Back to Journals » International Journal of Nanomedicine » Volume 9 » Issue 1
Nanotechnology versus stem cell engineering: in vitro comparison of neurite inductive potentials
Authors Morano M, Wrobel S, Fregnan F, Ziv-Polat O, Shahar A, Ratzka A, Grothe C, Geuna S, Haastert-Talini K
Received 29 July 2014
Accepted for publication 24 September 2014
Published 14 November 2014 Volume 2014:9(1) Pages 5289—5306
DOI https://doi.org/10.2147/IJN.S71951
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Thomas Webster
Michela Morano,1,* Sandra Wrobel,2,3,* Federica Fregnan,1 Ofra Ziv-Polat,4 Abraham Shahar,4 Andreas Ratzka,2 Claudia Grothe,2,3 Stefano Geuna,1 Kirsten Haastert-Talini2,3
1Department of Clinical and Biological Sciences, Università Degli Studi di Torino, Orbassano, Piemonte, Italy; 2Institute of Neuroanatomy, Hannover Medical School, Hannover, Lower-Saxony, Germany; 3Center for Systems Neuroscience (ZSN), Hannover, Lower-Saxony, Germany; 4NVR Research Ltd, Ness-Ziona, Israel
*These authors contributed equally to this work and share first authorship
Purpose: Innovative nerve conduits for peripheral nerve reconstruction are needed in order to specifically support peripheral nerve regeneration (PNR) whenever nerve autotransplantation is not an option. Specific support of PNR could be achieved by neurotrophic factor delivery within the nerve conduits via nanotechnology or stem cell engineering and transplantation.
Methods: Here, we comparatively investigated the bioactivity of selected neurotrophic factors conjugated to iron oxide nanoparticles (np-NTFs) and of bone marrow-derived stem cells genetically engineered to overexpress those neurotrophic factors (NTF-BMSCs). The neurite outgrowth inductive activity was monitored in culture systems of adult and neonatal rat sensory dorsal root ganglion neurons as well as in the cell line from rat pheochromocytoma (PC-12) cell sympathetic culture model system.
Results: We demonstrate that np-NTFs reliably support numeric neurite outgrowth in all utilized culture models. In some aspects, especially with regard to their long-term bioactivity, np-NTFs are even superior to free NTFs. Engineered NTF-BMSCs proved to be less effective in induction of sensory neurite outgrowth but demonstrated an increased bioactivity in the PC-12 cell culture system. In contrast, primary nontransfected BMSCs were as effective as np-NTFs in sensory neurite induction and demonstrated an impairment of neuronal differentiation in the PC-12 cell system.
Conclusion: Our results evidence that nanotechnology as used in our setup is superior over stem cell engineering when it comes to in vitro models for PNR. Furthermore, np-NTFs can easily be suspended in regenerative hydrogel matrix and could be delivered that way to nerve conduits for future in vivo studies and medical application.
Keywords: iron oxide nanoparticles, conjugated neurotrophic factors, bone marrow-derived mesenchymal stem cells, genetic cell engineering, neurite outgrowth
Introduction
Tissue engineering of peripheral nerves is an active field in research and development. Tissue-engineered nerves could become a valuable alternative to autologous nerve grafts, which are used as the gold standard for peripheral nerve reconstruction surgeries.1 This type of surgery is indicated whenever complete transection injuries of a peripheral nerve cannot be repaired by tension-free end-to-end coaptation of the severed nerve ends.2 Tissue engineering of peripheral nerves in order to bridge nerve defects and provide an optimized regenerative milieu is a complex effort that can only be achieved in a multidisciplinary setting.3 One important task is the delivery of regeneration-promoting molecules such as neurotrophic factors (NTFs) into the nerve defect or, more specifically, their application together with the artificial nerve graft. The main drawback in NTF application is the short half-life time, causing minimal efficacy of single NTF application at the time of reconstructive surgery or systemic application. Different ways to ensure extended availability of added NTFs at the site of nerve reconstruction have been attempted in recent years, including gene therapy via transplanted Schwann cells4 or nanotechnology approaches.5
Ex vivo gene therapy can be used to genetically induce the overexpression of selected NTFs in cells that are later transplanted as part of tissue-engineered nerve grafts.6 The usefulness of this approach has been proven already for transplanted Schwann cells overexpressing different isoforms of fibroblast growth factor-2 (FGF-2) in the rat sciatic nerve model.7,8 Schwann cells are crucially involved in successful peripheral nerve regeneration (PNR), but they are not easy to harvest and propagate for cell transplantation strategies. Therefore, mesenchymal stem cells (MSCs) have been addressed as an easy to access and potentially unlimited cell source for tissue-engineered nerve grafts.9 In this study, we performed stem cell engineering by nonviral genetic modification of bone marrow-derived mesenchymal stem cells (BMSCs), resulting in overexpression of selected NTFs. Another option to ensure availability of NTFs within an artifical nerve graft is the conjugation of the proteins to nanoparticles and their delivery within a hydrogel matrix for axonal regeneration. This strategy was also evaluated in the current work as an alternative strategy to cell-based delivery of NTFs.
Three well-defined NTFs have been analyzed in the presented study. Nerve growth factor (NGF) is known as the neurotrophin with the strongest effect on sensory neurite outgrowth with regard to both axonal elongation and sprouting.10 The second neurotrophin analyzed was glia-derived neurotrophic factor (GDNF), which exerts the most prominent effects on regenerating motor neurons.11 The growth factor FGF-2, in particular its low molecular weight isoform FGF-218kDa, has demonstrated significant support on axonal elongation along with reduced sprouting.12 For the study presented, all three NTFs named were conjugated to iron oxide nanoparticles while overexpression of exclusively FGF-218kDa or GDNF was induced in BMSCs. Sensory neurite outgrowth assays as well as the cell line from rat pheochromocytoma cells (PC-12) neuronal differentiation assay were utilized to quantitatively analyze the regeneration-promoting potential of NTF-overexpressing BMSCs or nanoparticle-conjugated NTFs in comparison with free recombinant NTF proteins.
Materials and methods
Synthesis of np-NTFs
Dextran-coated iron oxide magnetic nanoparticles of, on average, 10 nm dry diameters were prepared as described previously.13 The proteins βNGF, GDNF, and FGF-218kDa (PeproTech Asia, Rehovot, Israel) were covalently conjugated to the dextran coating layer through the divinyl sulfone binding reagent, as described previously.5,14,15 Utilizing the appropriate enzyme-linked immunosorbent assay kits (PeproTech Asia, Rehovot, Israel, and Boster Immunoleader, Pleasanton, CA, USA), the concentration of each protein conjugated to the nanoparticles was determined. Stock aqueous nanoparticle suspensions, with a final concentration of 10 ng/μL of each NTF (np-NTF), were used for the in vitro evaluations.
In vitro techniques
The harvest of material for tissue and cell culture was approved by the local animal protection committees in Torino, Piemonte, Italy, and Hannover, Lower-Saxony, Germany. Figure 1 summarizes details about the different culture systems used in this study.
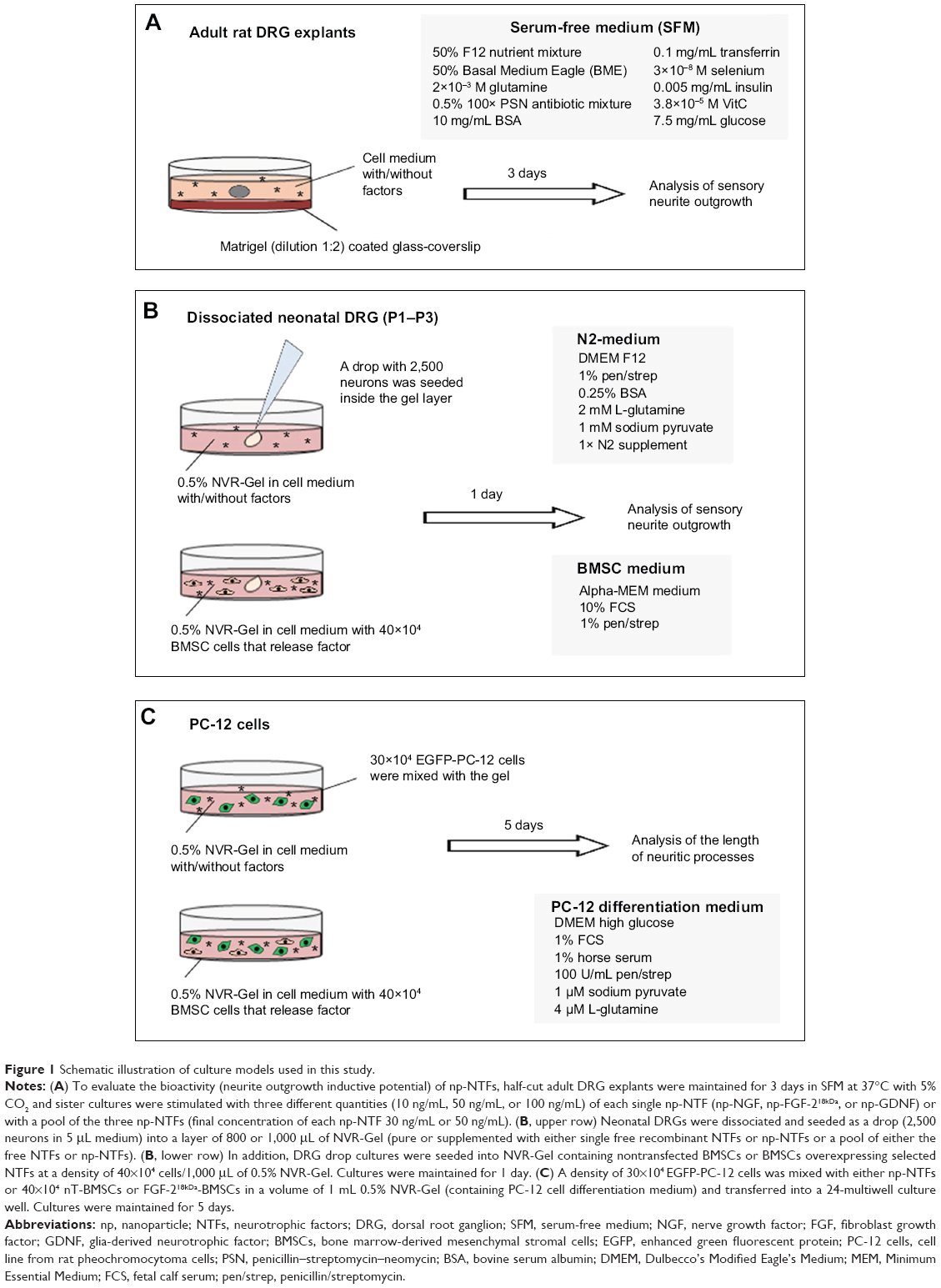 | Figure 1 Schematic illustration of culture models used in this study. |
Adult dorsal root ganglion explant cultures
The spinal column of adult Hannover Wistar rats (approximately 250 g body weight, Harlan, Verona, Italy) was removed after euthanasia. To reach the spinal cord, the vertebral bodies were cut off and removed. Using fine scissors, the vertebral canal was opened through a double cut on both sides of the vertebral bodies. On both sides of the spinal cord, dorsal root ganglia (DRGs) were reached along the posterior roots and extracted from the intervertebral foramina. Harvested DRGs were maintained in Leibovitz’s medium (Invitrogen, Monza, Italy) for 1 hour under sterile conditions. The connective tissue capsules were then mechanically reduced and the explants were half cut. The DRG explants were cultured on matrigel-coated (BD Biosciences, Bedford, MA, USA) glass coverslips in serum-free medium (SFM16): 50% (v/v) F12 nutrient mixture, 50% (v/v) Basal Medium Eagle, 2×10−3 M glutamine, 0.5% (v/v) 100× penicillin–streptomycin–neomycin antibiotic mixture (all Invitrogen), 10 mg/mL bovine albumin serum, 1×10−4 M putrescine, 0.1 mg/mL transferrin, 3×10−8 selenium, 0.005 mg/mL insulin, 3.8×10−5 M vitamin C, 7.5 mg/mL glucose (all Sigma, Milan, Italy) in culture-grade water (Figure 1A).
To evaluate the stability of NTF bioactivity in vitro, free recombinant GDNF (Peprotech, Hamburg, Germany) and np-GDNF were selected, and adult DRG explant cultures were either incubated with fresh or preincubated pure SFM or supplemented with either GDNF type. Preincubated SFM formulations were prepared by incubating the medium for 1 week or 2 weeks at 37°C and 5% CO2. Afterward, this medium was used for DRG explant cultures.
Preparation of NVR-Gel for neurite outgrowth assays
NVR-Gel17 was used as a matrix for seeding drops of dissociated neonatal rat DRG (Figure 1B). NVR-Gel is composed of high molecular weight hyaluronic acid (3×106 Da, BTG Polymers, Kiryat Malachi, Israel) and laminin (Sigma, Rehovot, Israel). For cell cultivation, hyaluronic acid of 1% was further diluted 1:1 either with N2 cell culture medium (see Figure 1B) or PC12 differentation medium (see Figure 1C) to a final concentration of 0.5%. Laminin was then added (10 ng/mL) to complete the NVR-Gel composition. For bioactivity assays, NVR-Gel was either left pure or enriched with 50 ng/mL of single free recombinant NTF (NGF, Invitrogen, Darmstadt, Germany; FGF-2 and GDNF, Peprotech, Hamburg Germany) or single np-NTFs or a pool of either the free NTFs or np-NTFs (50 ng/mL each). In the alternative setting, nontransfected BMSCs or BMSCs overexpressing selected NTFs (see next section) were seeded into the NVR-Gel (Figure 1B).
Harvest and culture of rat BMSCs
Isolation and culturing of rat BMSCs from the femur bone marrow of adult Hannover Wistar rats were performed as described.18 Therefore, BMSCs were cultured in noncoated cell culture flasks in Minimum Essential Medium Alpha (Gibco, Darmstadt, Germany) supplemented with 15% fetal calf serum (FCS) (PAA Laboratories GmbH, Cölbe, Germany) and 1% penicillin/streptomycin (pen/strep, Gibco). Propagated cell cultures were processed to genetic modification or left nontransfected and maintained in Minimum Essential Medium Alpha supplemented with 10% FCS and 1% pen/strep (BMSC culture medium).
The nonviral plasmids for genetic modification of BMSCs were derived from the pCAGGS-empty vector.19 The plasmids encoding for enhanced green fluorescent protein (EGFP, pCAGGS-EGFP-Flag) and FGF-218kDa (pCAGGS-FGF-218kDa-Flag, NCBI GenBank accession NM_019305.2, 533-994 bp) have been previously described.20,21 The pCAGGS-GDNF-Flag plasmid was constructed by polymerase chain reaction-based cloning of the rat GDNF coding sequence (NCBI GenBank accession NM_019139.1, 50-682 bp) using primers introducing MfeI and XbaI cloning sites and removing the stop codon. The in-frame cloned C-terminal 3× Flag tag of the pCAGGS-Flag vector backbone enabled convenient detection of all three proteins (EGFP-Flag, FGF-218kDa-Flag, GDNF-Flag) with the same antibody (anti-Flag M2, F1804 Sigma-Aldrich, Seelze, Germany). Internal reference numbers for the plasmids are given in parentheses for pCAGGS-empty (R399, used for control transfections), pCAGGS-EGFP-FLAG (R412), pCAGGS-FGF-218kDa-FLAG (R417), and pCAGGS-GDNF-FLAG (R415). For the purpose of cell transfection, plasmid deoxyribonucleic acid (DNA) has been purified using the QIAfilter Plasmid Maxi Kit (Qiagen, Hilden, Germany).
For the nonviral transfection, 2.5×106 viable (trypan blue negative) BMSCs were suspended in 90 μL basic transfection solution (Human MSC Kit, LONZA, Cologne, Germany) and mixed with 5 μg of the selected plasmid DNA in AMAXA specific cuvettes using the program A-33 (AMAXA II device, LONZA). The reaction was stopped by adding 900 μL RPMI 1640 medium (Gibco) supplemented with 10% FCS, and the cell suspension was then transferred into new 15 mL falcon tubes. After centrifugation (5 minutes, 235× g), cell pellets were prepared for seeding into 0.5% NVR-Gel (based on either N2 medium or on PC-12 differentiation medium, see Figure 1B and 1C). The transfection efficiency, as exemplarily determined for EGFP-Flag-BMSCs in immunofluorescence microscopy, reached about 60% (Figure S1 in “Supplementary materials”). We assume the same transfection rate also for the other transfections performed.
Drop cultures of dissociated neonatal DRG (sensory neurite outgrowth assay)
Neonatal DRGs were harvested from P1–P3 Hannover Wistar or Sprague Dawley rat pups (breeding pairs obtained from Janvier, Le Gernest-Saint Isle, France). Briefly, after decapitation, the skin on the back was removed and the vertebral canal was longitudinally opened from the neck to the tail. The spinal cord was taken off and the DRGs were collected from intervertebral foramina at both sides of the vertebral canal. DRGs were kept for up to 1 hour in Hank’s balanced salt solution (w/o Mg and Ca, PAA Laboratories GmbH) prior to enzymatic dissociation at 37°C for 20 minutes in Hank’s balanced salt solution added with 0.125% trypsin (Gibco) and 0.05% DNase (Roche, Mannheim, Germany). Then, an additional 0.0075% collagenase type IV (0.1%, PAA Laboratories) was added and the dissociation continued for another 20 minutes. DRGs were then mechanically dissociated using a fire-polished glass pipette before dissociation was stopped by adding 2.5 mL N2 cell culture medium (Dulbecco’s Modified Eagle’s Medium [DMEM] F-12, 1% pen/strep, 0.25% bovine serum albumin, 2 mM L-glutamine, 1 mM sodium pyruvate, 1× N2 supplement; all PAA Laboratories) supplemented with 3% fetal bovine serum. Cells were then counted and placed as drop culture into the middle of 0.5% NVR-Gel (containing N2 medium) (Figure 1B).
Neuronal differentiation assay with PC-12 cells
During propagation of the PC-12 cell line from rat phaeochromocytoma cells of sympathoadrenal origin, a specific proliferation medium was used: DMEM high glucose + 10% horse serum + 5% FCS + 4 μM L-glutamine + 1 μM sodium pyruvate + 100 U/mL pen/strep.22 To analyze the neurite inductive potential of np-NTFs, np-NGF or np-FGF-218kDa was mixed into 0.5% NVR-Gel containing PC-12 cell differentation medium: DMEM high glucose + 1% horse serum + 1% FCS + 4 μM L-glutamine + 1 μM sodium pyruvate + 100 U/mL pen/strep. To analyze the neurite inductive potential of nontransfected (nT-BMSCs) and FGF-218kDa-overexpressing BMSCs (FGF-218kDa-BMSCs), the respective cells were seeded into 0.5% NVR-Gel containing PC-12 cell differentiation medium (Figure 1C). In order to enable specific monitoring of neuronal differentiation of PC-12 cells, the latter were nonvirally transfected to express EGFP. Therefore, pCAGGS-EGFP-FLAG (R412) was introduced to the cells using the AMAXA nucleofection technique similar to the procedures described (Cell Line Nucleofection Kit, program U-29, LONZA). For the neurite outgrowth assay, EGFP-PC-12 cells were mixed with np-NTFs or nT-BMSCs or FGF-218kDa-BMSCs. After 5 days of cultivation, the lengths of neuritic processes of EGFP-PC-12 cells were analyzed with fluorescence microscopy (Olympus IX 70 microscope, Olympus, Hamburg, Germany).
Sodium dodecyl sulfate gel electrophoresis and Western blot analyses
Overexpression of selected NTFs by nonvirally transfected BMSCs was controlled using Western blot analyses of cell lysates. Therefore, nT-BMSCs, FGF-218kDa-BMSCs, and GDNF-BMSCs, as well as empty vector-transfected BMSCs (empty-BMSCs), were cultured with BMSC culture medium as listed in Figure 1C (passage 3, 4, and 5). After 24 hours, proteins were extracted from sister culture cells and samples prepared for Western blotting. Additionally, cell culture supernatants (300 μL) were collected 24 hours, 72 hours, and 120 hours after seeding of nT-BMSCs or after genetic modification (empty-BMSCs, FGF-218kDa-BMSCs, or GDNF-BMSCs) and concentrated using the speedvac system Con 1000 (LTF Labortechnik, Wasserburg, Germany). A calculated protein concentration of 50 μg was prepared in Laemmli buffer following sodium dodecyl sulfate polyacrylamide gel electrophoresis (15% gels). The separated proteins were electrophoretically transferred to a nitrocellulose membrane (Amersham Bioscience, Freiburg, Germany) as described.20 Immunodetection of FGF-218kDa or GDNF was performed in the Enhanced Chemiluminescence Imager System system (ECL, Intas, Science Imaging, Göttingen, Germany) using monoclonal anti-FGF-2 antibody (1:1,000; MerckMillipore, Darmstadt, Germany) or polyclonal anti-GDNF antibody (1:100; Santa Cruz, Heidelberg, Germany) followed by incubation with the respective antimouse immunoglobulin G (IgG) or antirabbit IgG horseradish peroxidase-coupled secondary antibodies (1:4,000; both Amersham Bioscience Europe, Freiburg, Germany). Protein signals were detected in a horseradish peroxidase chemiluminescent reaction (Immobilon, Merck-Millipore, Schwalbach, Germany).
Immunocytochemistry
Transfection efficiency for nonviral genetic modification of BMSCs was determined using EGFP-Flag BMSCs. Cells were fixed for 30 minutes in 4% paraformaldehyde (PFA, Sigma-Aldrich). Prior to immunostaining, fixation was stopped by washing with phosphate-buffered saline.
Adult DRG explant cultures were also fixed for 15 minutes in 4% PFA, while neonatal DRG drop cultures and PC-12 cell cultures in 0.5% NVR-Gel were fixed for 30 minutes in 6% PFA.
Primary antibodies used were anti-βIII-tubulin (monoclonal mouse, 1:400, Sigma-Aldrich), anti-S100 antibody (polyclonal goat, 1:1,000, Dako, Glostrup, Denmark), and anti-Flag antibody (1:1,000, Sigma-Aldrich). Secondary antibodies used were antimouse IgG Alexa-Fluor-488 conjugated (for adult DRG explants and BMSC assays, 1:500–1:2,000, Molecular Probes, Eugene, OR, USA) or antimouse IgG Alexa-Fluor-555 conjugated (for neonatal DRG drop cultures and EGFP-Flag-BMSCs, 1:500, Invitrogen) and rabbit antigoat IgG Alexa-Fluor-488 (1:500; Invitrogen). All primary antibodies were incubated overnight at 4°C and secondary antibodies for 1 hour at room temperature.
In addition, to identify BMSCs, the actin filaments were stained for 30 minutes with phalloidin-tritc fluorescence stain (1:500; Sigma-Aldrich).
All samples were stained with the nuclear marker 4′,6-diamidino-2-phenylindole (1:1,000, 10 minutes, room temperature; Sigma-Aldrich).
Quantification of sensory neurite outgrowth
Adult DRG explants were scanned horizontally and images were merged in order to obtain three-dimensional pictures. The samples were observed with an LSM 510 confocal laser microscopy system (Zeiss, Jena, Thuringia, Germany), which incorporates two lasers (argon and HeNe) and is equipped with an inverted Axiovert 100 M microscope. All images captured were digitally adjusted for optimal resolution.
During photomicrography, entire neonatal DRG drop cultures were taken in six or more overlapping image sections using the 4× objective of an inverted fluorescence microscope (Olympus IX 70, Olympus, Hamburg, Germany). The image sections were then automatically combined using Adobe Photoshop software to obtain a new aligned picture of the entire neuritic network.
Images of both adult DRG explants and neonatal DRG drop cultures were processed using image analysis software (ImageJ, National Institutes of Health, Bethesda, MD, USA) to evaluate the number of extending neurites in certain distances from the center of the cultures. For each sample, the perimeter of the DRG explant or the DRG drop was manually drawn (Polygon selection tool) and automatically enlarged (Edit–Selection–Enlarge tool) to create concentric curves at a fixed distance (50–200 μm for adult DRG and 500–1,100 μm for neonatal DRG) between each other. The number of intersections of neuritic processes with the different curves was then counted and represented as mean values. Additionally, the maximum neurite lengths originating from neonatal DRG drop cultures were analyzed for the ten longest neurites.
Statistical analysis
Three and four biological replicates were performed for experiments with adult DRG explants and neonatal DRG drop cultures, respectively. Each culture condition was analyzed for five adult DRG explants or two to three neonatal DRG drop cultures in each biological replicate. PC-12 cell assays were carried out in three biological replicates with two sister cultures per condition in each replicate. Western blot analyses were performed in three biological replicates. Results are reported as a mean ± standard deviation. Intergroup comparisons were made using one-way or two-ways analysis of variance followed by Bonferroni posttest. A probability value lower than 0.05 was considered as statistically significant.
Results
Np-NTFs demonstrate a prolonged bioactivity in vitro in comparison with free NTFs
To evaluate whether the bioactivity of NTFs is improved by conjugation of the proteins to nanoparticles in comparison with free recombinant proteins, we cultured adult rat DRG explants over 3 days in the presence of fresh and preincubated medium formulations prepared with 50 ng/mL of free NTFs or NTFs conjugated to iron oxide nanoparticles (np-NTFs) and determined the numbers of extending neurites at certain distances from the DRG explant. GDNF was selected as reference NTF because it has demonstrated high bioactivity on adult DRG explant cultures, although with levels below NGF treatment (see Figure S2A). This made GDNF a candidate NTF to also detect minimal changes in bioactivity over time. As is depicted in Figure 2, incubation of adult DRG explant cultures for 3 days with fresh free GDNF or np-GDNF induced significantly increased numeric neurite outgrowth (Figure 2A) and extension of neuritic processes (Figure 2B) in comparison with control conditions with fresh medium without GDNF supplementation. Representative photomicrographs are given in Figure 2C. Fresh free GDNF additionally demonstrated an increased bioactivity over that of fresh np-GDNF. While the neurite outgrowth was not significantly affected after 1-week preincubation of medium formulations (data not shown), prolonged bioactivity of np-GDNF compared with free GDNF was clearly detectable after a 2-week preincubation period. After this period, free GDNF no longer increased numeric neurite outgrowth when compared with control conditions, and we detected a significant reduction of bioactivity by 65% of the bioactivity of fresh free GDNF. In contrast, np-GDNF revealed its bioactivity also after a 2-week preincubation period, as demonstrated by the maintained neurite outgrowth inductive activity, which was then still significantly increased over control conditions (Figure 2A and B).
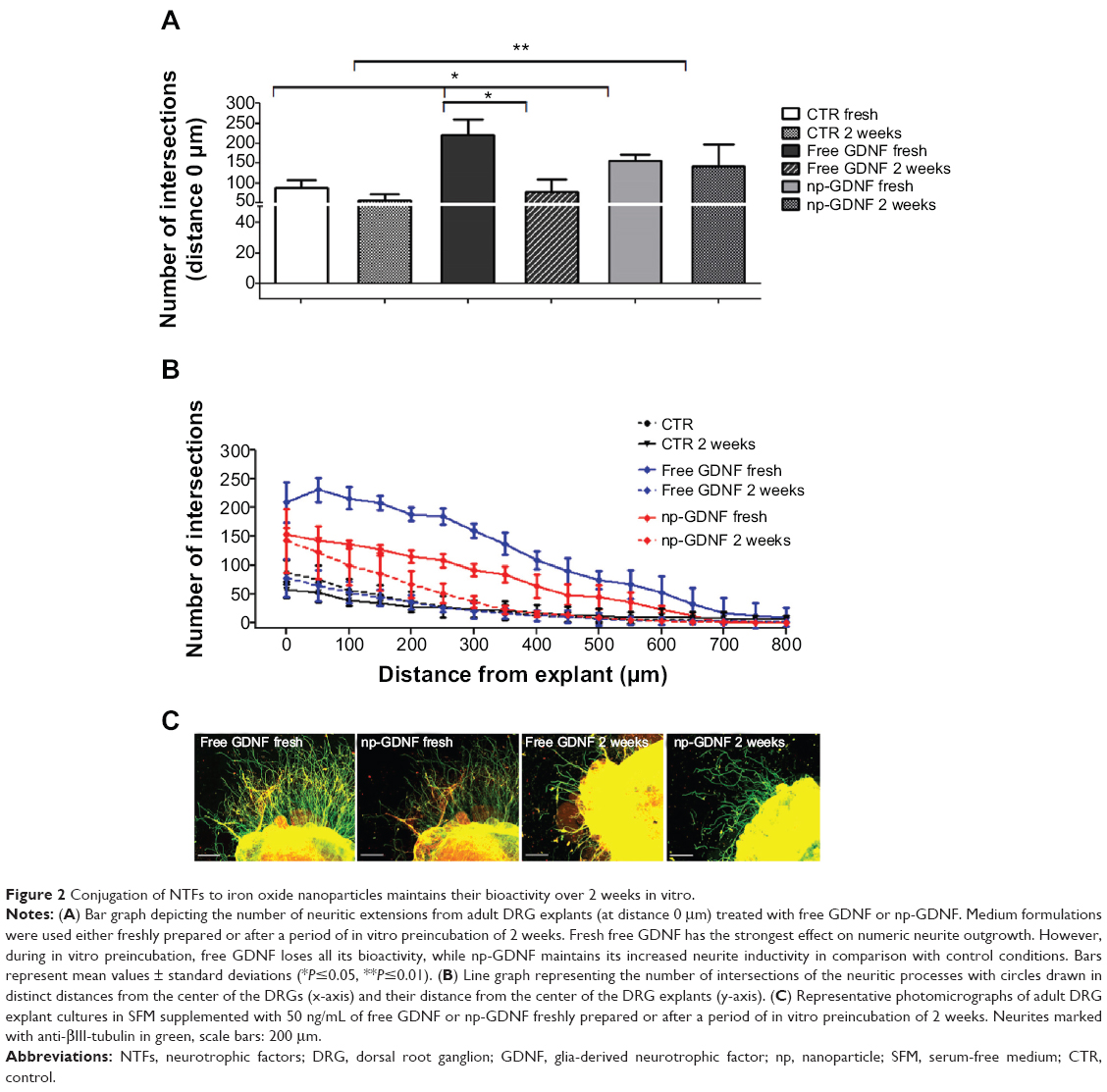 | Figure 2 Conjugation of NTFs to iron oxide nanoparticles maintains their bioactivity over 2 weeks in vitro. |
Additionally, we demonstrated that at a certain dosage, all np-NTFs tested (np-NGF, np-FGF-218kDa, and np-GDNF) significantly increased adult sensory neurite outgrowth over the values seen in control cultures (cultured in SFM medium alone). All np-NTFs further demonstrated a dose dependency of their neurite inductive effects. The respective data are presented as supplementary material, including Figure S2.
np-NTFs exert the same bioactivity as free recombinant proteins when incorporated into a regenerative hydrogel matrix
After we could demonstrate that np-NTFs exert a neurite inductive effect on adult DRG explants, we further compared effects of free NTFs and np-NTFs on neurite outgrowth from neonatal DRG preparations placed into the nerve regenerative matrix NVR-Gel. NVR-Gel is composed of high molecular weight hyaluronic acid and laminin and has been described as regenerative matrix for nervous system repair.17,23 The next experiment was performed in order to determine whether the suspension of np-NTFs in the regenerative NVR-Gel matrix still allows them to exert inductive effects on neurite outgrowth, as seen for their suspension in medium. Therefore, the in vitro neurite outgrowth assay was analyzed after placing dissociated neonatal DRG drop cultures into 0.5% NVR-Gel supplemented with np-NTFs. To further elucidate whether conjugation of NTFs to iron oxide nanoparticles has an impact on the bioactivity of the NTF proteins, we compared in this experiment the effects of np-NTFs with those of free recombinant NTF proteins. As is depicted in Figure 3, the supplementation of NVR-Gel with either free NTF or np-NTF resulted in significant effects on neurite outgrowth induction from neonatal DRG drop cultures (Figure 3A, C, and E). Both free NGF and np-NGF induced significantly higher numeric neurite outgrowth in distances up 700 μm from the center of the DRG drop culture than the other NTFs (Figure 3A and B). However, analysis of the ten longest neurites extending from each DRG drop culture did not reveal any significant difference among all treatments (Figure 3G). No statistically significant difference was detectable between the neurite outgrowth inductive effects of np-NTFs compared with free NTFs, suggesting that when incorporated into a regenerative hydrogel matrix, np-NTFs are able to exert the same bioactivity as free NTFs. Interestingly, for FGF-218kDa, an even slightly increased neurite inductive potential over short distances was visible for np-FGF-218kDa in comparison with free FGF-218kDa (Figure 3C and D). The opposite was seen when comparing np-GDNF with free GDNF (Figure 3E and F).
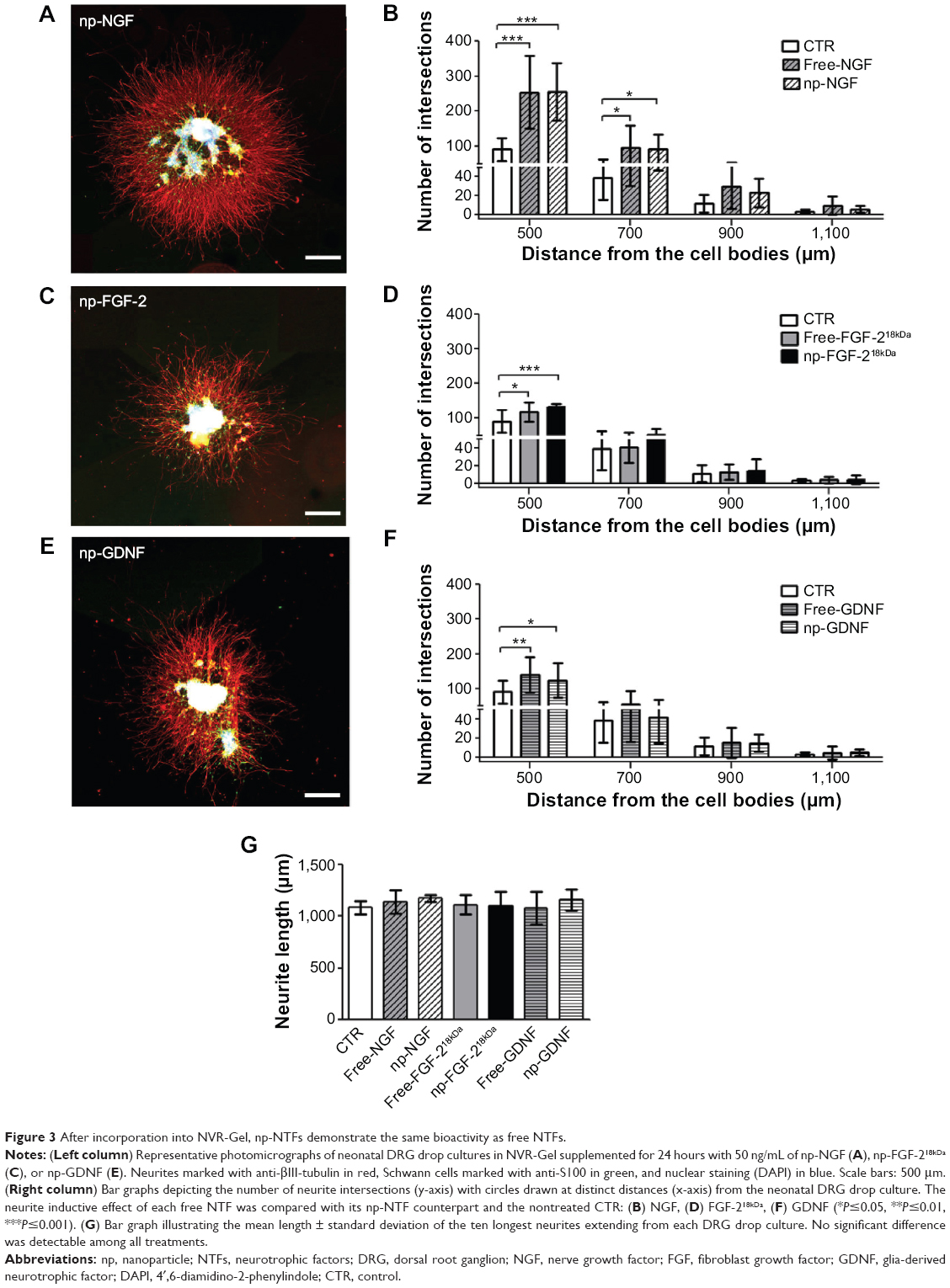 | Figure 3 After incorporation into NVR-Gel, np-NTFs demonstrate the same bioactivity as free NTFs. |
Because we could demonstrate that np-NGF exerts the most prominent neurite inductive effect in both adult DRG explant cultures and neonatal DRG drop cultures, we were interested in whether this effect could be further increased by a possible synergistic effect of the other np-NTFs. Therefore, all three np-NTFs (np-NGF + np-FGF-218kDa + np-GDNF) were mixed to an np-NTF pool. As is shown in the supplementary material, including Figures S3 and S4, we observed that neither np-NTF pool concentration was able to significantly induce neurite outgrowth over the value achieved with the treatment with np-NGF alone.
BMSCs overexpressing NTFs induce less sensory neurite outgrowth than np-NTFs
BMSCs were nonvirally transfected to induce overexpression of NTFs. Successful genetic modification was monitored in Western blot analysis from cell lysates and cell culture supernatants (Figure 4). Cell lysates of engineered BMSCs reveal strong expression of the respective FGF-218kDa-Flag or pro-GDNF-Flag (Figure 4A). Cell culture supernatants collected from cultures of GDNF-BMSCs contain increased levels of pro-GDNF with increasing culture time as well as mature GDNF protein after 72–120 hours of culture (Figure 4B). On the contrary, nT-BMSCs culture supernatants contain low levels of endogenous pro-GDNF, while culture supernatants of the empty-BMSCs demonstrate a decreased expression of pro-GDNF (Figure 4B). With regard to FGF-2, neither endogenous FGF-218kDa nor FGF-218kDa-Flag was detectable in cell culture supernatants of nT-BMSCs or genetically engineered BMSCs (data not shown).
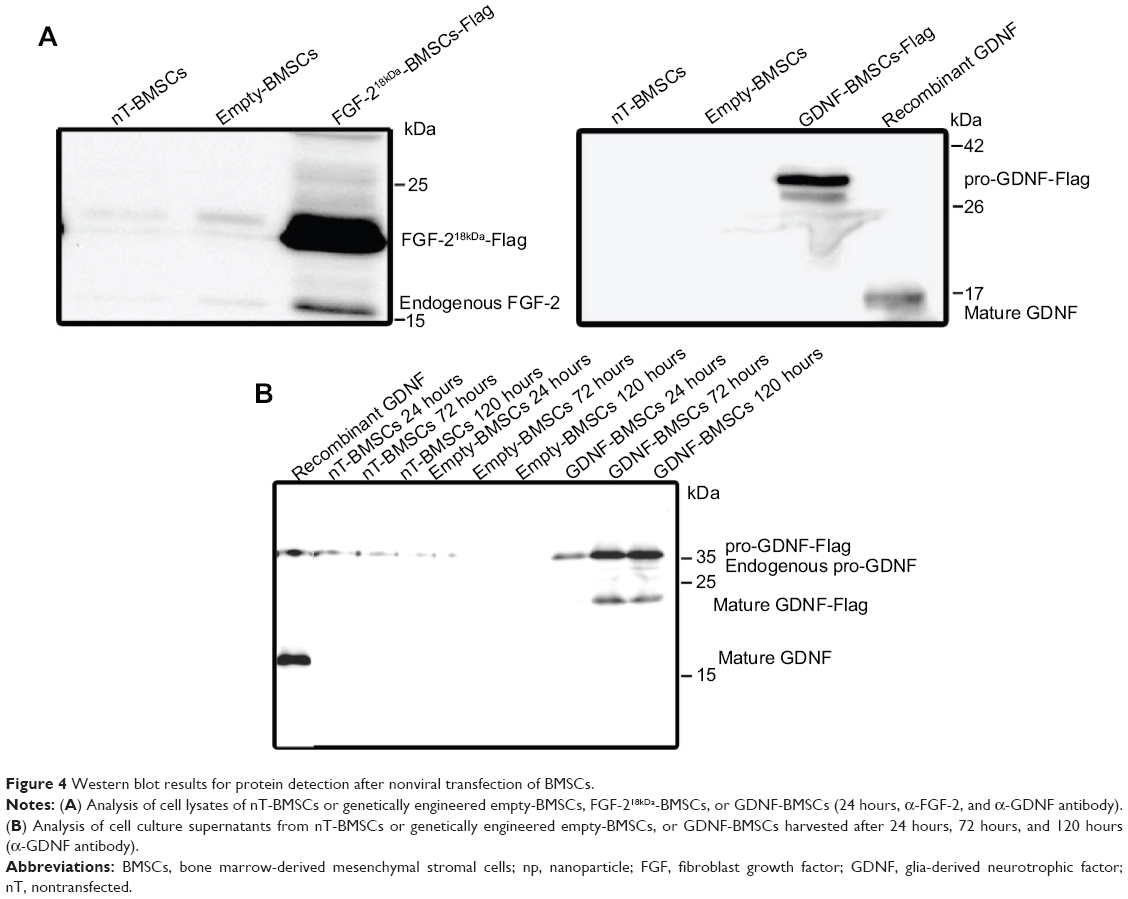 | Figure 4 Western blot results for protein detection after nonviral transfection of BMSCs. |
Genetically engineered BMSCs were suspended in NVR-Gel and analyzed with respect to their neurite inductive effect on neonatal DRG drop cultures. As is depicted in Figure 5, all types of genetically engineered BMSCs, but especially nT-BMSCs, demonstrated nonsignificant increase in numeric neurite outgrowth from DRG drop cultures over the value achieved in pure NVR-Gel (control [CTR]), and no significant difference was detected between the different cell types analyzed (Figure 5A). The maximum neurite lengths were almost not affected by the presence of nT-BMSCs (Figure 5C) in comparison with control levels (CTR), while engineered BMSCs (FGF-218kDa-BMSCs and GDNF-BMSCs) demonstrated nonsignificant impairment of distance outgrowth, which was most prominent in the presence of GDNF-BMSCs (Figure 5B). When comparing numeric neurite outgrowth from DRG drop cultures in the presence of engineered BMSCs with the incubation with np-FGF-218kDa or np-GDNF, respectively, no significant differences were detectable (Figure 6A). However, it is noteworthy that while nT-BMSCs had a very similar effect on maximum neurite lengths as np-NTFs, co-culture with engineered BMSCs, especially GDNF-BMSCs, significantly impaired distance outgrowth of neurites (Figure 6B).
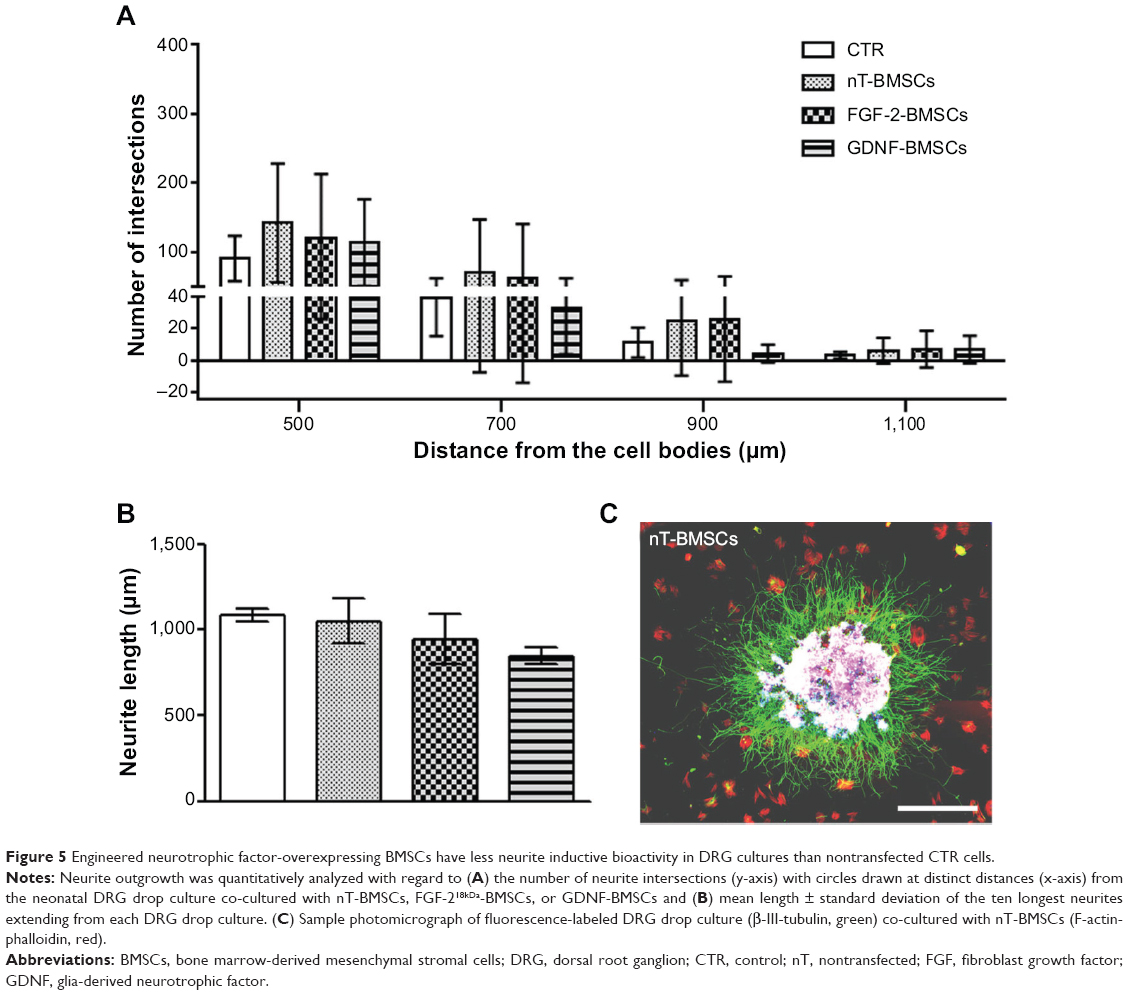 | Figure 5 Engineered neurotrophic factor-overexpressing BMSCs have less neurite inductive bioactivity in DRG cultures than nontransfected CTR cells. |
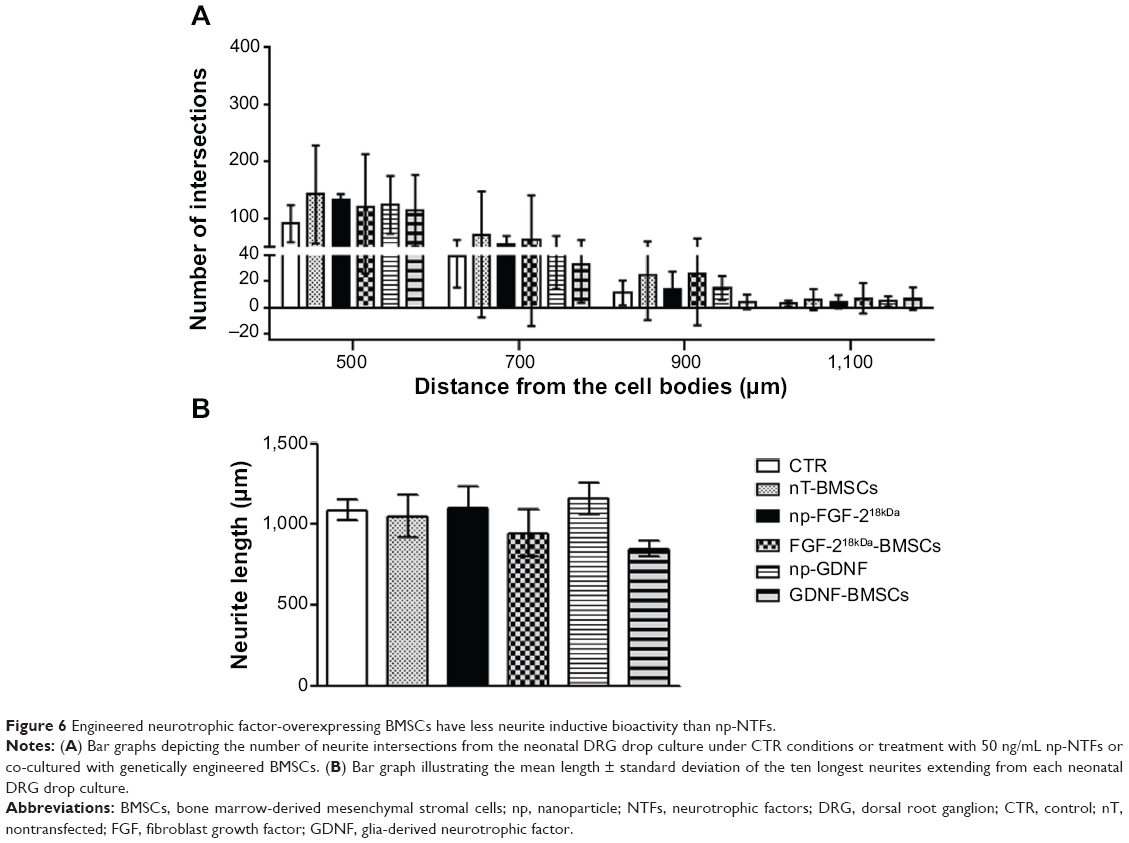 | Figure 6 Engineered neurotrophic factor-overexpressing BMSCs have less neurite inductive bioactivity than np-NTFs. |
Neurite formation by PC-12 cells is supported to a higher extent in the presence of FGF-218kDa-BMSCs than in the presence of np-FGF-218kDa
In order to directly compare the neurite inductive potential of np-NTFs and engineered NTF-BMSCs again in another cell system, we utilized the PC-12 cell neuronal differentiation assay as a third in vitro model. PC-12 cells are highly sensitive for treatment with NGF or FGF-218kDa. Therefore, free NGF, np-NGF, free FGF-218kDa, np-FGF-218kDa, or FGF-218kDa-BMSCs were incorporated in NVR-Gel together with PC-12 cells. As is shown in Figure 7, 50 ng/mL of np-NGF demonstrated a slightly stronger neurite outgrowth inductive potential than 50 ng/mL free NGF (positive control condition) or free FGF-218kDa and np-FGF-218kDa (Figure 7A). Interestingly, the presence of nT-BMSCs significantly impaired neurite outgrowth from PC-12 cells when compared with all other conditions, while FGF-218kDa-BMSCs (Figure 7B) demonstrated a significantly increased effect on neuronal differentiation of PC-12 cells in comparison with free FGF-218kDa or np-FGF-218kDa.
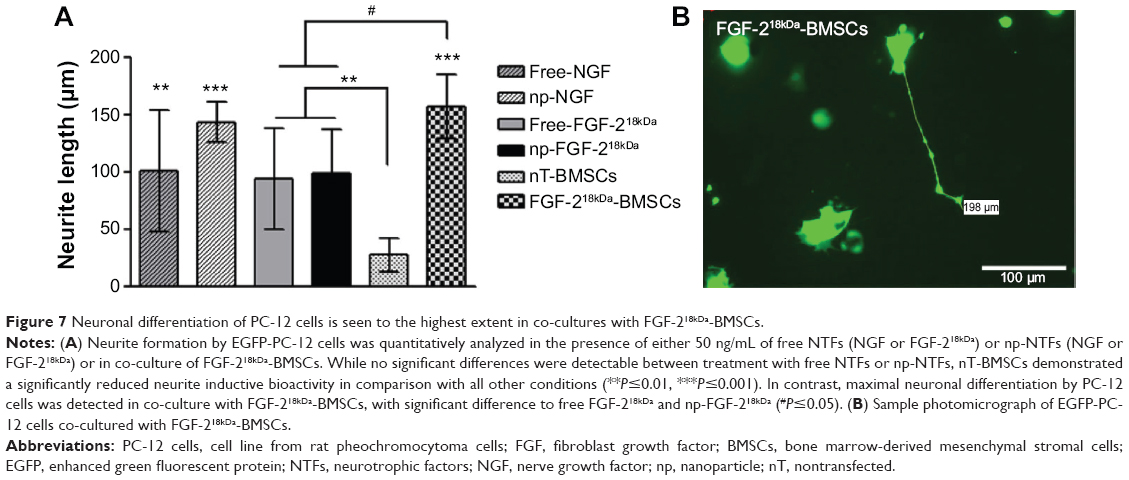 | Figure 7 Neuronal differentiation of PC-12 cells is seen to the highest extent in co-cultures with FGF-218kDa-BMSCs. |
Discussion
In this study, we compared in vitro two different possible approaches to deliver NTFs within artificial grafts for peripheral nerve reconstruction. The first was the nanotechnology approach of conjugation of NTFs with iron oxide nanoparticles, which was supposed to increase the stability of the conjugated NTFs, but also to ensure local and slow release of NTFs in the lumen of nerve conduits.5 The second approach we studied was stem cell engineering. Here, we induced NTF overexpression in BMSCs with nonviral genetic modification. The genetically engineered BMSCs could potentially be transplanted within tissue-engineered nerve conduits, as has been successfully done before with genetically engineered Schwann cells.4
In order to compare the feasibility of optimized NTF delivery increasing the neurite outgrowth in vitro, we utilized adult DRG explant organotypic cultures as well as neonatal DRG drop cultures. The DRG neurite outgrowth model is widely used to assess the neurite outgrowth in a specific environment.24,25
Using adult DRG explant cultures, we demonstrate that the bioactivity of np-NTFs is comparable with that of free NTFs when incorporated into a regenerative hydrogel matrix. The latter is a good prerequisite for future in vivo application. From previous studies, we already have proof that NTFs conjugated to iron oxide nanoparticles demonstrate prolonged stability over free NTFs.14 Here, we clearly demonstrate that not only the stability of the proteins but also, most importantly, their bioactivity is stabilized by conjugation of NTFs to nanoparticles. Our study comparing the neurite inductive potential of fresh with preincubated medium formulations evidenced that only minimal reduction of bioactivity occurs for np-GDNF during a 2-week preincubation period, while free GDNF lost its neurite inductive activity in the same time span.
We could further demonstrate that np-NTFs are able to stimulate neuronal outgrowth in a dose-dependent manner and that np-NTFs have similar bioactivity as free NTFs when suspended in a regenerative hydrogel matrix. The latter system mimics a way to apply np-NTFs with nerve conduits for peripheral nerve reconstruction surgery. As hydrogel matrix, we used NVR-Gel, which has been described before as a suitable matrix for neuronal cultures17 and which seemed to be appropriate as luminal filler for artificial nerve conduits. Using neonatal DRG drop cultures we demonstrate that np-NTFs are effective in inducing the numeric neurite outgrowth activity also when suspended in NVR-Gel. In NVR-Gel enriched with np-NTFs, the elongation of neurites, however, seems not to be stimulated. We suppose that the presence of the gel slowed down neurite elongation, but further analysis using long-term cultures is necessary to elucidate how neurite distance outgrowth is influenced by NVR-Gel.
With regard to potential in vivo application of np-NTFs, another finding from our study has to be considered. The dose dependency of np-NTF-induced sensory neurite outgrowth varied among the NTFs investigated. While 50 ng/mL of np-NGF or np-GDNF induced maximal neurite outgrowth among the concentrations chosen, the lower concentration of only 10 ng/mL np-FGF-218kDa showed a trend to be more efficient than higher concentrations (data presented as supplementary material). All np-NTFs investigated, however, demonstrated an induction of neurite outgrowth activity at the concentration of 50 ng/mL, which can be concluded as a useful concentration for future in vivo application. From the acquired in vitro data, it cannot be finally decided which np-NTFs would give maximum support of axonal regeneration in vivo. DRG neuron populations are heterogeneous with regard to the quality of sensory transmission, and different growth factors act selectively on different subpopulations of DRG neurons.26–29 Therefore, an emerging concept in the field of artificial nerve graft development is to use a combination of NTFs with the aim to obtain a widespread effect on all sensory, motor, and sympathetic neuronal subpopulations.30–32 It is, however, still under debate which NTF combination will result in the most successful support of PNR while simultaneously avoiding negative influences in the process.33 Therefore, we also investigated the effect of an np-NTF pool on the neurite outgrowth activity in adult DRG explant and neonatal DRG drop cultures. In our hands, an np-NTF pool of np-NGF, np-FGF-218kDa, and np-GDNF did not demonstrate higher efficiency in promoting neurite outgrowth than 50 ng/mL np-NGF alone. This indicates that there is no synergistic effect among these three selected NTFs. However, considering the limits of an in vitro approach, it is possible that in our culture systems, 50 ng/mL np-NGF treatment already induced the maximum neurite outgrowth with no possibility to demonstrate a synergy for the additionally added np-FGF-218kDa or np-GDNF.
Evaluation of the second innovative approach of NTF delivery to nerve conduits and stem cell engineering showed that primary nT-BMSCs have a higher potency to induce sensory neurite outgrowth than engineered NTF-BMSCs. The basis for this may be the secretome of BMSCs that provides a variety of growth factors to the co-cultured neurons.34 We assume that the secretome has, to some extent, been negatively influenced by the genetic modification during BMSC engineering. Further analysis is needed to elucidate this in more detail prior to potential in vivo application, but, although only seen in one out of three independent analyses, the Western blot analysis shown in Figure 4B already indicates that the GDNF expression might be altered between nT-BMSCs and genetically engineered empty-BMSCs.
When comparing the two innovative approaches of nanotechnology and stem cell engineering with regard to sensory neurite outgrowth, however, np-NTFs have demonstrated a slightly increased bioactivity over NTF-BMSCs. Additionally, using a different neuronal differentiation model,22,35 the PC-12 cell neurite outgrowth induction assay, NTF-BMSCs demonstrated an increased bioactivity over np-NTFs. We therefore assume that the secretome together with the overexpressed FGF-218kDa was, in this setting, specifically supportive for PC-12 cell sympathetic neurite outgrowth.
One last point has to be considered when thinking of future in vivo application of nanotechnological NTF delivery to peripheral nerve guidance conduits. The conduits will be sutured between the transected nerve ends and, especially the distal nerve end, will undergo Wallerian degeneration. As this process will provide Schwann cell ingrowth into the nerve conduits, we were also interested in whether Schwann cell migration is modified in our in vitro models. The data are not shown because we have no hints that Schwann cell migration is modified by adding np-NTFs or BMSCs to the system. Whether synergistic effects would develop for np-NTFs delivered within the conduits and those additionally liberated during Wallerian degeneration of the distal nerve end, is only accessible in vivo and must be the subject of future studies.
Conclusion
Both approaches demonstrated their potential to induce neurite outgrowth in different neuronal models. They may be chosen specifically in the context of peripheral nerve tissue engineering. However, the variability of the BMSC secretome upon genetic engineering of the cells needs to be studied in more detail prior to their in vivo application. Our comparative in vitro study provides evidence that np-NTFs have long-term bioactivity and reliably support neurite outgrowth in vitro also after periods when free NTFs have already lost their bioactivity. With regard to their therapeutical use, iron oxide nanoparticles are considered to be nontoxic and are already in use for various biomedical applications, such as diagnostics, cell labeling, magnetic resonance imaging, and X-ray contrast agents, and for hyperthermia.5 Therefore, np-NTFs are good candidates for the in vivo evaluation of their PNR-promoting potential in future studies.
Acknowledgments
This project has received funding from the European Union’s Seventh Programme for Research, Technological Development and Demonstration under Grant Agreement No 278612. For excellent technical assistance, we thank Maike Wesemann from the Institute of Neuroanatomy at Hannover Medical School. For support in bone marrow-derived mesenchymal stem cell cultures, we thank Antonio J Salgado from the University of Minho, Braga, Portugal.
Disclosure
The authors report no conflicts of interest in this work.
References
Tos P, Ronchi G, Geuna S, Battiston B. Future perspectives in nerve repair and regeneration. Int Rev Neurobiol. 2013;109:165–192. | ||
Deumens R, Bozkurt A, Meek MF, et al. Repairing injured peripheral nerves: bridging the gap. Prog Neurobiol. 2010;92(3):245–276. | ||
Geuna S, Gnavi S, Perroteau I, Tos P, Battiston B. Tissue engineering and peripheral nerve reconstruction: an overview. Int Rev Neurobiol. 2013;108:35–57. | ||
Haastert K, Grothe C. Gene therapy in peripheral nerve reconstruction approaches. Curr Gene Ther. 2007;7(3):221–228. | ||
Ziv-Polat O, Skaat H, Shahar A, Margel S. Novel magnetic fibrin hydrogel scaffolds containing thrombin and growth factors conjugated iron oxide nanoparticles for tissue engineering. Int J Nanomedicine. 2012;7:1259–1274. | ||
Madduri S, Gander B. Schwann cell delivery of neurotrophic factors for peripheral nerve regeneration. J Peripher Nerv Syst. 2010;15(2):93–103. | ||
Haastert K, Lipokatic E, Fischer M, Timmer M, Grothe C. Differentially promoted peripheral nerve regeneration by grafted Schwann cells over-expressing different FGF-2 isoforms. Neurobiol Dis. 2006;21(1):138–153. | ||
Haastert K, Ying Z, Grothe C, Gomez-Pinilla F. The effects of FGF-2 gene therapy combined with voluntary exercise on axonal regeneration across peripheral nerve gaps. Neurosci Lett. 2008;443(3):179–183. | ||
Oliveira JT, Mostacada K, de Lima S, Martinez AM. Bone marrow mesenchymal stem cell transplantation for improving nerve regeneration. Int Rev Neurobiol. 2013;108:59–77. | ||
Johnson EO, Charchanti A, Soucacos PN. Nerve repair: experimental and clinical evaluation of neurotrophic factors in peripheral nerve regeneration. Injury. 2008;39 Suppl 3:S37–S42. | ||
Catrina S, Gander B, Madduri S. Nerve conduit scaffolds for discrete delivery of two neurotrophic factors. Eur J Pharm Biopharm. 2013;85(1):139–142. | ||
Klimaschewski L, Hausott B, Angelov DN. The pros and cons of growth factors and cytokines in peripheral axon regeneration. Int Rev Neurobiol. 2013;108:137–171. | ||
Molday RS, MacKenzie D. Immunospecific ferromagnetic iron-dextran reagents for the labeling and magnetic separation of cells. J Immunol Methods. 1982;52(3):353–367. | ||
Ziv-Polat O, Shahar A, Levy I, et al. The role of neurotrophic factors conjugated to iron oxide nanoparticles in peripheral nerve regeneration: in vitro studies. Biomed Res Int. 2014;2014:1–10. | ||
Shahar A, Ziv-Polat O, Margel S, Freier T. Developing a customized tissue-engineered implant for peripheral nerve reconstruction: role of nanoparticles. In: Bhoop BS, ed. Medical Nanobiotechnology. Vol 5. Houston, TX: Studium Press LLC; 2014. | ||
Fueshko S, Wray S. LHRH cells migrate on peripherin fibers in embryonic olfactory explant cultures: an in vitro model for neurophilic neuronal migration. Dev Biol. 1994;166(1):331–348. | ||
Shahar A, Nevo Z, Rochkind S, inventors. Cross-linked hyaluronic acid-laminin gels and use thereof in cell culture and medical implants. United States patent US 00243732001. November 13, 2001. | ||
Wrobel S, Serra SC, Ribeiro-Samy S, et al. In vitro evaluation of cell-seeded chitosan films for peripheral nerve tissue engineering. Tissue Eng Part A. 2014;20(17–18):2339–2349. | ||
Niwa H, Yamamura K, Miyazaki J. Efficient selection for high-expression transfectants with a novel eukaryotic vector. Gene. 1991;108(2):193–199. | ||
Ratzka A, Kalve I, Ozer M, et al. The colayer method as an efficient way to genetically modify mesencephalic progenitor cells transplanted into 6-OHDA rat model of Parkinson’s disease. Cell Transplant. 2012;21(4):749–762. | ||
Ratzka A, Baron O, Grothe C. FGF-2 deficiency does not influence FGF ligand and receptor expression during development of the nigrostriatal system. PLoS One. 2011;6(8):e23564. | ||
van Bergeijk J, Haastert K, Grothe C, Claus P. Valproic acid promotes neurite outgrowth in PC12 cells independent from regulation of the survival of motoneuron protein. Chem Biol Drug Des. 2006;67(3):244–247. | ||
Rochkind S, Shahar A, Fliss D, et al. Development of a tissue-engineered composite implant for treating traumatic paraplegia in rats. Eur Spine J. 2006;15(2):234–245. | ||
Melli G, Hoke A. Dorsal root ganglia sensory neuronal cultures: a tool for drug discovery for peripheral neuropathies. Expert Opin Drug Discov. 2009;4(10):1035–1045. | ||
Bilsland J, Rigby M, Young L, Harper S. A rapid method for semi-quantitative analysis of neurite outgrowth from chick DRG explants using image analysis. J Neurosci Methods. 1999;92(1–2):75–85. | ||
Benedetti M, Levi A, Chao MV. Differential expression of nerve growth factor receptors leads to altered binding affinity and neurotrophin responsiveness. Proc Natl Acad Sci U S A. 1993;90(16):7859–7863. | ||
Bennett DL, Michael GJ, Ramachandran N, et al. A distinct subgroup of small DRG cells express GDNF receptor components and GDNF is protective for these neurons after nerve injury. J Neurosci. 1998;18(8):3059–3072. | ||
Wright DE, Snider WD. Neurotrophin receptor mRNA expression defines distinct populations of neurons in rat dorsal root ganglia. J Comp Neurol. 1995;351(3):329–338. | ||
Xiao J, Wong AW, Willingham MM, et al. BDNF exerts contrasting effects on peripheral myelination of NGF-dependent and BDNF-dependent DRG neurons. J Neurosci. 2009;29(13):4016–4022. | ||
Sharma HS. Neurotrophic factors in combination: a possible new therapeutic strategy to influence pathophysiology of spinal cord injury and repair mechanisms. Curr Pharm Des. 2007;13(18):1841–1874. | ||
Madduri S, di Summa P, Papaloizos M, Kalbermatten D, Gander B. Effect of controlled co-delivery of synergistic neurotrophic factors on early nerve regeneration in rats. Biomaterials. 2010;31(32):8402–8409. | ||
Blesch A, Fischer I, Tuszynski MH. Gene therapy, neurotrophic factors and spinal cord regeneration. Handb Clin Neurol. 2012;109:563–574. | ||
Madduri S, Gander B. Growth factor delivery systems and repair strategies for damaged peripheral nerves. J Control Release. 2012;161(2):274–282. | ||
Ribeiro CA, Salgado AJ, Fraga JS, Silva NA, Reis RL, Sousa N. The secretome of bone marrow mesenchymal stem cells-conditioned media varies with time and drives a distinct effect on mature neurons and glial cells (primary cultures). J Tissue Eng Regen Med. 2011;5(8):668–672. | ||
van Bergeijk J, Rydel-Konecke K, Grothe C, Claus P. The spinal muscular atrophy gene product regulates neurite outgrowth: importance of the C terminus. Faseb J. 2007;21(7):1492–1502. |
Supplementary materials
Neurotrophic factors conjugated to iron oxide nanoparticles induce a dose-dependent neurite outgrowth from adult dorsal root ganglion explants
Adult rat dorsal root ganglion (DRG) explants were cultured over 3 days in the presence of increasing amounts of neurotrophic factors conjugated to iron oxide nanoparticles (np-NTFs), and the numbers of extending neurites measured at certain distances from the DRG explant. As is depicted in Figure S2, at a certain dosage, all np-NTFs tested significantly increased adult sensory neurite outgrowth over the values seen in control cultures (cultured in serum-free medium alone). All np-NTFs demonstrated a dose dependency of their neurite inductive effects. The strongest effect on numeric neurite outgrowth (Figure S2A) was seen for the treatment with 50 ng/mL of either np-NGF or np-glia-derived neurotrophic factor (GDNF). Lower or higher medium concentrations of both np-NGF and np-GDNF showed reduced effects with significantly lower neurite numbers for the reduced concentration of 10 ng/mL of np-NGF and the elevated concentration of 100 ng/mL of np-GDNF, respectively. Only minimal differences in the numeric neurite outgrowth were detectable for the treatment with different concentrations of np-fibroblast growth factor (FGF)-218kDa. While a significant increase of neurite numbers over control levels was already detectable for the lowest concentration of 10 ng/mL np-FGF-218kDa, the elevated concentrations no longer induced significant effects on neurite outgrowth. As is depicted in Figure S2B, the strongest effect on neurite elongation was detected under treatment with np-NGF by which, again, 50 ng/mL induced the maximum neurite lengths in the highest number of neurites. But treatment with 50 ng/mL np-FGF-218kDa also induced outgrowth of longer neurites, up to a distance of 450 μm from the center of the DRG, than all other treatments except the one with np-NGF. Under control conditions, outgrowing neurites reached a distance of only 300 μm from the DRG center.
Combination of three different neurotrophic factors conjugated to np-NTFs fails to further increase the neurite inductive effect of np-NGF alone
Because a concentration of 50 ng/mL np-NGF had demonstrated the most prominent neurite inductive effect, we were interested in whether this effect could be further increased by a possible synergistic effect of the other np-NTFs. Therefore, all three np-NTFs (np-NGF + np-FGF-218kDa + np-GDNF) were mixed to an np-NTF pool of two different concentrations: 30 ng/mL (each np-NTF) and 50 ng/mL (each np-NTF). As is demonstrated in Figure S3, we observed that neither np-NTF pool concentration was able to significantly induce neurite outgrowth over the value achieved with the treatment with 50 ng/mL np-NGF alone. The numeric neurite outgrowth from adult DRG explants was significantly increased over control levels in both np-NTF pool treatments, which were not significantly different from each other (Figure S3A). Significantly longer neurites than under control conditions were induced by both np-NTF pool treatments as well (Figure S3B). However, treatment with 50 ng/mL np-NGF alone demonstrated a tendency to induce an even more increased number of long neurites (up to >500 μm from the center of the DRGs).
Additionally to testing the neurite inductive effects of np-NTFs, we again analyzed the bioactivity of an np-NTF pool (50 ng/mL of each np-NGF, np-FGF 218kDa, and np-GDNF) suspended in NVR-Gel. Similar to the results after incubation of adult DRG explant cultures with an np-NTF pool, we observed no synergistic effect. As is shown in Figure S4, the neurite inductive potential of the np-NTFs pool was similar to that of 50 ng/mL np-NGF alone.
 | Figure S1 Nontransfected and EGFP-Flag-BMSCs in culture. |
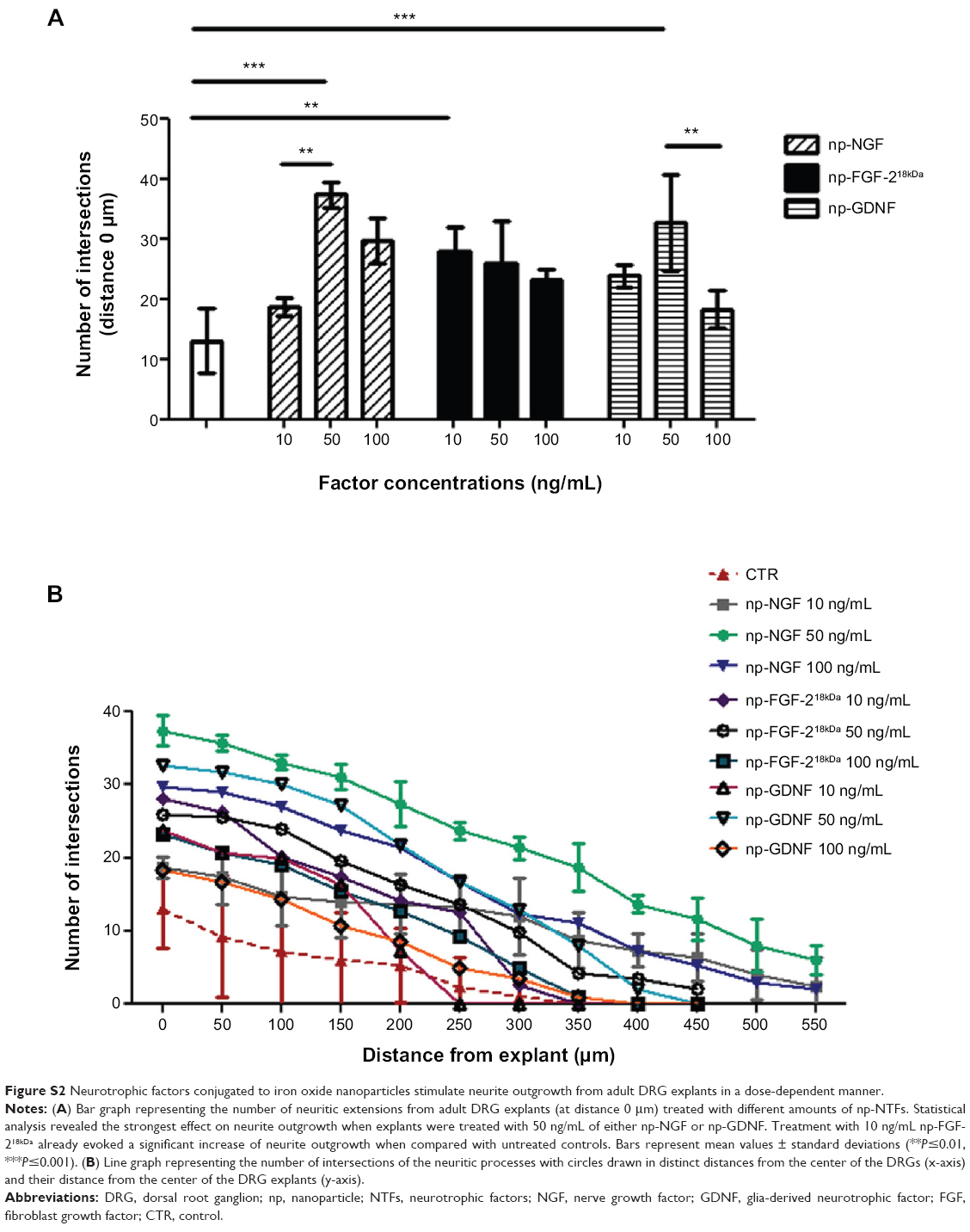 | Figure S2 Neurotrophic factors conjugated to iron oxide nanoparticles stimulate neurite outgrowth from adult DRG explants in a dose-dependent manner. |
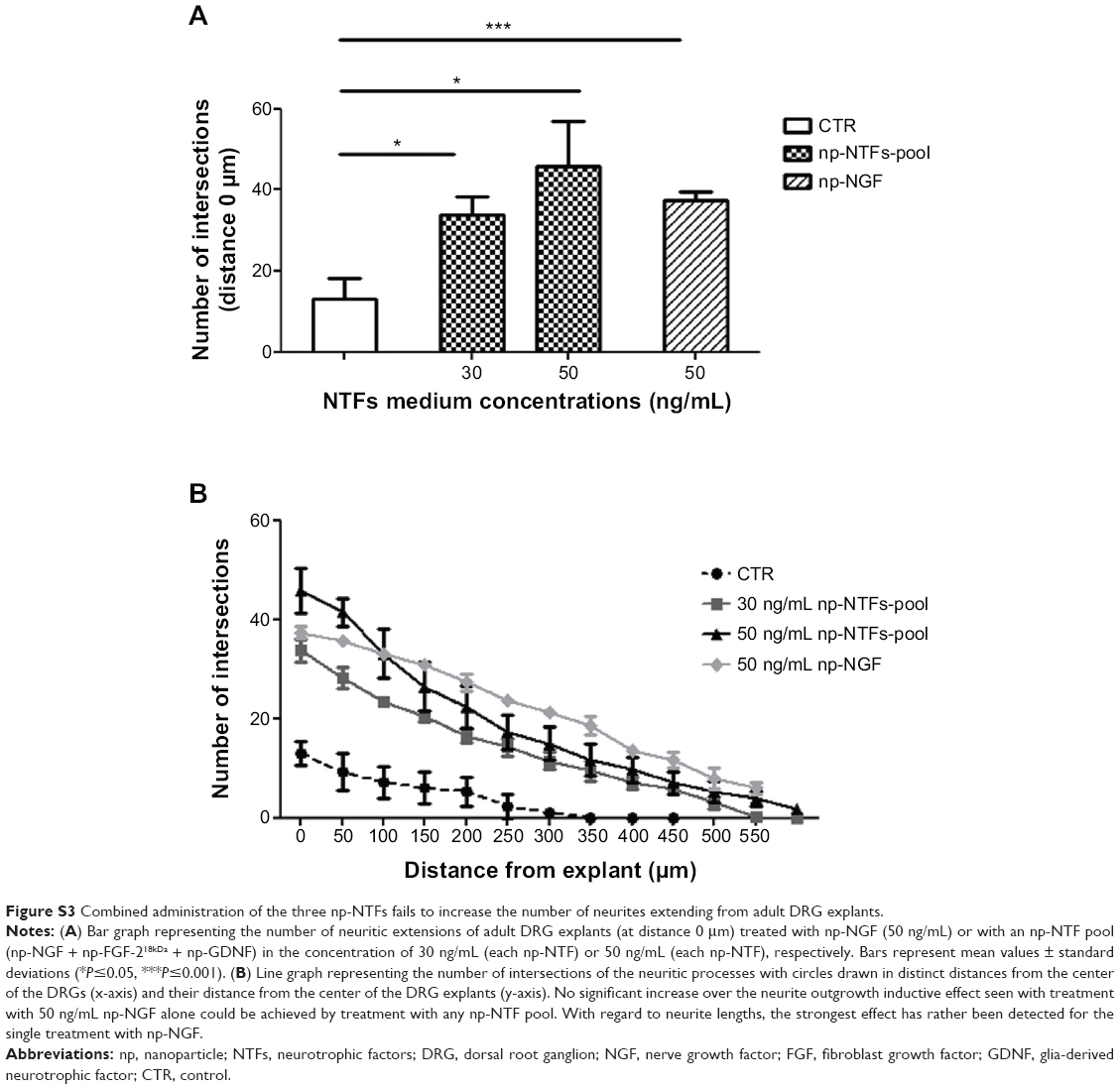 | Figure S3 Combined administration of the three np-NTFs fails to increase the number of neurites extending from adult DRG explants. |
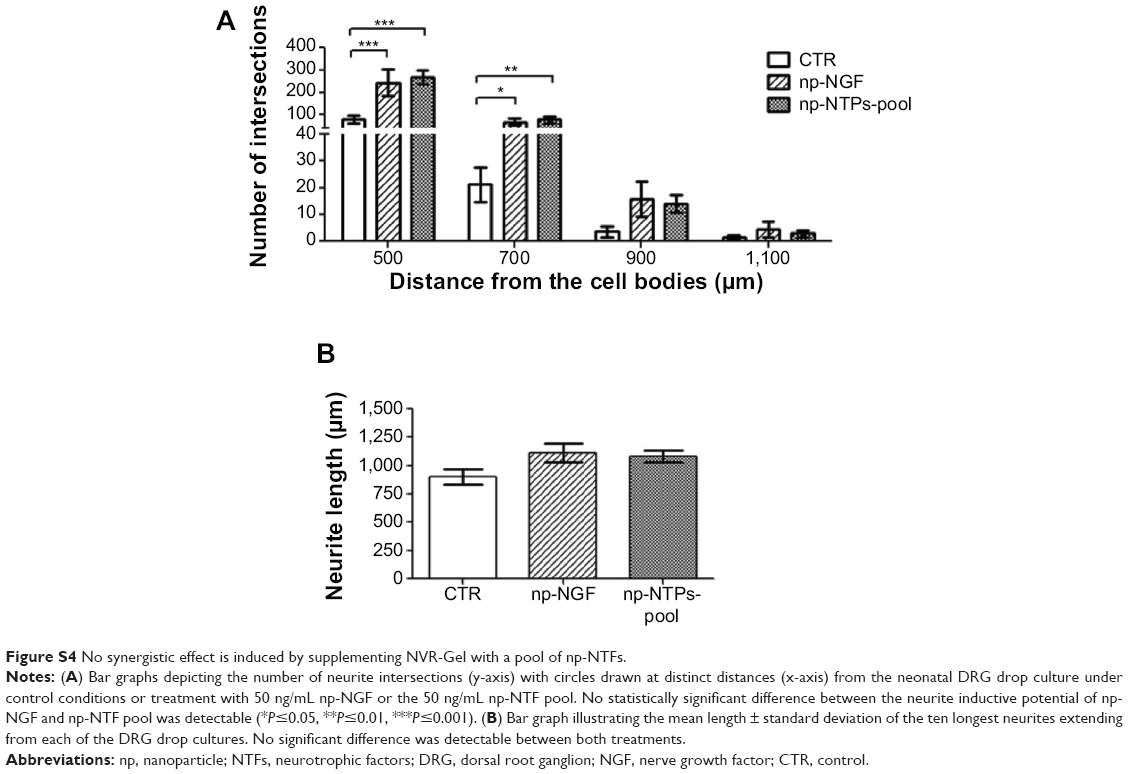 | Figure S4 No synergistic effect is induced by supplementing NVR-Gel with a pool of np-NTFs. |
 © 2014 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2014 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.