J Pediatr. 2017;84(8):618–623. doi:10.1007/s12098-017-2371-8
3. Ambartsumyan L, Smith C, Kapur RP. Diagnosis of Hirschsprung disease. Pediatr Dev Pathol. 2020;23(1):8–22. doi:10.1177/1093526619892351
4. Ruiz Aja E, Vega Hernandez L, Martinez Ezquerra N, De Diego Garcia E, Perez Marrodan A, Alvarez-Buhilla PL. Genetic, population and phenotypic characteristics of patients with Hirschsprung disease. Cir Pediatr. 2012;25(3):135–139.
5. Wang XJ, Camilleri M. Hirschsprung disease: insights on genes, penetrance, and prenatal diagnosis. Neurogastroenterol Motil. 2019;31(11):e13732. doi:10.1111/nmo.13732
6. Butler Tjaden NE, Trainor PA. The developmental etiology and pathogenesis of Hirschsprung disease. Transl Res. 2013;162(1):1–15. doi:10.1016/j.trsl.2013.03.001
7. Borrego S, Wright FA, Fernandez RM, et al. A founding locus within the RET proto-oncogene may account for a large proportion of apparently sporadic Hirschsprung disease and a subset of cases of sporadic medullary thyroid carcinoma. Am J Hum Genet. 2003;72(1):88–100. doi:10.1086/345466
8. Xie X, He Q, Huang L, et al. Associations of SLC6A20 genetic polymorphisms with Hirschsprung’s disease in a Southern Chinese population. Biosci Rep. 2019;39(8). doi:10.1042/BSR20182290
9. Fadista J, Lund M, Skotte L, et al. Genome-wide association study of Hirschsprung disease detects a novel low-frequency variant at the RET locus. Eur J Hum Genet. 2018;26(4):561–569. doi:10.1038/s41431-017-0053-7
10. Heanue TA, Pachnis V. Enteric nervous system development and Hirschsprung’s disease: advances in genetic and stem cell studies. Nat Rev Neurosci. 2007;8(6):466–479. doi:10.1038/nrn2137
11. Wang H, Li Q, Zhang Z, Xiao P, Li L, Jiang Q. Functional studies on novel RET mutations and their implications for genetic counseling for Hirschsprung Disease. Front Genet. 2019;10:924. doi:10.3389/fgene.2019.00924
12. Wang Y, Jiang Q, Cai H, et al. Genetic variants in RET, ARHGEF3 and CTNNAL1, and relevant interaction networks, contribute to the risk of Hirschsprung disease. Aging (Albany NY). 2020;12(5):4379–4393. doi:10.18632/aging.102891
13. Wu W, Lu L, Xu W, et al. Whole exome sequencing identifies a novel pathogenic RET variant in Hirschsprung disease. Front Genet. 2018;9:752. doi:10.3389/fgene.2018.00752
14. Emison ES, McCallion AS, Kashuk CS, et al. A common sex-dependent mutation in a RET enhancer underlies Hirschsprung disease risk. Nature. 2005;434(7035):857–863. doi:10.1038/nature03467
15. Wu TT, Tsai TW, Chu CT, et al. Low RET mutation frequency and polymorphism analysis of the RET and EDNRB genes in patients with Hirschsprung disease in Taiwan. J Hum Genet. 2005;50(4):168–174. doi:10.1007/s10038-005-0236-x
16. Sanchez-Mejias A, Fernandez RM, Lopez-Alonso M, Antinolo G, Borrego S. New roles of EDNRB and EDN3 in the pathogenesis of Hirschsprung disease. Genet Med. 2010;12(1):39–43. doi:10.1097/GIM.0b013e3181c371b0
17. Luzon-Toro B, Torroglosa A, Nunez-Torres R, et al. Comprehensive analysis of NRG1 common and rare variants in Hirschsprung patients. PLoS One. 2012;7(5):e36524. doi:10.1371/journal.pone.0036524
18. Garcia-Barcelo MM, Tang CS, Ngan ES, et al. Genome-wide association study identifies NRG1 as a susceptibility locus for Hirschsprung’s disease. Proc Natl Acad Sci U S A. 2009;106(8):2694–2699. doi:10.1073/pnas.0809630105
19. Trochet D, O’Brien LM, Gozal D, et al. PHOX2B genotype allows for prediction of tumor risk in congenital central hypoventilation syndrome. Am J Hum Genet. 2005;76(3):421–426. doi:10.1086/428366
20. Pingault V, Bondurand N, Kuhlbrodt K, et al. SOX10 mutations in patients with Waardenburg-Hirschsprung disease. Nat Genet. 1998;18(2):171–173. doi:10.1038/ng0298-171
21. Tang CS, Cheng G, So MT, et al. Genome-wide copy number analysis uncovers a new HSCR gene: NRG3. PLoS Genet. 2012;8(5):e1002687. doi:10.1371/journal.pgen.1002687
22. Kim JH, Cheong HS, Sul JH, et al. A genome-wide association study identifies potential susceptibility loci for Hirschsprung disease. PLoS One. 2014;9(10):e110292. doi:10.1371/journal.pone.0110292
23. Mercer TR, Dinger ME, Mattick JS. Long non-coding RNAs: insights into functions. Nat Rev Genet. 2009;10(3):155–159. doi:10.1038/nrg2521
24. Dykes IM, Emanueli C. Transcriptional and post-transcriptional gene regulation by long non-coding RNA. Genom Proteom Bioinf. 2017;15(3):177–186. doi:10.1016/j.gpb.2016.12.005
25. Li Y, Zhou L, Lu C, et al. Long non-coding RNA FAL1 functions as a ceRNA to antagonize the effect of miR-637 on the down-regulation of AKT1 in Hirschsprung’s disease. Cell Prolif. 2018;51(5):e12489. doi:10.1111/cpr.12489
26. Shen Z, Du C, Zang R, et al. Microarray expression profiling of dysregulated long non-coding RNAs in Hirschsprung’s disease reveals their potential role in molecular diagnosis. Neurogastroenterol Motil. 2016;28(2):266–273. doi:10.1111/nmo.12722
27. Su Y, Wen Z, Shen Q, et al. Long non-coding RNA LOC100507600 functions as a competitive endogenous RNA to regulate BMI1 expression by sponging miR128-1-3p in Hirschsprung’s disease. Cell Cycle. 2018;17(4):459–467. doi:10.1080/15384101.2017.1403688
28. Rinn JL, Kertesz M, Wang JK, et al. Functional demarcation of active and silent chromatin domains in human HOX loci by noncoding RNAs. Cell. 2007;129(7):1311–1323. doi:10.1016/j.cell.2007.05.022
29. Kumar V, Westra HJ, Karjalainen J, et al. Human disease-associated genetic variation impacts large intergenic non-coding RNA expression. PLoS Genet. 2013;9(1):e1003201. doi:10.1371/journal.pgen.1003201
30. Zhao J, Zhu Y, Xie X, et al. Pleiotropic effect of common PHOX2B variants in Hirschsprung disease and neuroblastoma. Aging (Albany NY). 2019;11(4):1252–1261. doi:10.18632/aging.101834
31. Zhang Y, Xie X, Zeng J, et al. Association of NRG1 and AUTS2 genetic polymorphisms with Hirschsprung disease in a South Chinese population. J Cell Mol Med. 2018;22(4):2190–2199. doi:10.1111/jcmm.13498
32. He J, Wang F, Zhu J, et al. Association of potentially functional variants in the XPG gene with neuroblastoma risk in a Chinese population. J Cell Mol Med. 2016;20(8):1481–1490. doi:10.1111/jcmm.12836
33. He J, Yang T, Zhang R, et al. Potentially functional polymorphisms in the LIN28B gene contribute to neuroblastoma susceptibility in Chinese children. J Cell Mol Med. 2016;20(8):1534–1541. doi:10.1111/jcmm.12846
34. He J, Zou Y, Liu X, et al. Association of common genetic variants in pre-microRNAs and neuroblastoma susceptibility: a two-center study in Chinese children. Mol Ther Nucleic Acids. 2018;11:1–8. doi:10.1016/j.omtn.2018.01.003
35. He J, Zhang X, Zhang J, et al. LMO1 super-enhancer polymorphism rs2168101 G>T correlates with decreased neuroblastoma risk in Chinese children. J Cancer. 2018;9(9):1592–1597. doi:10.7150/jca.24326
36. Wang KC, Yang YW, Liu B, et al. A long noncoding RNA maintains active chromatin to coordinate homeotic gene expression. Nature. 2011;472(7341):120–124. doi:10.1038/nature09819
37. Quagliata L, Matter MS, Piscuoglio S, et al. Long noncoding RNA HOTTIP/HOXA13 expression is associated with disease progression and predicts outcome in hepatocellular carcinoma patients. Hepatology. 2014;59(3):911–923. doi:10.1002/hep.26740
38. Xu LM, Chen L, Li F, et al. Over-expression of the long non-coding RNA HOTTIP inhibits glioma cell growth by BRE. J Exp Clin Cancer Res. 2016;35(1):162. doi:10.1186/s13046-016-0431-y
39. Xie H, Zhu D, Xu C, et al. Long none coding RNA HOTTIP/HOXA13 act as synergistic role by decreasing cell migration and proliferation in Hirschsprung disease. Biochem Biophys Res Commun. 2015;463(4):569–574. doi:10.1016/j.bbrc.2015.05.096
40. Wang BG, Xu Q, Lv Z, et al. Association of twelve polymorphisms in three onco-lncRNA genes with hepatocellular cancer risk and prognosis: a case-control study. World J Gastroenterol. 2018;24(23):2482–2490. doi:10.3748/wjg.v24.i23.2482
41. Hu P, Qiao O, Wang J, et al. rs1859168 A > C polymorphism regulates HOTTIP expression and reduces risk of pancreatic cancer in a Chinese population. World J Surg Oncol. 2017;15(1):155. doi:10.1186/s12957-017-1218-0
42. Richards EJ, Permuth-Wey J, Li Y, et al. A functional variant in HOXA11-AS, a novel long non-coding RNA, inhibits the oncogenic phenotype of epithelial ovarian cancer. Oncotarget. 2015;6(33):34745–34757. doi:10.18632/oncotarget.5784
43. Gui H, Tang WK, So MT, et al. RET and NRG1 interplay in Hirschsprung disease. Hum Genet. 2013;132(5):591–600. doi:10.1007/s00439-013-1272-9
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.